Пред.
След.
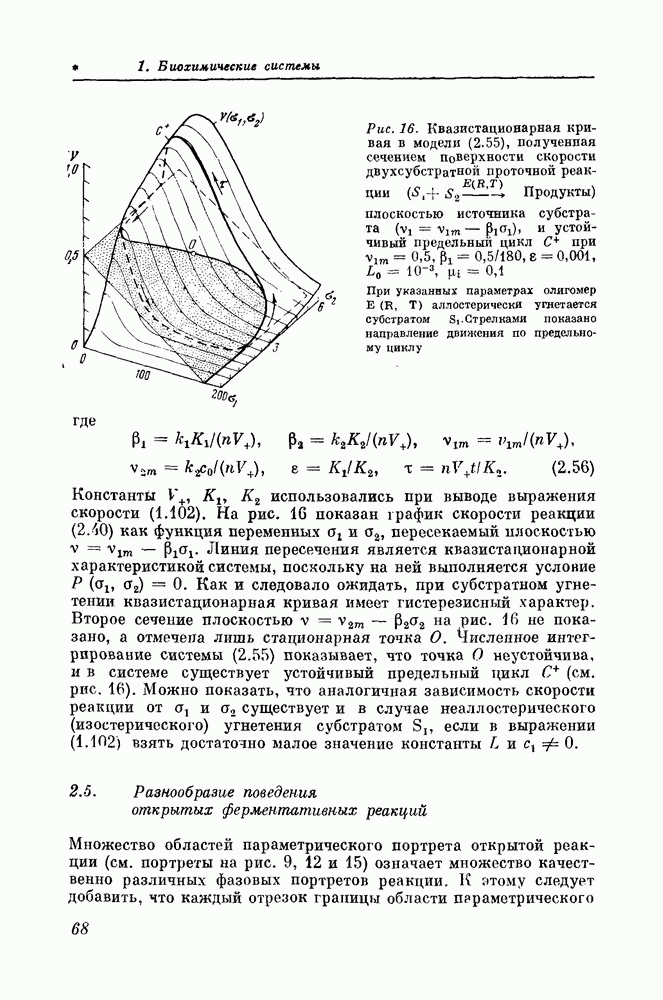
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
9.4. Голографический синтез объемного изображения поверхности клеточной мембраныРеология клеточных мембранРеология рассматривает вопросы течения и деформации сплошных сред. Все тела, встречающиеся в природе, обладают в той или иной степени текучестью. Если для многих «простых» жидкостей вязкость постоянна (это заметил еще Ньютон), то в коллоидах вязкость становится переменной величиной. Возникновение в яшдких дисперсных системах и растворах полимеров пространственных структур, образуемых сцеплением частиц или макромолекул, вызывает резкое повышение вязкости. В «структурированных жидкостях» работа внешней силы затрачивается не только на преодоление обычной (ньютоновской) вязкости, но и на разрушение структуры. Вязкость цитоплазмы клеток прежде всего связана со структурой составляющих ее биополимеров и органоидов. Абсолютная вязкость цитоплазмы колеблется от 2 до Плазматические мембраны клеток обладают заметной эластичностью и низким поверхностным натяжением В физической механике существует теория упругих пленок, основанная на работах К. Ф. Гаусса и У. Гиббса, но все попытки приложения этой теории к описанию мембран живых клеток не увенчались успехом. Возникает вопрос: можно ли к цитоплазме и мембранам живой клетки (т. е. гетерогенным системам, обладающим свойствами гомеостаза, имеющим внутри себя обратные связи) применять понятия физической механики, а именно понятие вязкости, коэффициентов поверхностного натяжения, модуля упругости и т. д.? Клетка является динамической системой, поэтому необходимо исследовать не абсолютные значения коэффициентов, используемых в физической механике, а локальное изменение этих величин во времени и пространстве. При этом следует учитывать три момента. 1. Упаковка молекул фосфолипидов и белков наьязывает мембране, состоящей из них, такую форму, при которой электрический заряд внутри слоя будет минимальным. Поэтому такая пленка легко искривляется в очень малых пределах (пока но произойдет некоторое ее сжатие), а затем для ее дальнейшего искривления требуются большие усилия. Таким образом, изменения вязкости и коэффициента поверхностного натяжения имеют нелинейный характер. 2. Мембрана живой клетки находится на грани фазового перехода от кристаллической структуры к аморфной, т. е. коэффициент вязкости по поверхности мембраны меняется от точки к точке и находится в состоянии динамического равновесия, по-видимому, управляемого белками-«скрепками». Другими словами, часть участков мембраны переходит в аморфное состояние, в свою очередь другая часть переходит в кристаллическое. Это условие также делает классическую теорию гибких пленок несправедливой для биологических мембран. 3. Известно утверждение «мембрана рождает мембрану», т. е. наличие мембраны служит «затравкой» для дальнейшего увеличения ее поверхности, а рост мембраны никак не учитывается в существующей теории. Чтобы избежать перечисленных выше трудностей в применении классической теории упругих пленок к биологическим мембранам, было предложено [25] выделить и отдельно описывать три типа реакции мембран при воздействии на них: первый — изменение формы без изменения площади поверхности при низком модуле упругости (т. е. чистая деформация); второй — увеличение поверхности без изменения формы (т. е. чистый рост) и, наконец, третий — локальное изменение кривизны мембраны за счет местного молекулярного воздействия (например, разжижение мембраны, локальное изменение вязкости). Во втором случае мембране приписывается большой коэффициент упругости. Она как бы пружинит, противодействуя изменению формы. Очевидно, что третий тип реакции мембран активный, а поэтому наиболее сложный. Экспериментально определить изменение коэффициента вязкости по поверхности мембрапы чрезвычайно трудно, поэтому о нем судят косвенно по изменению эластических констант под действием осмотического давления. Однако даже такое грубое приближение дает возможность вычислить величины взаимодействующих сил, возникающих в молекулярных слоях мембран за счет электростатических зарядов. В работе [26] в основу классификации положены физико-химические феномены, поддерживающие целостность мембраны при растягивающих воздействиях. При разрыве мембран возможны два варианта разрушения — лизис и фрагментация. Фрагментация требует в 1000 раз меньше энергии, чем лизис. На основании этого указывалось на существование трех физико-химических факторов, удерживающих целостность мембраны и противодействующих ее разрушению: 1) поверхностное стягивание в результате химического контакта за счет ближайшего окружения, находящегося вне мембраны (например, гидрофобные взаимодействия); 2) поддержание устойчивости оболочки за счет слабых сил молекулярного сцепления внутри мембраны (двумерные, жидкостные характеристики); 3) поддержание устойчивости оболочки за счет сильных молекулярных взаимодействий (свойства участков мембран, близкие к свойствам твердого тела). Таким образом, наиболее слабым местом, определяющим разрыв мембран, является сцепление между двумя липидными слоями. Эти два слоя начинают «скользить» друг относительно друга, приводя к фрагментации мембран [26]. В настоящее время задача реологии клеточных мембран только поставлена. Для ее решения необходимы чувствительные методы локального определения кривизны мембран. Одним из таких методов служит непосредственное наблюдение формы клеточных мембран. Описанный ниже метод, разработанный в Институте биологической физики АН СССР [27], позволяет определять локальную кривизну клеточных мембран по электронно-микроскопическим изображениям клеток. Синтез голограммы по набору ракурсных снимковСинтез голограмм из некогерентно записанных снимков производится в случаях, когда непосредственная запись объемных изображений затруднительна или невозможна из-за отсутствия в соответствующем диапазоне источников когерентного излучения (например, в электронной микроскопии, рентгенографии). Снимки, полученные с помощью сканирующего электронного микроскопа, благодаря значительной глубине фокуса обладают заметным «эффектом объемности». Однако более точная оценка размеров выступающих деталей и рельефа на них затруднительна из-за отсутствия параллакса. В основе метода лежит особенность нашего зрения, позволяющая заменить трехмерный предмет, рассматриваемый одним глазом с данной точки, соответствующим двумерным изображением, полученным методом центрального проецирования с центром в этой же точке. При наблюдении обоими глазами двух плоских изображений, отличающихся на соответствующий ракурсный угол, можно получить стереопару и образ объемного предмета. Составная голограмма позволяет запасти целый набор таких стереопар в значительном угловом диапазоне. Это создает при рассматривании восстановленного изображения полную иллюзию реального предмета. С помощью специальных методов могут быть измерены геометрические параметры поверхности объекта. Процесс синтеза голограмм состоит из двух этапов: получения двумерных изображений объекта под различными углами зрении и трансформирования этих изображений в голограмму. Двумерные изображения, являющиеся исходным материалом для синтеза голограммы, обычно представляют собой фотографии объекта, снятые с помощью некогерентных источников в видимом [28—30], рентгеновском диапазонах длин волн [31, 32] или в электронном пучке [33]. Эти снимки получают, поворачивая объект вокруг оси, проходящей через центр объекта, либо перемещая регистрирующую систему вокруг объекта. Информация о различных ракурсах объекта объединяется затем на одной голограмме путем последовательного экспонирования участков голограммы или многократным экспонированием всей голограммы с соответствующими двумерными изображениями совместно с опорным пучком. Между экспозициями производятся взаимные перемещения элементов системы регистрации голограммы, перемещение маски с прямоугольным отверстием или угловое перемещение двумерного изображения (поворот самой голограммы). Восстановленное изображение представляет собой объемное изображение. При синтезе голограммы большое значение имеет максимальный угол обзора реконструированного трехмерного изображения. В методе, предложенном в Институте биологической физики АН СССР [27], синтез составной голограммы осуществляется по набору ракурсных снимков, полученных на сканирующем электронном микроскопе с сохранением нормального бинокулярного параллакса в одном (например, горизонтальном) направлении, а диапазон углов зрения увеличивается за счет записи голограмм ракурсных снимков не только по горизонтали, но и по вертикали. При просматривании составной голограммы от одного края до другого по вертикали создается эффект вращения трехмерного изображения. Потеря вертикального параллакса несущественна для трехмерного восприятия. Составная голограмма представляет собой прямоугольную матрицу из
Рис. 110. Синтез составной голограммы а — оптическая схема установки: 1 — лазер, 2 — затвор с реле времени, з — полупрозрачное зеркало, 4,5 — зеркало, 6,7 — микрообъективы с точечными диафрагмами, 8 — диапозитив с рассеивателем, 9 — опорный пучок, 10 - предметный пучок, 11 - передвигаемая щель, 12 — голограммная пластинка; Оптическая схема установки приведена на рис. изучаемого микрообъекта получены на сканирующем электронном микроскопе Ориентация диапозитивов должна быть нормальной к линии, соединяющей центр диапозитивов с центром отверстия в маске. Индикатриса рассеяния матового стекла обеспечивает перепое информации в данную точку голограммы со всех точек диапозитива. Изучение геометрии изображения объекта производилось следующим образом. Светящаяся точка (торец световода диаметром От чего зависит разрешение данного метода? При переносе информации с диапозитивов на элементы одного из горизонтальных рядов голограммной пластины центр щели, через которую происходит экспонирование, располагается в той части плоскости, которая соответствовала угловому положению объекта при получении двумерного диапозитива. Последующие ряды матрицы записываются аналогично, но соответствуют сдвинутым на некоторый угол
где гониометра микроскопа при получении ракурсных снимков; Размеры диапозитивов, вводимых в систему синтеза составных голограмм, должны быть достаточными для обеспечения рассматривания с расстояния наилучшего зрения без потери четкости изображения, сформированного электронным микроскопом, а кроме того, не слишком большими, так как при этом увеличиваются габариты всей системы для синтеза составных голограмм. При выборе размеров элементов голограммной матрицы приходится идти на компромисс между необходимостью плавного изменения ракурса объемного изображения при переходе от элемента к элементу матрицы и возможностью наблюдения всего объекта через один элемент голограммной пластины. Чем меньше будет ракурсных снимков (а следовательно, и синтезированных с них элементов составной голограммы), тем ниже будет разрешение деталей рассматриваемого объекта. Наблюдение изображения одним глазом через более чем один элемент матрицы приводит к искажению изображения. Изображения, синтезируемые разными элементами матрицы, соответствуют разным точкам наблюдения. Ограниченность числа точек наблюдения определяет предел разрешения наблюдаемых деталей. Рассмотрим еще один метод — построение трехмерных моделей структур ферментов на основе анализа их электронно-микроскопических изображений. В качестве примера возьмем фермент — лейцинаминопептидазу. Работа но уточнению структуры этого фермента была проведена в Институте биологической физики АН СССР совместно с сотрудниками Центрального института молекулярной биологии АН ГДР [34].
|
 |
Оглавление
|








































































































































































































































































































 Она меняется в различных частях клетки и в разные периоды клеточного цикла. В пределах от
Она меняется в различных частях клетки и в разные периоды клеточного цикла. В пределах от  до
до  вязкость цитоплазмы постоянна, вне этих пределов она увеличивается [23].
вязкость цитоплазмы постоянна, вне этих пределов она увеличивается [23].
 дин/см). Клетки легко деформируются, но при снятии нагрузки они, как правило, приобретают первоначальную форму, определяемую белковым каркасом. Бактериальные и животные клетки благодаря белковому каркасу способны выдержать большое осмоти ческое давление внутри клетки
дин/см). Клетки легко деформируются, но при снятии нагрузки они, как правило, приобретают первоначальную форму, определяемую белковым каркасом. Бактериальные и животные клетки благодаря белковому каркасу способны выдержать большое осмоти ческое давление внутри клетки  атм). Если с помощью лизоцима убрать белковый каркас и оставить только плазматическую мембрану, то клетка приобретает шаровидную форму. Это объясняется тем, что эластичная плазматическая мембрана стремится образовать форму с минимальным запасом энергии [24].
атм). Если с помощью лизоцима убрать белковый каркас и оставить только плазматическую мембрану, то клетка приобретает шаровидную форму. Это объясняется тем, что эластичная плазматическая мембрана стремится образовать форму с минимальным запасом энергии [24].
 прямоугольных элементов.
прямоугольных элементов.

 два снимка мнимого трехмерного изображения нейрона, образующего стереопару (сняты фотоаппаратом с расстояния 25 см от мнимого изображения при базисе
два снимка мнимого трехмерного изображения нейрона, образующего стереопару (сняты фотоаппаратом с расстояния 25 см от мнимого изображения при базисе  линии равной кривизны в изображении изолированного нейрона большого прудовика
линии равной кривизны в изображении изолированного нейрона большого прудовика  паса stapnalis, полученные в результате измерения восстановленного изображения методом «светящейся сетки»
паса stapnalis, полученные в результате измерения восстановленного изображения методом «светящейся сетки»
 Регистрация голограммы осуществляется с помощью гелий-неонового лазера 1 с длиной волны излучения
Регистрация голограммы осуществляется с помощью гелий-неонового лазера 1 с длиной волны излучения  Экспозиция голограммных элементов матрицы производилась с помощью затвора 2, управляемого реле времени. Размеры элемента голограммы
Экспозиция голограммных элементов матрицы производилась с помощью затвора 2, управляемого реле времени. Размеры элемента голограммы  Размеры диапозитивов
Размеры диапозитивов  Расстояние от голограммной пластинки до диапозитива 20 см, от наблюдателя до голограммной пластинки 5 см. Для получения голограммы использовались пластинки типа
Расстояние от голограммной пластинки до диапозитива 20 см, от наблюдателя до голограммной пластинки 5 см. Для получения голограммы использовались пластинки типа  Восстановленное с голограммы изображение обладает эффектом трехмерности. Два изображения, образующие стереопару, показаны на рис. 110, б. В качестве примера рассмотрим поверхность изолированного нейрона большого прудовика Limnaea stapnalis. Ракурсные снимки
Восстановленное с голограммы изображение обладает эффектом трехмерности. Два изображения, образующие стереопару, показаны на рис. 110, б. В качестве примера рассмотрим поверхность изолированного нейрона большого прудовика Limnaea stapnalis. Ракурсные снимки
 работающем в режиме вторичной электронной эмиссии, с помощью гониометра с определенным угловым шагом
работающем в режиме вторичной электронной эмиссии, с помощью гониометра с определенным угловым шагом  Предельный диапазон углов наклона гониометра — 5—46° был расширен с помощью специально изготовленного устройства поворота объекта. На объект напылялся в вакуумной камере тонкий слой углерода, а затем серебра. Снимки (диапозитивы) устанавливались на гониометрический столик и освещались через матовое стекло. Затем последовательно голографировались на элементах составной голограммы через прямоугольную щель, расположенную вплотную к голограммной пластинке. Размеры щели могли регулироваться в двух взаимно перпендикулярных направлениях, а сама щель перемещалась с помощью микрометрического винта.
Предельный диапазон углов наклона гониометра — 5—46° был расширен с помощью специально изготовленного устройства поворота объекта. На объект напылялся в вакуумной камере тонкий слой углерода, а затем серебра. Снимки (диапозитивы) устанавливались на гониометрический столик и освещались через матовое стекло. Затем последовательно голографировались на элементах составной голограммы через прямоугольную щель, расположенную вплотную к голограммной пластинке. Размеры щели могли регулироваться в двух взаимно перпендикулярных направлениях, а сама щель перемещалась с помощью микрометрического винта.
 совмещалась при бинокулярном наблюдении с характерными точками изображения с помощью трехкоординатного устройства перемещения, снабженного шкалой отсчета. Для изучения топографии поверхности объекта использовались также светящиеся сетки, перемещаемые с определенным шагом в направлении наблюдения. Для каждого положения сетки определялся контур поверхности объекта, совпадающий при бинокулярном наблюдении с плоскостью сетки. Светящаяся сетка изготавливалась фотографическим способом и подсвечивалась через матовое стекло. На рис. 110, в представлены контуры одинаковой кривизны поверхности объекта, снятые с помощью светящейся сетки.
совмещалась при бинокулярном наблюдении с характерными точками изображения с помощью трехкоординатного устройства перемещения, снабженного шкалой отсчета. Для изучения топографии поверхности объекта использовались также светящиеся сетки, перемещаемые с определенным шагом в направлении наблюдения. Для каждого положения сетки определялся контур поверхности объекта, совпадающий при бинокулярном наблюдении с плоскостью сетки. Светящаяся сетка изготавливалась фотографическим способом и подсвечивалась через матовое стекло. На рис. 110, в представлены контуры одинаковой кривизны поверхности объекта, снятые с помощью светящейся сетки.
 ракурсам обзора объекта. Полный диапазон углов обзора объекта при этом составляет
ракурсам обзора объекта. Полный диапазон углов обзора объекта при этом составляет

 число элементов ряда матриц,
число элементов ряда матриц,  угловой шаг
угловой шаг
 число рядов голограммной матрицы.
число рядов голограммной матрицы.