Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
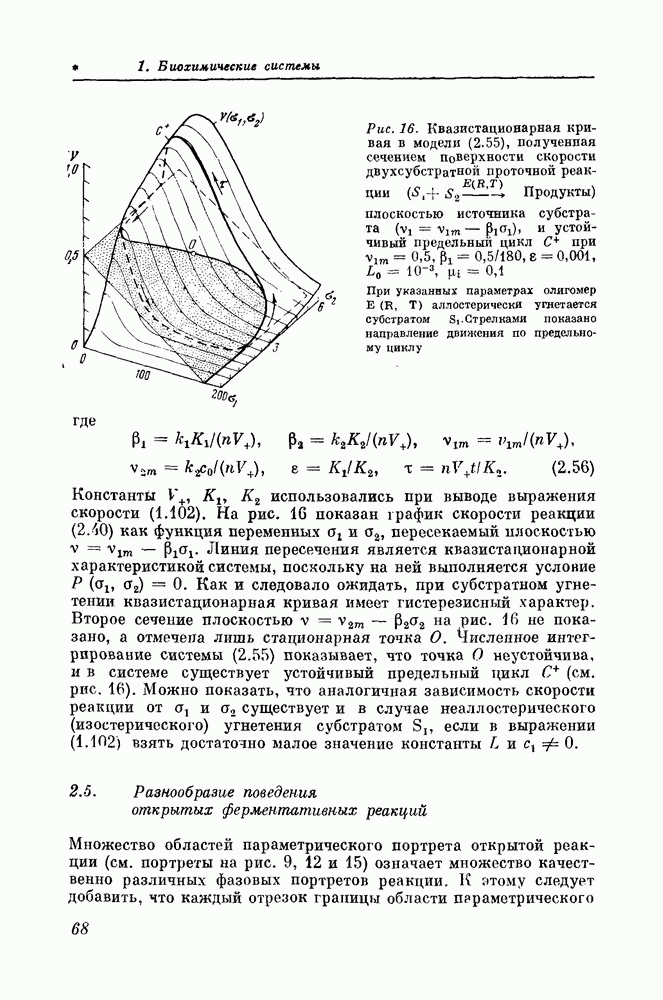
2.5. Разнообразие поведения открытых ферментативных реакцийМножество областей параметрического портрета открытой реакции (см. портреты на рис. 9, 12 и 15) означает множество качественно различных фазовых портретов реакции. К атому следует добавить, что каждый отрезок границы области параметрического портрета, заключенный между двумя соседними пересечениями границ, каждая точка пересечения или касания таких границ, а также точки вырождении, лежащие на границах (например, точки, в которых В свою очередь, каждый фазовый портрет обычно представляет собой совокупность фазовых траекторий различного типа — траекторий, представляющих различные типы поведения моделируемой реакции во времени в зависимости от начальных условий. В общей сложности открытая ферментативная реакция, имеющая гистерезисную квазистационарную входную (выходную) характеристику, может иметь в зависимости от значений параметров или начальных значений концентраций сотни качественно различных форм динамического поведения. Если реакция имеет больше чем три стационарных состояния и (или) описывается большим числом уравнений, то число возможных типов динамического поведения должно быть еще большим. Каждая из исследованных ферментативных реакций отличается большим разнообразием динамического поведения даже тогда, когда имеет гистерезисную квазистационарную характеристику и лишь одно стационарное состояние (рис. 17). Если единственное стационарное состояние устойчиво, то реакция обычно эквифинальна: при любых начальных условиях в ней устанавливается одно и то же стационарное значение скорости. Однако достижение этого значения при нестационарных начальных значениях концентраций реагентов имеет различный характер — это может быть либо монотонное приближение к стационарному значению (рис. 17,1), либо немонотонное релаксационное приближение (рис. 17, 2), либо, наконец, релаксационное движение, сменяющееся затухающими колебаниями (рис. 17, 4). Единственное устойчивое стационарное состояние не всегда гарантирует зквифинальность реакции. Действительно, если состояние окружено двумя предельными циклами, как на фазовом портрете (см. рис. 13, в), то в зависимости от начальных условий реакция окажется после переходного процесса либо в неколебательном, либо в автоколебательном стационарном состоянии (рис. 17,5). Форма автоколебаний может быть различной: от почти гармонической (рис. 17, 6) — такая форма характерна для случаев, и которых представляющая точка параметрического портрета находится вблизи линий нейтральности, до резко релаксационной (рис. 17, 7). Релаксационные колебания неизбежны для реакций, в которых концентрации реагирующих веществ отличаются на несколько порядков друг от друга, причем большие концентрации в таких случаях обычно изменяются во времени пилообразно (рис. 17, 8), а малые — разрывно (рис. 17, 7).
Рис. 17. Формы динамического поведения проточной реакции, имеющей одно стационарное состояние и гистерезисную квазистационарную характеристику, при различных начальных условиях (1-8) или при различной силе возмущения реакции (9—12) 1 — изменение скорости реакции во времени; 2 — устойчивое стационарное значение скорости; 3 — неустойчивое стационарное значение спорости. Стрелками показаны моменты приложения возмущения Особый интерес представляют переходные процессы, вызванные однократным импульсным возмущением одного из параметров, например возмущением скорости поступления субстрата. Если стационарная точка располагается на нижней устойчивой ветви гистерезисной входной характеристики, как на рис. И вблизи нижнего экстремума, то кратковременное увеличение скорости поступления субстрата может вызывать слабый всплеск скорости либо спайкообразное изменение большой амплитуды, завершающееся асимптотическим возвращением реакции в исходное стационарное состояние (рис. 17, 9). Процесс генерации спайкообразного импульса во всех деталях подобен генерации потенциала действии мембранами нервных и мышечных клеток (см. главу 4). В частности, возбуждающий стимул, отклоняющий реакцию от стационарного состояния на некоторое время, должен преодолеть некоторый порог возбуждения. Подпороговый стимул вызывает слабый всплеск скорости, сверхпороговый — импульс стандартной формы (спайк), возникающий через некоторый латентный период после начала действия стимула. Повторное возбуждение характеризуется периодом рефрактерности: еще не пришедшая в стационарное состояние реакция имеет тем более высокий порог возбуждения, чем меньше времени истекло с момента генерации предыдущего спайка. Если стационарное состояние реакции представляется на фазовой плоскости устойчивым фокусом, то возвращение реакции к такому состоянию после возбуждения может носить характер затухающих колебаний (рис. 17, 10). Если же стационарное состояние неустойчиво, но так, что стационарная точка окружена малым предельным циклом, расположенным в окрестности нижнего экстремума входной характеристики, то реакция по-прежнему в состоянии генерировать импульс в ответ на сверхпороговый стимул (рис. 17, 11). Однако в этом случае порог возбуждения периодически изменяется во времени. Если устойчивая стационарная точка располагается на устойчивой верхней ветви вблизи верхнего экстремума входной характеристики, то кратковременное уменьшение скорости поступления субстрата в реакцию способно вызывать генерацию импульса отрицательной полярности (рис. 17, 12). Процессы возбуждения и генерации такого импульса не отличаются от тех же процессов в случае генерации положительного импульса. На рис. 18 представлены некоторые типы динамического поведения открытой реакции, имеющей из-за гистерезиса квазистационарной характеристики три альтернативных стационарных состояния. В зависимости от начальных условий в такой реакции может монотонно (рис. 18,7) или немонотонно устанавливаться одно из двух устойчивых альтернативных стационарных значений скорости. Одно из двух стационарных значений скорости может быть автоколебательным (рис. 18, 4, 5). Если одна из устойчивых стационарных точек окружена неустойчивым предельным циклом (см. рис. 13, в), то в зависимости от начальных условий в реакции устанавливается одно из двух устойчивых стационарных значений скорости (рис. 18, 6). Если фазовый портрет реакции подобен портрету на рис. 13, е, то в такой реакции при одних начальных условиях устанавливается устойчивое стационарное значение скорости, а при других — автоколебания скорости с большой амплитудой (рис. 18, 7). В случае, когда фазовый портрет реакции подобен портрету на рис. 13, д, в зависимости от начальных условий она может оказаться в одном из трех устойчивых состояний: в одной из двух неколебательных стационарных состояний, сильно отличающихся друг от друга стационарными значениями скорости реакции, либо в автоколебательном состоянии, в котором скорость колеблется с большой амплитудой (рис. 18, 8). Для реакции с тремя стационарными состояниями характерно наличие порогов для внешних возмущений, вызывающих смену состояний: подпороговое возмущение вызывает лишь незначительное временное отклонение от устойчивого стационарного
Рис. 18. Возможные типы динамического поведения проточной реакции, имеющей три альтернативных стационарных состояния 1 — изменения скорости реакции во времени, соответствующие трем стационарным точкам состояния, а сверхпороговое — переход реакции в новое состояние (рис. 18, 9—12). При этом возможны различные варианты переходов: из неколебательного в другое неколебательное состояние (рис. 18, 5), из неколебательного в колебательное (рис. 18, 10), из колебательного в неколебательное (рис. 18, 11), из колебательного состояния большой амплитуды в одно из двух неколебательных стационарных состояний, отличающихся установившимся значением скорости (рис. 18, 12). Стоит подчеркнуть тот факт, что все перечисленные формы динамического поведения могут быть получены в математической модели любой открытой ферментативной реакции, имеющей гистерезисную квазистационарную характеристику, если только на величины параметров не наложены какие-либо физические ограничения. Иными словами, широкий спектр динамического поведения проточных реакций инвариантен по отношению к молекулярным механизмам, создающим гистерезис квазистационарной характеристики. Выше упоминалось о большом сходстве между «возбуждением» открытой реакции и электрическим возбуждением биологических мембран. Это сходство основано на общей нелинейности — на гистерезисе квазистационарных характеристик, которые имеют возбудимые реакции и мембраны (см. главу 4). Между тем подобные гистерезисные характеристики (N- или S-образные характеристики, характеристики с падающим участком, характеристики с отрицательным сопротивлением, трением и т. п.) широко распространены в системах самой различной природы. И всякий раз, когда открытая система произвольной физической природы обнаруживает гистерезис квазистационарной характеристики, она обладает тем же набором динамических свойств, какие были выявлены у других гистерезисных систем. Прекрасные примеры таких систем, встречающихся в физике, электронике, физической химии, электрохимии, электронике и в химии открытых гомогенных реакций, а также доказательства сходства динамического поведения всех этих совершенно различных систем можно найти в публикациях Франка [76—79].
|
 |
Оглавление
|








































































































































































































































































































 на границе
на границе  или точка возврата границы
или точка возврата границы  дают свои особые, качественно отличные от других фазовые портреты.
дают свои особые, качественно отличные от других фазовые портреты.


 (см. рис. 7 и 13); 2 — устойчивое стационарное значение скорости; 3 — неустойчивое стационарное значение скорости. Стрелками показаны моменты приложения возмущений различной силы
(см. рис. 7 и 13); 2 — устойчивое стационарное значение скорости; 3 — неустойчивое стационарное значение скорости. Стрелками показаны моменты приложения возмущений различной силы