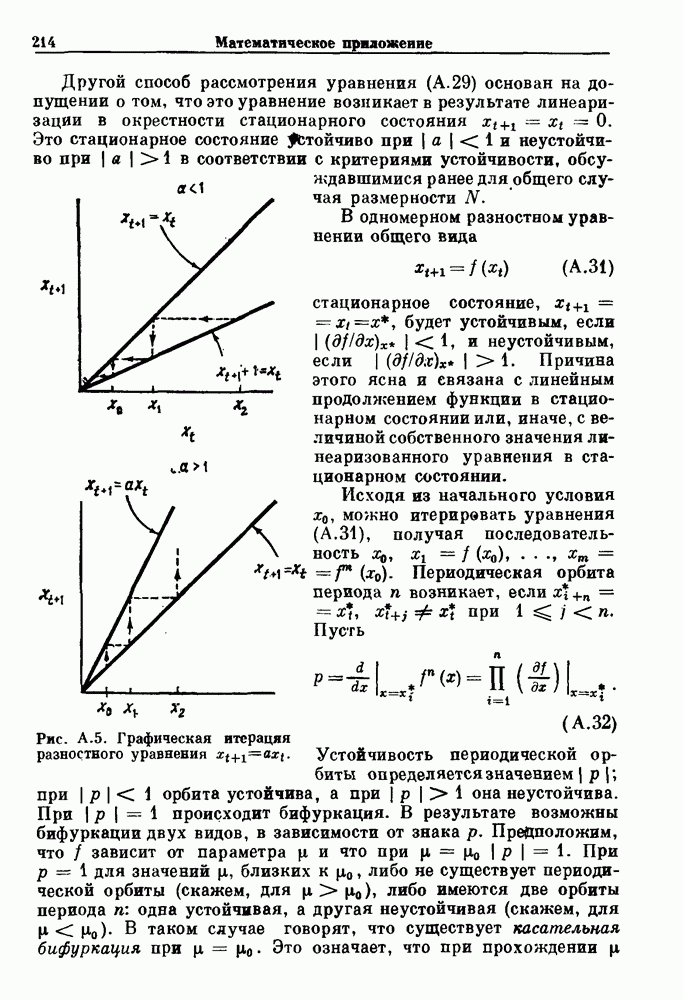
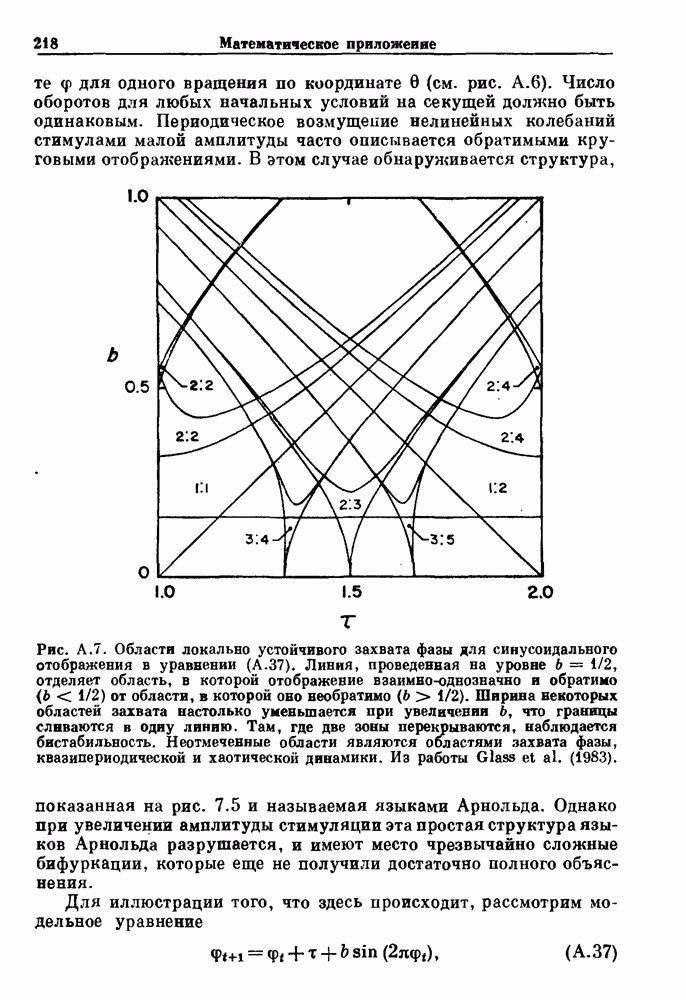
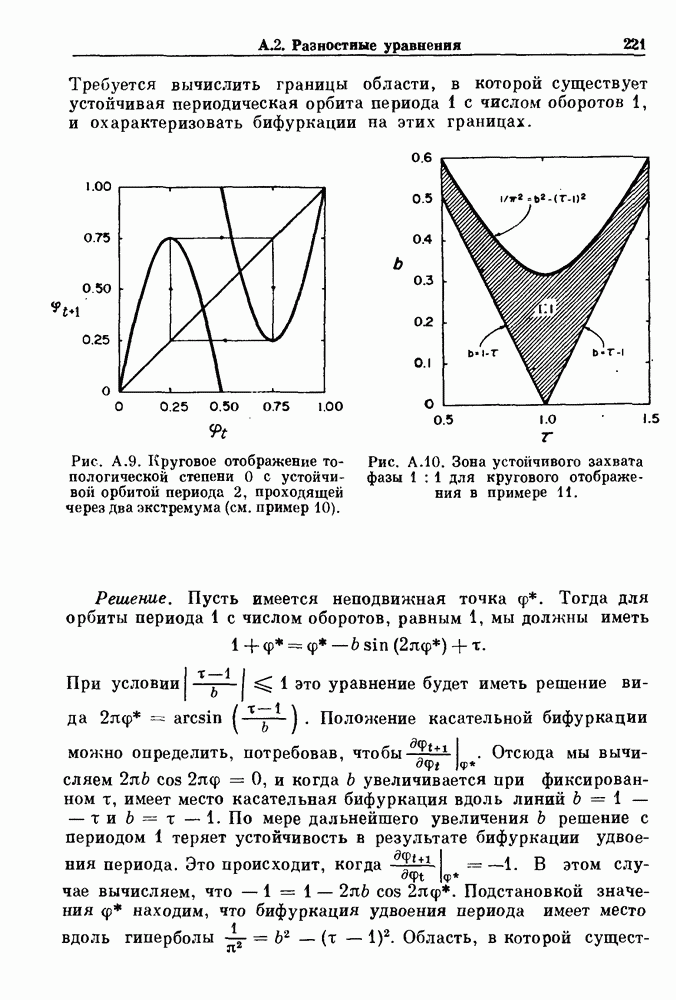
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
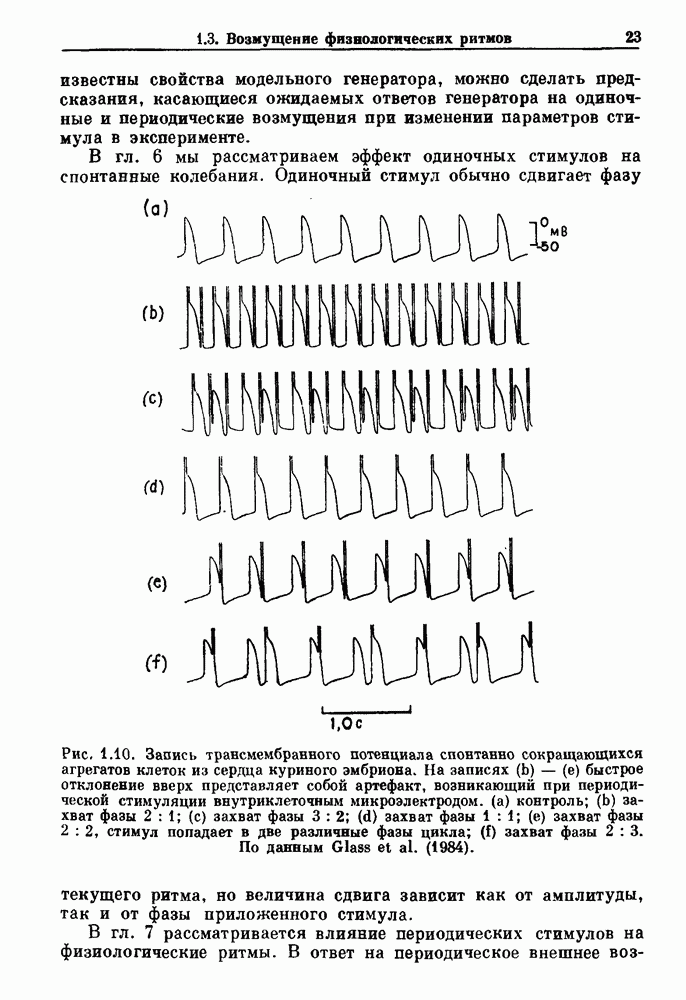
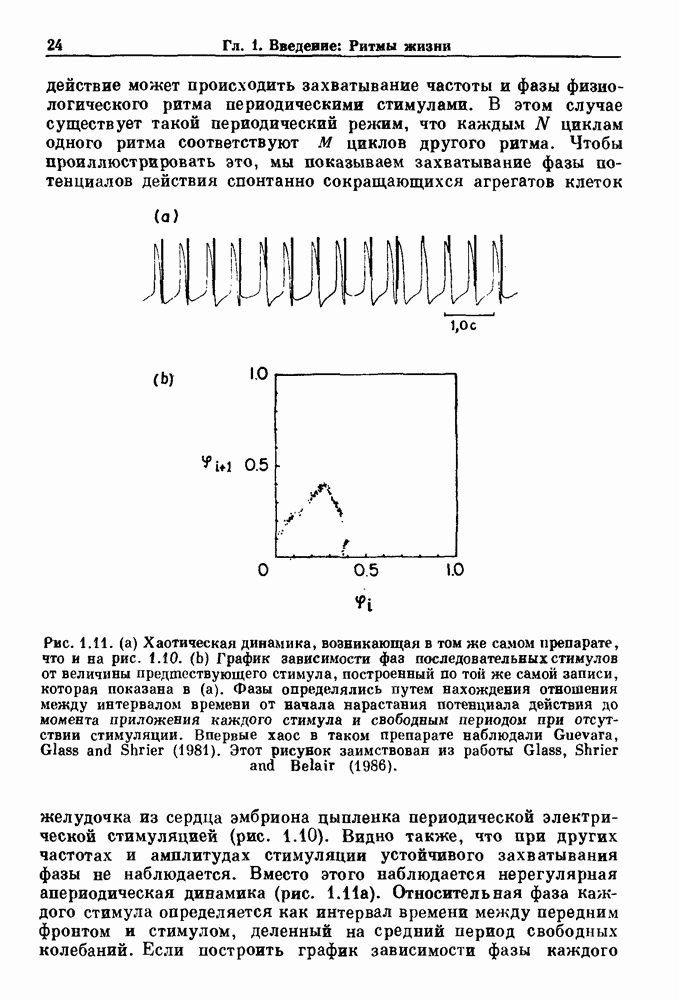
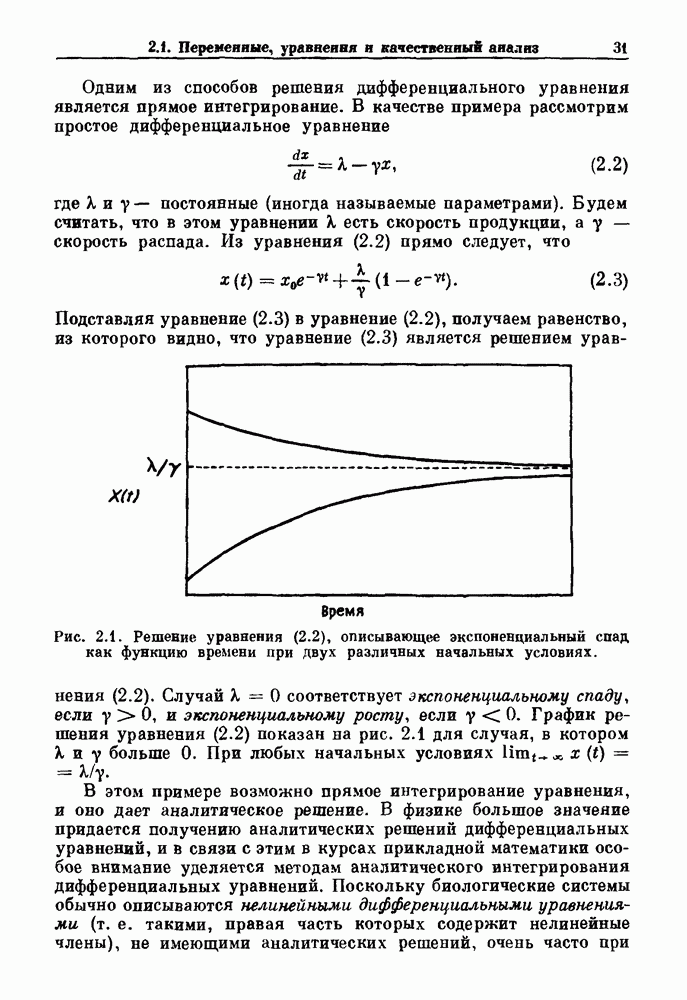
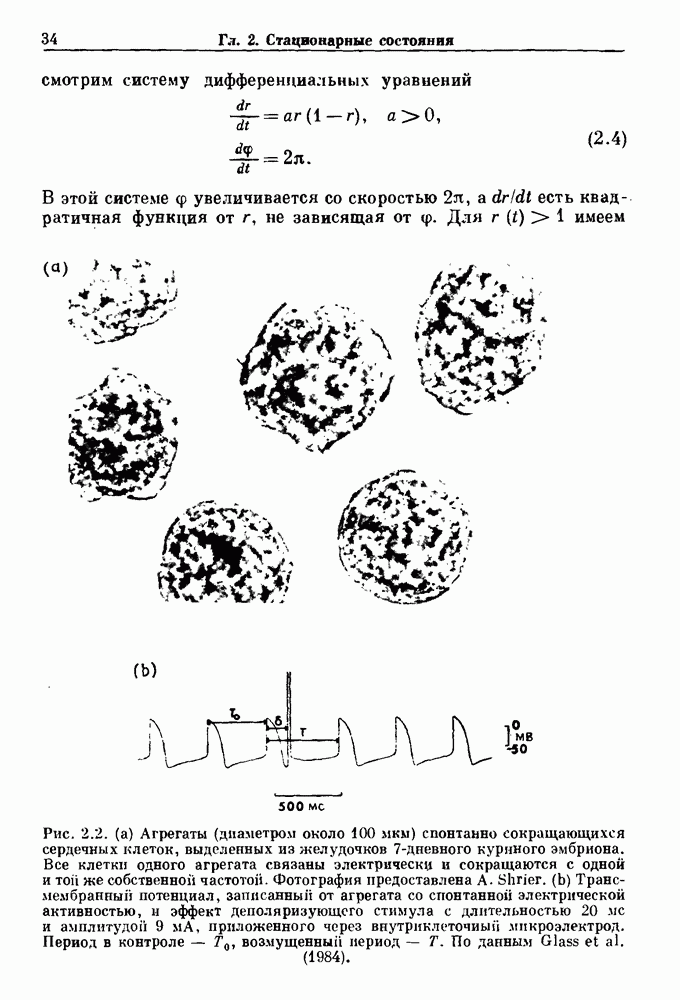
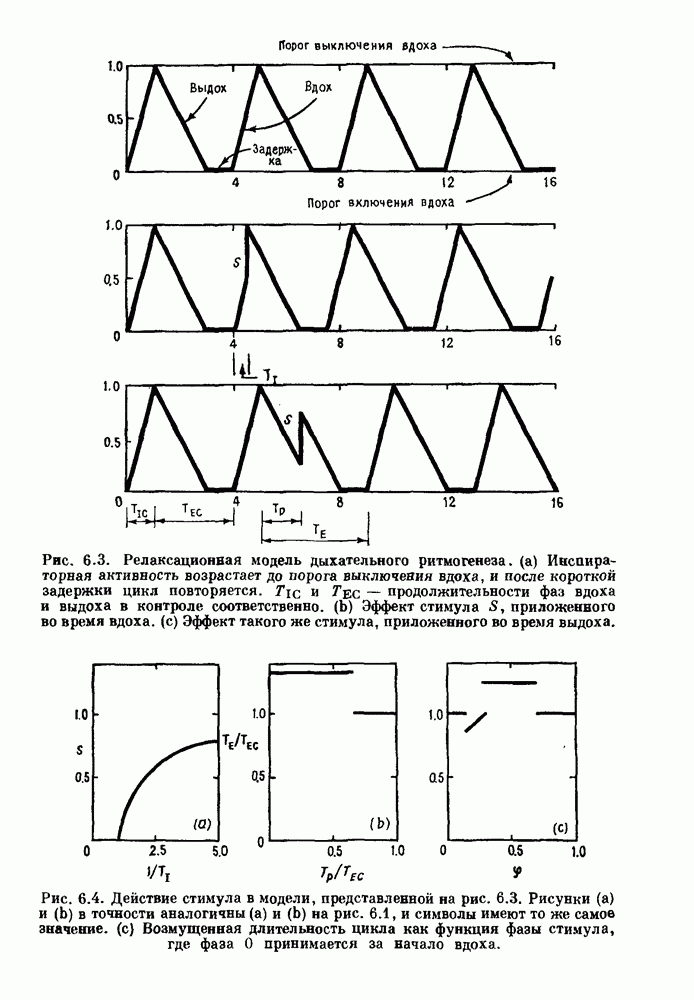
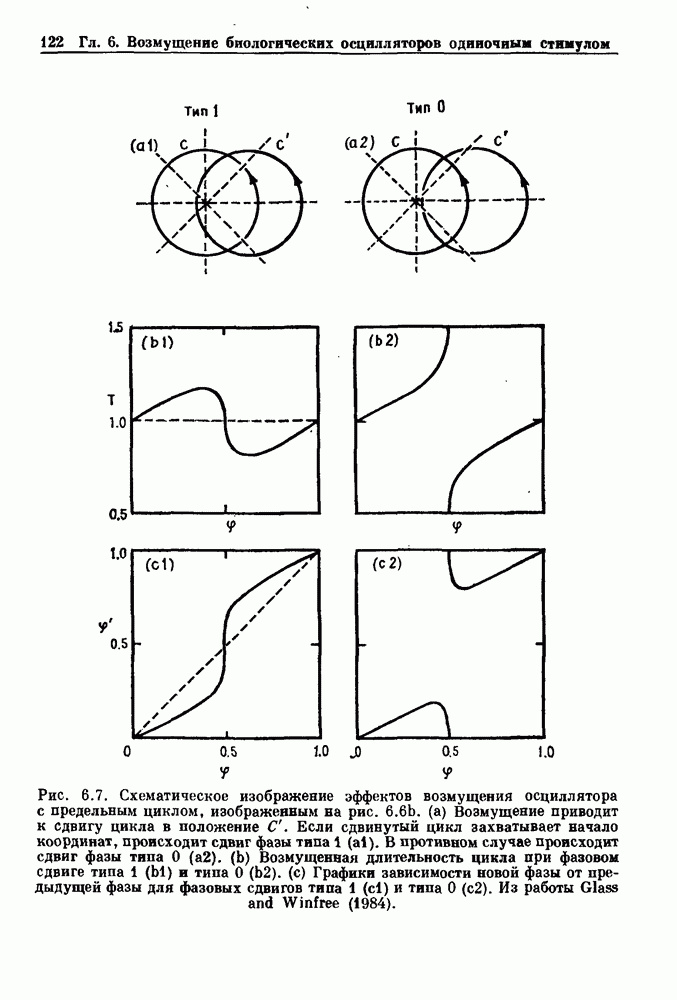
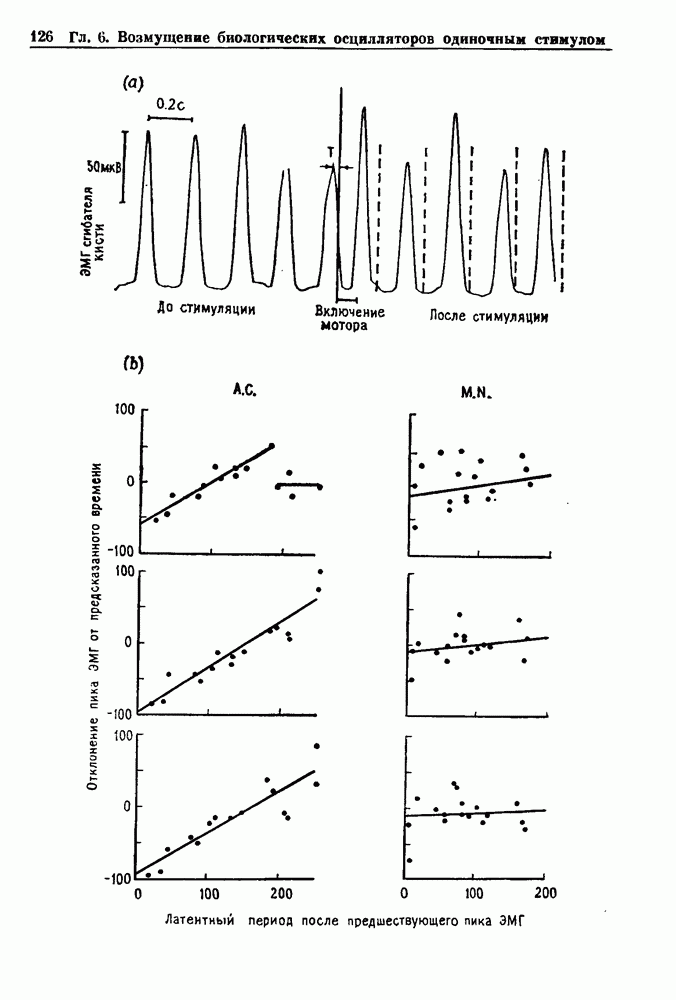
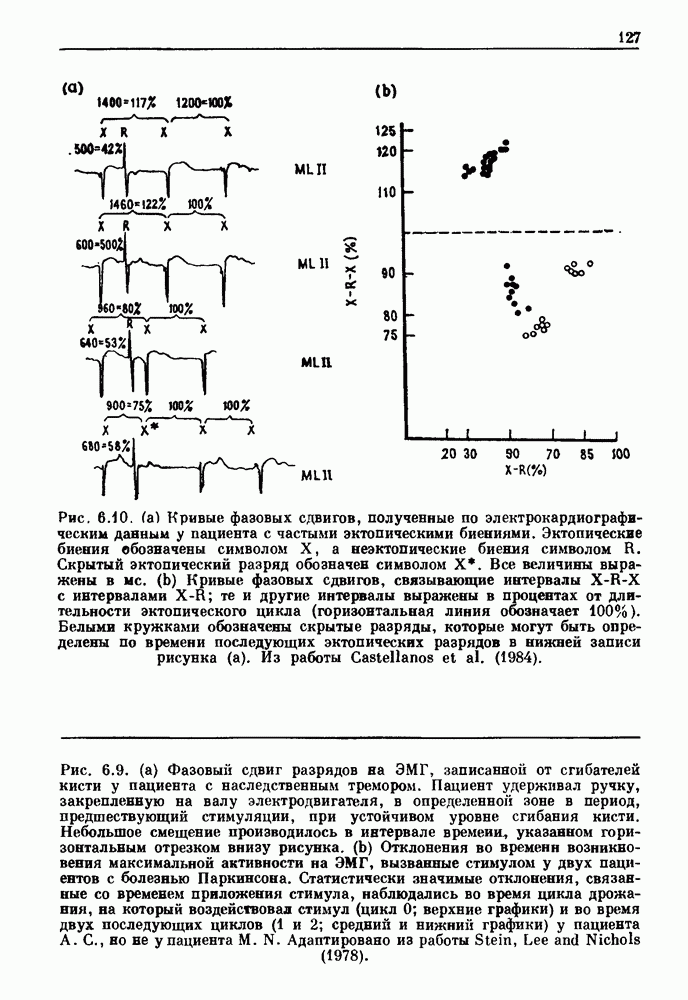
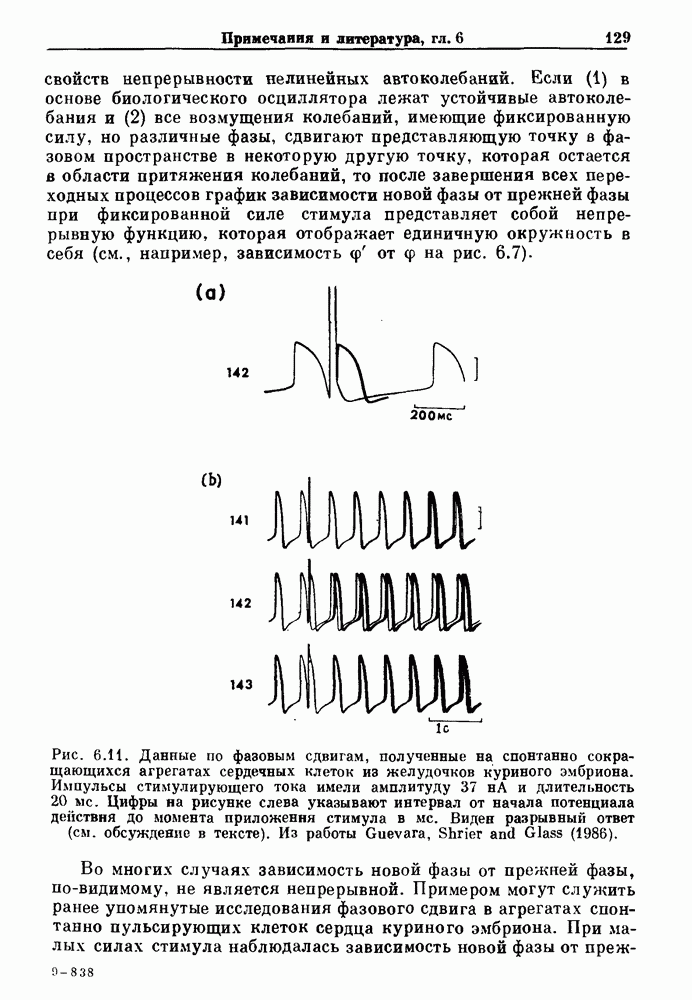
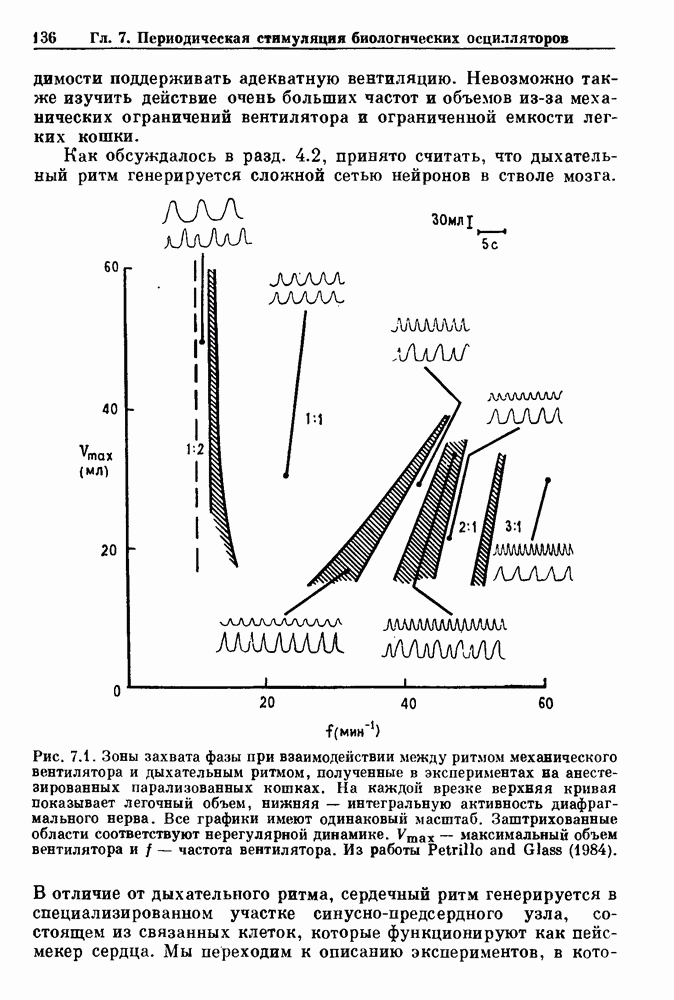
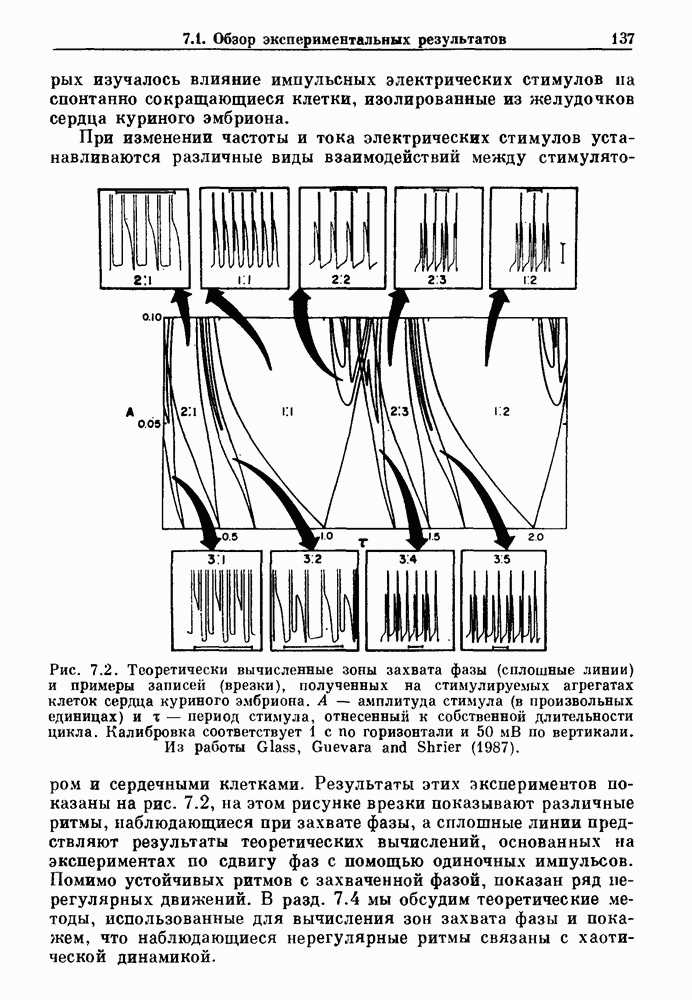
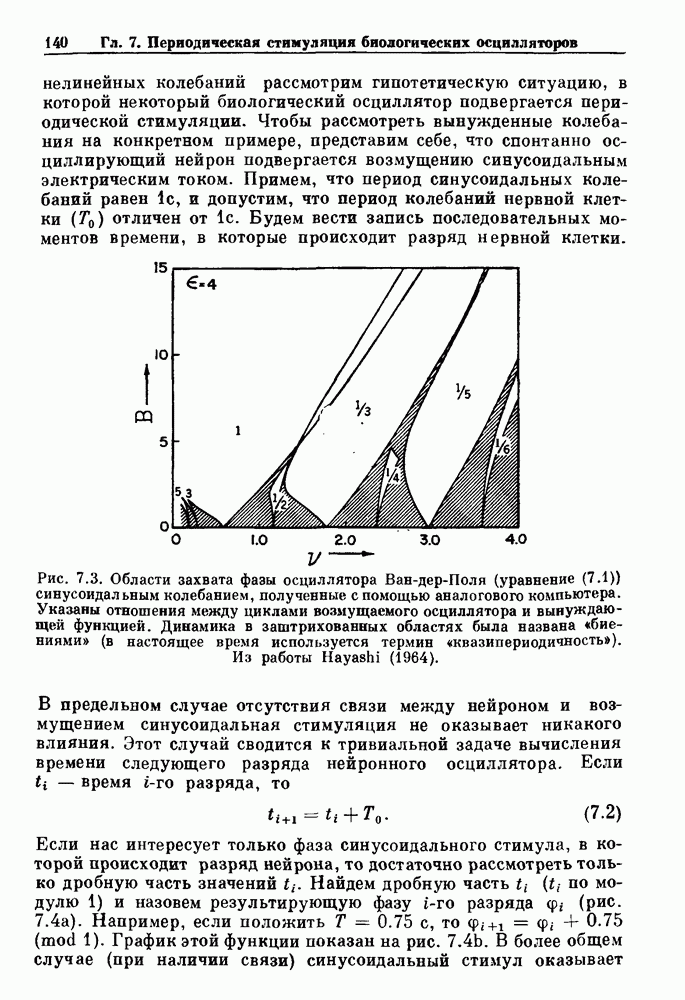
Примечания и литература, глава 66.1. Обзор экспериментальных результатовОбширная литература по фазовым сдвигам в биологических ритмах, вызванным возмущением, подробно обсуждается в работах Winfree (1980, 1987а, b). Описание рефлекса Геринга — Брейера можно найти в оригинальной работе Вгеиег (1868). Adrian (1933) в своей классической работе продемонстрировал роль афферентных волокон блуждающего нерва в модуляции рефлекса. Дальнейшие исследования роли афферентных волокон блуждающего нерва в физиологии дыхания были выполнены многими другими авторами, и мы использовали важные результаты работ Clark and Euler (1972) и Knox (1973). Iscoe and V anner (1980) изучали длительный временной сдвиг дыхательного ритма, вызванный стимуляцией блуждающего нерва. Современный обзор дыхательного ритмогенеза, включающий рефлекторные механизмы блуждающего нерва, см. в монографии Feldman (1986). Фазовый сдвиг в препаратах волокон Пуркинье описан в статье Jalife and Мое (1976, 1979), обзор литературы на эту тему можно найти в работе Jalife и Michaels (1985). 6.2. Фазовый сдвиг в релаксационных моделяхМногие исследователи использовали релаксационные модели для описания респираторной активности. Примерами могут служить работы Bradley et а) (1975), Herczynski and Karczewski (1976), Remmers (1976), Cohen and Feldman (1977), Baconnier et al (1983) и Petrillo and Glass (1984). Столь же многочислены работы, в которых в качестве моделей центрального генератора дыхательного ритма предложены нелинейные дифференциальные уравнения, имеющие решения в виде предельного цикла (Feldman and Cowan (1975); Geman and Miller (1976), Finchman and Liassides (1978); Pham Dinh et al. (1983); Baconnier et al. (1983)). В некоторых предельных случаях автогенератор можно аппроксимировать релаксационной моделью. Впервые эта мысль была высказана при обсуждении регуляции митоза в работе Tyson and Sachsenmaier (1978). Другой пример такого соответствия, относящийся к дыханию, дается у Petrillo and Glass (1984). 6.3. Сдвиг фазы автогенераторовОсновные источники по теории фазовых сдвигов предельных циклов — это книги и статьи Winfree (1975, 1977, 1980, 1987 a,b), в которых можно найти обширную информацию по истории вопроса. примеры и ссылки на другие источники. Теоретическое обсуждение фазовых сдвигов автоколебаний проводится также в работах Pavlidis (1973), Guckenheimer (1975), Kavato and Suzuki (1978) и Kavato (1981). В этой книге данные излагаются в соответствии со статьей Glass and Winfree (1984). Van Meerwijk et al. (1984) применили теорию к экспериментам по фазовым сдвигам сердечного ритма в агрегатах клеток из сердца куриного эмбриона. Возможно, что теория применима также и для объяснения генерации дыхательного ритма. Изучение действия стимуляции верхнего гортанного нерва на дыхательный ритм свидетельствует о том, что при слабых стимулах происходят фазовые сдвиги типа 1, а при сильных стимулах — сдвиги типа0 (Paydarfar, Eldridge and Kiley (1986)). Подобные результаты были получены в экспериментах, в которых электрические стимулы прикладывались непосредственно к мозгу (Paydarfar and Eldridge (1987)). 6.4. Фазовый сдвиг в различных системахЭксперименты по фазовым сдвигам были выполнены для очень большого числа других систем, как уже было отмечено ранее. Особый интерес вызывают исследования жевания (J.P. Lund, Ros-signol and Murakami (1981)) и локомоции (Forssberg et al. (1976)). Фазовый сдвиг использовался в качестве диагностического средства для выявления паркинсонизма (R.B. Stein, Lee and Nichols (1978) и Lee and R.B. Stein (1981)). Анализ модулированных пара-систолических ритмов, основанный на фазовых сдвигах, обсуждался в работах Jalife andMoe (1976, 1979) и Мое et al. (1977). Использование этих идей в клинике было рассмотрено у Furuse, Matsuo and Saigusa (1981), Jalife, Anizelevitch and Мое (1982), Nauet al. (1982) и Castellanos et al. (1984). Хороший обзор был недавно сделан Jalife and Michaelis (1985). Дополнительное обсуждение теории модулированной парасистолы проводится в разд. 7.5. 6.5. Трудности в применении топологической теорииТопологическая теория фазовых сдвигов, кратко изложенная в разд. 6.3, — довольно тонкая вещь, и, к сожалению, часто неправильно истолковывается в экспериментальной литературе. Например, Jalife and Antzelevitch (1979) утверждают: «Его iWinfree] теория, в которой используется метод дифференциальной топологии, предсказывает, что если фазовый сдвиг пейсмекера в ответ на возмущение происходит по некоторому определенному типу [т. е. по типу 1 при малой силе стимула и по типу 0 при большой силе стимула], то должны существовать характерные величина стимула и время его приложения, при которых пейсмекерная активность полностью уничтожается». Даже если это рассуждение и привело Jalife and Antzelevitch (1979) к открытию прерывания потенциалов действия одиночным стимулом, это утверждение тем не менее ошибочно, поскольку может существовать неустойчивая фазовая сингулярность. В этом случае колебания всегда восстанавливаются после возмущения. Количественные характеристики фазовых сдвигов, предсказанных на основании дифференциальных уравнений типа Ходжкина—Хаксли, моделирующих поведение возбудимой мембраны, были даны в работах Best (1979), Chay and Lee (1984, 1985) и Clay, Guevara and Shrier (1984). На основании исследований этих авторов можно ожидать, что наблюдение топологических свойств функций фазовых переходов (например, поведение типа 1 или типа 0) может быть сопряжено с трудностями из-за очень большой КФП. Отсюда возникает возможность объяснения кажущейся разрывности в динамике фазовых переходов, наблюдавшихся в работе Guevara, Shrier and Glass (1986). Теоретические доводы в пользу непрерывности КФП приводятся у Winfree (1983b, 1987b).
|
 |
Оглавление
|