Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
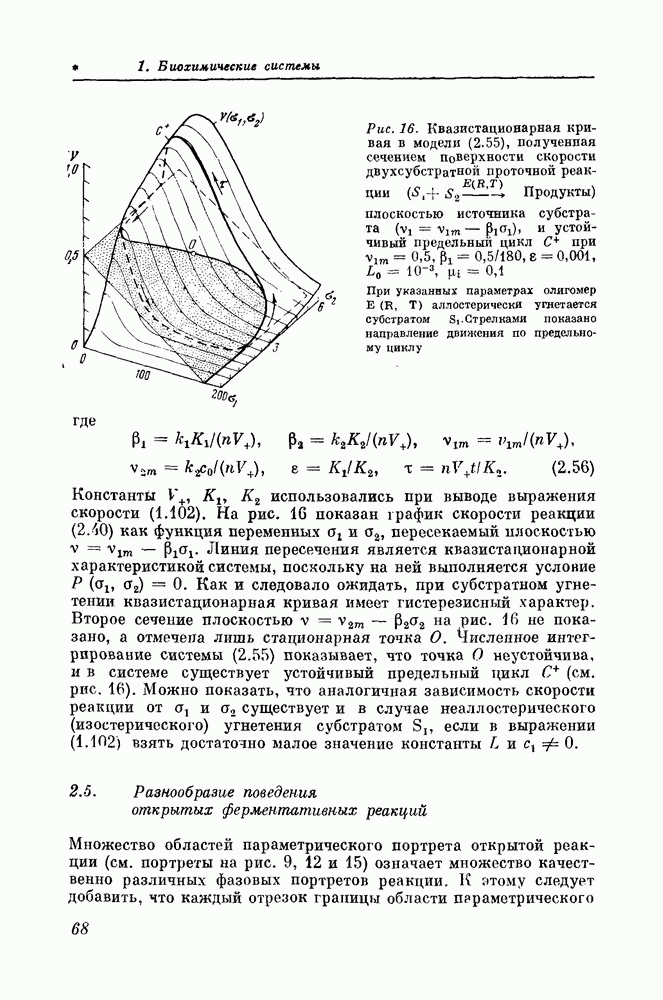
Глава третья. МОДЕЛИРОВАНИЕ ПОЛИФЕРМЕНТНЫХ СИСТЕМ3.1. Полиферментные системы и их моделиТермин «полиферментная система» используется в биохимической литературе для обозначения совокупности взаимосвязанных реакций, выполняющих некоторую биохимическую функцию. Важным для экспериментального и теоретического анализа свойством полиферментных систем является их относительная автономность. Это прежде всего относится к энергетическому метаболизму. Напротив, в анаэробных условиях гликолиз оказывается единственным процессом, обеспечивающим клетку энергией, и служит в основном источником пир овин оградной кислоты, окисление которой в митохондриях обеспечивает клетку энергией. В отсутствие глюкозы образование АТФ митохондриями осуществляется за счет окисления кетонных тел, жирных кислот и аминокислот. Существенная автономность позволяет проводить моделирование полиферментных систем по отдельности, без связей с остальным клеточным метаболизмом, Конечно, такое изолированное рассмотрение полиферментных систем имеет смысл только до тех пор, пока не возникает необходимость исследования их взаимодействий друг с другом. Важным этапом в теоретическом анализе полиферментной системы является изучение особенностей поведения реакций, составляющих эту систему. В предыдущей главе было показано, что угнетение проточной ферментативной реакции субстратом или активация продуктом могут приводить к автоколебаниям. Очевидно, что если одна из реакций полиферментной системы угнетается своим субстратом или активируется продуктом, то можно ожидать, что в такой полиферментной системе могут наблюдаться автоколебания, множественность стационарных состояний и гистерезисные переходы между ними. Так, например, одночастотные колебания в гликолитической системе [87—100] объясняются продуктной активацией ключевой фосфофруктокиназной реакции посредством АМФ или фруктозо-1,6-бисфосфатом [80—86, 90, 100, 106—108]. Однако нередко полиферментные системы проявляют свойства, отсутствующие у составляющих их реакций. В гликолитической системе, например, наблюдаются при определенных условиях двух-частотные колебания [89, 90, 92, 94]. Исследование этого явления с помощью математических моделей [109—113] показало, что двухчастотные колебания объясняются взаимодействием двух гликолитических одночастотных генераторов. Одним из них служит фосфофруктокиназная реакция, активируемая зденозинмонофосфатом, другим — сложная система реакций, вся нижняя часть гликолитической системы, в которой колебания возникают в результате активации пируваткиназы фруктозо-1,6-бисфосфатом или(и) субстратного угнетения глицеральдегидфосфатдегидрогеназы [110, 113]. Таким образом, двухчастотные колебания в гликолизе — типичный «системный эффект». Как бы мы детально не изучали свойства отдельных гликолитических реакций, мы не поймем механизм этого явления до тех пор, пока не рассмотрим, как взаимодействуют друг с другом реакции в гликолитической системе. Другим примером системного эффекта может служить множественность стационарных состояний, возникающая в полиферментной системе при наличии в ней перекрестного угнетения ферментов своими продуктами. Это явление впервые было рассмотрено на простой кинетической модели в 1948 г. Дельбрюком [114]. Позднее Спэнглер и Снелл [115, 116], несколько модифицировав эту кинетическую модель, показали, что перекрестное угнетение может вызвать также и автоколебания. Жакоб и Моно [117], развив идею Дельбрюка, предложили целую серию кинетических моделей для обьяснения триггерных и колебательных явлений, связанных с механизмами генетической регуляции. В последние годы различные варианты моделей, описывающих взаимоотношения между генами-регуляторами и генами-операторами, были предметом тщательного теоретического анализа. Такие исследования проводились Андреевым [118], Чернавским и сотр. [119—121], Ратнером [122, 123] и другими исследователями [124—128]. Еще одним известным из теоретических расчетов системы эффектом являются автоколебания в системах реакций, контролируемых механизмом отрицательной обратной связи [129—140]. Во всех цитированных до сих пор теоретических работах 1106—140] неустойчивость стационарных состояний, необходимая для существования колебаний или переключений между альтернативными состояниями, была результатом специфической аллостерической регуляции активности ферментов и генов. В еще более яркой форме системный эффект проявляется тогда, когда полиферментная система вовсе не имеет аллостерических регуляторных механизмов и все взаимодействия между ее реакциями чисто стехиометрические. Как показали исследования [145—149] моделей механизмов стехиометрической регуляции энергетического метаболизма, на стехиометрическом уровне взаимоотношений между реакциями могут создаваться практически все важнейшие свойства полиферментных систем, которые ранее приписывались исключительно механизмам аллостерической регуляции. Поскольку энергетический метаболизм и в особенности его анаэробная часть — гликолиз — изучены в экспериментальном отношении значительно более подробно, чем любая другая часть клеточного обмена, в этой главе будут рассмотрены лишь математические модели энергетического метаболизма.
|
 |
Оглавление
|