Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
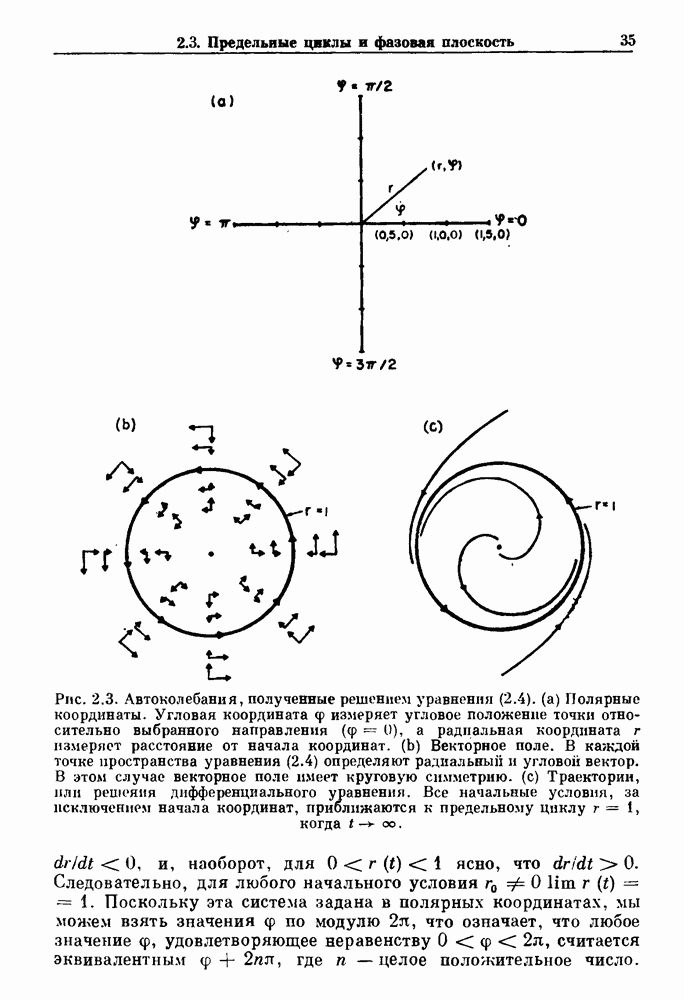
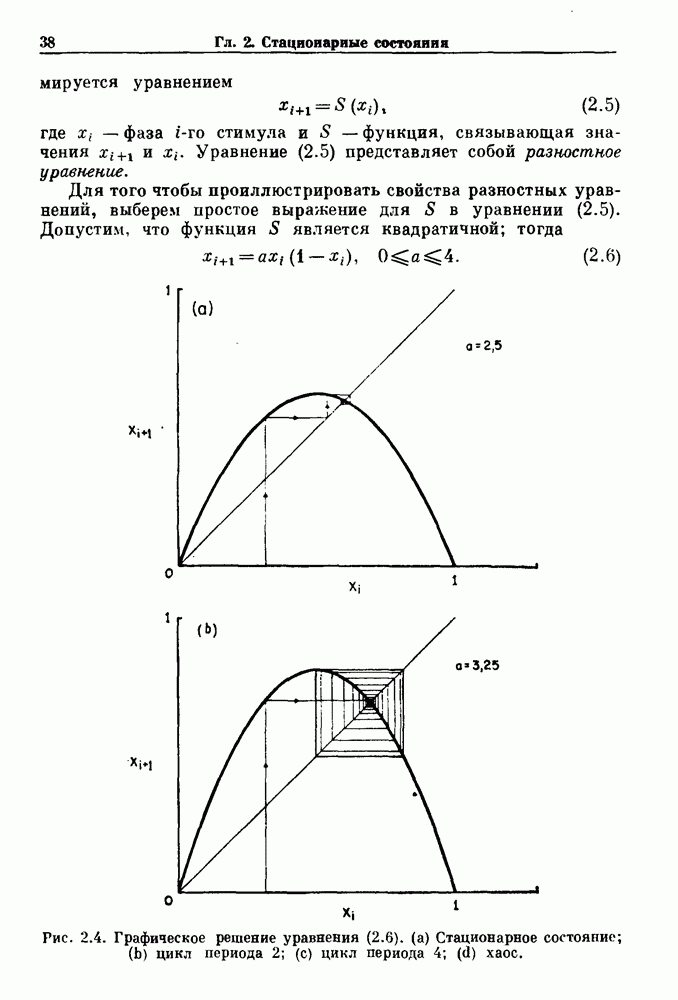
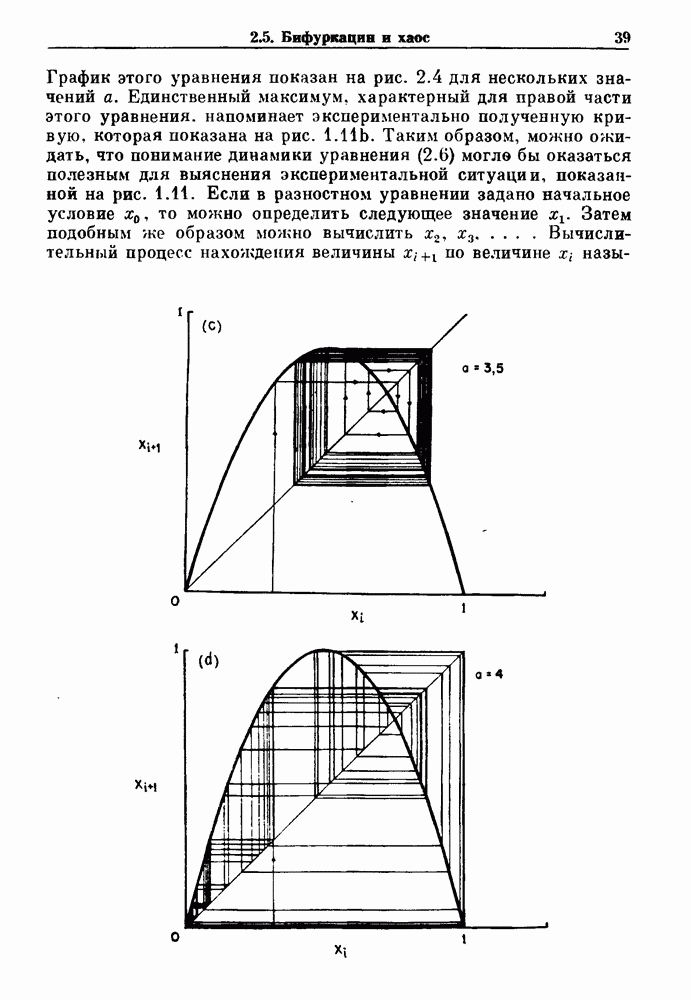
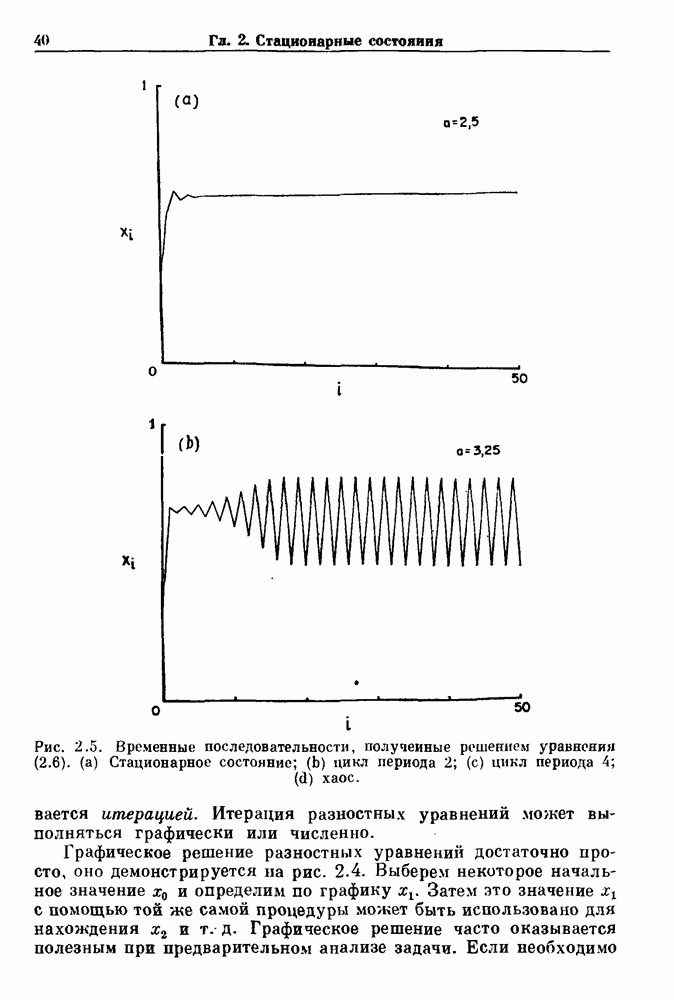
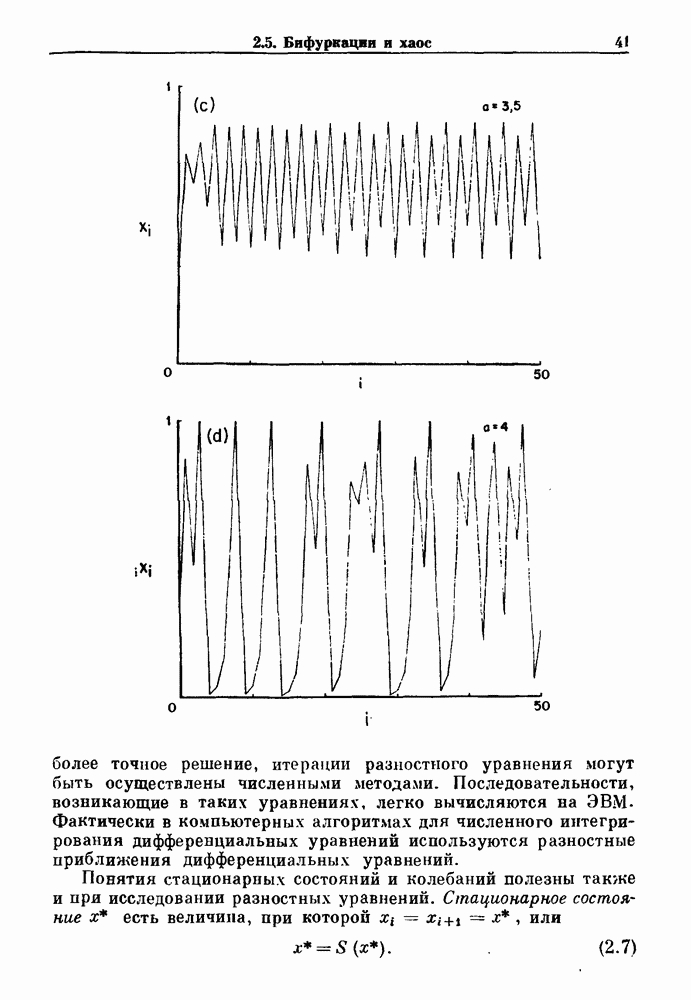
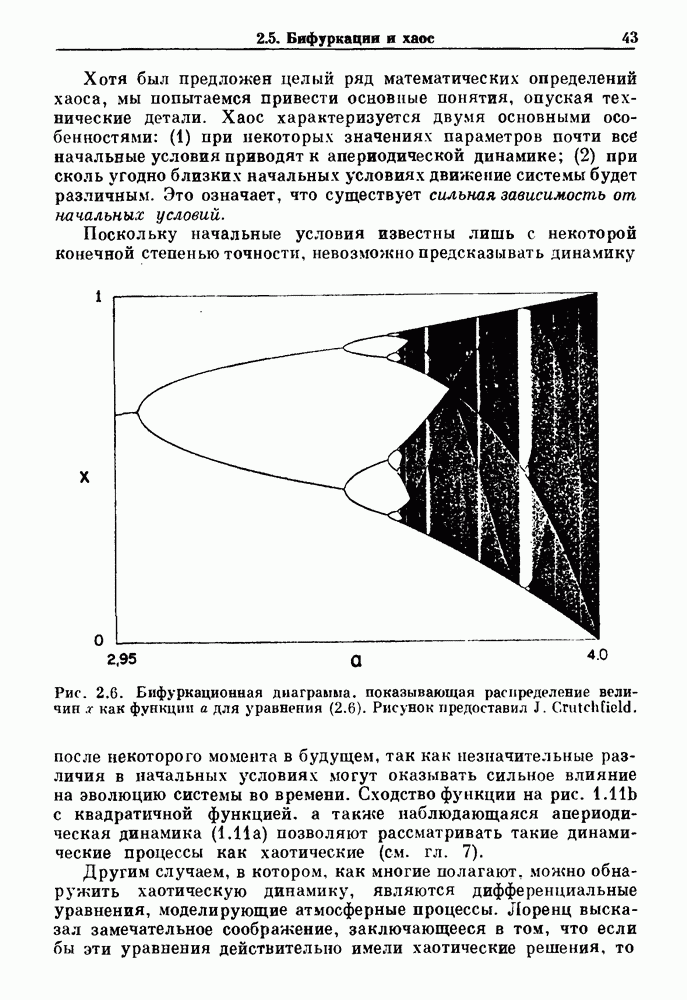
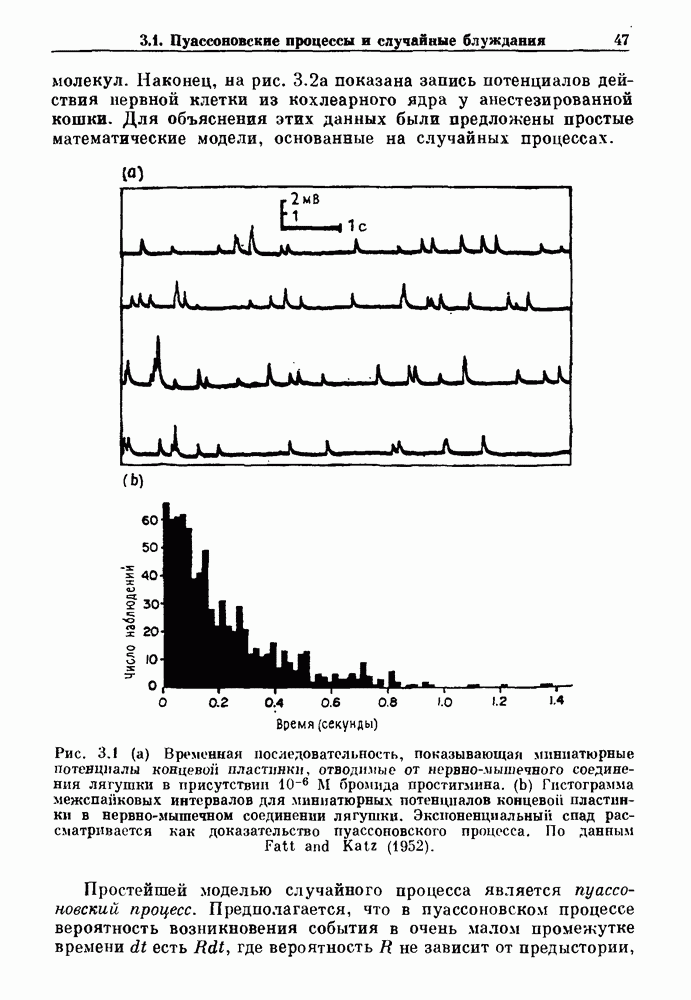
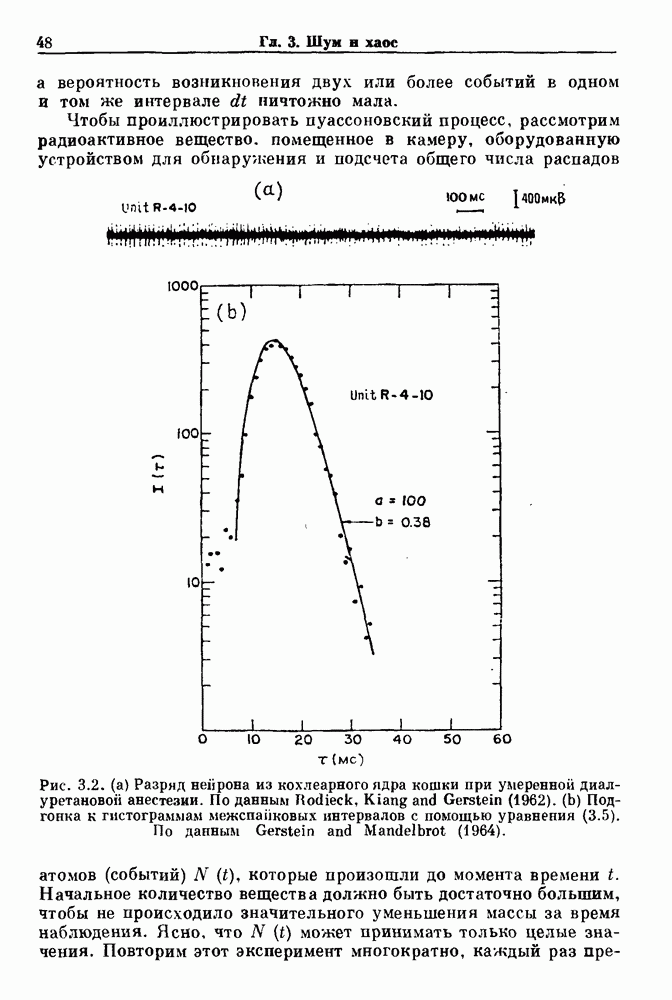
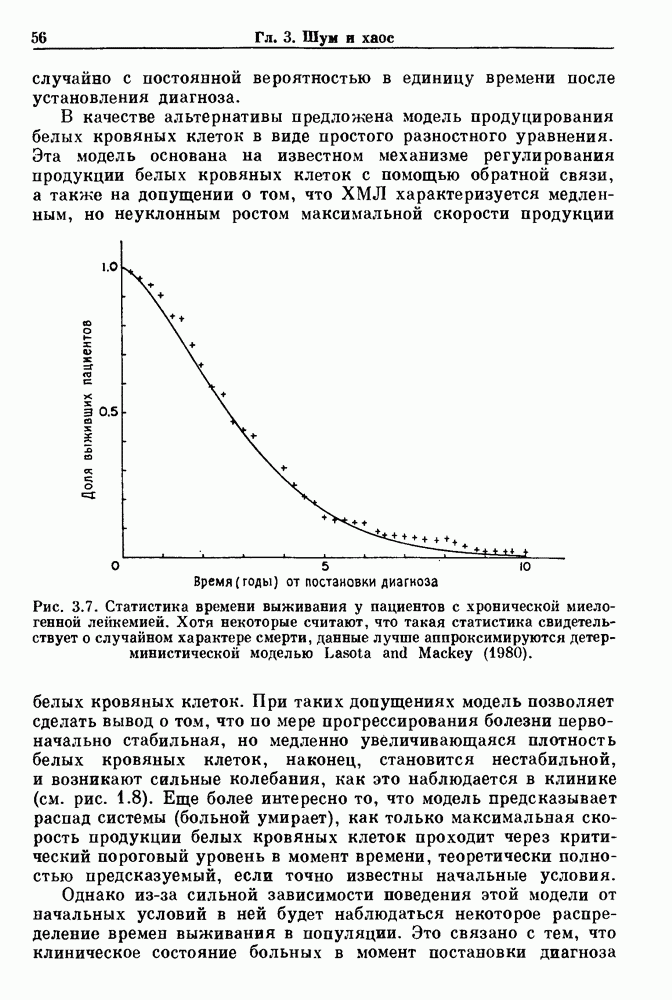
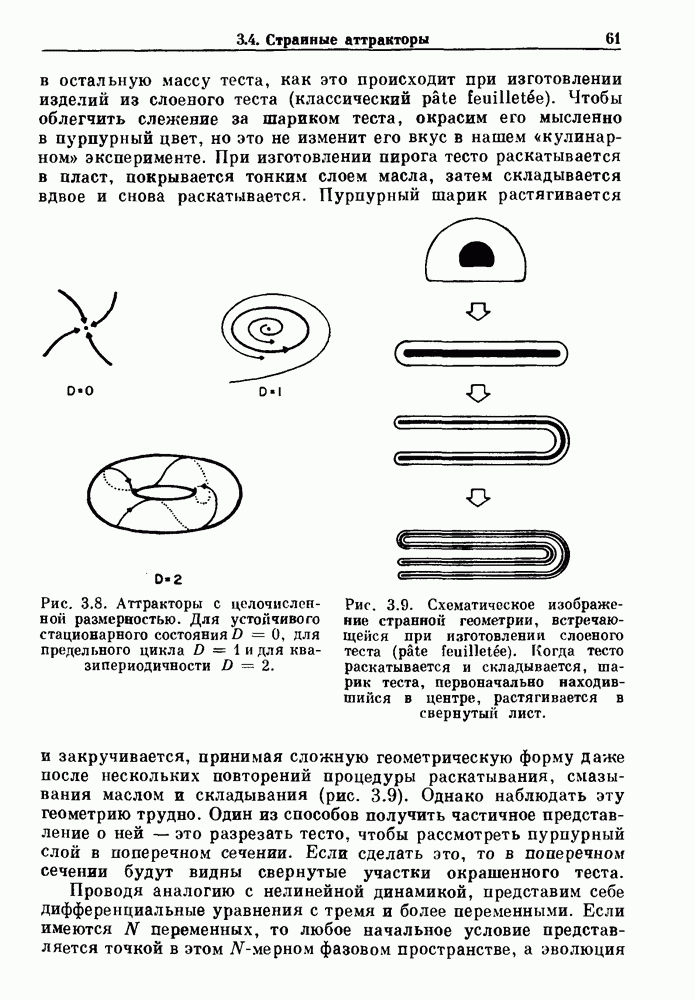
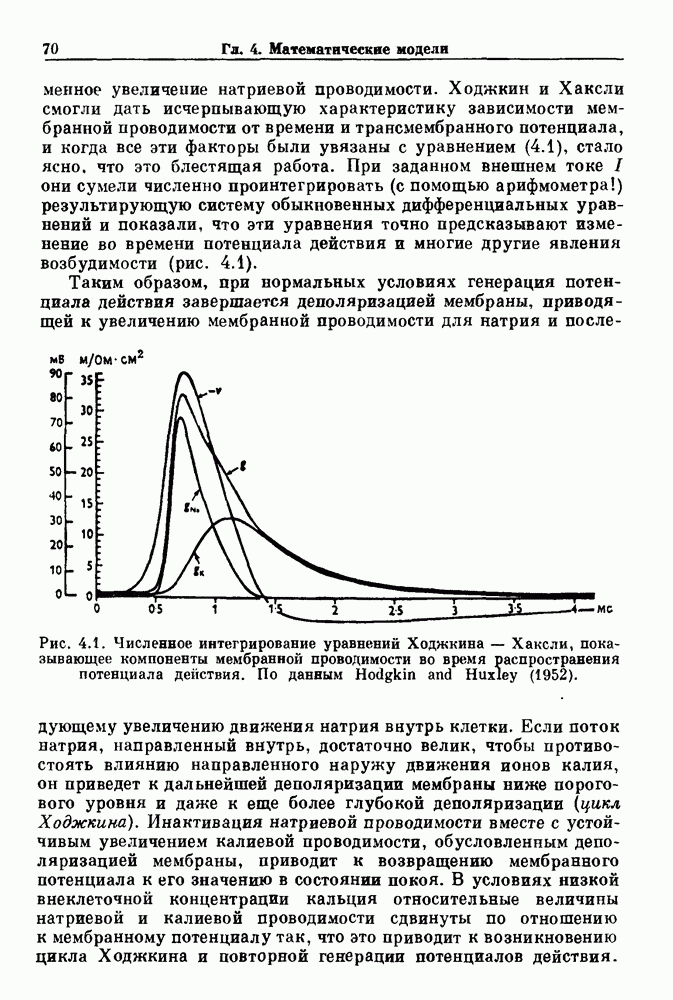
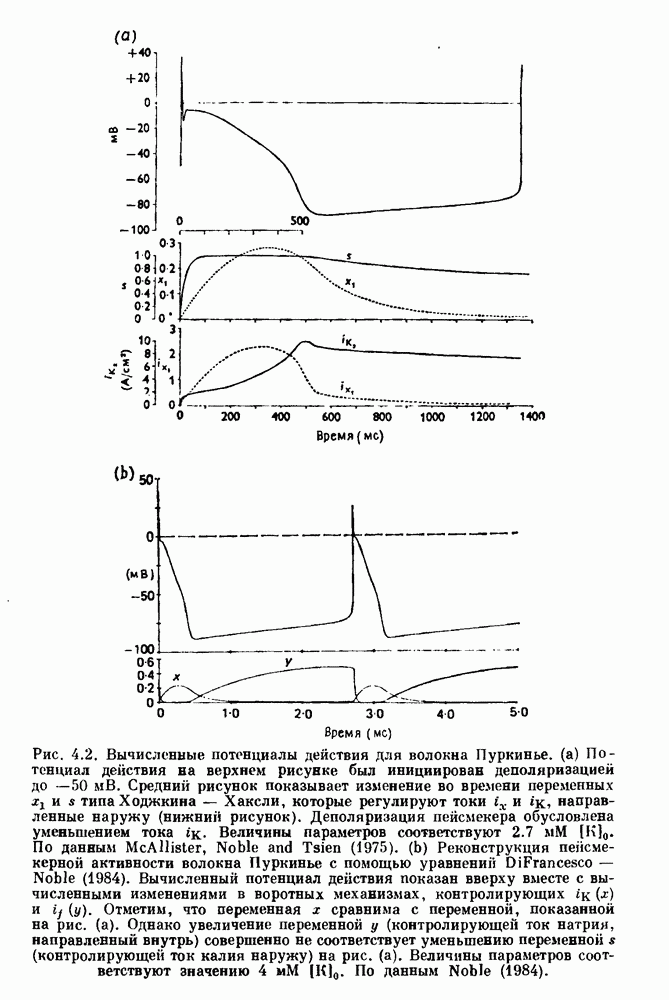
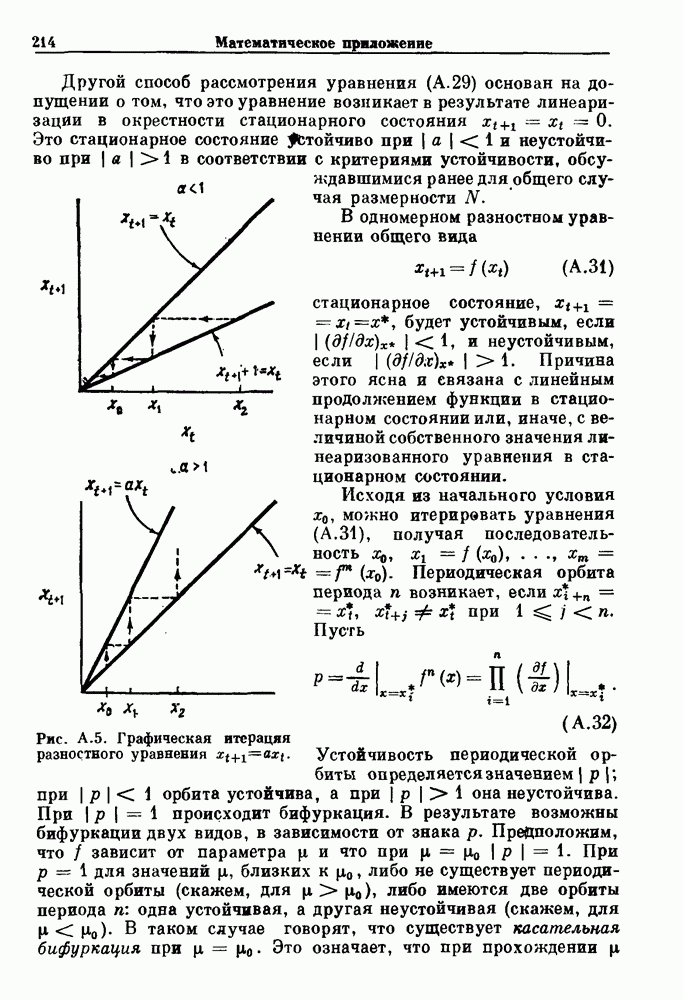
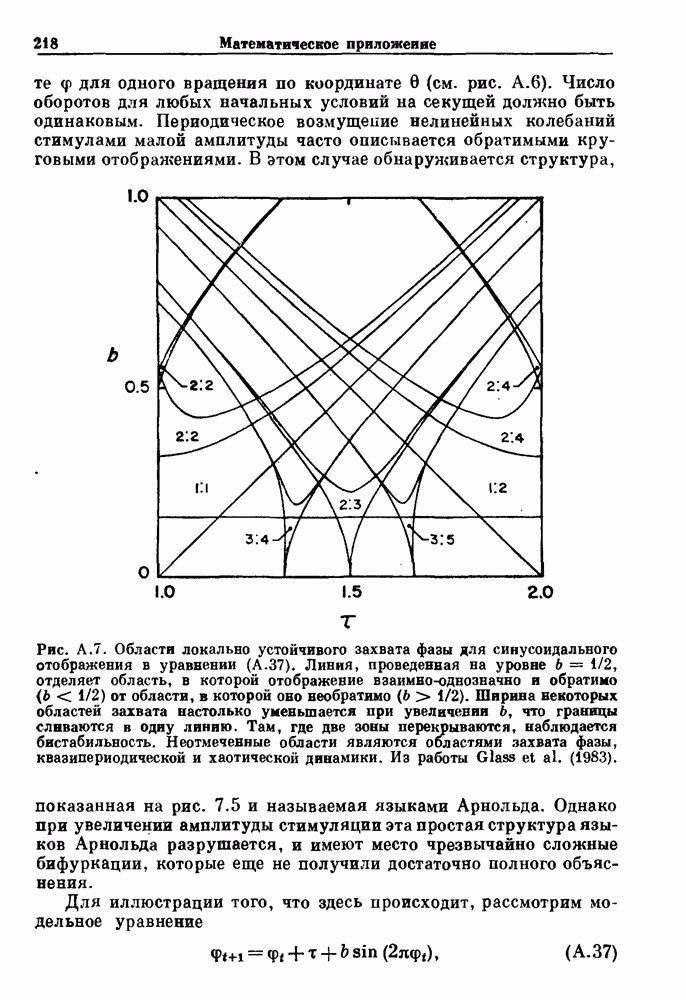
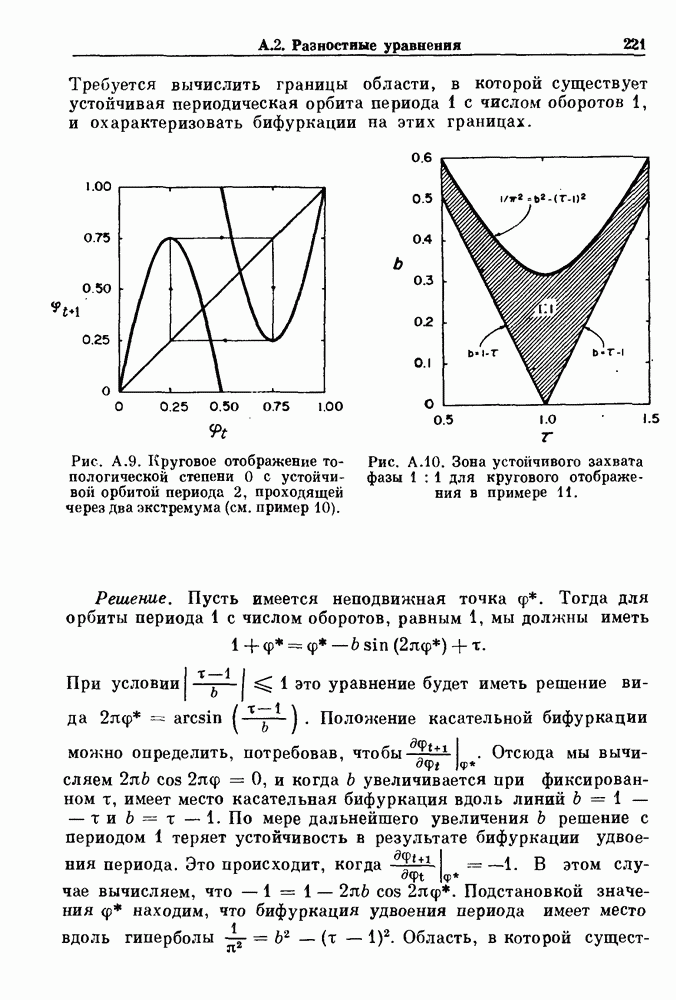
Глава 3. Шум и хаосДанные, получаемые при экспериментальном изучении физиологических систем, часто не остаются постоянными во времени, но их изменение не имеет регулярного, периодического характера. Такие нерегулярности часто связаны с шумом, или случайными стохастическими процессами, которые, в принципе, совершенно непредсказуемы, если не считать их статистических свойств. Например, шум может быть обусловлен случайными тепловыми флуктуациями или другими воздействиями окружающей среды. В отличие от шума хаос возникает в детерминированных системах. В разд. 3.1 мы обсуждаем пуассоновские процессы и случайные блуждания, которые обычно связаны с шумом. В разд. 3.2 мы рассматриваем хаотическое поведение, которое может порождаться простыми разностными уравнениями, и показываем, что иногда такое хаотическое поведение может иметь сходство с поведением, обычно приписываемым шуму. Мы приводим несколько примеров, в которых в настоящее время невозможно определить однозначно, обусловлен ли динамический процесс шумом или хаотическим поведением. В разд. 3.3 мы рассматриваем несколько методов, используемых для идентификации хаотического поведения. В последнее время находят применение две статистические меры хаоса — число Ляпунова и размерность, которые обсуждаются в разд. 3.4. 3.1. Пуассоновские процессы и случайные блужданияДля того чтобы ввести понятия шума и случайных процессов, рассмотрим три примера из нейрофизиологии, в которых динамическое поведение моделировалось случайным процессом. На рис. 1.5 показан ток, протекающий через одиночный ионный канал мембраны. Принято считать, что дискретные скачки соответствуют моментам открывания каналов в мембране нервного волокна. На рис. 3.1 а показаны процессы деполяризации мембраны в мышце лягушки (называемые миниатюрными потенциалами концевой пластинки). В каждом случае деполяризация связана с высвобождением в пресинаптическую щель небольшого количества медиатора ацетилхолина, содержащего около тысячи молекул. Наконец, на рис. 3.2 а показана запись потенциалов действия нервной клетки из кохлеарного ядра у анестезированной кошки. Для объяснения этих данных были предложены простые математические модели, основанные на случайных процессах.
Рис. 3.1 (а) Временная последовательность, показывающая миниатюрные потенциалы концевом пластинки, отводимые от нервно-мышечного соединения лягушки в присутствии Простейшей моделью случайного процесса является пуассоновский процесс. Предполагается, что в пуассоновском процессе вероятность возникновения события в очень малом промежутке времени а вероятность возникновения двух или более событий в одном и том же интервале Чтобы проиллюстрировать пуассоновский процесс, рассмотрим радиоактивное вещество, помещенное в камеру, оборудованную устройством для обнаружения и подсчета общего числа распадов атомов (событий) (см. скан) Рис. 3.2. (а) Разряд нейрона из кохлеарного ядра кошки при умеренной диал-уретановой анестезии. По данным Hodieck, Kiang and Gerstein (1962). (b) Подгонка к гистограммам межспапковых интервалов с помощью уравнения (3.5). По данным Gerstein and Mandelbrot (1964). Начальное количество вещества должно быть достаточно большим, чтобы не происходило значительного уменьшения массы за время наблюдения. Ясно, что его в тот момент, когда происходит первый распад. Если обозначить через
из которого мы немедленно получаем (см. уравнение 2.2)
где С есть некоторая произвольная постоянная. Поскольку вероятность того, что распад не произойдет в момент времени
Продолжая эти рассуждения, можно вывести другие свойства пуассоновского процесса. Вероятность
Далее, вероятность того, что интервал между некоторым событием и последующим
Уравнение (3.4) называется распределением Пуассона, а уравнение (3.5) представляет собой плотность вероятности пуассоновского процесса. Из уравнения (3.5) легко вычислить, что среднее время между событиями для пуассоновского процесса равно Часто считают, что если гистограмма интервалов между событиями представляет собой экспоненту, то имеет место пуассоновский процесс. Так, полагают, что открывание каналов (рис. 1.5) и миниатюрные потенциалы концевых пластинок (рис. 3.1) представляют собой пуассоновский процесс, поскольку экспериментальные данные хорошо аппроксимируются единственной экспонентой. Однако так как между событиями возможна корреляция, которая не обнаруживается с помощью функции распределения интервалов между событиями, то необходимо определить статистику множественных событий, но в большинстве исследований такой анализ не предпринимается. Хотя простой пуассоновский процесс является прекрасной моделью радиоактивного распада, неудивительно, что в физиологических системах статистический анализ данных часто обнаруживает значительные расхождения со статистикой пуассоновского процесса.
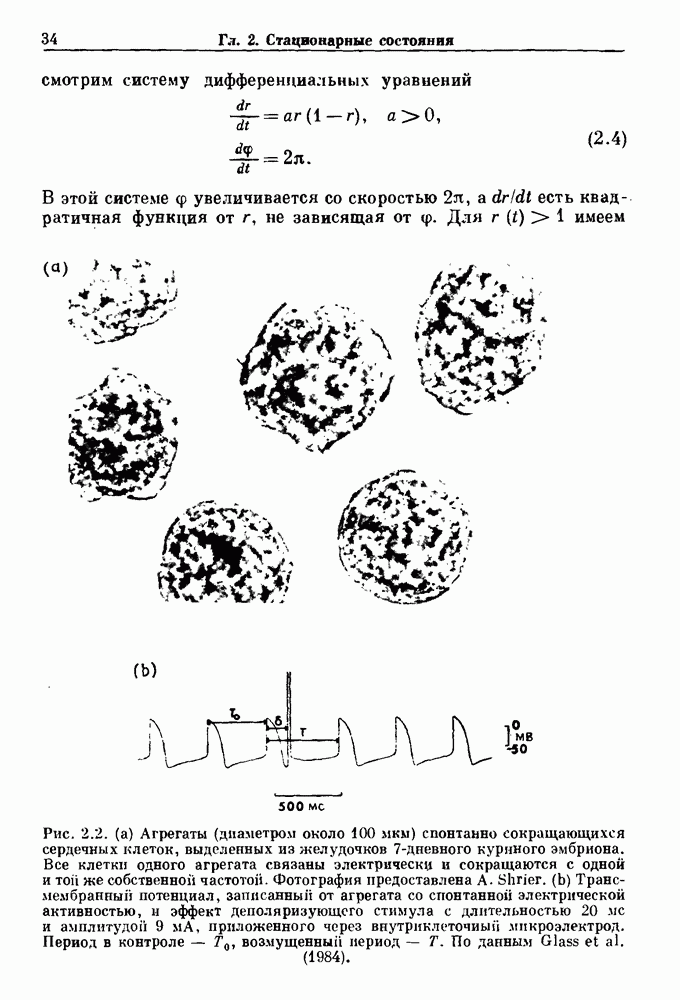
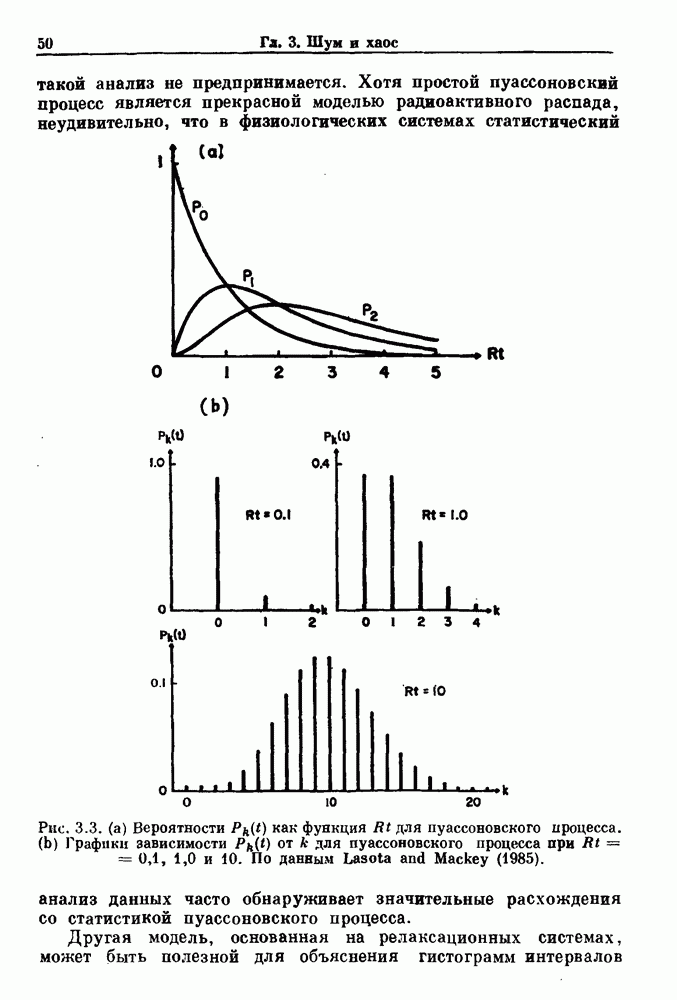
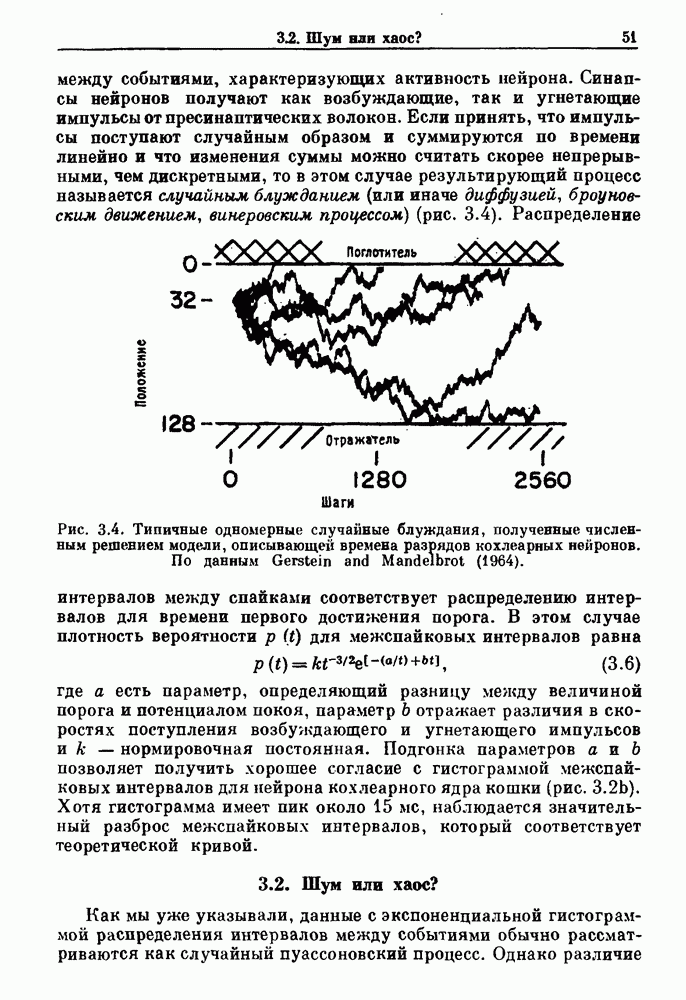
Рис. 3.3. (а) Вероятности Другая модель, основанная на релаксационных системах, может быть полезной для объяснения гистограмм интервалов между событиями, характеризующих активность нейрона. Синапсы нейронов получают как возбуждающие, так и угнетающие импульсы от пресинаптических волокон. Бели принять, что импульсы поступают случайным образом и суммируются по времени линейно и что изменения суммы можно считать скорее непрерывными, чем дискретными, то в этом случае результирующий процесс называется случайным блужданием (или иначе диффузией, броуновским движением, винеровским процессом) (рис. 3.4).
Рис. 3.4. Типичные одномерные случайные блуждания, полученные численным решением модели, описывающей времена разрядов кохлеарных нейронов. По данным Gerstein and Mandelbrot (1964). Распределение интервалов между спайками соответствует распределению интервалов для времени первого достижения порога. В этом случае плотность вероятности
где а есть параметр, определяющий разницу между величиной порога и потенциалом покоя, параметр b отражает различия в скоростях поступления возбуждающего и угнетающего импульсов и А: — нормировочная постоянная. Подгонка параметров а и b позволяет получить хорошее согласие с гистограммой межепайковых интервалов для нейрона кохлеарного ядра кошки (рис. 3.2b). Хотя гистограмма имеет пик около 15 мс, наблюдается значительный разброс межепайковых интервалов, который соответствует теоретической кривой.
|
 |
Оглавление
|















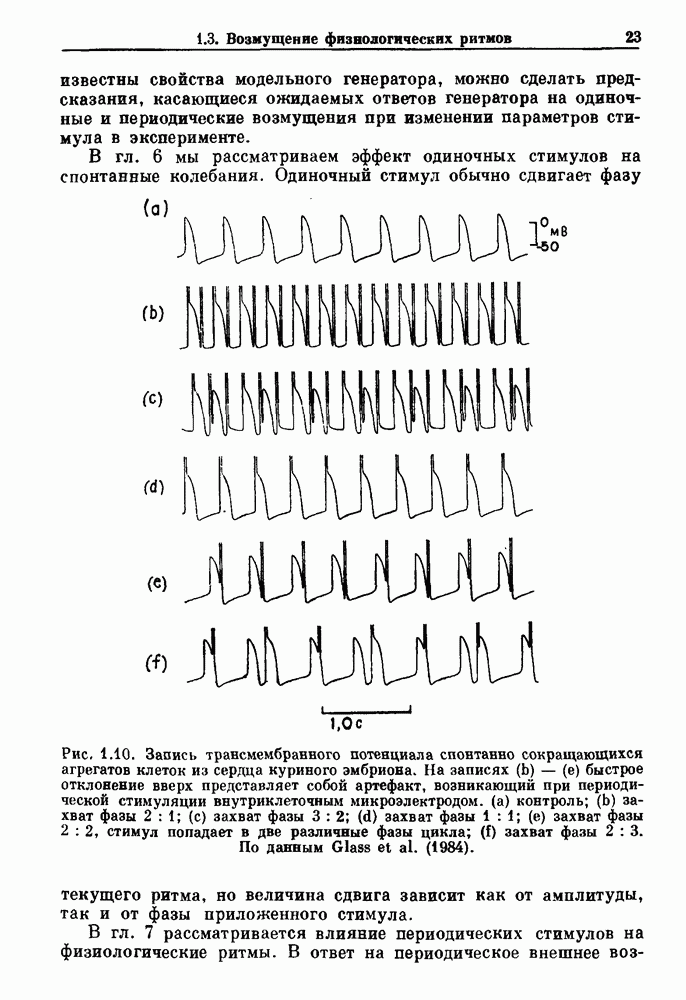
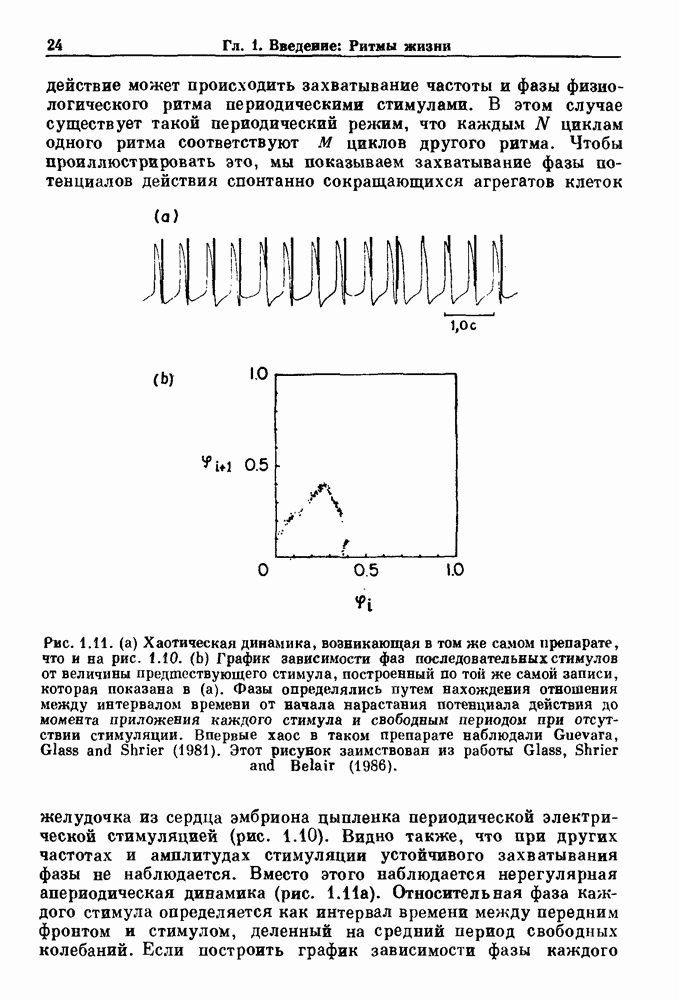






























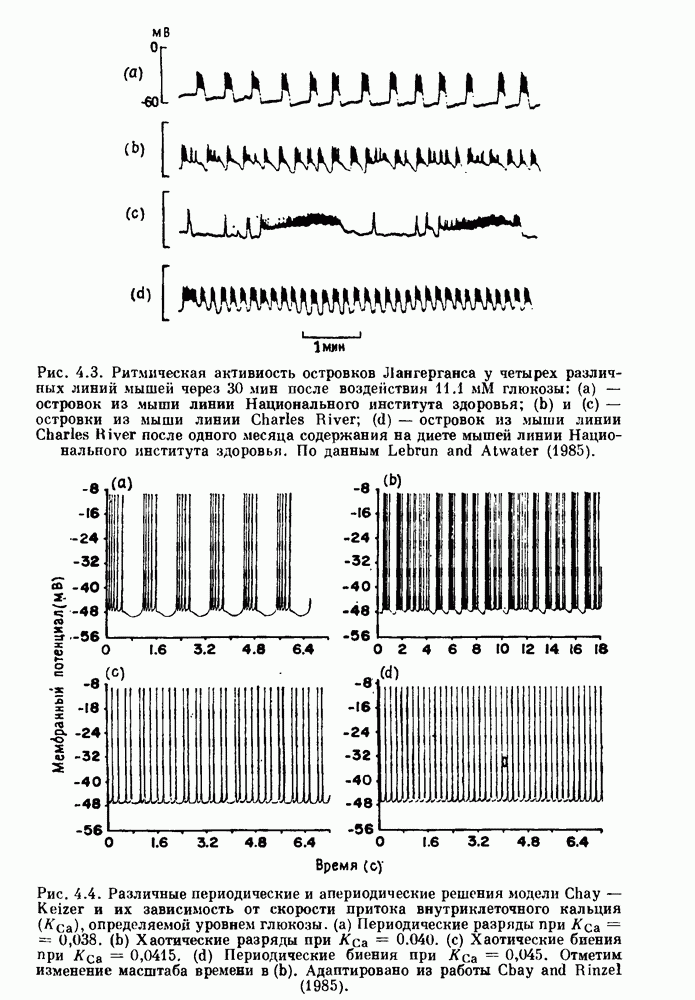








































































































































 бромида простигмина. (b) Гистограмма межепанковых интервалов для миниатюрных потенциалов концевой пластинки в нервно-мышечном соединении лягушки. Экспоненциальный спад рассматривается как доказательство пуассоновского процесса. По данным
бромида простигмина. (b) Гистограмма межепанковых интервалов для миниатюрных потенциалов концевой пластинки в нервно-мышечном соединении лягушки. Экспоненциальный спад рассматривается как доказательство пуассоновского процесса. По данным 
 есть
есть  где вероятность R не зависит от предыстории,
где вероятность R не зависит от предыстории,
 ничтожно мала.
ничтожно мала.
 , которые произошли до момента времени t.
, которые произошли до момента времени t.
 может принимать только целые значения. Повторим этот эксперимент многократно, каждый раз прерывая
может принимать только целые значения. Повторим этот эксперимент многократно, каждый раз прерывая
 вероятность того, что в данном эксперименте распад не произойдет до момента времени
вероятность того, что в данном эксперименте распад не произойдет до момента времени  и если принять, что отдельные распады не зависят друг от друга, то вероятность
и если принять, что отдельные распады не зависят друг от друга, то вероятность  удовлетворяет дифференциальному уравнению
удовлетворяет дифференциальному уравнению

 (3.2)
(3.2)
 , равна 1, постоянная С должна быть равна 1. Таким образом,
, равна 1, постоянная С должна быть равна 1. Таким образом,

 того, что на интервале времени t произойдет точно к событий, определяется выражением
того, что на интервале времени t произойдет точно к событий, определяется выражением

 событием окажется между t и
событием окажется между t и  есть
есть  где
где

 , а дисперсия равна
, а дисперсия равна  На рис. 3.3 а показана зависимость
На рис. 3.3 а показана зависимость  от t для k = 0, 1 и 2, а на рис. 3.3b показана зависимость
от t для k = 0, 1 и 2, а на рис. 3.3b показана зависимость  от к для
от к для  .
.

 как функция
как функция  для пуассоновского процесса. (b) Графики зависимости
для пуассоновского процесса. (b) Графики зависимости  от к для пуассоновского процесса при
от к для пуассоновского процесса при  . По данным Lasota and Mackey (1985).
. По данным Lasota and Mackey (1985).

 для межепайковых интервалов равна
для межепайковых интервалов равна
