Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
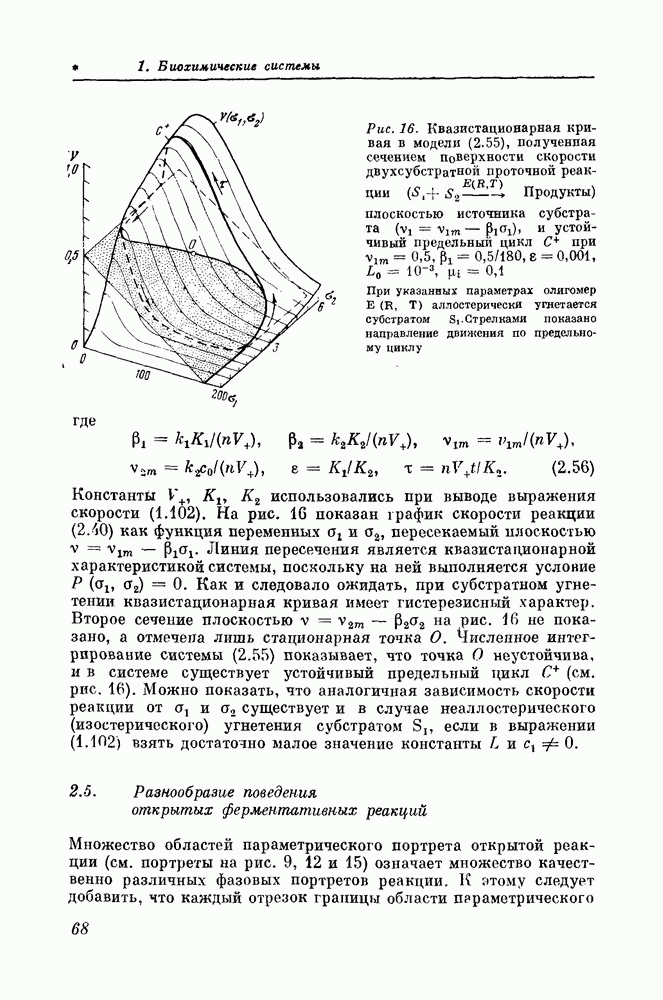
9.3. Модель трехмерной структуры отростка бактериофагаСистема «вирус — клетка»Существуют несколько сотен бактериофагов или вирусов, поражающих бактерии [12, 13]. До недавнего времени основные усилия были сосредоточены на исследовании фагов с отростками, способных размножаться на клетках Escherichia coli, относящихся к штамму В. В настоящее время из них лучше других изучены семь фагов: от Все четные фаги имеют сходное строение (рис. 106). Их различие состоит в величине и толщине отростка, состоящего из стержня и чехла. В очень кратком изложении взаимодействие фага с бактериальной клеткой выглядит так: фаг встречает клетку, затем с помощью своих хвостовых нитей «проверяет» по химическому составу внешней мембраны вид клетки. Если клетка соответствует виду фага, то он прикрепляется к ней снаружи своей базальной пластинкой. Затем базальная пластинка с помощью литического фермента в месте своего крепления к клетке растворяет небольшую зону внешней клеточной мембраны. Чехол хвоста сокращается, выталкивая ДНК (или РНК для некоторых видов фагов) из головы фага внутрь клетки.
Рис. 106. Схема строения бактериофага С этого момента биохимический аппарат клетки в течение двух десятков минут работает под контролем фаговой ДНК. Сначала синтезируются особые ферменты, которые блокируют синтез собственных белков и собственной ДНК клетки и стимулируют синтез фаговой ДНК и фаговых белков. Затем внутри клетки из фаговых белков начинается сборка компонентов новых фагов (хвостов, голов) и целых фагов. Приблизительно через 20 мин весь имевшийся в распоряжении клетки-хозяина биохимический материал будет использован для построения фагов. Оболочка клетки разрывается, лизируется и 100— 200 потомков фага готовы к встрече с новыми бактериальными клетками. Как мы видим, вся проблема взаимодействия вируса и клетки распадается на большое количество отдельных проблем: проблема исследования рецепторных участков мембран (иммунохимическая проблема узнавания), молекулярных механизмов передачи информации ДНК - РНК - белок, морфогенеза (самосборки фагов) и, наконец, биологической подвижности. Сведения о пространственной молекулярной реорганизации отростка бактериофага при сокращении чрезвычайно важны, так как это сократительная структура однократного действия, она не обладает сопутствующими любому двигательному акту системами расслабления и регулирования и потому в наиболее элементарном виде моделирует поведение более сложных двигательных систем. Точные количественные результатывосстановленияпространственной организации такой структуры но электронно-микроскопическим изображениям представляют большой интерес. Для того чтобы увидеть, какова внутренняя структура растянутого хвоста бактериофага и как она изменяется при сокращении чехла, нам пришлось отказаться от фагов группы бактериях Bacillus mycoides и имеет отросток почти в два раза длиннее, чем фаг группы Модель структуры растянутого отростка бактериофагаСтруктура растянутого отростка бактериофага Для отбора качественных электронно-микроскопических изображений и точной оценки параметров спирали отростка фага По положению характерных рефлексов были определены основные параметры растянутой структуры отростка: радиус чехла
(кликните для просмотра скана) Электронно-микроскопическое изображение чехла оказалось эквивалентным 51 проекции элементарной асимметричной группировки. Фотометрирование электронно-микроскопического изображения отростка фага проводилось на микроденситометре при дискретном перемещении предметного столика с шагом Синтез сечений диска осуществлялся методом модифицированного обратного проектирования на квадратной сетке с шагом 6 А в пространстве объекта. Использовалась УЦВМ с оперативной памятью объемом 16 двенадцатиразрядных килослов. Одно из сечений диска, сфотографированное с экрана дисплея Модель структуры сокращенного отростка бактериофагаСложности в исследовании сокращенных форм отростков бактериофагов обычно связываются со значительными вариациями в структуре за счет ее деформации. Это проявляется в первую очередь в плохом качестве картин оптической дифракции, которую они дают. С сокращенного отростка фага На электронно-микроскопическом снимке сокращенной структуры можно выделить два основных семейства спиралей (рис. 108, а): спирали с малым периодом, наклоненные под углом 15° к горизонтальной оси, и спирали с большим периодом, составляющие с вертикальной осью видимо, предельным по разрешению деталям в изображении. Его можно обнаружить только при больших экспозициях и развернутой относительно отростка маске. Исходя из элементарных геометрических соотношений, можно показать, что при полученных параметрах дифракционной картины узлы обратной решетки с ординатами Правило отбора [7] для такой структуры имеет вид: На основании данных дифракционного анализа была осуществлена трехмерная реконструкция сокращенного отростка методом модифицированного обратного проектирования Рассчитанное сечение показано на рис. Хорошее совпадение формы впадин и выступов центральной части сечения и внутренней части чехла, их «комплементарный» характер могут свидетельствовать о плотном прилегании этих частей друг к другу в растянутом отростке. Устранение зазора и более плотная упаковка субъединиц чехла уменьшают диаметр сокращенного отростка, равного 270 А, до поперечных размеров растянутой формы. Характерный вид имеет стержень в сечении. Форма шестиконечной структуры, с длиной лучей примерно 25—30 А, которую имеет стержень, ассоциируется с формой базальной пластинки, которую она приобретает, инициируя сокращение отростка (переход «шестиугольник — шестиконечная звезда») [20]. Сходство усиливается тем, что лучи звезды слегка наклонены в одну сторону. Весьма близкое изображение, полученное в результате цифровой фильтрации изображения базальной пластинки, приведено в работе [14]. Выполненная реконструкция позволяет говорить о том, что подобная форма характерна не только для центральной зоны базальной пластинки, но и для внутренней структуры сокращенного отростка. Центральный канал стержня имеет диаметр около 50 А. Это несколько больше диаметра канала растянутого отростка. Не вполне удовлетворительное совпадение формы при наложении изображений сечений ноличехлов и сокращенной формы базальной пластинки, отмеченное Амосом и Клугом [14], по-видимому, объясняется малым диаметром пустующей зоны в Рис. 108. (см. скан) Структура сокращенного отростка бактериофага сечении поличехлов. Реконструкция показывает, что структура, имеющая форму шестиконечной звезды с наклоненными лучами в центральной части базальной пластинки, вероятно, должна быть связана со стержнем и в результате сокращения полностью отделилась от чехла. Наличие зазора между стержнем и чехлом Рис. 108 (см. скан) (окончание) можно увидель и в радиальных распределениях плотности сокращенного отростка, стоящего на торце (рис. 4, б, в [14]). Возможно, что поличехлы имеют внутренний диаметр, несколько меньший, чем сокращенный чехол. Изменения формы субъединиц чехла связаны с изменением характера укладки полипептидной цепи. По данным реконструкции, каждая из субъединиц имеет два концевых утолщения неравной величины, выступающих на поверхности чехла, связанных между собой перемычкой, спрятанной внутри. Каждая из семейства спиралей с большим периодом образована Рис. 109. (см. скан) Сравнение структурных перестроек в растянутом и сокращенном отростках бактериофага совокупностью одного вида концевых утолщений. Различие в размерах в какой-то степени маскируется достаточно плотной упаковкой на поверхности чехла. Спирали малого периода сформированы путем чередования концевых утолщений обоих видов вдоль каждого витка (рис. 108, д). Изменение формы определяет изменение в положении и числе контактных групп в субъединицах. На периферии чехла проходят шесть внутренних спиральных каналов, вытянутых в тангенциальном направлении, суженных в центральной части и напоминающих цифру 8 с размерами Сравнение сечения сокращенного и растянутого отростков (рис. 109, а) позволяет предполагать, что растянутый отросток представляет собой неустойчивое состояние структуры как по длине, так и по ширине. Отросток растянут и одновременно компактно свернут. Причем стабилизирующей структурой является стержень. Косвенным подтверждением этому могут служить данные морфогенеза, согласно которым стержень является матрицей, на которой происходит сборка чехла [21]. Совокупность событий, приводящих к реорганизации структуры при сокращении, выглядит так. Реорганизация базалыюй пластинки сопровождается структурными перестройками в стержне, в результате которых он (или какая-то неописанная пока структура, плотно прилегающая к нему) приобретает форму шестиконечника с несколько наклоненными в одну сторону лучами. Возможно, что стержень имеет подобную форму и в растянутом отростке, но ее не удается наблюдать из-за плотного прилегания чехла и стержня. Разрыв определенных связей приводит к отделению чехла от стержня и образованию зазора между ними. Чехол разворачивается и одновременно укорачивается. Субъединицы чехла при этом, совершая одновременно две формы движения — по дуге увеличивающегося радиуса при уширении чехла и в вертикальном направлении при укорочении, в результате движутся по коническим винтовым линиям, совпадающим, возможно, с переходными спиралями в модели Муди [22]. Более детального анализа заслуживает характер перестроек, происходящих в субъедшшцах при сокращении. В сечениях сокращенного чехла можно идентифицировать участки, соответствующие внутренним спиральным каналам растянутой структуры. Они лежат в вершинах зазора, образовавшегося между чехлом и стержнем. Они расположены на расстоянии около 60 А. В результате сокращения и отделения чехла от стержня спиральные каналы оказываются незамкнутыми со стороны, обращенной к центру отростка. Кроме того, отделение субъединиц сокращенного чехла друг от друга происходит за счет разрыва боковых стенок спиральных каналов. Таким образом, можно более четко определить локализацию структуры в сечении растянутого чехла, из которой в результате разрыва определенных связей и конформационного перехода формируются субъединицы сокращенного чехла. Она ограничена местами потенциальных разрывов между субъединицами и внутренним спиральным каналом. На рис. 109, б совмещены усредненные по шести результатам восстановления изображения сечений субъединиц растянутого и сокращенного чехла (сплошная линия соответствует субъединице растянутого чехла, пунктирная — сокращенного). В обоих случаях субъединицы имеют близкую форму, напоминающую рыболовный крючок. Преобразование формы при сокращении должно включать в себя, по-видимому, движение большего концевого утолщения (вдоль направления указанного стрелкой) и некоторую модификацию концевых утолщений. Итак, на примере трехмерной модели отростка бактериофага показано, что методы восстановления объемных биологических структур дают наглядное представление о механике пространственных перестроек «биологических молекулярных машин». Теперь перейдем к изложению метода, позволяющего изучать рельеф поверхности клеток.
|
 |
Оглавление
|








































































































































































































































































































 до
до 

 так как они имеют короткий хвост и при сокращении чехла дифракционное изображение не позволяет определить параметры симметрии отростка и перейти на фаг группы
так как они имеют короткий хвост и при сокращении чехла дифракционное изображение не позволяет определить параметры симметрии отростка и перейти на фаг группы  Он паразитирует на
Он паразитирует на
 Что касается сокращенной структуры чехлов фагов группы
Что касается сокращенной структуры чехлов фагов группы  то в этом отношении значительный интерес представляет работа [14], посвященная трехмерной реконструкции сокращенного отростка бактериофага
то в этом отношении значительный интерес представляет работа [14], посвященная трехмерной реконструкции сокращенного отростка бактериофага  Пространственная модель сокращенного отростка получена путем комбинации результатов трехмерного восстановления поличехлов и изображения сокращенной формы базальной пластинки. И хотя близость структуры сокращенных чехлов и поличехлов выглядела достаточно убедительно, существующее функциональное различие в способах формирования этих структур оставляло открытым вопрос об их полной структурной идентичности. Кроме того, не совсем ясна структурная организация центральной зоны сечепий сокращенного отростка. Поэтому чрезвычайно полезной была трехмерная реконструкция растянутого и сокращенного отростков бактериофага Н-17 Bacillus mycoides [15, 16]. Сначала рассмотрим пространственную организацию растянутого отростка бактериофага
Пространственная модель сокращенного отростка получена путем комбинации результатов трехмерного восстановления поличехлов и изображения сокращенной формы базальной пластинки. И хотя близость структуры сокращенных чехлов и поличехлов выглядела достаточно убедительно, существующее функциональное различие в способах формирования этих структур оставляло открытым вопрос об их полной структурной идентичности. Кроме того, не совсем ясна структурная организация центральной зоны сечепий сокращенного отростка. Поэтому чрезвычайно полезной была трехмерная реконструкция растянутого и сокращенного отростков бактериофага Н-17 Bacillus mycoides [15, 16]. Сначала рассмотрим пространственную организацию растянутого отростка бактериофага 
 в какой-то степени напоминает форму растянутых отростков
в какой-то степени напоминает форму растянутых отростков  -четных фагов [17—18]. Контрастирование препаратов фага проводилось
-четных фагов [17—18]. Контрастирование препаратов фага проводилось  -ным раствором уранилацетата. Съемка осуществлялась на микроскопе
-ным раствором уранилацетата. Съемка осуществлялась на микроскопе  с увеличением
с увеличением  (рис. 107, а).
(рис. 107, а).
 было проведено исследование оптических дифракционных картин с нескольких десятков снимков на оптическом дифрактометре. Характерные особенности дифракционных картин растянутого отростка — наличие четкого меридианального рефлекса, зеркальной симметрии относительно меридиана и двух наиболее сильных слоевых линий, расположенных примерно на
было проведено исследование оптических дифракционных картин с нескольких десятков снимков на оптическом дифрактометре. Характерные особенности дифракционных картин растянутого отростка — наличие четкого меридианального рефлекса, зеркальной симметрии относительно меридиана и двух наиболее сильных слоевых линий, расположенных примерно на  расстояния от меридианального рефлекса до экватора.
расстояния от меридианального рефлекса до экватора.
 , величина заходности
, величина заходности  период прерывной спирали
период прерывной спирали  , толщина дисков 39 А. Нанесение на дифракционную картину обратной решетки (рис. 107, б) и системы слоевых линий позволило определить число субъединиц на период прерывной спирали
, толщина дисков 39 А. Нанесение на дифракционную картину обратной решетки (рис. 107, б) и системы слоевых линий позволило определить число субъединиц на период прерывной спирали  и число оборотов непрерывной спирали на период прерывной
и число оборотов непрерывной спирали на период прерывной  Для проверки рассчитывалось распределение функций Бесселя по слоевым линиям для спирали с параметрами
Для проверки рассчитывалось распределение функций Бесселя по слоевым линиям для спирали с параметрами  (см. выражение 9.7).
(см. выражение 9.7).

 и с прямоугольной щелью в плоскости изображения размером
и с прямоугольной щелью в плоскости изображения размером 
 показано на рис. 107, в. Модель отростка бактериофага
показано на рис. 107, в. Модель отростка бактериофага  показана на рис.
показана на рис.  Она собрана из дисков, повернутых друг относительно друга на угол, равный
Она собрана из дисков, повернутых друг относительно друга на угол, равный  рад. Каждый диск имеет центральный канал диаметром около 30 А. На расстоянии 60 А от центра диска расположены шесть спиральных каналов диаметром 20—30 А. На периферии диска расположены шестьсубъединиц, которые соединяются с центральной частью диска мостиками. Субъединица в радиальном направлении имеет размер около 40 А, а в тангенциальном — около 80 А.
рад. Каждый диск имеет центральный канал диаметром около 30 А. На расстоянии 60 А от центра диска расположены шесть спиральных каналов диаметром 20—30 А. На периферии диска расположены шестьсубъединиц, которые соединяются с центральной частью диска мостиками. Субъединица в радиальном направлении имеет размер около 40 А, а в тангенциальном — около 80 А.
 были отобраны четкие дифракционные картины, позволяющие установить основные параметры структуры. В процессе сокращения растянутый отросток претерпевает значительные изменения.
были отобраны четкие дифракционные картины, позволяющие установить основные параметры структуры. В процессе сокращения растянутый отросток претерпевает значительные изменения.
 На дифракционной картине (рис. 108, б) этим спиралям соответствуют системы рефлексов на 1-й, 16-й и 17-й слоевых линиях. Причем спирали с малым периодом обладают
На дифракционной картине (рис. 108, б) этим спиралям соответствуют системы рефлексов на 1-й, 16-й и 17-й слоевых линиях. Причем спирали с малым периодом обладают  -кратной симметрией, а спирали с большим периодом —
-кратной симметрией, а спирали с большим периодом —  -кратной симметрией. Эти данные согласуются с результатами, полученными на сокращенном отростке фага
-кратной симметрией. Эти данные согласуются с результатами, полученными на сокращенном отростке фага  [19]. На 33-й слоевой линии на отдельных дифракционных картинах можно наблюдать слабый меридианальный рефлекс, соответствующий,
[19]. На 33-й слоевой линии на отдельных дифракционных картинах можно наблюдать слабый меридианальный рефлекс, соответствующий,
 расположенные на линиях
расположенные на линиях  действительно будут лежать на меридиане
действительно будут лежать на меридиане  соответственно ординаты рефлексов на 1-й и 16-й слоевых линиях (рис. 108, в).
соответственно ординаты рефлексов на 1-й и 16-й слоевых линиях (рис. 108, в).
 где
где  Таким образом, каждая из шести прерывных спиралей с основным периодом около 660 А содержит 33 субъединицы в 10 оборотах.
Таким образом, каждая из шести прерывных спиралей с основным периодом около 660 А содержит 33 субъединицы в 10 оборотах.

 Отличительная особенность его состоит в следующем. В сокращенной структуре произошло полное отделение чехла от стержня и между ними образовался зазор, ширина которого составляет примерно 30—35 А, в отличие от структуры растянутого чехла, где между стержнем и чехлом имелись связи.
Отличительная особенность его состоит в следующем. В сокращенной структуре произошло полное отделение чехла от стержня и между ними образовался зазор, ширина которого составляет примерно 30—35 А, в отличие от структуры растянутого чехла, где между стержнем и чехлом имелись связи.
 а — электронно-микроскопическая фотография бактериофага (лучшим по качеству оказалось изображение отростка, при сокращении чехла которого произошло отделение последнего от головки);
а — электронно-микроскопическая фотография бактериофага (лучшим по качеству оказалось изображение отростка, при сокращении чехла которого произошло отделение последнего от головки);  дифракционная картина от сокращенного отростка; в — график
дифракционная картина от сокращенного отростка; в — график  порядок функции Бесселя, I — номера слоевых линий) и радиальная проекция структуры;
порядок функции Бесселя, I — номера слоевых линий) и радиальная проекция структуры;  рассчитанное на УЦВМ сечение сокращенного отростка на экране дисплея (внизу — гистограмма распределений плотности в сечении с указанием уровня ограничения); в — трехмерная модель секращенного отростка
рассчитанное на УЦВМ сечение сокращенного отростка на экране дисплея (внизу — гистограмма распределений плотности в сечении с указанием уровня ограничения); в — трехмерная модель секращенного отростка
 а — сечения растянутого (слева) и сокращенного (справа) отростка; б - усредненные изображения сечений субъединицы растянутого и сокращенного чехла
а — сечения растянутого (слева) и сокращенного (справа) отростка; б - усредненные изображения сечений субъединицы растянутого и сокращенного чехла
