Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
3.3. Простая модель стехиометрической структуры энергетического метаболизмаГликолиз и окислительное фосфорилирование имеют автокаталитическую, точнее рефлексивно-каталитическую, стехиометрическую структуру: их продукт АТФ участвует в «активации» субстратов окисления, используемых для фосфорилирования АДФ. В гликолизе активация заключается в фосфорилировании гексоз, в окислительном фосфорилировании — в ацилировании кофермента А (липолиз) или карбоксилировании пирувата. Энергозависимый транспорт субстратов окисления через клеточную или митохондриальную мембраны также можно рассматривать как один из этапов активации. Совокупность реакций, участвующих в энергетическом метаболизме, можно упрощенно представить кинетической моделью [141—143], показанной на схеме (3.1)
В этой модели
и что существует линейный интеграл
Учитывая допущения (3.2), (3.3) и считая, что реакции протекают в среде идеального перемешивания, термо- и рН-статирования, можно получить следующую безразмерную форму математической модели энергетического метаболизма:
где
В модели (3.4) использованы безразмерные переменные и параметры:
Если значения
в которой
— выходная характеристика и
— стехиометрическая эффективность источника энергии, состоящего из реакций первой — четвертой схемы (3.1). График
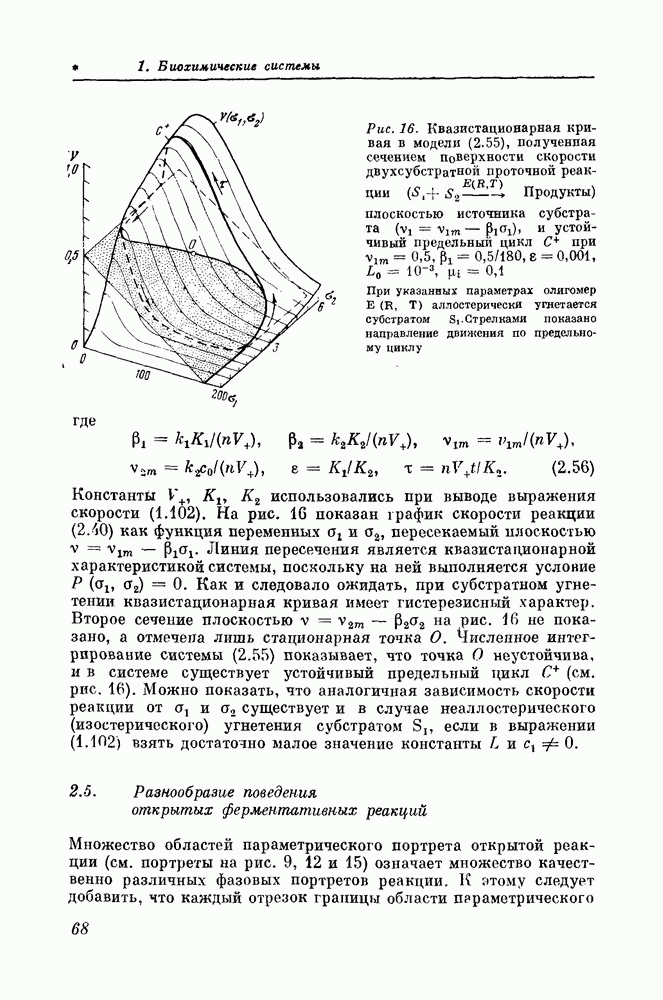
(кликните для просмотра скана) выходной характеристики при фиксированном значении а показан на рис. 23, а. Из рисунка видно, что с возрастанием
и граница нейтральности
построены при помощи параметрического задания в виде
В областях 1 и 3 параметрического портрета все стационарные состояния неустойчивы и модель (3.7) имеет устойчивый предельный цикл (см. рис. 24, а). При исследовании модели энергетического метаболизма были использованы методы, рассмотренные в главе 2 для случая открытых ферментативных реакций. Такие методы, как графическое исследование стационарных состояний, анализ их устойчивости и построение параметрического портрета, оказываются полезными для анализа самых различных процессов, протекающих в проточных условиях. Важнейшей количественной характеристикой любого энергетического метаболизма является его нагрузочная характеристика [144, 147]. Под нагрузочной характеристикой подразумевается зависимость стационарной концентрации АТФ от активности
Рис. 25. Параметрический портрет модели (3.7) энергетического метаболизма (3.1), построенный в плоскости параметров источника
Рис. 26. Семейство стационарных нагрузочных характеристик энергетического метаболизма (3.1), полученных с помощью модели (3.7) при различных активностях утечки обобщенной АТФазы (нагрузки), представляющей суммарную активность всех биохимических процессов, потребляющих АТФ. Семейство таких нагрузочных характеристик, построенных с помощью модели (3.7) для энергетического метаболизма (3.1), приведено на рис. 26. На этом рисунке показана зависимость стационарной концентрации Другой заметной особенностью нагрузочной характеристики является ее гистерезис. Этот гистерезис приводит к тому, что при перегрузке (т. е. при Модель энергетического метаболизма (3.7) является обобщением различных модификаций модели Лотки [166—167]. При Нетрудно заметить, что двойной предельный переход
Здесь штрих-пунктирной стрелкой обозначено автокаталитическое участие АТФ в собственном синтезе. Подобное свертывание сложной полиферментной системы к одной эквивалентной реакции открывает большие возможности для упрощения анализа клеточного метаболизма. При выводе модели (3.7) использовали простейшие гипотезы относительно скоростей
В работе [144] была проанализирована модель стехиометрической структуры энергетического метаболизма, учитывающая как насыщение всех реакций субстратами, так и реакцию (3.14). Исследование этой более полной модели показало, что ее нагрузочная характеристика также имеет четко выраженное плато и гистерезис. При этом гистерезис нагрузочной характеристики может возникать по независимым причинам: 1) из-за дополнительной нелинейности, вносимой в модель реакцией (3.14), 2) из-за конкуренции между генераторной ступенью и утечкой за интермедиат 12 и 3) из-за насыщения нагрузки аденозинтрифосфатом, как и в модели (3.7). В работах [145—154] было продолжено исследование различных вариантов кинетической модели (3.1) и было показано, что во всех ее вариантах неизменно обнаруживается плато нагрузочной характеристики и что в энергетическом метаболизме имеется множество различных стехиометрических взаимодействий, порождающих гистерезис этой характеристики. В некоторых вариантах модели (3.1) была учтена зависимость скорости генераторной ступени от концентрации неорганического фосфата [145—149]. В таких вариантах плато нагрузочной характеристики в широком диапазоне нагрузок имеет положительный наклон, т. е. АТФ растет с увеличением нагрузки. Таким образом, участие фосфата в работе энергетического метаболизма в качестве стехиометрического регулятора приводит к своеобразной перекомпенсации нагрузки: чем больше активность обобщенной АТФазы, тем больше неорганического фосфата и, следовательно, тем интенсивнее работает генераторная ступень. В итоге, в некотором диапазоне нагрузок концентрация АТФ не только не уменьшается, но даже растет с увеличением нагрузки. В публикациях [152—154] были проанализированы варианты модели (3.1), детально описывающие рециркуляцию интермедиатов, существующую в цикле Кребса. Анализ этих вариантов модели (3.1) показал, что нагрузочная характеристика аэробного энергетического метаболизма в условиях интенсивного протока субстратов через цикл Кребса также имеет плато и гистерезис. Таким образом, из анализа стехиометрической структуры энергетического метаболизма, проведенного здесь с помощью модели (3.1), а также с помощью различных обобщений этой модели [141 — 154], следует, что плато и гистерезис нагрузочной характеристики являются фундаментальными свойствами любого энергетического метаболизма, имеющего автокаталитическую природу.
|
 |
Оглавление
|









































































































































































































































































































 субстрат, и
субстрат, и  - интермедиаты, Р - продукт окисления,
- интермедиаты, Р - продукт окисления,  скорость источника,
скорость источника,  скорость инициаторной ступени, в которой на активацию одной молекулы
скорость инициаторной ступени, в которой на активацию одной молекулы  расходуется молекул
расходуется молекул  скорость распада на
скорость распада на  молекул
молекул  скорость генераторной ступени, в которой окисление одной молекулы
скорость генераторной ступени, в которой окисление одной молекулы  до
до  сопровождается фосфорилированием
сопровождается фосфорилированием  молекул АДФ,
молекул АДФ,  - скорость утечки,
- скорость утечки,  скорость гидролиза АТФ нагрузкой,
скорость гидролиза АТФ нагрузкой,  скорость образования АТФ альтернативным источником. При анализе схемы (3.1) примем, что неорганический фосфат и окисляющий
скорость образования АТФ альтернативным источником. При анализе схемы (3.1) примем, что неорганический фосфат и окисляющий  кофактор имеются в избытке. Примем также, что скорости реакций схемы (3.1) могут быть описаны простыми выражениями
кофактор имеются в избытке. Примем также, что скорости реакций схемы (3.1) могут быть описаны простыми выражениями





 достаточно малы, то при помощи предельного перехода
достаточно малы, то при помощи предельного перехода  модель (3.4) сводится к системе двух дифференциальных уравнений:
модель (3.4) сводится к системе двух дифференциальных уравнений:




 скорость продукции АТФ вначале возрастает, а затем, достигнув максимума, падает. При больших
скорость продукции АТФ вначале возрастает, а затем, достигнув максимума, падает. При больших  скорость
скорость  может принимать отрицательные значения, т. е. энергетический метаболизм становится потребителем АТФ. Это связано с тем, что при большом
может принимать отрицательные значения, т. е. энергетический метаболизм становится потребителем АТФ. Это связано с тем, что при большом  скорость
скорость  мала
мала  мало) и фосфорилированный интермедиат 12 оттекает без синтеза АТФ. Уравнение для квазистационарных состояний
мало) и фосфорилированный интермедиат 12 оттекает без синтеза АТФ. Уравнение для квазистационарных состояний  легко решается графически посредством нахождения точек пересечения графика выходной характеристики с линией
легко решается графически посредством нахождения точек пересечения графика выходной характеристики с линией  На рис. 23, б показаны три квазистационарных состояния, а при больших или, напротив, малых а должно быть только одно квазистационарное состояние. Следовательно, квазистационарная кривая в плоскости
На рис. 23, б показаны три квазистационарных состояния, а при больших или, напротив, малых а должно быть только одно квазистационарное состояние. Следовательно, квазистационарная кривая в плоскости  должна иметь гистерезисный характер (рис. 24).
должна иметь гистерезисный характер (рис. 24).  -образность квазистационарной входной характеристики делает возможным существование не только одного (рис. 24, а), но и трех (рис. 24, б) стационарных состояний, которые находятся проведением на плоскости
-образность квазистационарной входной характеристики делает возможным существование не только одного (рис. 24, а), но и трех (рис. 24, б) стационарных состояний, которые находятся проведением на плоскости  прямой
прямой  в соответствии с уравнением
в соответствии с уравнением  На параметрическом портрете в плоскости
На параметрическом портрете в плоскости  область существования трех альтернативных стационарных состояний имеет клинообразный вид (рис. 25). Граница этой области
область существования трех альтернативных стационарных состояний имеет клинообразный вид (рис. 25). Граница этой области




 при
при  На портрете проведены линии нейтральности
На портрете проведены линии нейтральности  и линии моностационарности
и линии моностационарности  Стационарное состояние О (см. рис. 24, б) неустойчиво в области 1 и устойчиво в областях 4 и 6. Состояние
Стационарное состояние О (см. рис. 24, б) неустойчиво в области 1 и устойчиво в областях 4 и 6. Состояние  рис. 24, а) неустойчиво в областях 3 и 4 и устойчиво в областях 5 и 6. Состояние
рис. 24, а) неустойчиво в областях 3 и 4 и устойчиво в областях 5 и 6. Состояние  будучи седлом неустойчиво во всех областях
будучи седлом неустойчиво во всех областях  своего существования
своего существования

 (цифры на кривых)
(цифры на кривых)  безразмерная стационарная концентрация АТФ, удовлетворяющая условию
безразмерная стационарная концентрация АТФ, удовлетворяющая условию  относительная активность АТФазы (нагрузки). Параметры:
относительная активность АТФазы (нагрузки). Параметры:  Пунктир — неустойчивый участок нагрузочной характеристики
Пунктир — неустойчивый участок нагрузочной характеристики
 от безразмерной активности АТФазы
от безразмерной активности АТФазы  при различных относительных активностях утечки
при различных относительных активностях утечки  Кривые
Кривые  получены на основании условия стационарности
получены на основании условия стационарности  для модели (3.7). Как видно из рис. 26, при малой активности утечки нагрузочная характеристика обладает важной особенностью: с изменением нагрузки в широком диапазоне
для модели (3.7). Как видно из рис. 26, при малой активности утечки нагрузочная характеристика обладает важной особенностью: с изменением нагрузки в широком диапазоне  где
где  критическое значение нагрузки) стационарная концентрация АТФ меняется очень слабо
критическое значение нагрузки) стационарная концентрация АТФ меняется очень слабо  Таким образом, стехиометрическая структура энергетического метаболизма (3.1) способна без участия каких-либо аллостерических регуляторных связей с высокой точностью поддерживать стабильную концентрацию АТФ в широком диапазоне изменений нагрузки.
Таким образом, стехиометрическая структура энергетического метаболизма (3.1) способна без участия каких-либо аллостерических регуляторных связей с высокой точностью поддерживать стабильную концентрацию АТФ в широком диапазоне изменений нагрузки.
 ) в энергетическом метаболизме скачком устанавливается очень низкая стационарная концентрация АТФ, определяемая лишь активностью альтернативного источника
) в энергетическом метаболизме скачком устанавливается очень низкая стационарная концентрация АТФ, определяемая лишь активностью альтернативного источника  Для возвращения перегруженного энергетического метаболизма в рабочее состояние — в состояние активной стабилизации [АТФ] - необходимо резко уменьшить нагрузку
Для возвращения перегруженного энергетического метаболизма в рабочее состояние — в состояние активной стабилизации [АТФ] - необходимо резко уменьшить нагрузку 
 модель сводится к модификации модели Лотки, рассмотренной Павлидисом и Козменом [156], при
модель сводится к модификации модели Лотки, рассмотренной Павлидисом и Козменом [156], при  к модели Хиггинса [107] и, наконец, при
к модели Хиггинса [107] и, наконец, при  к оригинальной модели Лотки [167].
к оригинальной модели Лотки [167].
 использованный при выводе модели (3.7) и исключающий концентрации интермедиатов и 12 как несущественные быстрые переменные, редуцирует кинетическую модель (3.1) до существенно более простой модели, показанной на схеме:
использованный при выводе модели (3.7) и исключающий концентрации интермедиатов и 12 как несущественные быстрые переменные, редуцирует кинетическую модель (3.1) до существенно более простой модели, показанной на схеме:

 считая, что скорости этих реакций линейно зависят от концентраций соответствующих интермедиатов и коферментов. Кроме того, при выводе не учитывалось, что в клетке имеется фермент, аденилаткиназа
считая, что скорости этих реакций линейно зависят от концентраций соответствующих интермедиатов и коферментов. Кроме того, при выводе не учитывалось, что в клетке имеется фермент, аденилаткиназа  катализирующий быструю обратимую реакцию
катализирующий быструю обратимую реакцию
