Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
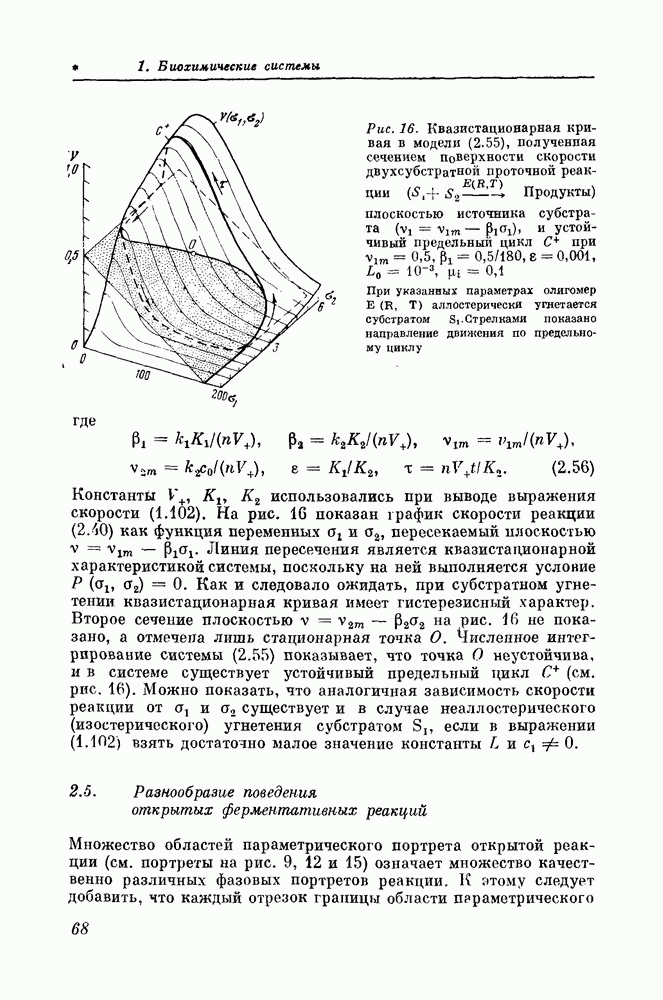
4.3. Исследование основных режимов мембраны аксона кальмара с помощью качественных методовСвойства полной модели Ходжкина — Хаксли, как правило, изучались с помощью расчетов на ЦВМ. Так как модели второго порядка хорошо описывают свойства моделей четвертого порядка, оценки свойств решений исходных уравнений можно легко получить по графикам нуль-изоклин уравнений второго порядка. Ниже будет продемонстрировано, как самые различные электрофизиологические явления могут быть объяснены не только качественно, но и количественно в терминах фазовых портретов редуцированных моделей. Спонтанная активность. Известно, что в клетках, не обладающих спонтанной активностью, последняя может быть вызвана, например, увеличением натриевой или уменьшением калиевой
Рис. 31. Нуль-изоклины
Рис. 32. Нуль-изоклины модели
Рис. 33. Нуль-изоклины системы проводимости Анализ основан на том, что спонтанная активность в модели четвертого порядка Изоклины системы Обратим внимание, что, как следует из формулы (4.6), на форму изоклины Устойчивая деполяризация мембраны. Увеличение натриевого тока (например, за счет сдвига Исследование устойчивости деполяризации в модели Ходжкина — Хаксли, в принципе, ничем не отличается от исследования спонтанной активности. Здесь необходимо определить, при каких изменениях параметров модели мембраны Повторные ответы. Известно [21, 22], что в гигантском аксоне кальмара и в других мембранах автоколебания могут возникать под действием внешнего тока; в этом случае их называют повторными ответами. А возможны ли повторные ответы в модели Ходжкина — Хаксли? В каком диапазоне токов? Каков их механизм? Рис. 34. (см. скан) Фазовые портреты системы
Рис. 35. Нуль-изоклины системы Пунктиром показано положение нуль-изоклин Ответы на эти вопросы можно легко получить с помощью семейства нуль-изоклин модели правую и вновь становится устойчивой. Повторные ответы прекращаются, возникает устойчивая деполяризация под током (рис. 35). Интересно, что на некоторых биологических мембранах (перехват Ранье, модифицированный аконитином [36], препараты папиллярной мышцы сердца [37] и др.) повторные ответы не удается вызвать ни Скачкообразная деполяризация. При плавном повышении концентрации Триггерные свойства мембраны. Аксон кальмара при некоторых экспериментальных условиях, например при внутриклеточной перфузии Аккомодация. Этим термином называют свойство мембраны увеличивать порог возбуждения при длительном действии тока. Выделяют три типа мембран [33]: абсолютно аккомодирующие (при стимуляции линейно нарастающим током
Рис. 36. Скачкообразная деполяризация в модели
Рис. 37. Триггерные свойства в модели наклоном Тип аккомодационной кривой и зависимость аккомодации от параметров в уравнениях исследуемых значений
|
 |
Оглавление
|









































































































































































































































































































 модели
модели  Стационарная точка лежит на левой ветви
Стационарная точка лежит на левой ветви  устойчива
устойчива

 а — исходная модель Нобла
а — исходная модель Нобла  спонтанно активна (в модели
спонтанно активна (в модели  особая точка
особая точка  лежит на средней ветви изоклины
лежит на средней ветви изоклины  и неустойчива);
и неустойчива);  модель
модель  без спонтанной активности (в модели
без спонтанной активности (в модели  имеется устойчивая особая точка
имеется устойчивая особая точка  она определяет потенциал покоя)
она определяет потенциал покоя)

 соответствующие трем различным состояниям мембраны, описываемой уравнениями
соответствующие трем различным состояниям мембраны, описываемой уравнениями  а — покой (особая точка
а — покой (особая точка  устойчива, лежит на левой ветви изоклины
устойчива, лежит на левой ветви изоклины  ); б - спонтанная активность (особая точка
); б - спонтанная активность (особая точка  неустойчива, лежит на средней ветви), пунктиром показала проекция предельного цикла системы
неустойчива, лежит на средней ветви), пунктиром показала проекция предельного цикла системы  на плоскость
на плоскость  ; в — устойчивая деполяризация (особая точка
; в — устойчивая деполяризация (особая точка  устойчива, расположена на правой ветви), о: — параметры системы соответствуют нормальной мембране;
устойчива, расположена на правой ветви), о: — параметры системы соответствуют нормальной мембране;  график функции
график функции  сдвинут соответственно на
сдвинут соответственно на  влево
влево
 Однако при этом автоколебания возникают далеко не всегда; например, значительное увеличение натриевой проводимоси может и не привести к появлению спонтанной активности [19]. Система
Однако при этом автоколебания возникают далеко не всегда; например, значительное увеличение натриевой проводимоси может и не привести к появлению спонтанной активности [19]. Система  позволяет легко определить, когда какой случай реализуется.
позволяет легко определить, когда какой случай реализуется.
 возникает тогда, когда в модели второго порядка
возникает тогда, когда в модели второго порядка  точка покоя теряет устойчивость (при этом появляется предельный цикл, и возникают автоколебания). Для релаксационных систем с достаточной для анализа степенью точности устойчивость может быть определена по нуль-изоклинам: особые точки, расположенные на средней ветви изоклины
точка покоя теряет устойчивость (при этом появляется предельный цикл, и возникают автоколебания). Для релаксационных систем с достаточной для анализа степенью точности устойчивость может быть определена по нуль-изоклинам: особые точки, расположенные на средней ветви изоклины  неустойчивы; расположенные на левой или правой ветви — устойчивы.
неустойчивы; расположенные на левой или правой ветви — устойчивы.
 при параметрах, соответствующих модели аксона, показаны на рис. 33, а. Особая точка устойчива (расположена на левой ветви), и мембрана не обладает спонтанной активностью. При изменении параметров изоклины деформируются. Если при этом особая точка станет неустойчивой (сместится с левой ветви изоклины
при параметрах, соответствующих модели аксона, показаны на рис. 33, а. Особая точка устойчива (расположена на левой ветви), и мембрана не обладает спонтанной активностью. При изменении параметров изоклины деформируются. Если при этом особая точка станет неустойчивой (сместится с левой ветви изоклины  на среднюю и не появится новых устойчивых особых точек), то возникает спонтанная активность (рис. 33, б).
на среднюю и не появится новых устойчивых особых точек), то возникает спонтанная активность (рис. 33, б).
 влияет только зависимость
влияет только зависимость  остальные параметры влияют на форму изоклины
остальные параметры влияют на форму изоклины  На рис. 33, б приведен пример возникновения спонтанной активности при сдвиге графика
На рис. 33, б приведен пример возникновения спонтанной активности при сдвиге графика  как целого (такие сдвиги возникают, например, при уменьшении концентрации ионов
как целого (такие сдвиги возникают, например, при уменьшении концентрации ионов  в наружном растворе). Точно так же можно проследить влияние на спонтанную активность и других входящих в систему (4.4) параметров [24] (некоторые из примеров см. на рис. 34).
в наружном растворе). Точно так же можно проследить влияние на спонтанную активность и других входящих в систему (4.4) параметров [24] (некоторые из примеров см. на рис. 34).
 на
на  влево) приводит к появлению спонтанной активности. Однако если натриевый ток увеличить еще значительней, то спонтанная активность исчезает, и возникает бесконечно длящийся потенциал действия с затухающими колебаниями на плато. Этот эффект называется устойчивой деполяризацией мембраны.
влево) приводит к появлению спонтанной активности. Однако если натриевый ток увеличить еще значительней, то спонтанная активность исчезает, и возникает бесконечно длящийся потенциал действия с затухающими колебаниями на плато. Этот эффект называется устойчивой деполяризацией мембраны.
 в модели
в модели  возникает устойчивая особая точка (фокус или узел) на правой ветви (рис.
возникает устойчивая особая точка (фокус или узел) на правой ветви (рис.  При этом удается не только качественно проследить появление устойчивой деполяризации, но и оценить значения параметров, при которых она возникает.
При этом удается не только качественно проследить появление устойчивой деполяризации, но и оценить значения параметров, при которых она возникает.
 при изменении параметров а — натриевой активации
при изменении параметров а — натриевой активации  калиевой активации
калиевой активации  в — натриевой инактивации
в — натриевой инактивации 

 при изменении внешнего тока
при изменении внешнего тока  (цифры на кривых)
(цифры на кривых)
 при токах, вызывающих повторные ответы
при токах, вызывающих повторные ответы
 Внешний ток
Внешний ток  входит только в выражение для изоклины
входит только в выражение для изоклины  Поэтому при увеличении внешнего тока изменяется лишь она (рис. 35). Повторные ответы возникают, когда особая точка, смещаясь с левой ветви изоклины
Поэтому при увеличении внешнего тока изменяется лишь она (рис. 35). Повторные ответы возникают, когда особая точка, смещаясь с левой ветви изоклины  на среднюю, становится неустойчивой. При дальнейшем увеличении тока особая точка смещается со средней ветви на
на среднюю, становится неустойчивой. При дальнейшем увеличении тока особая точка смещается со средней ветви на
 каких внешних токах. Фазовый портрет модели второго порядка показывает, что возможны две причины отсутствия повторных ответов: 1. При увеличении тока особая точка все время остается на левой ветви изоклины
каких внешних токах. Фазовый портрет модели второго порядка показывает, что возможны две причины отсутствия повторных ответов: 1. При увеличении тока особая точка все время остается на левой ветви изоклины  Однако вряд
Однако вряд  этот случай имеет место в мембранах: обычно можно подать столь большой внешний ток, что особая точка покидает левую ветвь (об этом можно судить по появлению скачка потенциала). 2. Хотя при увеличении тока особая точка и покидает левую ветвь, но автоколебания не возникают, так как раньше успела возникнуть еще одна особая точка на правой ветви (устойчивая). Этот случай обычно и реализуется в мембранах.
этот случай имеет место в мембранах: обычно можно подать столь большой внешний ток, что особая точка покидает левую ветвь (об этом можно судить по появлению скачка потенциала). 2. Хотя при увеличении тока особая точка и покидает левую ветвь, но автоколебания не возникают, так как раньше успела возникнуть еще одна особая точка на правой ветви (устойчивая). Этот случай обычно и реализуется в мембранах.
 во внешней среде потенциал покоя аксона кальмара и других возбудимых мембран может изменяться скачком (феномен Тасаки [30]). Пока ставились лишь численные эксперименты на ЦВМ, считалось, что классической теорией Ходжкина — Хаксли скачкообразную деполяризацию объяснить нельзя [31]. Однако фазовый портрет редуцированной системы позволяет легко подобрать параметры, при которых этот эффект воспроизводится в полных уравнениях Ходжкина — Хаксли (рис. 36, а): необходимо, чтобы при изменении калиевого равновесного потенциала
во внешней среде потенциал покоя аксона кальмара и других возбудимых мембран может изменяться скачком (феномен Тасаки [30]). Пока ставились лишь численные эксперименты на ЦВМ, считалось, что классической теорией Ходжкина — Хаксли скачкообразную деполяризацию объяснить нельзя [31]. Однако фазовый портрет редуцированной системы позволяет легко подобрать параметры, при которых этот эффект воспроизводится в полных уравнениях Ходжкина — Хаксли (рис. 36, а): необходимо, чтобы при изменении калиевого равновесного потенциала  особая точка, определяющая потенциал покоя, скачком переходила с левой ветви на правую ветвь изоклины
особая точка, определяющая потенциал покоя, скачком переходила с левой ветви на правую ветвь изоклины  минуя неустойчивую ветвь.
минуя неустойчивую ветвь.
 [32], может вести себя как триггер с двумя устойчивыми состояниями: раздражение током, направленным попеременно наружу и внутрь, приводит к перебросу из одного устойчивого состояния в другое. Это настолько отличается от поведения нормальной мембраны, что Тасаки [31] был вынужден искать объяснение вне рамок теории Ходжкина — Хаксли. Однако и здесь с помощью нуль-изоклин модели
[32], может вести себя как триггер с двумя устойчивыми состояниями: раздражение током, направленным попеременно наружу и внутрь, приводит к перебросу из одного устойчивого состояния в другое. Это настолько отличается от поведения нормальной мембраны, что Тасаки [31] был вынужден искать объяснение вне рамок теории Ходжкина — Хаксли. Однако и здесь с помощью нуль-изоклин модели  легко найти параметры, при которых этот эффект воспроизводится в модели Ходжкина — Хаксли (рис. 37).
легко найти параметры, при которых этот эффект воспроизводится в модели Ходжкина — Хаксли (рис. 37).
 с малым
с малым

 при увеличении внеклеточной концентрации
при увеличении внеклеточной концентрации  а — зависимость потенциала покоя в модели
а — зависимость потенциала покоя в модели  от
от  исходной модели
исходной модели  сдвинуты на
сдвинуты на  соответственно);
соответственно);  нуль-изоклнны системы
нуль-изоклнны системы  при тех же значениях параметров. Стрелками показана траектория особой точки при из менении
при тех же значениях параметров. Стрелками показана траектория особой точки при из менении  определяющая потенциал покоя. При
определяющая потенциал покоя. При  мембрана находится в покоящемся состоянии, при
мембрана находится в покоящемся состоянии, при  стационарная точка на левой ветви исчезает, и мембрана скачком переходит в устойчиво деполяризованное состояние. Цифры на кривых
стационарная точка на левой ветви исчезает, и мембрана скачком переходит в устойчиво деполяризованное состояние. Цифры на кривых 

 Потенциалы мембраны в модели
Потенциалы мембраны в модели  сдвинуты на
сдвинуты на  соответственно) (а); нуль-изоклины системы
соответственно) (а); нуль-изоклины системы  при тех же значениях параметров
при тех же значениях параметров  Имеются два устойчивых значения потенциала покоя мембраны:
Имеются два устойчивых значения потенциала покоя мембраны:  (особая точка
(особая точка  (особая точка
(особая точка  Из покоящегося состояния в устойчиво деполяризованное и обратно мембрана переводится импульсами тока противоположной полярности (показано стрелками)
Из покоящегося состояния в устойчиво деполяризованное и обратно мембрана переводится импульсами тока противоположной полярности (показано стрелками)
 ответ не возникает;
ответ не возникает;  называют минимальным градиентом); частично аккомодирующие (порог срабатывания растет лишь в некотором диапазоне изменений а, а затем постоянен); неаккомодирующие (порог срабатывания постоянен и не зависит от а).
называют минимальным градиентом); частично аккомодирующие (порог срабатывания растет лишь в некотором диапазоне изменений а, а затем постоянен); неаккомодирующие (порог срабатывания постоянен и не зависит от а).
 просто определяется по семейству нуль-изоклин уравнений
просто определяется по семейству нуль-изоклин уравнений  построенному при различных токах
построенному при различных токах  [24]. Если особая точка системы для всех
[24]. Если особая точка системы для всех
 остается на левой ветви изоклины
остается на левой ветви изоклины  то мембрана абсолютно аккомодирующая. Если при увеличении
то мембрана абсолютно аккомодирующая. Если при увеличении  особая точка покидает левую ветвь, то тип аккомодационной кривой зависит от соотношения между током реобазы
особая точка покидает левую ветвь, то тип аккомодационной кривой зависит от соотношения между током реобазы  и током возникновения повторных ответов
и током возникновения повторных ответов  При
При  наблюдается частичная аккомодация, при
наблюдается частичная аккомодация, при  аккомодация отсутствует.
аккомодация отсутствует.