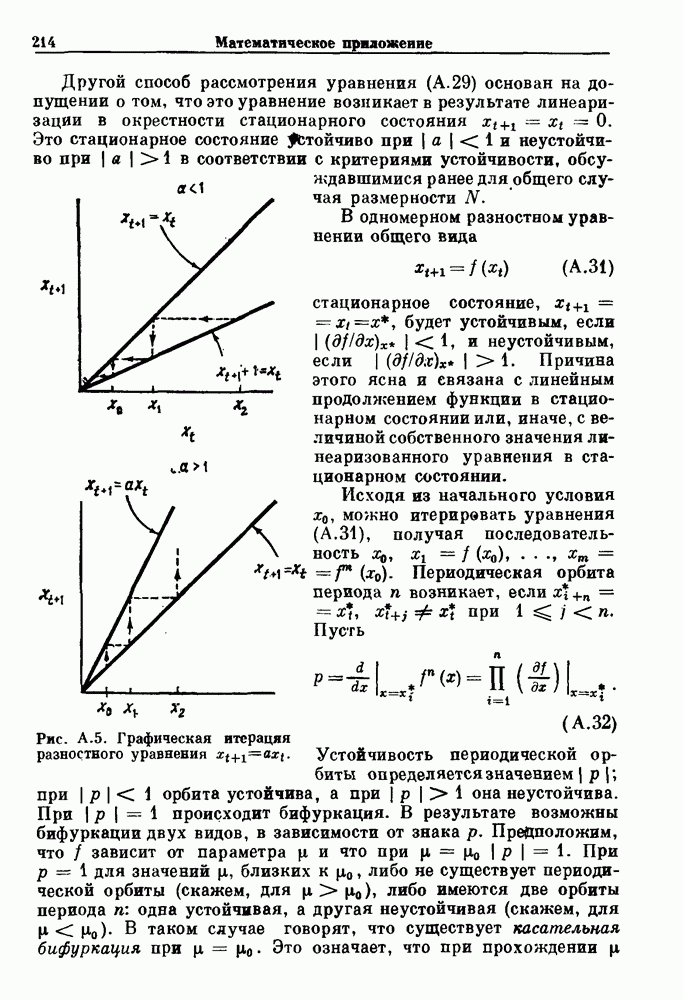
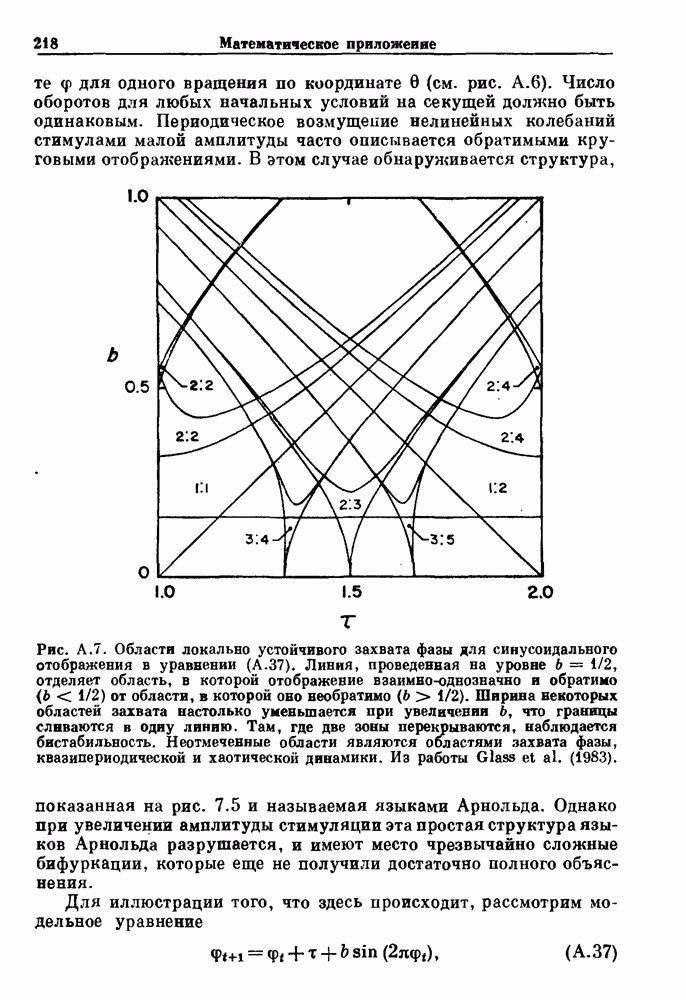
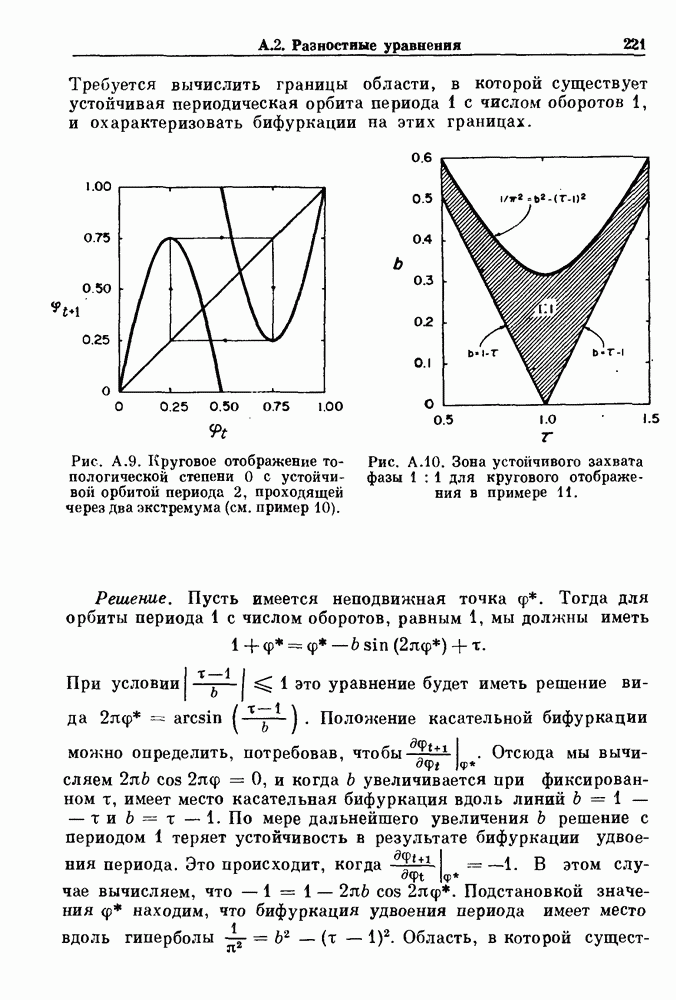
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
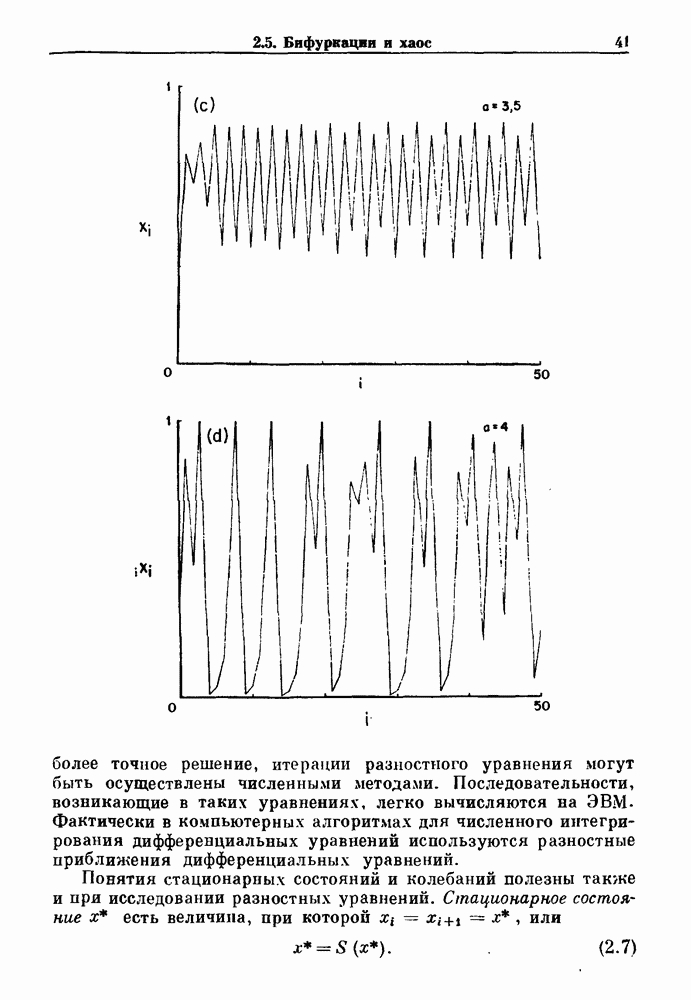
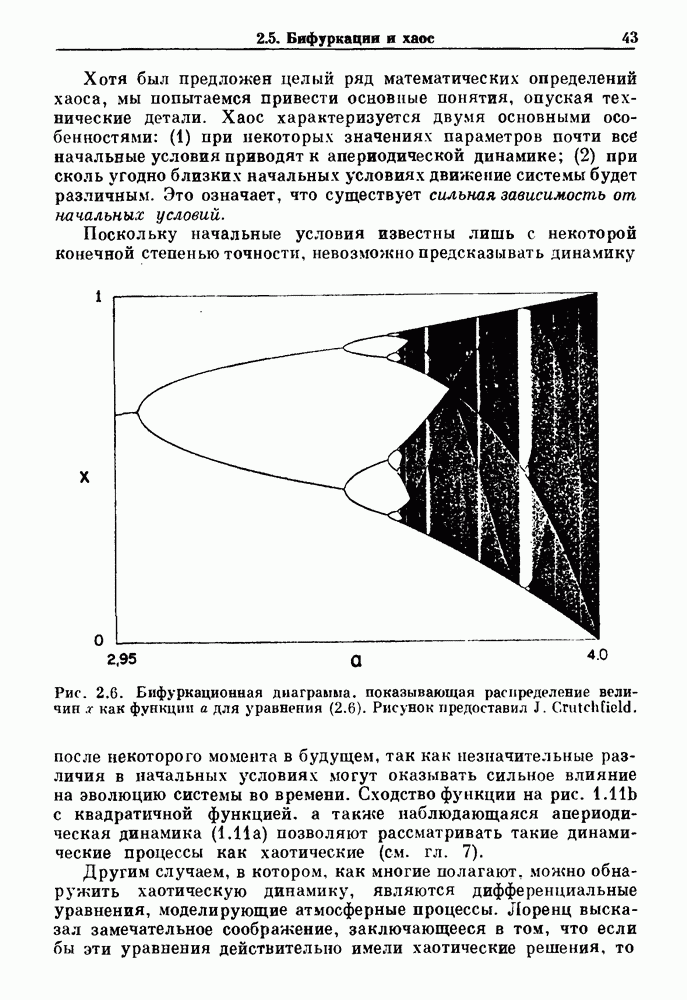
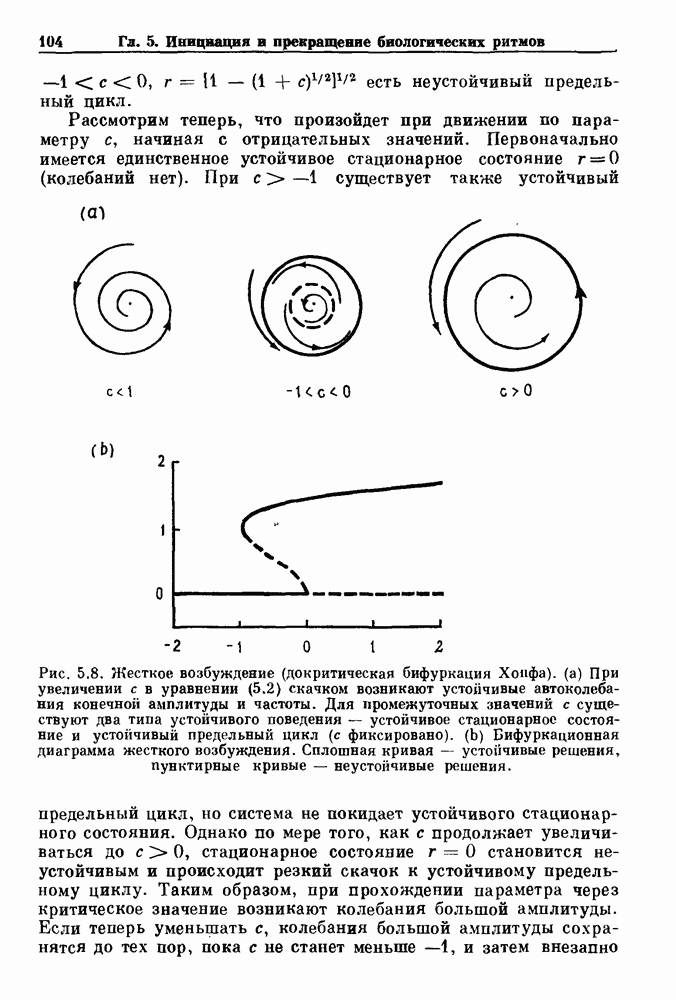
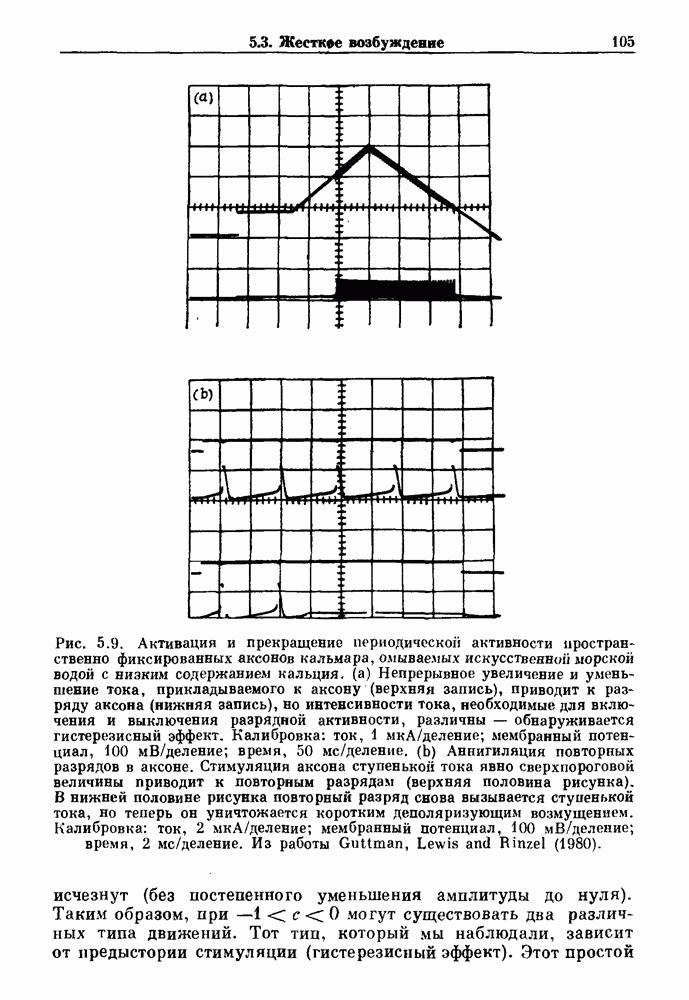
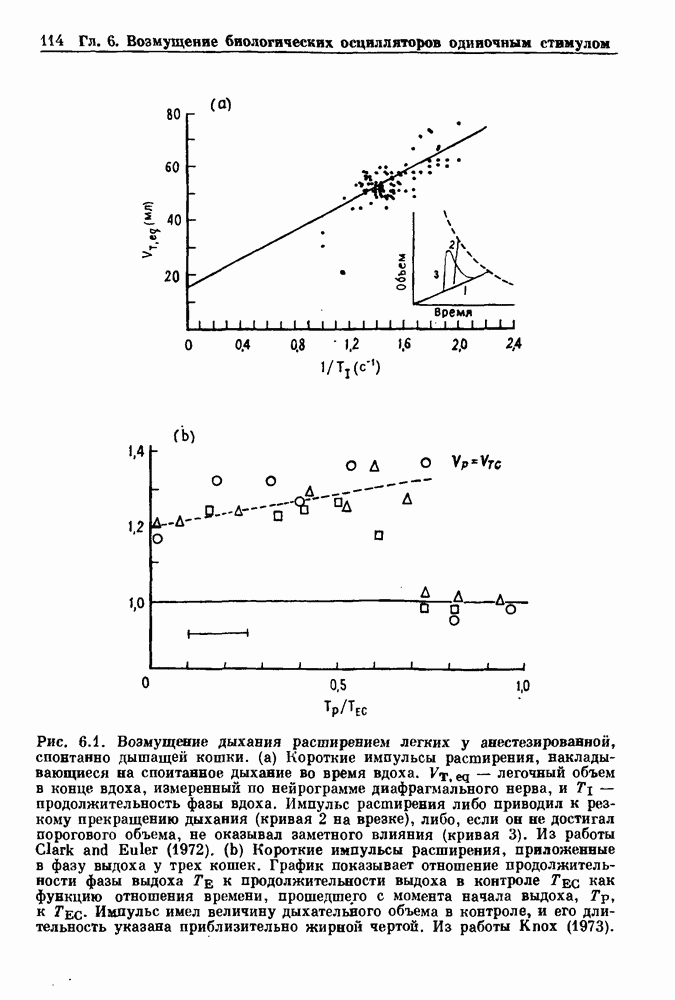
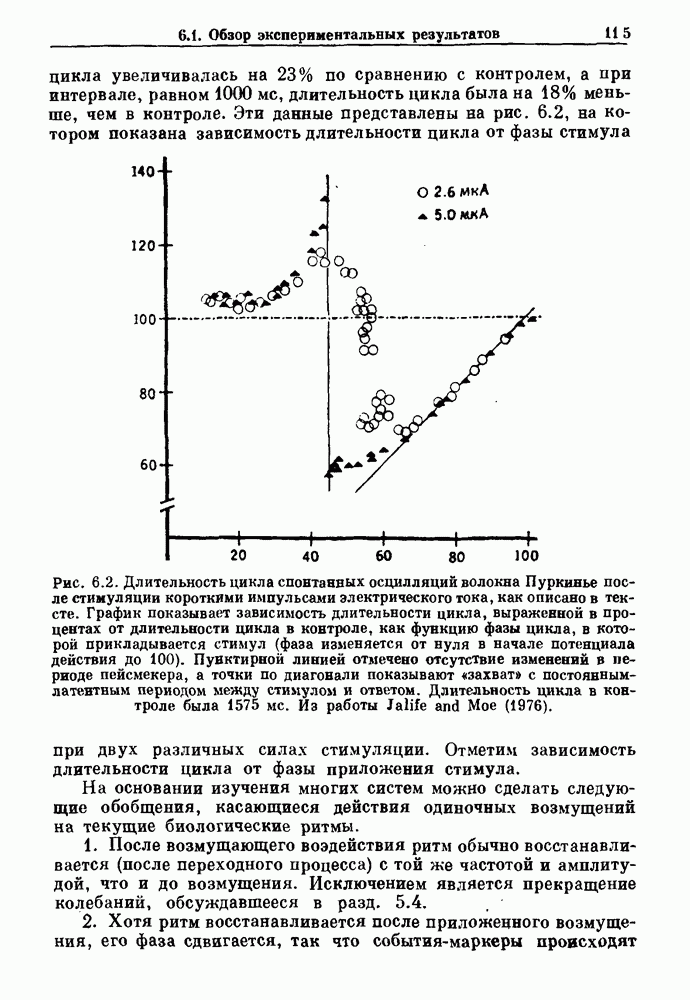
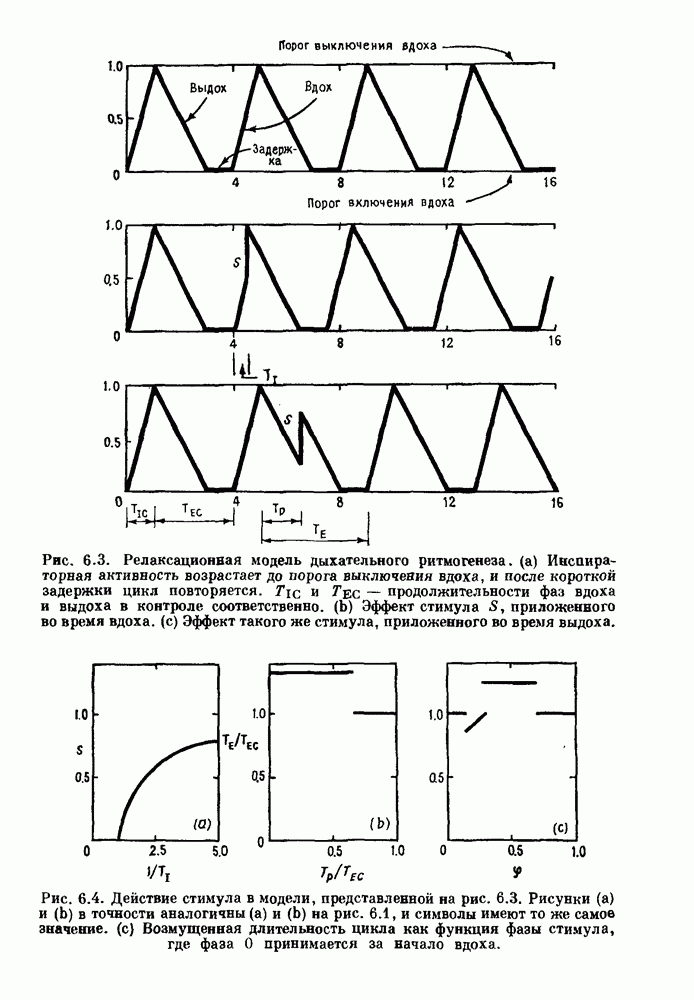
4.6. Колебания в системах с комбинированными обратными связями и временными задержкамиСистемы с положительной обратной связью — это такие системы, в которых отклонения от стационарного состояния стремятся возрасти до некоторого значения. Несмотря на то, что таким отклонениям не придается сколько-нибудь существенного значения в функционировании биологических систем из-за их предполагаемой неустойчивости, часто встречаются ситуации, при которых биологическая обратная связь представляет собой комбинацию положительной и отрицательной обратной связи. Такой тип обратной связи называется комбинированной обратной связью. В качестве примера системы с комбинированной обратной связью, в которой временные задержки играют важную роль, рассмотрим простую модель регуляции образования белых клеток крови. Принято считать, что существует самоподдерживающаяся полипотентная популяция стволовых клеток, способная продуцировать резервные стволовые клетки для эритроцитарной (красные клетки), миелоидной (белые клетки) и мегакариоцитарной (пластинки) клеточных линий. По мере созревания миелоидных стволовых клеток они приобретают морфологические характеристики, позволяющие идентифицировать их как пролиферирующие предшественники миелоидных клеток. В процессе дальнейшего созревания пролиферативная активность этих клеток прекращается, клетки входят в фазу зрелости и их ядра выталкиваются. Зрелые нейтрофилы (один из типов белых клеток крови, возникающих из миелоидных стволовых клеток) высвобождаются из костного мозга в кровь, где они быстро разрушаются случайным образом (время полужизни этих клеток у человека 7 часов). Общее время, необходимое для созревания и высвобождения распознаваемой миелоидной клетки-предшественника составляет у человека примерно 6 дней. Гормональное регулирование происходит на уровне между циркулирующей нейтрофильной массой и миелоидными стволовыми клетками. Уменьшение числа циркулирующих нейтрофилов приводит к образованию и высвобождению гормона грануло-поэтина (ГП). ГП затем действует на популяцию миелоидных стволовых клеток, увеличивая их пролиферативную активность и, в конце концов, увеличивая поток клеток в популяцию распознаваемых предшественников миелоцитов. Пусть х — плотность циркулирующих нейтрофилов (в клетках на кг веса тела), а у — скорость случайного распада нейтрофилов
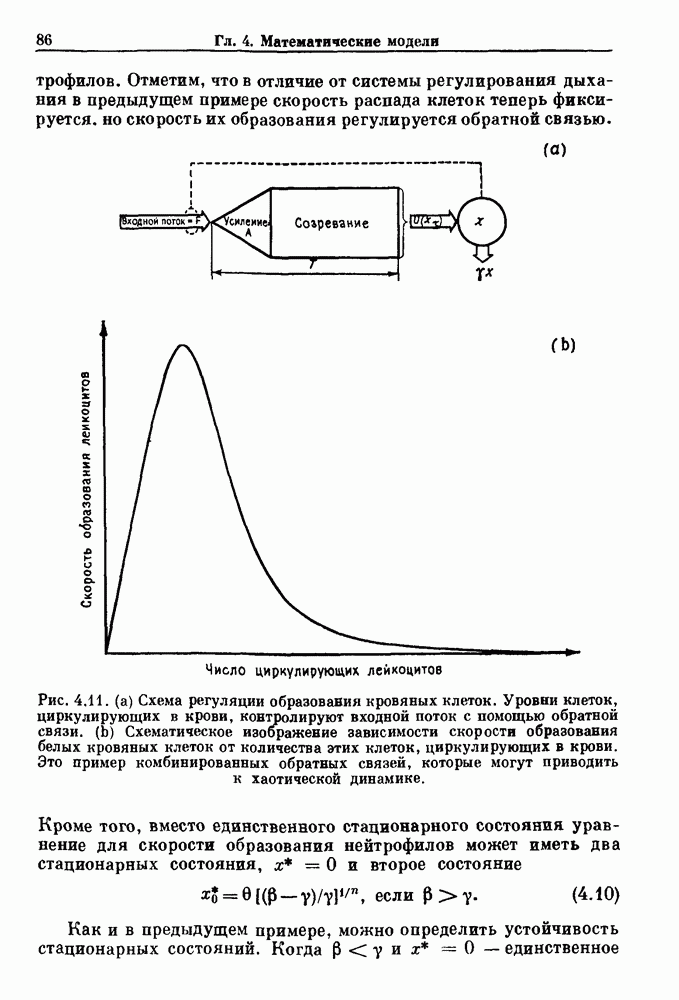
где В широком диапазоне изменений уровня циркулирующих нейтрофилов скорость образования нейтрофилов v падает с увеличением их плотности. Однако благодаря действию различных факторов можно ожидать, что при очень низких уровнях нейтрофилов скорость их образования будет падать, приближаясь к нулю. Таким образом, для v мы выбираем унимодальную функцию (см. рис. 4.l1b)
где Объединяя уравнения (4.7) и (4.8), получаем
нелинейное дифференциальное уравнение с временной задержкой, описывающее динамику изменения числа циркулирующих нейтрофилов Отметим, что в отличие от системы регулирования дыхания в предыдущем примере скорость распада клеток теперь фиксируется. но скорость их образования регулируется обратной связью. (см. скан) Рис. 4.11. (а) Схема регуляции образования кровяных клеток. Уровни клеток, циркулирующих в крови, контролируют входной поток с помощью обратной связи. (b) Схематическое изображение зависимости скорости образования белых кровяных клеток от количества этих клеток, циркулирующих в крови. Это пример комбинированных обратных связей, которые могут приводить к хаотической динамике. Кроме того, вместо единственного стационарного состояния уравнение для скорости образования нейтрофилов может иметь два стационарных состояния,
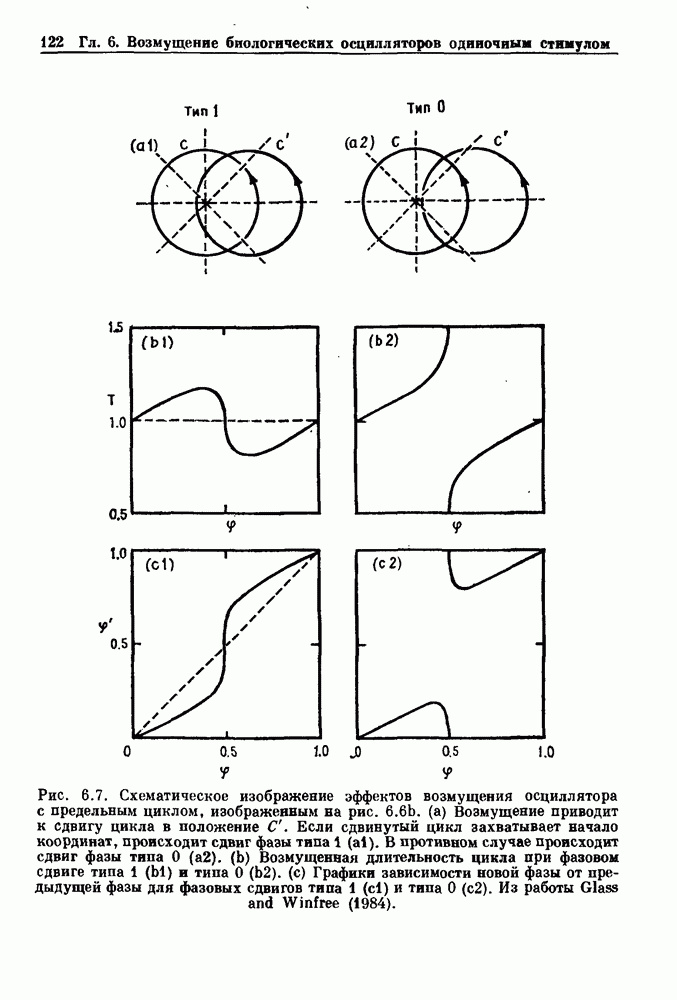
Как и в предыдущем примере, можно определить устойчивость стационарных состояний. Когда стационарное состояние, оно всегда устойчиво, как и следует ожидать в ситуации, когда максимальная скорость притока клеток меньше скорости разрушения нейтрофилов. В случае когда Для здоровых людей получены следующие оценки значений параметров: В качестве еще одного примера генерации периодического и апериодического поведения в системах с временной задержкой и комбинированной обратной сиязью рассмотрим процесс рекуррентного угнетения, который был описан почти для всех типов нервной ткани у разных видов от низших беспозвоночных до человека. Этот процесс характеризуется тем, что пресинаптические клетки доставляют возбуждение к постсинаптическим клеткам. Постсинаптические клетки затем генерируют потенциалы действия, один из эффектов которых заключается в активации угнетающих промежуточных нейронов через коллатерали аксонов постсинаптических клеток.
Рис. 4.12. Схема сети с рекуррентным угнетенней. Эти промежуточные нейроны, в свою очередь, передают угнетающее воздействие обратно к постсинаптическим клеткам, от которых они получили первоначальную активацию. В исследовании, специально направленном на выяснение динамических свойств рекуррентной угнетающей цепи в гиппокампе, рассматривался комплекс, состоящий из пирамидальных клеток поля САЗ, мшистого волокна и корзинчатых клеток (рис. 4.12). В этой модели, сформулированной в рамках модели (4.2), Комплекс, состоящий из пирамидальных клеток поля САЗ, мшистого волокна и корзинчатых клеток, хорошо изучен, и соответствующие параметры этой системы можно легко оценить. Более того, известно, что ингибирующим нейротрансмиттером между корзинчатыми клетками и пирамидальными клетками поля САЗ является гамма-аминобутировая кислота (GABA) и что пенициллин связывается практически необратимо с рецепторами GABA на мембране пирамидальных клеток поля САЗ.
Рис. 4.13. Результат численного моделирования сети, представленной на рис. 4.12. N — мера плотности рецепторов. С уменьшением плотности ритм меняется от регулярных вспышек с различными периодами до постоянных, но нерегулярных разрядов. По данным Mackey and an der Heiden (1984). Таким образом, пенициллин может быть использован для титрования числа доступных рецепторов GAB А в популяции постсинаптических клеток, и поэтому естественно было бы изучить поведение модели этой системы по мере уменьшения плотности рецепторов GABA, соответствующего увеличению уровня пенициллина. На рис. 4.13 показано поведение этой простой модели рекуррентного угнетения как функции числа рецепторов GAB А. По мере уменьшения плотности рецепторов, соответствующего введению пенициллина, наблюдается прогрессивный сдвиг клеточной активности от регулярных всплесков активности с различными периодами до установившегося поведения, характеризующегося устойчивыми, но нерегулярными разрядами при малом числе рецепторов. Хаотическое поведение решений дифференциальных уравнений с временной задержкой и комбинированной обратной связью наблюдалось при моделировании различных явлений в физиологических и экологических системах. Несмотря на явную концептуальную простоту и физиологическую важность систем с обратной связью, содержащих временные задержки, математический анализ таких систем чрезвычайно труден. Один из перспективных подходов основан на представлении нелинейностей в уравнениях в виде кусочно-линейных функций. В таких случаях становится возможным прямое интегрирование уравнений при некоторых начальных условиях. Этот метод был применен к системам с комбинированной обратной связью, и оказалось возможным продемонстрировать целую иерархию бифуркационных переходов между периодическими решениями, а также между хаотическими непериодическими решениями при изменении величин параметров. Во многих других исследованиях были использованы кусочнопостоянные нелинейности или другие типы нелинейностей в дифференциальных уравнениях с временной задержкой для выяснения свойств их решений.
|
 |
Оглавление
|




















































































































































































 — приток новых нейтрофилов в кровь, измеряемый в клетках/кг/час, F (клетки/кг/час) — приток резервных миелоидных клеток-предшественников в популяцию распознаваемых предшественников миелоцитов и А — коэффициент усиления потока в данную популяцию (см. рис. 4.11 а). Тогда, исходя из вышеприведенного описания системы образования нейтрофилов, имеем
— приток новых нейтрофилов в кровь, измеряемый в клетках/кг/час, F (клетки/кг/час) — приток резервных миелоидных клеток-предшественников в популяцию распознаваемых предшественников миелоцитов и А — коэффициент усиления потока в данную популяцию (см. рис. 4.11 а). Тогда, исходя из вышеприведенного описания системы образования нейтрофилов, имеем

 — текущий приток клеток в кровь, возникающий в ответ на запрос, создавшийся в некоторый момент времени
— текущий приток клеток в кровь, возникающий в ответ на запрос, создавшийся в некоторый момент времени  в прошлом.
в прошлом.

 — параметры.
— параметры.

 и второе состояние
и второе состояние

 единственное
единственное
 и имеются два стационарных состояния, состояние
и имеются два стационарных состояния, состояние  всегда неустойчиво, в то время как второе стационарное состояние, задаваемое уравнением (4.10), может быть либо устойчивым, либо неустойчивым в зависимости от значений параметров
всегда неустойчиво, в то время как второе стационарное состояние, задаваемое уравнением (4.10), может быть либо устойчивым, либо неустойчивым в зависимости от значений параметров  . Условие устойчивости второго стационарного состояния имеет сложный вид, и мы не приводим его здесь (см. математическое приложение). Достаточно сказать, что увеличение
. Условие устойчивости второго стационарного состояния имеет сложный вид, и мы не приводим его здесь (см. математическое приложение). Достаточно сказать, что увеличение  и/или
и/или  может привести к дестабилизации стационарного состояния, и в том случае, когда это происходит, число циркулирующих нейтрофилов колеблется с периодом, значение которого лежит в интервале от
может привести к дестабилизации стационарного состояния, и в том случае, когда это происходит, число циркулирующих нейтрофилов колеблется с периодом, значение которого лежит в интервале от  до
до  Изменения скорости случайного разрушения нейтрофилов имеют более сложный характер, так как оказывается, что неустойчивость может возникать либо при увеличении, либо при уменьшении у, в зависимости от значений других параметров. Такой анализ, однако, далек от выявления всей возможной динамики, которая может возникать в уравнении (4.9). Для того чтобы исследовать более сложную динамику, мы должны отказаться от имеющихся аналитических методов в пользу численного интегрирования.
Изменения скорости случайного разрушения нейтрофилов имеют более сложный характер, так как оказывается, что неустойчивость может возникать либо при увеличении, либо при уменьшении у, в зависимости от значений других параметров. Такой анализ, однако, далек от выявления всей возможной динамики, которая может возникать в уравнении (4.9). Для того чтобы исследовать более сложную динамику, мы должны отказаться от имеющихся аналитических методов в пользу численного интегрирования.
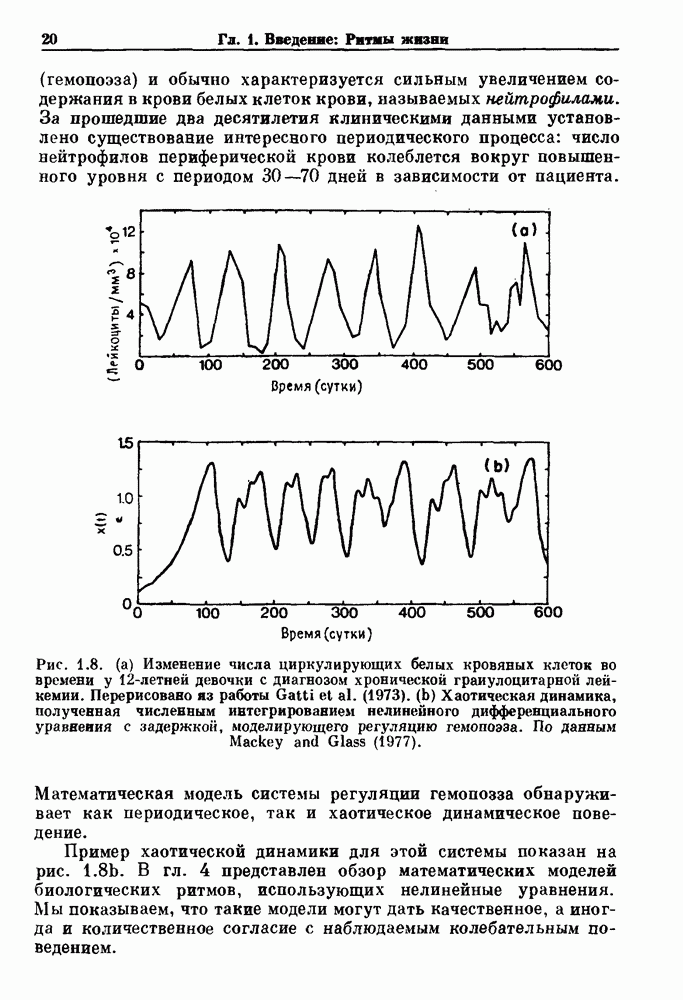
 суток. При таких значениях параметров анализ устойчивости предсказывает, что число нейтрофилов в стационарном состоянии не должно оставаться постоянным. Действительно, численное интегрирование уравнения (4.9) при этих значениях показывает наличие мягко возникающих колебаний числа нейтрофилов с периодом около 20 суток. На рис. 1.8b показаны результаты такого же численного интегрирования при увеличении
суток. При таких значениях параметров анализ устойчивости предсказывает, что число нейтрофилов в стационарном состоянии не должно оставаться постоянным. Действительно, численное интегрирование уравнения (4.9) при этих значениях показывает наличие мягко возникающих колебаний числа нейтрофилов с периодом около 20 суток. На рис. 1.8b показаны результаты такого же численного интегрирования при увеличении  до 20 суток. Отметим чрезвычайную нерегулярность решения полностью детерминированного уравнения (4.9). Эта простая модель, представленная уравнением (4.9), еще раз демонстрирует возможность существования хаоса в детерминированной, непрерывной во времени системе. В данном случае уровни циркулирующих нейтрофилов случайны просто вследствие уравнений, описывающих их собственную эволюцию. Как обсуждалось в гл. 1, рассматриваемые здесь динамические свойства обнаруживают по крайней мере качественное сходство с данными, полученными при определении числа белых кровяных клеток у пациента с хронической миелоидной лейкемией (XMЛ). Некоторые исследователи полагают, что XMЛ обычно сопровождается увеличением переходного времени
до 20 суток. Отметим чрезвычайную нерегулярность решения полностью детерминированного уравнения (4.9). Эта простая модель, представленная уравнением (4.9), еще раз демонстрирует возможность существования хаоса в детерминированной, непрерывной во времени системе. В данном случае уровни циркулирующих нейтрофилов случайны просто вследствие уравнений, описывающих их собственную эволюцию. Как обсуждалось в гл. 1, рассматриваемые здесь динамические свойства обнаруживают по крайней мере качественное сходство с данными, полученными при определении числа белых кровяных клеток у пациента с хронической миелоидной лейкемией (XMЛ). Некоторые исследователи полагают, что XMЛ обычно сопровождается увеличением переходного времени  между фазами созревания клеток и, таким образом, оказывается возможной связь между теоретической моделью и предложенными механизмами заболевания.
между фазами созревания клеток и, таким образом, оказывается возможной связь между теоретической моделью и предложенными механизмами заболевания.

 характеризует частоту разряда популяции пирамидальных клеток поля САЗ. «Образование» X переменной х целиком обусловлено возбуждающей активностью популяции мшистых волокон. Однако Распад х определяется двумя различными процессами: (естественным угасанием активности, которое происходит из-за злектротонических свойств мембраны пирамидальных клеток САЗ, и (2) нелинейным типом обратной связи, аналогичным тому, который был использован для описания образования нейтрофилов (уравнение (4.8)). Такая связь обусловлена существованием рекуррентного угнетающего пути, образованного корзинчатыми клетками. Кроме того, в генерации рекуррентного угнетения имеется временная задержка, обусловленная временем проведения импульсов и синаптическими задержками в петле обратной связи.
характеризует частоту разряда популяции пирамидальных клеток поля САЗ. «Образование» X переменной х целиком обусловлено возбуждающей активностью популяции мшистых волокон. Однако Распад х определяется двумя различными процессами: (естественным угасанием активности, которое происходит из-за злектротонических свойств мембраны пирамидальных клеток САЗ, и (2) нелинейным типом обратной связи, аналогичным тому, который был использован для описания образования нейтрофилов (уравнение (4.8)). Такая связь обусловлена существованием рекуррентного угнетающего пути, образованного корзинчатыми клетками. Кроме того, в генерации рекуррентного угнетения имеется временная задержка, обусловленная временем проведения импульсов и синаптическими задержками в петле обратной связи.
