2.7. Эффект депонирования
При математическом моделировании механизма одночастотных гликолитических колебаний [87—100] было замечено, что участие быстрой обратимой реакции в биохимическом автогенераторе увеличивает период автоколебаний примерно в  раз (К — константа равновесия обратимой реакции) [72, 83, 84]. Это явление имеет важное значение для теории клеточных часов — еще не идентифицированной автоколебательной биохимической системы, создающей колебания с периодом
раз (К — константа равновесия обратимой реакции) [72, 83, 84]. Это явление имеет важное значение для теории клеточных часов — еще не идентифицированной автоколебательной биохимической системы, создающей колебания с периодом  [84, 101—104].
[84, 101—104].
Эффект депонирования, или замедление колебаний, вызываемое быстрой обратимой реакцией, имеет простое объяснение.
Пусть в биохимической системе имеется вещество  участвующее в различных обменных реакциях. Внесем в эту систему реакцию быстрого обратимого депонирования вещества в его резервную, метаболически неактивную форму
участвующее в различных обменных реакциях. Внесем в эту систему реакцию быстрого обратимого депонирования вещества в его резервную, метаболически неактивную форму 

Здесь  константы скорости. Теперь суммарная концентрация вещества
константы скорости. Теперь суммарная концентрация вещества  стала равной
стала равной  Если реакция (2.67) быстрая, то в ней за короткое время устанавливается равновесие, при котором
Если реакция (2.67) быстрая, то в ней за короткое время устанавливается равновесие, при котором

С участием условия (2.68) суммарная концентрация вещества  равна
равна  где
где  константа равновесия реакции (2.67). Таким образом, внесение в биохимическую систему реакции (2.67) эквивалентно увеличению в
константа равновесия реакции (2.67). Таким образом, внесение в биохимическую систему реакции (2.67) эквивалентно увеличению в  раз суммарной концентрации вещеста
раз суммарной концентрации вещеста  а это означает увеличение во столько же раз постоянной времени для изменения
а это означает увеличение во столько же раз постоянной времени для изменения  для того чтобы синтезировать или расходовать увеличенное в К 1 раз количество вещества, надо в
для того чтобы синтезировать или расходовать увеличенное в К 1 раз количество вещества, надо в  раз больше времени.
раз больше времени.
В клетках депонирование метаболитов и их превращение из резервной в метаболически активную форму осуществляется отдельными ферментами. Анализ эффекта депонирования в таких сложных случаях может быть проведен лишь с помощью математических моделей [72, 105], так как из-за нелинейности кинетики ферментативных реакций не удается в явном виде вычислить фактор замедления, т. е. величину, на которую увеличивается постоянная времени концентрации депонируемого вещества.
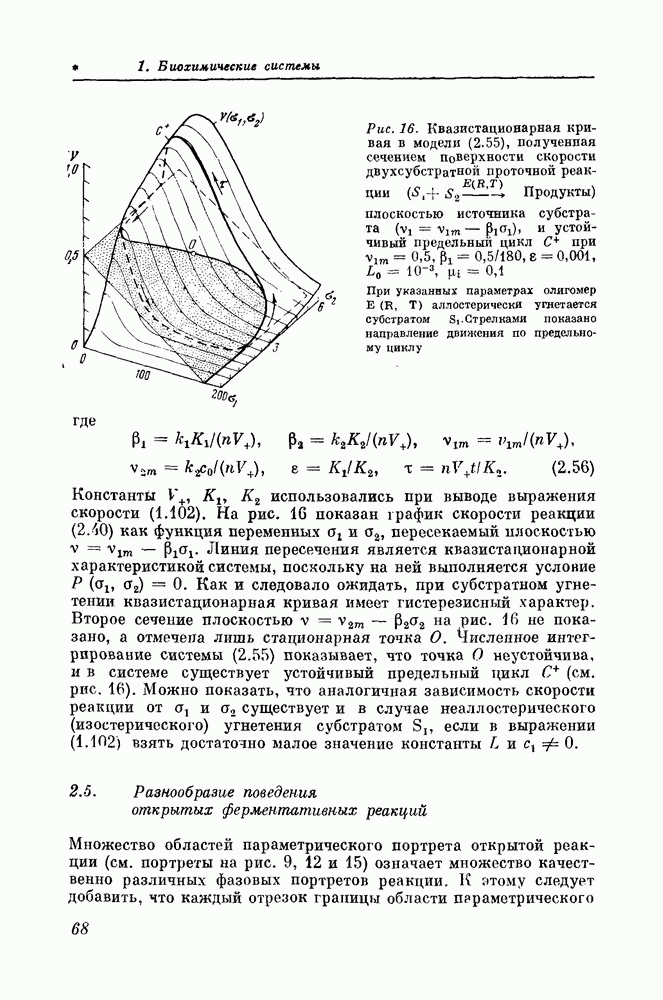
Детальное исследование моделей показывает, что ферментативные реакции в проточных условиях обладают большим разнообразием динамического поведения, которое никогда не наблюдается
в непроточных условиях обычных биохимических экспериментов с выделенными из клетки ферментами. Сравнивая модели, описывающие совершенно различные молекулярные механизмы, контролирующие активности ферментов (механизмы активации и угнетения ферментов субстратами или продуктами), нетрудно заметить тем не менее поразительную общность динамического поведения реакций в проточных условиях. Эта общность объясняется тем, что все рассмотренные молекулярные механизмы регуляции активности ферментов, быть может совершенно не сопоставимые биохимически, создают однотипную нелинейность — гистерезис квазистационарной входной или выходной характеристики.
В главах 3 и 4 будет показано, что подобный гистерезис может возникать также и в полиферментных системах и возбудимых клеточных мембранах. И хотя молекулярные механизмы, создающие гистерезис в полиферментной системе и мембране, также совершенно различны, этот гистерезис приводит к тому же разнообразию форм динамического поведения, какое было продемонстрировано в этой главе.