Пред.
След.
Макеты страниц
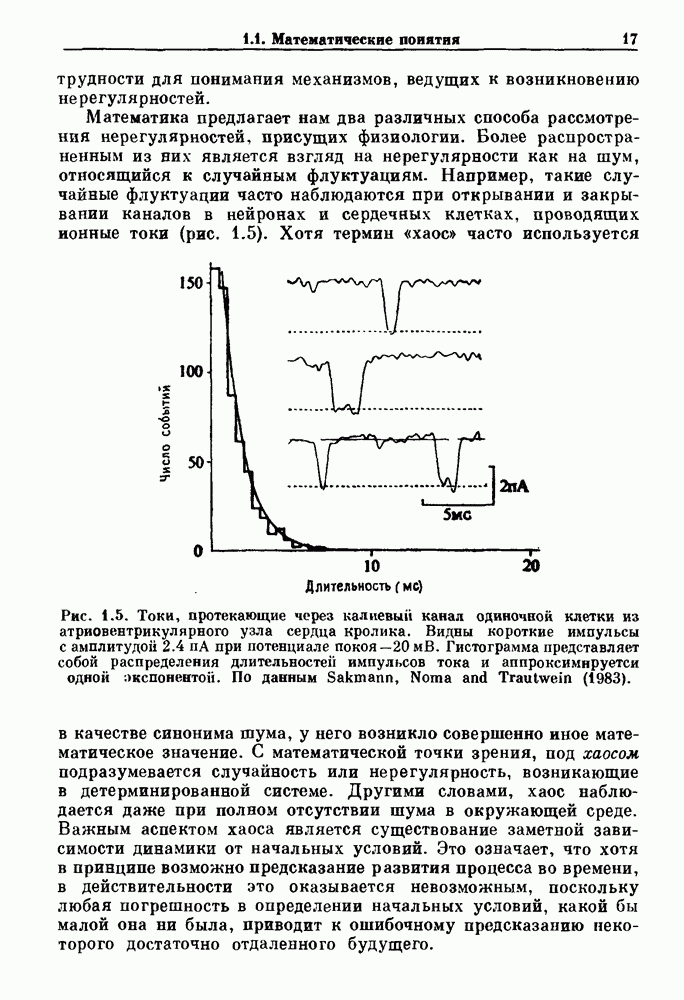
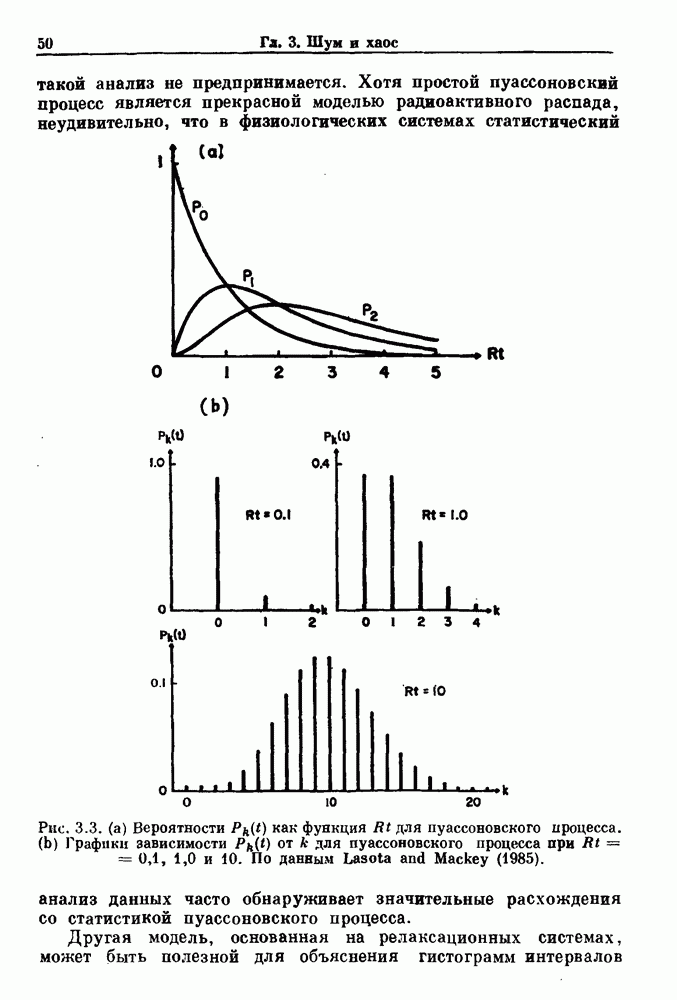
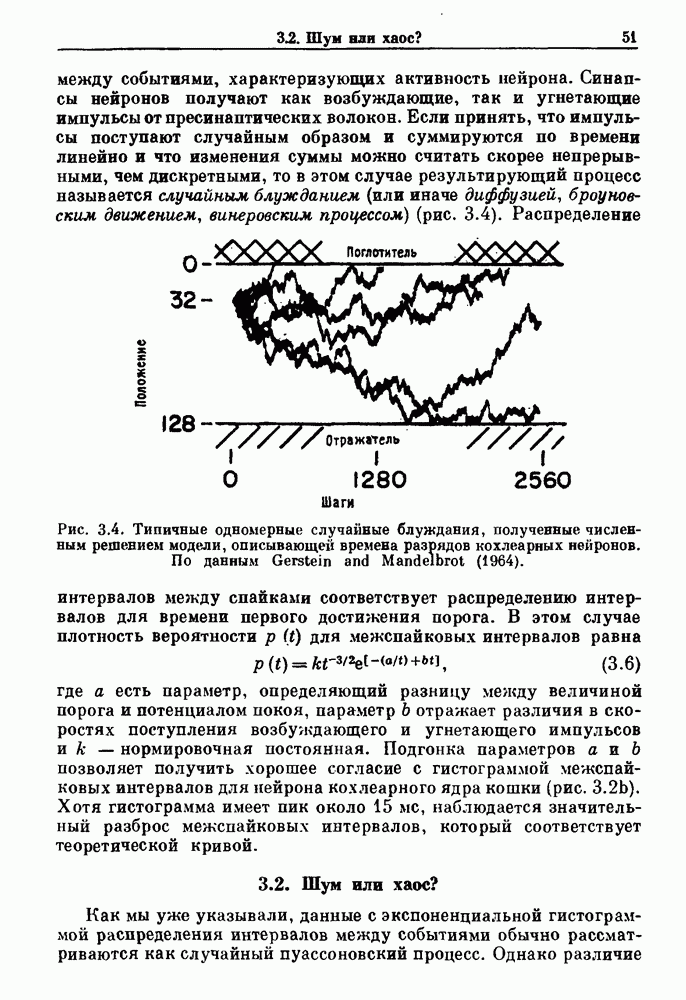
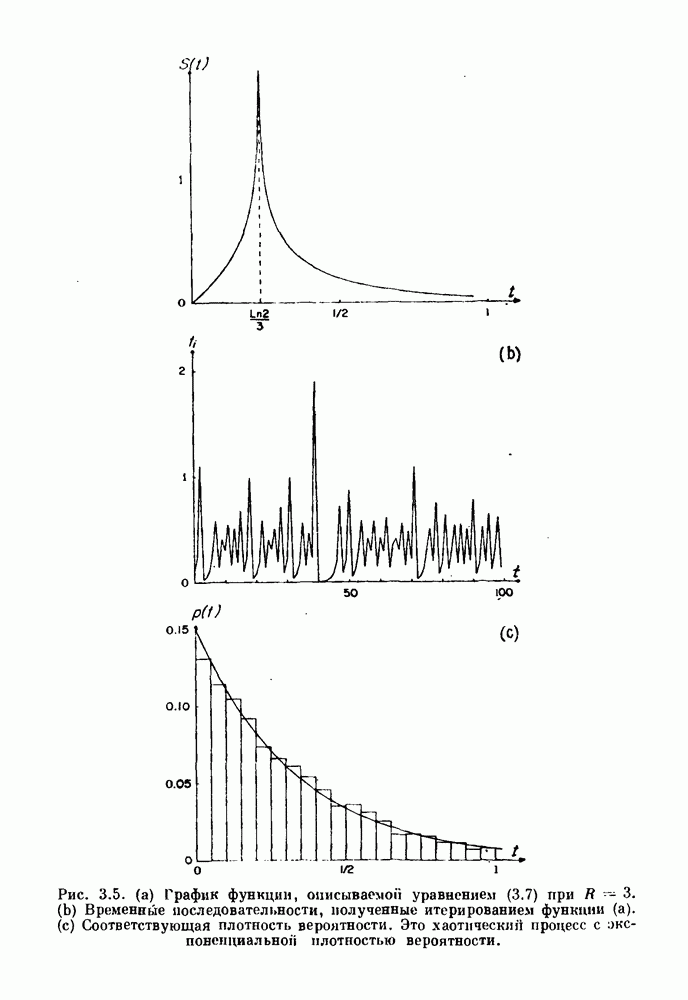
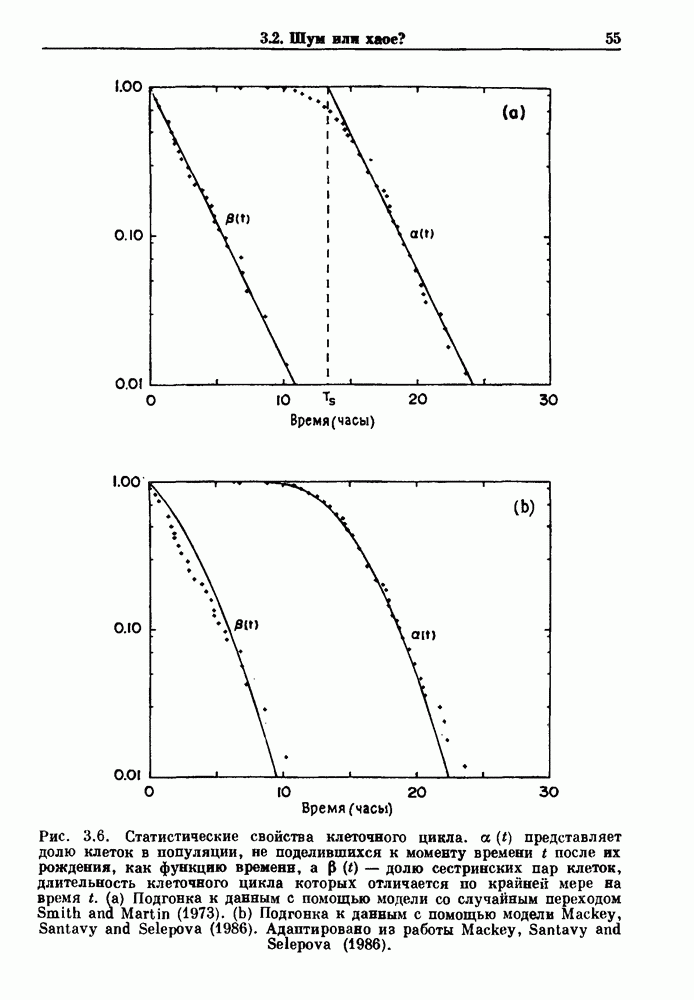
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
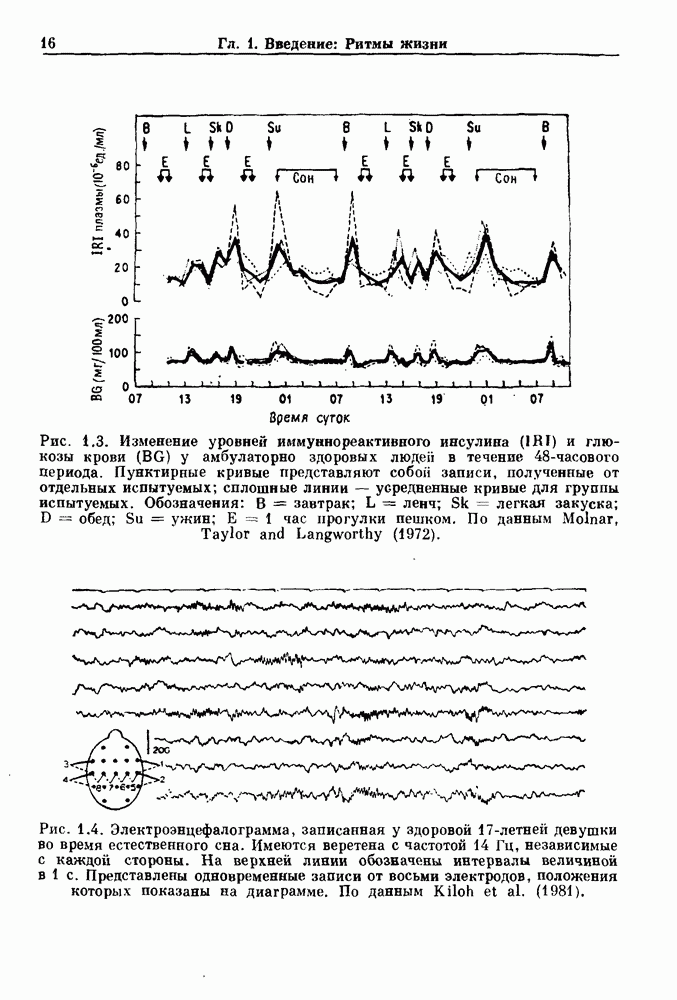
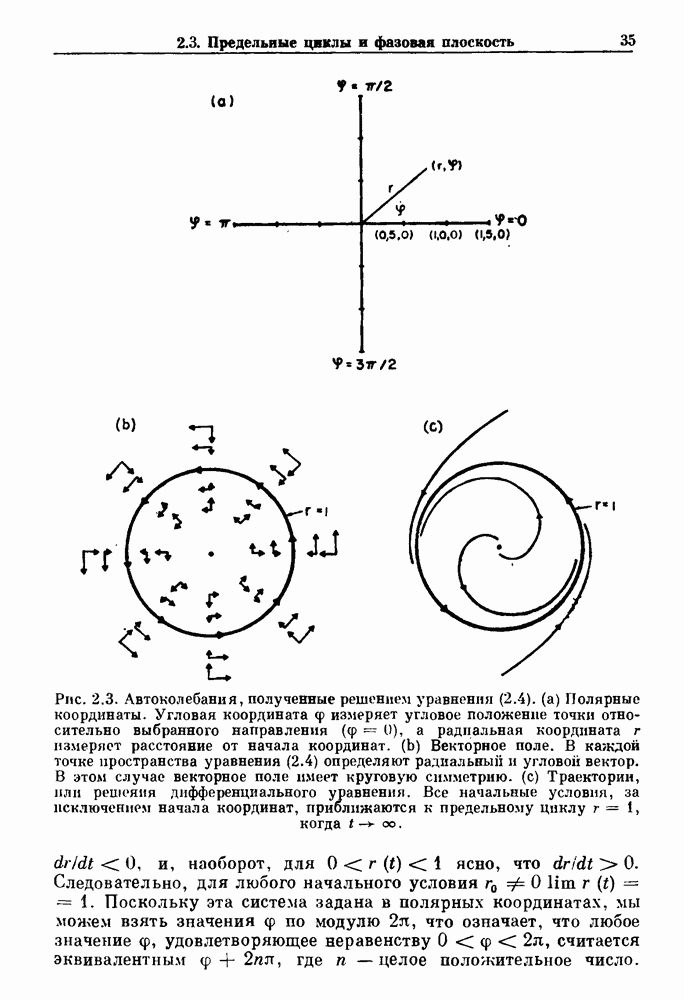
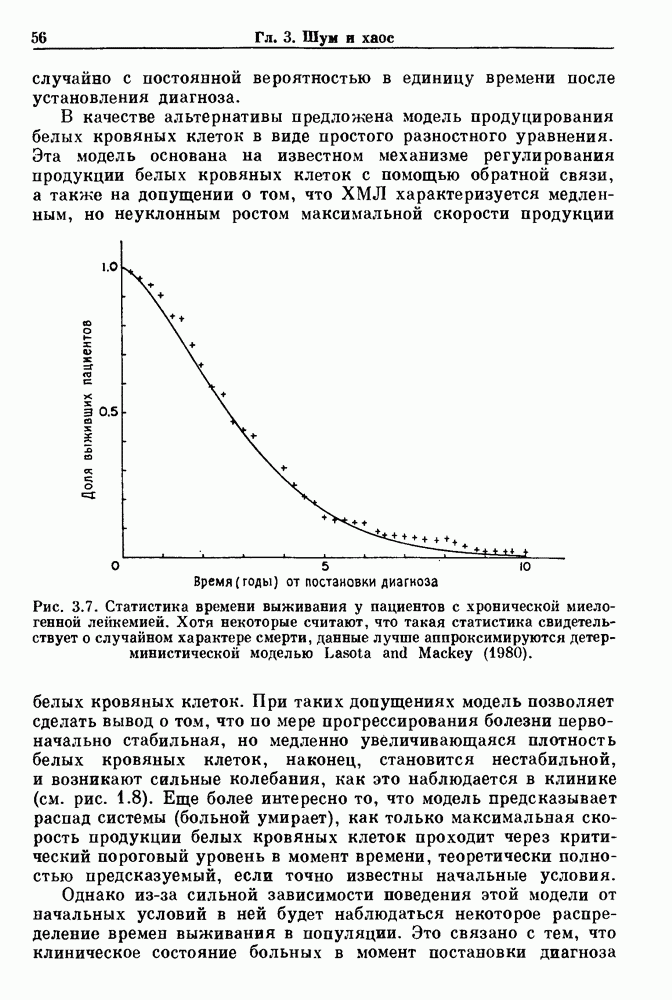
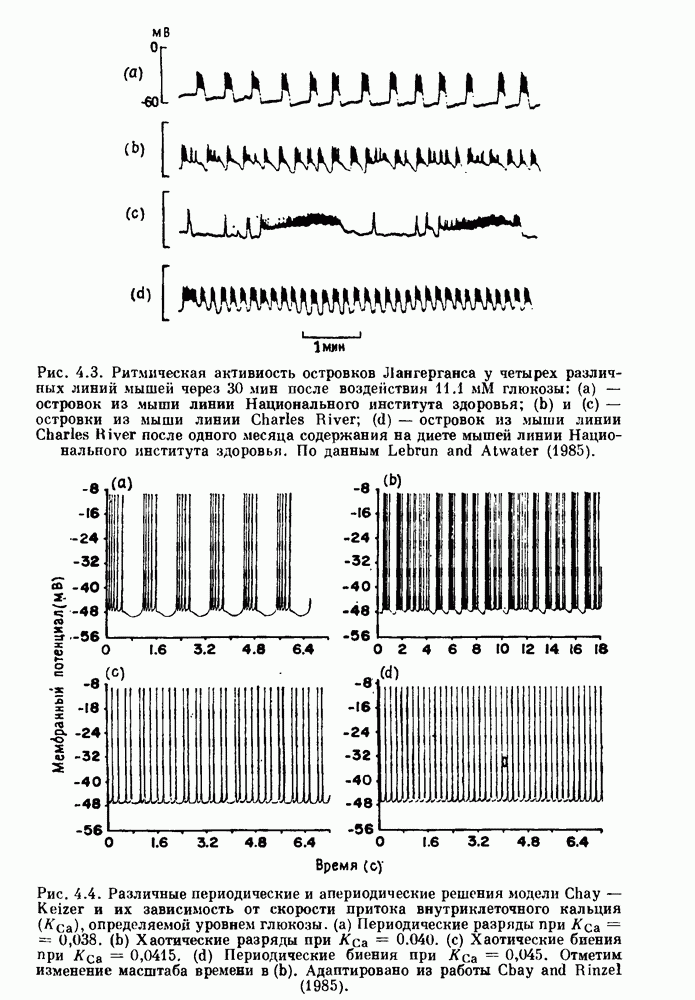
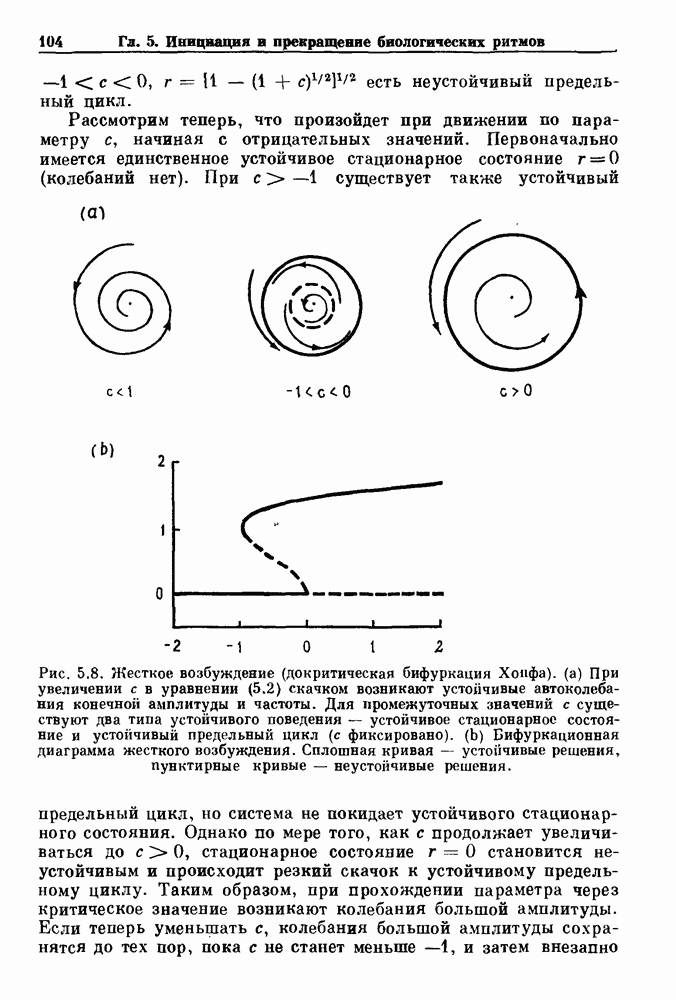
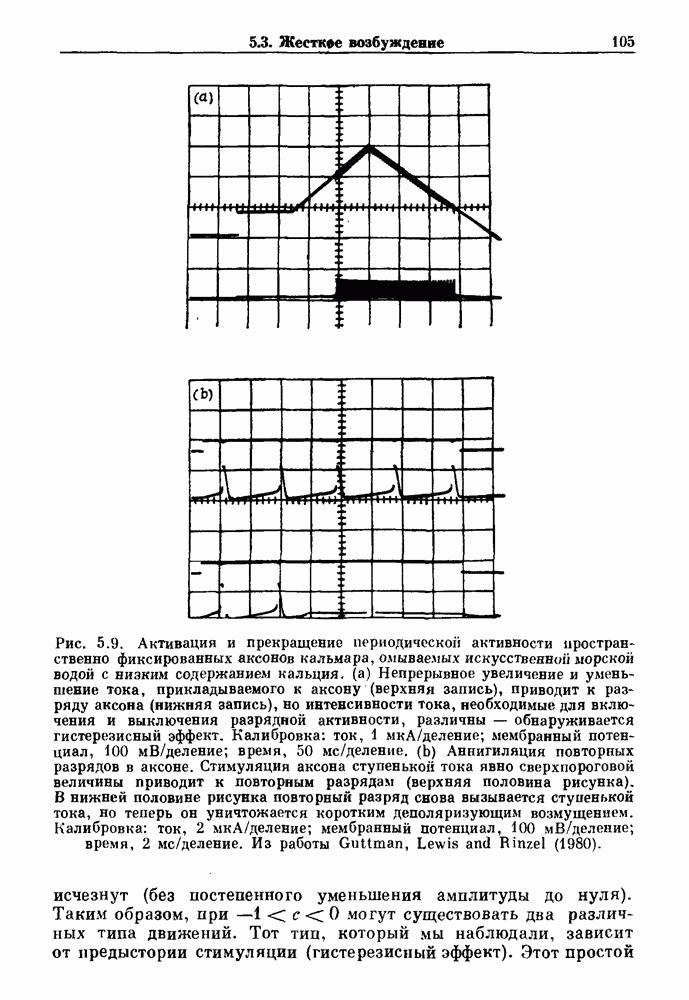
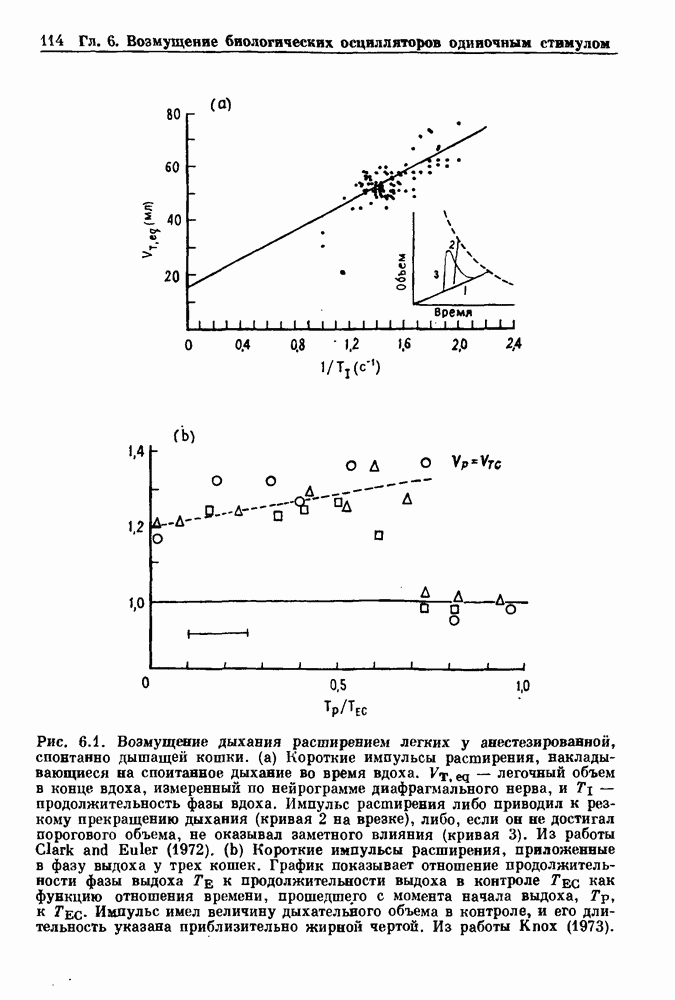
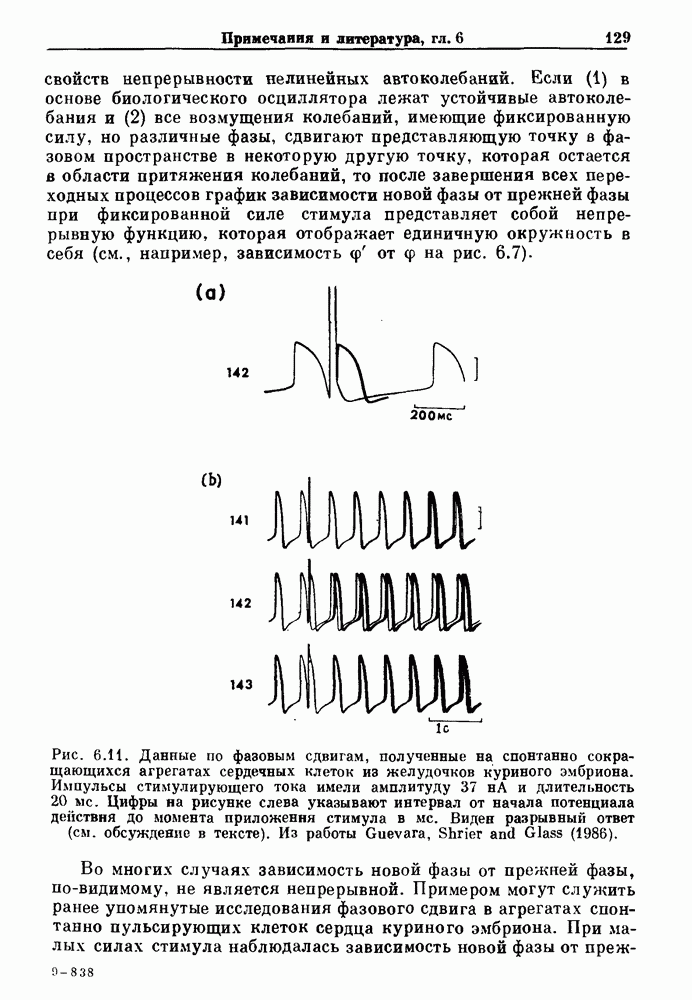
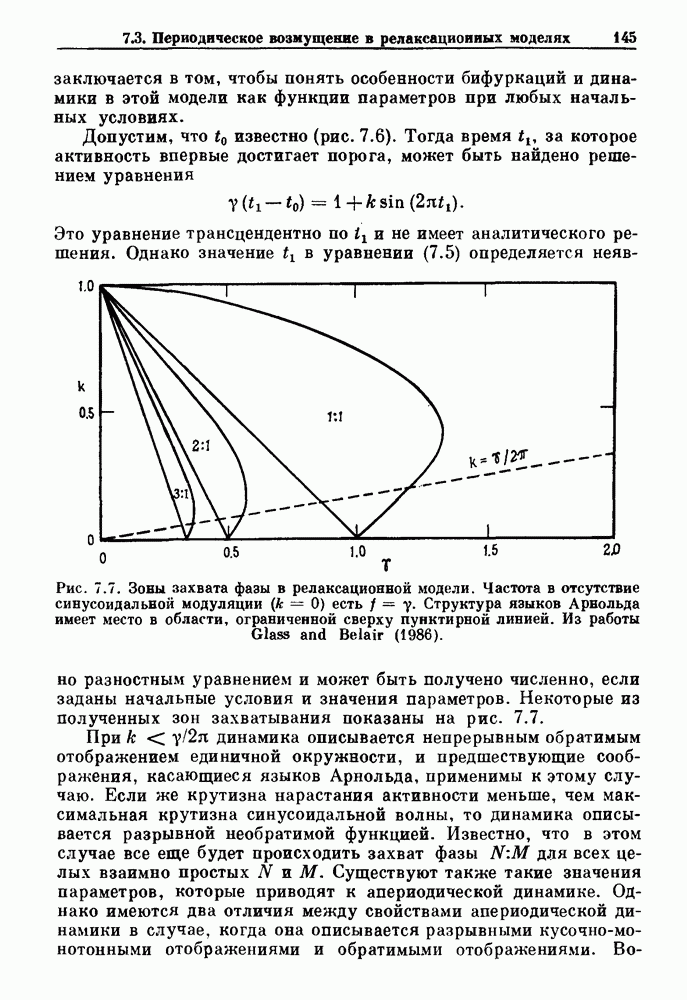
Глава 7. Периодическая стимуляция биологических осцилляторовПериодическая стимуляция спонтанно осциллирующих физиологических процессов оказывает мощное воздействие на эндогенный ритм. При изменении частоты и амплитуды периодической стимуляции устанавливаются разнообразные типы связей между стимулом и спонтанным осциллятором. В некоторых случаях происходит захват частоты и фазы спонтанного ритма стимулом, так что на каждые N циклов стимула приходится М циклов спонтанного ритма, и спонтанные колебания происходят в определенной фазе (или фазах) периодического стимула (захват фазы 7.1. Обзор экспериментальных результатовОдна из экспериментальных парадигм в изучении физиологических осцилляторов состоит в том, что осциллятор подвергается воздействию периодических стимулов, в то время как физиологические условия поддерживаются по возможности постоянными. Мы не пытаемся подытожить всю обширную информацию, имеющуюся по этому вопросу; вместо этого мы обсуждаем два характерных случая: механическую вентиляцию легких у животных и воздействие на сердечный ритм периодической стимуляции с использованием внутриклеточного микроэлектрода. На основании рассмотрения этих случаев делаются обобщения, применимые к широкому классу экспериментальных систем. Со времен Геринга и Брейера известно, что периодическое расширение легких механическим вентилятором может привести к захвату фазы эндогенного дыхательного ритма у млекопитающих ритмом вентилятора. Полагают, что захват фазы осуществляется через рефлексы Геринга—Брейера, при которых расширение легких угнетает вдох и удлиняет выдох (см. гл. 6). Афферентная активность рецепторов растяжения легких передается по афферентным волокнам блуждающего нерва. Захват дыхательного ритма может изучаться с помощью механической вентиляции легких животного при различных объемах и частотах вентилятора в условиях поддержания постоянных уровней анестезии, температуры тела и концентрации газов в крови. Эксперименты выполнялись на парализованных, анестезированных пентобарбиталом взрослых кошках. Активность дыхательного центра прослеживалась с помощью отведения активности одной из ветвей диафрагмального нерва. У нормального непарализованного животного диафрагмальный нерв иннервирует диафрагму, и его активация вызывает сокращение диафрагмы, что приводит ко вдоху. У парализованного животного нервно-мышечная передача между диафрагмальным нервом и диафрагмой блокируется, и расширение легких происходит исключительно в результате механической вентиляции. Однако афферентная активность рецепторов растяжения легких по-прежнему передается по блуждающему нерву. Необходимо предпринимать специальные меры для поддержания постоянной концентрации газов в крови при изменении частоты и амплитуды вентилятора. Если животное подвергается гипервентиляции, активность дыхательного центра исчезает из-за низкого уровня При изменении объема и частоты вентилятора устанавливаются различные взаимодействия между вентилятором и активностью диафрагмального нерва. Возникающие при этом ритмы располагаются в координатах «объем вентилятора» — «частота вентилятора» упорядоченным образом, как показано на рис. 7.1. На врезках представлены типичные записи объемов вентилятора и активности диафрагмального нерва, соответствующие различным случаям захвата фазы, а также ритмам без захвата фазы (возникающим в заштрихованных областях). В этих экспериментах не могут быть использованы малые частоты и объемы вентиляции из-за необходимости поддерживать адекватную вентиляцию. Невозможно также изучить действие очень больших частот и объемов из-за механических ограничений вентилятора и ограниченной емкости легких кошки. Как обсуждалось в разд. 4.2, принято считать, что дыхательный ритм генерируется сложной сетью нейронов в стволе мозга.
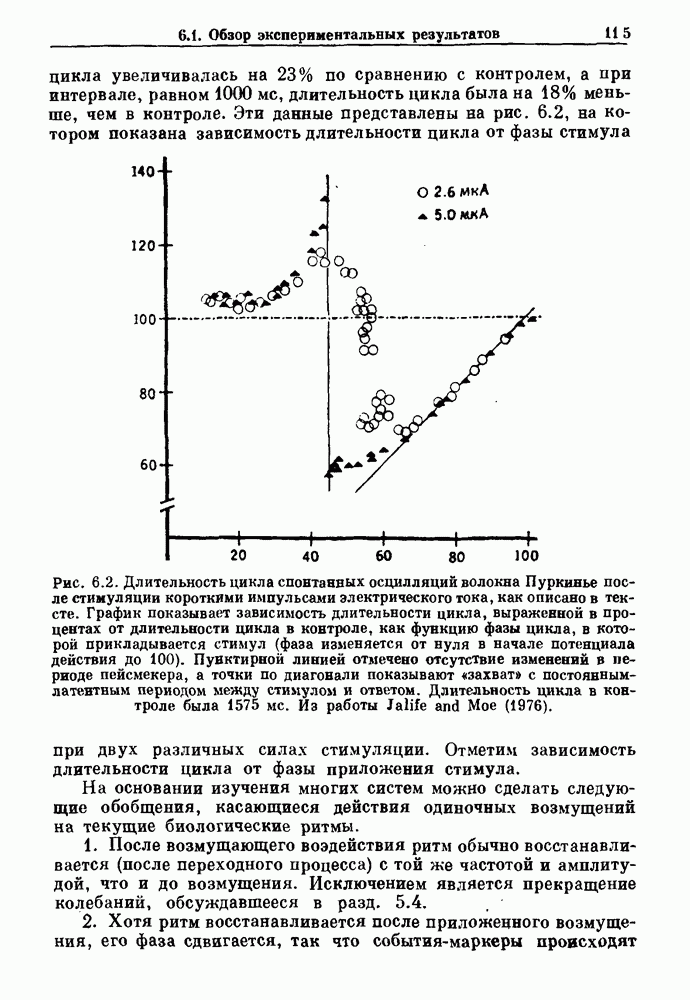
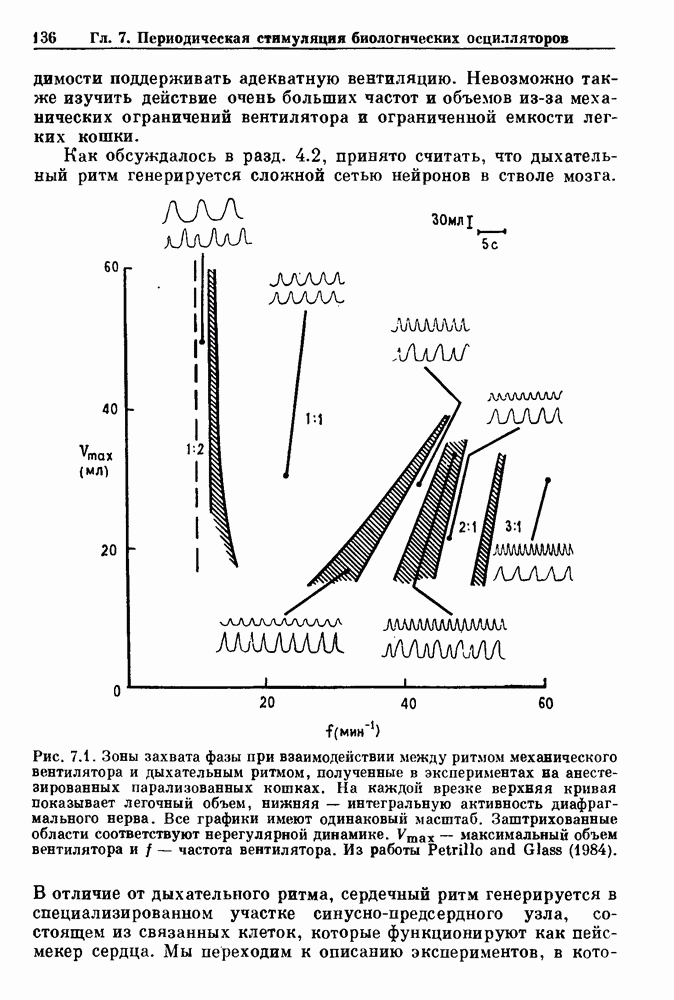
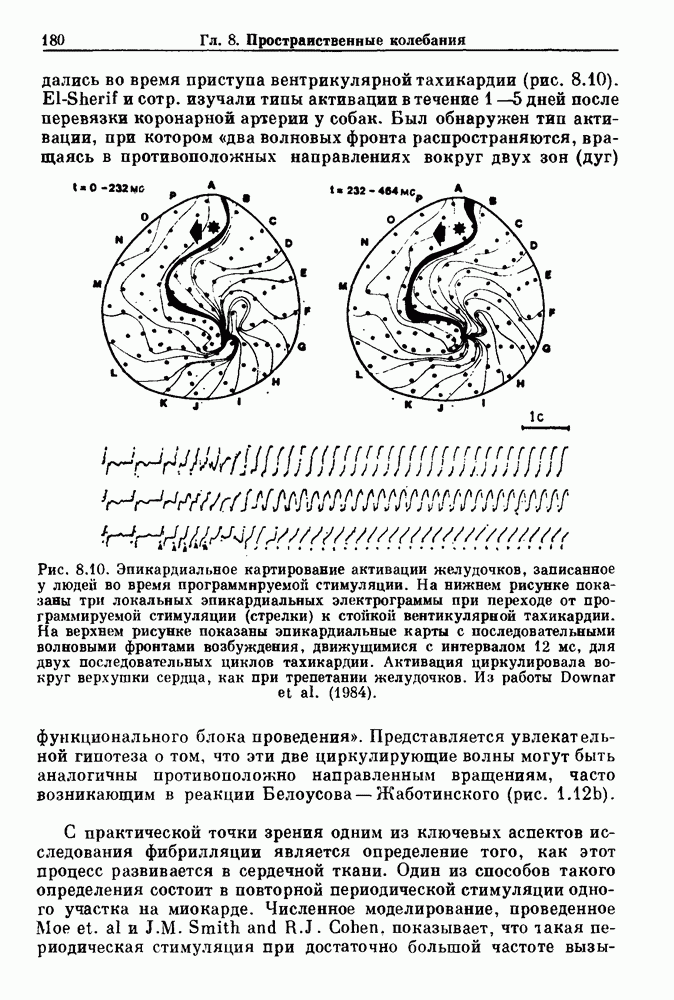
Рис. 7.1. Зоны захвата фазы при взаимодействии между ритмом механического вентилятора и дыхательным ритмом, полученные в экспериментах на анестезированных парализованных кошках. На каждой врезке верхняя кривая показывает легочный объем, нижняя — интегральную активность диафрагмального нерва. Все графики имеют одинаковый масштаб. Заштрихованные области соответствуют нерегулярной динамике. Ктах — максимальный объем вентилятора и В отличие от дыхательного ритма, сердечный ритм генерируется в специализированном участке синусно-предсердного узла, состоящем из связанных клеток, которые функционируют как пейсмекер сердца. Мы переходим к описанию экспериментов, в которых изучалось влияние импульсных электрических стимулов на спонтанно сокращающиеся клетки, изолированные из желудочков сердца куриного эмбриона. При изменении частоты и тока электрических стимулов устанавливаются различные виды взаимодействий между стимулятором и сердечными клетками.
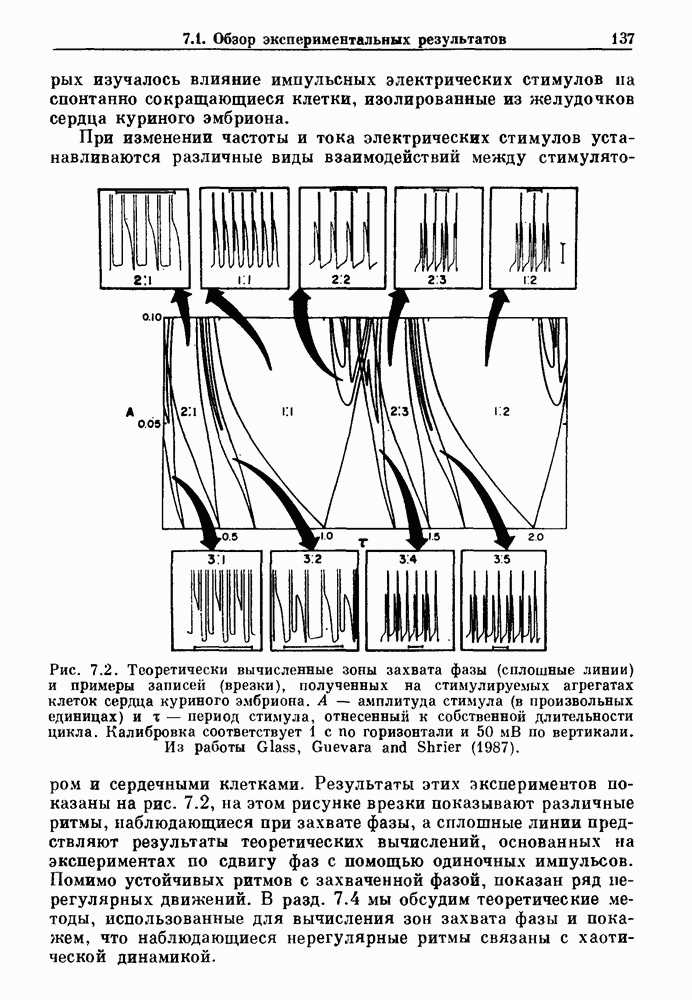
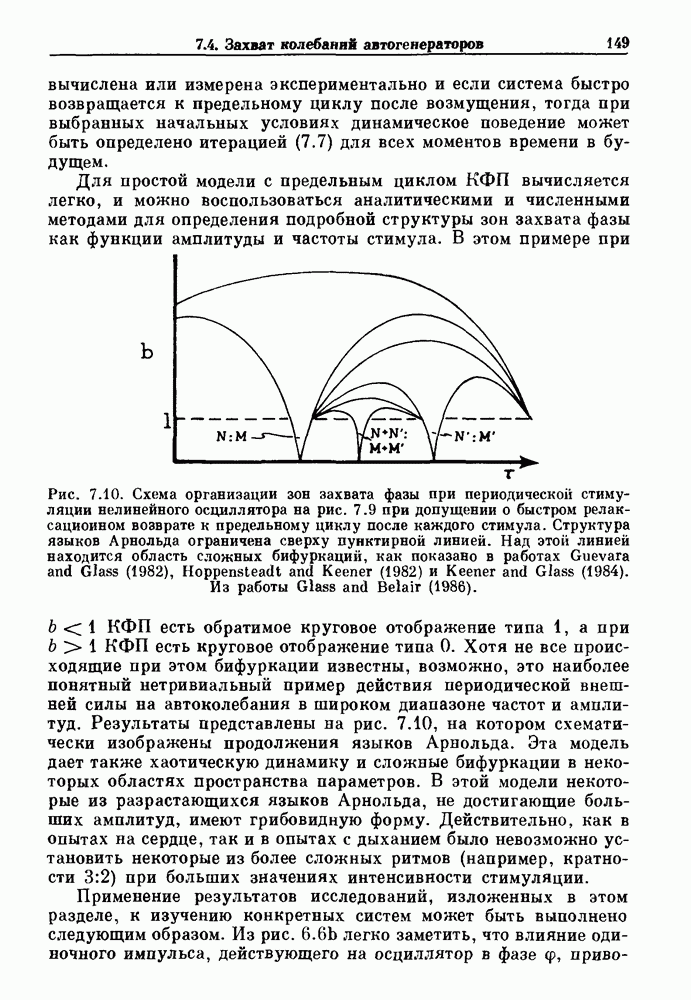
Рис. 7.2. Теоретически вычисленные зоны захвата фазы (сплошные линии) и примеры записей (врезки), полученных на стимулируемых агрегатах клеток сердца куриного эмбриона. А — амплитуда стимула (в произвольных единицах) их — период стимула, отнесенный к собственной длительности цикла. Калибровка соответствует 1 с по горизонтали и 50 мВ по вертикали. Из работы Glass, Guevara and Shrier (1987). Результаты этих экспериментов показаны на рис. 7.2, на этом рисунке врезки показывают различные ритмы, наблюдающиеся при захвате фазы, а сплошные линии предствляют результаты теоретических вычислений, основанных на экспериментах по сдвигу фаз с помощью одиночных импульсов. Помимо устойчивых ритмов с захваченной фазой, показан ряд нерегулярных движений. В разд. 7.4 мы обсудим теоретические методы, использованные для вычисления зон захвата фазы и покажем, что наблюдающиеся нерегулярные ритмы связаны с хаотической динамикой. Несмотря на то, что эти результаты были получены на двух сильно отличающихся физиологических системах с использованием различных способов периодической стимуляции, обнаружились некоторые поразительные особенности, общие для обеих систем. Ниже приводятся обобщения, применимые к большому числу экспериментов по периодическому воздействию внешней силы на биологические осцилляторы. 1. Наиболее часто наблюдающиеся устойчивые зоны захвата фазы соответствуют небольшой кратности циклов вынуждающего стимула и свободного ритма (т. е., 2:1, 3:2, 1:1, 2:3, 1:2). Хотя можно наблюдать и другие отношения N: М с большими величинами N и М, они занимают меньшие области в пространстве параметров частота — амплитуда и, следовательно, могут легко ускользать от внимания или маскироваться шумом. 2. Устойчивые ритмы существуют в определенных областях плоскости частота—амплитуда. Принято связывать число оборотов 3. При очень низких амплитудах стимуляции трудно поддерживать устойчивый захват фазы. 4. При изучении областей амплитудно-частотного пространства параметров, расположенных между зонами устойчивого захвата фазы, обычно можно найти такие значения параметров стимуляции, которые приводят к нерегулярной динамике. Несмотря на некоторое сходство, между сердечной и дыхательной системами существуют различия. Например, область кратности 2:2, наблюдаемая при периодической стимуляции сердечных клеток, не обнаруживалась в случае механической вентиляции легких кошки. Целью теоретического изучения захват фазы является обеспечение теоретической основы для понимания сходства и различия между разными объектами. В идеале исследователи хотели бы делать предсказания о захвате фазы при изменении параметров стимуляции исходя из механизмов ритмогенеза и связи между стимулятором и свободным ритмом. На практике математический анализ периодического воздействия внешней силы на нелинейные осцилляторы оказывается чрезвычайно сложным, и детальные количественные характеристики движения таких систем были получены лишь в нескольких специальных случаях. Теоретические исследования показывают, что хотя существует большое сходство между динамическим поведением различных систем, выявляются также и различия при достаточно подробном его изучении. Чтобы подкрепить это утверждение, мы рассмотрим динамическое поведение различных модельных систем при действии на них периодической стимуляции. Мы также обсудим захват фазы биологических ритмов в ряде клинических случаев.
|
 |
Оглавление
|















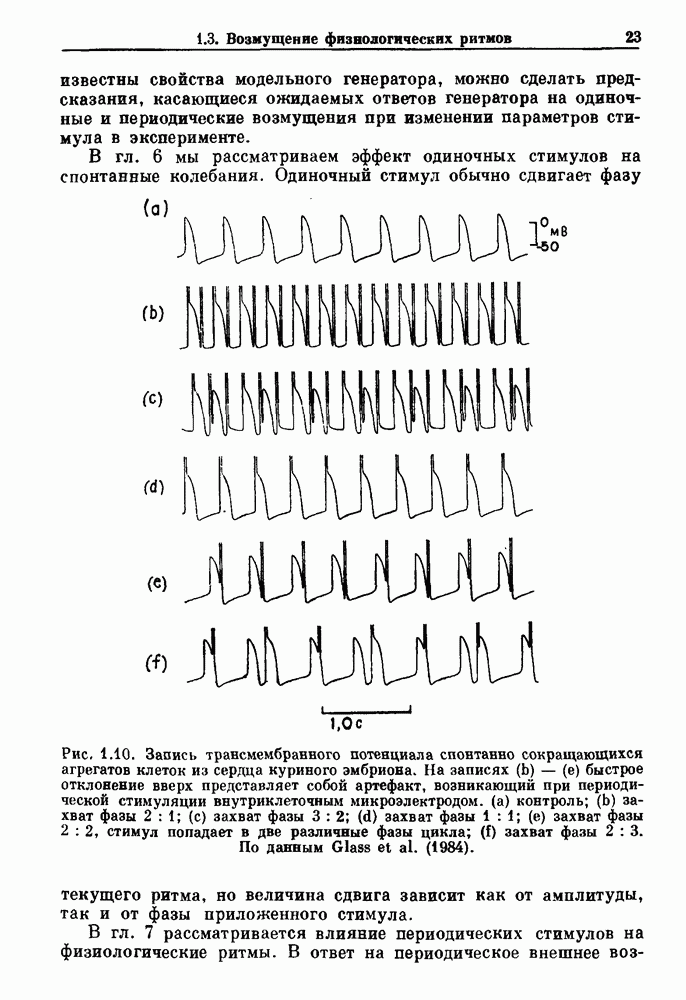
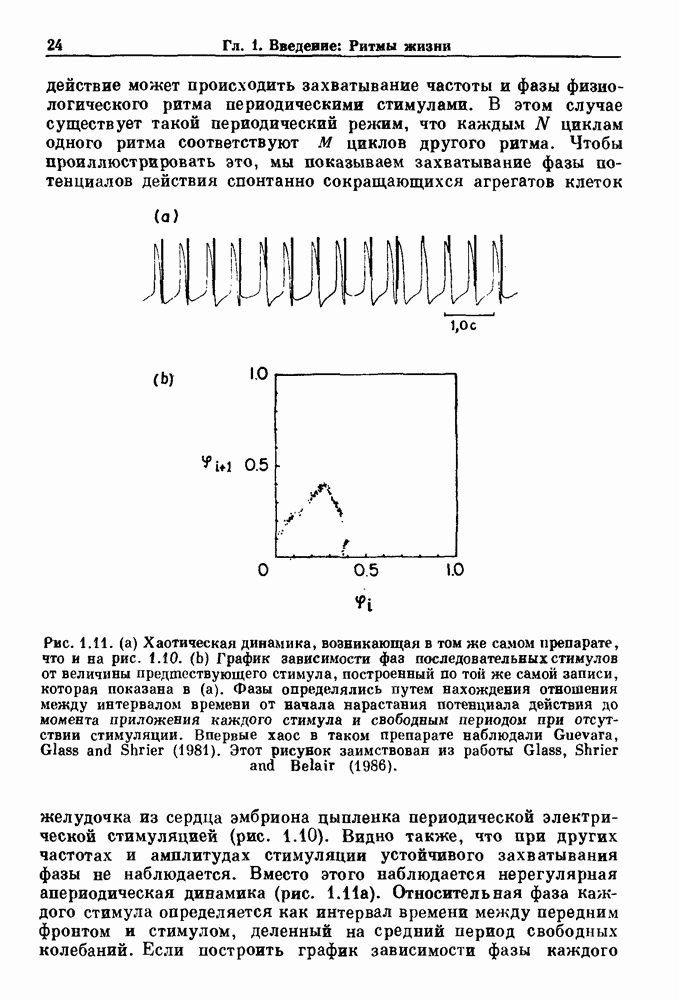






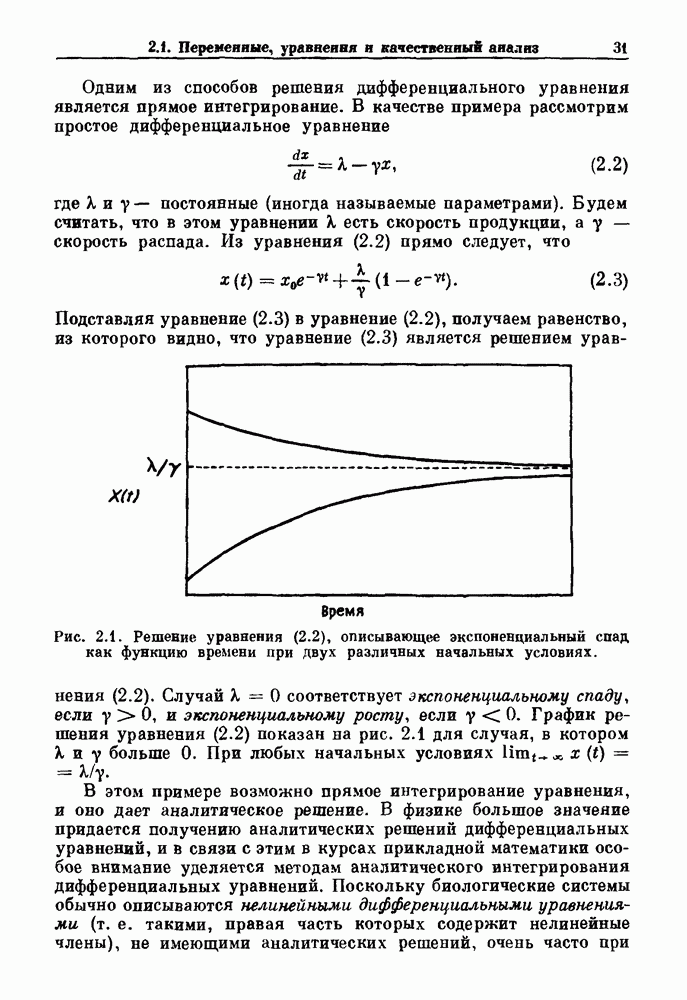


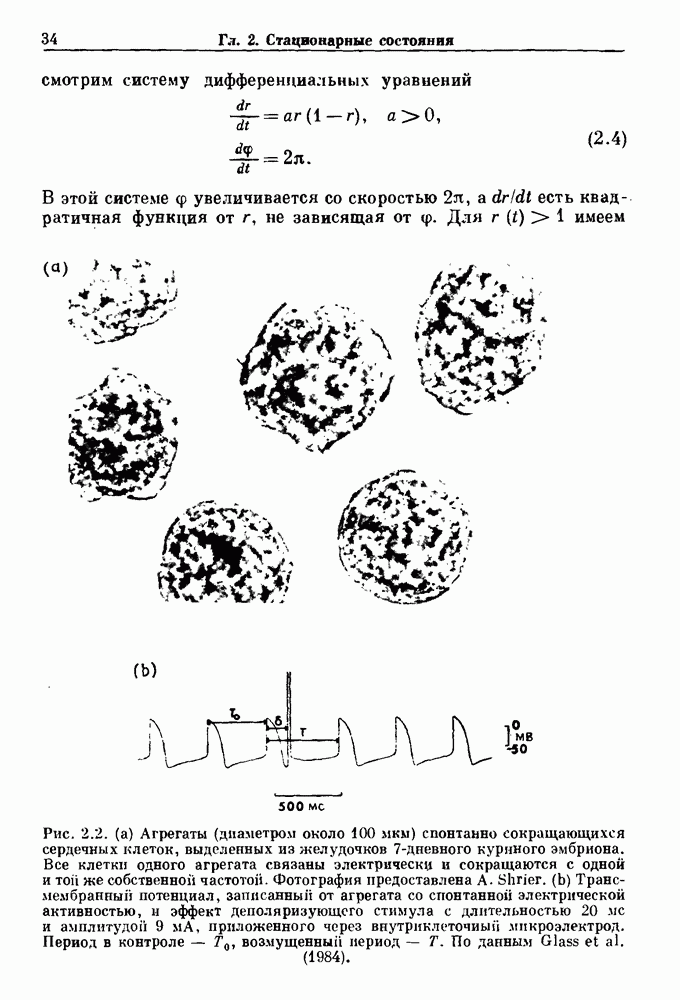





















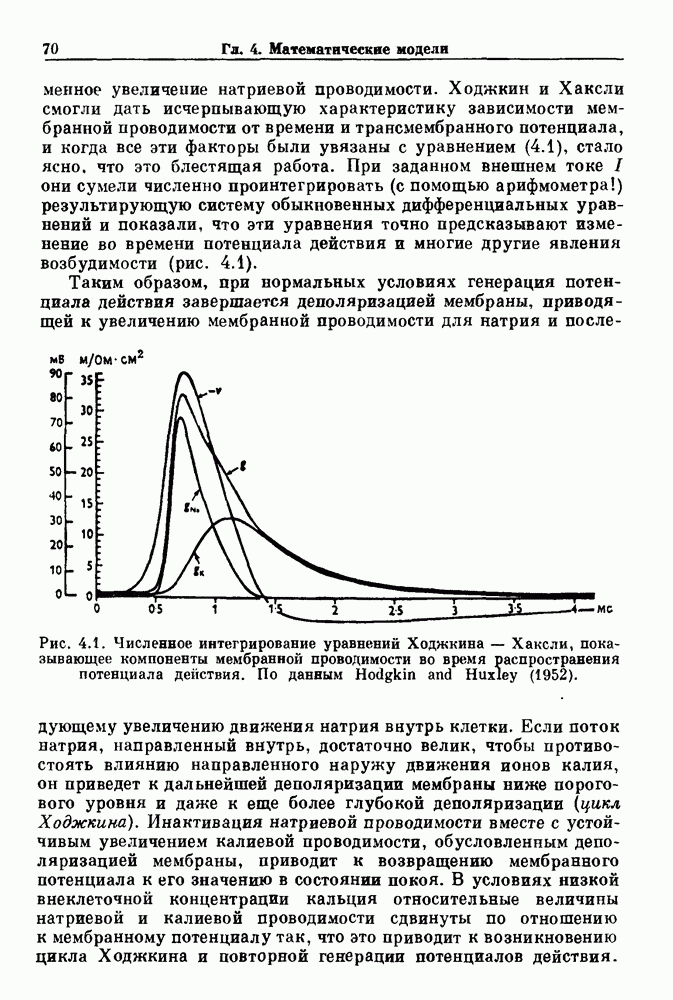

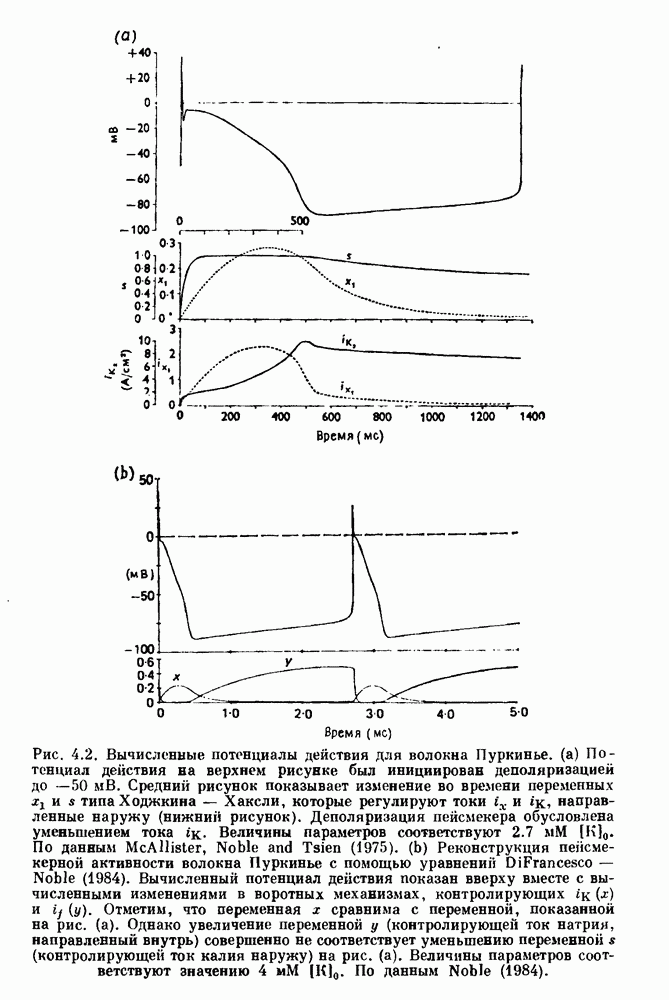
















































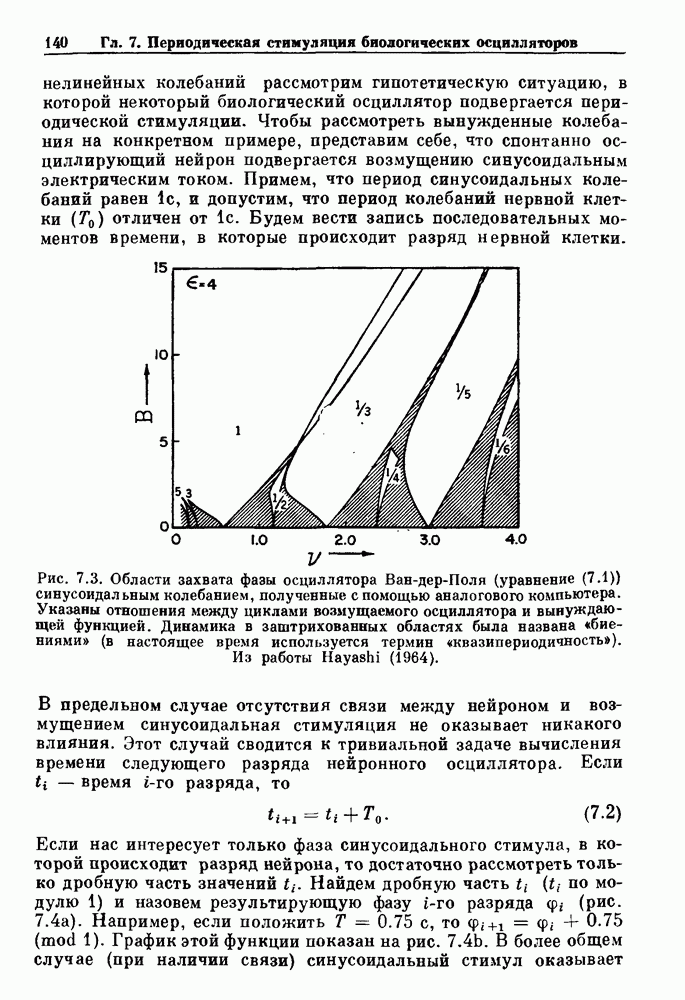
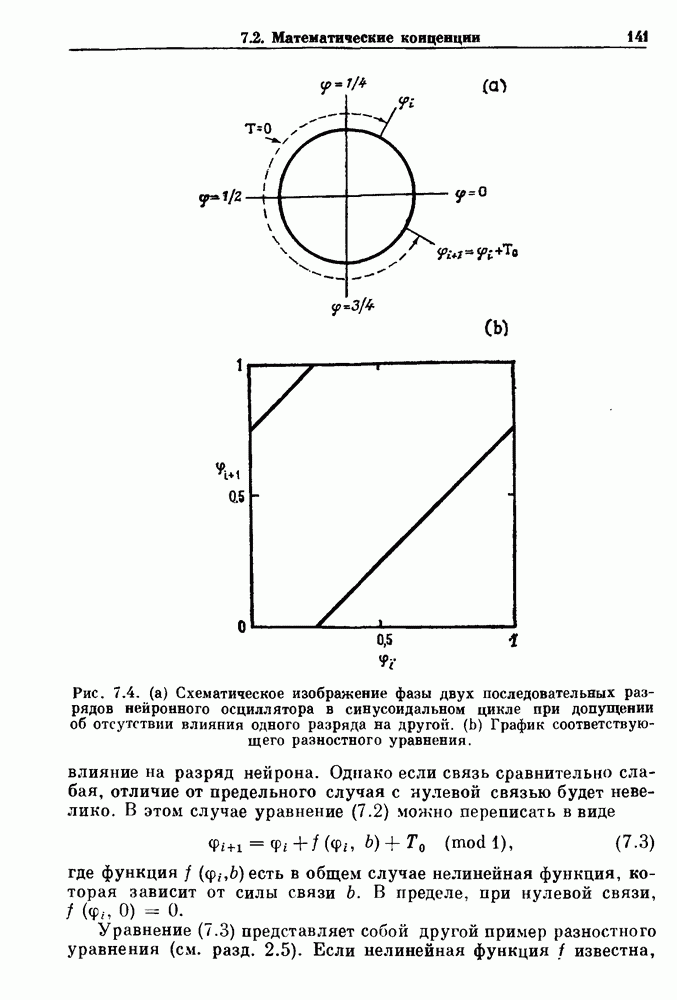





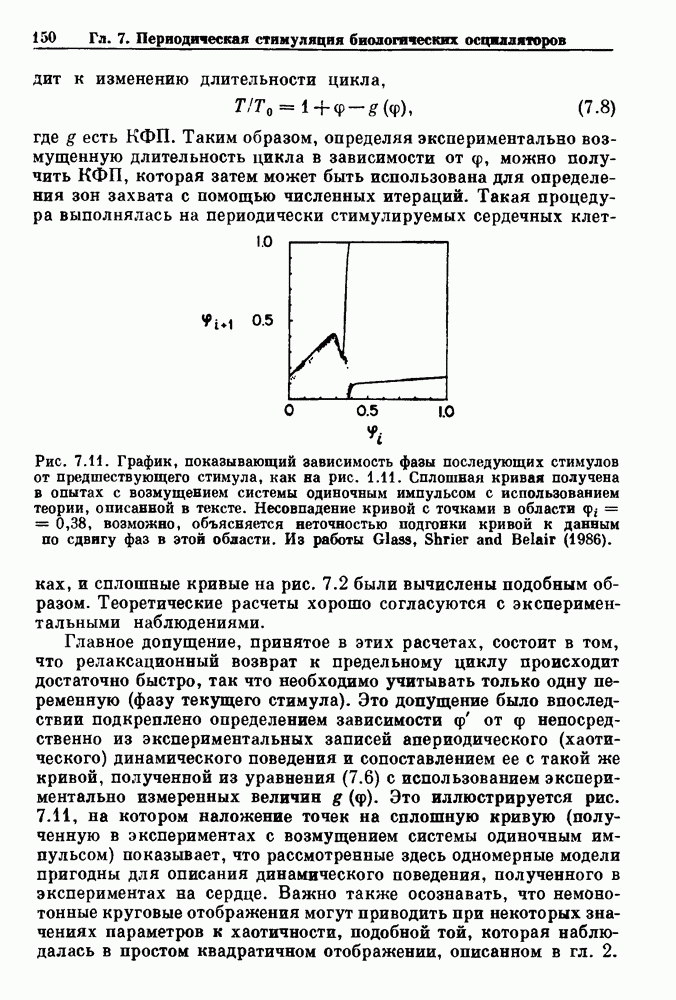


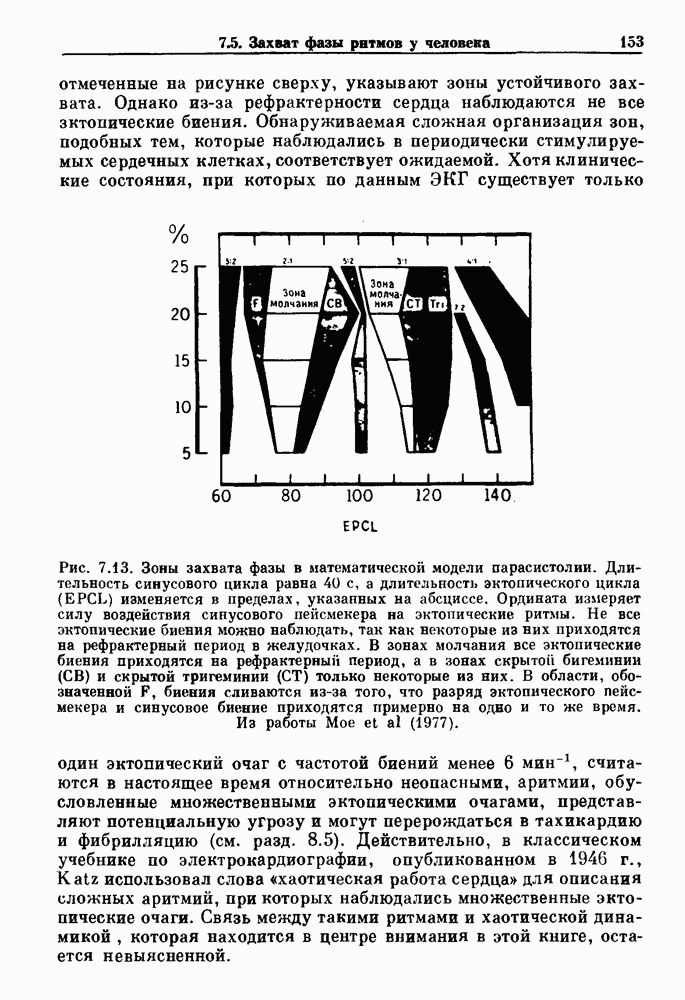
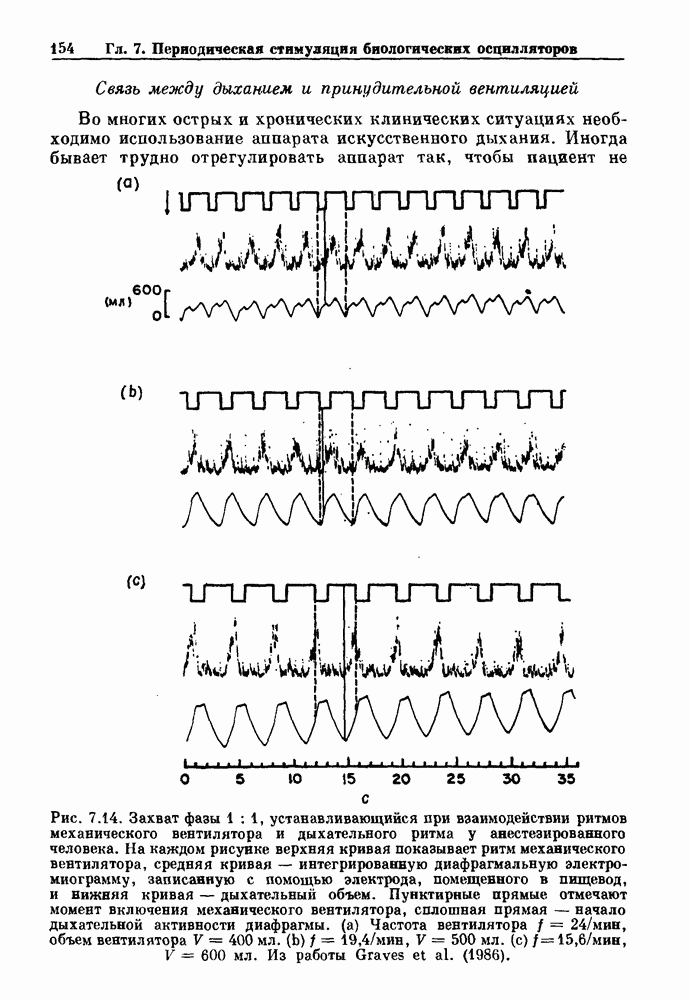
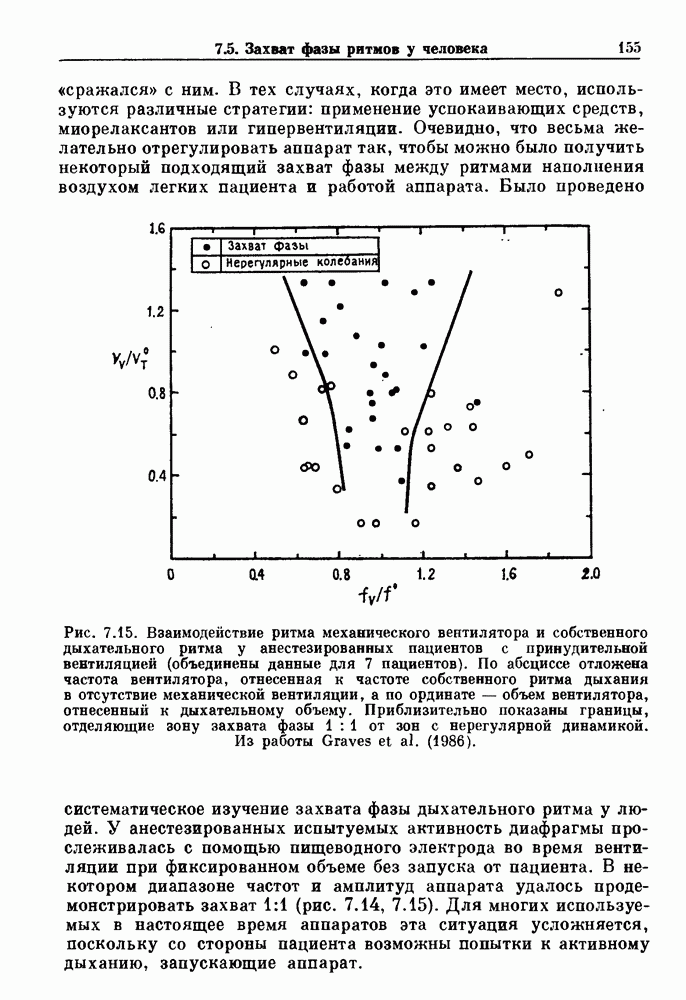







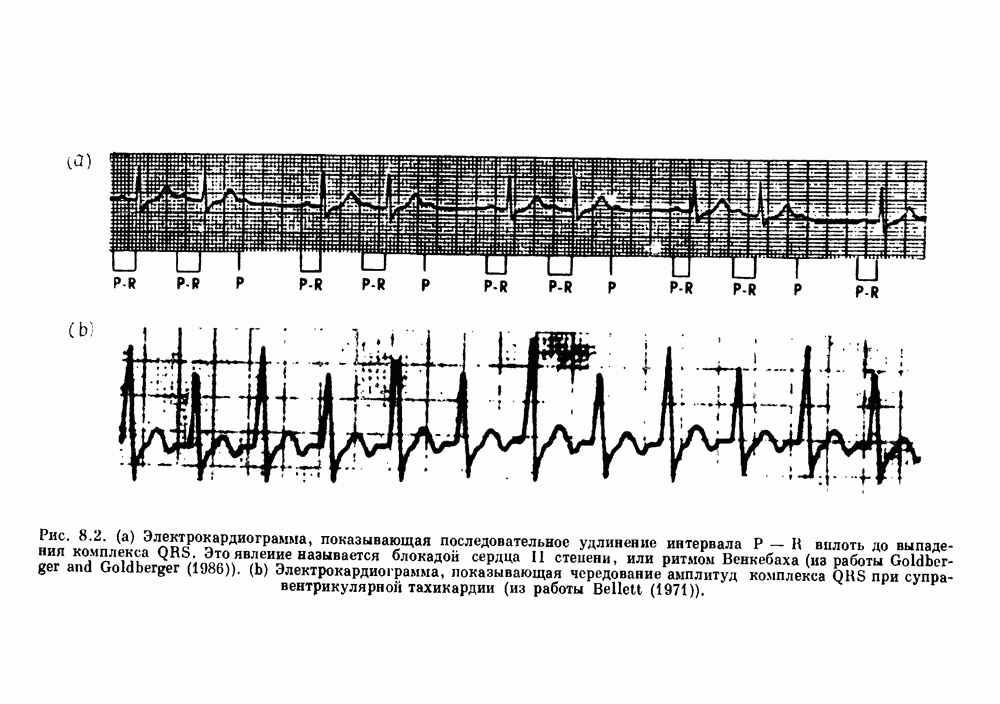


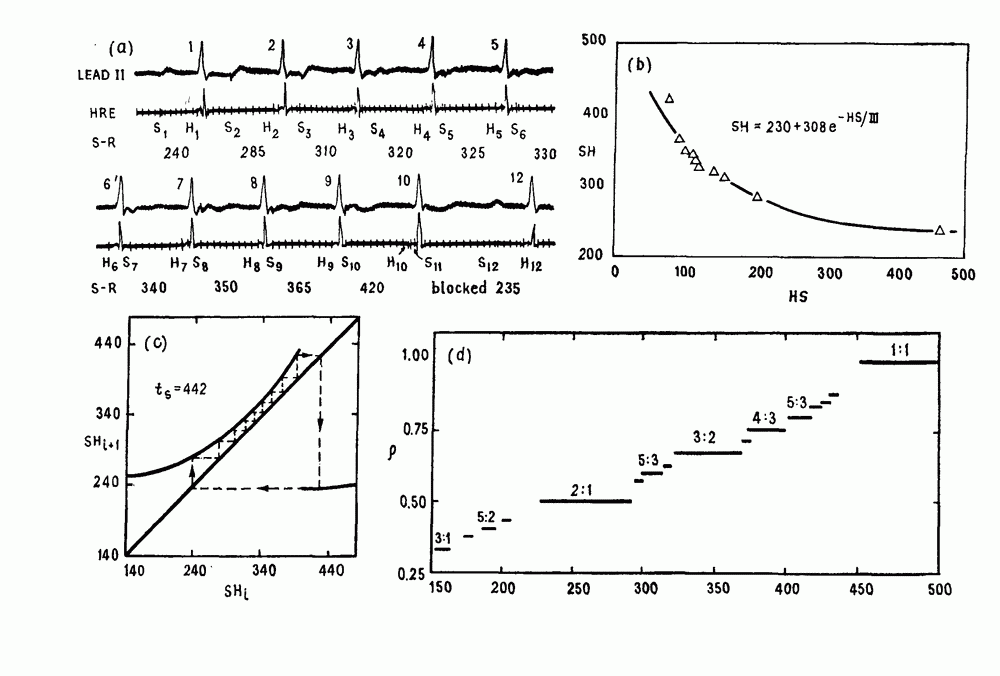

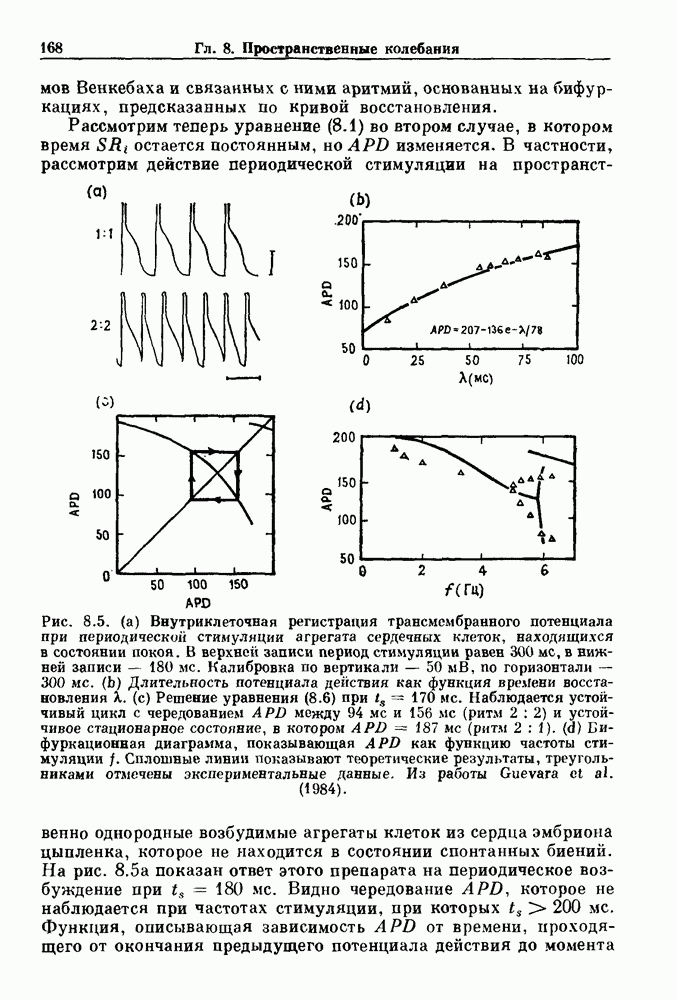

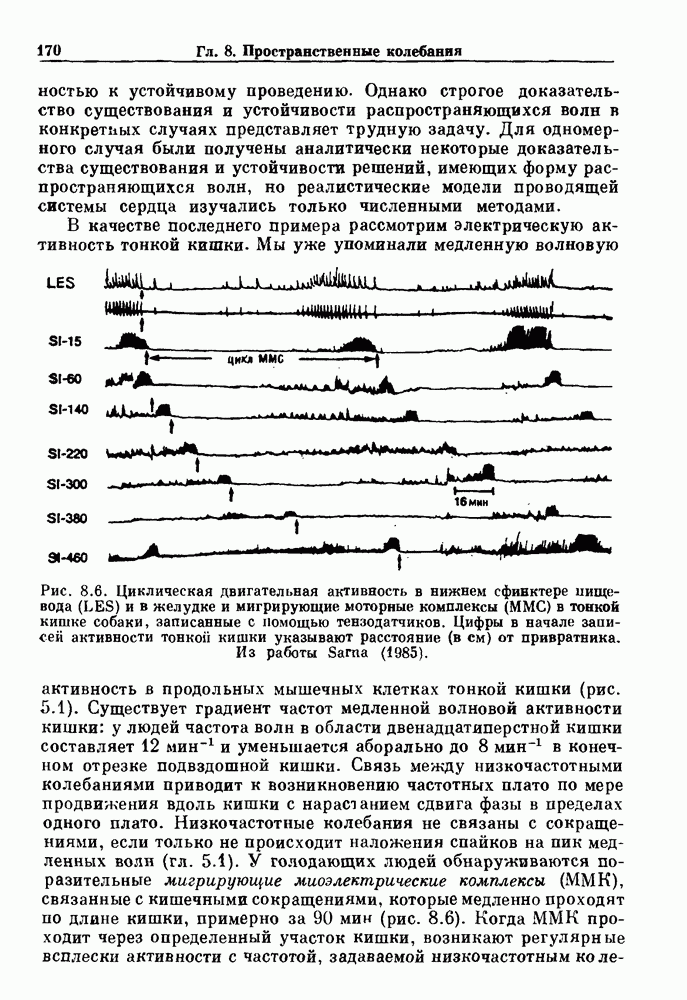





































































 ). Помимо ритмов с захватом фазы, можно наблюдать также нерегулярные или апериодические движения, при которых фиксированные соотношения фаз и регулярно повторяющиеся циклические процессы не наблюдаются. В разд. 7.1 мы обсуждаем основные наблюдения в экспериментах по захвату фазы, уделяя особое внимание экспериментам на сердечной и дыхательной системах. В разд. 7.2 мы развиваем математические концепции, необходимые для теоретического анализа явления захвата фазы. Эти концепции используются при анализе захвата фазы в релаксационных моделях в разд. 7.3 и в моделях с предельным циклом в разд. 7.4. В разд. 7.5 мы обсуждаем некоторые явления, связанные с захватом фазы у людей.
). Помимо ритмов с захватом фазы, можно наблюдать также нерегулярные или апериодические движения, при которых фиксированные соотношения фаз и регулярно повторяющиеся циклические процессы не наблюдаются. В разд. 7.1 мы обсуждаем основные наблюдения в экспериментах по захвату фазы, уделяя особое внимание экспериментам на сердечной и дыхательной системах. В разд. 7.2 мы развиваем математические концепции, необходимые для теоретического анализа явления захвата фазы. Эти концепции используются при анализе захвата фазы в релаксационных моделях в разд. 7.3 и в моделях с предельным циклом в разд. 7.4. В разд. 7.5 мы обсуждаем некоторые явления, связанные с захватом фазы у людей.
 Один из методов поддержания постоянного уровня газов в крови заключается в одновременном изменении частоты и объема вентилятора, так что результирующая вентиляция в единицу времени поддерживается постоянной. Однако этот метод не может быть использован, если нужно получить информацию о взаимодействии между респираторным ритмом и искусственной вентиляцией в зависимости от ее частоты и объема (изменяемых независимо). В этом случае можно использовать другой метод, заключающийся в добавлении
Один из методов поддержания постоянного уровня газов в крови заключается в одновременном изменении частоты и объема вентилятора, так что результирующая вентиляция в единицу времени поддерживается постоянной. Однако этот метод не может быть использован, если нужно получить информацию о взаимодействии между респираторным ритмом и искусственной вентиляцией в зависимости от ее частоты и объема (изменяемых независимо). В этом случае можно использовать другой метод, заключающийся в добавлении  к вдыхаемому газу для поддержания постоянных физиологических уровней
к вдыхаемому газу для поддержания постоянных физиологических уровней  даже при больших объемах и частотах вентилятора.
даже при больших объемах и частотах вентилятора.

 — частота вентилятора. Из работы Petrillo and Glass (1984).
— частота вентилятора. Из работы Petrillo and Glass (1984).

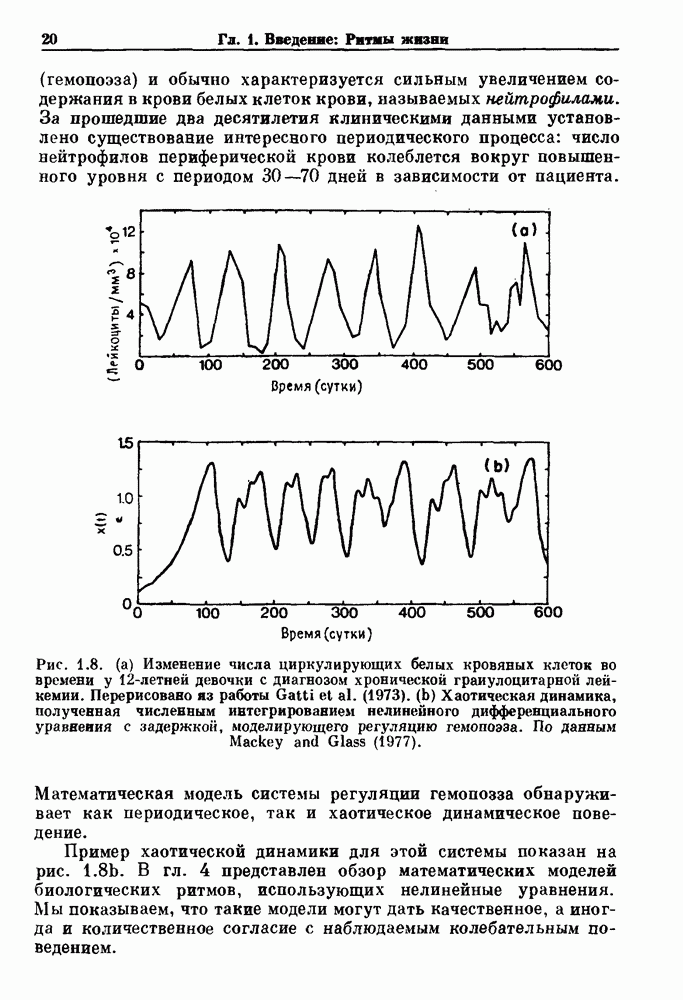
 с ритмом
с ритмом  Таким образом,
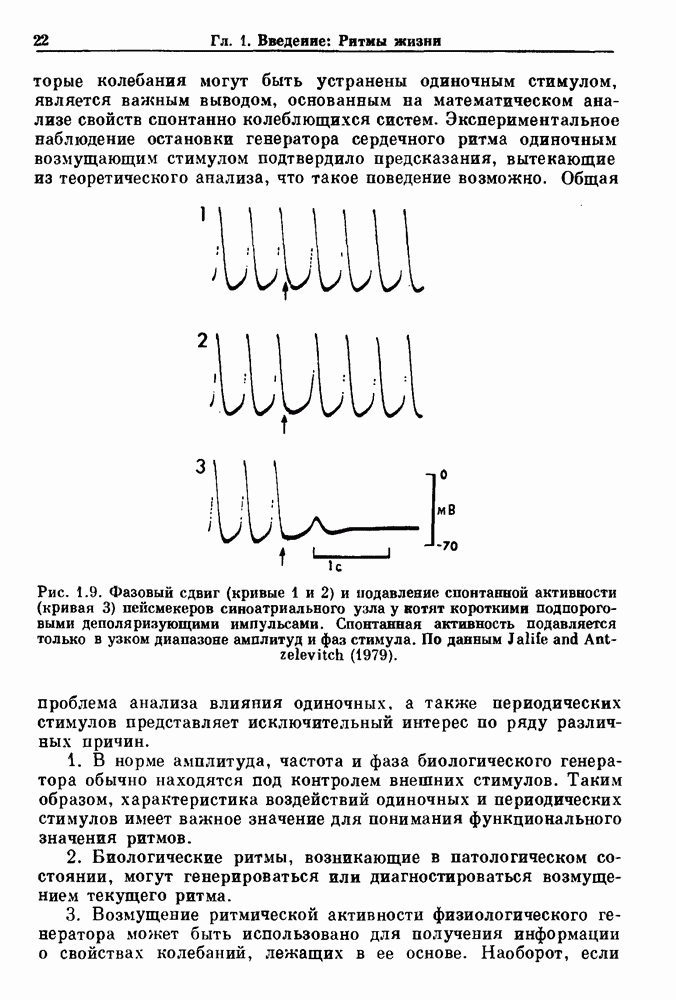
Таким образом,  уменьшается с увеличением частоты стимуляции при фиксированной амплитуде стимула.
уменьшается с увеличением частоты стимуляции при фиксированной амплитуде стимула.