Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
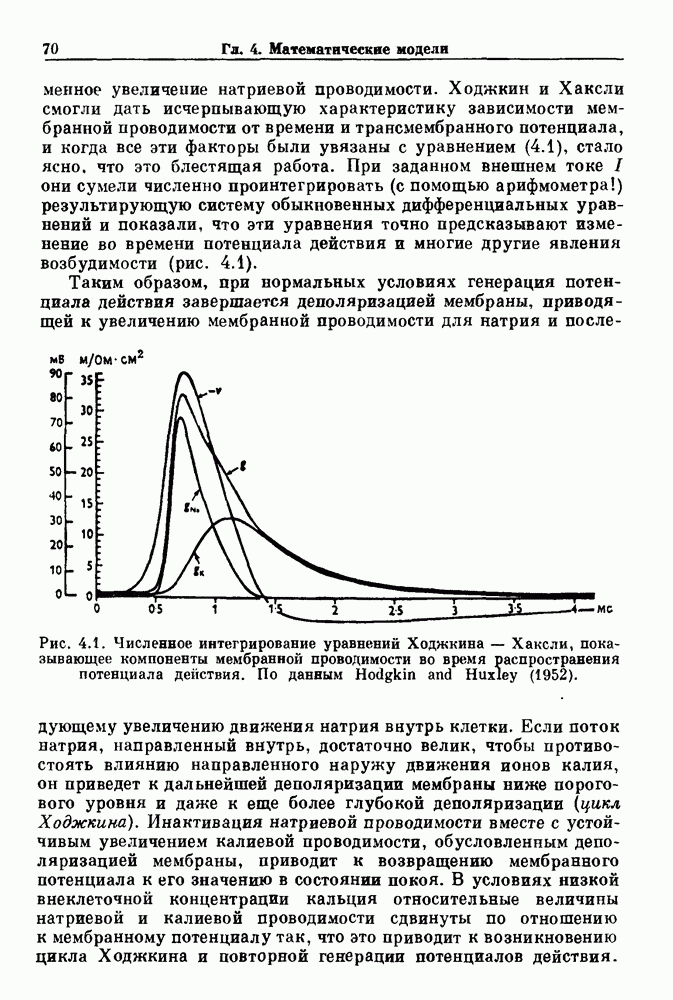
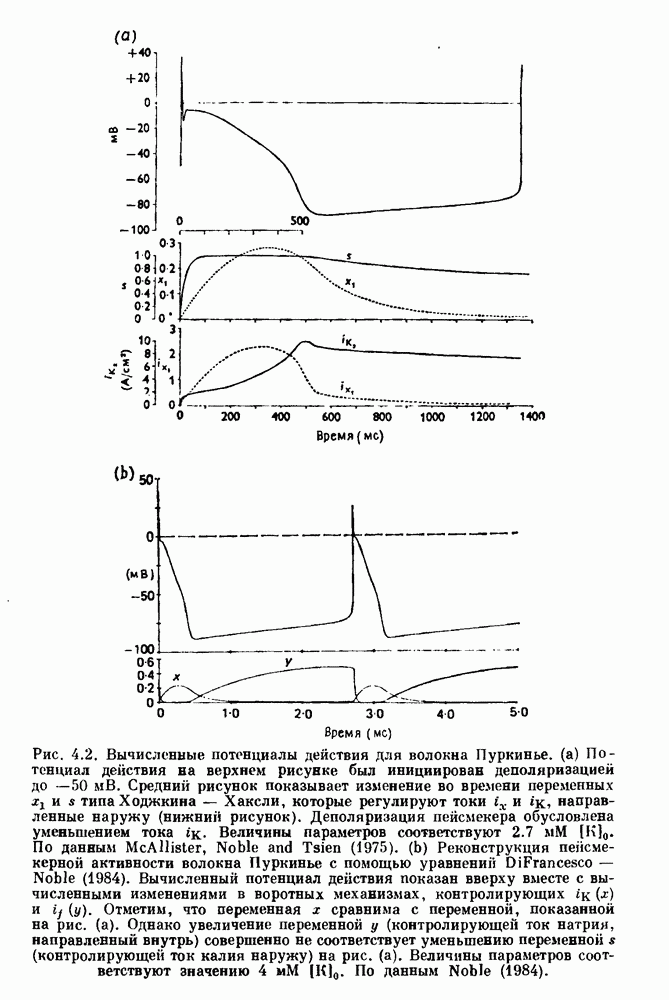
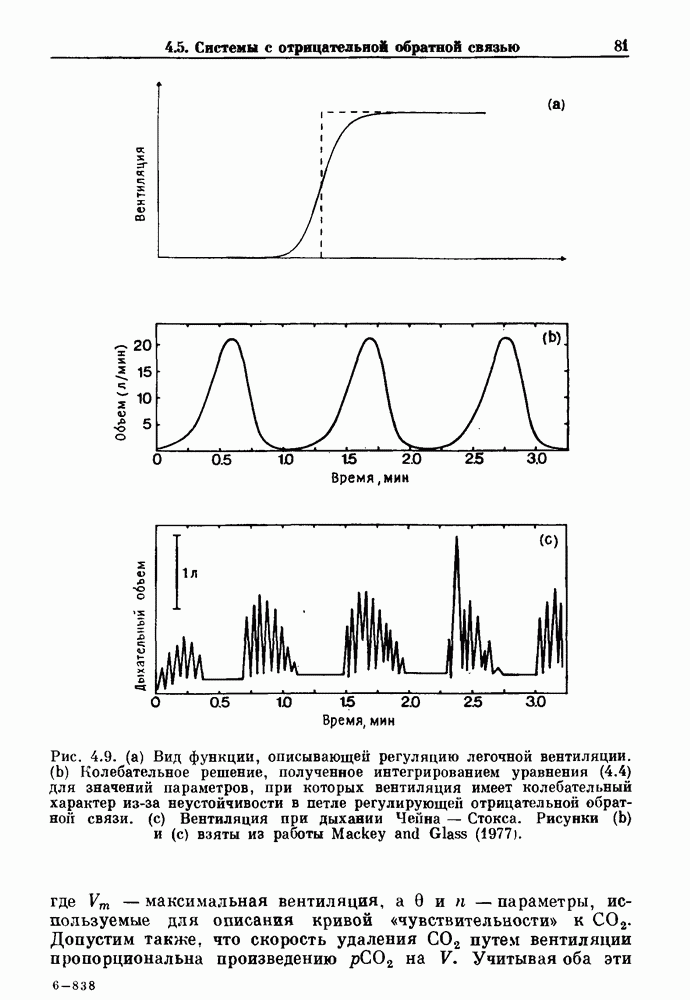
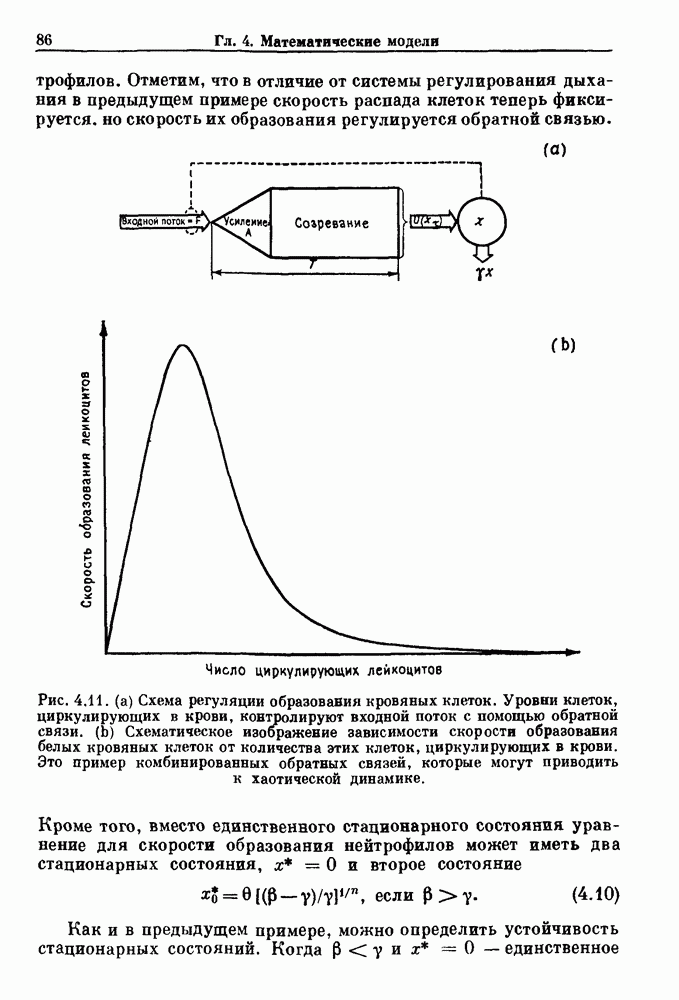
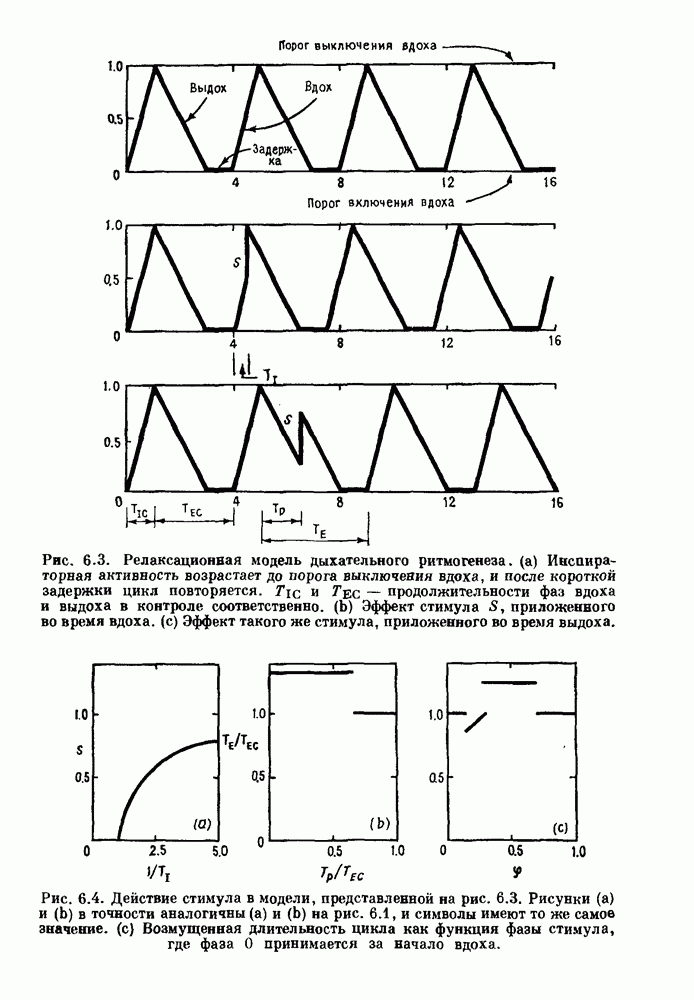
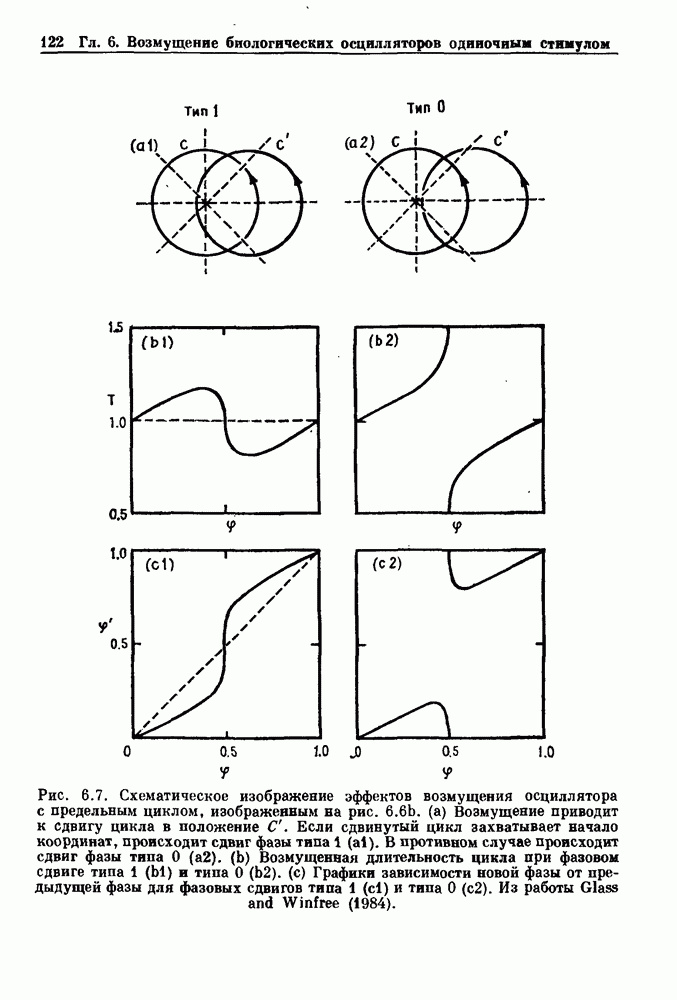
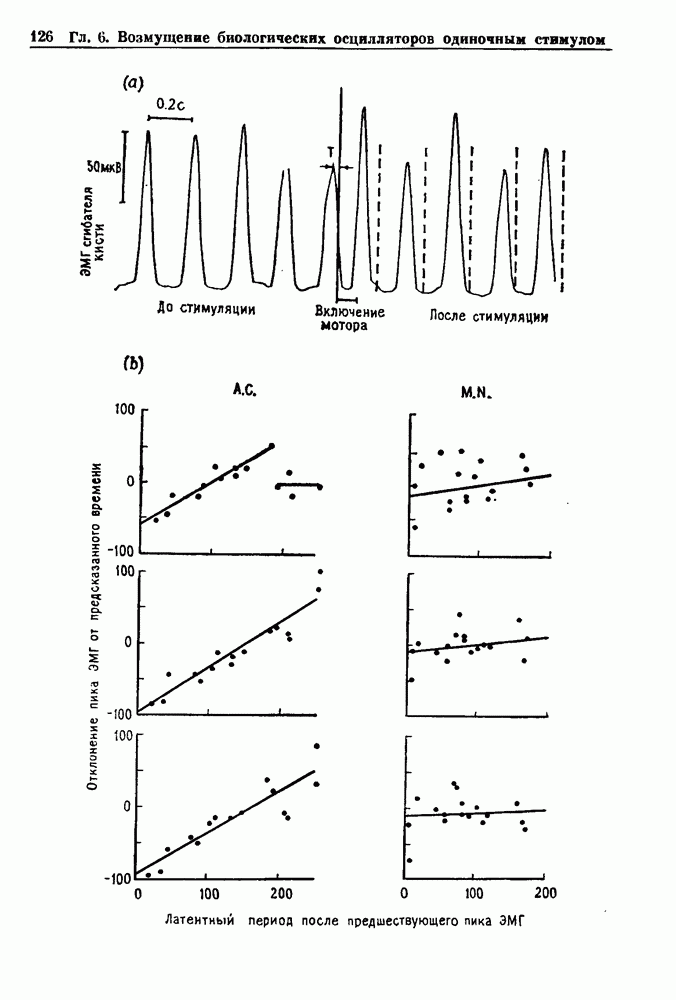
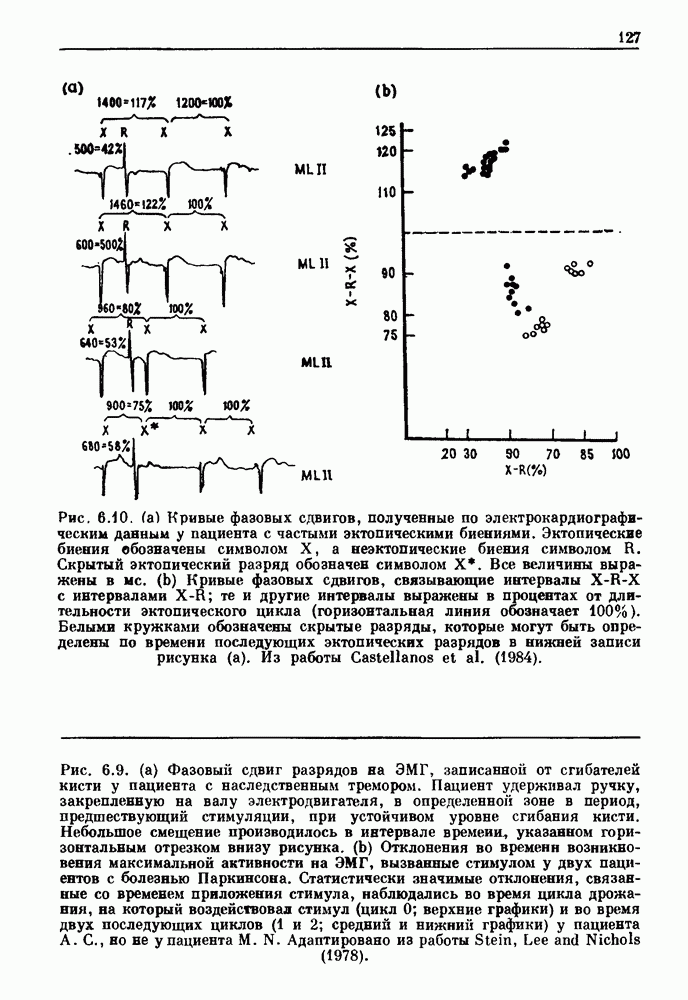
Примечания и литература, глава 44.1. Пейсмекерные колебанияРабота, имеющая решающее значение для выяснения природы процесса возбуждения в классической возбудимой ткани, гигантском аксоне кальмара, была выполнена Ходжкином и Хаксли (Hodgkin and Huxley (1952)). Позднее Хаксли расширил эти исследования, изучив влияние растворов с низким содержанием кальция. Многие авторы использовали этот подход для характеристики других возбудимых тканей. McAllister, Noble and Tsien (1975) изучали потенциалы действия в сердце, но некоторые их результаты недавно были пересмотрены (Noble (1983, 1984); Di Francesco (1984); Di Francesco and Noble (1985)). Нерегулярная динамика пейсмекерной активности наблюдалась экспериментально в ряде нейронных систем (Chalazontis and Boisson (1978)), в -клетках поджелудочной железы (Lebrun and Atwater (1985)). и после воздействия биологически активных веществ на пейсмекерные клетки моллюска (Holden, Winlow and Haydon (1982)). Математические модели пейсмекерных колебаний в нейронах (Chay (1984), 4.2. Генераторы центрального типаИсследование генераторов центрального типа (ГЦТ) началось еще в начале этого века, когда Brown (1914) продемонстрировал наличие двигательной активности у кошек, у которых была устранена афферентная обратная связь. Однако четкого теоретического объяснения механизмов ГЦТ пока еще нет, и некоторые исследователи выражают скептическое отношение к роли теории (М. I. Cohen (1979)). Мы не пытаемся дать здесь полный обзор работ по ритмогенезу в различных системах, но отсылаем читателя к блестящему сборнику статей под ред. А. Н. Cohen, Rossignol and Gril-lner (1988). 4.3. Взаимное ингибированиеКонцептуальные и математические модели, основанные на взаимном ингибировании, были предложены в различных работах (Т. G. Brown (1914); Burns and Salmoiraghi (1960); Salmoiraghi and Burns (1960); Harmon (1964); Perkel and Mulloney (1974)). Колебания в системах с взаимным ингибированием продемонстрировали Selverston, Miller and Wadepuhl (1983) и Satterlie (1985). Взаимное ингибирование не обязательно приводит к колебаниям, но может порождать ситуацию, когда тот или другой из двух нейронов остается активным. Hopfield (1984) предположил, что такой нейронный переключатель может играть определенную роль в процессах памяти. 4.4. Последовательное деингибированиеПервое обсуждение последовательного деингибирования содержится в работах Szekely (1965) и Kling and Szekely (1968). Обсуждение и анализ таких систем с использованием булевой алгебры можно найти в статьях Glass and Young (1979) и Thomas (1979). Дифференциальные уравнения, моделирующие последовательное деингибирование и другие сложные взаимодействия систем, приводятся в работах Glass and Pasternack (1978а, b). Обзоры по механизмам дыхательного ритмогенеза содержатся в работах М. I. Cohen (1974, 1979), Richter and Ballantyne (1983), Euler (1986) и Feldman (1986). Из них можно заключить, что дыхательный ритм, генерируемый нейронной сетью, имеет сложную природу и связан с несколькими различными типами взаимодействующих клеток. Предположение о том, что системы последовательного деингибирования могут лежать в основе дыхательного ритмогенеза, высказано в работе Petrillo and Glass (1984). 4.5. Системы с отрицательной обратной связьюХорошо известно, что системы с отрицательной обратной связью способны давать колебания, когда временные задержки и или коэффициенты связи велики (Grodins (1963); Milhorn (1966); Stark (1968)). Однако многие исследования устойчивости таких систем используют линейные методы и, таким образом, имеют ограниченное применение для анализа нелинейных колебаний, которые могут наблюдаться вне линейной области уравнений. Модель дыхания Чейна—Стокса, которую мы рассматривали, можно найти в работах Mackey and Glass (1977) и Glass and Mackey (1979a); там подробно описаны способы оценки значений параметров. Другие, более сложные математические модели были разработаны для объяснения изменений в концентрации проводится в работе Waggener et al. (1984). Недавний обзор Pack and Millman (1986) по регулированию дыхания у пожилых людей содержит обширную литературу. Колебательная неустойчивость в системах с отрицательной обратной связью имеет отношение к возникновению колебаний во многих других системах физиологического регулирования (Grodins (1963); Milhorn (1966)). Другие интересные примеры были получены при изучении динамики зрачкового рефлекса (Stark (1968, 1984)), периодической автоиммунной гемолитической анемии (Mackey (1979b)), периодической кататонической шизофрении (Gronin-Skanlon (1974)), рефлекса растяжения (Lippold (1970)), тремора (Merton, Morton and Rashbass (1967)) и кровяного давления (Sagawa, Carrier and Guyton (1962); Hosomi and Hayashida (1984)). Множественная отрицательная обратная связь с задержкой может порождать сложные ритмы (Glass, Beuter and Larocque (1988)). 4.6. Колебания в системах с комбинированными обратными связями и временными задержкамиРассмотренная здесь модель регуляции образования белых кровяных клеток предложена Mackey and Glass (1977) и Glass and Mackey (1979a). Общее обсуждение схемы построения системы гемопоэза можно найти у Wintrobe (1976), Mackey (1979а) и Que-senberry and Levitt (1979). Данные Wazewska-Czyzewska (1984) свидетельствуют о том, что при очень низких уровнях нейтрофилов скорость их продукции падает, приближаясь к нулю. Данные, указывающие на увеличение времени созревания клеток при хронической миелогенной лейкемии, содержатся в работах Killman et al. (1963), Ogawa et al. (1970), M. L. Greenberg et al. (1972) и Gavosto (1974). Ряд исследователей разработали модели различной сложности для описания динамики рекуррентного ингибирования (Mates and Horowitz (1976); Kaczmarek and Babloyantz (1977); Traub and Wong (1981); Knowles et al. (1985)). Рассмотренная здесь модель предложена Mackey and an der Heiden (1984). Другие исследователи изучали хаотическое поведение решений дифференциальных уравнений с временной задержкой в моделях с комбинированной обратной связью, описывающих регуляцию эритропоэза (Wazewska—Czyzewska and Lasota (1976); Lasota (1977)); в системах общего физиологического регулирования (ап der Heiden (1979)); в модели динамики действия допамина (King, Barchas and Huberman (1984)); в других моделях рекуррентного ингибирования (an der Heiden, Mackey and Walther (1981)) и в экологии (J. F. Perez, Malta and Coutinho (1978); May (1980)). Метод замены нелинейностей кусочно-постоянными, кусочно-линейными или другими специальными функциями использовался в работах Glass and Mackey (1979a), Peters (1980), an der Heiden and Mackey (1982), Saupe (1982), an der Heiden (1985) и Walther (1985). Указанный метод позволил этим исследователям доказать некоторые очень интересные свойства решений уравнений. Экспериментальное исследование механизма комбинированной обратной связи в зрачковом рефлексе описано в работе Longting and Milton (1988). Интересный побочный результат исследований заключается в том, что уравнения, первоначально предложенные нами для моделирования образования белых кровяных клеток, были широко использованы математиками и физиками для проверки алгоритмов вычисления размерности и числа Ляпунова (см. разд. 3.4) по временным последовательностям (Farmer (1982); Grassberger and Pro-caccia (1983); Wolf et al. (1985); Le Berre et al. (1987); Kostelich and Swinney (1987)).
|
 |
Оглавление
|















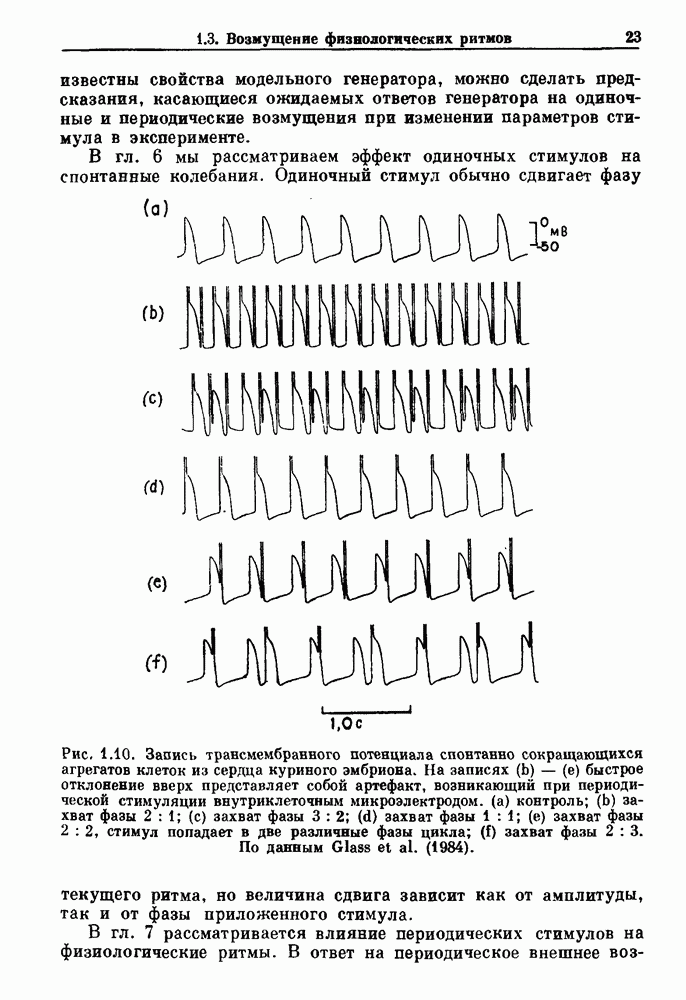
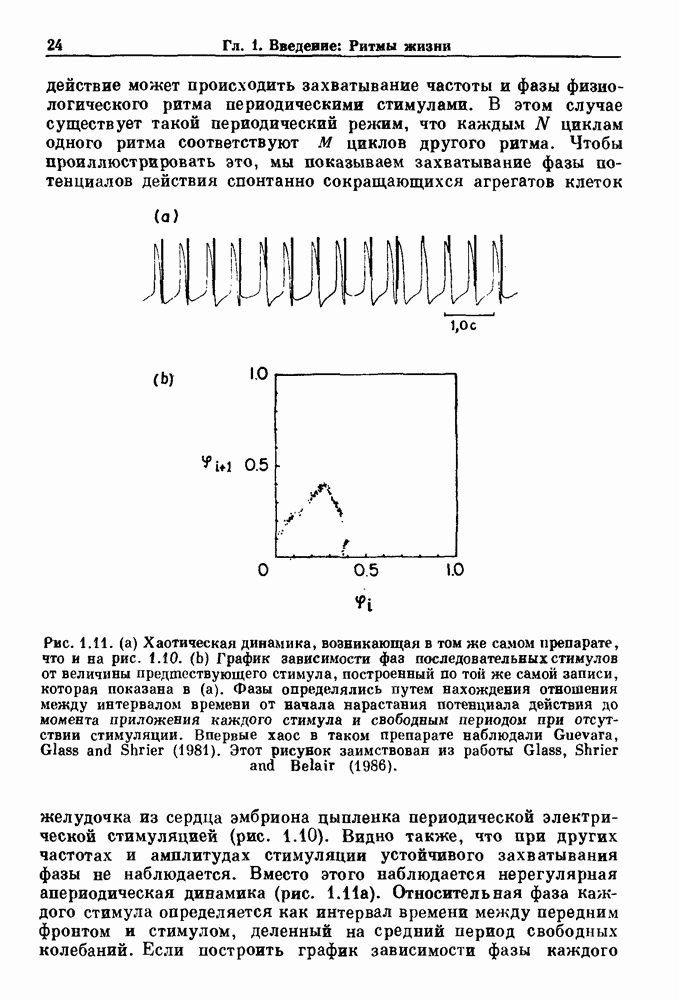






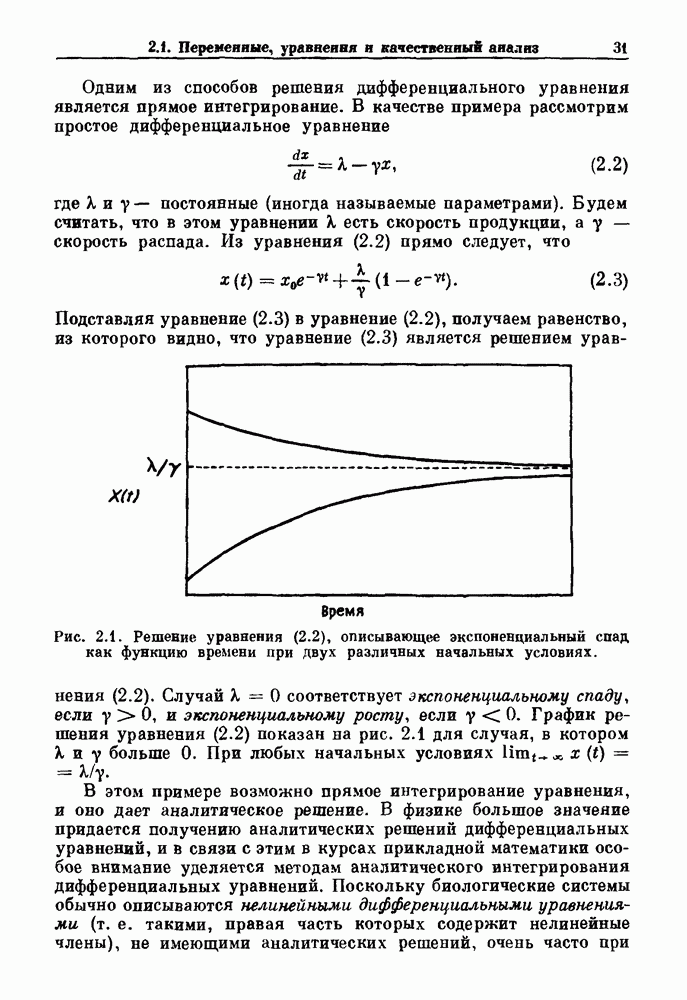


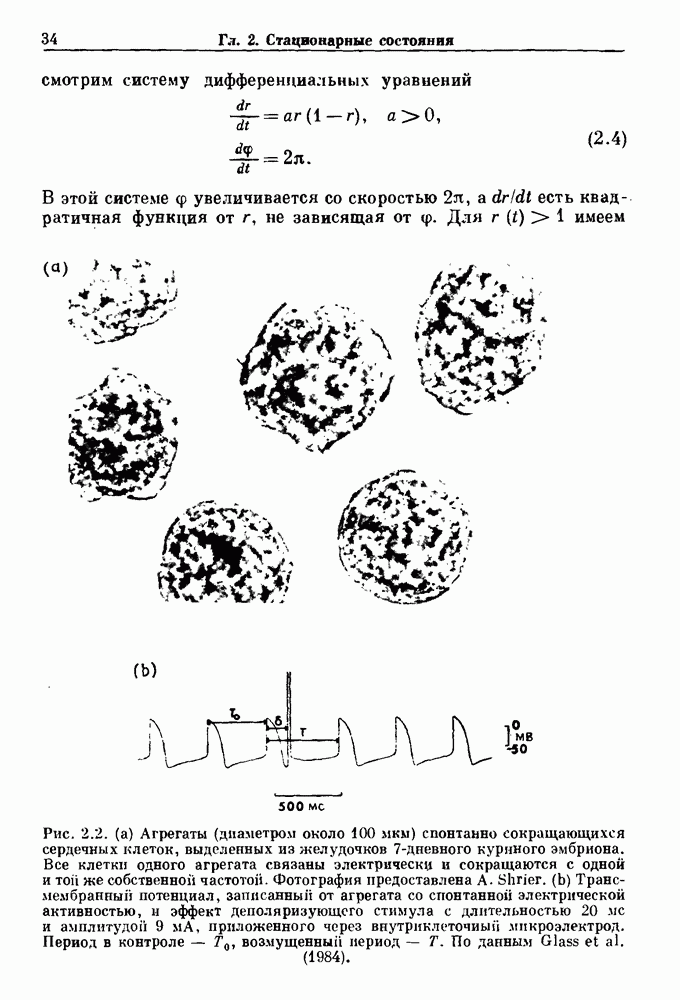
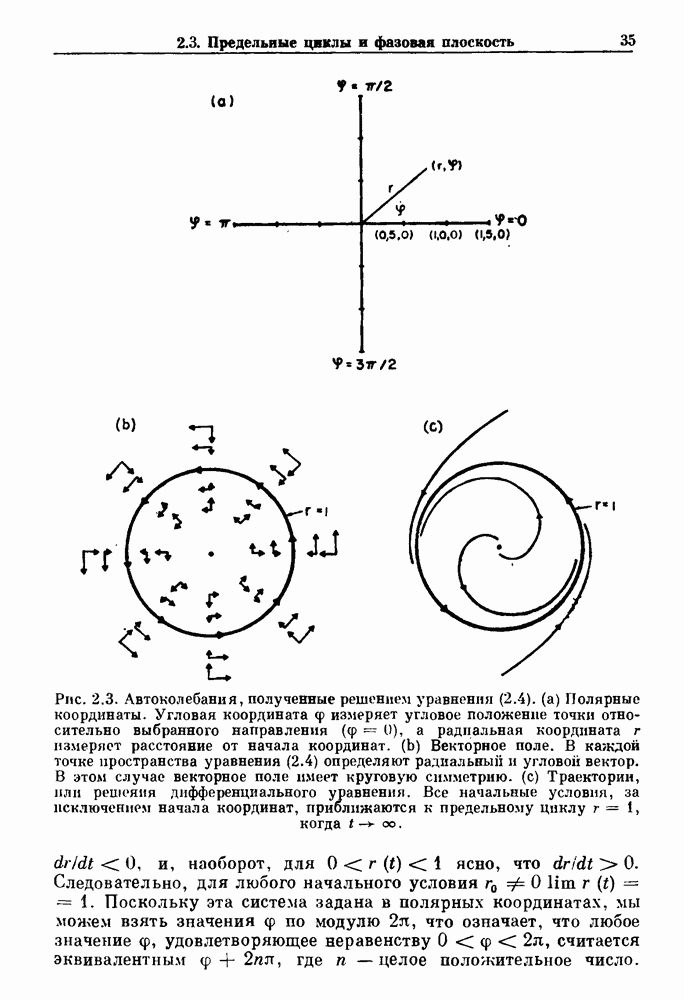


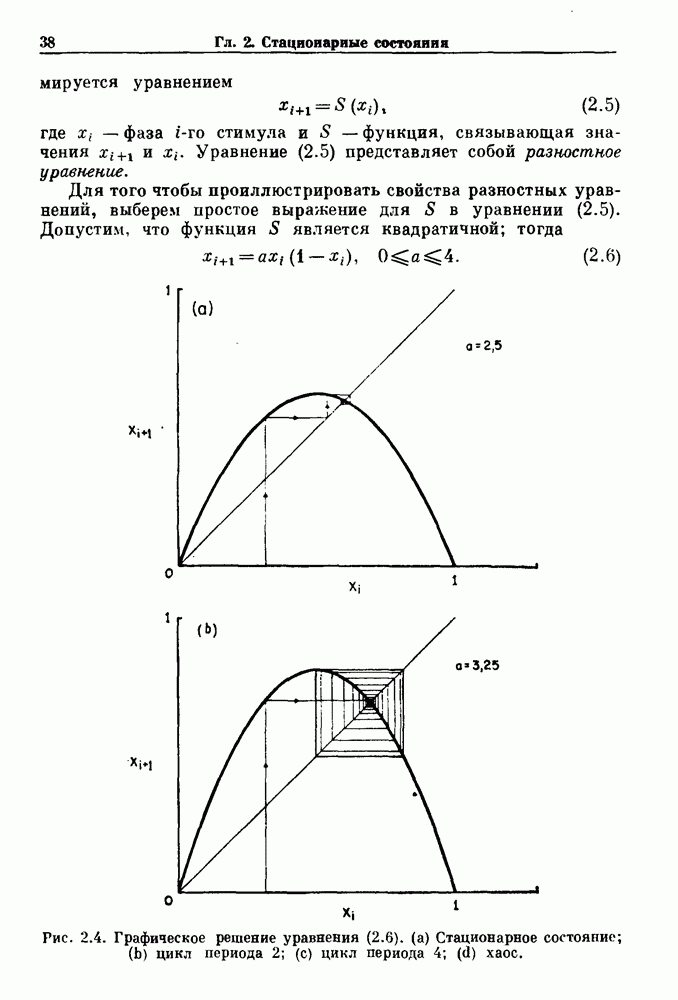
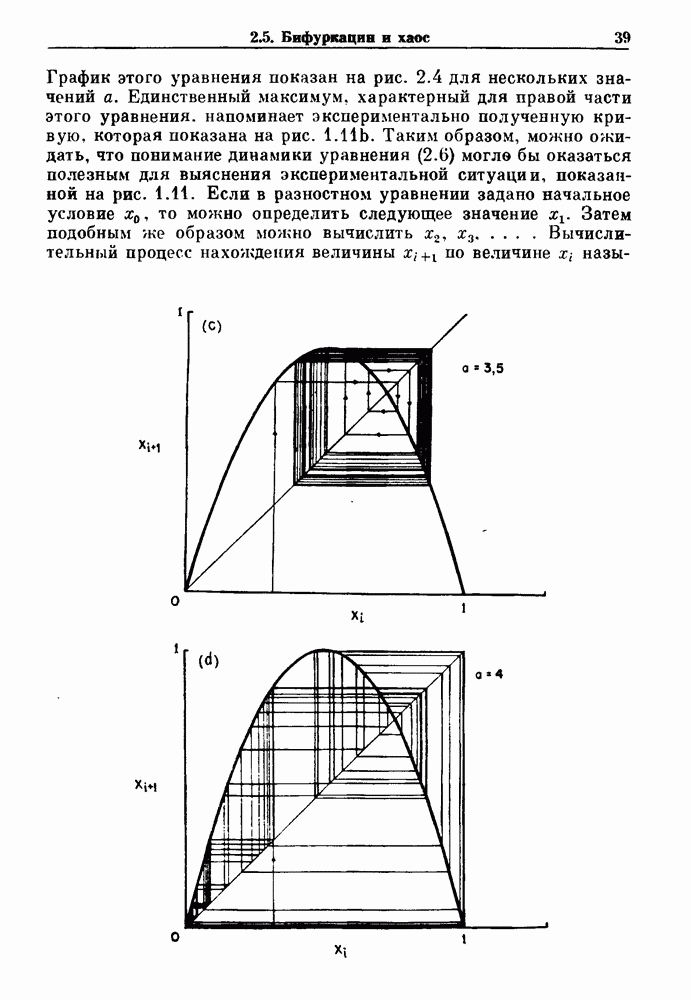
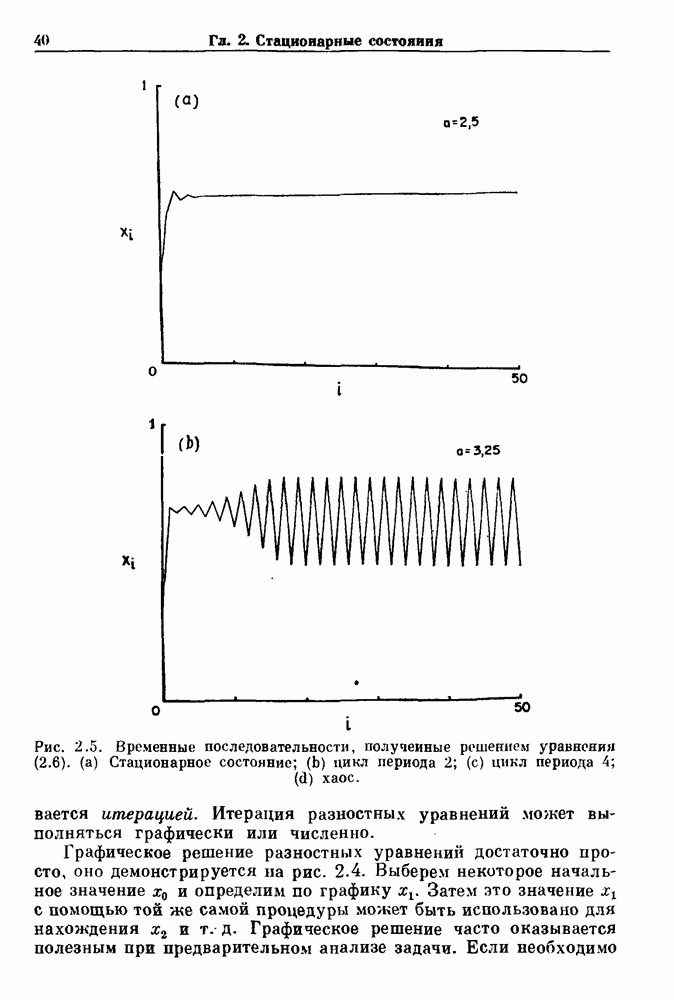





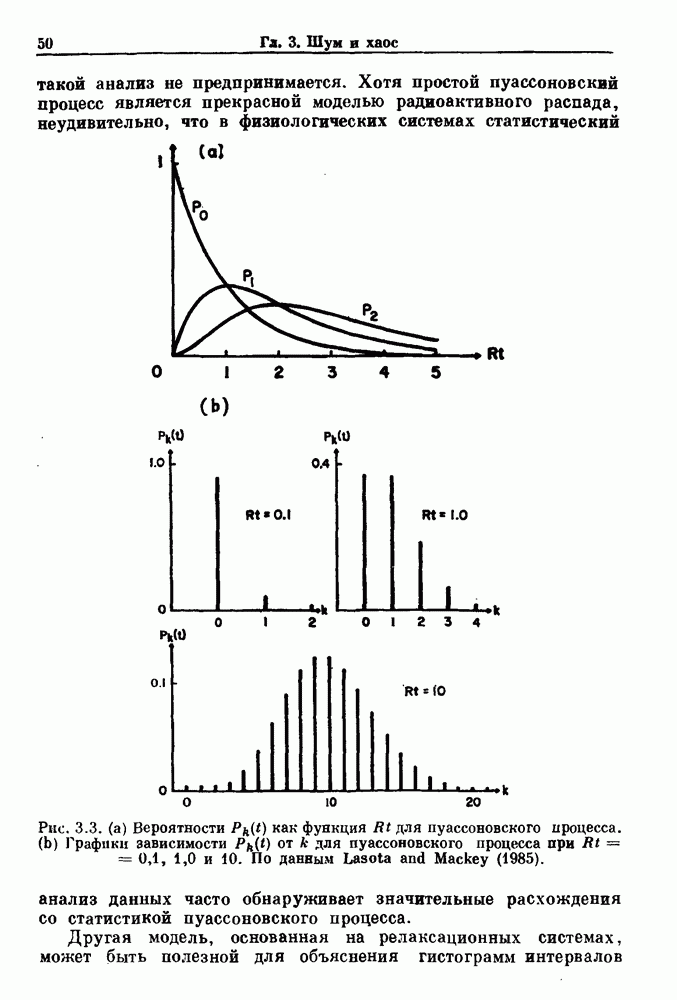
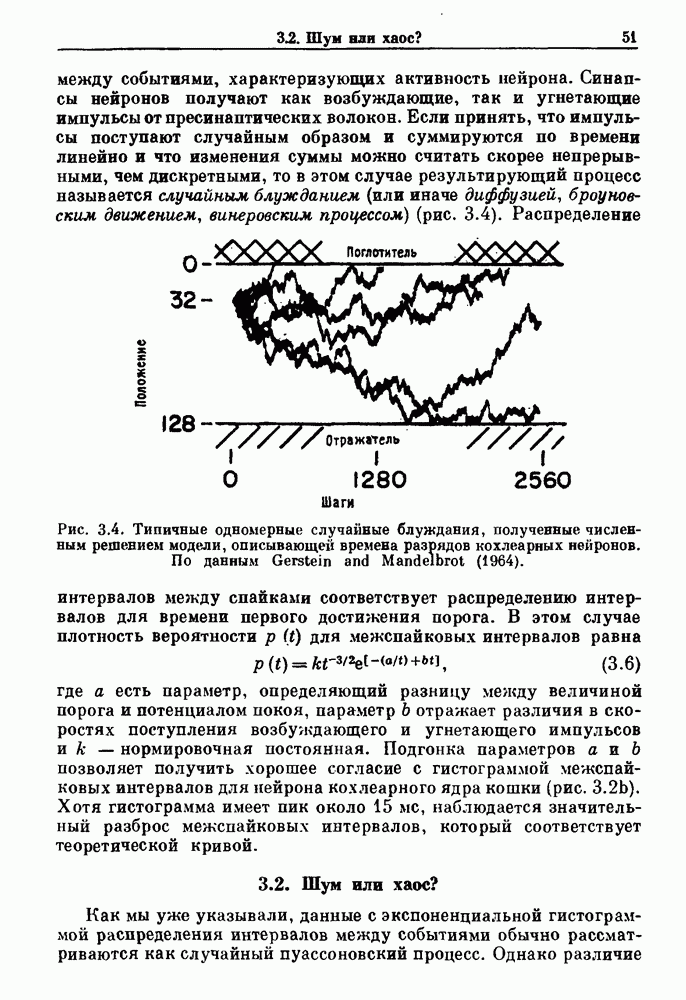

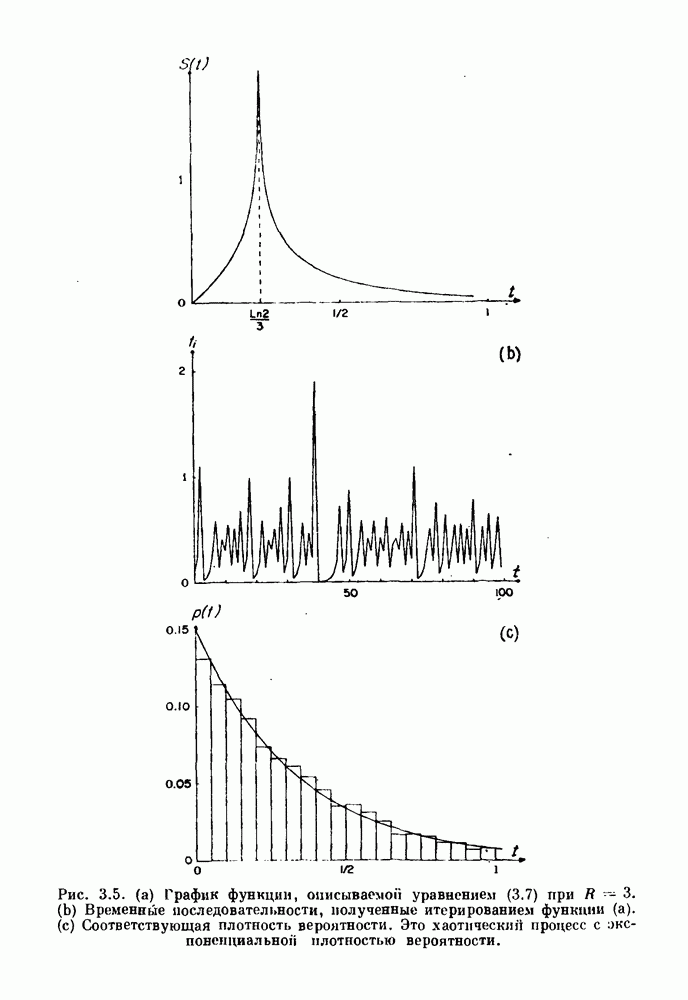

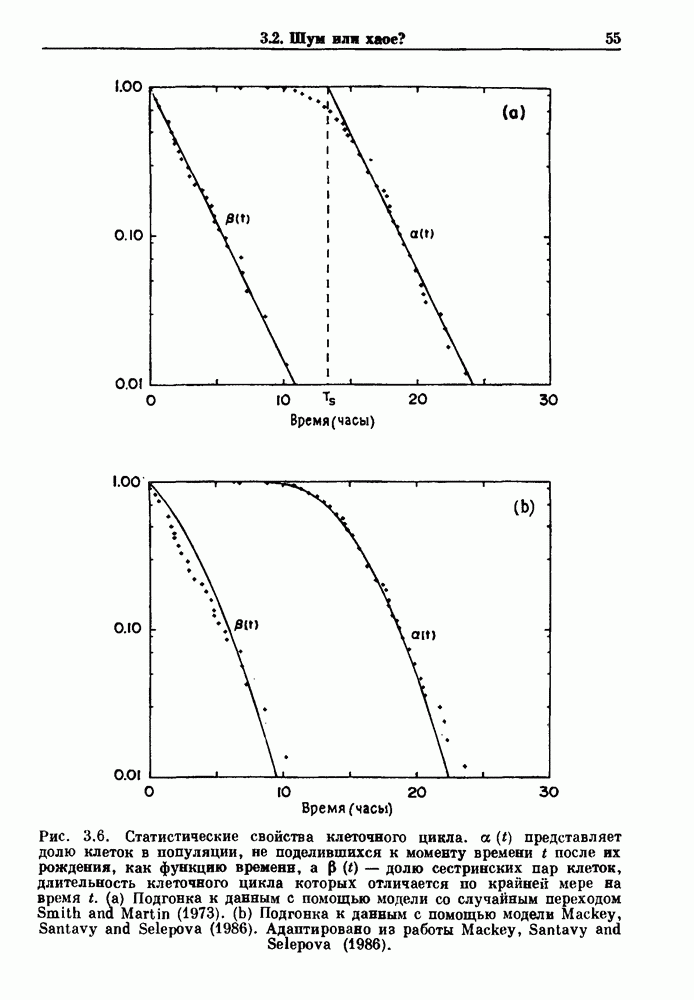
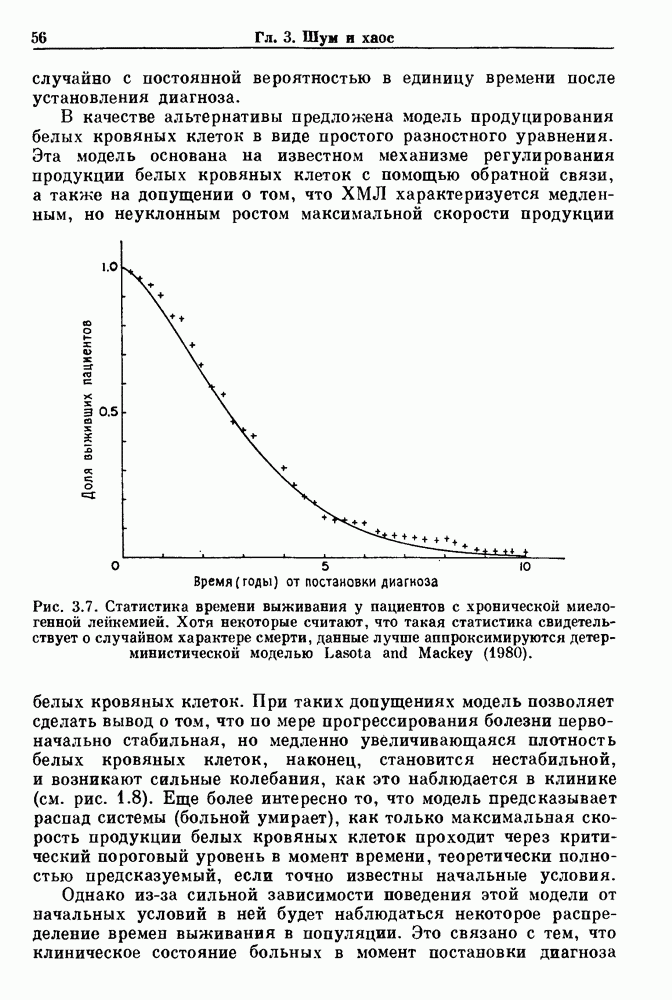




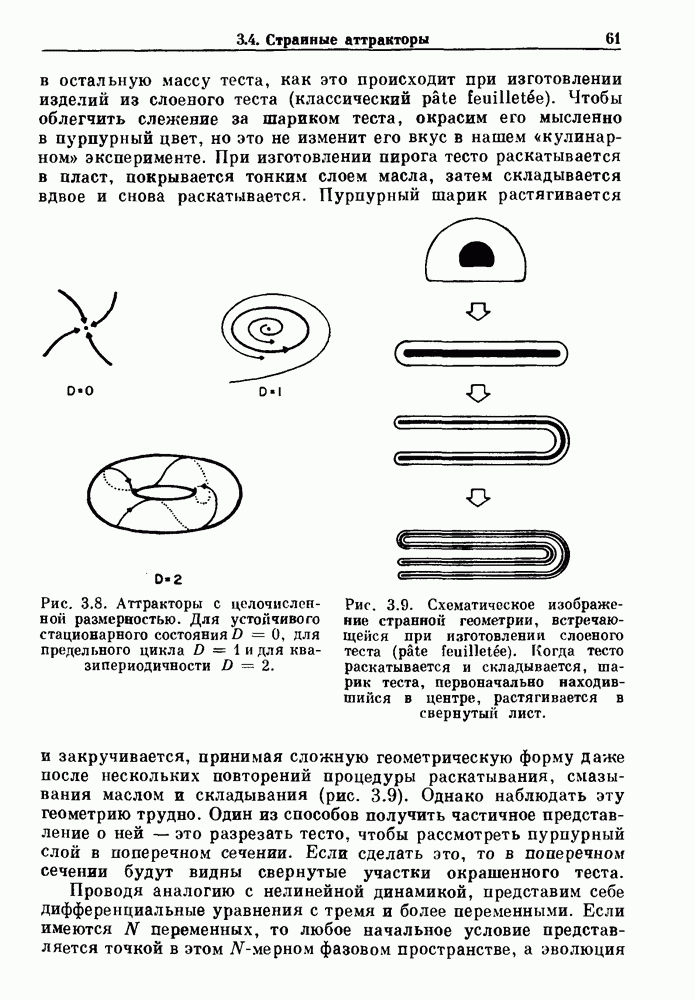









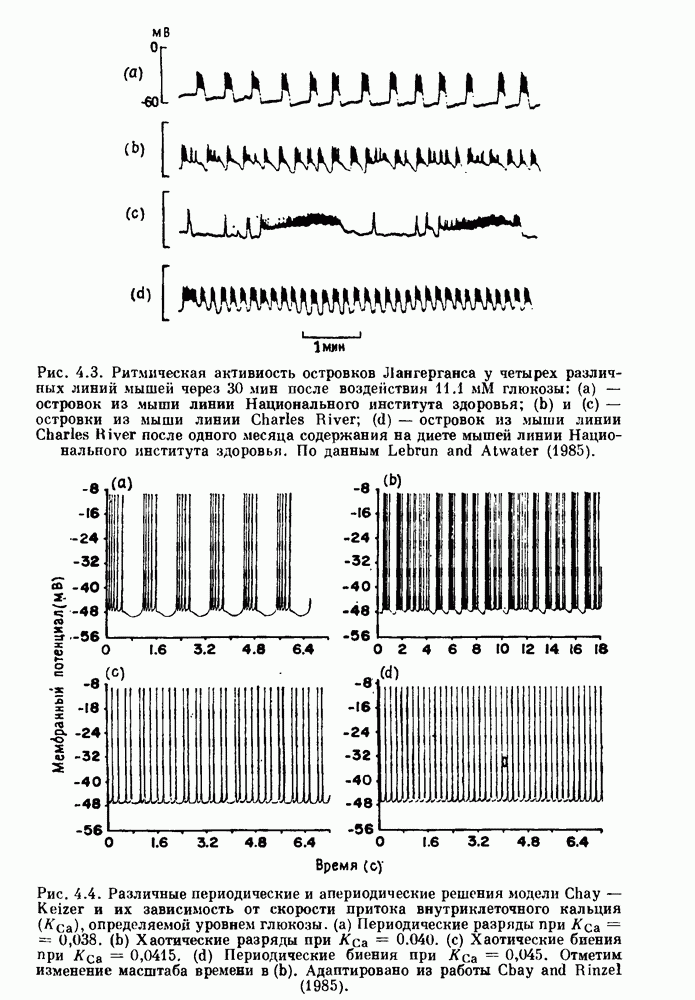























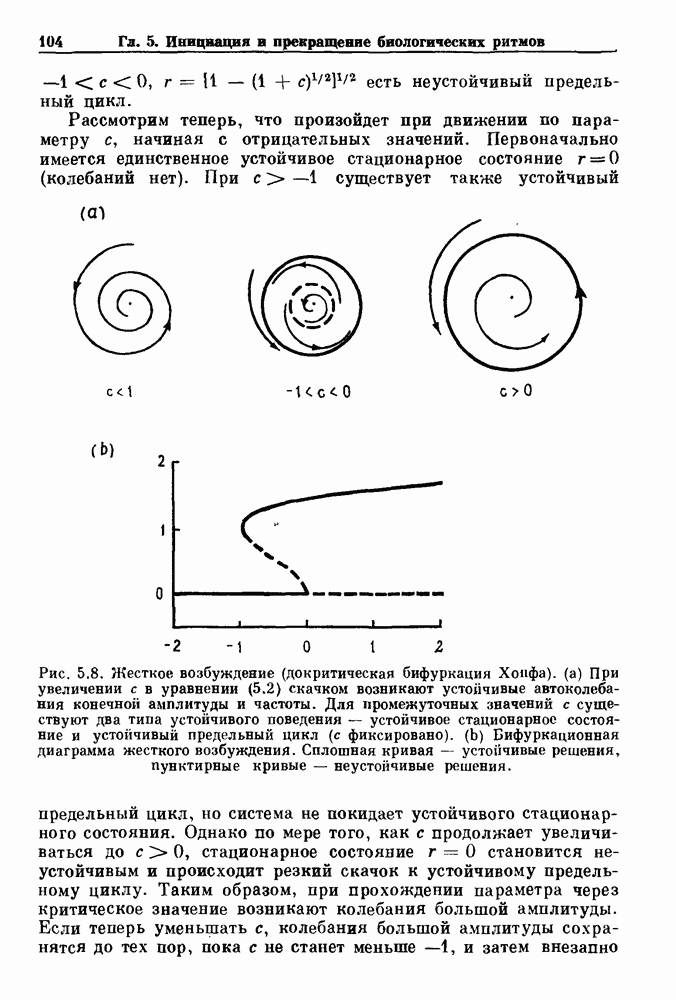
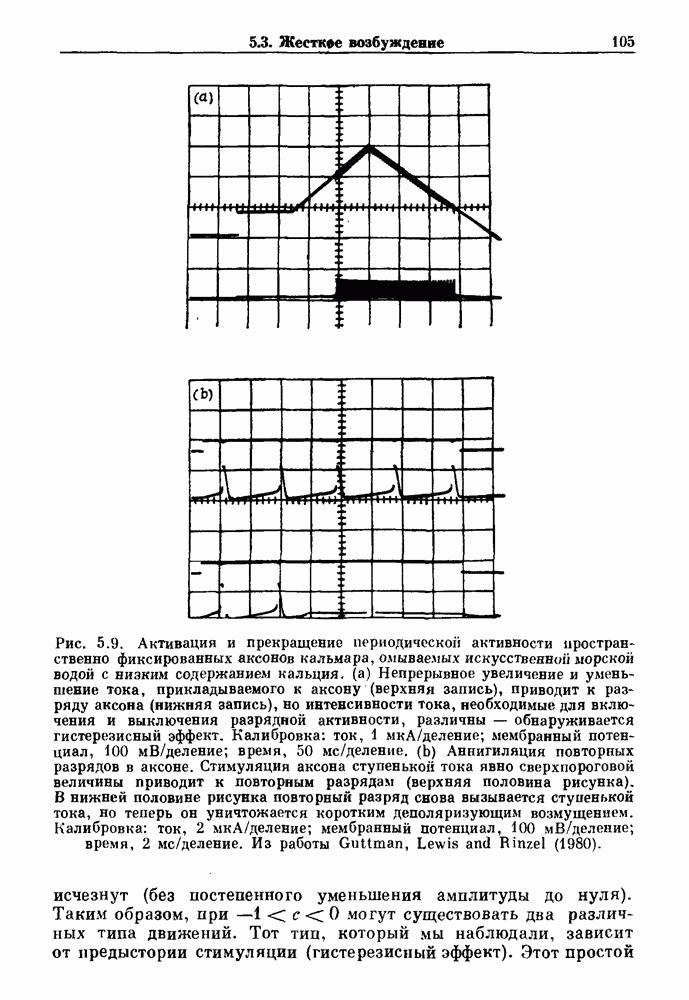








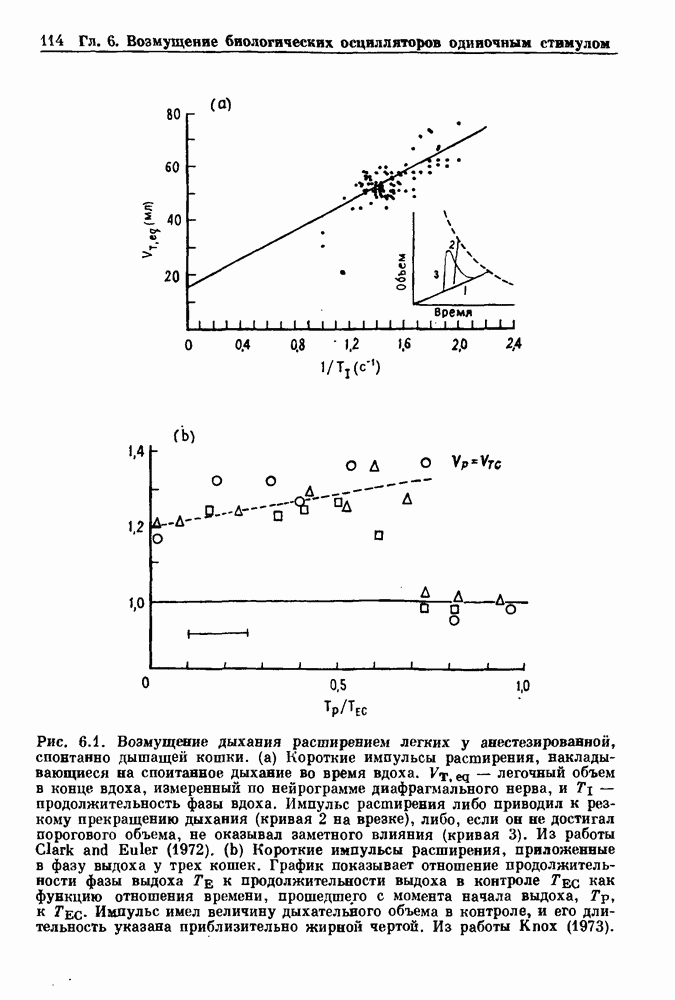
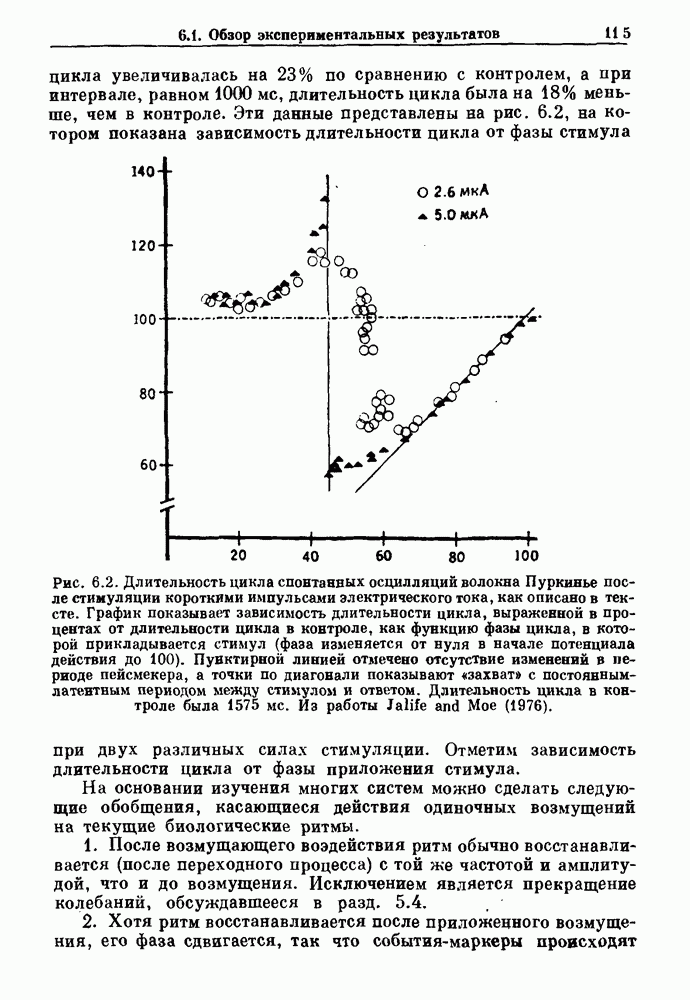

















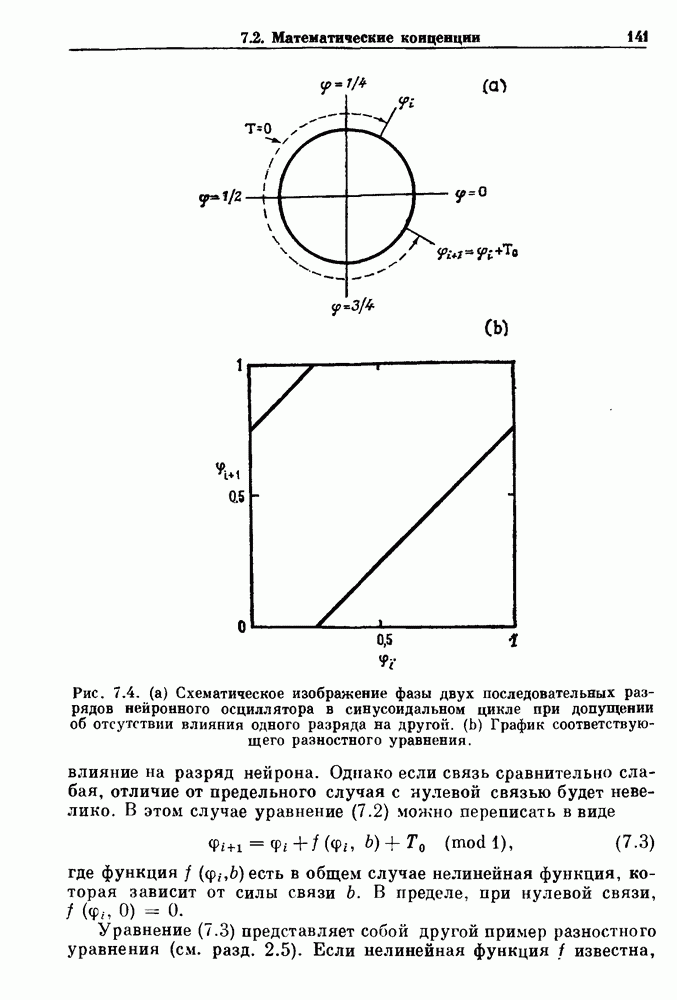



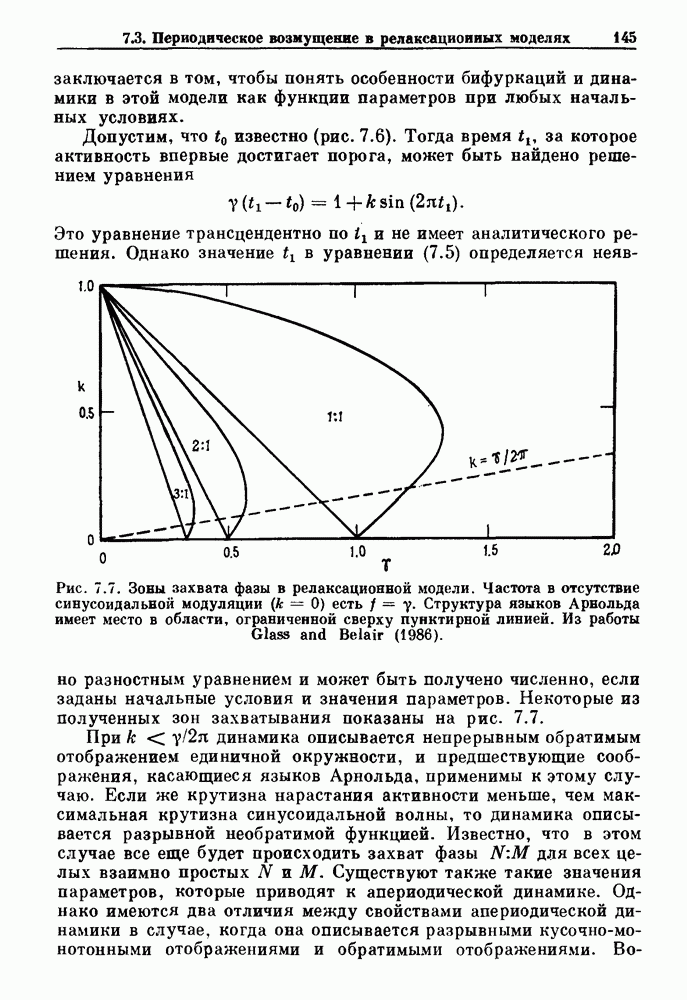
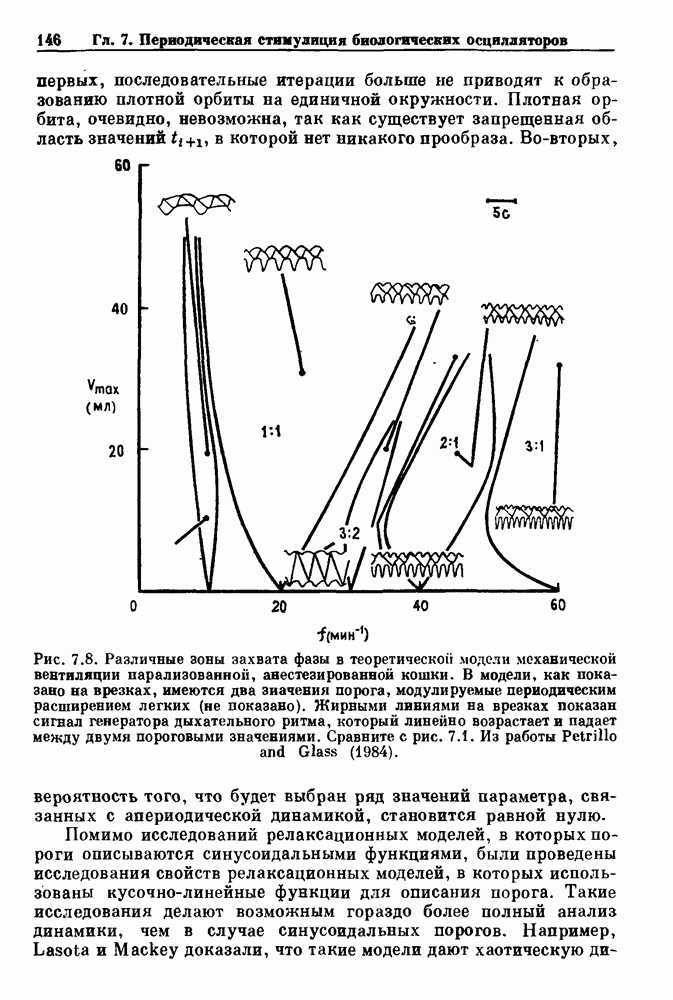


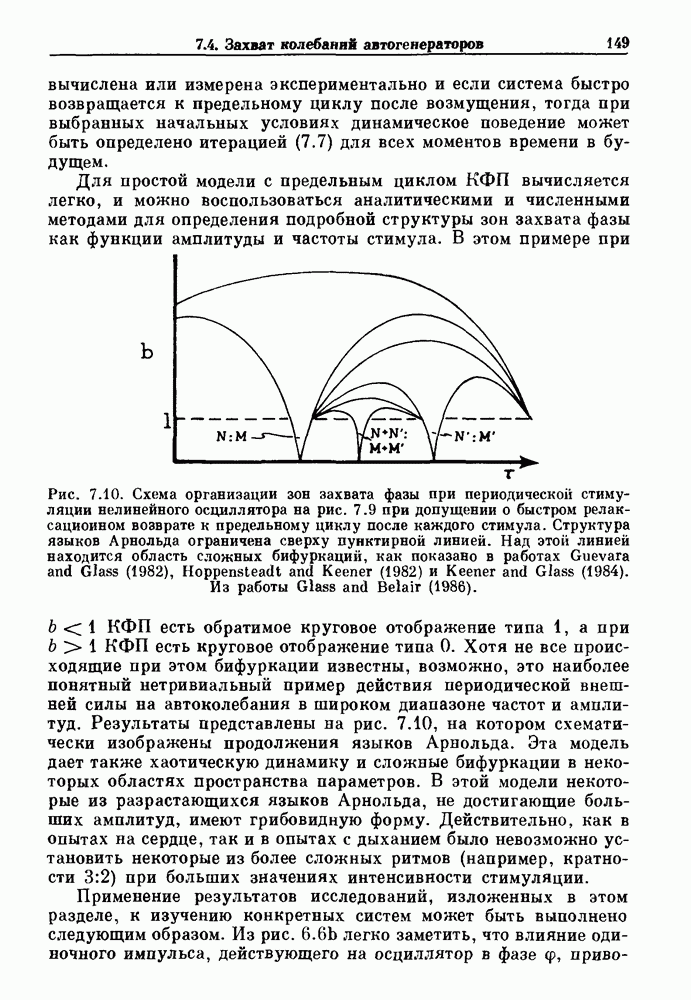
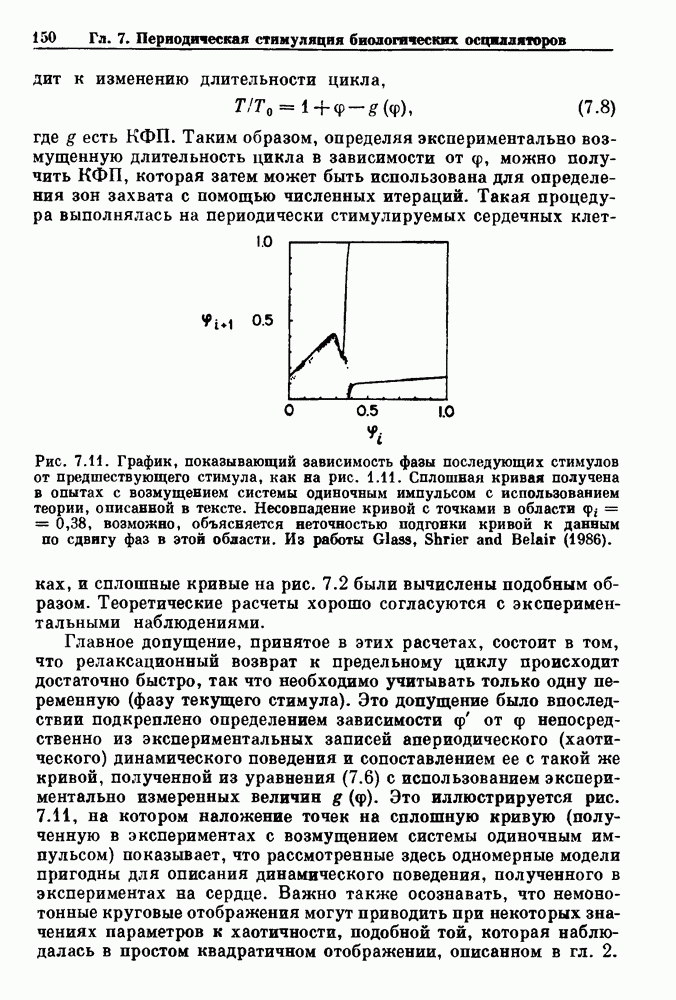


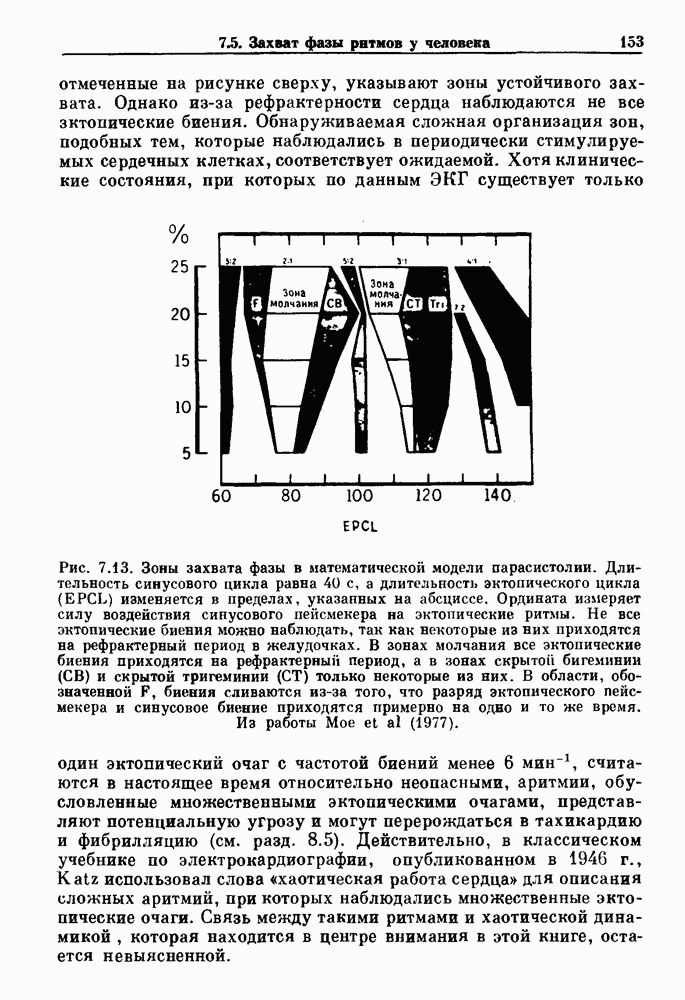
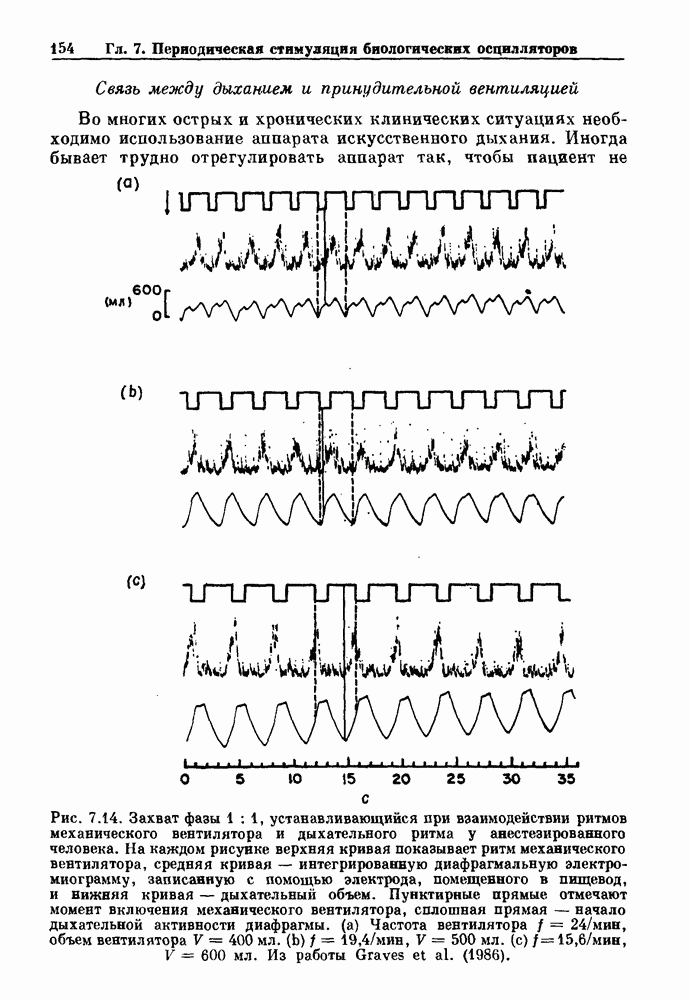
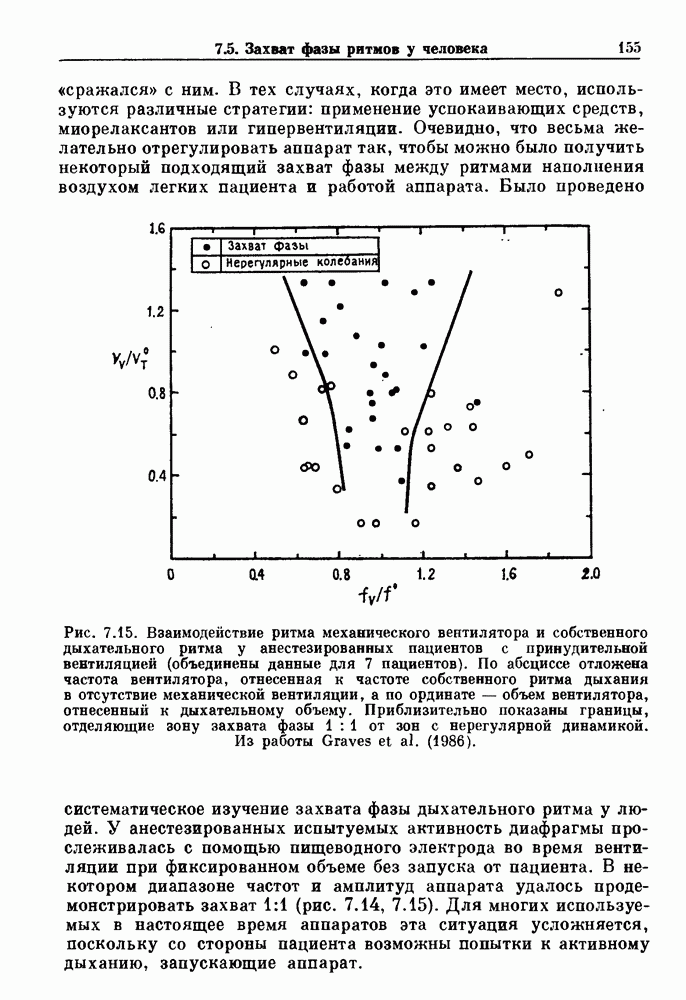







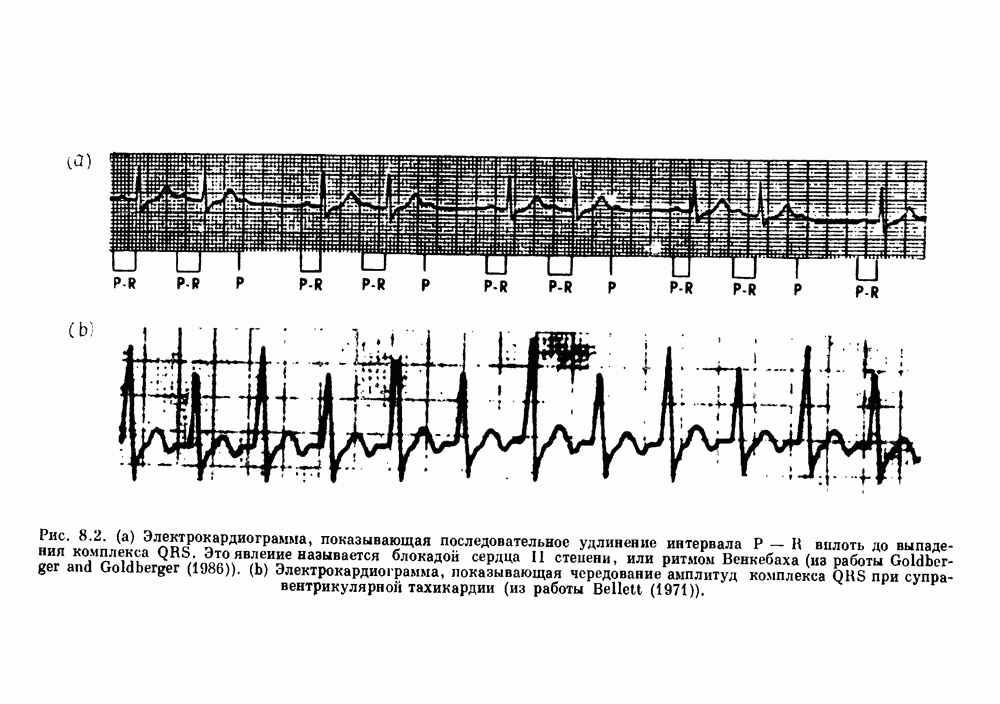


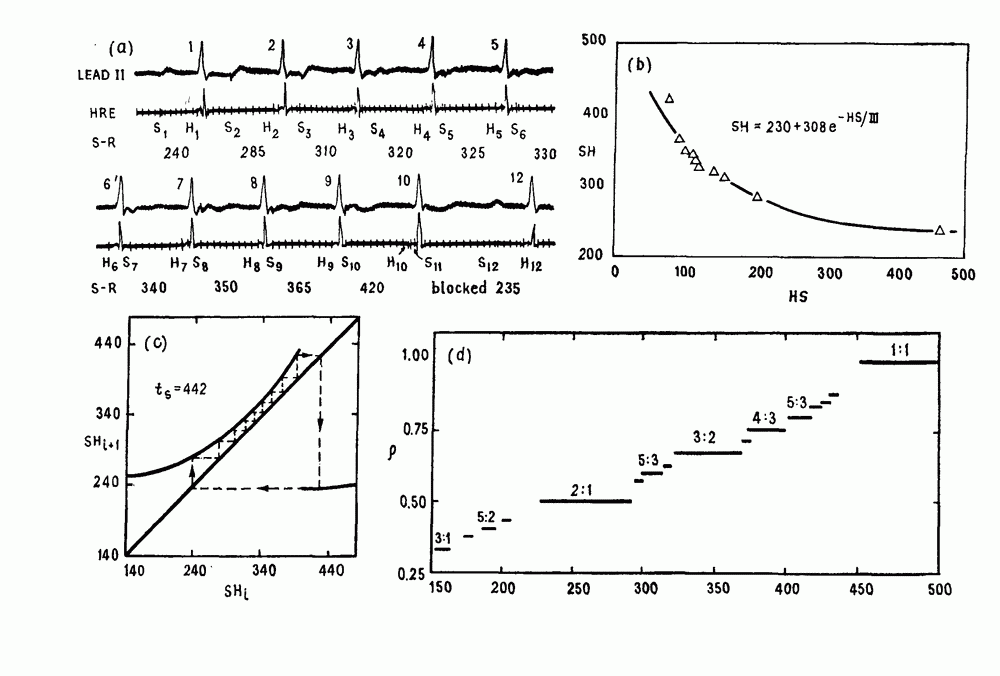

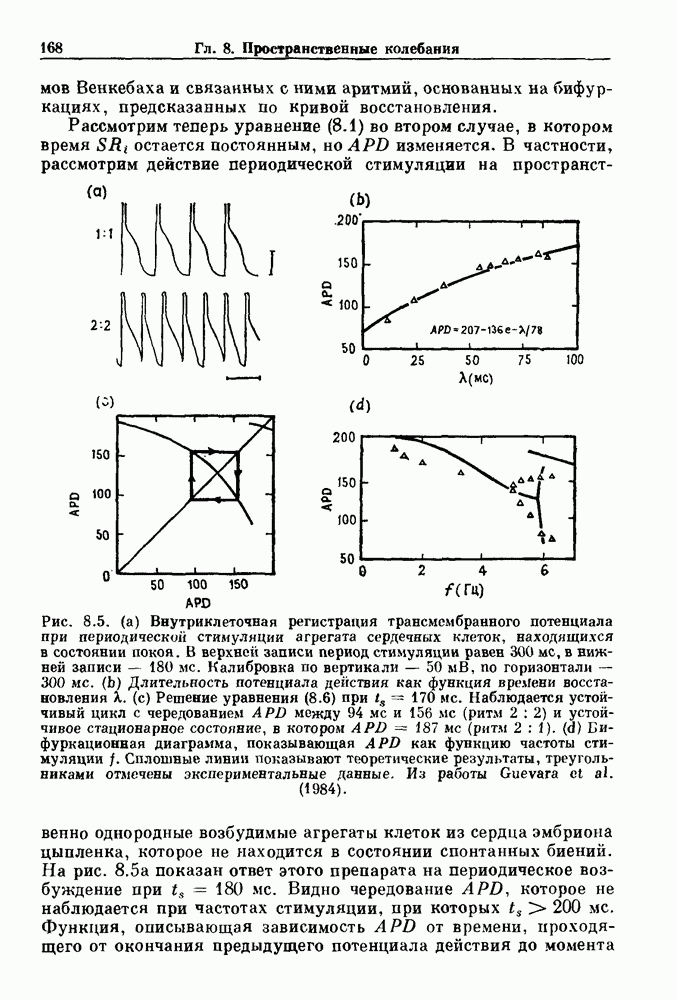

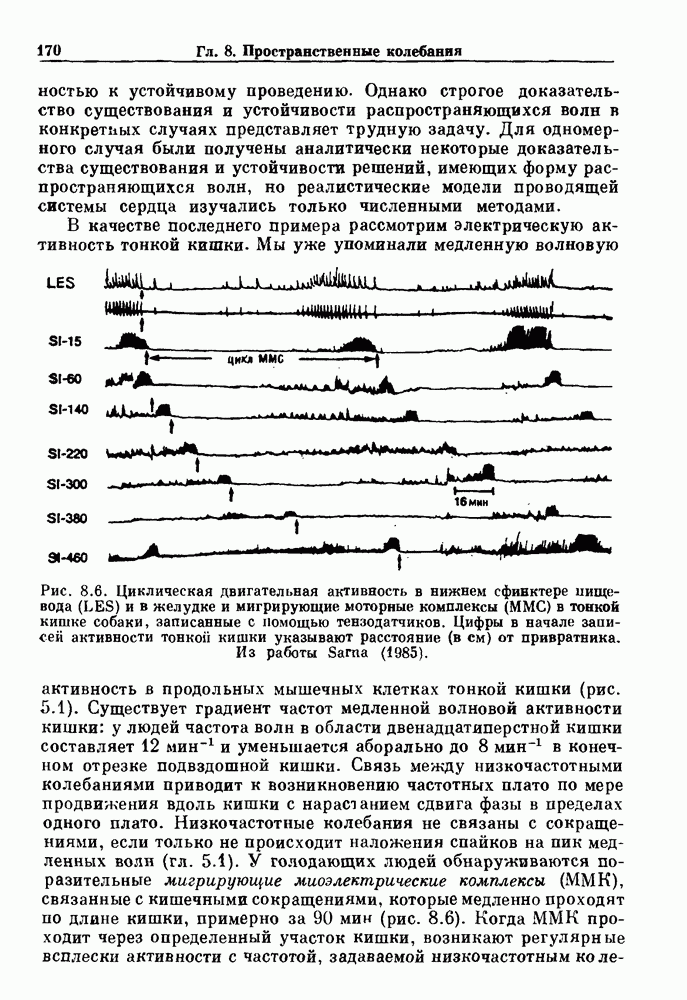







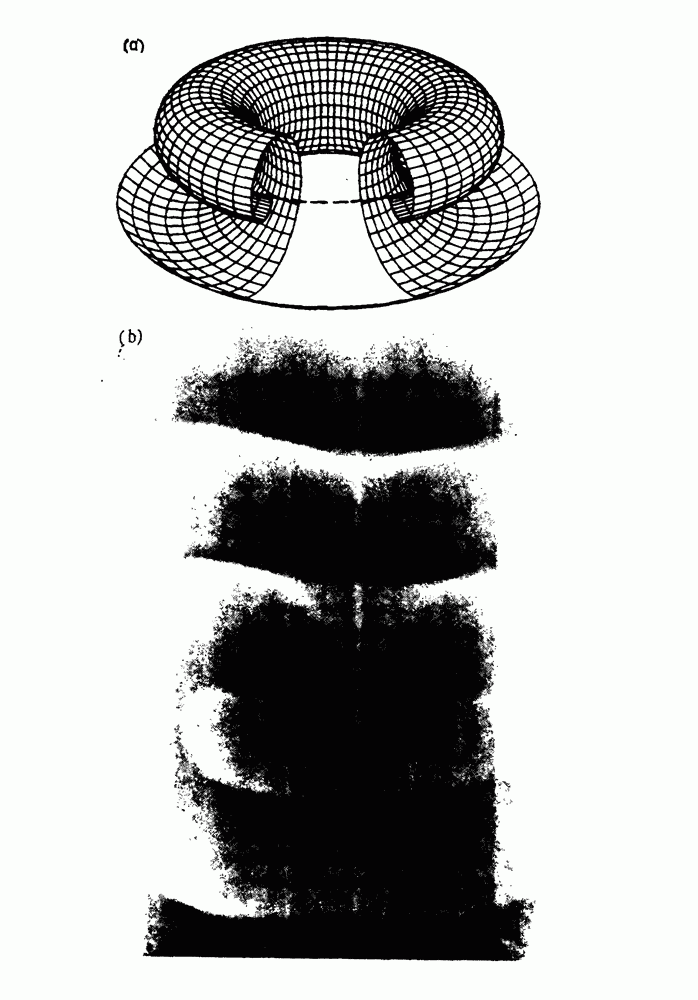

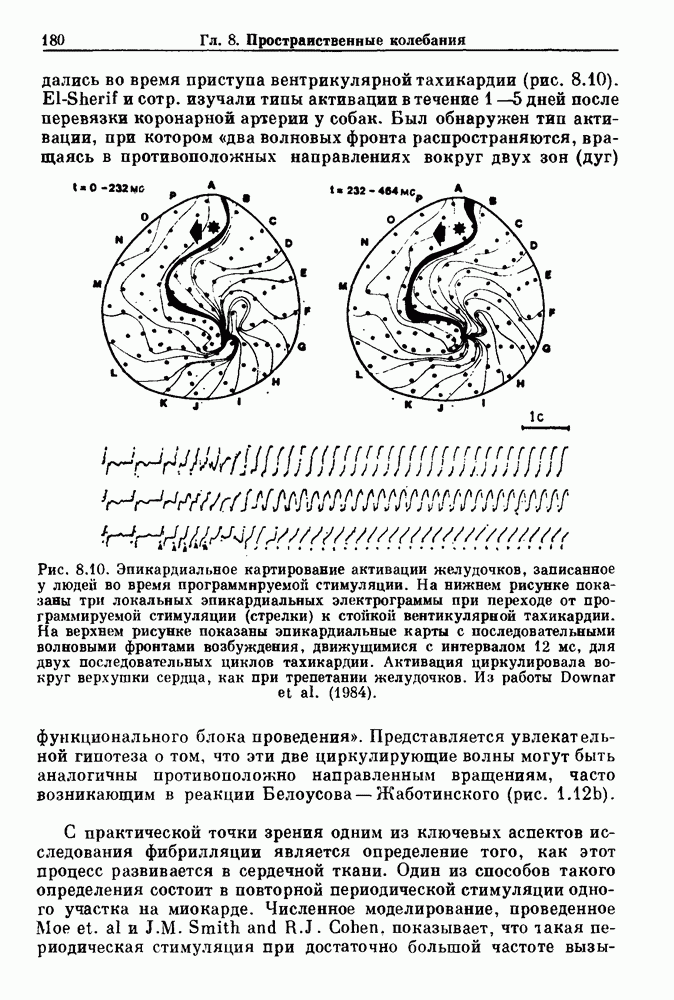


























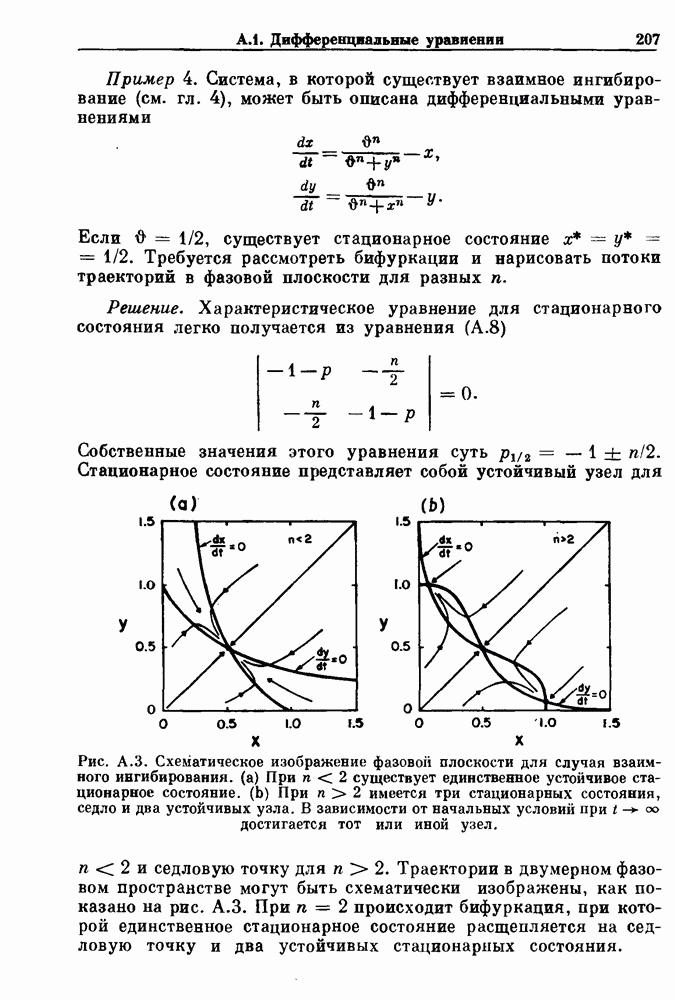
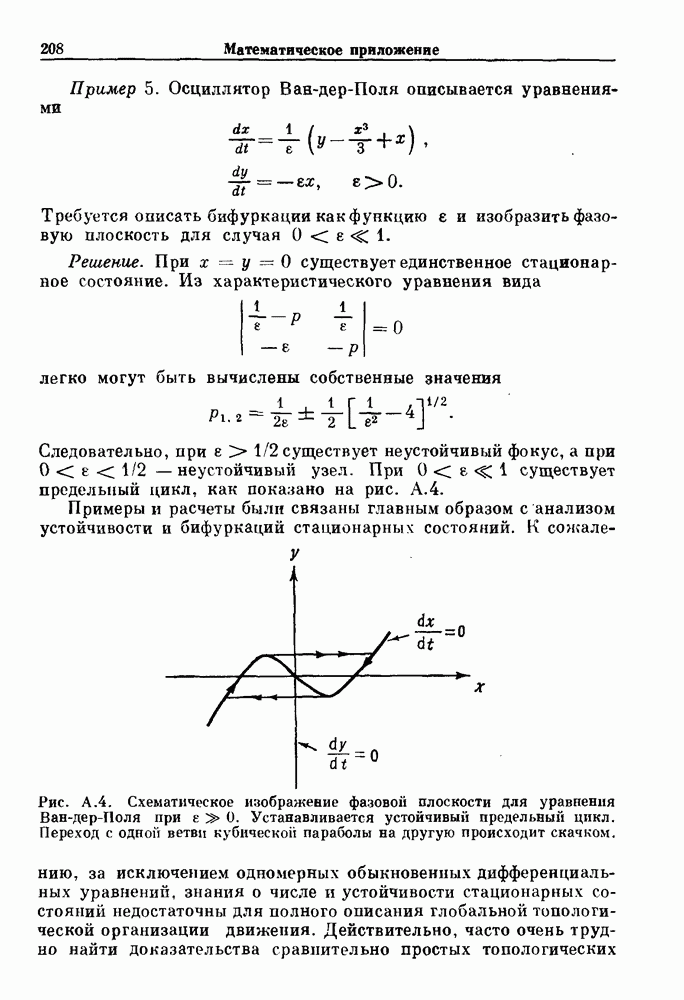



































 -клетках поджелудочной железы (Chay and Rinzel (1985)) и в клетках слизевика (Martiel and Goldbeter (1985) также обнаруживают хаос. По-видимому, единого общего механизма пейсмекерных колебаний не существует (Berridge and Rapp (1979); Noble (1983)).
-клетках поджелудочной железы (Chay and Rinzel (1985)) и в клетках слизевика (Martiel and Goldbeter (1985) также обнаруживают хаос. По-видимому, единого общего механизма пейсмекерных колебаний не существует (Berridge and Rapp (1979); Noble (1983)).
 и
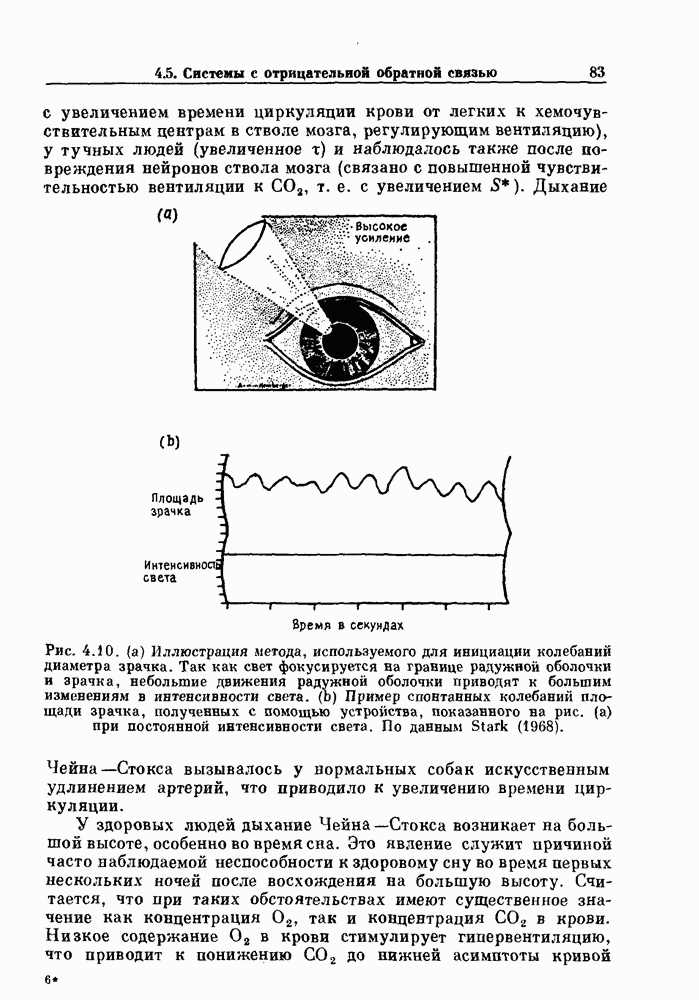
и  , но детальный теоретический анализ устойчивости в этих случаях обычно невозможен (Longobardo, Cherniack and Fishman (1966); Milhorn (1966); Khoo et al. (1982)). С медицинской точки зрения дыхание Чейна—Стокса может быть связано с неврологическими нарушениями или болезнями сердца (Dowell et al. (1971). (Lam-bersten (1974)). Дыхание Чейна — Стокса создавалось у собак путем искусственного увеличения задержки кровообращения после окси-генации крови (Guyton, Crowell and Moore (1956)). Обсуждения дыхания Чейна — Стокса, возникающего на большой высоте,
, но детальный теоретический анализ устойчивости в этих случаях обычно невозможен (Longobardo, Cherniack and Fishman (1966); Milhorn (1966); Khoo et al. (1982)). С медицинской точки зрения дыхание Чейна—Стокса может быть связано с неврологическими нарушениями или болезнями сердца (Dowell et al. (1971). (Lam-bersten (1974)). Дыхание Чейна — Стокса создавалось у собак путем искусственного увеличения задержки кровообращения после окси-генации крови (Guyton, Crowell and Moore (1956)). Обсуждения дыхания Чейна — Стокса, возникающего на большой высоте,