Пред.
След.
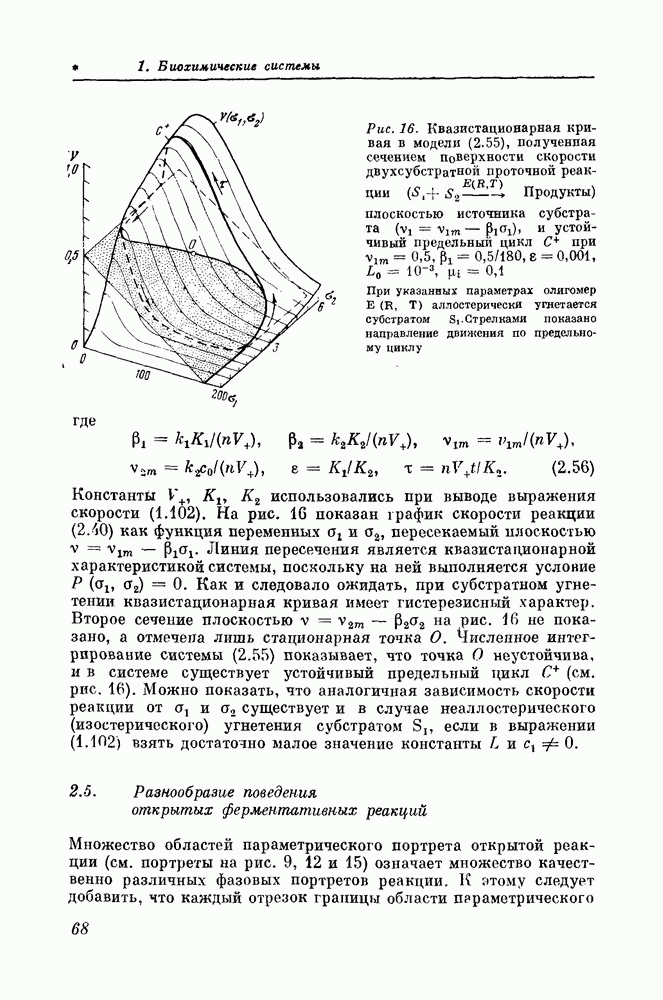
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
4.4. Исследование основных режимов мембраны волокна Пуркинье сердца с помощью качественных методовДля мембраны аксона кальмара качественные методы оказались довольно эффективным инструментом исследования. В сердечной мышце потенциал действия примерно в 100 раз продолжительнее, чем в аксоне кальмара, а релаксационность значительно больше (длительность потенциала действия Покой, спонтанная активность, повторные ответы, устойчивая деполяризация и триггерные свойства в модели мембраны волокна Пуркинье анализируются точно так же, как и в модели аксона кальмара. Границы изменения параметров, при которых в уравнениях Нобла возникают эти режимы, определяются по графикам нуль-изоклин системы Два типа автоколебаний. В мембране волокна Пуркинье экспериментально обнаружены два типа автоколебаний [34]: нормальные (потенциал меняется в диапазоне от —100 до Их появление просто объясняется с помощью системы «Аконитиновые» потенциалы действия. Известно, что алкалоид аконитин вызывает потенциалы действия (рис. 40, а) с колебаниями на плато 135]. Ионный механизм этого явления до конца неясен, поскольку аконитин одновременно меняет несколько различных характеристик мембраны [36].
(кликните для просмотра скана)
Рис. 39. Рождение нового предельного цикла в уравнениях
Рис. 40. «Аконитиновые» потенциалы действия, связанные с жестким режимом возбуждения в уравнениях Нобла Система
Рис. 41. Отсутствие аккомодации в модели волокна Пуркинье а — рассчитанные на ЦВМ изменения мембранного потенциала в ответ на прямоугольный толчок (1) и на линейно нарастающие токи
Рис. 42. Отсутствие анодно-размыкательного ответа в модели Нобла лишь при уменьшении Аккомодация. По фазовому портрету системы току реобазы Прямые расчеты на ЦВМ системы Анодное размыкательное возбуждение. В большинстве возбудимых мембран потенциал действия возникает не только в ответ на включение катодного тока, но и при размыкании анодного тока. Однако нуль-изоклины системы Аналогично по фазовому портрету, построенному по записям ионных токов (глава 5), было предсказано отсутствие анодного размыкательного возбуждения в мембране волокна предсердия лягушки. В дальнейшем это экспериментально подтвердилось [45]. Причина различного поведения моделей Ходжкина — Хаксли и Нобла при выключении анодного тока легко устанавливается при сравнении фазовых портретов редуцированных систем. В мембране Ходжкина — Хаксли при большом гиперполяризующем токе новое состояние равновесия оказывается ниже минимума изоклины Аналогично с помощью качественных методов но фазовому портрету системы
|
 |
Оглавление
|








































































































































































































































































































 а длительность переднего фронта
а длительность переднего фронта  Это дает основание ожидать, что и здесь применение качественных методов окажется успешным, а точность анализа по нуль-изоклинам редуцированных моделей будет еще выше.
Это дает основание ожидать, что и здесь применение качественных методов окажется успешным, а точность анализа по нуль-изоклинам редуцированных моделей будет еще выше.
 с точностью до долей
с точностью до долей  (см. табл. 2, рис. 38). Ввиду того, что эти эффекты качественно те же, что и для модели Ходжкина — Хаксли, далее основное внимание будет уделено специфическим явлениям, характерным именно для модели Нобла.
(см. табл. 2, рис. 38). Ввиду того, что эти эффекты качественно те же, что и для модели Ходжкина — Хаксли, далее основное внимание будет уделено специфическим явлениям, характерным именно для модели Нобла.
 и колебания на уровне плато потенциала действия (от —50 до
и колебания на уровне плато потенциала действия (от —50 до 
 На фазовом портрете видно, что кроме исходного предельного цикла (рис. 39, а, в) при изменении параметров в системе
На фазовом портрете видно, что кроме исходного предельного цикла (рис. 39, а, в) при изменении параметров в системе  может рождаться «малый» предельный цикл (рис. 39, б, г). Колебания потенциала мембраны на этих предельных циклах качественно соответствуют экспериментально наблюдаемым (рис. 40, а).
может рождаться «малый» предельный цикл (рис. 39, б, г). Колебания потенциала мембраны на этих предельных циклах качественно соответствуют экспериментально наблюдаемым (рис. 40, а).


 соответствующее скачкообразной смене типов автоколебаний в модели
соответствующее скачкообразной смене типов автоколебаний в модели  потенциалы действия в
потенциалы действия в  сдвинута на
сдвинута на  соответственно);
соответственно);  проекции предельных циклов системы
проекции предельных циклов системы  на плоскость
на плоскость  (цифры на кривых — величины
(цифры на кривых — величины  пунктирные кривые — нуль-изоклины уравнений
пунктирные кривые — нуль-изоклины уравнений 

 а — мембранный потенциал (стрелкой указан момент стимуляции);
а — мембранный потенциал (стрелкой указан момент стимуляции);  нуль-изоклины уравнений
нуль-изоклины уравнений  (пунктирные кривые) в норме и при сдвиге
(пунктирные кривые) в норме и при сдвиге  на
на  (цифры на кривых — величины
(цифры на кривых — величины  Сплошная кривая — проекция траектории изображающей точки в
Сплошная кривая — проекция траектории изображающей точки в  на фазовую плоскость. В системе
на фазовую плоскость. В системе  стационарная точка
стационарная точка  устойчива, и существует предельный цикл
устойчива, и существует предельный цикл
 позволяет исследовать механизмы возникновения аконитиновых потенциалов действия. Потенциалы действия такой формы появляются, бели на фазовой плоскости системы есть «малый» предельный цикл и в отличие от случая, рассмотренного в предыдущем пункте, на левой ветви сохраняется стационарная точка (рис. 40, б). Поскольку такой фазовый портрет воаникает
позволяет исследовать механизмы возникновения аконитиновых потенциалов действия. Потенциалы действия такой формы появляются, бели на фазовой плоскости системы есть «малый» предельный цикл и в отличие от случая, рассмотренного в предыдущем пункте, на левой ветви сохраняется стационарная точка (рис. 40, б). Поскольку такой фазовый портрет воаникает

 различной крутизны
различной крутизны  соответственно);
соответственно);  аккомодационная кривая; при уменьшении крутизны линейно нарастающего тока повышения порога срабатывания не наблюдается
аккомодационная кривая; при уменьшении крутизны линейно нарастающего тока повышения порога срабатывания не наблюдается

 Для сравнения показано возникновение ответа в модели Ходжкина — Хаксли
Для сравнения показано возникновение ответа в модели Ходжкина — Хаксли  фазовые портреты редуцированных уравнений Ходжкина-Хаксли Н - Н2 и Нобла
фазовые портреты редуцированных уравнений Ходжкина-Хаксли Н - Н2 и Нобла  при действии гиперполяризующего тока (пунктирная кривая — изоклина
при действии гиперполяризующего тока (пунктирная кривая — изоклина  Траектория представляющей точки после размыкания тока показана стрелкой
Траектория представляющей точки после размыкания тока показана стрелкой
 можно сделать вывод, что основным механизмом, ответственным за появления «зконитиновых»
можно сделать вывод, что основным механизмом, ответственным за появления «зконитиновых»  является уменьшение инактивации натриевого тока.
является уменьшение инактивации натриевого тока.
 видно, что в модели Нобла повторные ответы возникают при токах, равных
видно, что в модели Нобла повторные ответы возникают при токах, равных
 Поэтому (см. раздел 4.3) аккомодация в этой модели должна отсутствовать.
Поэтому (см. раздел 4.3) аккомодация в этой модели должна отсутствовать.
 подтвердили этот вывод (рис. 41). Экспериментальная проверка показала [37], что в волокнах Пуркинье аккомодация выражена очень слабо либо отсутствует вовсе.
подтвердили этот вывод (рис. 41). Экспериментальная проверка показала [37], что в волокнах Пуркинье аккомодация выражена очень слабо либо отсутствует вовсе.
 предсказывают, что в мембране Нобла в отличие от других мембран анодное размыкателыюе возбуждение должно отсутствовать. Расчеты на ЦВМ полной системы
предсказывают, что в мембране Нобла в отличие от других мембран анодное размыкателыюе возбуждение должно отсутствовать. Расчеты на ЦВМ полной системы  подтверждают и этот вывод (рис. 42).
подтверждают и этот вывод (рис. 42).
 и после выключения тока изображающая точка попадает в область притяжения правой ветви нуль-изоклины
и после выключения тока изображающая точка попадает в область притяжения правой ветви нуль-изоклины  (рис. 42, в), а в мембране Нобла это невозможно ни при каких токах (рис. 42, г). Ионный механизм отсутствия анодного размыкательного возбуждения в мембране Нобла заключается в том, что после выключения гиперполяризующего тока потенциал покоя достигается с таким же калиевым током, какой имеет место в покоящейся мембране.
(рис. 42, в), а в мембране Нобла это невозможно ни при каких токах (рис. 42, г). Ионный механизм отсутствия анодного размыкательного возбуждения в мембране Нобла заключается в том, что после выключения гиперполяризующего тока потенциал покоя достигается с таким же калиевым током, какой имеет место в покоящейся мембране.
 анализируются зависимость формы ответов мембраны Нобла от момента раздражения, латентность
анализируются зависимость формы ответов мембраны Нобла от момента раздражения, латентность  возникновения ответов, жесткие режимы автоколебаний, возникающие при взаимодействии двух мембран (эхо), и другие явления [26].
возникновения ответов, жесткие режимы автоколебаний, возникающие при взаимодействии двух мембран (эхо), и другие явления [26].