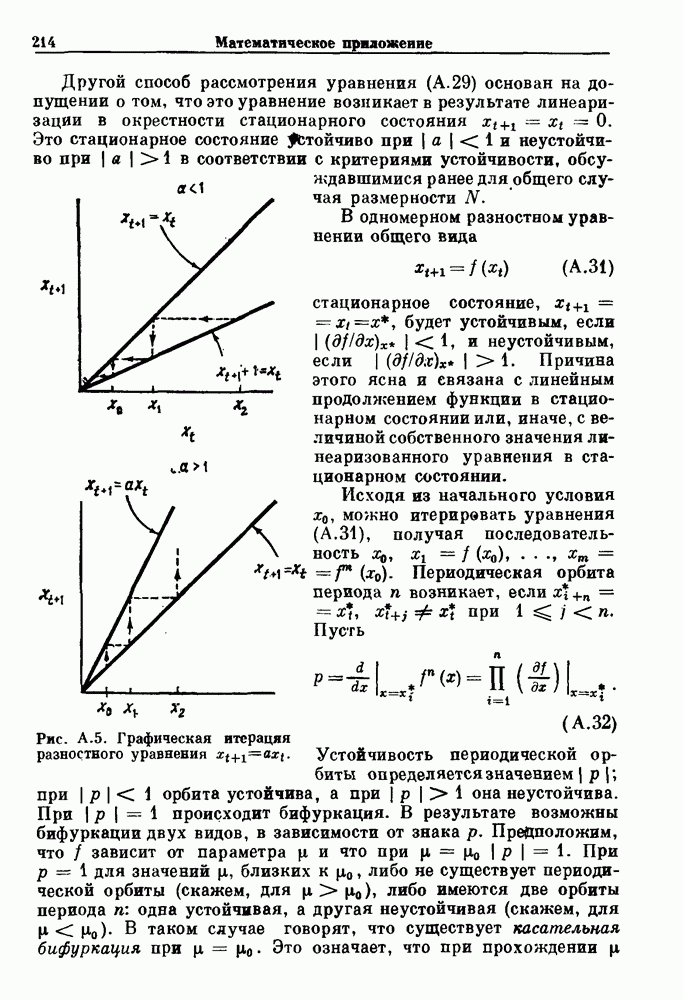
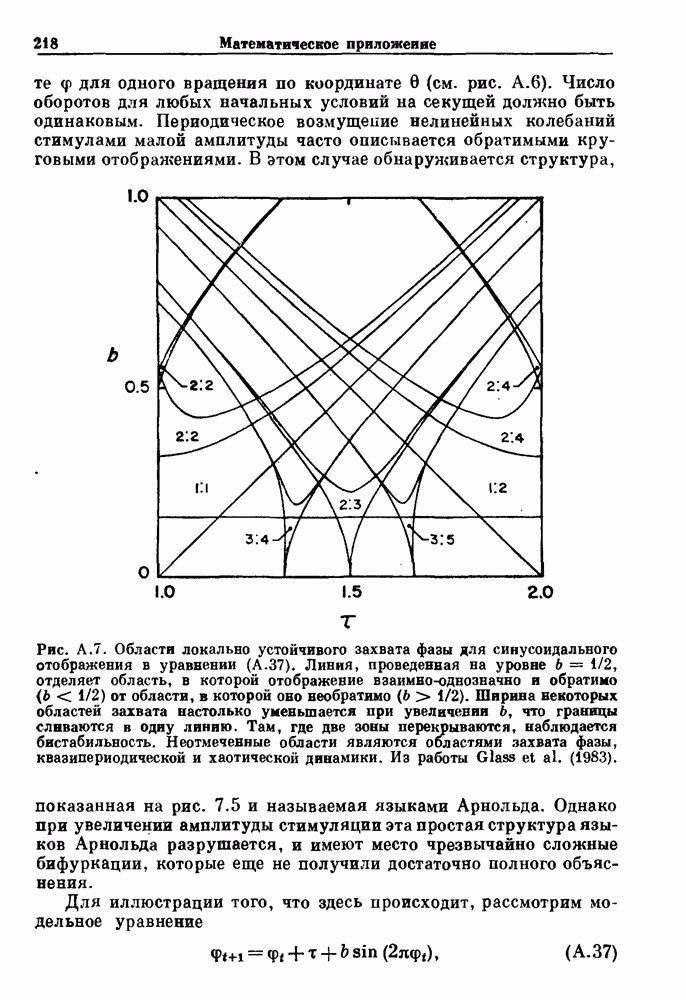
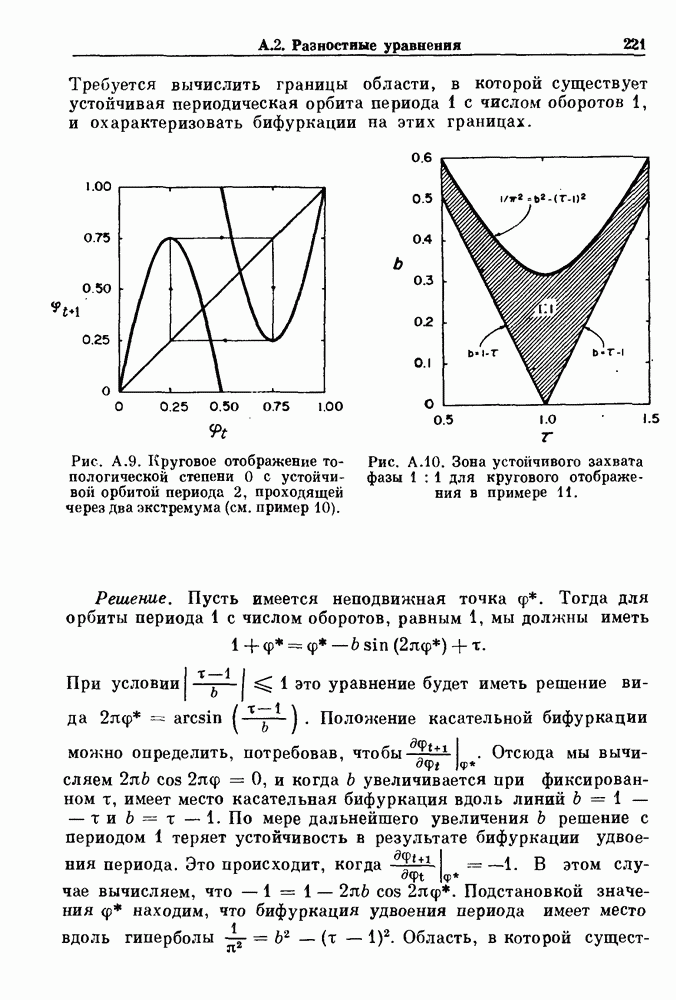
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
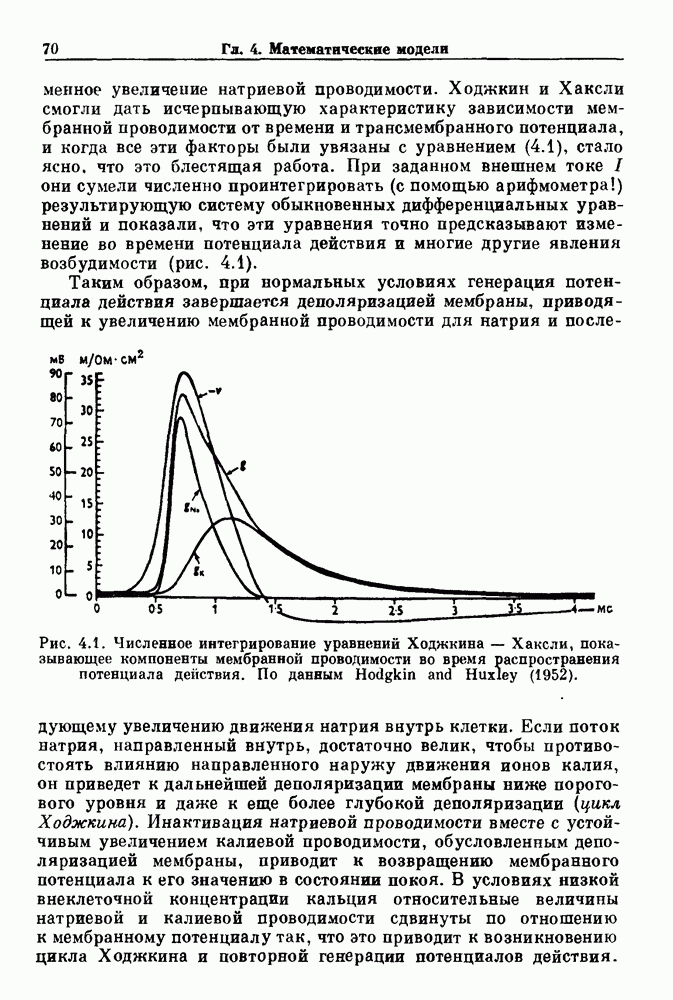
ZADANIA.TO
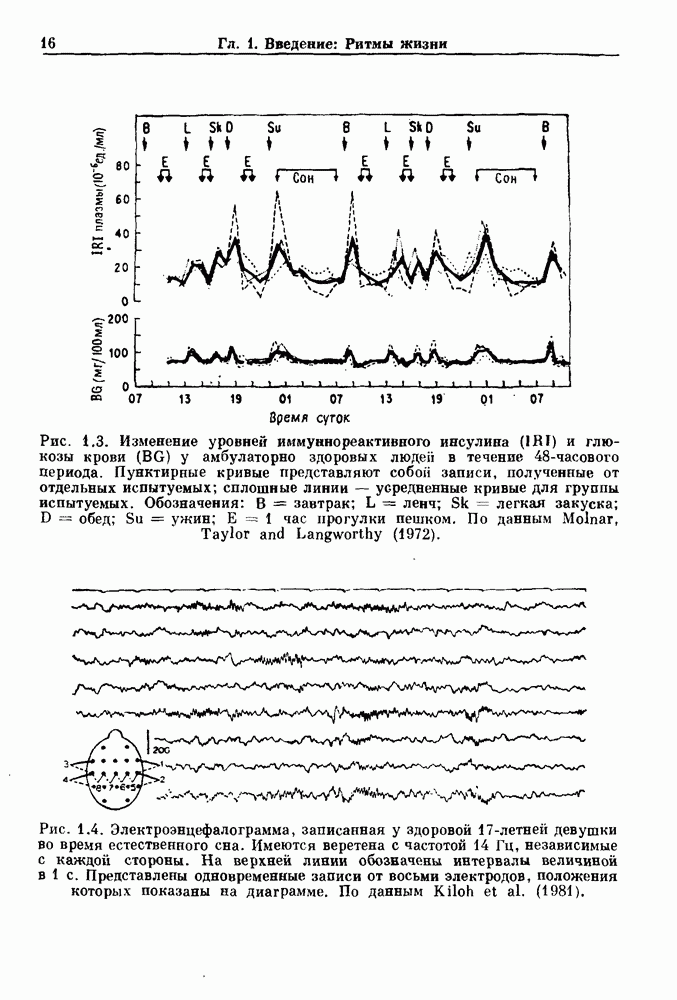
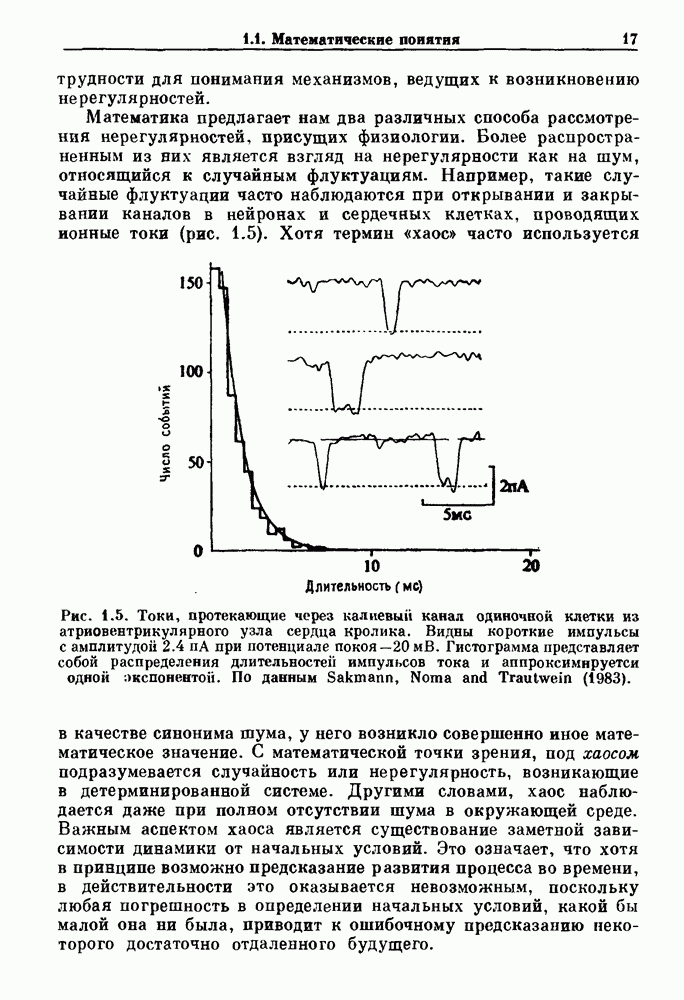
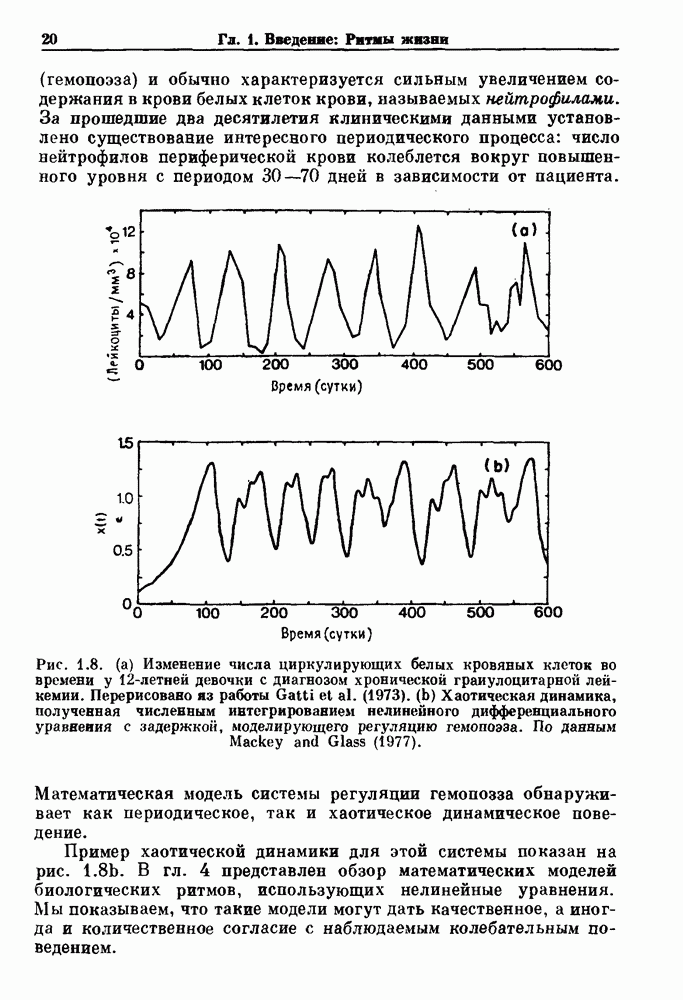
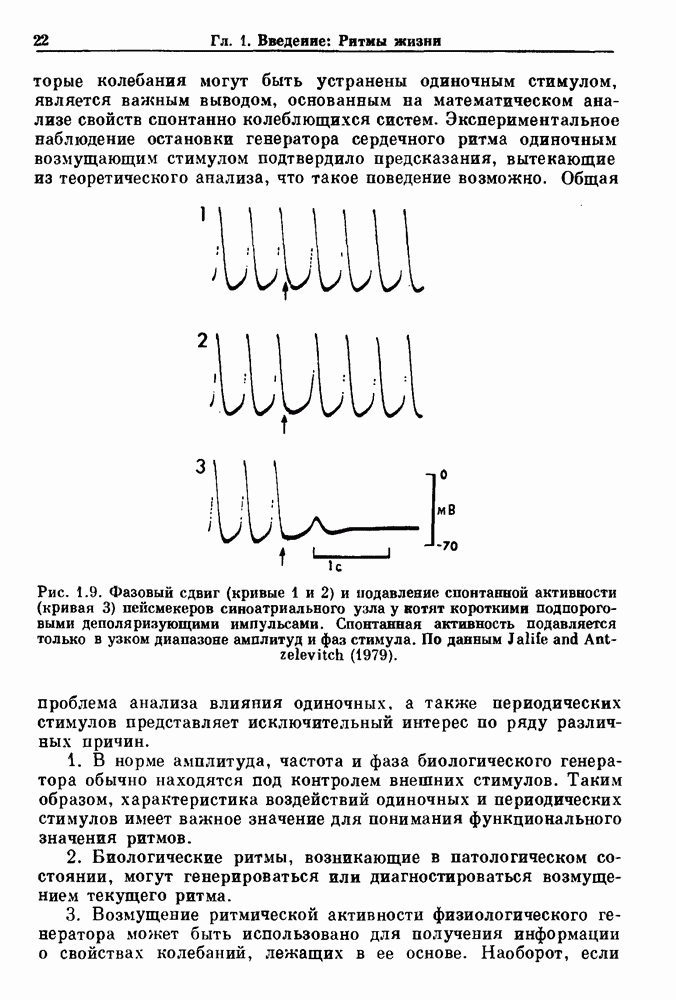
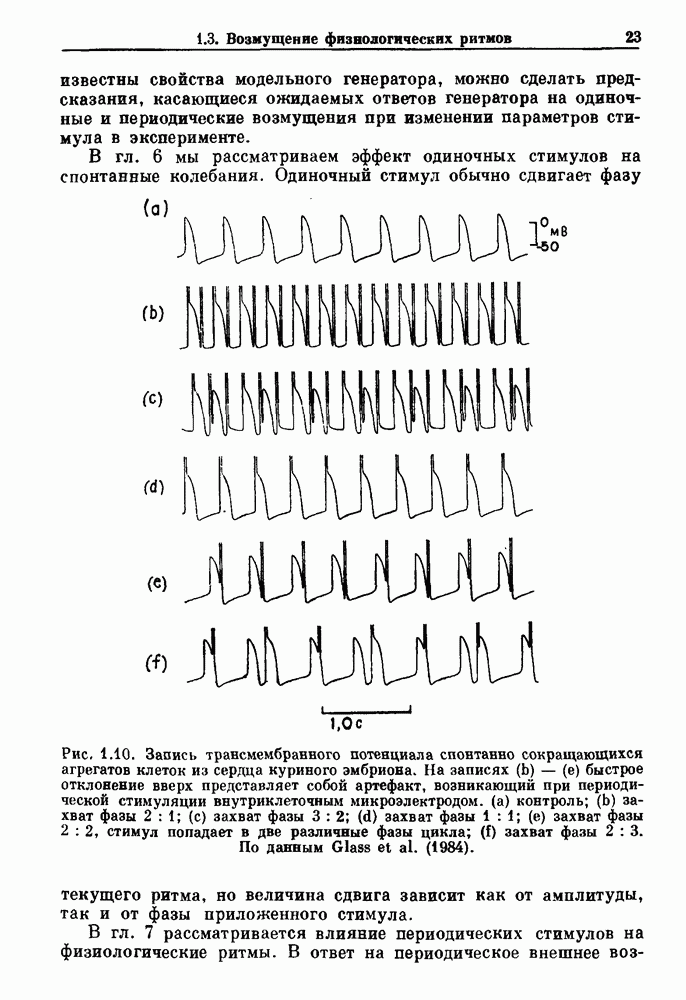
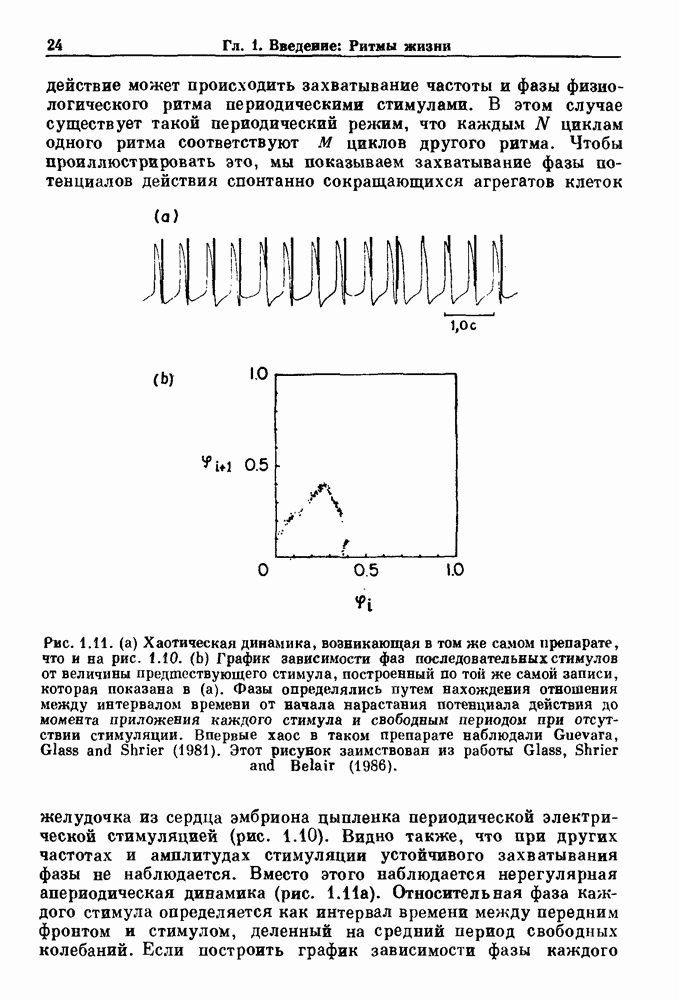
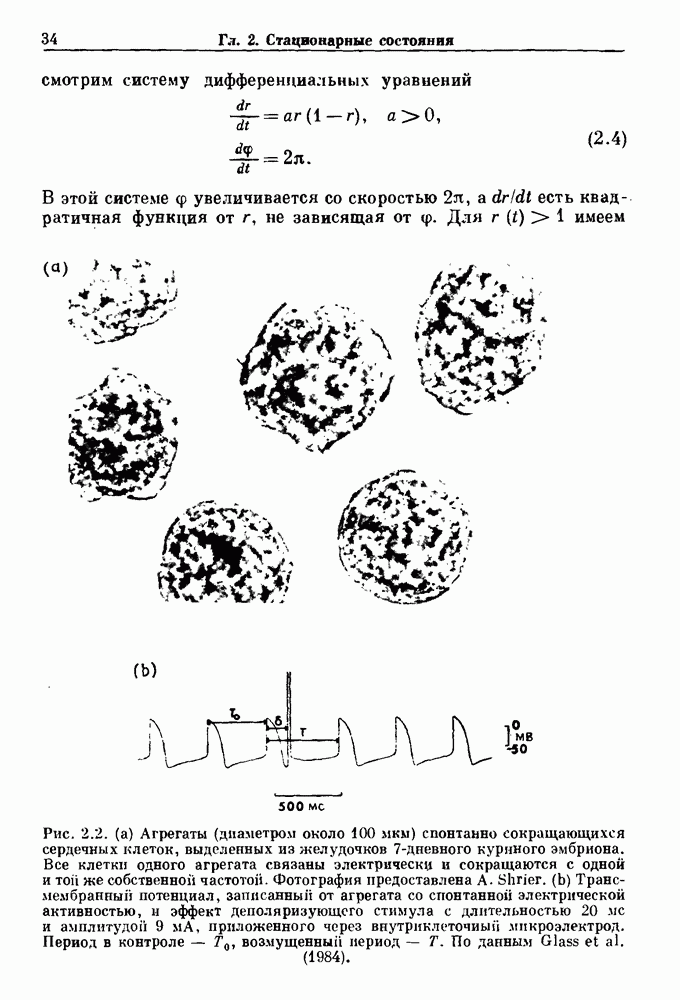
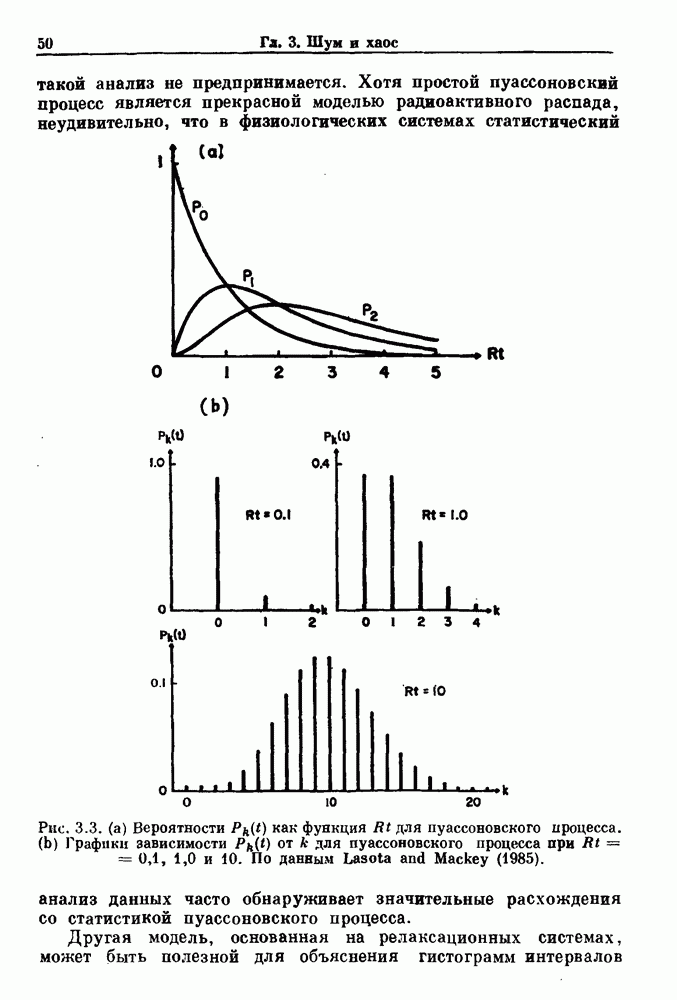
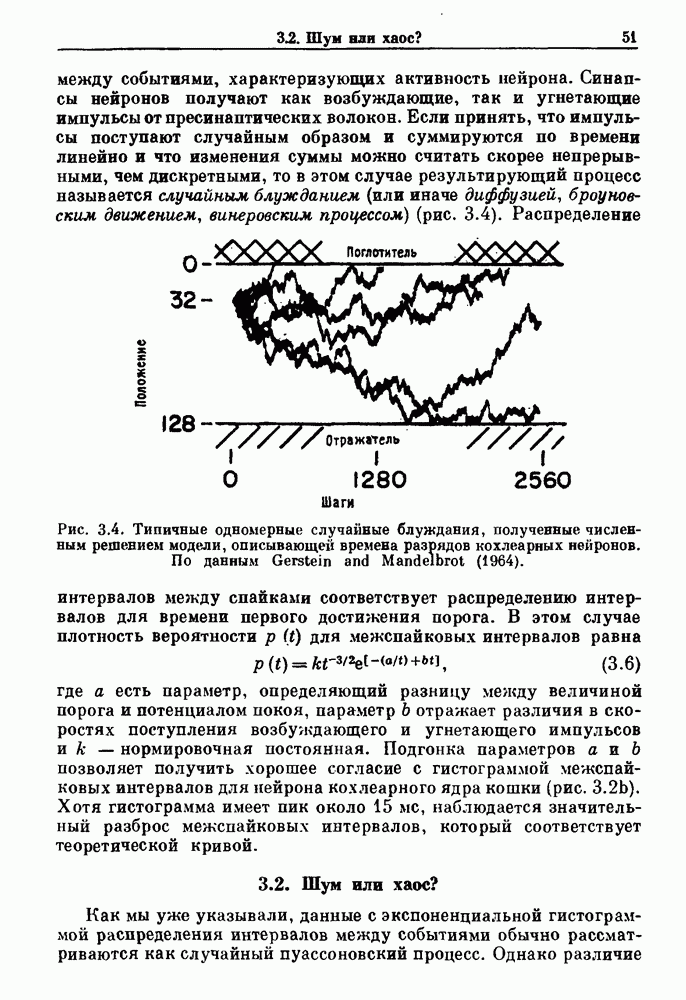
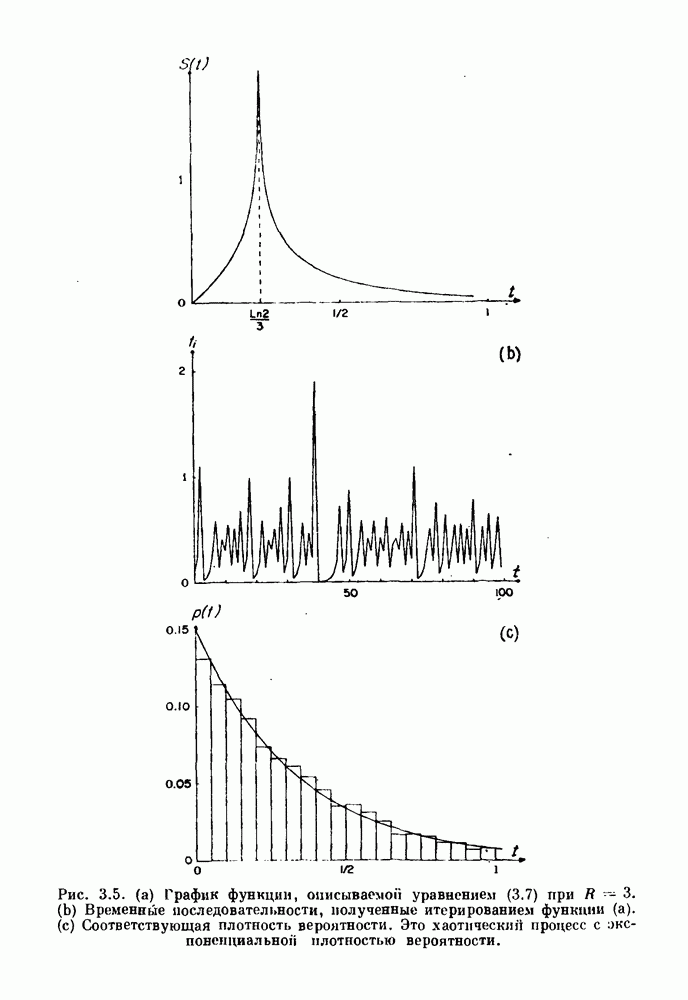
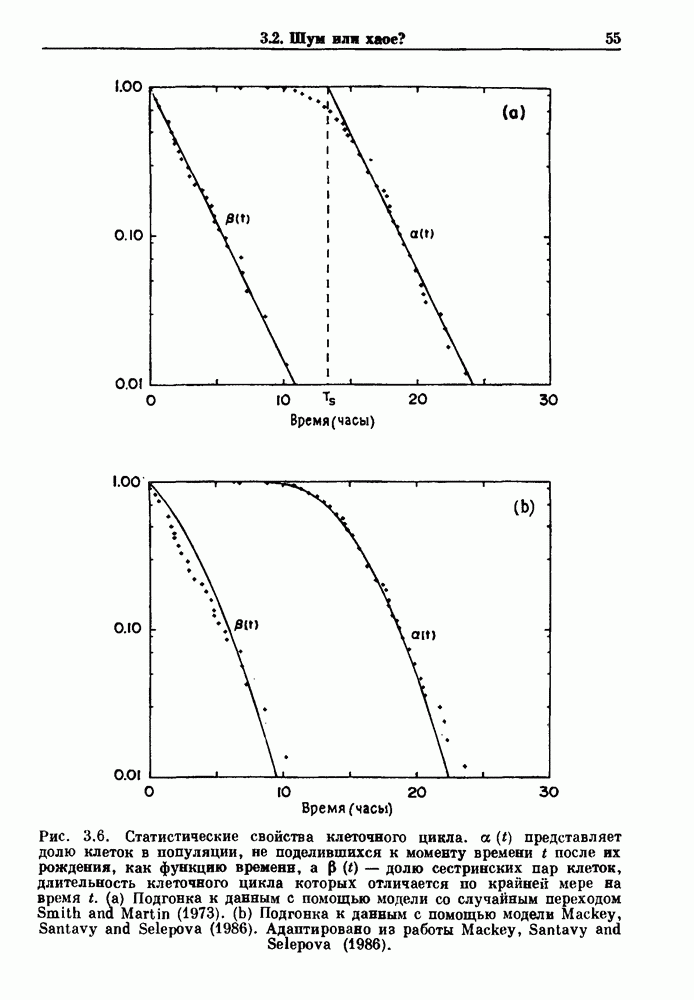
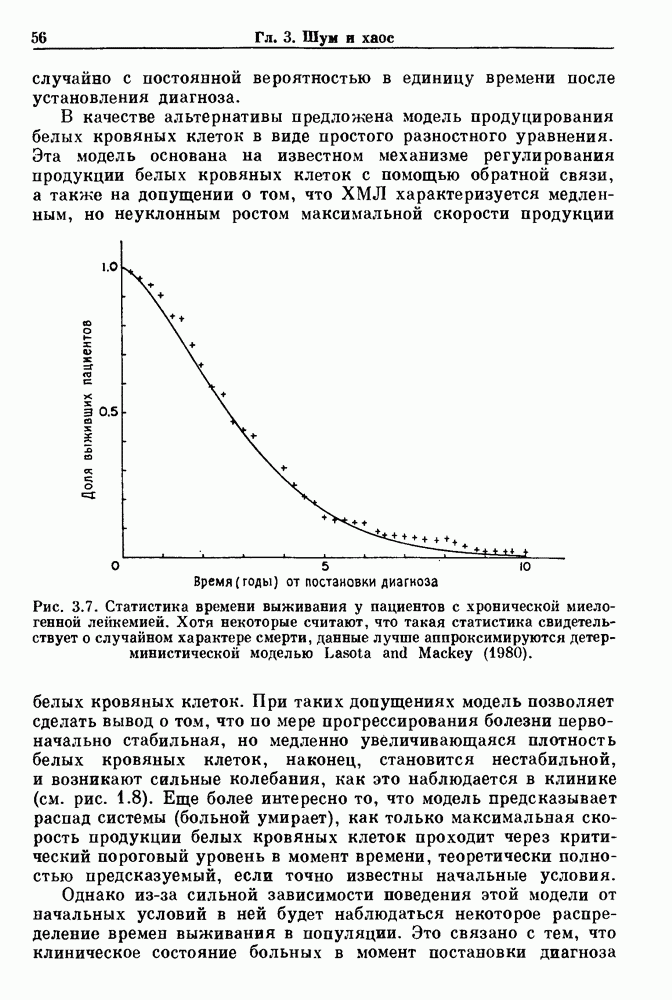
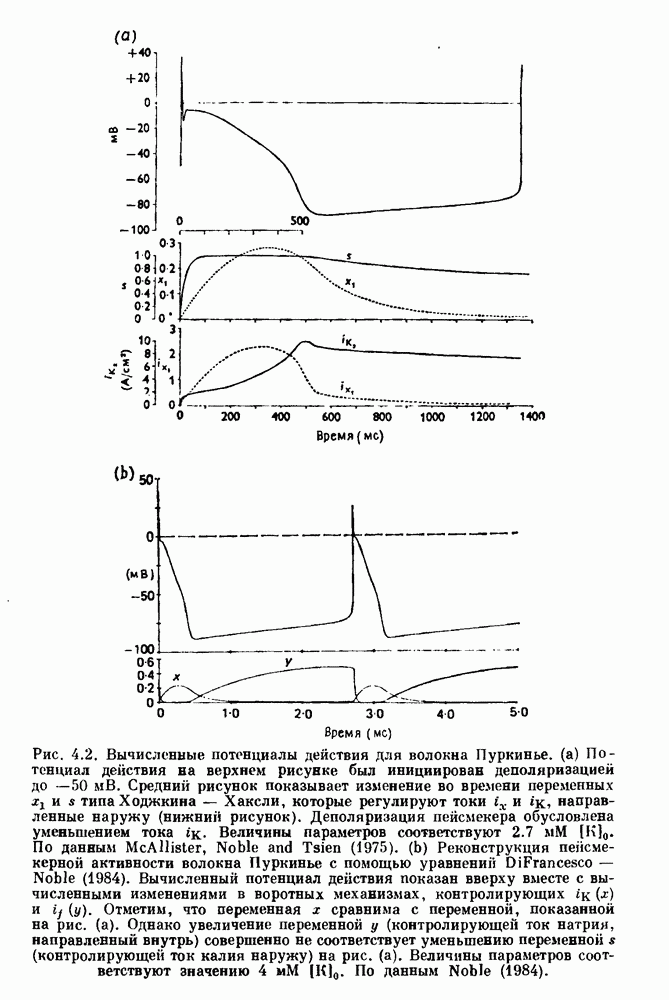
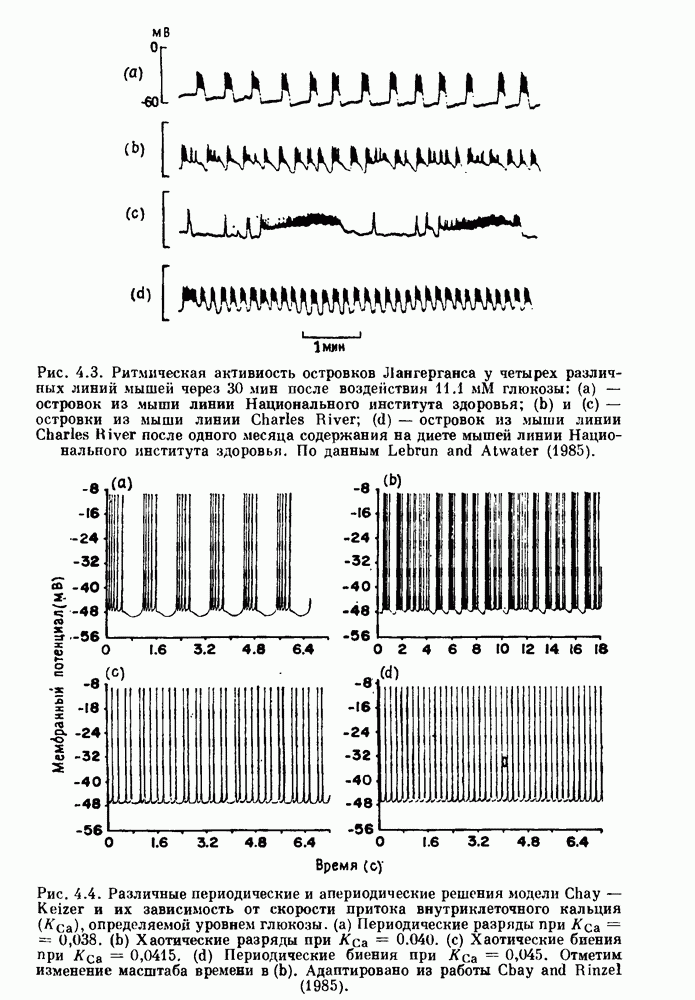
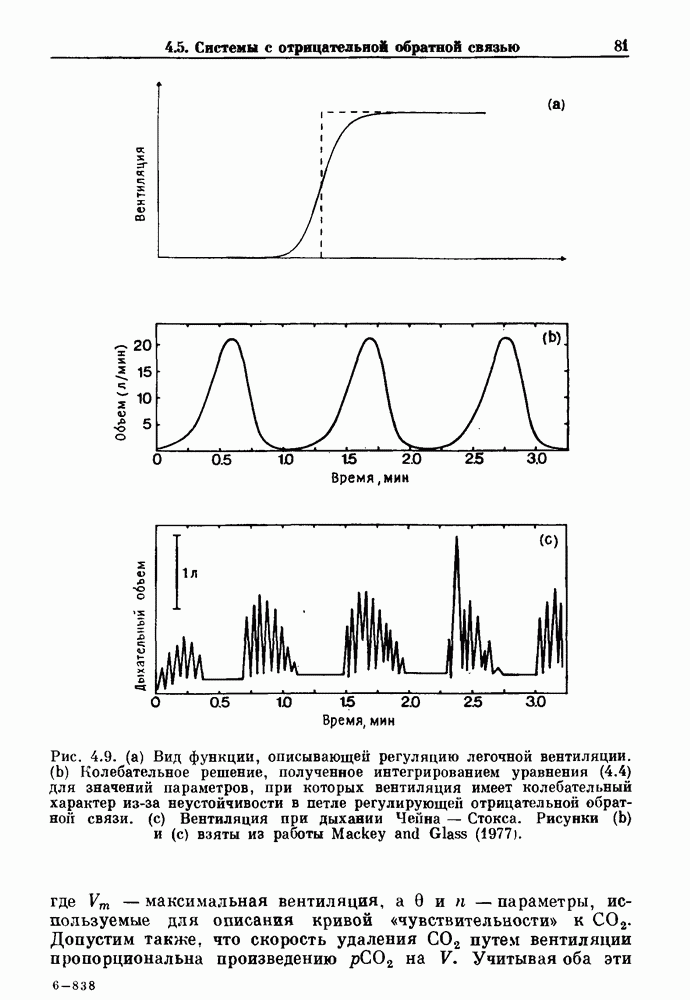
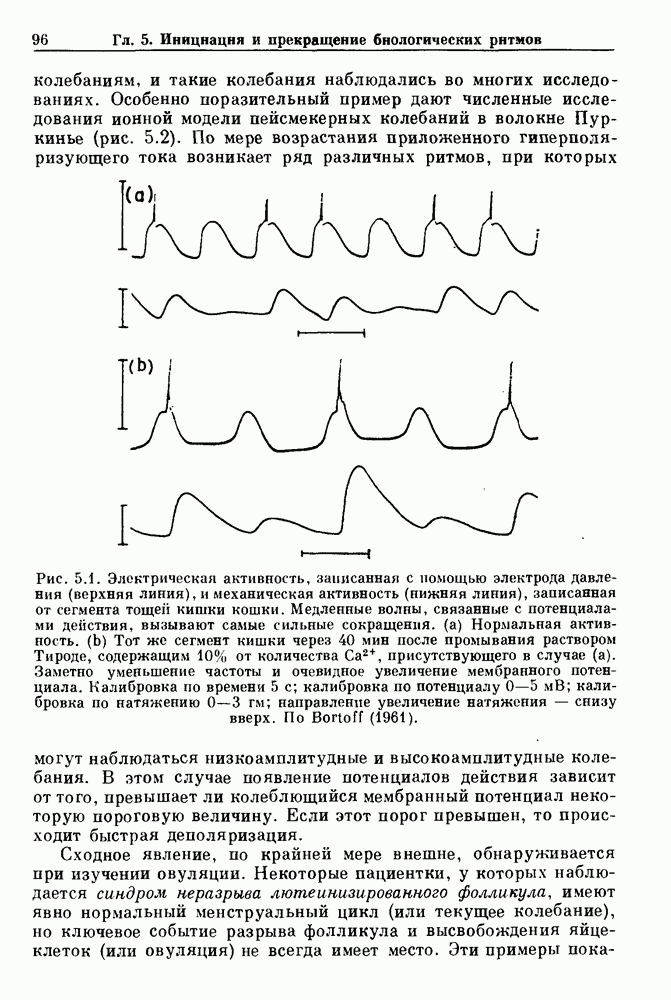
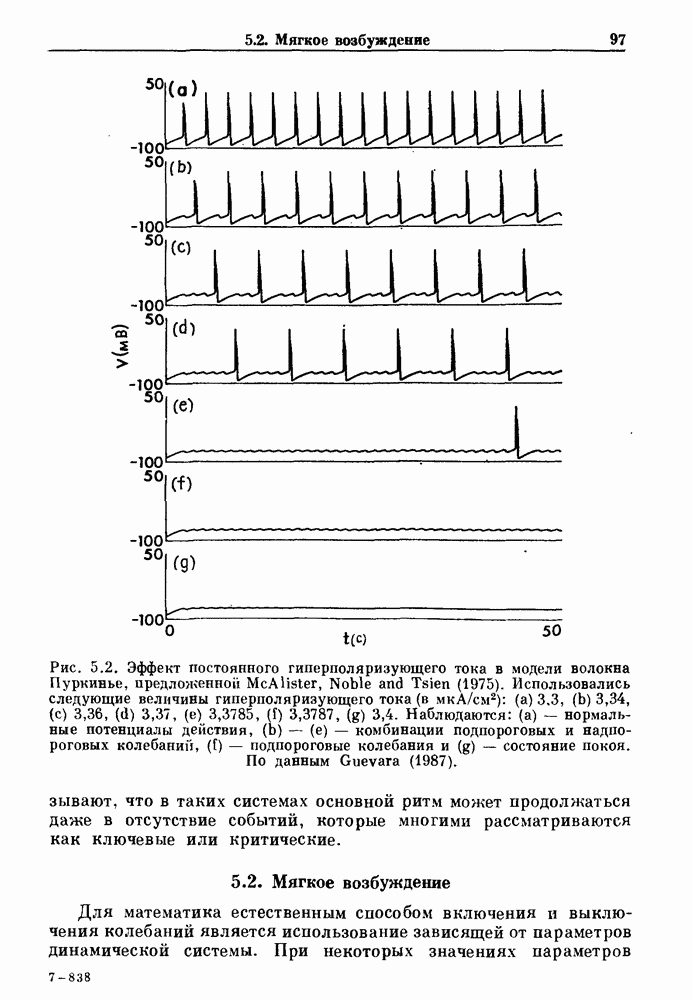
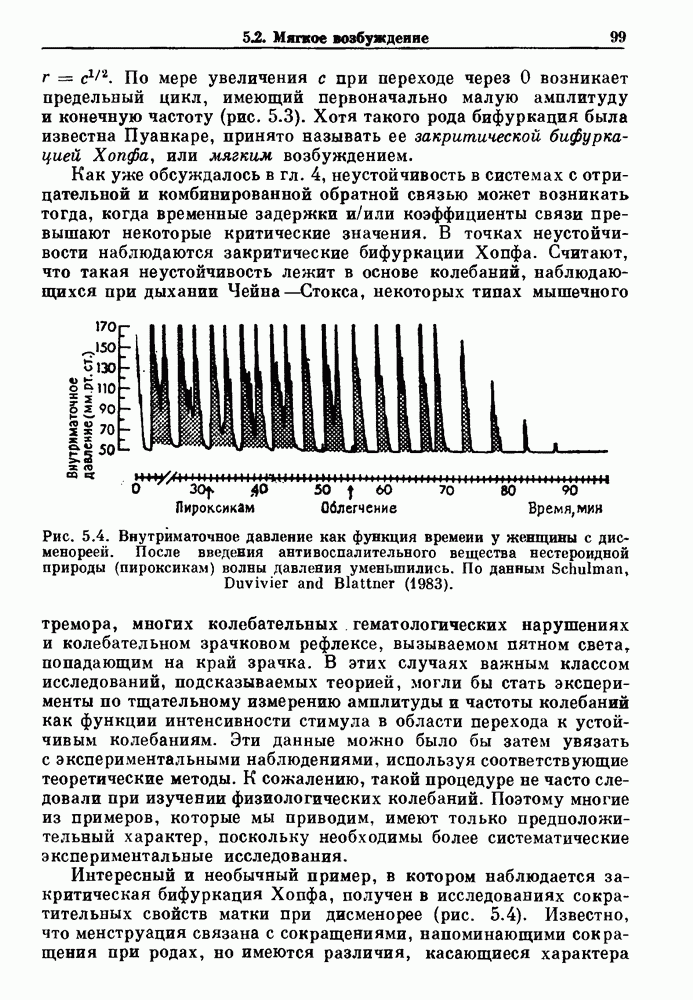
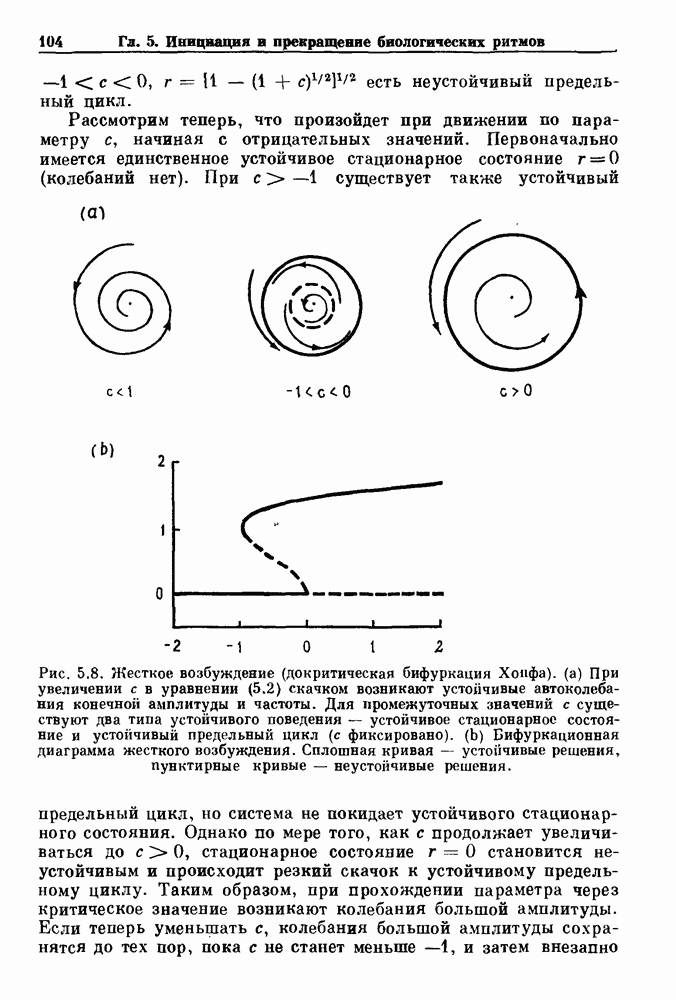
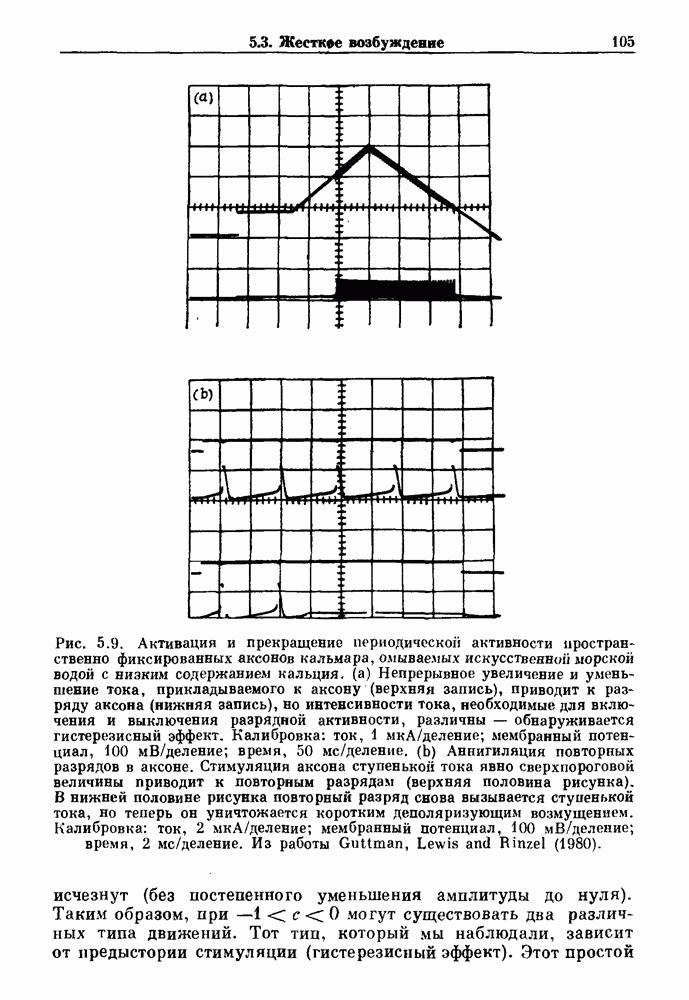
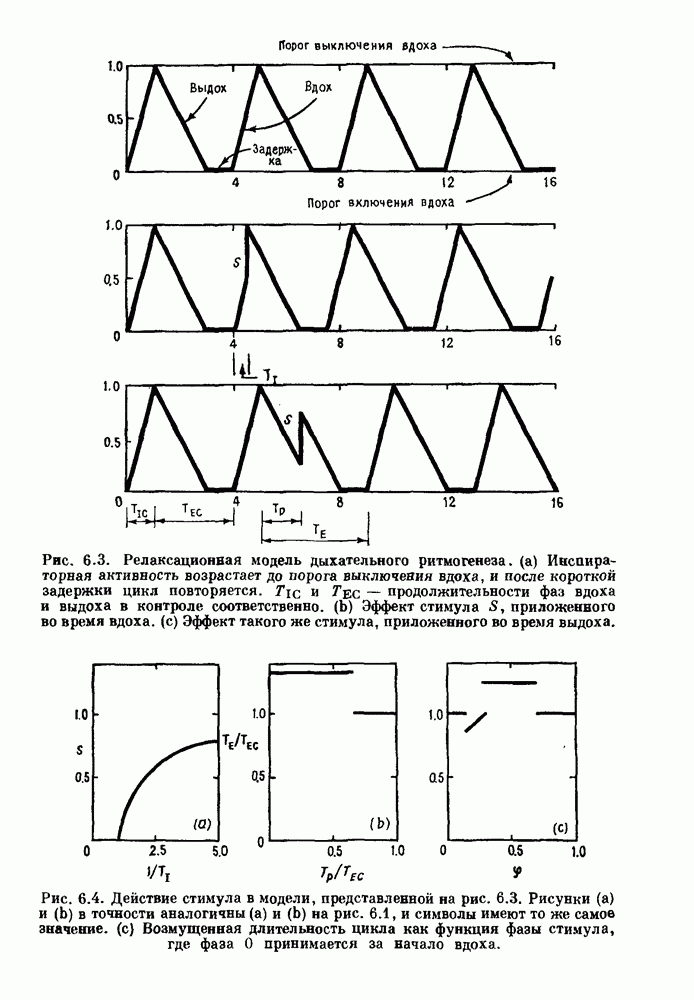
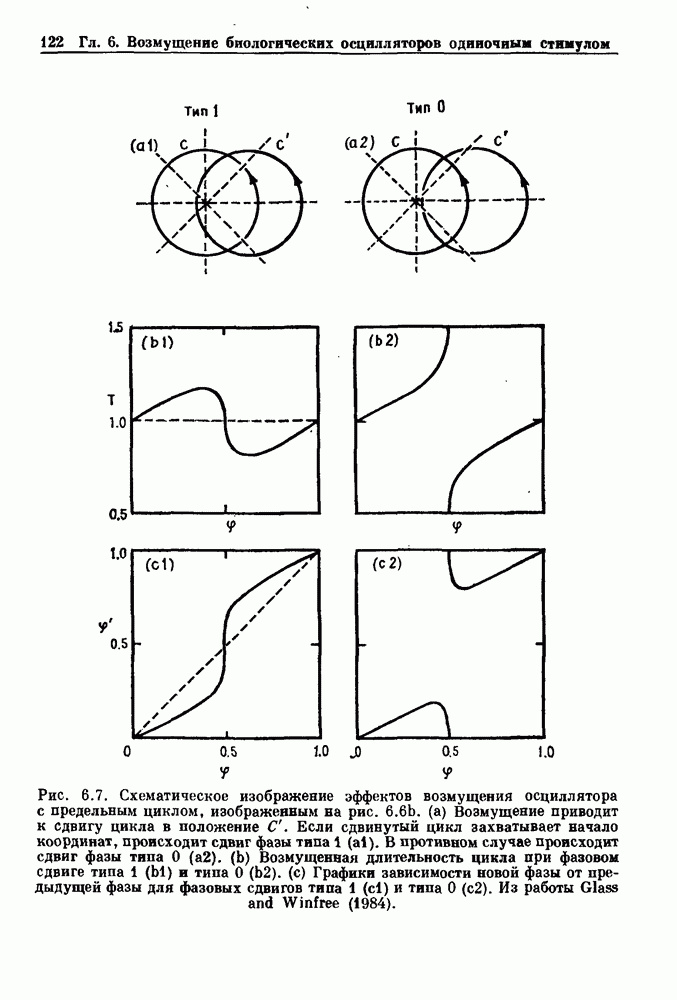
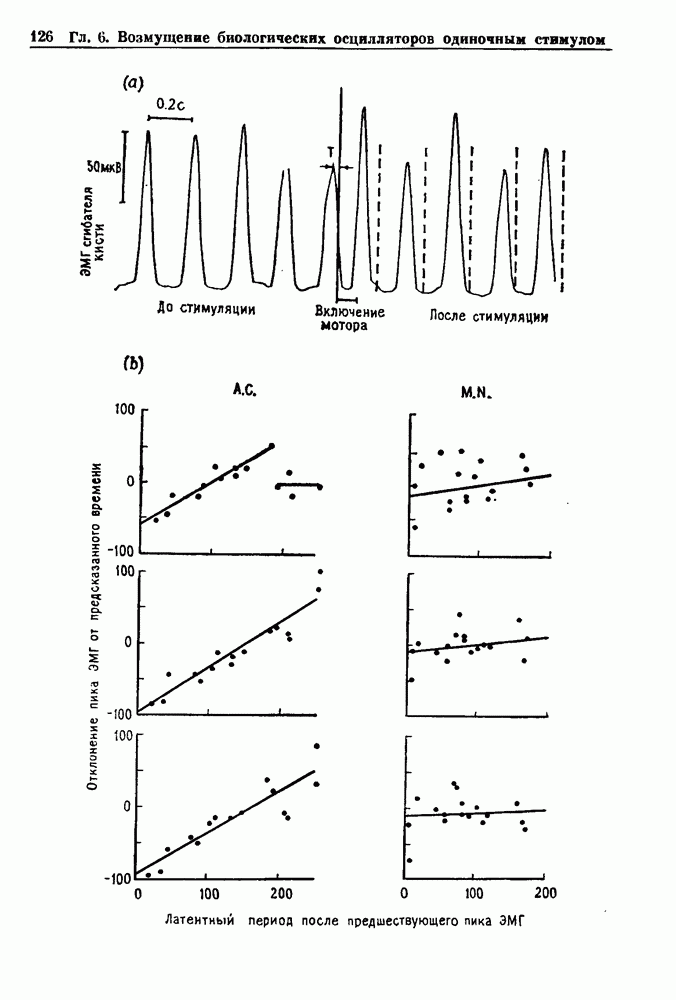
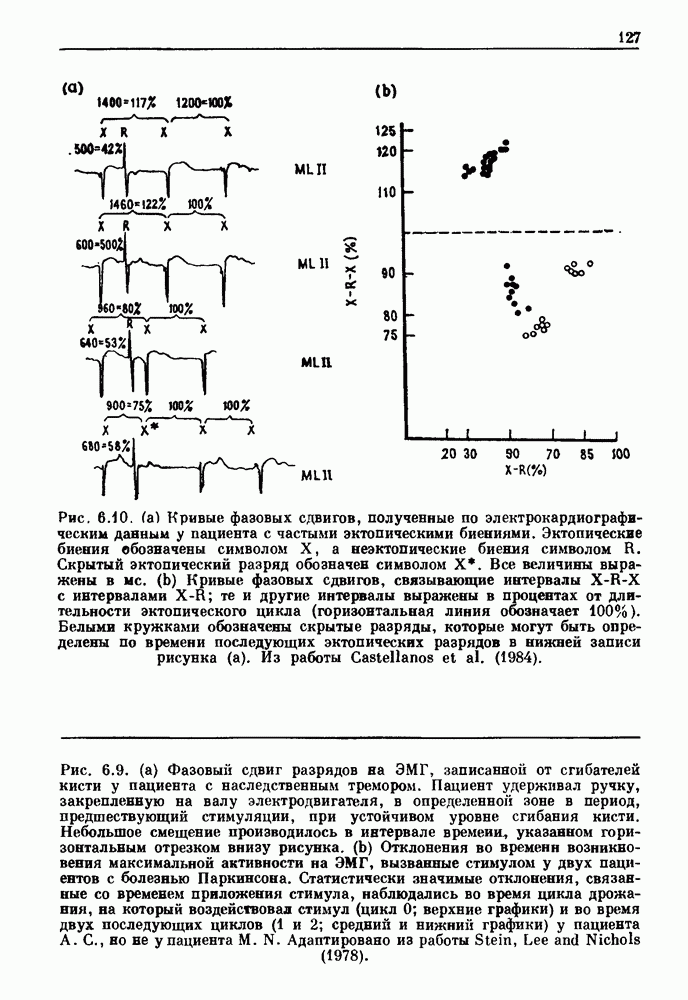
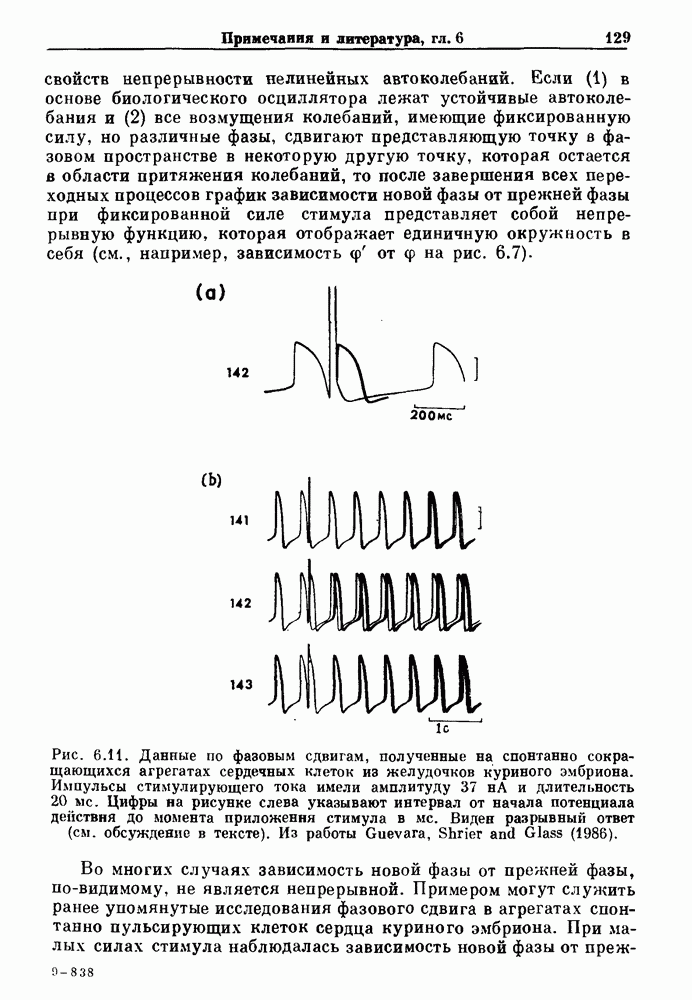
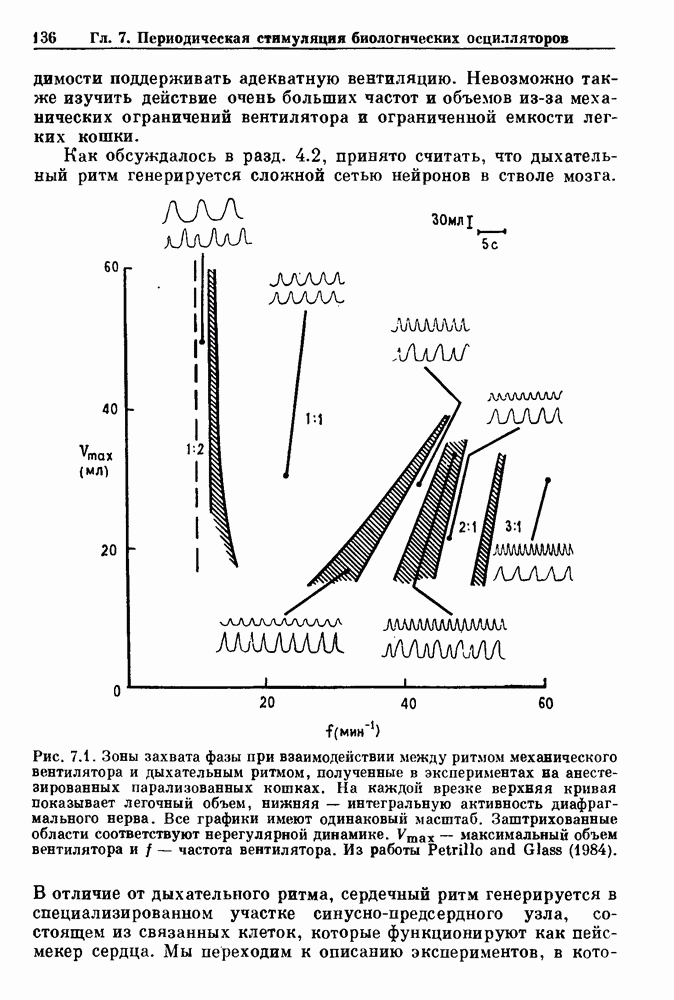
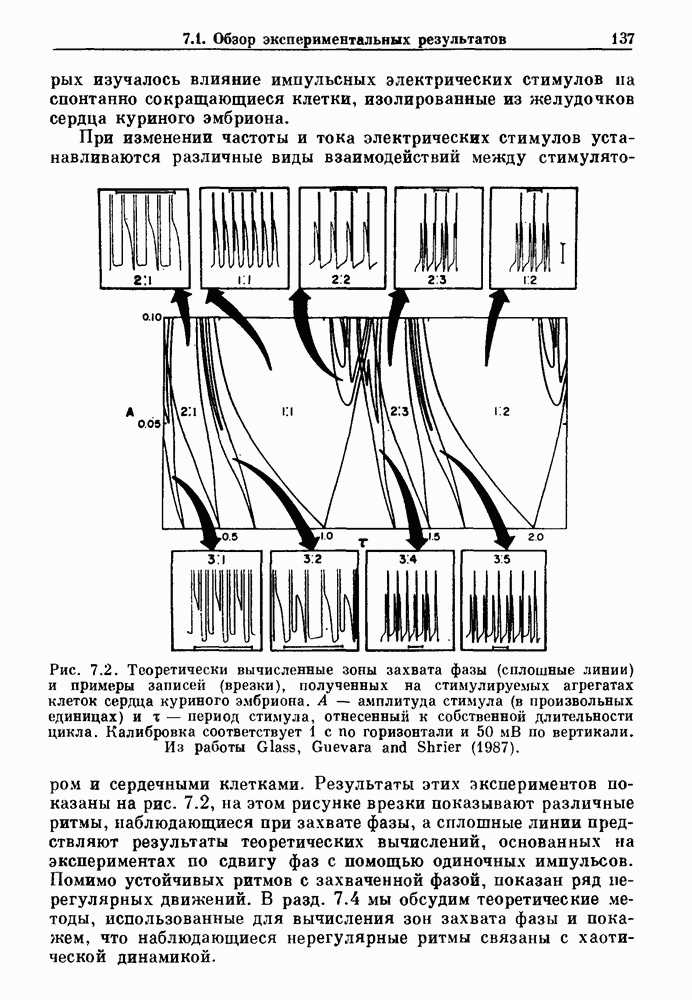
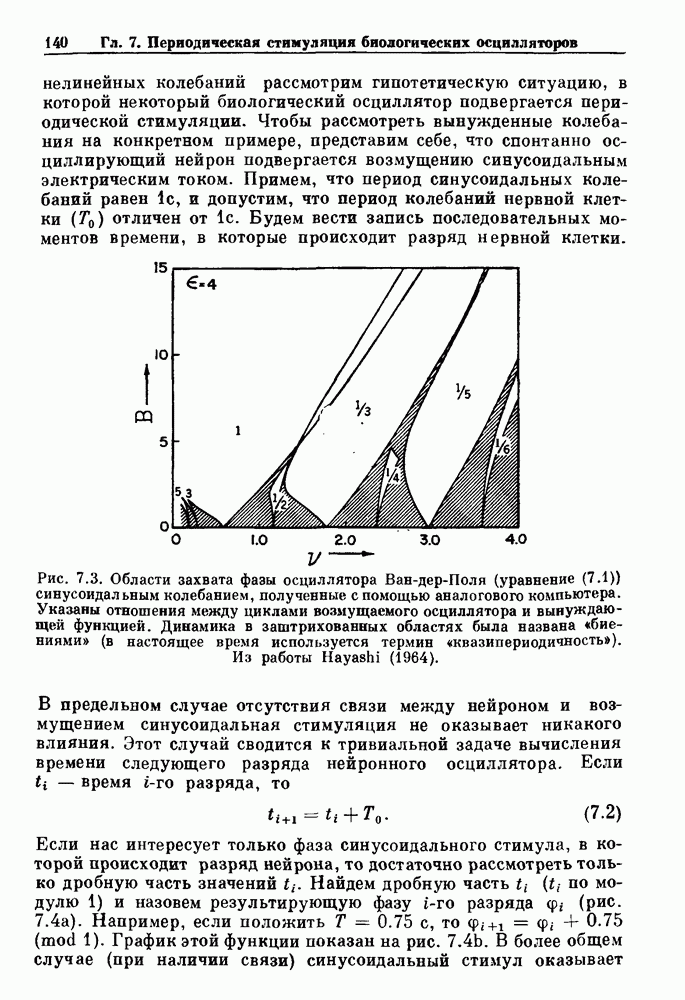
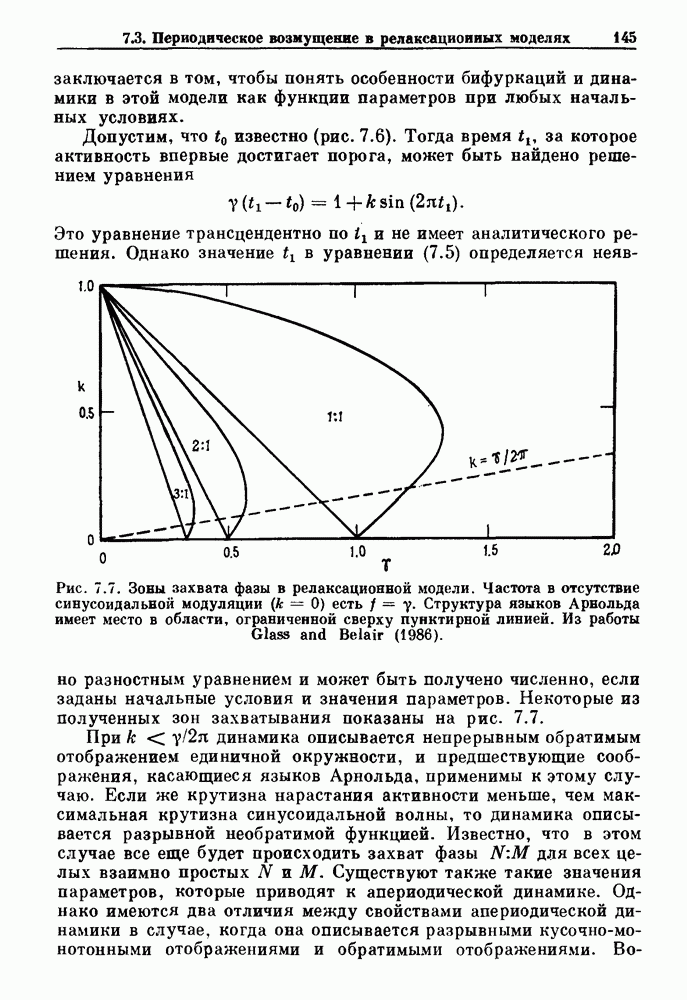
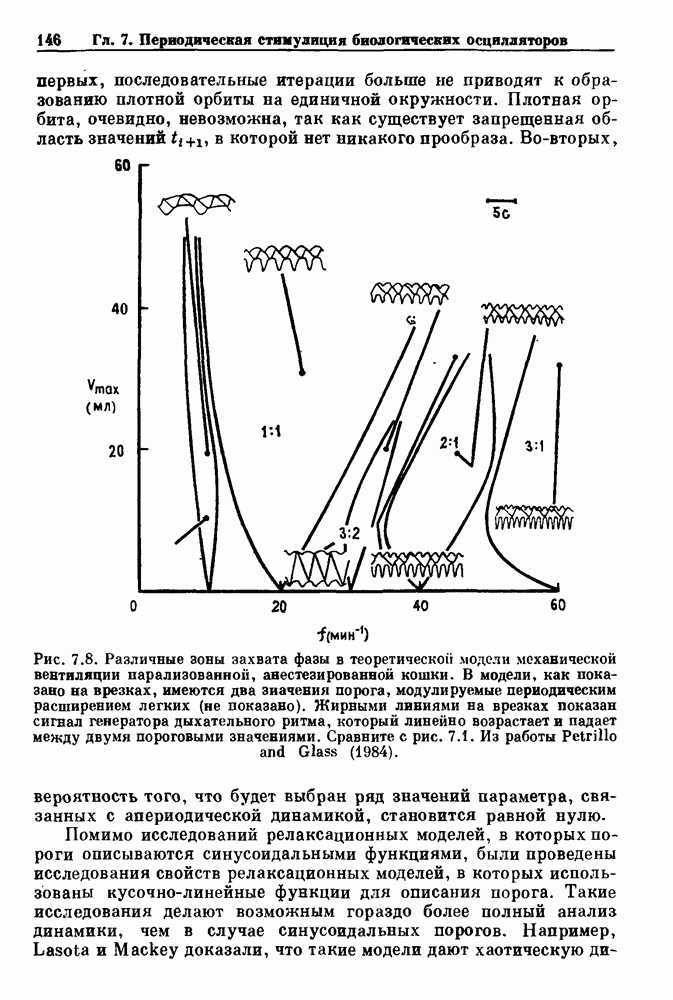
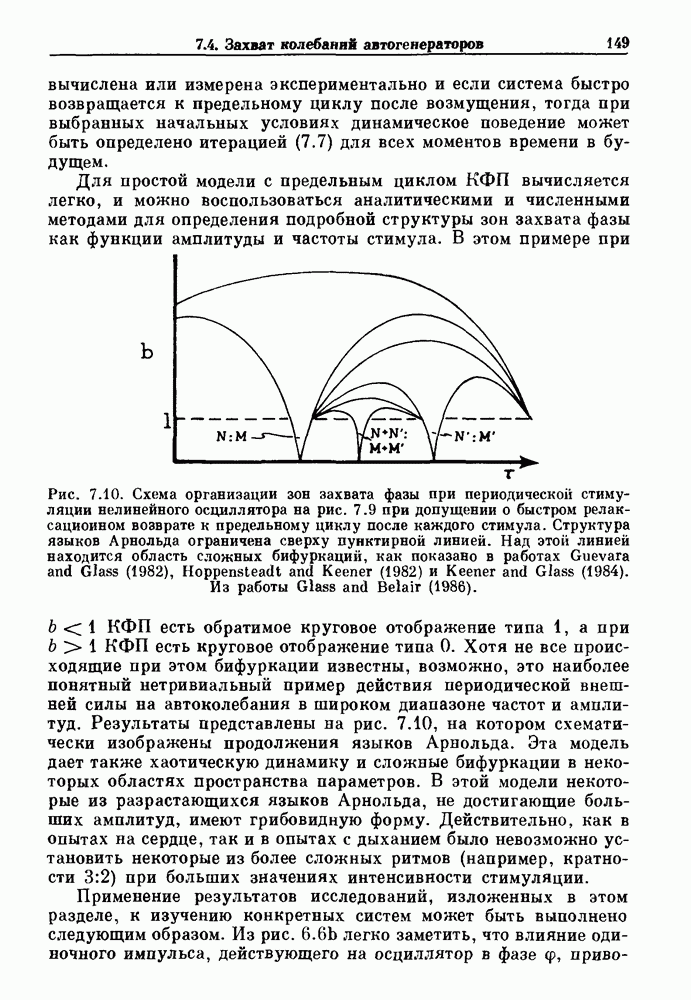
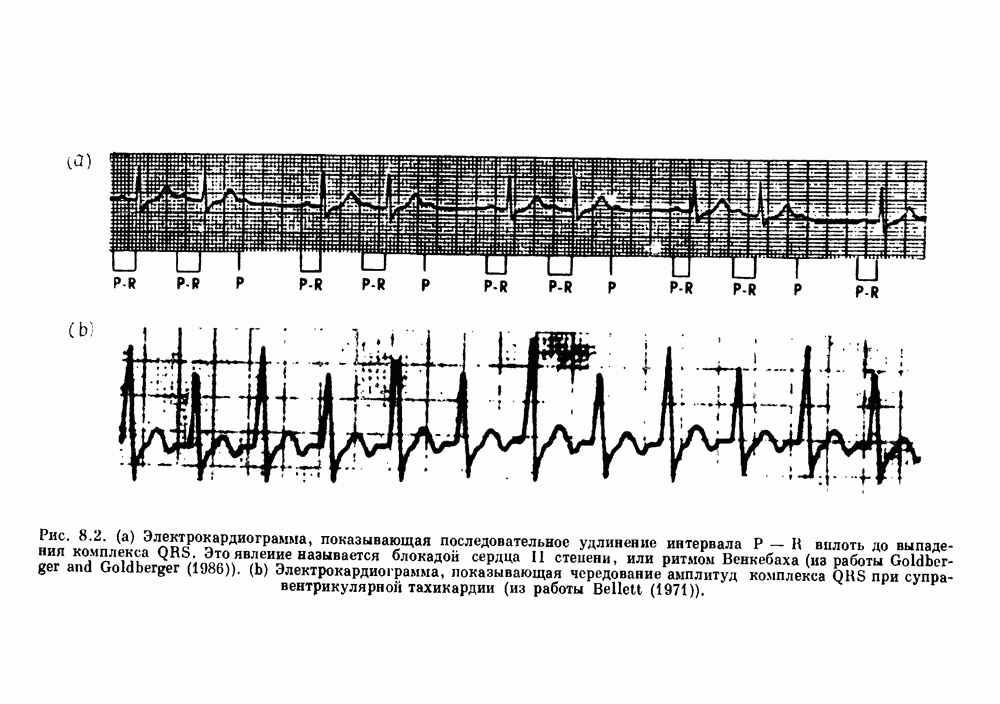
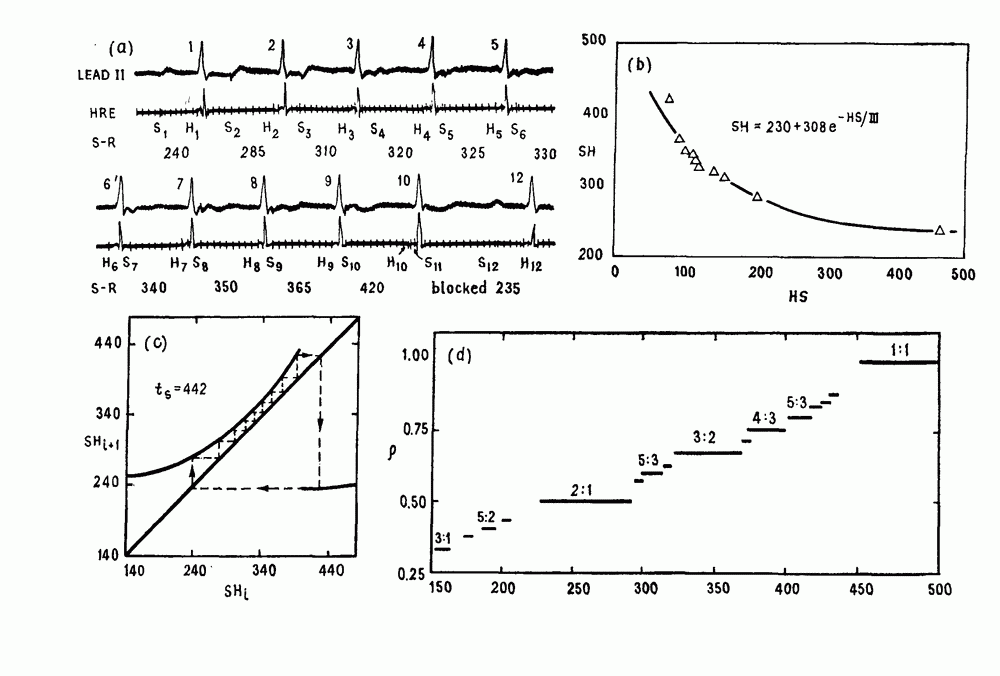
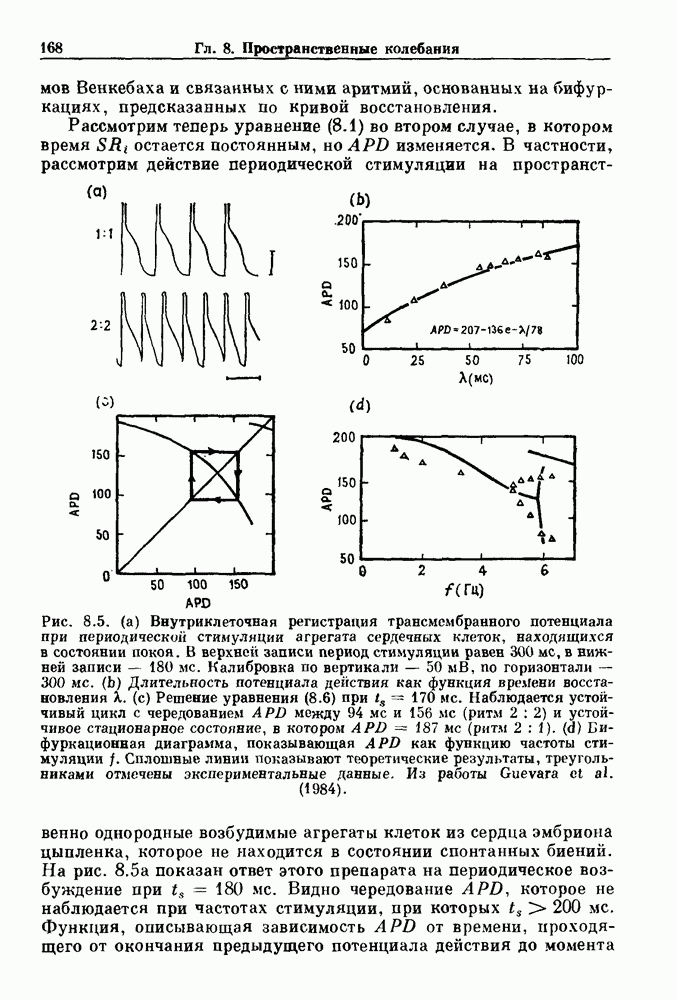
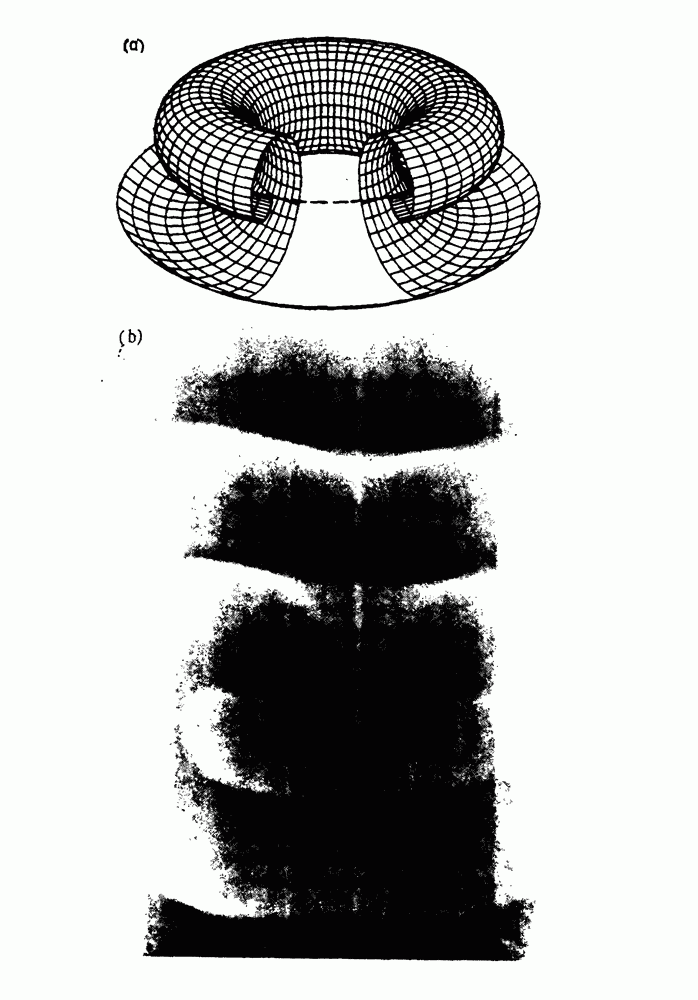
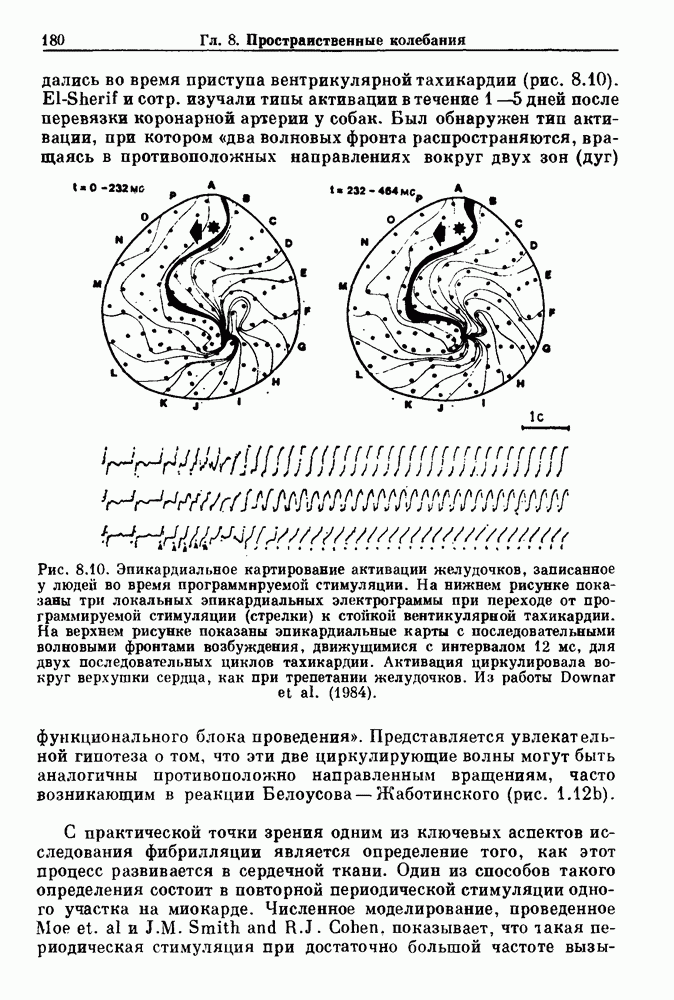
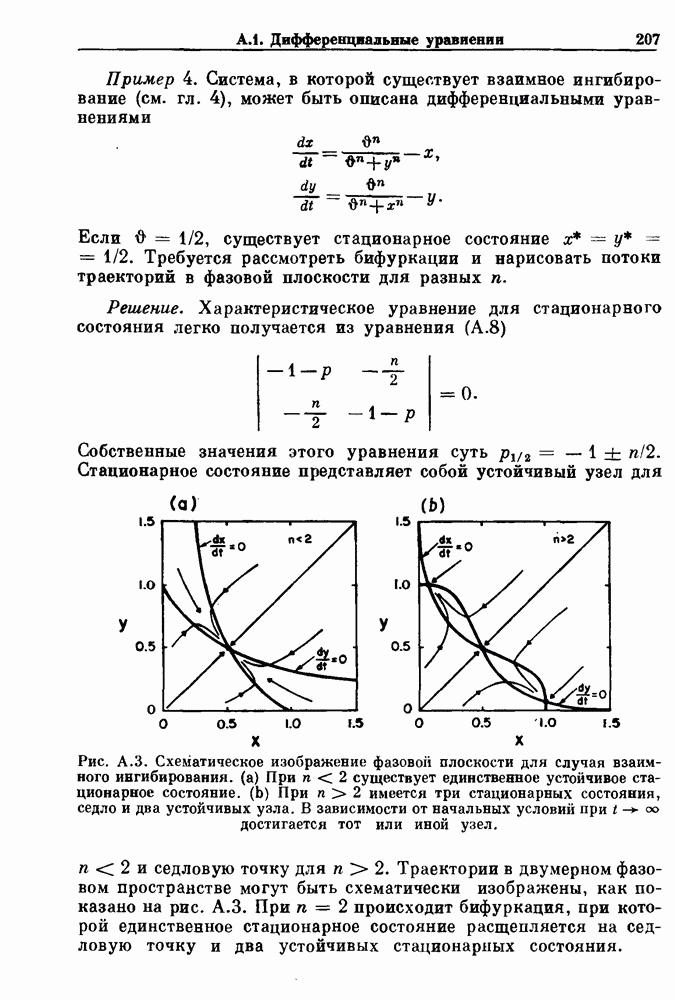
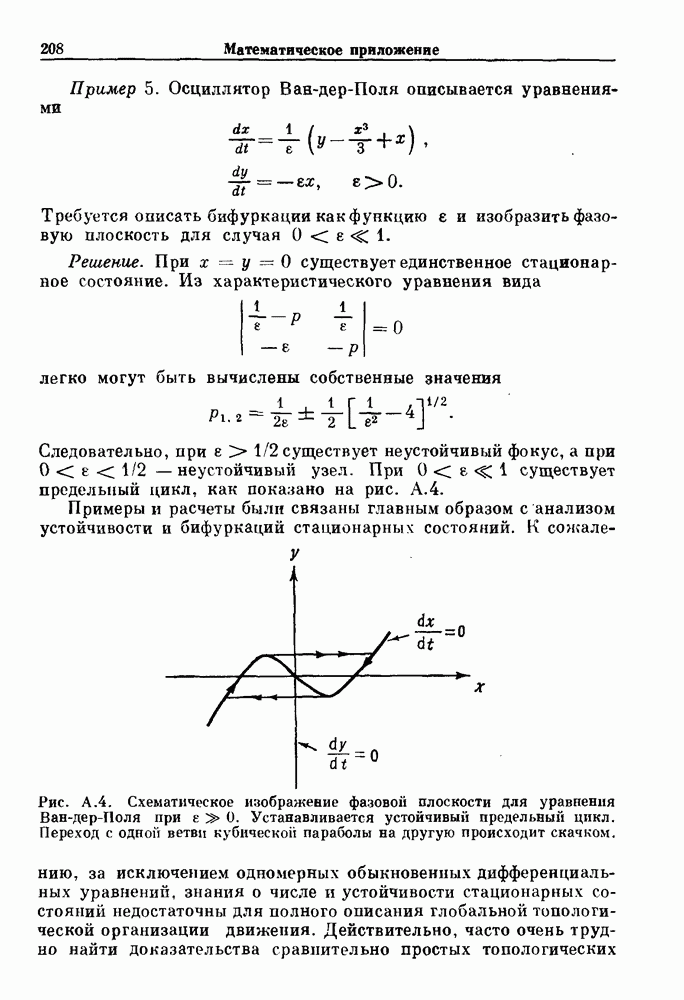
Глава 6. Возмущение биологических осцилляторов одиночным стимуломВ этой главе мы обсуждаем экспериментальные и теоретические результаты, касающиеся действия одиночных стимулов на биологические осцилляторы. В разд. 6.1 мы суммируем результаты таких исследований и, в частности, в качестве иллюстрации основных проблем приводим эксперименты по возмущению дыхательных и сердечных ритмов. В разд. 6.2 мы рассматриваем эксперименты по сдвигу фаз, используя релаксационные модели, а в разд. 6.3 описываем фазовые сдвиги, индуцируемые одиночным стимулом в моделях с предельным циклом. Некоторые возможные применения метода фазовых сдвигов в различных областях обсуждаются в разд. 6.4. Теория фазовых сдвигов в автоколебательных системах использует топологический подход. Практические проблемы, связанные с применением такого подхода, обсуждаются в разд. 6.5. 6.1. Обзор экспериментальных результатовВ этом разделе мы кратко обсуждаем сдвиги фаз дыхательных и сердечных ритмов и затем перечисляем основные свойства, обнаруженные в других системах. В качестве примера данных, получаемых в экспериментах с возмущением системы, рассмотрим возмущение дыхательного цикла принудительным расширением легких. Дыхательный цикл подразделяется на фазу вдоха, когда активность диафрагмального нерва велика и легкие расширяются, и фазу выдоха, когда активность мала или совсем отсутствует, а объем легких уменьшается. Длительность фазы вдоха ритма часто прослеживается путем регистрации усиливающей вдох активности диафрагмального нерва, который иннервирует диафрагму. Систематические исследования рефлексов Геринга—Брейера проводились при изучении действия расширения легких в различных фазах цикла у анестезированных, спонтанно дышащих кошек. При этом производилась запись активности диафрагмального нерва. Если достаточно сильное расширение легкого вызывается во время вдоха, то вдох прерывается. Если построить график зависимости пороговой величины объема В качестве второго примера рассмотрим действие возмущения на изолированные волокна Пуркинье, являющиеся частью проводящей системы сердца, у собак. После выделения из сердца волокно Пуркинье протягивалось через три камеры. В камере 1 вызывалась спонтанная колебательная активность волокна путем перфузии раствора Тироде с низким содержанием калия, в который добавлялся эпинефрин. Камера 2 содержала раствор декстрозы и хлорида кальция, который блокировал активное распространение волны, но не влиял на распространение электротонической активности. Камера 3 содержала раствор Тироде с высокой концентрацией калия, который увеличивал период спонтанных колебаний, но все же допускал распространение волны. Этот препарат с «сахарозным мостиком» рассматривался в качестве модели in vitro эктопического, т. е. аномального пейсмекерного участка (камера 1), способного отвечать на электрические стимулы (камера 3) через электротонические взаимодействия, распространяющиеся через область с пониженной проводимостью (камера 2). Электрические стимулы подавались в камеру 3 с различными интервалами на протяжении пейсмекерного цикла камеры 1. Если электрический стимул подавался от 0 до 230 мс после потенциала действия, длительность цикла изменялась мало. Если же стимул подавался спустя 800 мс после потенциала действия, длительность
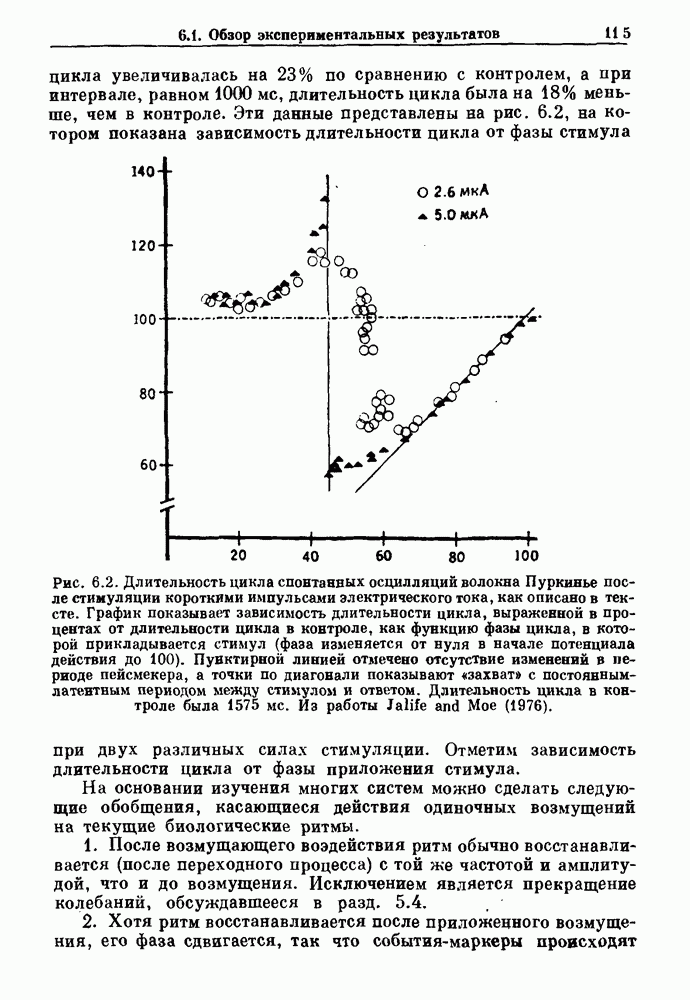
Рис. 6.1. Возмущение дыхания расширением легких у анестезированной, спонтанно дышащей кошки, (а) Короткие импульсы расширения, накладывающиеся на спонтанное дыхание во время вдоха. цикла увеличивалась на 23% по сравнению с контролем, а при интервале, равном 1000 мс, длительность цикла была на 18% меньше, чем в контроле. Эти данные представлены на рис. 6.2, на котором показана зависимость длительности цикла от фазы стимула при двух различных силах стимуляции.
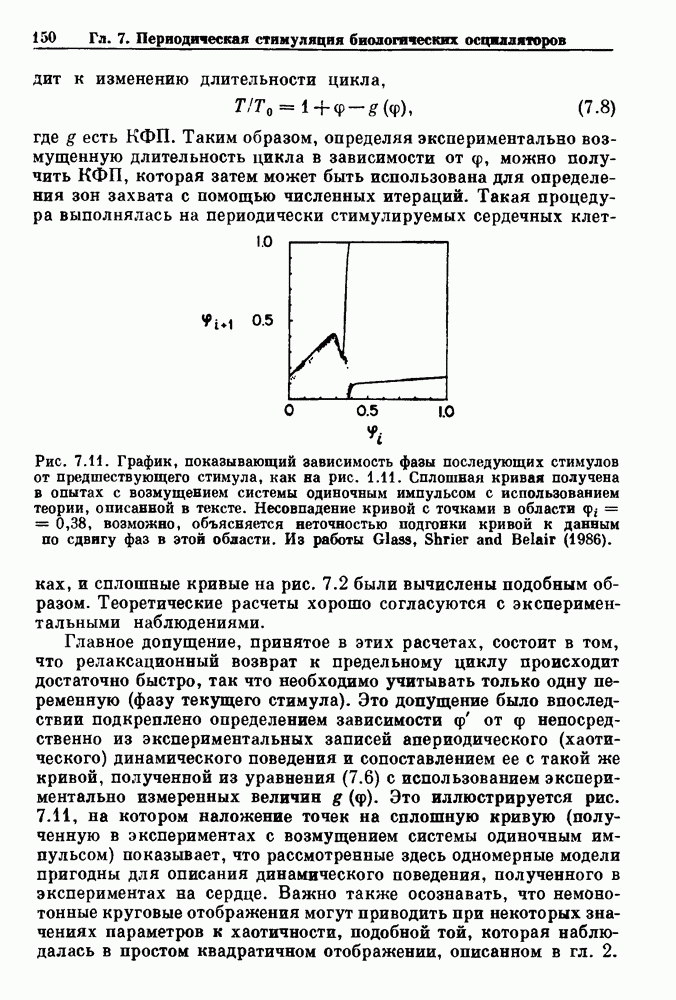
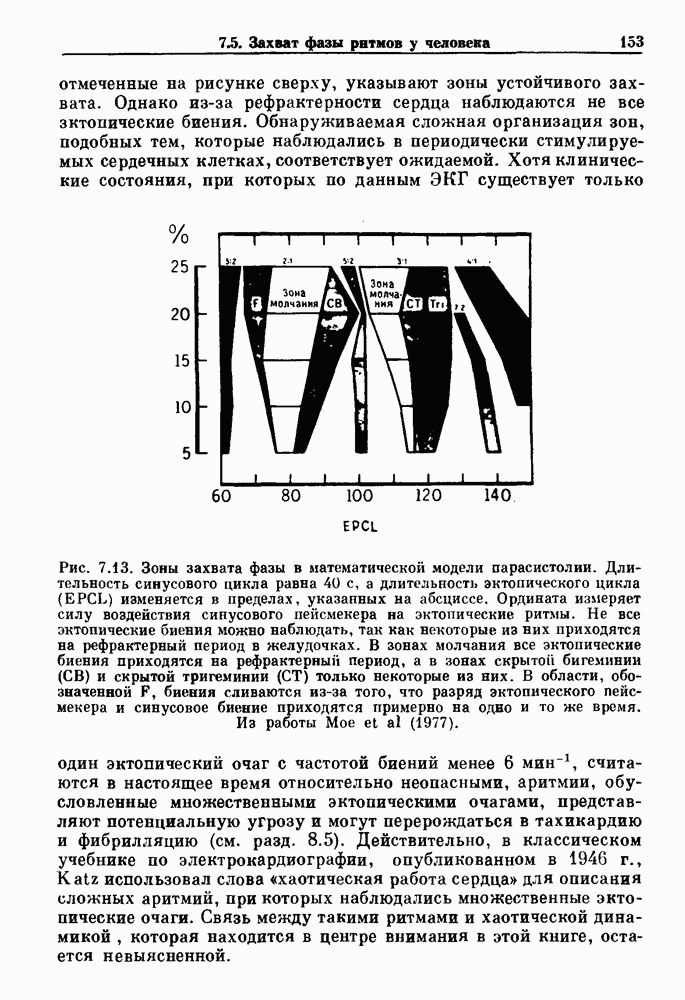
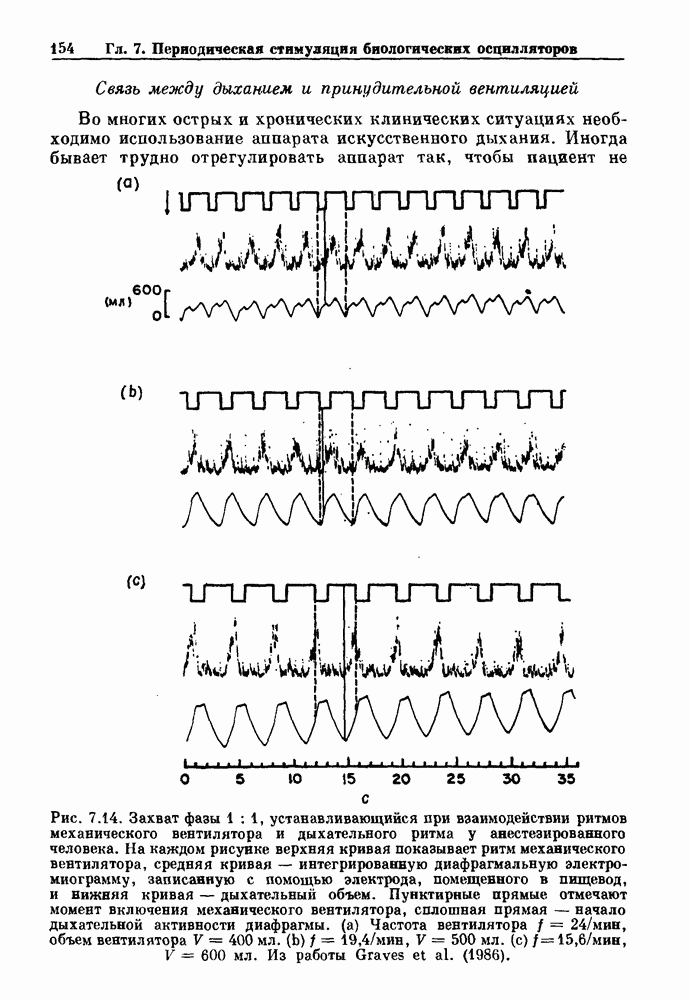
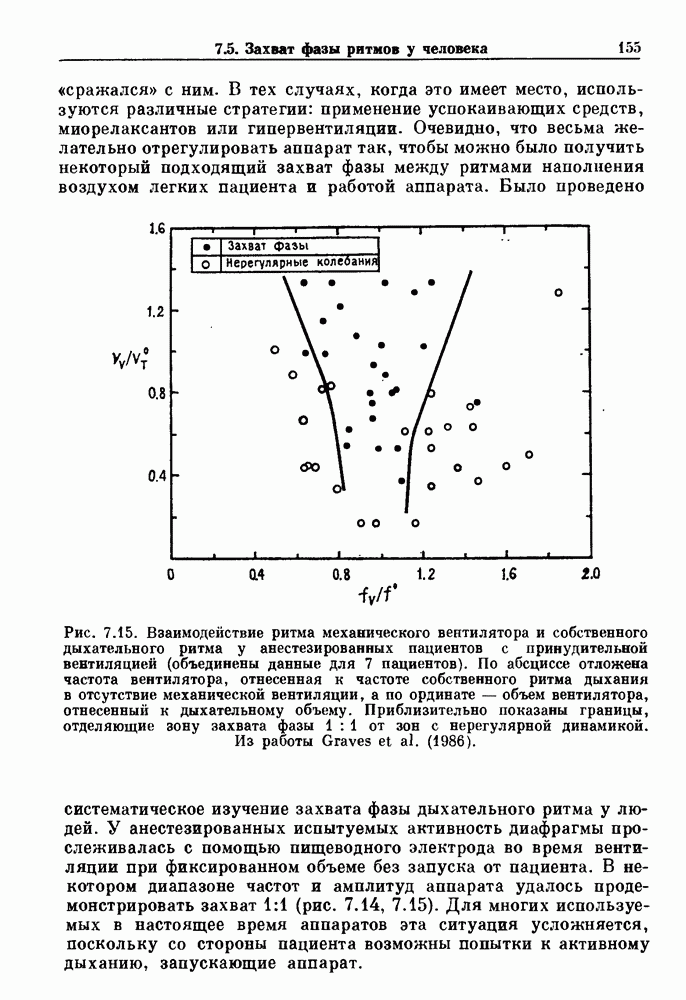
Рис. 6.2. Длительность цикла спонтанных осцилляций волокна Пуркинье после стимуляции короткими импульсами электрического тока, как описано в тексте. График показывает зависимость длительности цикла, выраженной в процентах от длительности цикла в контроле, как функцию фазы цикла, в которой прикладывается стимул (фаза изменяется от нуля в начале потенциала действия до 100). Пунктирной линией отмечено отсутствие изменений в периоде пейсмекера, а точки по диагонали показывают «захват» с постоянным-латентным периодом между стимулом и ответом. Длительность цикла в контроле была 1575 мс. Из работы Jalife and Мое (1976). Отметим зависимость длительности цикла от фазы приложения стимула. На основании изучения многих систем можно сделать следующие обобщения, касающиеся действия одиночных возмущений на текущие биологические ритмы. 1. После возмущающего воздействия ритм обычно восстанавливается (после переходного процесса) с той же частотой и амплитудой, что и до возмущения. Исключением является прекращение колебаний, обсуждавшееся в разд. 5.4. 2. Хотя ритм восстанавливается после приложенного возмущения, его фаза сдвигается, так что события-маркеры происходят в моменты времени, отличные от тех, в которые они наблюдались бы в отсутствие возмущения. 3. Одиночный стимул может приводить либо к удлинению, либо к укорочению продолжительности возмущенного цикла, в зависимости от фазы стимула. 4. На графике зависимости длительности цикла от. фазы стимула часто наблюдаются явные разрывы при некоторых амплитудах стимула, как показано на рис. 6.lb и 6.2. В следующих двух разделах мы обсудим эти четыре экспериментальных наблюдения в рамках модели релаксационного генератора и модели автогенератора и покажем, как применение этих двух классов моделей может быть разграничено в эксперименте.
|
 |
Оглавление
|























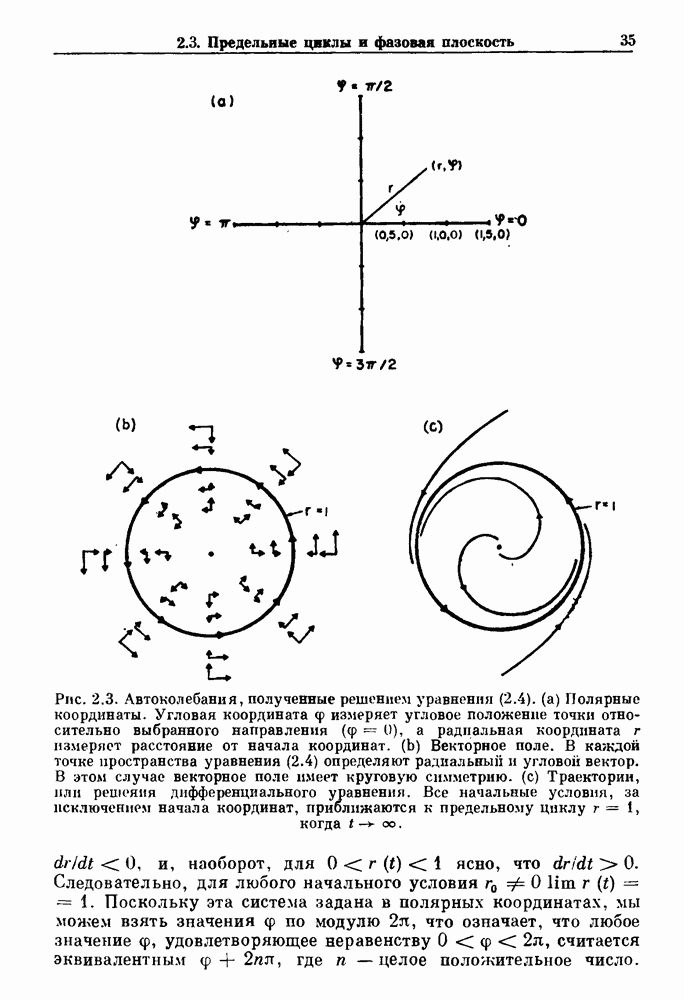


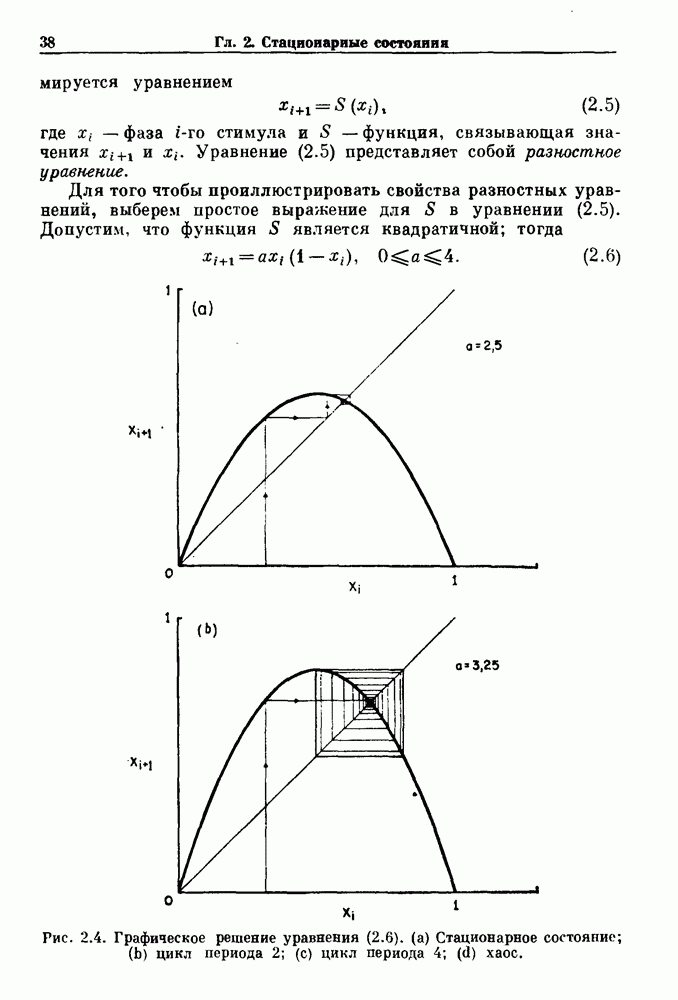
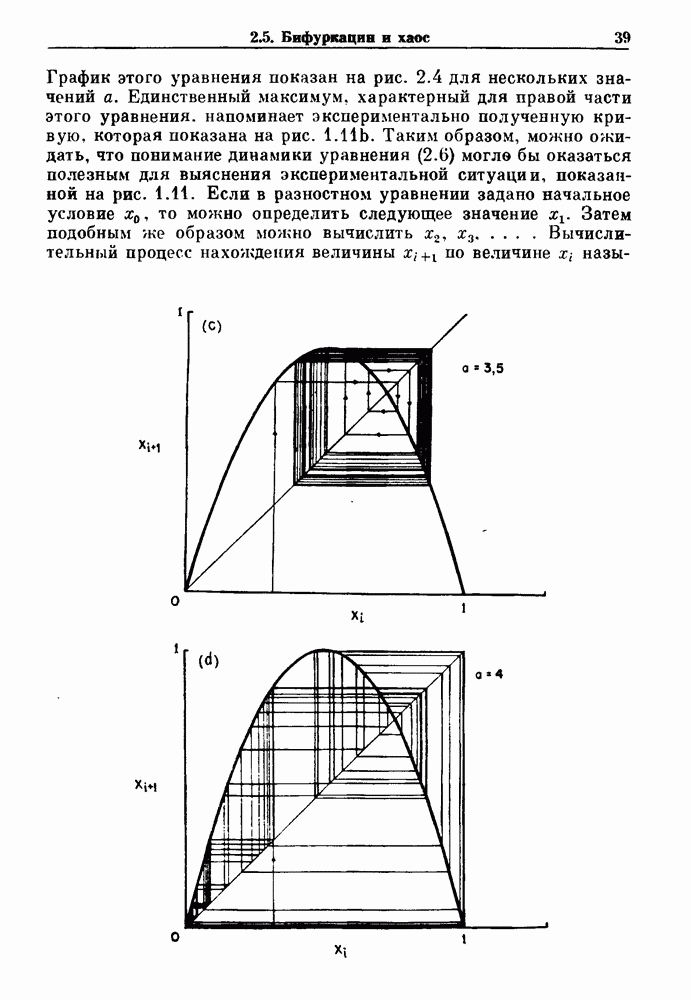
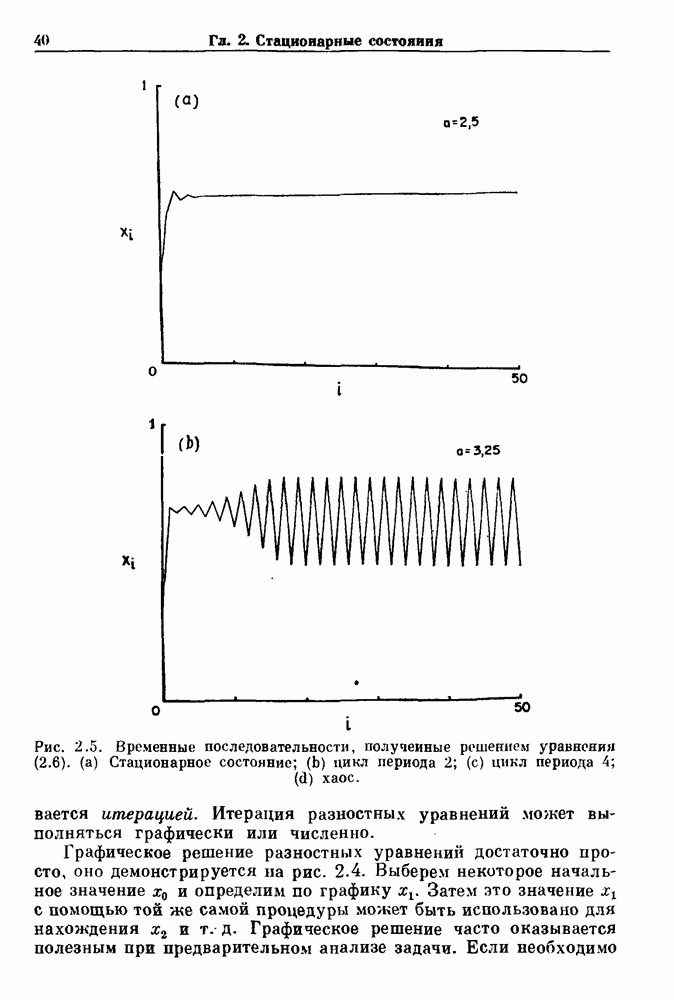











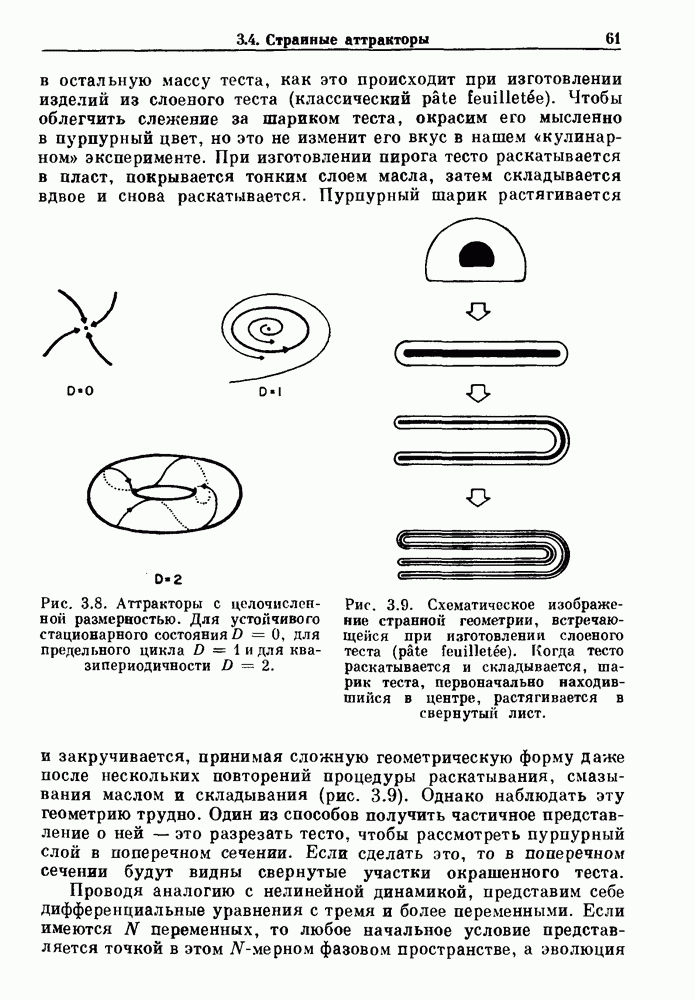































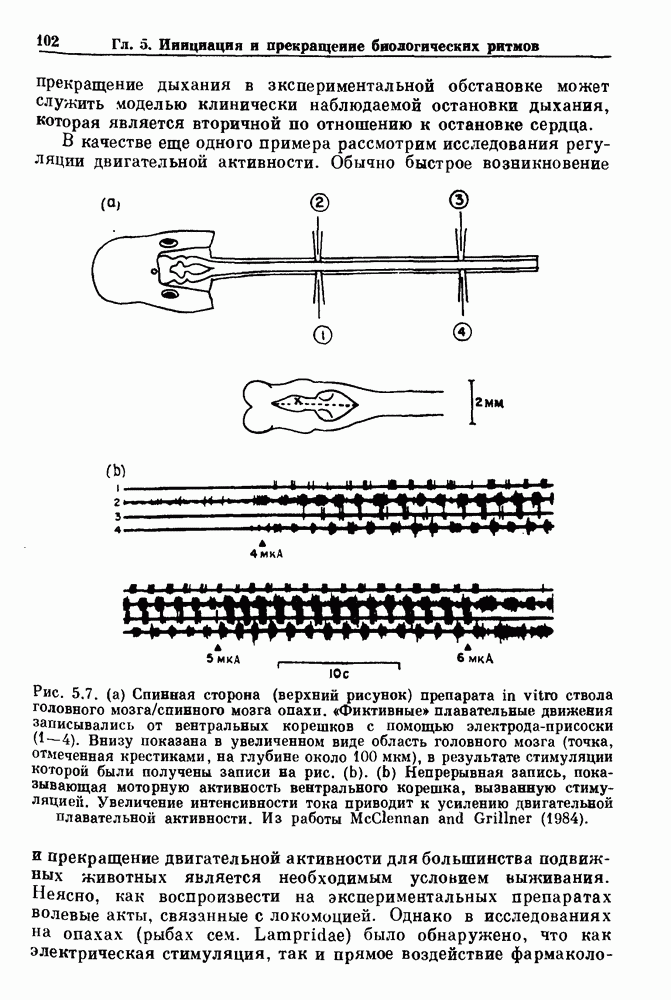









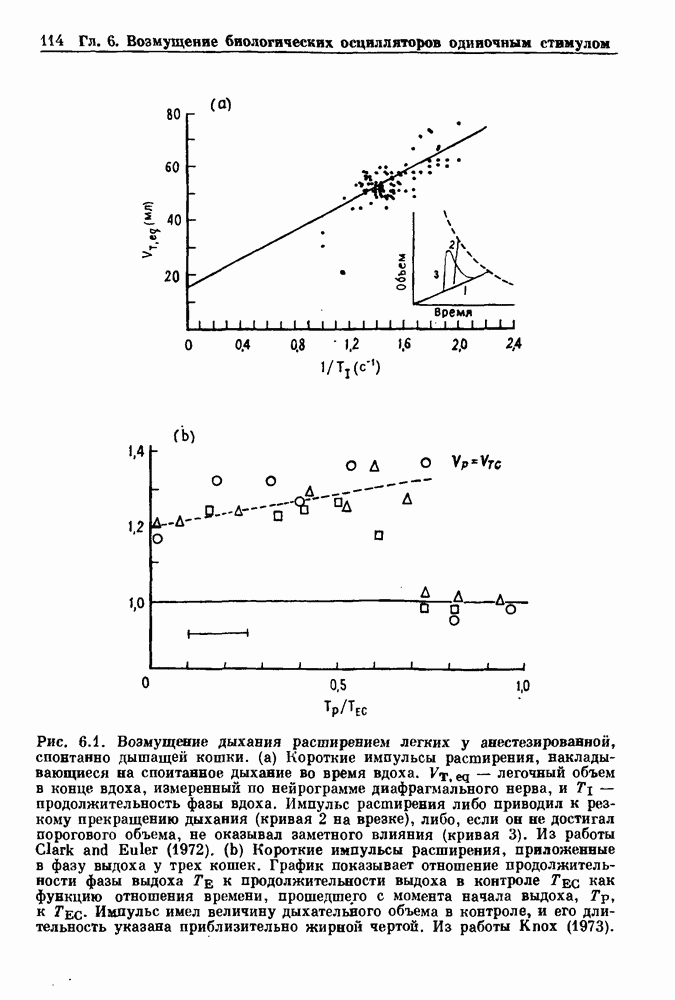

















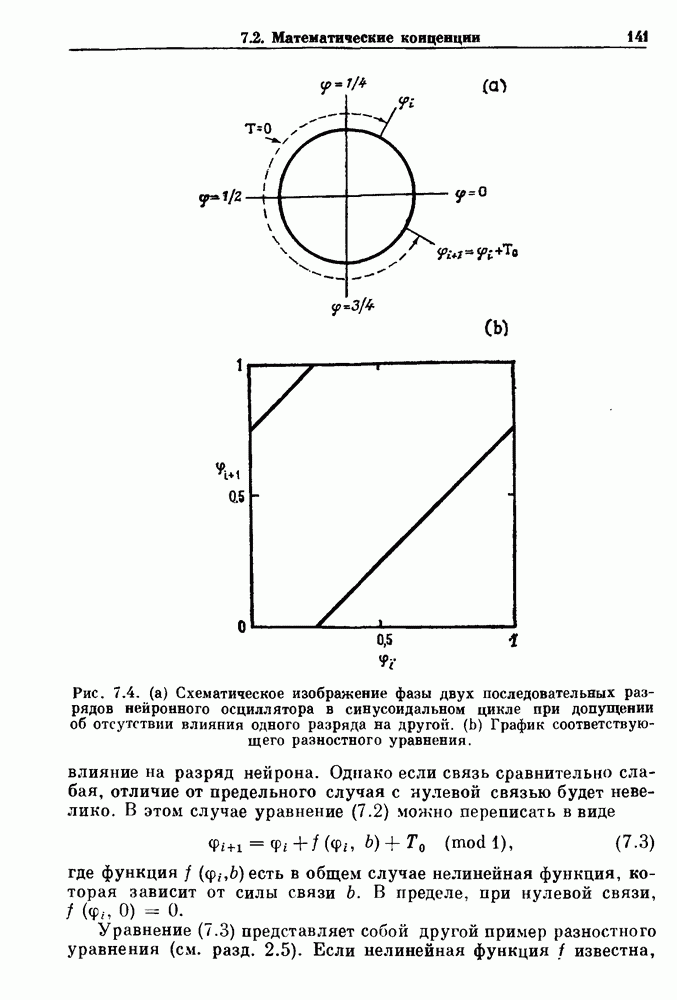























































































 и длительность фазы выдоха (ТЕ) изменяются при расширении легких. В прошлом веке Геринг и Брейер показали, что расширение легких, вызванное во время вдоха, приводит к укорочению фазы вдоха в респираторном цикле, в то время как расширение легких, вызванное во время выдоха, продлевает фазу выдоха. Рефлексы Геринга—Брейера существуют в норме и осуществляются с помощью афферентных волокон блуждающего нерва. Если перерезать блуждающий нерв, то частота дыхания уменьшится, а дыхательный объем увеличится. В современных исследованиях активность центрального генератора дыхательного
и длительность фазы выдоха (ТЕ) изменяются при расширении легких. В прошлом веке Геринг и Брейер показали, что расширение легких, вызванное во время вдоха, приводит к укорочению фазы вдоха в респираторном цикле, в то время как расширение легких, вызванное во время выдоха, продлевает фазу выдоха. Рефлексы Геринга—Брейера существуют в норме и осуществляются с помощью афферентных волокон блуждающего нерва. Если перерезать блуждающий нерв, то частота дыхания уменьшится, а дыхательный объем увеличится. В современных исследованиях активность центрального генератора дыхательного
 необходимой для прерывания вдоха, от
необходимой для прерывания вдоха, от  то он будет иметь вид, показанный на рис. 6.1а, и можно заметить, что подпороговые импульсы оказывают слабое влияние на продолжительность вдоха. Дополнительные эксперименты по измерению длительности возмущенной фазы выдоха по отношению к контролю в зависимости от момента времени, в который проводилась стимуляция (рис. 6.lb), показывает, что длительность выдоха и, следовательно, длительность возмущенного цикла увеличиваются при расширении легких, вызванном во время выдоха. Хотя исследования, иллюстрируемые рис. 6.lb, не имеют прямого отношения к длительным фазовым сдвигам, вызываемым возмущением, известно, что при стимуляции эфферентных волокон легких фаза сдвигается. Это означает, что после нескольких циклов характер разрядов диафрагмального нерва будет не таким, как в отсутствие стимуляции.
то он будет иметь вид, показанный на рис. 6.1а, и можно заметить, что подпороговые импульсы оказывают слабое влияние на продолжительность вдоха. Дополнительные эксперименты по измерению длительности возмущенной фазы выдоха по отношению к контролю в зависимости от момента времени, в который проводилась стимуляция (рис. 6.lb), показывает, что длительность выдоха и, следовательно, длительность возмущенного цикла увеличиваются при расширении легких, вызванном во время выдоха. Хотя исследования, иллюстрируемые рис. 6.lb, не имеют прямого отношения к длительным фазовым сдвигам, вызываемым возмущением, известно, что при стимуляции эфферентных волокон легких фаза сдвигается. Это означает, что после нескольких циклов характер разрядов диафрагмального нерва будет не таким, как в отсутствие стимуляции.

 легочный объем в конце вдоха, измеренный по нейрограмме диафрагмального нерва, к
легочный объем в конце вдоха, измеренный по нейрограмме диафрагмального нерва, к  продолжительность фазы вдоха. Импульс расширения либо приводил к резкому прекращению дыхания (кривая 2 на врезке), либо, если он не достигал порогового объема, не оказывал заметного влияния (кривая 3). Из работы Clark and Euler (1972). (b) Короткие импульсы расширения, приложенные в фазу выдоха у трех кошек. График показывает отношение продолжительности фазы выдоха Те к продолжительности выдоха в контроле
продолжительность фазы вдоха. Импульс расширения либо приводил к резкому прекращению дыхания (кривая 2 на врезке), либо, если он не достигал порогового объема, не оказывал заметного влияния (кривая 3). Из работы Clark and Euler (1972). (b) Короткие импульсы расширения, приложенные в фазу выдоха у трех кошек. График показывает отношение продолжительности фазы выдоха Те к продолжительности выдоха в контроле  как функцию отношения времени, прошедшего с момента начала выдоха,
как функцию отношения времени, прошедшего с момента начала выдоха,  к
к  Импульс имел величину дыхательного объема в контроле, и его длительность указана приблизительно жирной чертой. Из работы Knox (1973).
Импульс имел величину дыхательного объема в контроле, и его длительность указана приблизительно жирной чертой. Из работы Knox (1973).
