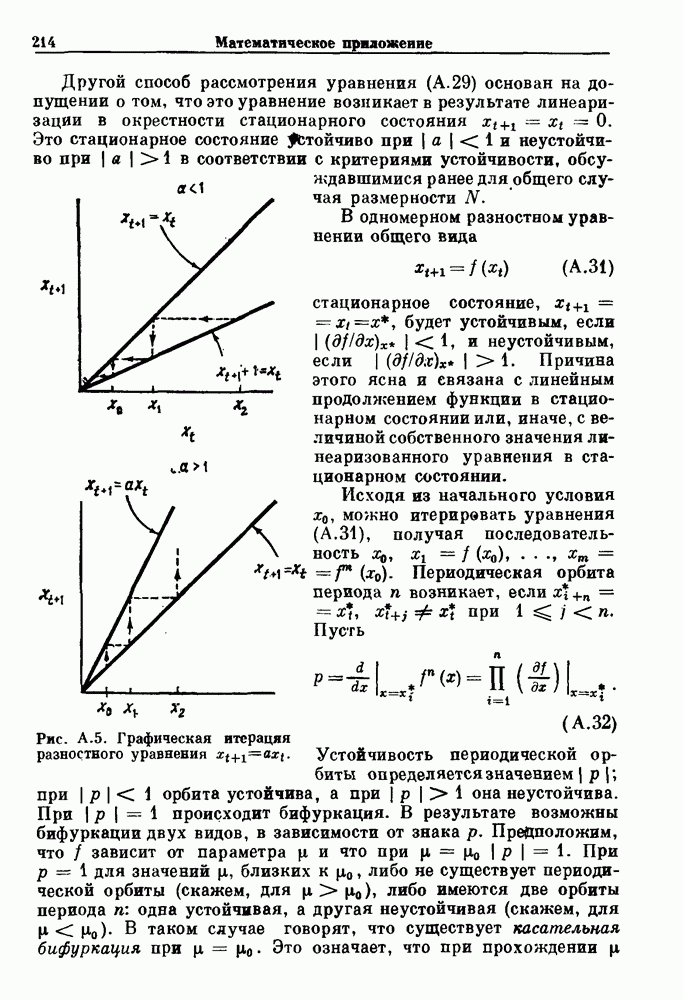
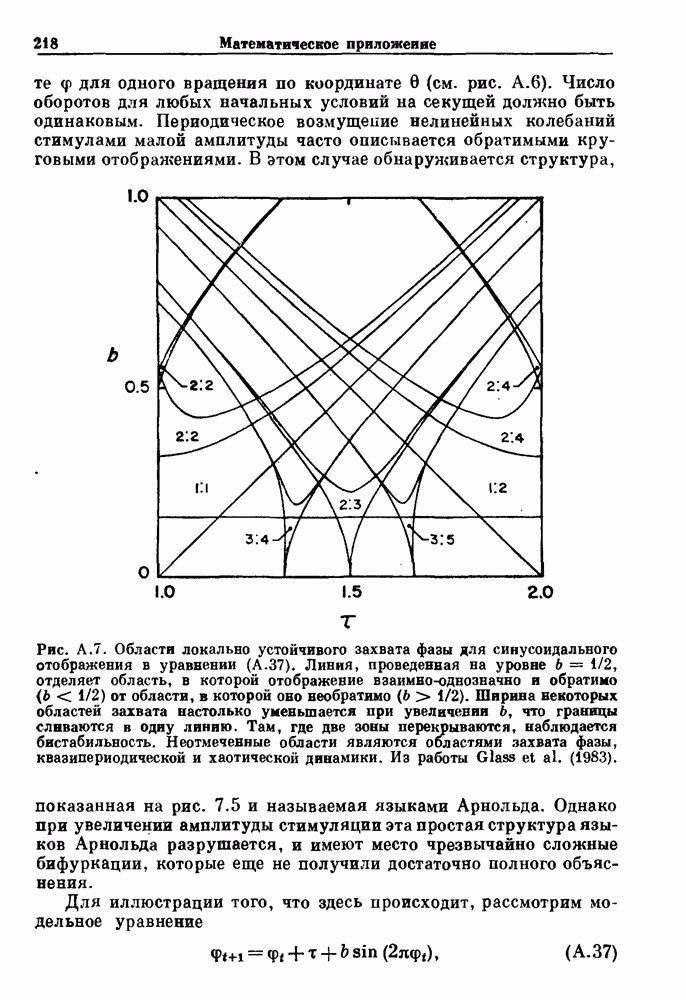
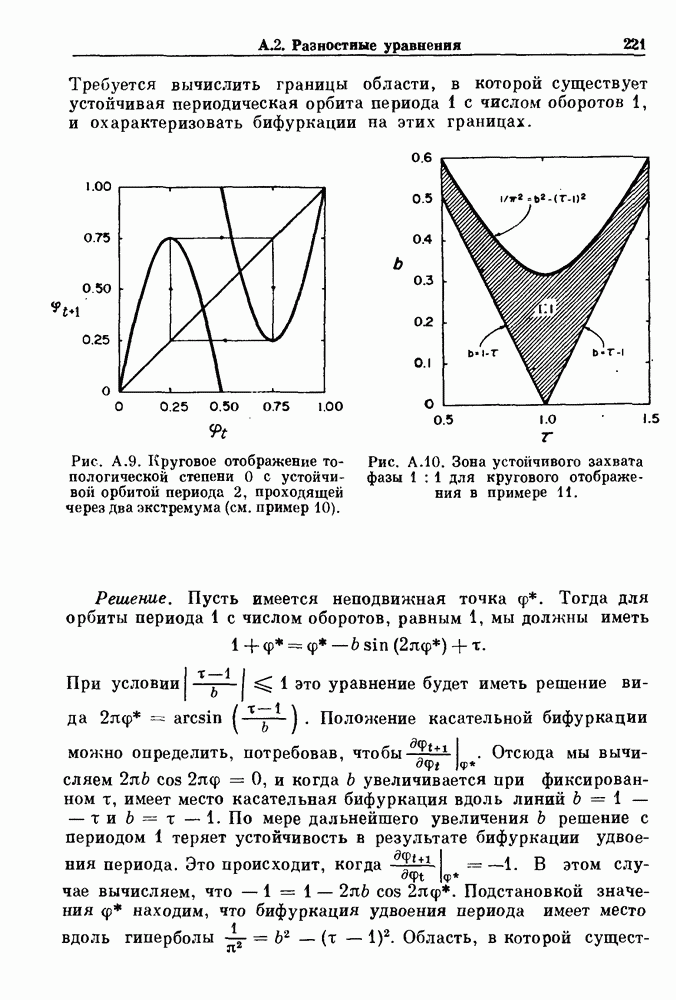
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
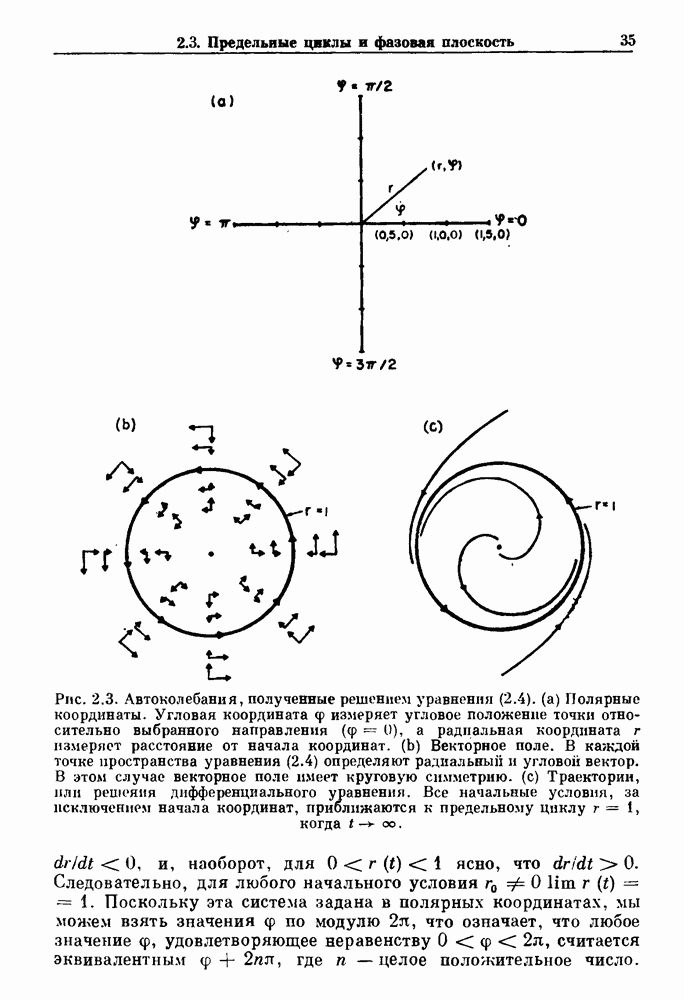
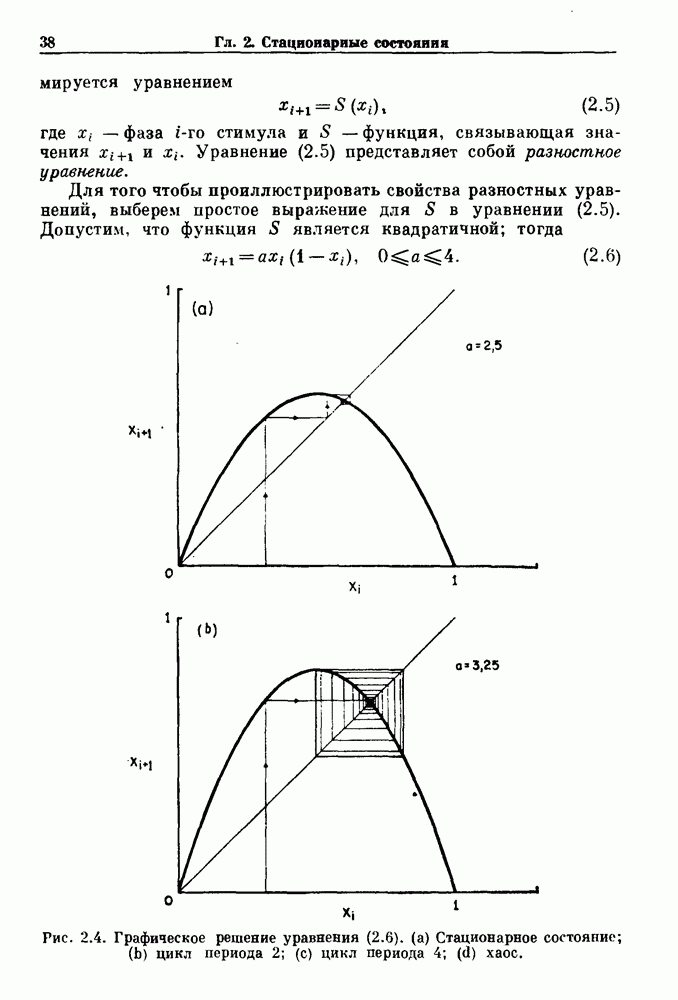
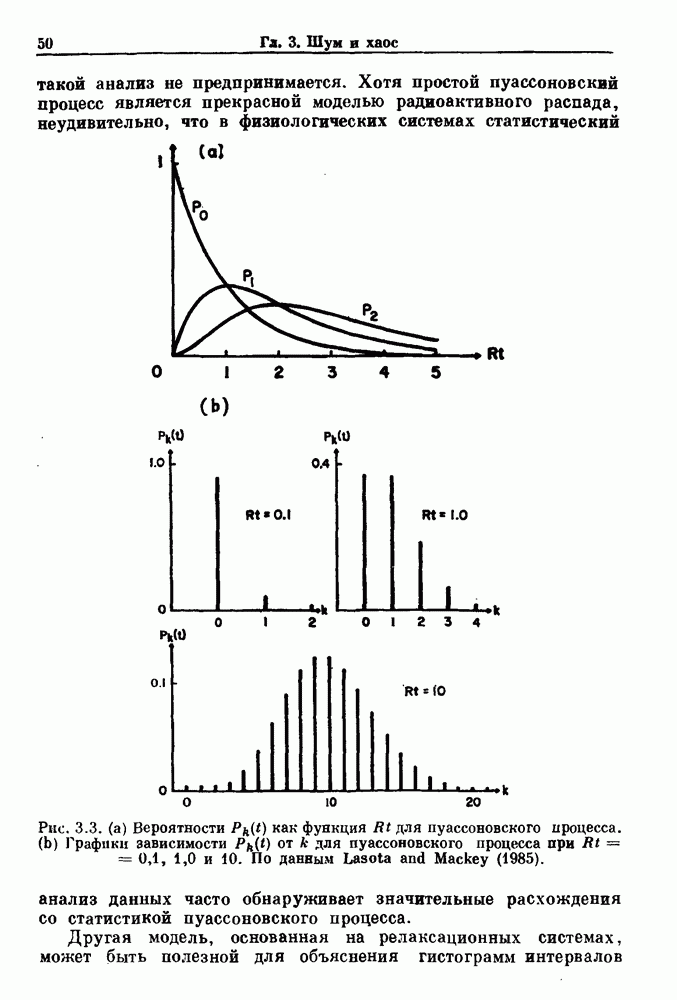
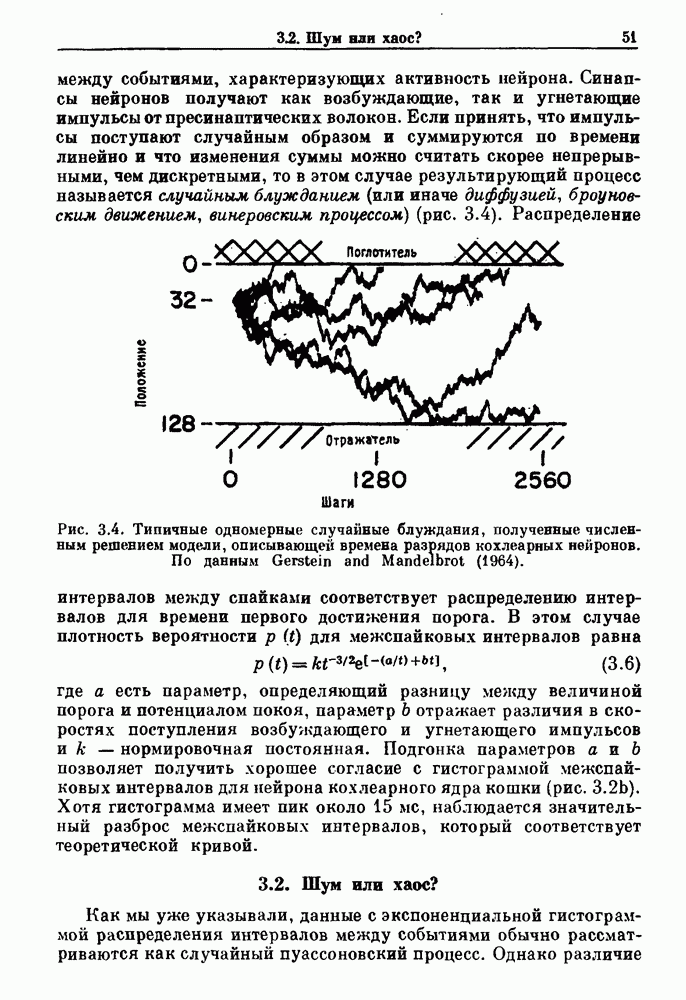
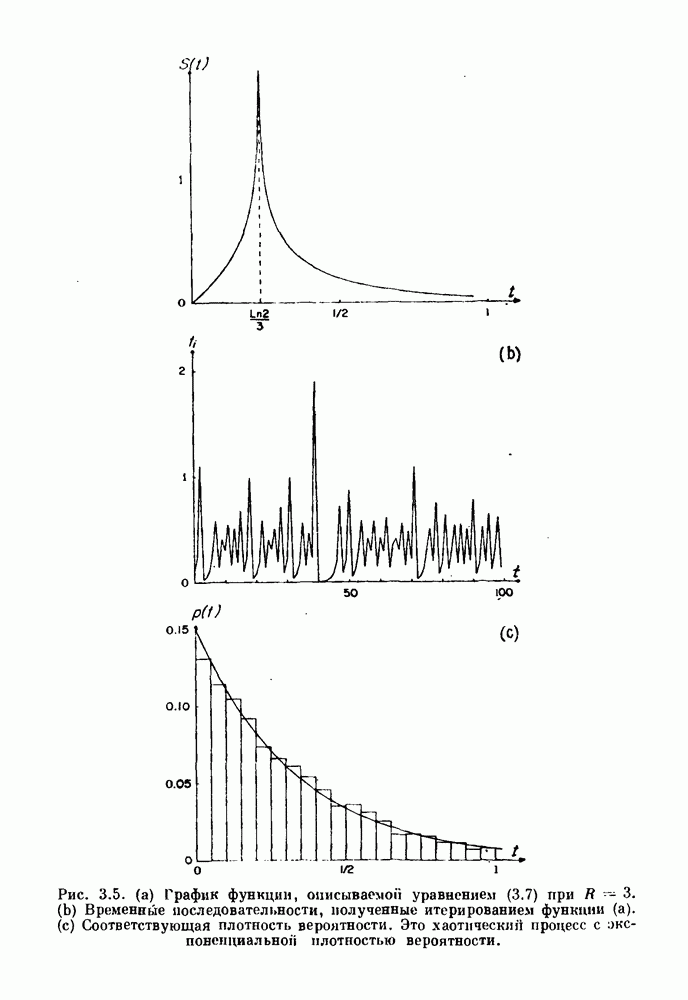
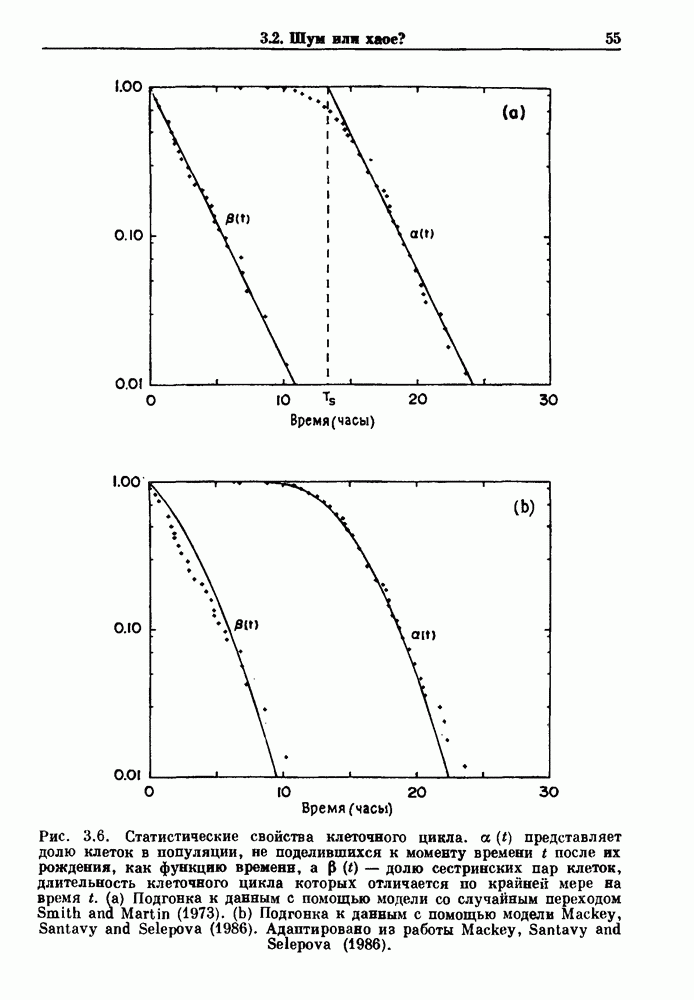
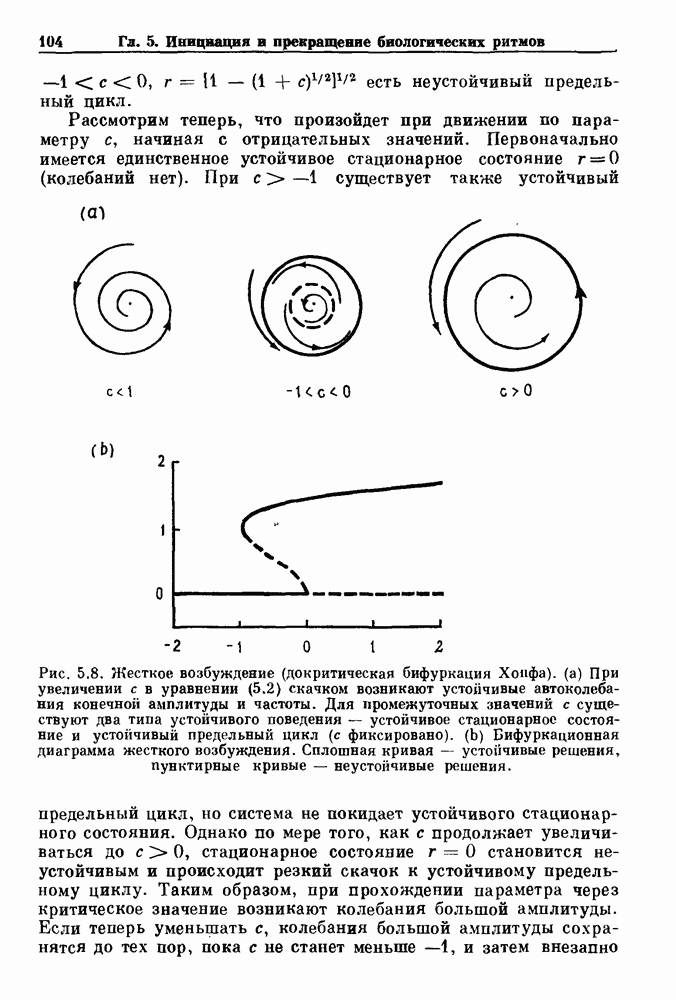
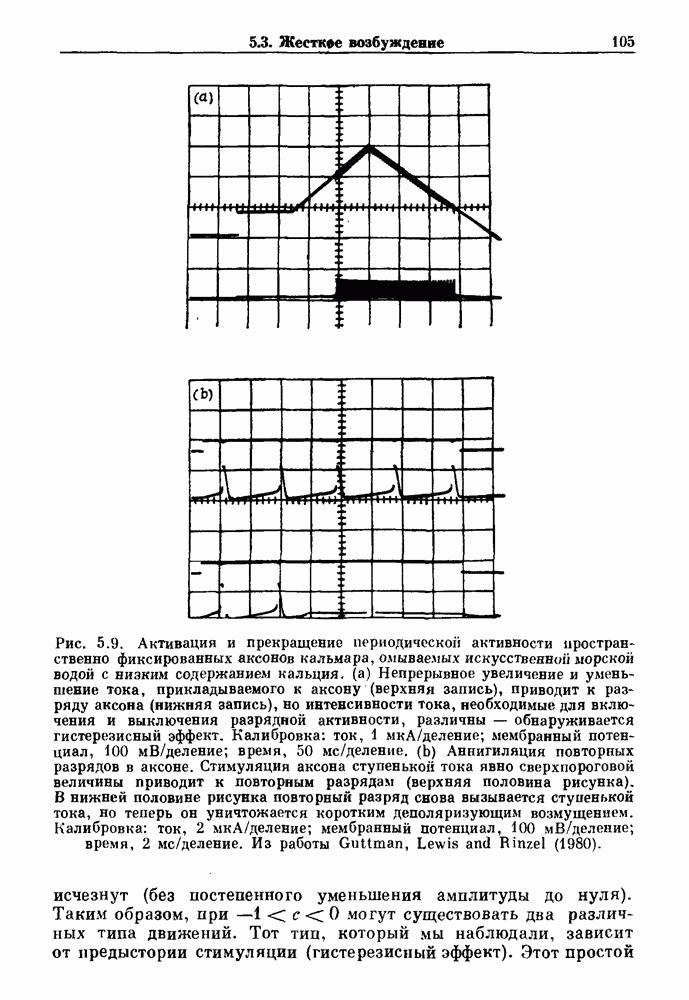
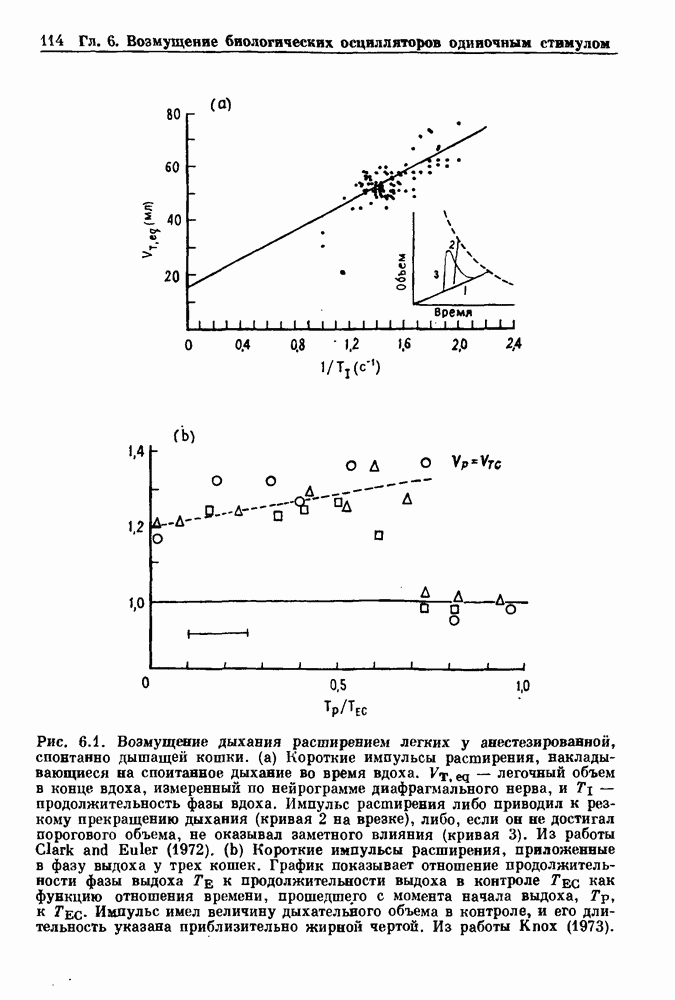
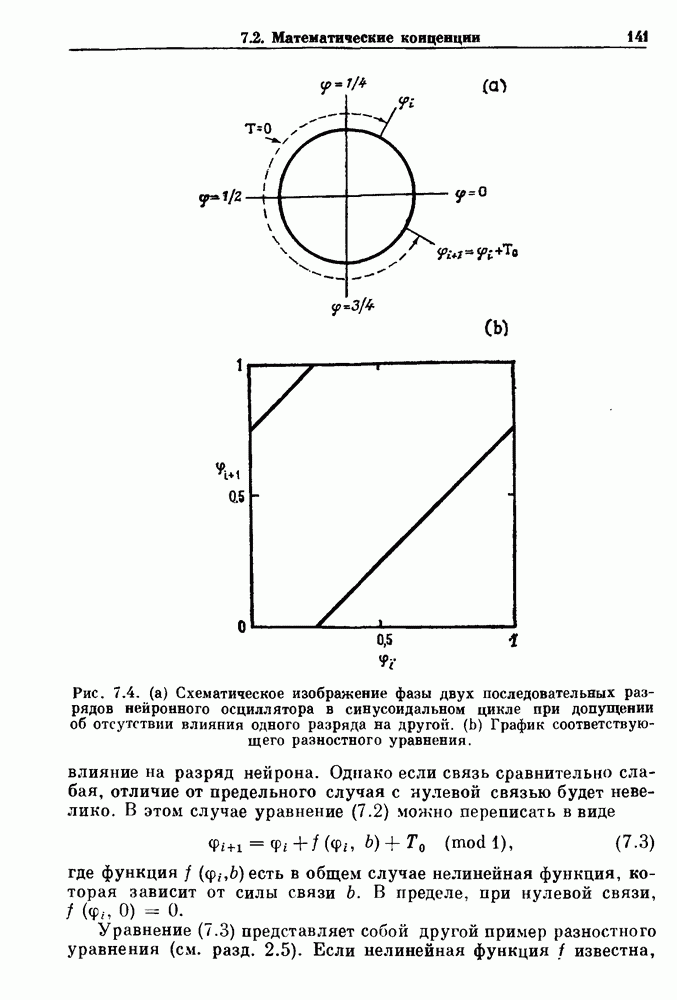
7.3. Периодическое возмущение в релаксационных моделяхПериодическая модуляция может быть включена в релаксационные модели двумя различными способами. Во-первых, можно принять, что периодически модулируется активность, но порог сохраняется постоянным.
Рис. 7.6. Релаксационная модель с синусоидально модулируемым порогом. После достижения порога происходит мгновенный возврат к нулю. Начиная с некоторого начального условия В альтернативном случае периодической модуляции может подвергаться порог. В дальнейшем мы будем обсуждать эффекты модуляции порога. Одним из побуждений к анализу периодического возмущения в релаксационных моделях послужили экспериментальные исследования механической вентиляции у кошек. Анализ осциллограмм на рис. 7.1, в частности ритмов 1:2 и 3:2, показывает, что сильное расширение легких совпадает с преждевременным прекращением вдоха. Это служит удивительно наглядной иллюстрацией применимости к этой системе релаксационной модели с модуляцией порога. Как мы уже обсуждали в гл. 6, рефлексы Геринга—Брейера могут моделироваться с помощью релаксационных моделей. Рассмотрим простую релаксационную модель с периодически модулируемым порогом (рис. 7.6). Для конкретности и простоты допустим, что порог описывается синусоидальной функцией
Допустим, что активность увеличивается линейно, с крутизной 7, до пороговой величины и затем устанавливается в 0. Наша цель заключается в том, чтобы понять особенности бифуркаций и динамики в этой модели как функции параметров при любых начальных условиях. Допустим, что
Это уравнение трансцендентно по 11 и не имеет аналитического решения.
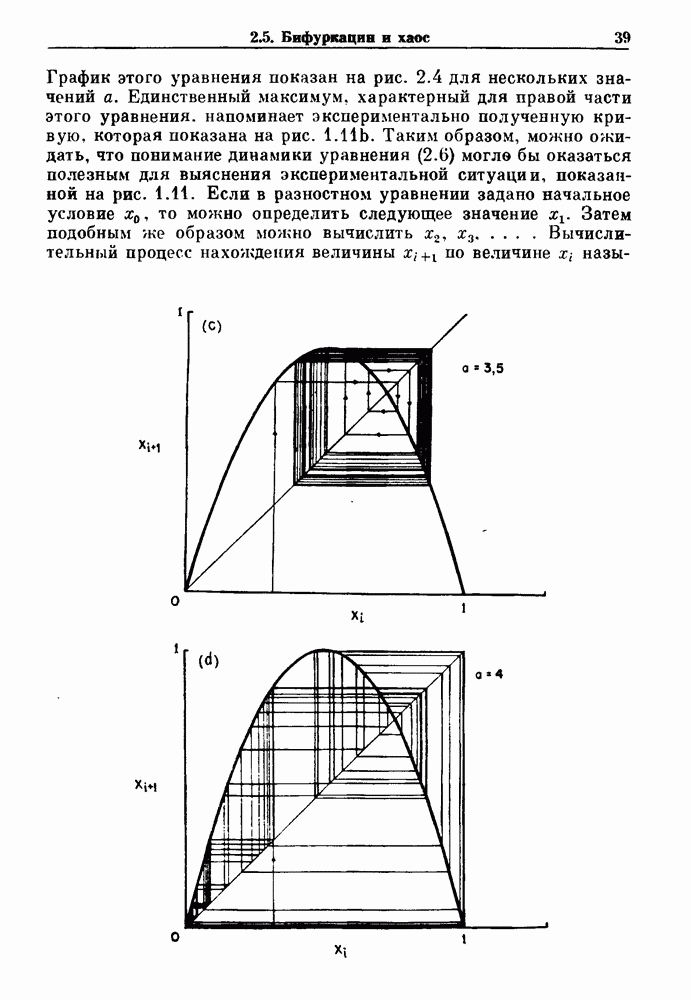
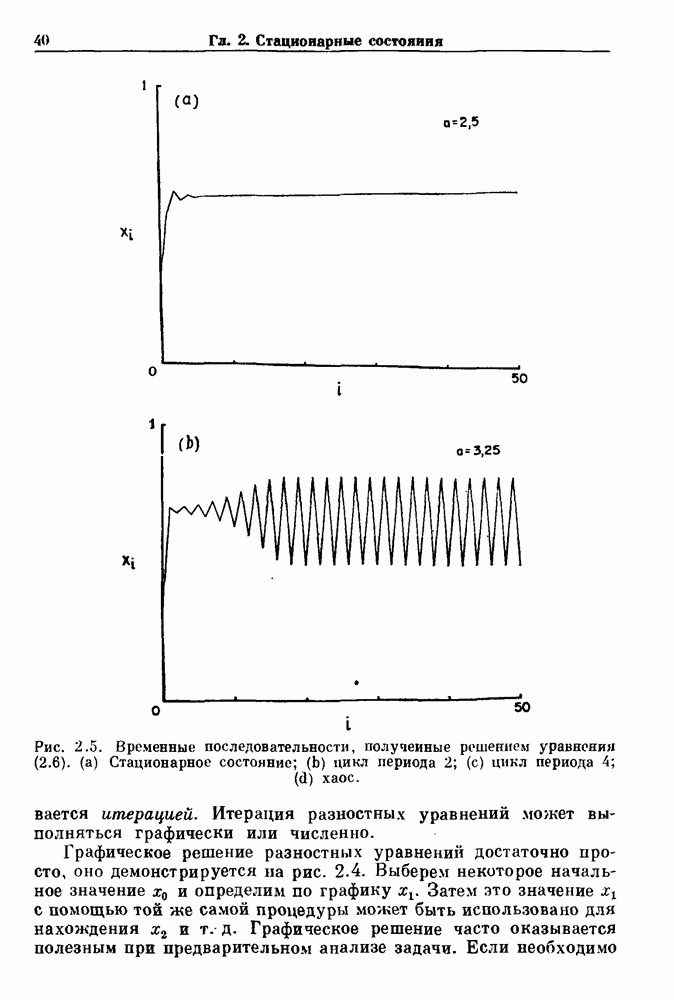
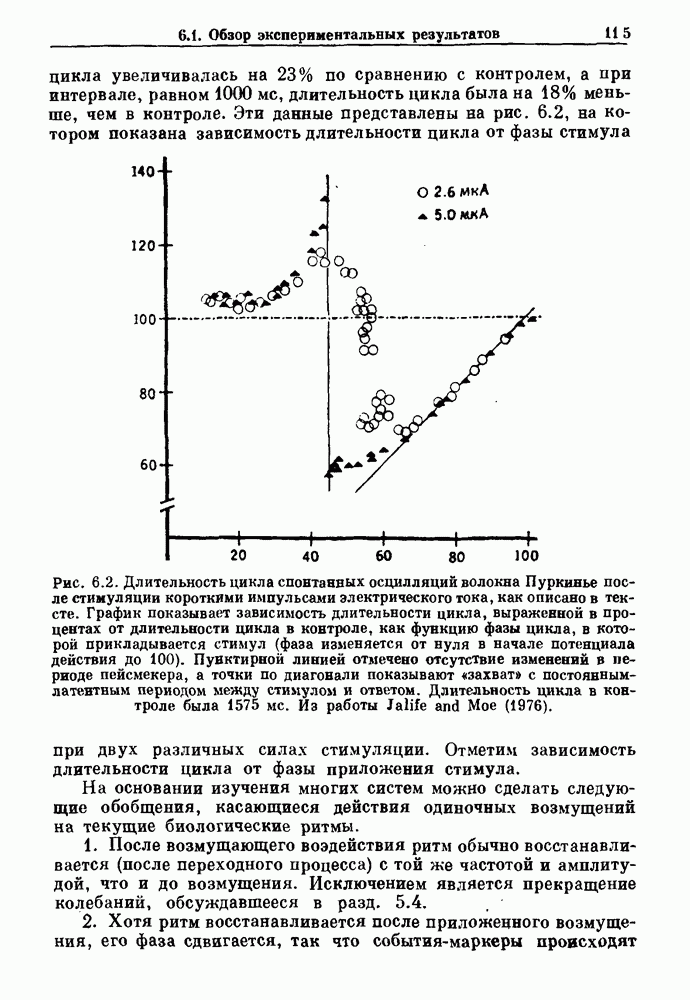
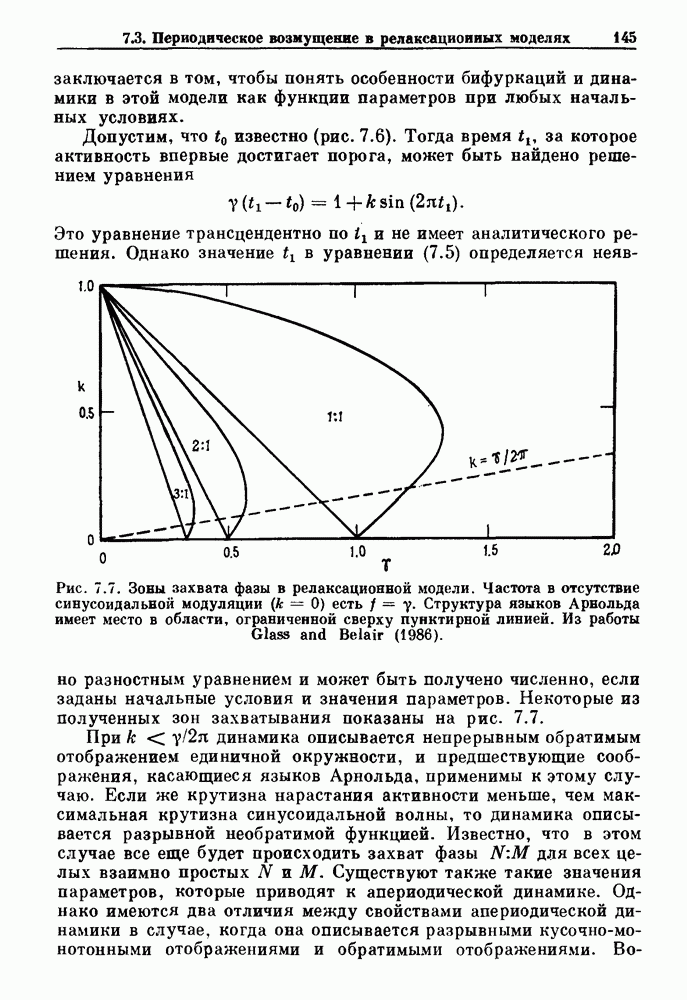
Рис. 7.7. Зоны захвата фазы в релаксационной модели. Частота в отсутствие синусоидальной модуляции Однако значение в уравнении (7.5) определяется неявно разностным уравнением и может быть получено численно, если заданы начальные условия и значения параметров. Некоторые из полученных зон захватывания показаны на рис. 7.7. При Во-первых, последовательные итерации больше не приводят к образованию плотной орбиты на единичной окружности. Плотная орбита, очевидно, невозможна, так как существует запрещенная область значений +1, в которой нет никакого прообраза.
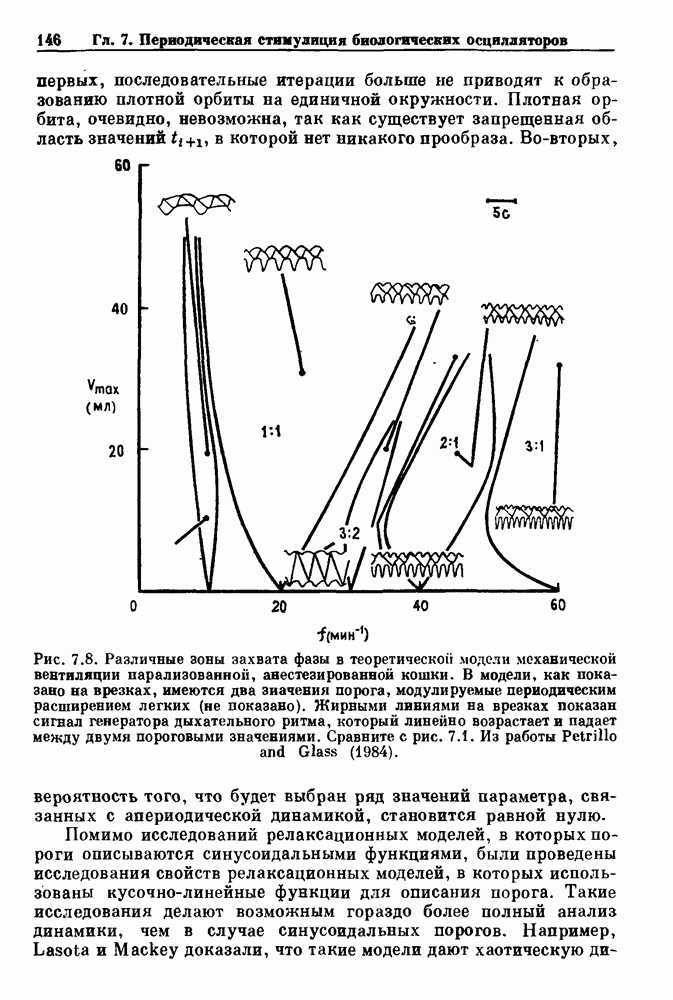
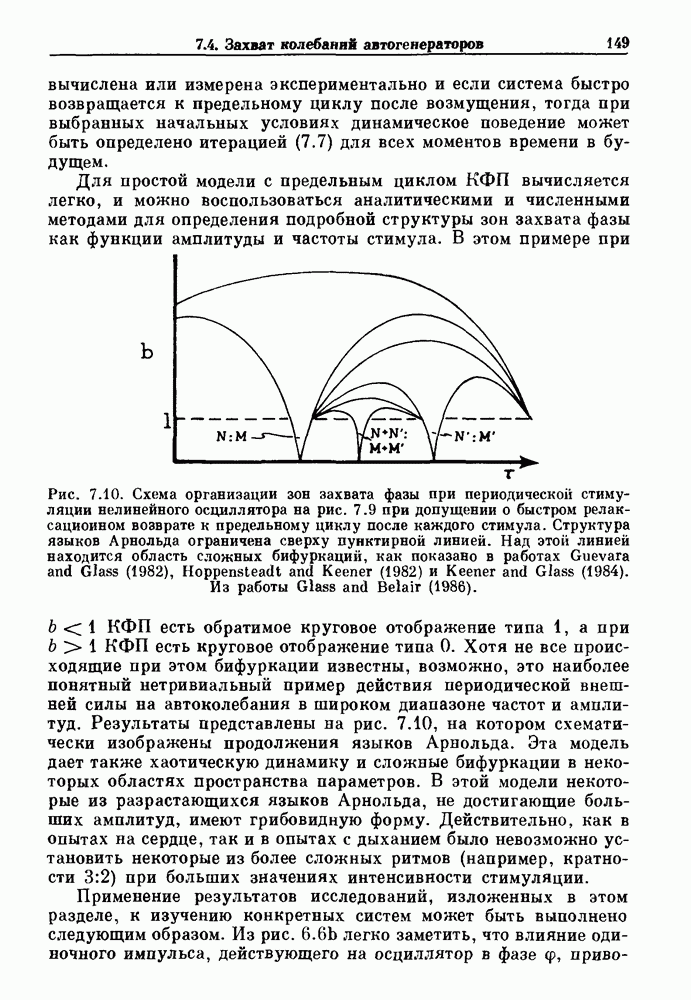
Рис. 7.8. Различные зоны захвата фазы в теоретической модели механической вентиляции парализованной, анестезированной кошки. В модели, как показано на врезках, имеются два значения порога, модулируемые периодическим расширением легких (не показано). Жирными линиями на врезках показан сигнал генератора дыхательного ритма, который линейно возрастает и падает между двумя пороговыми значениями. Сравните с рис. 7.1. Из работы Petrillo and Glass (1984). Во-вторых, вероятность того, что будет выбран ряд значений параметра, связанных с апериодической динамикой, становится равной нулю-Помимо исследований релаксационных моделей, в которых пороги описываются синусоидальными функциями, были проведены исследования свойств релаксационных моделей, в которых использованы кусочно-линейные функции для описания порога. Такие исследования делают возможным гораздо более полный анализ динамики, чем в случае синусоидальных порогов. Например, Lasota и Mackey доказали, что такие модели дают хаотическую динамику в некоторых областях значений параметров. Значение этого открытия заключается в возможности наблюдения хаоса даже в чрезмерно упрощенных моделях периодического возмущения колебаний. На основе этого открытия можно предположить, что появление хаотической динамики, по крайней мере в некоторой области параметров стимуляции, может быть очень общим явлением для периодически возмущаемых физиологических осцилляторов. Petrillo и Glass попытались построить более реалистичные релаксационные модели для описания захвата дыхательного ритма. Они предположили, что два порога модулируются объемом механического вентилятора. Продолжительность вдоха и выдоха представлена активностью, осциллирующей между двумя пороговыми уровнями. Существует некоторая задержка между моментом достижения порога вдоха и началом вдоха. Эта модель имеет пять параметров, которые могут быть получены из экспериментальных данных. Численные исследования модели (рис. 7.8) дают хорошее согласие с результатами экспериментов, показанными на рис. 7.1. Однако подробное изучение бифуркаций в этой модели не проводилось. Другая попытка построения релаксационных моделей для описания захвата биологических ритмов была предпринята Daan и сотр. для случая циркадианных ритмов. Авторы допускали существование синусоидально модулируемых порогов. С помощью такой модели они могли дать частичное объяснение зависимости продолжительности сна от времени засыпания. Систематический анализ зон захвата фазы 1:1 (соответствующих нормальному циркадианному ритму), а также других возможных зон захвата в зависимости от значений параметров не проводился. В заключение отметим, что периодически возмущаемые модели дают концептуально простое средство моделирования захвата фазы в биологических системах. Даже самые простые модели способны генерировать чрезвычайно сложные формы динамического поведения, которые еще не полностью объяснены.
|
 |
Оглавление
|















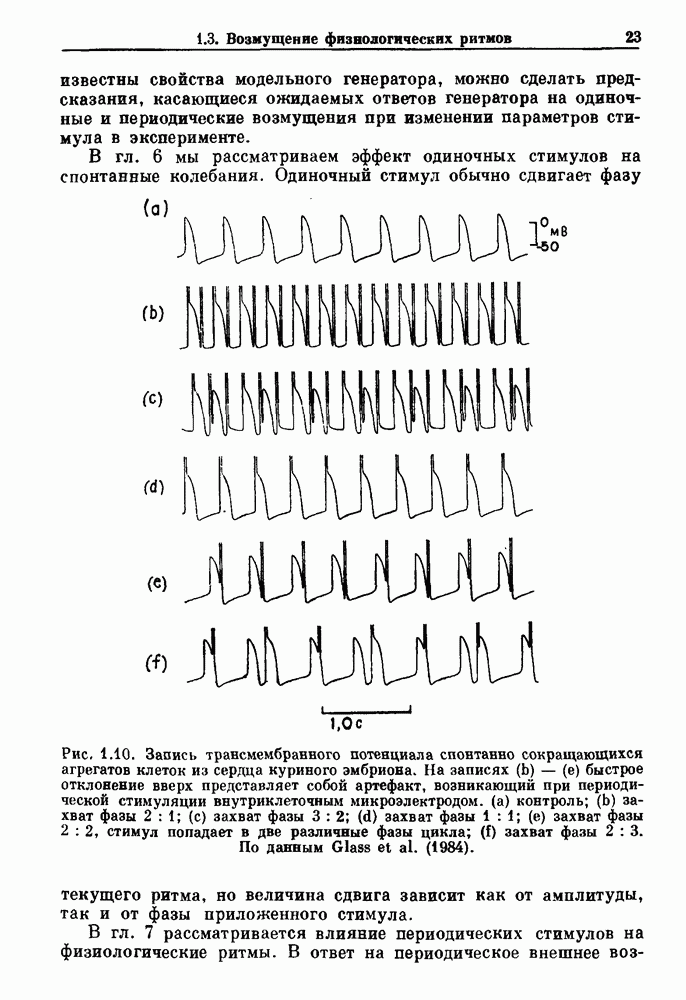



















































































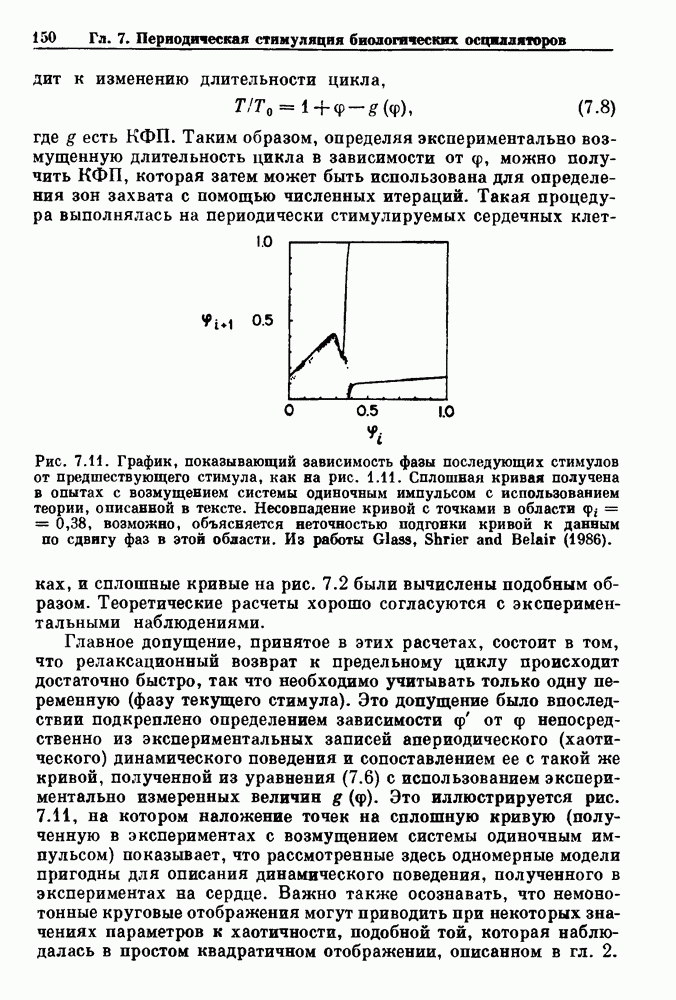



















































































 порог впервые достигается в момент времени
порог впервые достигается в момент времени  который может быть вычислен из уравнения (7.5). Показан захват фазы
который может быть вычислен из уравнения (7.5). Показан захват фазы  Адаптировано из работы Glass and Mackey (1979b).
Адаптировано из работы Glass and Mackey (1979b).

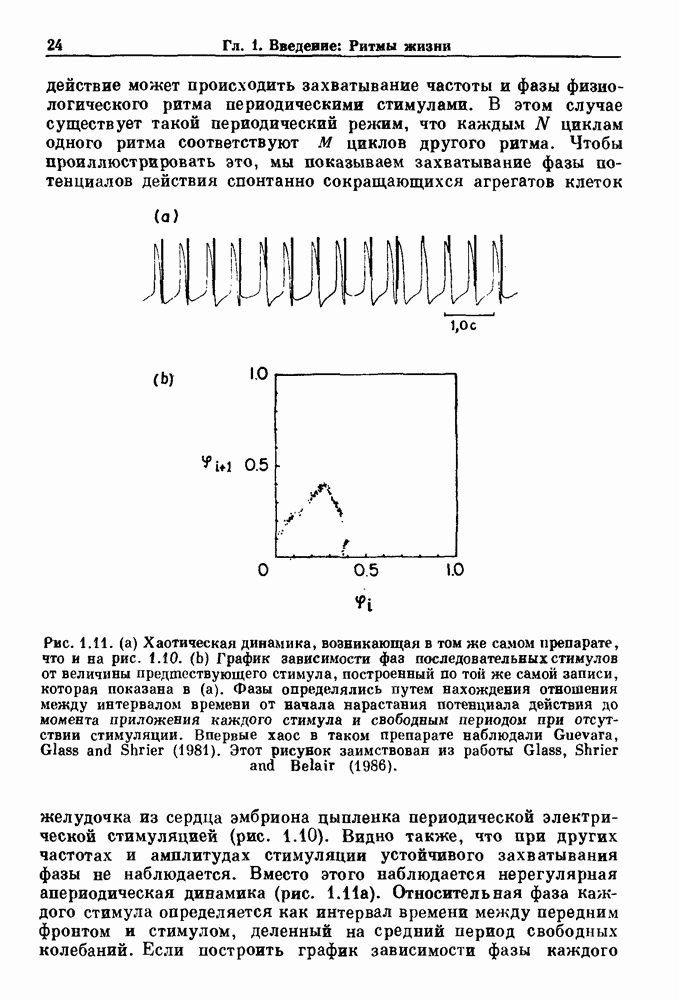
 известно (рис. 7.6). Тогда время
известно (рис. 7.6). Тогда время  за которое активность впервые достигает порога, может быть найдено решением уравнения
за которое активность впервые достигает порога, может быть найдено решением уравнения


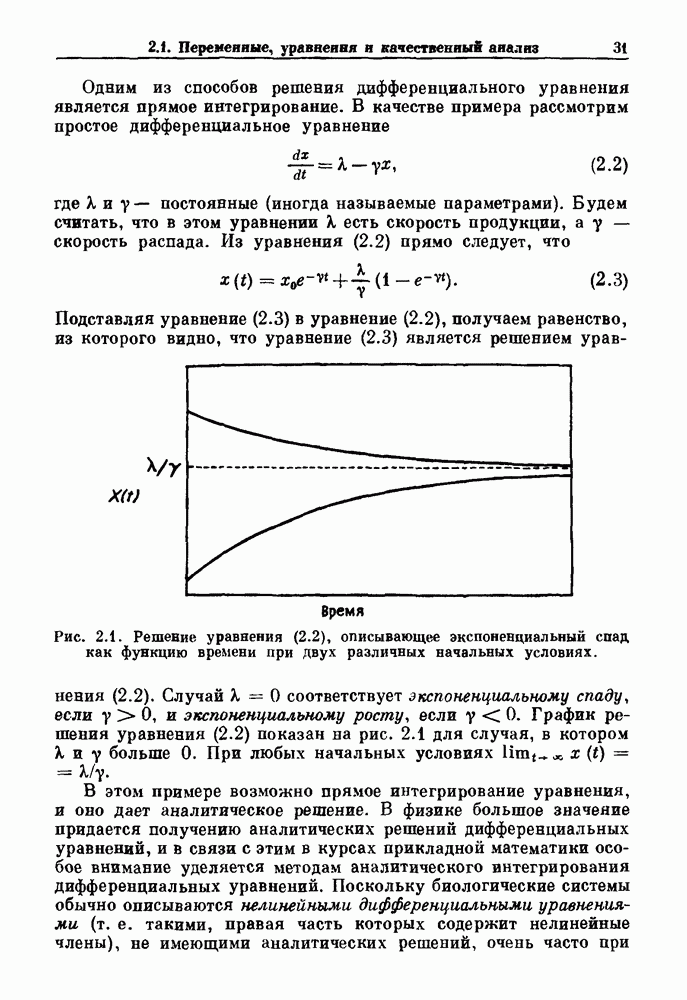
 есть
есть  Структура языков Арнольда имеет место в области, ограниченной сверху пунктирной линией. Из работы Glass and Belair (1986).
Структура языков Арнольда имеет место в области, ограниченной сверху пунктирной линией. Из работы Glass and Belair (1986).
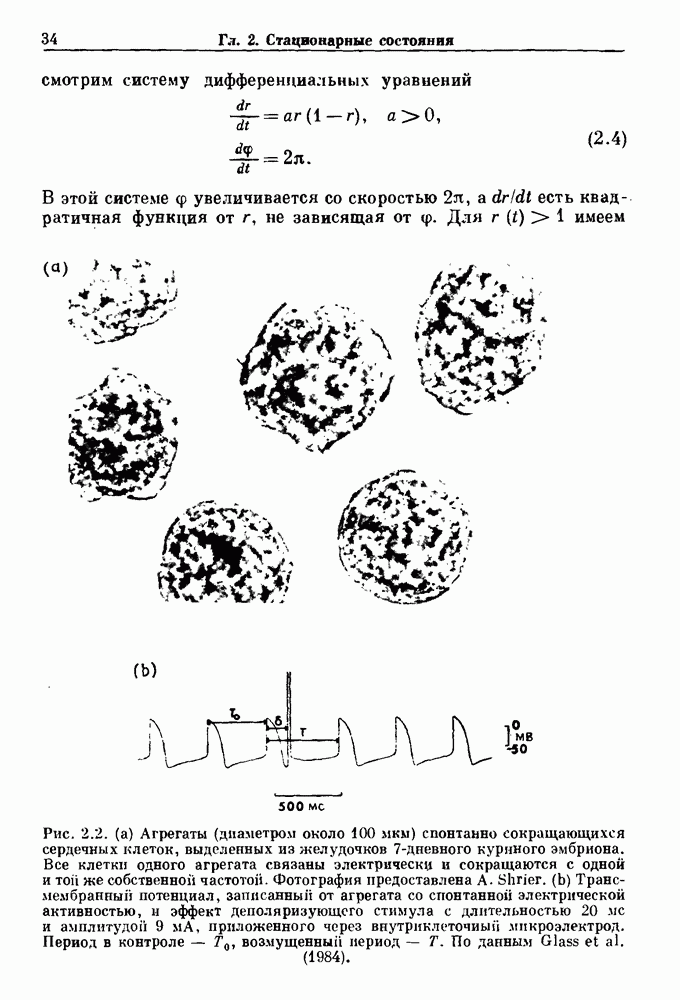
 динамика описывается непрерывным обратимым отображением единичной окружности, и предшествующие соображения, касающиеся языков Арнольда, применимы к этому случаю. Если же крутизна нарастания активности меньше, чем максимальная крутизна синусоидальной волны, то динамика описывается разрывной необратимой функцией. Известно, что в этом случае все еще будет происходить захват фазы
динамика описывается непрерывным обратимым отображением единичной окружности, и предшествующие соображения, касающиеся языков Арнольда, применимы к этому случаю. Если же крутизна нарастания активности меньше, чем максимальная крутизна синусоидальной волны, то динамика описывается разрывной необратимой функцией. Известно, что в этом случае все еще будет происходить захват фазы  для всех целых взаимно простых N и М. Существуют также такие значения параметров, которые приводят к апериодической динамике. Однако имеются два отличия между свойствами апериодической динамики в случае, когда она описывается разрывными кусочно-монотонными отображениями и обратимыми отображениями.
для всех целых взаимно простых N и М. Существуют также такие значения параметров, которые приводят к апериодической динамике. Однако имеются два отличия между свойствами апериодической динамики в случае, когда она описывается разрывными кусочно-монотонными отображениями и обратимыми отображениями.
