Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
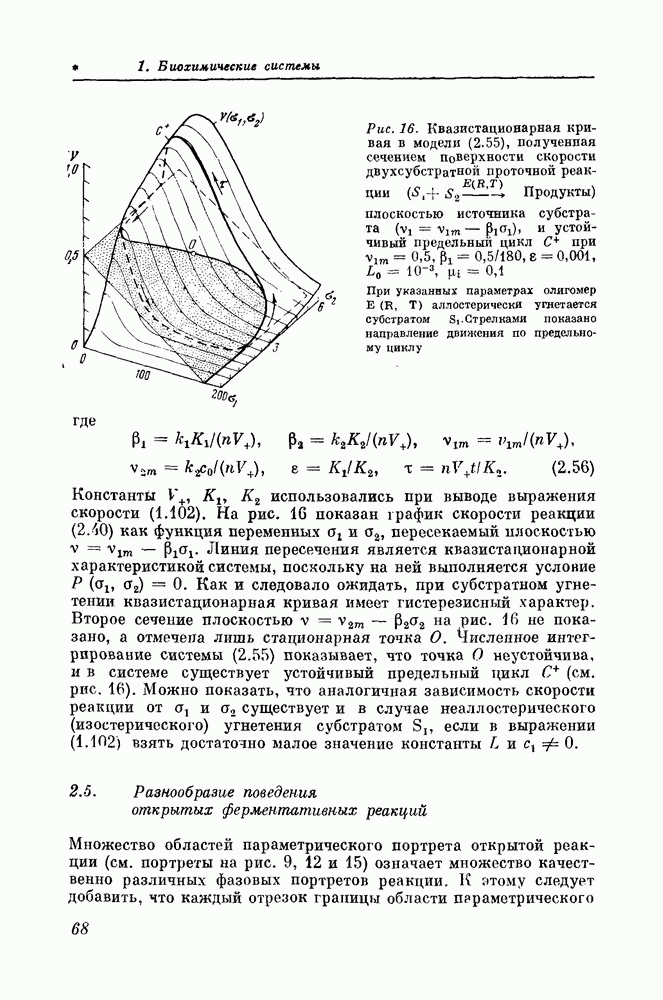
1.5. Модели реакций, катализируемых олигомерными ферментамиПри теоретическом обсуждении свойств регуляторных ферментов и катализируемых ими реакций наиболее часто используются модели Моно, Уаймена и Шанжё [25] (эта модель далее сокращенно называется моделью МУШ), Кошлэнда [52] и Фридена-Курганова [53, 54]. Эти модели учитывают олигомерную четвертичную структуру регуляторного фермента и способность протомеров менять свою конформацию при присоединении к ним аллостерических модификаторов. Модель МУШ [25] выведена в предположении, что олигомерный фермент имеет менять свое конформадионное состояние только совместно. Принято, что существуют только два альтернативных конформационных состояния — активное состояние Согласно модели Кошлэнда [52], присоединение субстрата к активному центру одной субъединицы способно изменять конформацию соседних субъединиц. По этой причине число всех возможных конформационных состояний молекулы фермента, а следовательно, и число параметров, определяющих эти состояния и вероятности переходов между ними, могут быть очень большими, что делает эту модель бесполезной для анализа свойств полиферментных систем. Модель Фридена — Курганова [53, 53] отличается от модели МУШ лишь тем, что совместные конформационные переходы сопровождаются обратимой диссоциацией молекулы фермента на идентичные субъединицы. Все упомянутые модели были выведены для описания простейшей ферментативной реакции, катализируемой отдельным активным центром олигомера, а именно необратимой односубстратной реакции. Между тем подавляющее большинство регуляторных ферментов катализирует двух- и трехсубстратные реакции, многие из которых существенно обратимы. Первая попытка обобщения модели МУШ на случай двухсубстратной необратимой реакции была предпринята Гольдштейном и Волькенштейном [55]. Недавно Поповой и Сельковым модель МУШ была обобщена на случай обратимой односубстратной реакции [56], а затем и наслучай ферментативной реакции произвольной сложности [571. Аналогичное обобщение выведено и для модели Фридена — Курганова [57]. Обобщение модели Моно — Уаймена — Шанжё на многосубстратные реакцииОсновываясь на результатах работы [57], рассмотрим ферментативное превращение
катализируемое ферментом-олигомером Молекула фермента состоит из
Из сказанного следует, что кинетическую схему ферментативной реакции (1.46) можно представить в весьма общем виде — в виде графа:
Здесь
Здесь
где перепишем выражение (1.49):
Рассмотрим первый сомножитель в выражении (1.51). Сравнивая выражения (1.50) и (1.51), нетрудно заметить, что
Из определения (1.52) ясно, что функция скорости есть скорость катализа реакции (1.46) ферментом, представленным только конформацией
Условимся считать, что
Введем регуляторную функцию
Из выражения (1.50) для
Таким образом, для получения выражения (1.46) для начальной квазистационарной скорости реакции достаточно рассмотреть три независимые реакции:
Из реакций (1.57) и (1.58) определяются функции скорости
Итак, с учетом введенных обозначений (1.52), (1.53) и (1.56) уравнение (1.51) можно переписать теперь в следующем виде:
Вывод функций
где
где
Выражение (1.63) еще более упрощается, если принятые ранее допущения дополнить еще одним — протомеры фермента При удовлетворении всех принятых допущений
а скорость описывается простым уравнением
В выражении (1.65) функции субстратов с активным центром в литературе [3,58] нет готовых формул для В качестве примера приведем выражения для начальной квазистационарной скорости обратимой моносубстратной реакции
Здесь
— коэффициенты неисключающего присоединения [25]. Предполагая, что фермент не меняет константу равновесия реакции, потребуем одновременного выполнения двух условий:
Отсюда вытекает термодинамическое ограничение на параметры
или в размерной форме
С учетом выражения (1.69) можно записать:
где
Из графа (1.11) находим
Теперь с учетом выражений (1.64), (1.66), (1.69) и (1.71) — (1.74) уравнение (1.65) для скорости реакции
Обобщение модели Фридена — Курганова для переходов типа тетрамер — димерПусть реакция (1.46) катализируется ферментом-олигомером
В положении равновесия реакции (1.76) имеем
Введя функнии
в виде, аналогичном предыдущему случаю:
Функция отношения
Стационарную концентрацию
или
Решив получившееся уравнение относительно
Пусть фермент удовлетворяет также и принятым ранее допущениям. В этом случае начальная квазистационарная скорость
где
Для рассмотренного в разделе 1.3 примера обратимой односубстратной реакции (1.14) выражения (1.83) и (1.84) принимают вид:
Полученные при довольно простых предположениях относительно механизма взаимодействия субстратов с активными центрами модели реакций, катализируемых олигомерными регуляторными ферментами, отличаются от одноцентровых моделей существенной нелинейностью. Такие модели способны описать многие характерные особенности поведения регулируемых ферментативных реакций при самой простой кинетике катализа, осуществляемого отдельным активным центром протомера.
|
 |
Оглавление
|








































































































































































































































































































 идентичных протомеров, которые могут
идентичных протомеров, которые могут
 и неактивное состояние
и неактивное состояние  между которыми происходят переходы, индуцированные аллостерическими модификаторами. Каждый протомер имеет один активный центр и один или несколько аллостерических центров, причем все центры (активные и аллостерические) считаются идентичными, а присоединение к ним молекул субстратов или модификаторов происходит статистически независимо. Присоединение субстратов к активным центрам фиксирует одно из двух конформационных состояний.
между которыми происходят переходы, индуцированные аллостерическими модификаторами. Каждый протомер имеет один активный центр и один или несколько аллостерических центров, причем все центры (активные и аллостерические) считаются идентичными, а присоединение к ним молекул субстратов или модификаторов происходит статистически независимо. Присоединение субстратов к активным центрам фиксирует одно из двух конформационных состояний.
 субстратов
субстратов  продуктов
продуктов 

 относительно которого примем несколько упрощающих допущений.
относительно которого примем несколько упрощающих допущений.
 протомеров (не обязательно идентичных), причем каждый
протомеров (не обязательно идентичных), причем каждый  протомер имеет два устойчивых конформадионных состояния
протомер имеет два устойчивых конформадионных состояния  соответствующие конформациям
соответствующие конформациям  фермента
фермента  Конформадионные переходы происходят совместно и только между формами фермента, свободными от субстратов:
Конформадионные переходы происходят совместно и только между формами фермента, свободными от субстратов:


 подграфы, описывающие всевозможные взаимодействия субстратов с ферментом в конформадии
подграфы, описывающие всевозможные взаимодействия субстратов с ферментом в конформадии  Если все
Если все  полная концентрация фермента), то к графу (1.48) применимо предположение о квазистационарности (см. раздел 1.3). В этом случае, используя формулу (1.15) и правило (1.20), можно записать выражение для начальной квазистационарной скорости реакции, описываемой графом (1.48):
полная концентрация фермента), то к графу (1.48) применимо предположение о квазистационарности (см. раздел 1.3). В этом случае, используя формулу (1.15) и правило (1.20), можно записать выражение для начальной квазистационарной скорости реакции, описываемой графом (1.48):

 фермент-субстратные комплексы, распадающиеся с константами скорости
фермент-субстратные комплексы, распадающиеся с константами скорости  на продукты реакции,
на продукты реакции,  определители подграфов
определители подграфов  с базой в узлах
с базой в узлах  соответственно,
соответственно,  определители подграфов
определители подграфов  Введем функцию отношения
Введем функцию отношения

 константа равновесия изомерных переходов (1.47), или аллостерическая функция. С учетом равенства (1.50)
константа равновесия изомерных переходов (1.47), или аллостерическая функция. С учетом равенства (1.50)


 Аналогично введем
Аналогично введем

 более активная конформация, чем
более активная конформация, чем  т. е.
т. е.

 как отношение
как отношение

 видно, что
видно, что


 и
и
 и стационарные концентрации
и стационарные концентрации  а из реакции (1.59) — аллостерическая функция
а из реакции (1.59) — аллостерическая функция 

 может оказаться довольно сложной задачей. Она упрощается, если предположить, что присоединение субстратов и продуктов к активным центрам фермента происходит статистически независимо. В этом случае
может оказаться довольно сложной задачей. Она упрощается, если предположить, что присоединение субстратов и продуктов к активным центрам фермента происходит статистически независимо. В этом случае

 функции скорости активных центров
функции скорости активных центров  протомера в конформации
протомера в конформации  Показано [57], что при сделанном предположении функция отношения
Показано [57], что при сделанном предположении функция отношения

 свободные активные центры
свободные активные центры  протомера в конформации
протомера в конформации  Начальная квазистационарная скорость на основе принятых предположений определяется как
Начальная квазистационарная скорость на основе принятых предположений определяется как

 идентичны.
идентичны.



 выводятся для одного активного центра. В этом случае весьма полезным может оказаться метод графов, если для выбранного механизма взаимодействия
выводятся для одного активного центра. В этом случае весьма полезным может оказаться метод графов, если для выбранного механизма взаимодействия

 протекающей по механизму (1.10). Согласно формуле (1.22) запишем выражения для
протекающей по механизму (1.10). Согласно формуле (1.22) запишем выражения для  в безразмерных переменных, определяемых соотношением (1.21):
в безразмерных переменных, определяемых соотношением (1.21):

 максимальная скорость прямой реакции, катализируемой одним протомером в форме
максимальная скорость прямой реакции, катализируемой одним протомером в форме  (по условию
(по условию  а
а







 принимает вид [56]:
принимает вид [56]:

 удовлетворяющим основным требованиям модели
удовлетворяющим основным требованиям модели  с той разницей, что согласованные конформационные переходы связаны с обратимой диссоциацией субъединиц. Рассмотрим частный случай такой диссоциации — диссоциацию активного фермента на две менее активные субъединицы
с той разницей, что согласованные конформационные переходы связаны с обратимой диссоциацией субъединиц. Рассмотрим частный случай такой диссоциации — диссоциацию активного фермента на две менее активные субъединицы


 аналогичные функциям (1.52) и (1.53), запишем начальную квазистационарную скорость реакции
аналогичные функциям (1.52) и (1.53), запишем начальную квазистационарную скорость реакции


 при этом имеет вид:
при этом имеет вид:

 определим из соотношения:
определим из соотношения:


 и подставив выражение для
и подставив выражение для  в выражение (1.80), получим
в выражение (1.80), получим



