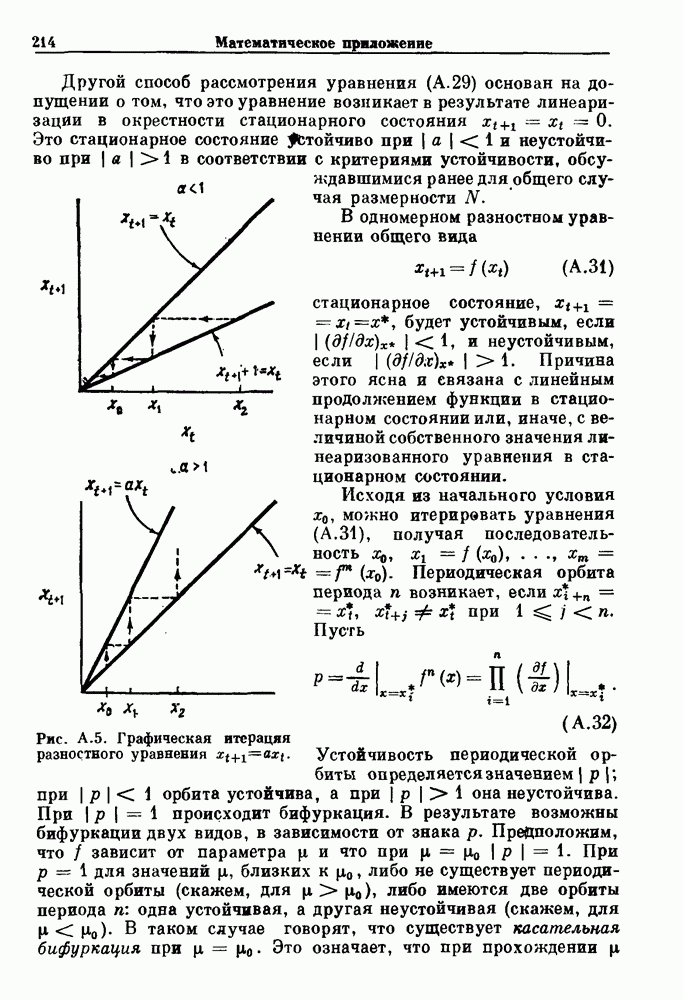
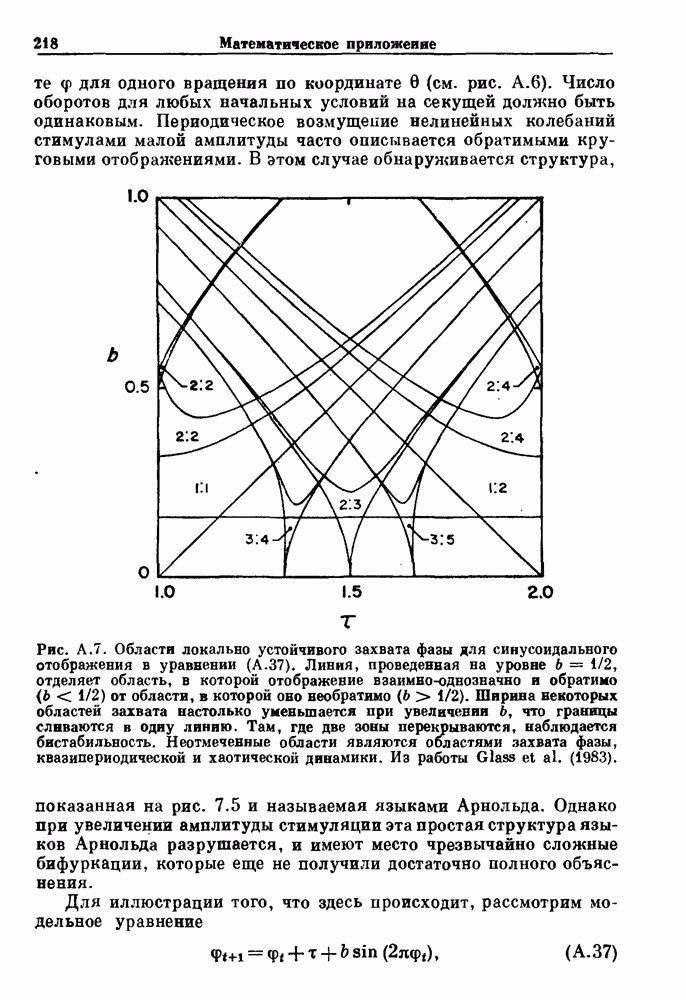
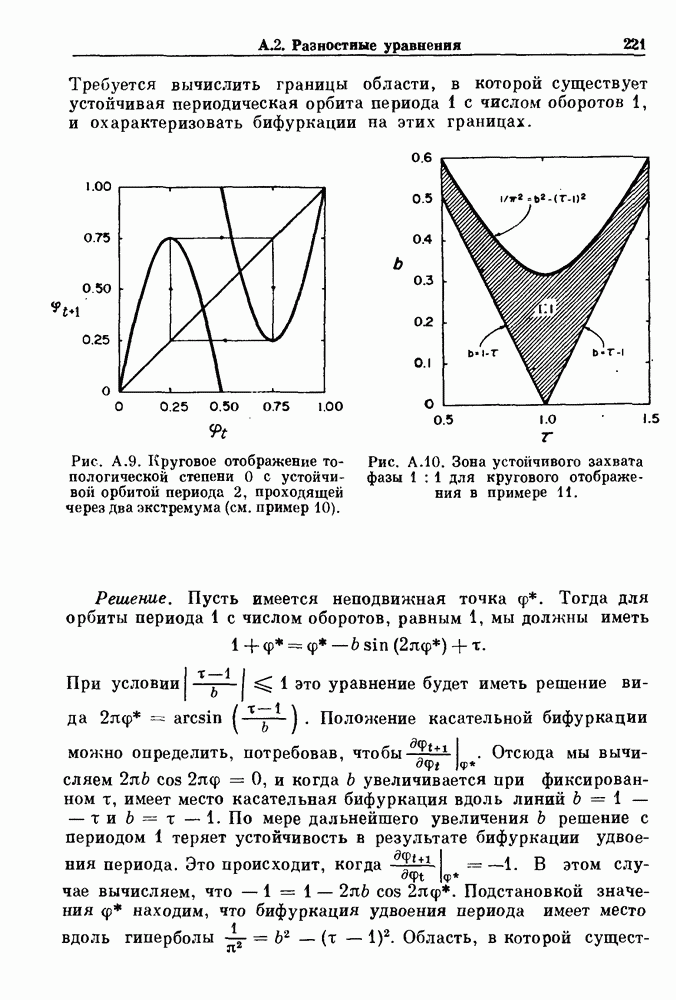
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
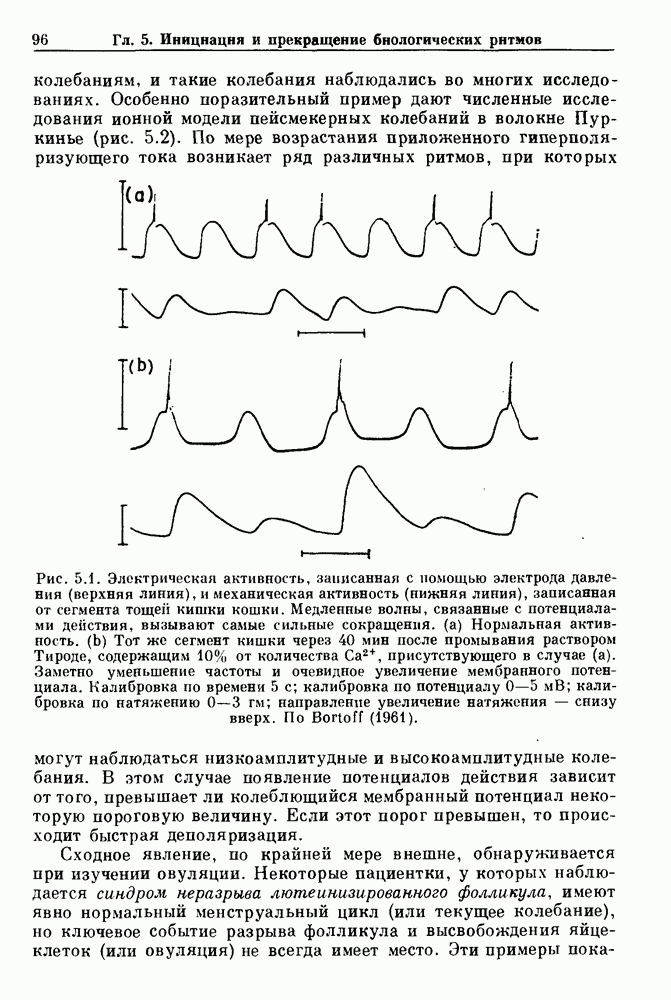
5.2. Мягкое возбуждениеДля математика естественным способом включения и выключения колебаний является использование зависящей от параметров динамической системы. При некоторых значениях параметров колебания существуют, тогда как при других значениях их нет. Простейший случай изображен схематически на рис. 5.3. При значениях параметра
Рис. 5.3. Мягкое возбуждение (закритическая бифуркация Хопфа). (а) Представление в фазовой плоскости решения уравнения (5.1). При увеличении с возникают устойчивые автоколебания низкой амплитуды и конечной частоты. (b) Бифуркационная диаграмма мягкого возбуждения. Сплошная кривая представляет устойчивые решения, пунктирная кривая — неустойчивые решения. Однако при
Используя качественный анализ, кратко описанный в гл. 2, можно легко заметить, что при с
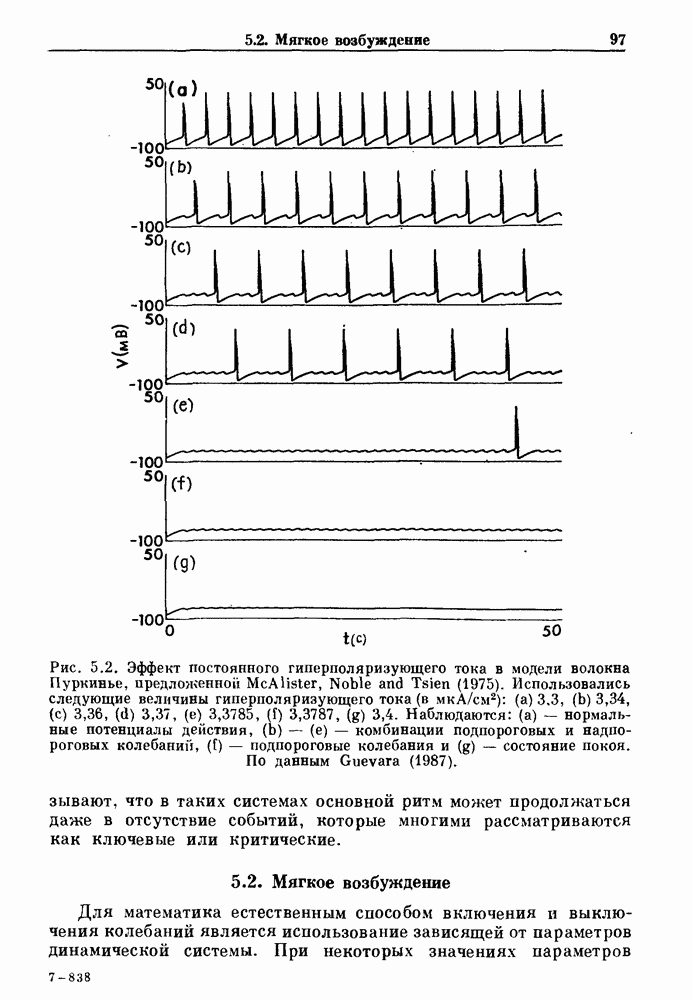
Как уже обсуждалось в гл. 4, неустойчивость в системах с отрицательной и комбинированной обратной связью может возникать тогда, когда временные задержки и/или коэффициенты связи превышают некоторые критические значения. В точках неустойчивости наблюдаются закритические бифуркации Хопфа. Считают, что такая неустойчивость лежит в основе колебаний, наблюдающихся при дыхании Чейна—Стокса, некоторых типах мышечного тремора, многих колебательных гематологических нарушениях и колебательном зрачковом рефлексе, вызываемом пятном света, попадающим на край зрачка.
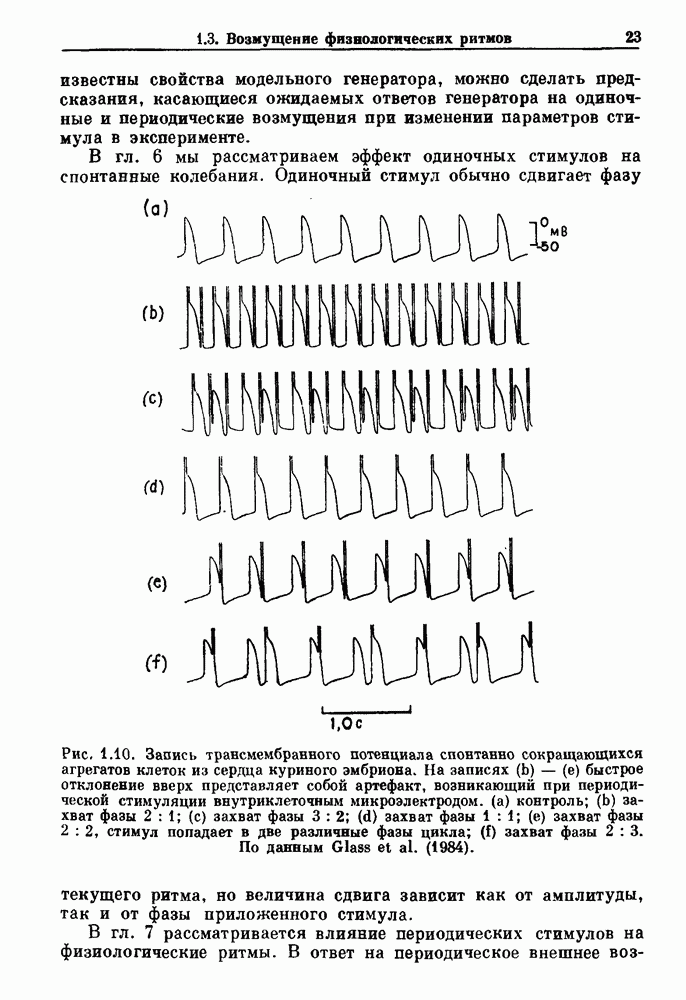
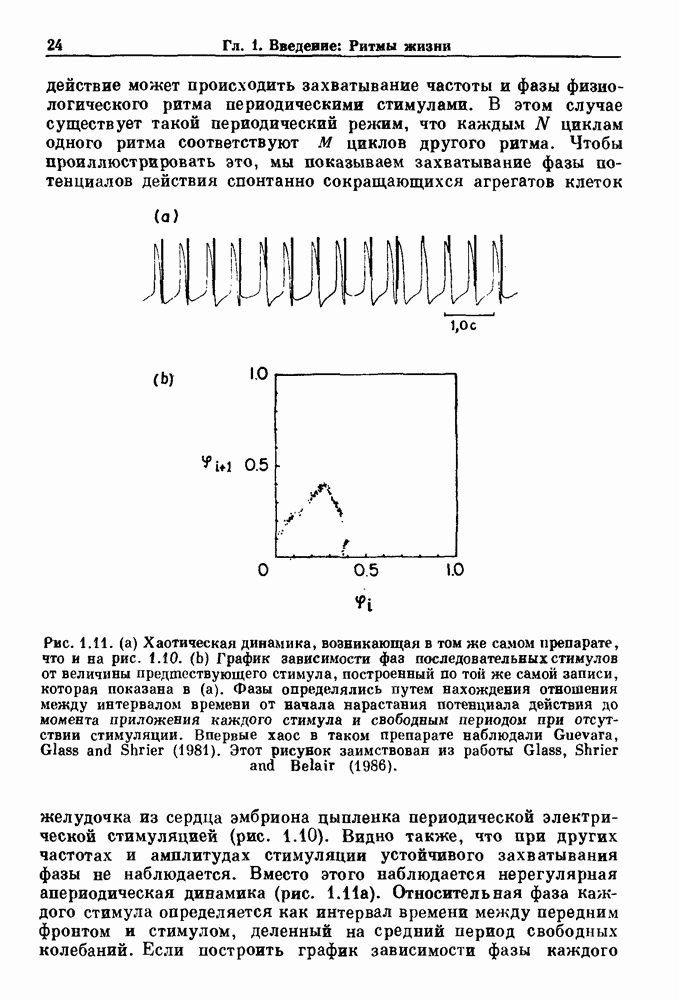
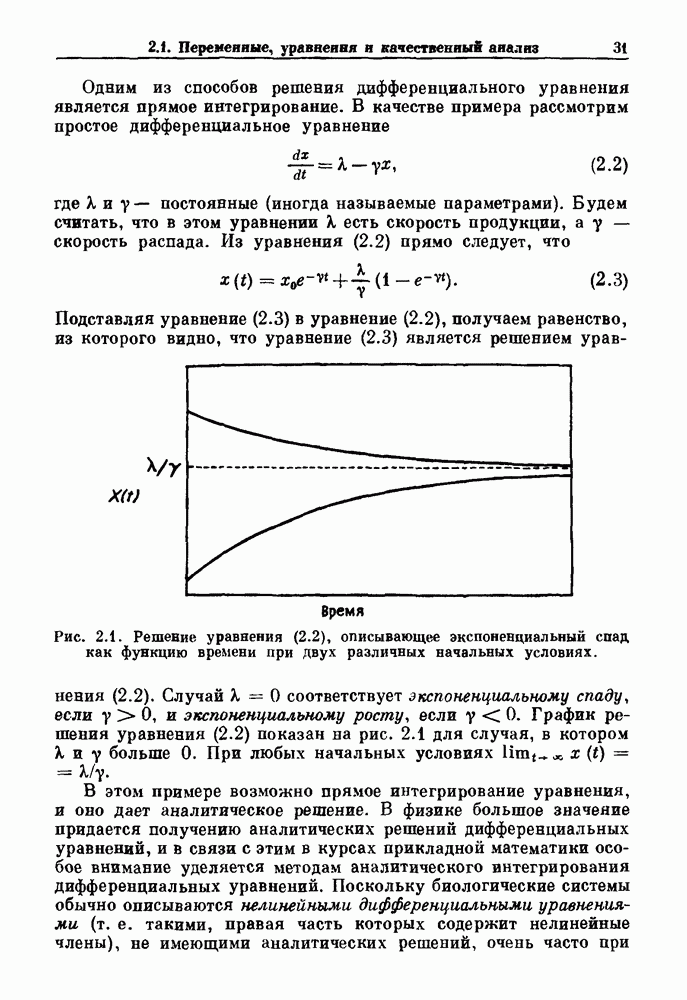
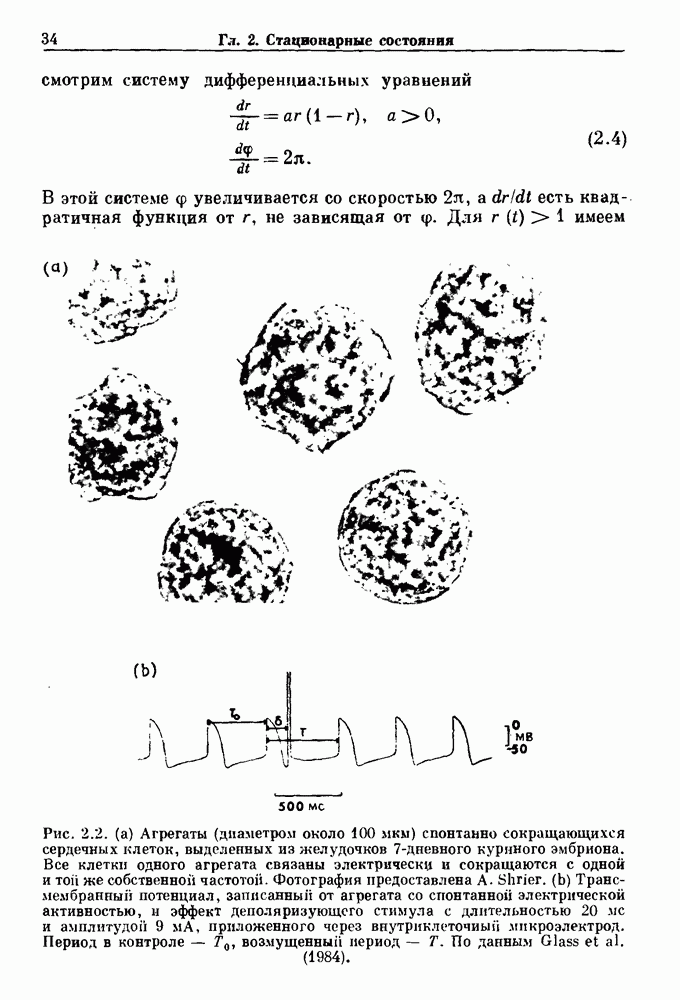
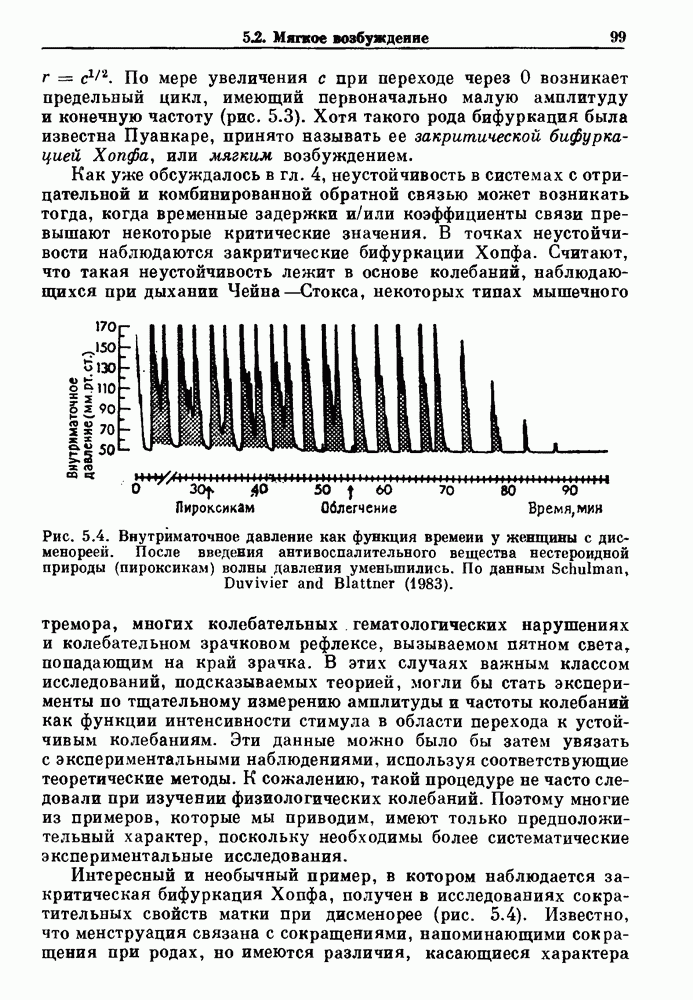
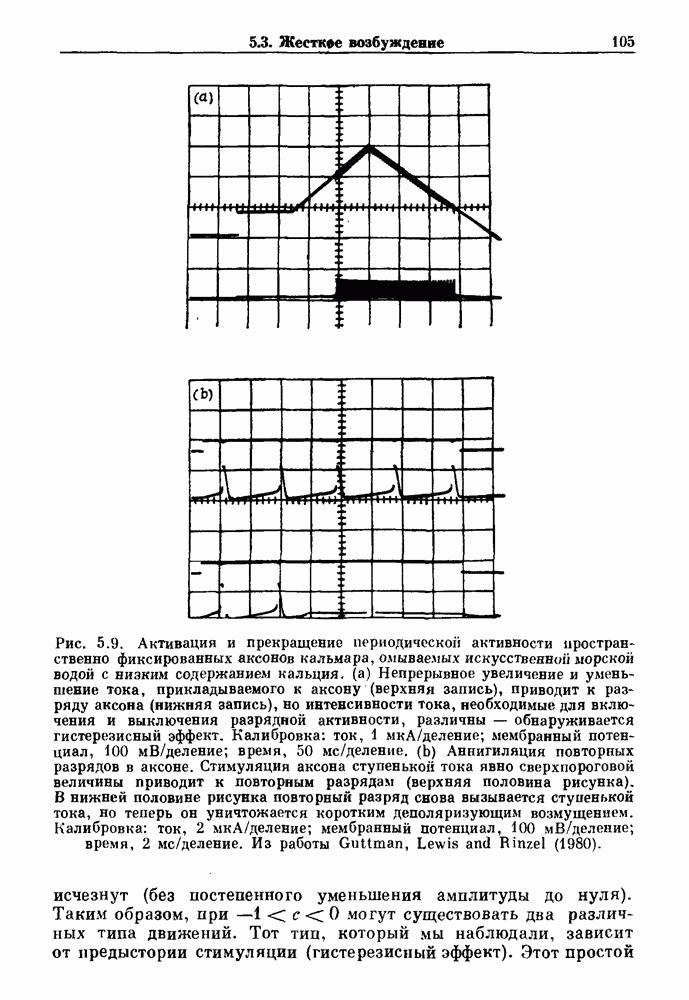
Рис. 5.4. Внутриматочное давление как функция времени у женщины с дисменореей. После введения антивоспалительного вещества нестероидной природы (пироксикам) волны давления уменьшились. По данным Schulman, Duvivier and Blattner (1983). В этих случаях важным классом исследований, подсказываемых теорией, могли бы стать эксперименты по тщательному измерению амплитуды и частоты колебаний как функции интенсивности стимула в области перехода к устойчивым колебаниям. Эти данные можно было бы затем увязать с экспериментальными наблюдениями, используя соответствующие теоретические методы. К сожалению, такой процедуре не часто следовали при изучении физиологических колебаний. Поэтому многие из примеров, которые мы приводим, имеют только предположительный характер, поскольку необходимы более систематические экспериментальные исследования. Интересный и необычный пример, в котором наблюдается закритическая бифуркация Хопфа, получен в исследованиях сократительных свойств матки при дисменорее (рис. 5.4). Известно, что менструация связана с сокращениями, напоминающими сокращения при родах, но имеются различия, касающиеся характера боли. У некоторых пациенток с болезненным протеканием менструации и сокращениями большой амплитуды измерялось с помощью катетера внутриматочное давление на пике дискомфорта. Затем перорально вводилось антивоспалительное вещество нестероидной природы (пироксикам), и внутриматочное давление прослеживалось в течение следующего часа. У одной из пациенток, как показано на рис. 5.4, первоначально отмечавшиеся периодические (или, возможно, хаотические) сокращения большой амплитуды угасали, и это было сопряжено с уменьшением дискомфорта. Облегчение боли было получено 69 % обследованных женщин. Уменьшение амплитуды, показанное на рис. 5.4, — это как раз то, что ожидается при закритической бифуркации Хопфа, когда регулирующий параметр проходит через свое критическое значение (на пример, как при уменьшении с на рис. 5.3). Однако предосторожность была бы здесь не лишней, так как дисменорея представляет собой сложное и малопонятное явление, которое, возможно, вызывается многими факторами. Кроме того, маточные сокращения большой амплитуды неоднозначно связаны с болью в матке. Таким образом, несмотря на существование возможности того, что наблюдаемый переход связан с бифуркацией Хопфа, ее роль в клинике остается спорной. Мягкое возбуждение возникает в системах, отличающихся от систем с отрицательной и комбинированной обратной связью. Примером может служить последовательное деингибирование. Так как последовательное деингибирование было предложено в качестве основы ритмогенеза в дыхательной системе и двигательной активности, интересно изучить экспериментальные данные по инициации и прекращению респираторных и локомоторных ритмов, чтобы выяснить наличие в них мягкого возбуждения при изменении параметров. Полное понимание механизма возникновения и прекращения вентиляции в настоящее время еще не достигнуто. Однако протекание гипервентиляции по сценарию мягкого возбуждения кажется наиболее вероятным. На рис. 5.5 показана активность диафрагмального нерва после гипервентиляции у анестезированной парализованной кошки при применении искусственного дыхания. Постепенное нарастание активности диафрагмального нерва, по-видимому, соответсгвует мягкому возбуждению. Местное охлаждение вентральной поверхности спинного мозга или гипоксия вызывают гораздо более резкие переходы от активного дыхания к его отсутствию. На рис. 5.6 прослеживается активность диафрагмального нерва у анестезированной парализованной кошки с билатеральной ваготомией и синоаортальной денервацией, у которой были перевязаны общие сонные артерии. Там, где указано стрелкой, создавалась непроходимость обеих позвоночных артерий, что закрывало ток крови к дыхательным центрам в стволе мозга. Это немедленно сопровождалось увеличением кровяного давления и небольшим увеличением амплитуды электрической активности диафрагмального нерва, после чего резко возникало апноэ (прекращение дыхания).
Рис. 5.5. Восстановление активности диафрагмального нерва после гипервентиляции у анестезированной парализованной кошки. Верхняя запись — легочный объем, нижняя запись — сглаженная нейрограмма диафрагмального нерва. Дрейф легочного объема при выключенном вентиляторе является артефактом. Записи предоставлены М. Bachoo and С. Polosa. Из работы Glass (1987).
Рис. 5.6. Эффект окклюзии позвоночной артерии у анестезированной парализованной кошки с билатеральной ваготомией, у которой были перевязаны общие сонные артерии. Окклюзия поддерживалась в течение периода времени, указанного стрелками. Верхняя запись — системное артериальное давление (SAP); нижняя запись — сглаженная нейрограмма диафрагмального нерва. Записи предоставлены М. Bachoo and С. Polosa. Из работы Glass (1987). В момент времени, указанный второй стрелкой (спустя примерно одну минуту), в позвоночных артериях восстанавливался нормальный кровоток путем устранения зажима, создающего окклюзию, и это приводило к быстрому возникновению активности диафрагмального нерва. Наши коллеги С. Polosa и М. Bachoo из университета McGill предположили, что резкое прекращение дыхания в экспериментальной обстановке может служить моделью клинически наблюдаемой остановки дыхания, которая является вторичной по отношению к остановке сердца. В качестве еще одного примера рассмотрим исследования регуляции двигательной активности. Обычно быстрое возникновение и прекращение двигательной активности для большинства подвижных животных является необходимым условием выживания.
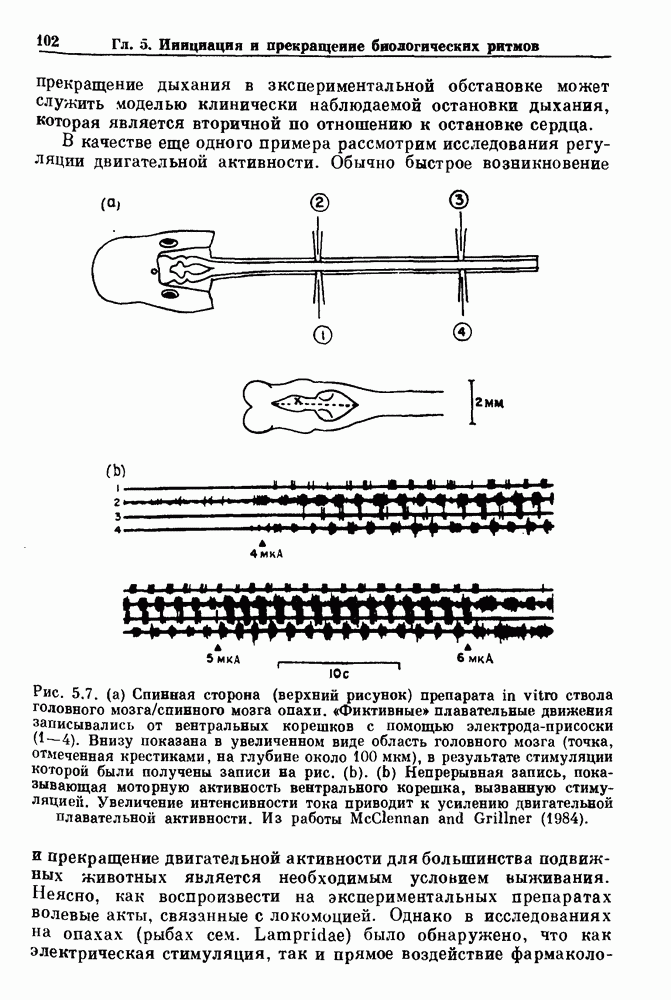
Рис. 5.7. (а) Спинная сторона (верхний рисунок) препарата in vitro ствола головного мозга/спинного мозга опахи. «Фиктивные» плавательные движения записывались от вентральных корешков с помощью электрода-присоски (1—4). Внизу показана в увеличенном виде область головного мозга (точка, отмеченная крестиками, на глубине около 100 мкм), в результате стимуляции которой были получены записи на рис. (b). (b) Непрерывная запись, показывающая моторную активность вентрального корешка, вызванную стимуляцией. Увеличение интенсивности тока приводит к усилению двигательной плавательной активности. Из работы McClennan and Grillner (1984). Неясно, как воспроизвести на экспериментальных препаратах волевые акты, связанные с локомоцией. Однако в исследованиях на опахах (рыбах сем. Lampridae) было обнаружено, что как электрическая стимуляция, так и прямое воздействие фармакологических веществ могут быть эффективными в индуцировании локомоции (или фиктивной локомодии). В качестве примера рассмотрим индукцию фиктивной локомоции у опах электрической стимуляцией (длительность импульсов Однако эта информация по регуляции дыхания и локомоции недостаточна для характеристики бифуркаций, лежащих в основе перехода от покоя к ритмической активности. Необходимо тщательно исследовать динамику как при увеличении, так и при уменьшении регулирующих параметров. Одинаковы или различны пороги возникновения и прекращения ритмической активности при изменении интенсивности стимуляции в сторону увеличения и в сторону уменьшения? При мягком возбуждении нет гистерезиса, и измеряемый порог будет иметь одну и ту же величину как при увеличении, так и при уменьшении регулирующих параметров. В другом случае, при наличии гистерезиса, возможен иной механизм.
|
 |
Оглавление
|























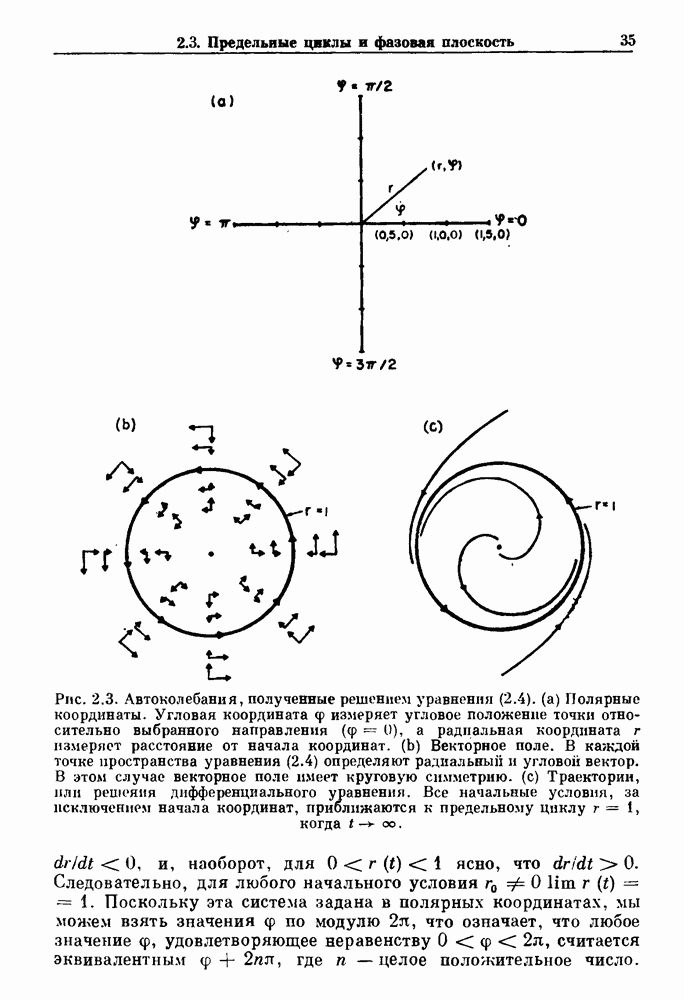


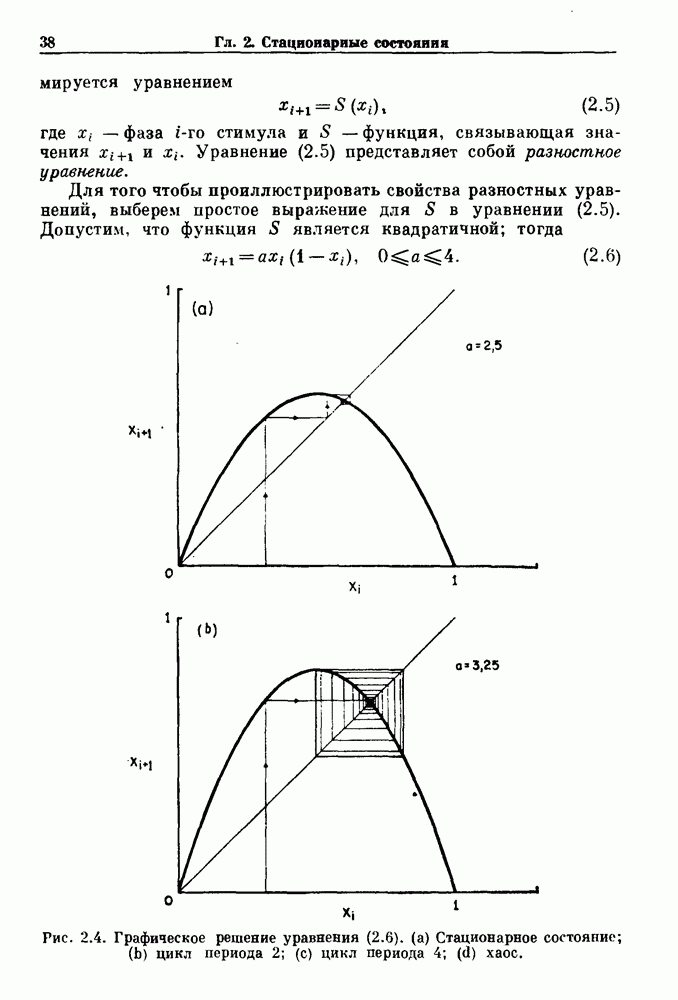
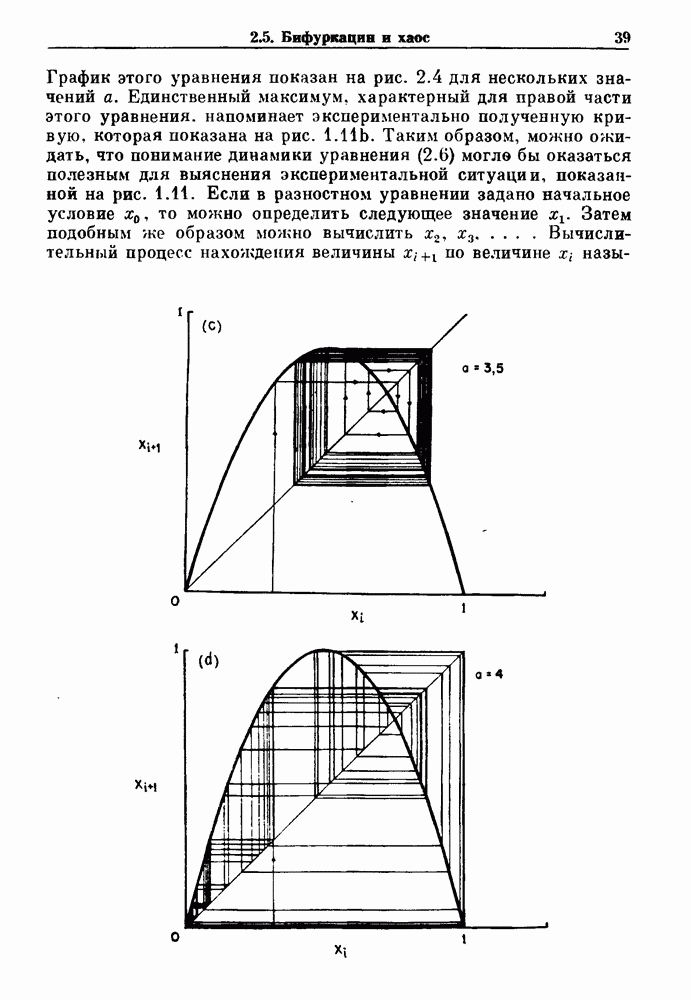
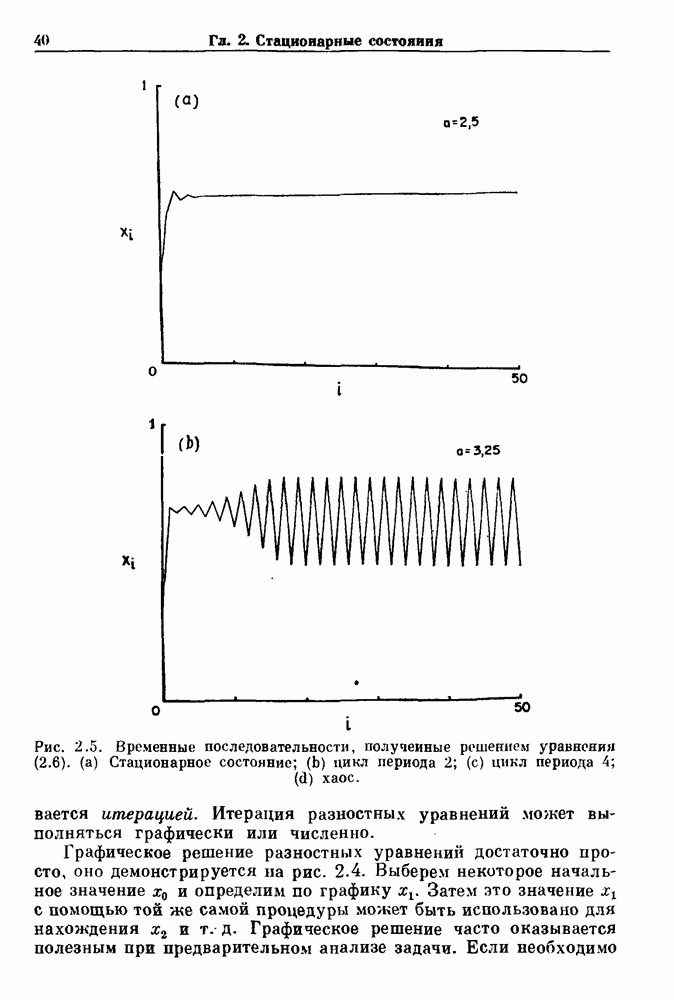
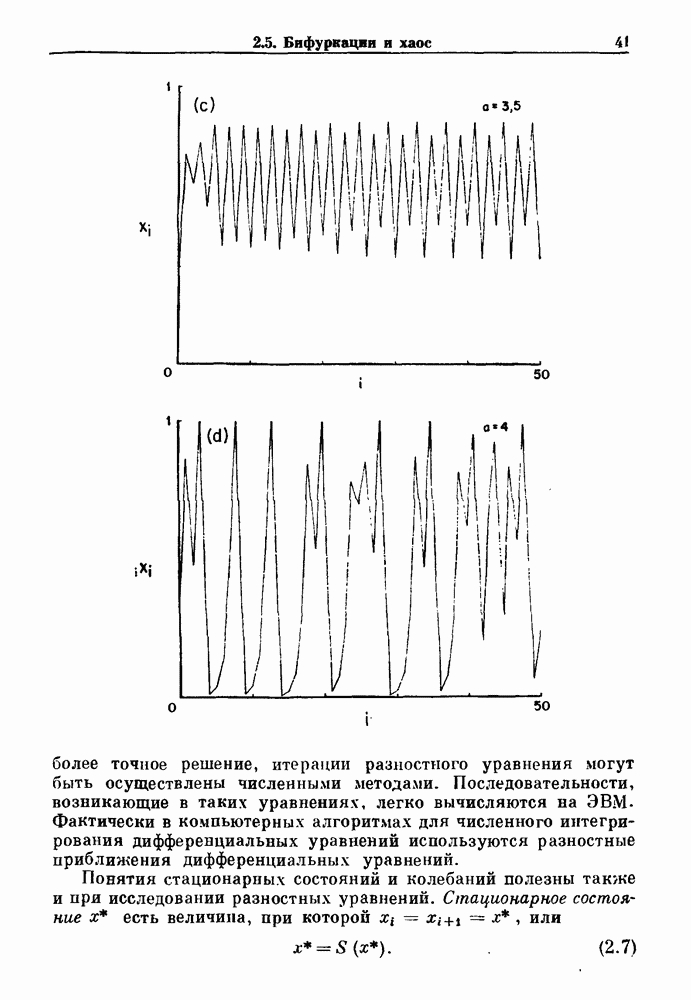

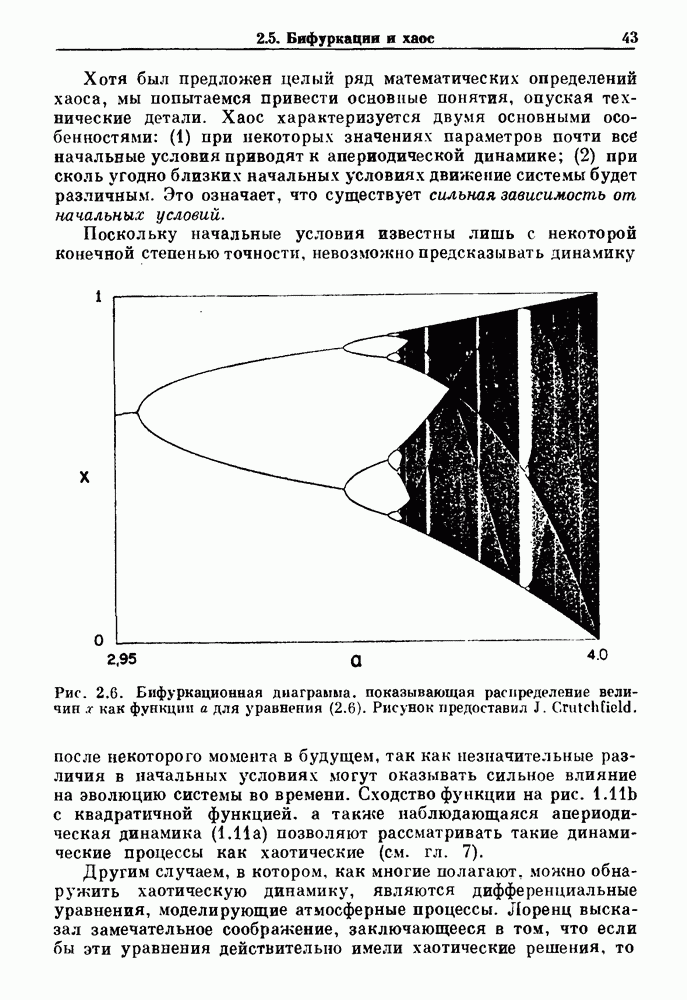



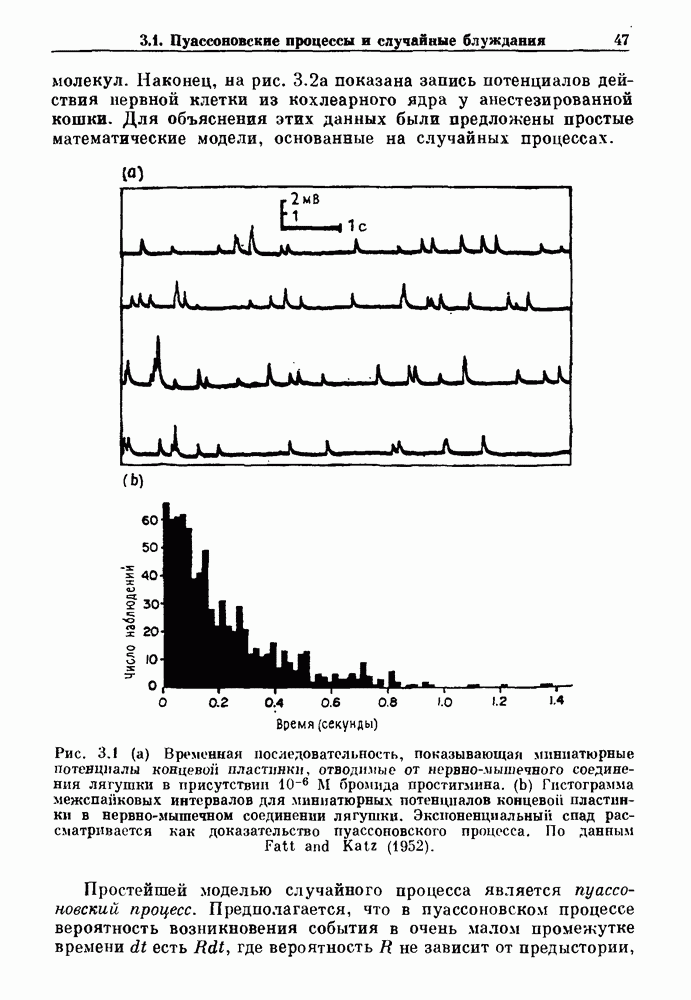
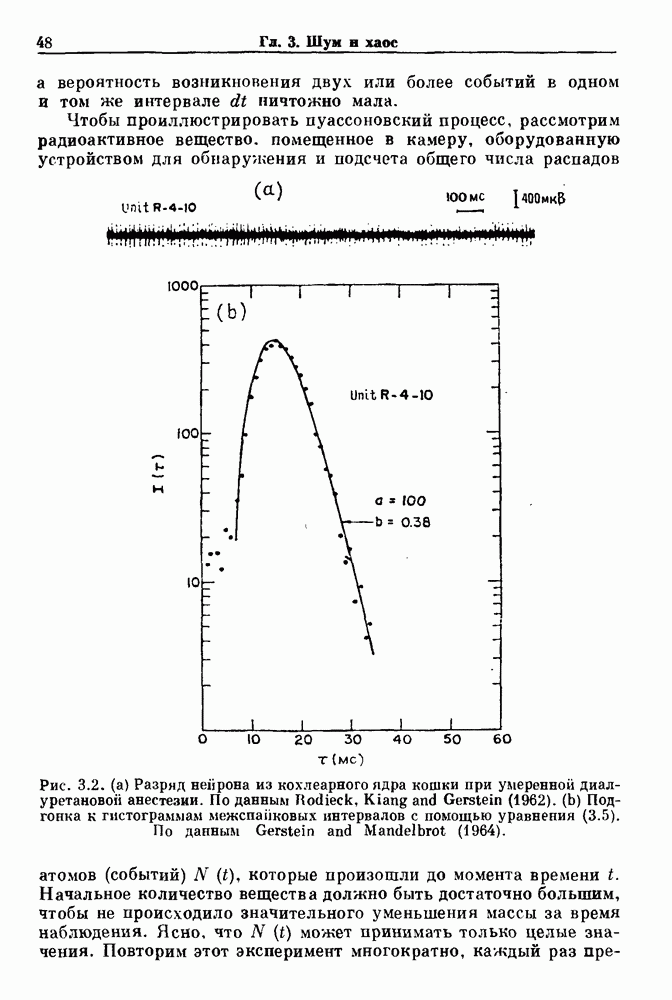







































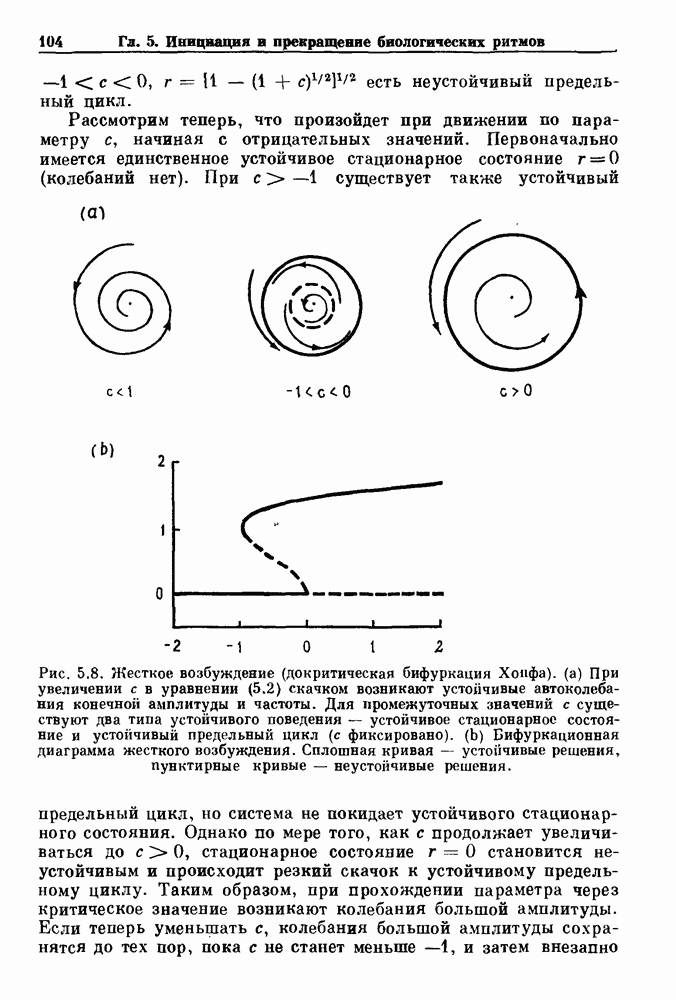

























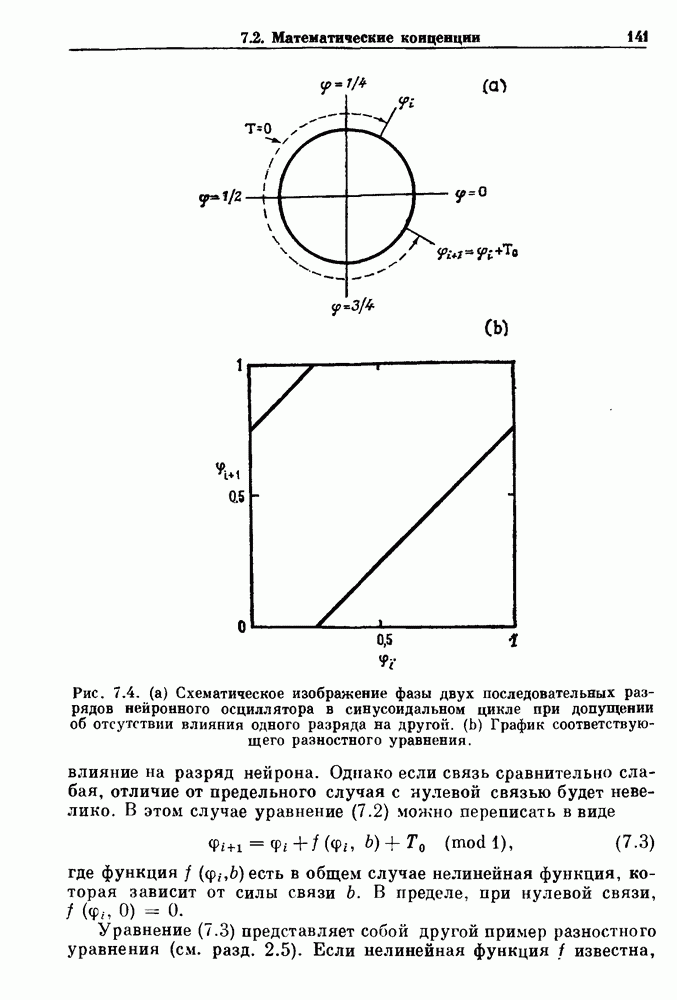



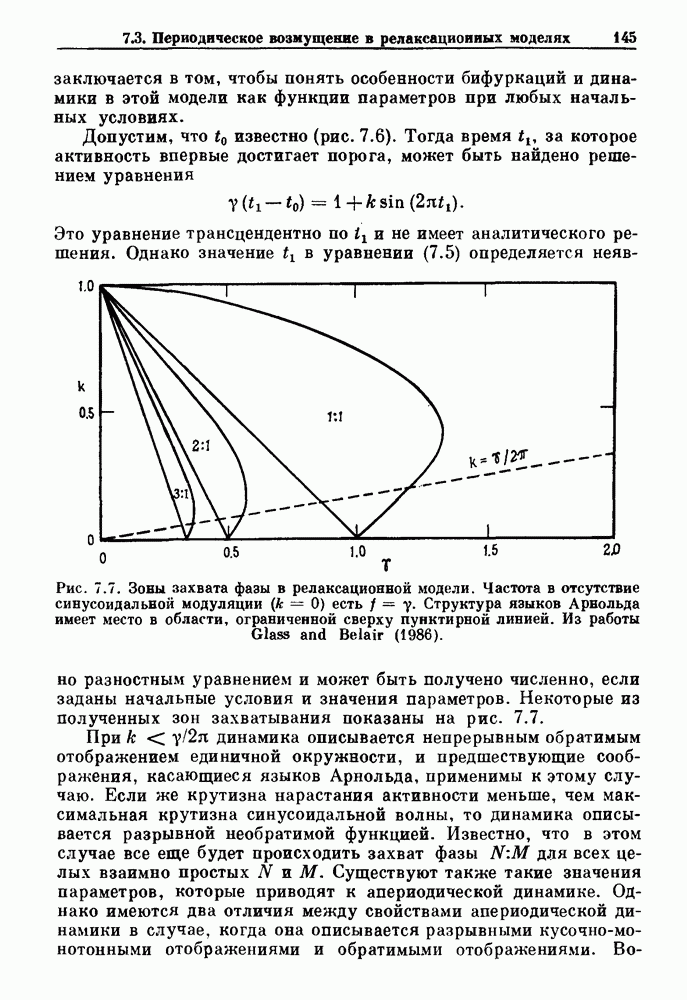
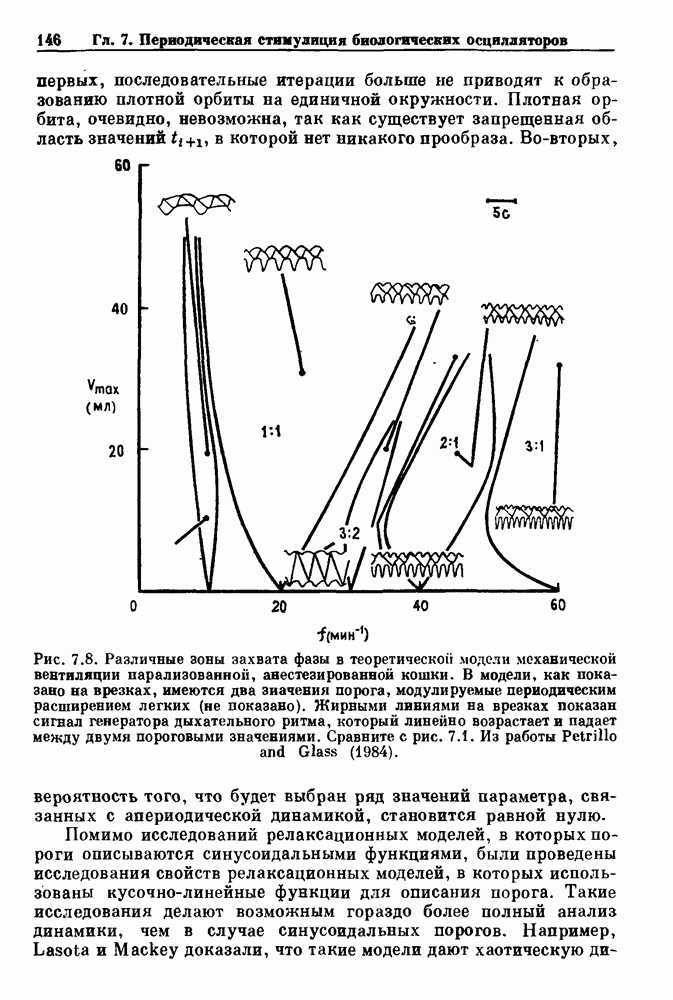


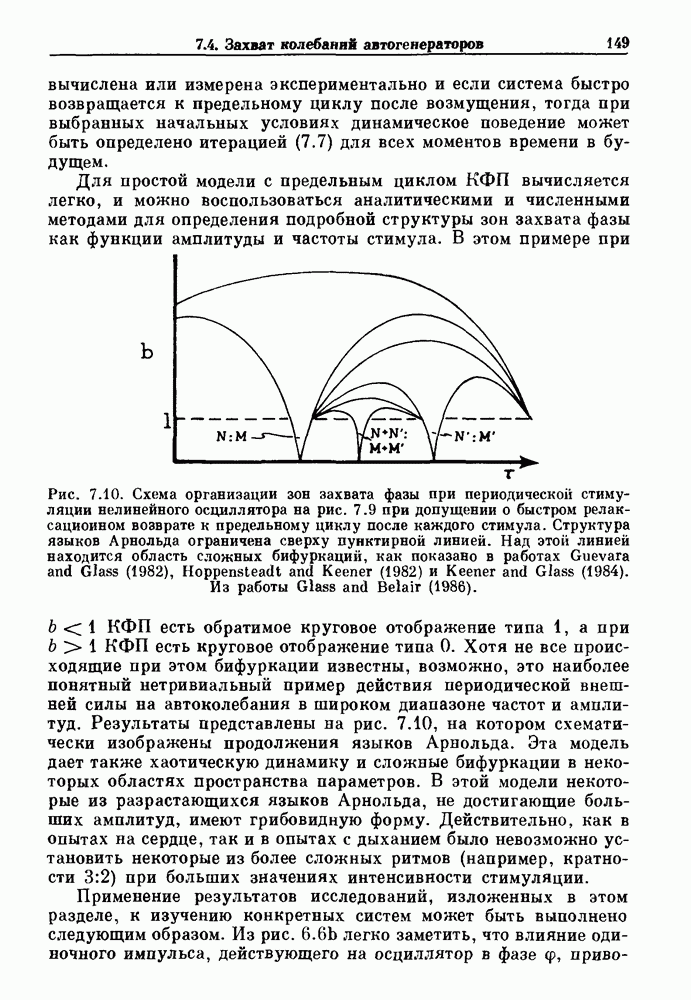


















































































 существует единственное стационарное состояние, которое является устойчивым и глобально притягивающим.
существует единственное стационарное состояние, которое является устойчивым и глобально притягивающим.

 стационарное состояние становится неустойчивым, и возникают устойчивые автоколебания. Конкретным примером системы дифференциальных уравнений, обнаруживающих такую бифуркацию, служат уравнения
стационарное состояние становится неустойчивым, и возникают устойчивые автоколебания. Конкретным примером системы дифференциальных уравнений, обнаруживающих такую бифуркацию, служат уравнения

 существует единственное стационарное состояние, а при
существует единственное стационарное состояние, а при  существует неустойчивое стационарное состояние
существует неустойчивое стационарное состояние  и устойчивый предельный цикл
и устойчивый предельный цикл
 По мере увеличения с при переходе через 0 возникает предельный цикл, имеющий первоначально малую амплитуду и конечную частоту (рис. 5.3). Хотя такого рода бифуркация была известна Пуанкаре, принято называть ее закритической бифуркацией Хопфа, или мягким возбуждением.
По мере увеличения с при переходе через 0 возникает предельный цикл, имеющий первоначально малую амплитуду и конечную частоту (рис. 5.3). Хотя такого рода бифуркация была известна Пуанкаре, принято называть ее закритической бифуркацией Хопфа, или мягким возбуждением.




 частота 20 Гц) ствола мозга (рис. 5.7). Ток силой 3 мкА был недостаточен для того, чтобы вызвать локомоцию, но при силе тока 4 мкА вызывалась ритмическая активность в вентральных двигательных корешках. При этом пороговом значении тока разряды начинались немедленно или несколько секунд спустя, и первый разряд мог возникать на любой стороне. Разрядная активность регулировалась величиной стимула.
частота 20 Гц) ствола мозга (рис. 5.7). Ток силой 3 мкА был недостаточен для того, чтобы вызвать локомоцию, но при силе тока 4 мкА вызывалась ритмическая активность в вентральных двигательных корешках. При этом пороговом значении тока разряды начинались немедленно или несколько секунд спустя, и первый разряд мог возникать на любой стороне. Разрядная активность регулировалась величиной стимула.