1.6. Свойства реакции ...
Рассмотрим реакцию

катализируемую ферментом  удовлетворяющим всем трем требованиям, изложенным в разделе 1.5. Примем, что стерические аналоги субстратов
удовлетворяющим всем трем требованиям, изложенным в разделе 1.5. Примем, что стерические аналоги субстратов  могут конкурентно связываться с активными центрами, а метаболиты-регуляторы связываются аллостерическими центрами фермента.
могут конкурентно связываться с активными центрами, а метаболиты-регуляторы связываются аллостерическими центрами фермента.
Не производя математических выкладок (они аналогичны выкладкам, проведенным в разделе 1.5), запишем выражение для безразмерной квазистационарной скорости реакции (1. 86):

уравнение (1.88) может описывать сигмоидальную зависимость  от
от  при выполнении условий
при выполнении условий

— кривую с максимумом (рис. 1, Б), отражающую субстратное угнетение, а при невыполнении условий (1.89) и (1.90) зависимость  может носить гиперболический или квазигиперболический характер (рис. 1, В). Условия (1.89) и (1.90) являются необходимыми, но не достаточными. Для получения достаточных условий необходимо решить систему нелинейных уравнений, определяющих граничные значения параметров.
может носить гиперболический или квазигиперболический характер (рис. 1, В). Условия (1.89) и (1.90) являются необходимыми, но не достаточными. Для получения достаточных условий необходимо решить систему нелинейных уравнений, определяющих граничные значения параметров.
Необратимая изомеризация фермент-субстратных комплексов
При  модель (1.87) описывает необратимую изомеризацию фермент-субстратных комплексов, происходящую в каждом активном центре олигомера. В этом случае модель (1.87) принимает вид:
модель (1.87) описывает необратимую изомеризацию фермент-субстратных комплексов, происходящую в каждом активном центре олигомера. В этом случае модель (1.87) принимает вид:

Хотя реакция протекает необратимо, продукт  точно так же, как и аналоги
точно так же, как и аналоги  конкурирует с субстратом
конкурирует с субстратом  за свободные активные центры. Введем новый масштаб измерения концентрации субстрата:
за свободные активные центры. Введем новый масштаб измерения концентрации субстрата:

и новые параметры

где

Замена переменных и параметров  приводит модель (1.91) к виду
приводит модель (1.91) к виду

Общий случай
Для его рассмотрения удобно представить скорость (1.87) в виде разности

где

 безразмерные скорости прямой и обратной реакций. По определению (1.98), скорость
безразмерные скорости прямой и обратной реакций. По определению (1.98), скорость  может быть как положительной
может быть как положительной  если
если  так и отрицательной
так и отрицательной  если
если 
В случае обратимой реакции большие концентрации продукта  угнетают прямую реакцию
угнетают прямую реакцию  Не столь очевидно, что малые концентрации продукта
Не столь очевидно, что малые концентрации продукта  способны изостерически активировать прямую реакцию. Необходимые условия существования эффекта изостерической продуктной активации [56, 61] определяются неравенствами
способны изостерически активировать прямую реакцию. Необходимые условия существования эффекта изостерической продуктной активации [56, 61] определяются неравенствами

Если в этих неравенствах положить  то они будут определять условия существования сигмоидальной зависимости начальной квазистационарной скорости обратной реакции
то они будут определять условия существования сигмоидальной зависимости начальной квазистационарной скорости обратной реакции  Отсюда вытекает простое правило для обратимых реакций, катализируемых олигомерными ферментами: субстрат, активирующий реакцию в прямом направлении, при обращении реакции становится продуктом-активатором. Это правило справедливо и в том случае, когда субстрат является аллостерическим активатором фермента. На рис. 3 показана зависимость скорости реакции
Отсюда вытекает простое правило для обратимых реакций, катализируемых олигомерными ферментами: субстрат, активирующий реакцию в прямом направлении, при обращении реакции становится продуктом-активатором. Это правило справедливо и в том случае, когда субстрат является аллостерическим активатором фермента. На рис. 3 показана зависимость скорости реакции  от
от  при различных
при различных  (рис. 3, а) и различных значениях
(рис. 3, а) и различных значениях  (рис. 3, б). Продуктная изостерическая активация наблюдается лишь в определенном диапазоне значений
(рис. 3, б). Продуктная изостерическая активация наблюдается лишь в определенном диапазоне значений  В противоположность ей аллостерическая продуктная активация не сменяется угнетением фермента при больших концентрациях
В противоположность ей аллостерическая продуктная активация не сменяется угнетением фермента при больших концентрациях 
Из рис. 3 видно, что прямая и обратная реакции ведут себя различно в зависимости от добавок продукта и аллостерических эффекторов. Так, если прямая реакция активируется своим продуктом  то обратная реакция только угнетается своим продуктом
то обратная реакция только угнетается своим продуктом  (см. рис. 3, а). Прямая и обратная реакции различаются и по чувствительности к действию аллостерических эффекторов, описываемому функцией
(см. рис. 3, а). Прямая и обратная реакции различаются и по чувствительности к действию аллостерических эффекторов, описываемому функцией  прямая реакция очень чувствительна к
прямая реакция очень чувствительна к

Рис. 5. Зависимость безразмерной скорости  реакции (1.86) от концентрации субстрата
реакции (1.86) от концентрации субстрата  различных значениях концентрации продукта
различных значениях концентрации продукта  (цифры на кривых), построенная но модели (1.87) при
(цифры на кривых), построенная но модели (1.87) при 
но оказывает сильное угнетающее воздействие на обратную реакцию в том же интервале значений а.
Кривые на рис. 4 построены для  При таком значении х обратная реакция термодинамически менее выгодна, чем прямая. Тем не менее, как видно из сопоставления двух семейств, представленных на рисунке, максимальная скорость обратной реакции при
При таком значении х обратная реакция термодинамически менее выгодна, чем прямая. Тем не менее, как видно из сопоставления двух семейств, представленных на рисунке, максимальная скорость обратной реакции при  больше, чем максимальная скорость прямой реакции при аналогичных условиях
больше, чем максимальная скорость прямой реакции при аналогичных условиях  Таким образом, олигомерный фермент может при определенных условиях катализировать термодинамически менее выгодное направление реакции
Таким образом, олигомерный фермент может при определенных условиях катализировать термодинамически менее выгодное направление реакции  с большей эффективностью и большей чувствительностью к действию изо- и аллостерических эффекторов, чем направление
с большей эффективностью и большей чувствительностью к действию изо- и аллостерических эффекторов, чем направление 
Еще один пример способности олигомерных ферментов эффективно катализировать термодинамически невыгодные превращения показан на семействе кривых  построенных на рис. 5 при различных значениях
построенных на рис. 5 при различных значениях  и при
и при  (предпочтительное связывание
(предпочтительное связывание  конформацией
конформацией  (конформация
(конформация  не способна связывать
не способна связывать  (конформация
(конформация  каталитически неактивна),
каталитически неактивна), 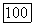 в два раза выше, чем
в два раза выше, чем 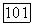 (равновесие
(равновесие 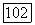 сильно смещено в пользу
сильно смещено в пользу 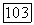 Как видно из рис. 5, из-за субстратного угнетения прямой реакции термодинамически выгодное превращение
Как видно из рис. 5, из-за субстратного угнетения прямой реакции термодинамически выгодное превращение 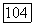 происходит с заметной скоростью лишь при малых концентрациях субстрата и при отсутствии продукта, тогда как обратное превращение
происходит с заметной скоростью лишь при малых концентрациях субстрата и при отсутствии продукта, тогда как обратное превращение 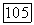 протекает с большой скоростью при любых значениях концентрации продукта
протекает с большой скоростью при любых значениях концентрации продукта 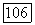
Подобными же свойствами олигомерного фермента можно объяснить парадоксальное сосуществование в клетках двух различных карбамоилфосфат-трансфераз 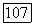 катализирующих реакцию
катализирующих реакцию

в противоположных направлениях. Анаболическая трансфераза, имеющая в различных организмах от 2 до 3 протомеров 111], катализирует термодинамически выгодную реакцию карбамоилирования орнитина. Анаболический фермент имеет гиперболическую кинетику по отношению к своим субстратам и конкурентно угнетается своими продуктами [64]. Катаболическая карбамоилфосфат-трансфераза, имеющая восемь протомеров и молекулярный вес 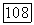 катализирует, как полагают [64], преимущественно термодинамически невыгодную обратную реакцию — фосфоролиз цитруллина. Катаболический фермент характеризуется исключительно высокой кооперативностью взаимодействий со всеми реактантами:
катализирует, как полагают [64], преимущественно термодинамически невыгодную обратную реакцию — фосфоролиз цитруллина. Катаболический фермент характеризуется исключительно высокой кооперативностью взаимодействий со всеми реактантами: 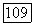 для любого субстрата в анаболическом или катаболическом направлении [64]. Именно этой высокой кооперативностью и объясняется «антитермодинамическое» действие катаболического фермента. Действительно, в соответствии со сформулированным выше правилом катаболический фермент должен кооперативно изостерически активироваться своими продуктами при протекании реакции в любом направлении. По этой причине при высокой концентрации субстратов и низкой концентрации продуктов анаболической реакции катаболический фермент не в состоянии катализировать эту реакцию из-за недостатка цитруллина и
для любого субстрата в анаболическом или катаболическом направлении [64]. Именно этой высокой кооперативностью и объясняется «антитермодинамическое» действие катаболического фермента. Действительно, в соответствии со сформулированным выше правилом катаболический фермент должен кооперативно изостерически активироваться своими продуктами при протекании реакции в любом направлении. По этой причине при высокой концентрации субстратов и низкой концентрации продуктов анаболической реакции катаболический фермент не в состоянии катализировать эту реакцию из-за недостатка цитруллина и 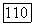 продуктов-активаторов. В этих же условиях анаболический фермент, напротив, максимально активен — субстратов достаточно, а продукты-ингибиторы практически не угнетают его. При высокой концентрации цитруллина и
продуктов-активаторов. В этих же условиях анаболический фермент, напротив, максимально активен — субстратов достаточно, а продукты-ингибиторы практически не угнетают его. При высокой концентрации цитруллина и 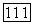 анаболический фермент изостерически угнетен упомянутыми продуктами, а катаболический фермент переводится в активную конформацию, способную в этих условиях катализировать фосфоролиз цитруллина. Переходу в активное конформационное состояние способствуют как большие концентрации цитруллина и
анаболический фермент изостерически угнетен упомянутыми продуктами, а катаболический фермент переводится в активную конформацию, способную в этих условиях катализировать фосфоролиз цитруллина. Переходу в активное конформационное состояние способствуют как большие концентрации цитруллина и  так и повышение концентраций орнитина и карбомоилфосфата, вызванное угнетением анаболического фермента.
так и повышение концентраций орнитина и карбомоилфосфата, вызванное угнетением анаболического фермента.










































































































































































































































































































 реакции (1.86) от безразмерной концентрации субстрата
реакции (1.86) от безразмерной концентрации субстрата  построенная с помощью модели (1.88) при различных значениях аллостерической функции L (цифры на кривых) и при
построенная с помощью модели (1.88) при различных значениях аллостерической функции L (цифры на кривых) и при 
 безразмерная концентрация аналога субстрата А, нормированная на свою константу диссоциации,
безразмерная концентрация аналога субстрата А, нормированная на свою константу диссоциации,  константа неисключающего присоединения для А. Рассмотрим частные случаи модели (1.87).
константа неисключающего присоединения для А. Рассмотрим частные случаи модели (1.87).
 модель (1.87) редуцируется к виду
модель (1.87) редуцируется к виду

 она может описывать сигмоидальную кривую, а также кривую с максимумом. Основным параметром в модели (1. 88), ответственным за эффект субстратной активации или угнетения, является коэффициент неисключающего присоединения
она может описывать сигмоидальную кривую, а также кривую с максимумом. Основным параметром в модели (1. 88), ответственным за эффект субстратной активации или угнетения, является коэффициент неисключающего присоединения  при
при  возможна сигмойдальная или гиперболическая зависимость
возможна сигмойдальная или гиперболическая зависимость  от
от  при 1 — гиперболическая кривая или кривая с максимумом. При выполнении условий
при 1 — гиперболическая кривая или кривая с максимумом. При выполнении условий


 реакции (1.86) от безразмерной концентрации субстрата
реакции (1.86) от безразмерной концентрации субстрата  при различных концентрациях продукта
при различных концентрациях продукта  (цифры на кривых) Кривые построены по модели (1.91) при а
(цифры на кривых) Кривые построены по модели (1.91) при а 
 модель (1.95) может описывать один из трех типов кривых: сигмоидальную кривую при
модель (1.95) может описывать один из трех типов кривых: сигмоидальную кривую при


 то с изменением этих концентраций может меняться тип кинетической зависимости. Например, для одних и тех же параметров:
то с изменением этих концентраций может меняться тип кинетической зависимости. Например, для одних и тех же параметров:  зависимость
зависимость  может описываться сигмоидальной кривой при
может описываться сигмоидальной кривой при  гиперболической — при
гиперболической — при  и кривой с максимумом — при
и кривой с максимумом — при  (рис. 2). Такая же эволюция типа зависимости
(рис. 2). Такая же эволюция типа зависимости  возможна и при изменении концентрации аналога А. Из рис. 2 видно, что с увеличением концентрации продукта скорость его образования резко возрастает. Такая изостерическая продуктная активация является частным случаем изостерической активации олигомерных ферментов аналогами субстрата
возможна и при изменении концентрации аналога А. Из рис. 2 видно, что с увеличением концентрации продукта скорость его образования резко возрастает. Такая изостерическая продуктная активация является частным случаем изостерической активации олигомерных ферментов аналогами субстрата  Заметим, что изостерическая активация в отличие от аллостерической, возникающей при уменьшении
Заметим, что изостерическая активация в отличие от аллостерической, возникающей при уменьшении  наблюдается в определенном диапазоне концентраций аналога субстрата и при больших концентрациях аналога сменяется изостерическим угнетением вследствие конкуренции с субстратом за активные центры фермента.
наблюдается в определенном диапазоне концентраций аналога субстрата и при больших концентрациях аналога сменяется изостерическим угнетением вследствие конкуренции с субстратом за активные центры фермента.

 реакции (1.86) от концентрации продукта
реакции (1.86) от концентрации продукта  при различных значениях концентрации субстрата с (цифры на кривых)
при различных значениях концентрации субстрата с (цифры на кривых)  и аллостерической константы L (цифры на кривых)
и аллостерической константы L (цифры на кривых)  Кривые построены по модели (1.87) при
Кривые построены по модели (1.87) при 

 прямой реакции
прямой реакции  и безразмерной скорости
и безразмерной скорости  обратной реакции
обратной реакции  от безразмерных концентраций
от безразмерных концентраций  и
и  при
при  ; цифры на кривых значения а
; цифры на кривых значения а
 обратная реакция практически нечувствительна к изменениям
обратная реакция практически нечувствительна к изменениям  в данных условиях (см. рис. 3, б). При
в данных условиях (см. рис. 3, б). При  прямая реакция уже не чувствительна к добавкам аллостерических ингибиторов, а обратная реакция угнетается по конкурентному типу. Подобное свойство обратимой реакции наблюдалось экспериментально [62, 63] в форме так называемого однонаправленного действия ингибиторов на обратимую НАДФ-зависимую глутаматдегидрогеназную
прямая реакция уже не чувствительна к добавкам аллостерических ингибиторов, а обратная реакция угнетается по конкурентному типу. Подобное свойство обратимой реакции наблюдалось экспериментально [62, 63] в форме так называемого однонаправленного действия ингибиторов на обратимую НАДФ-зависимую глутаматдегидрогеназную  реакцию. Это явление объясняется сильным различием в сродстве субстратов
реакцию. Это явление объясняется сильным различием в сродстве субстратов  к активным центрам конформаций
к активным центрам конформаций  Однонаправленное действие аналога А на реакцию (1.86) при
Однонаправленное действие аналога А на реакцию (1.86) при  одинаково хорошо присоединяется к конформациям
одинаково хорошо присоединяется к конформациям  и
и  связывается лишь конформацией
связывается лишь конформацией  показано на рис. 4. На рис. 4, а показана зависимость скорости прямой реакции
показано на рис. 4. На рис. 4, а показана зависимость скорости прямой реакции  от
от  построенная при различных концентрациях аналога а и при
построенная при различных концентрациях аналога а и при  а на рис. 4, б — аналогичная зависимость скорости обратной реакции
а на рис. 4, б — аналогичная зависимость скорости обратной реакции  при
при  Как видно из рисунка, аналог А практически не влияет на скорость прямой реакции в интервале концентраций
Как видно из рисунка, аналог А практически не влияет на скорость прямой реакции в интервале концентраций