Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
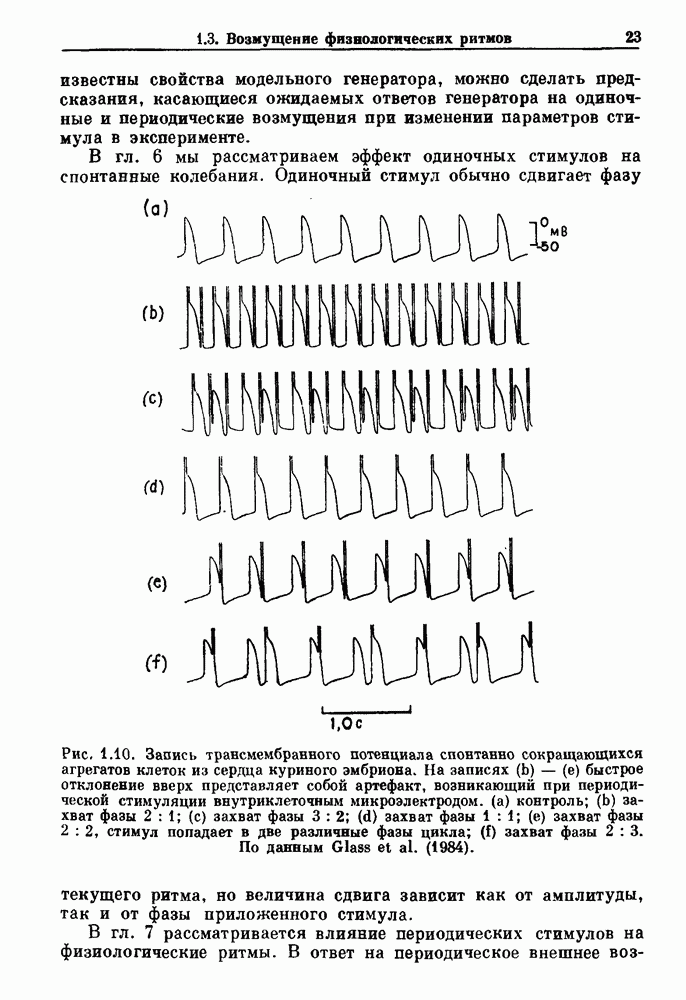
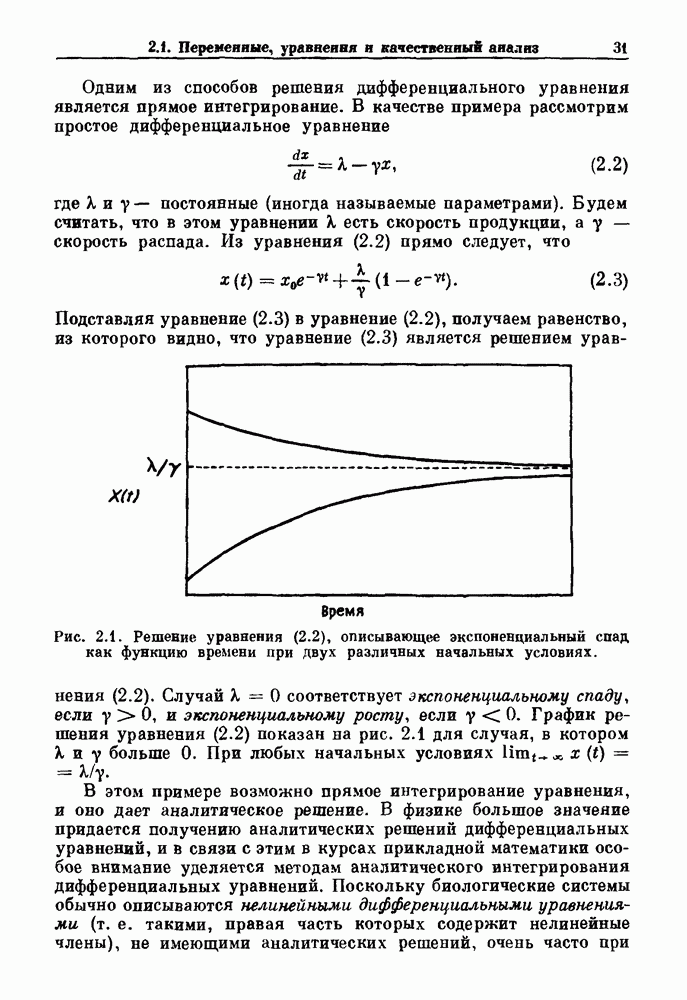
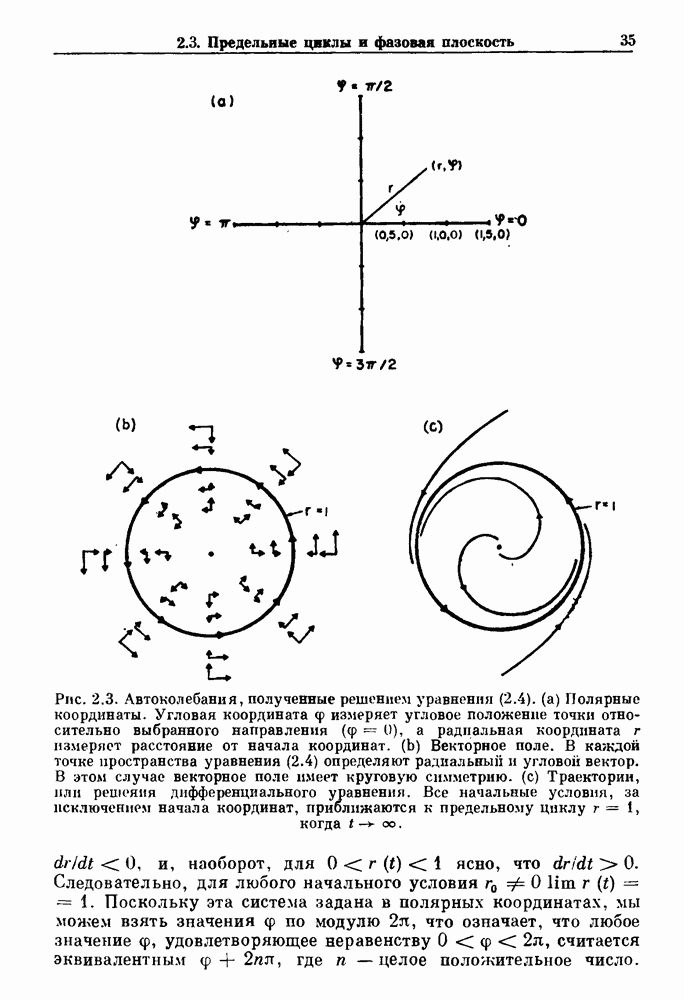
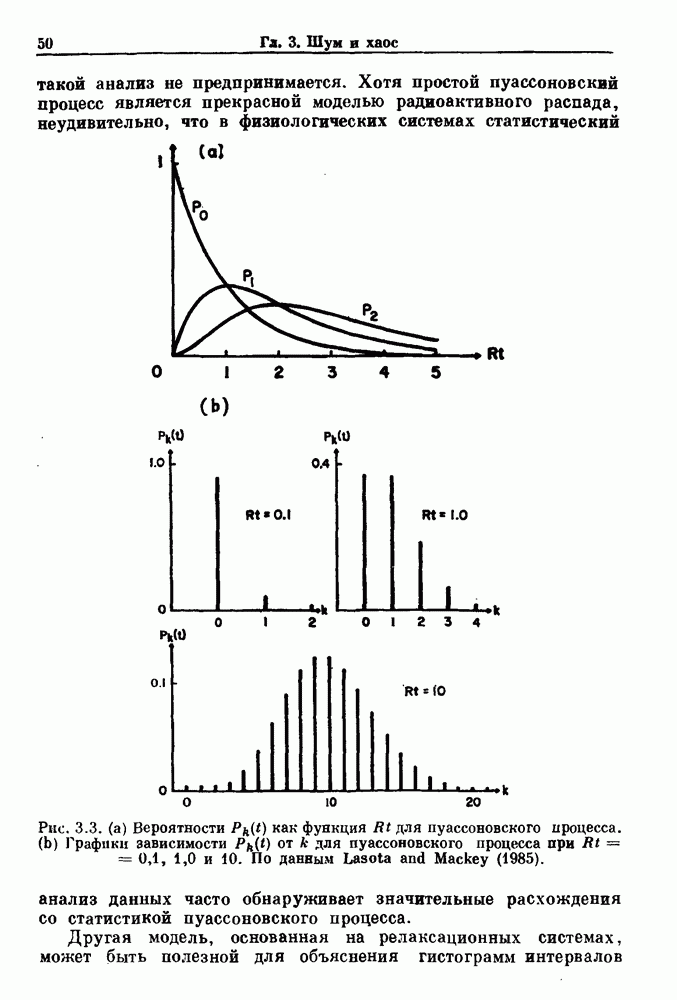
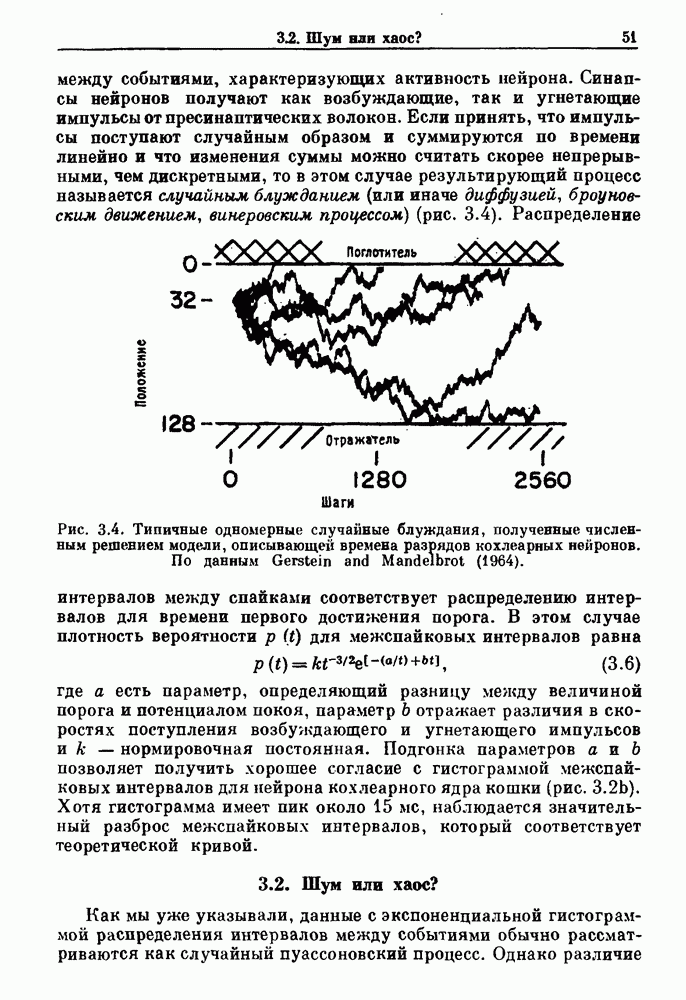
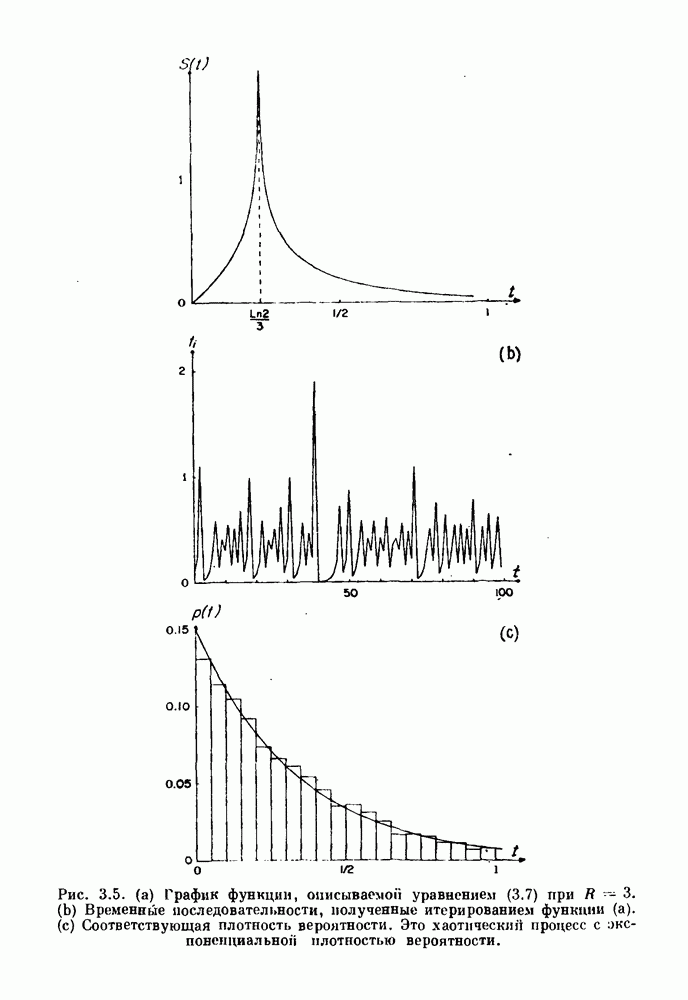
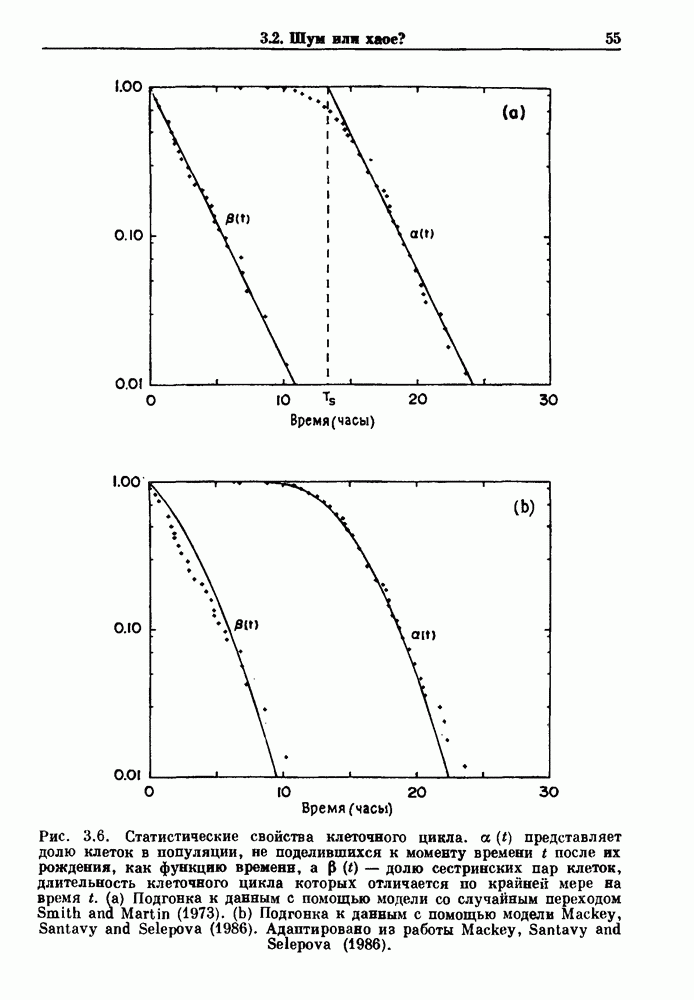
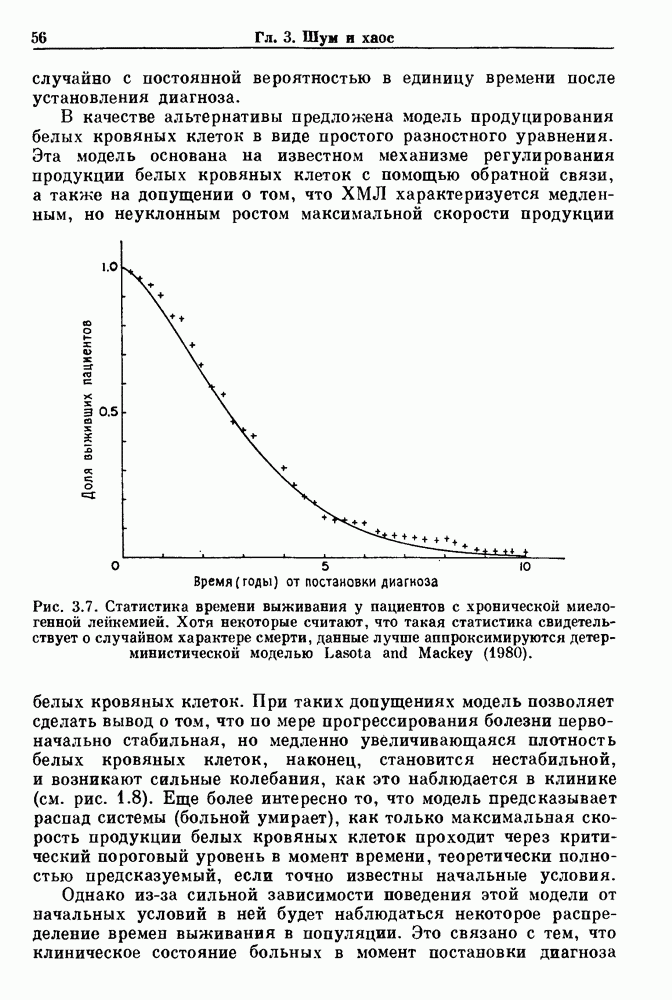
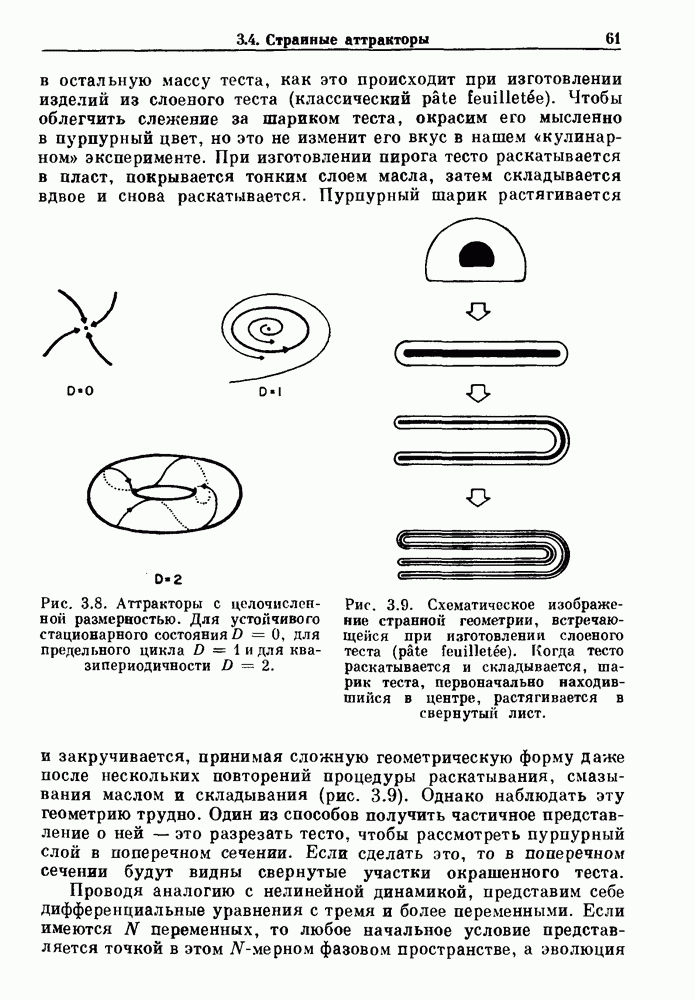
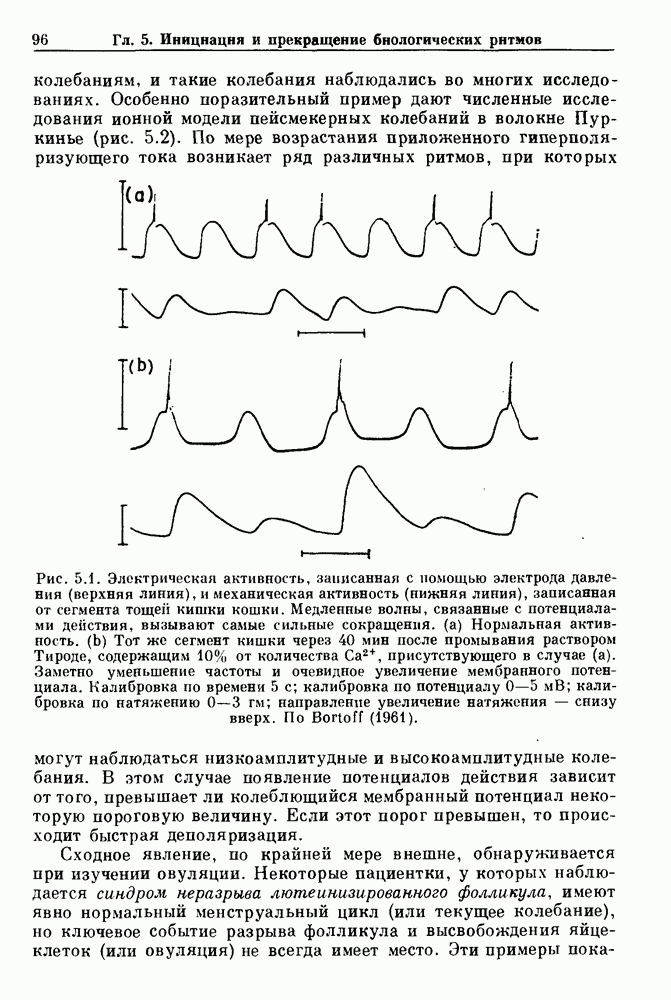
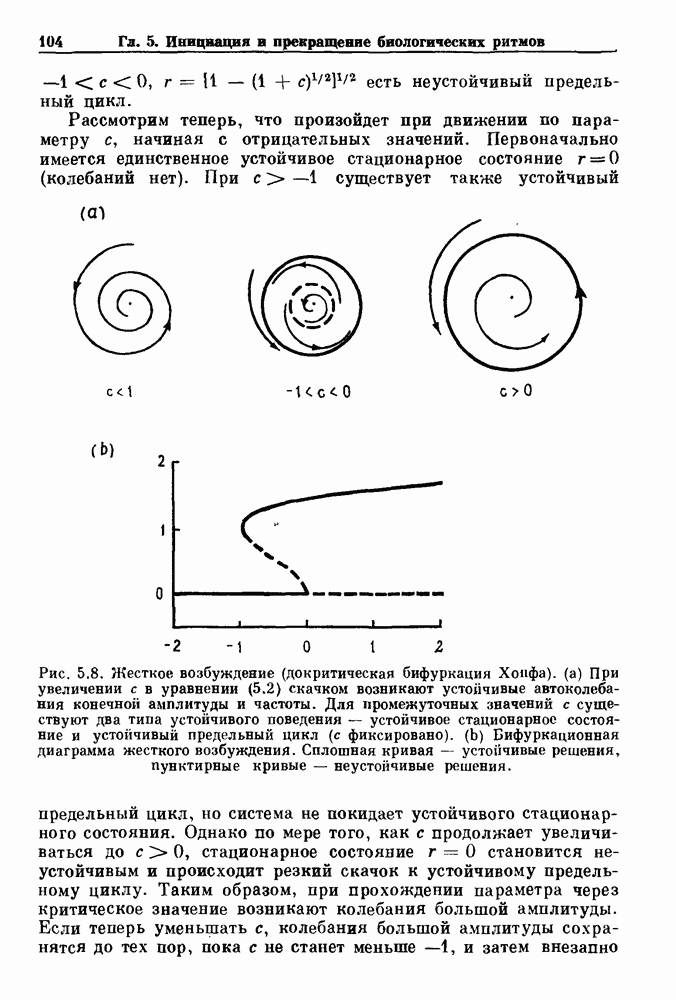
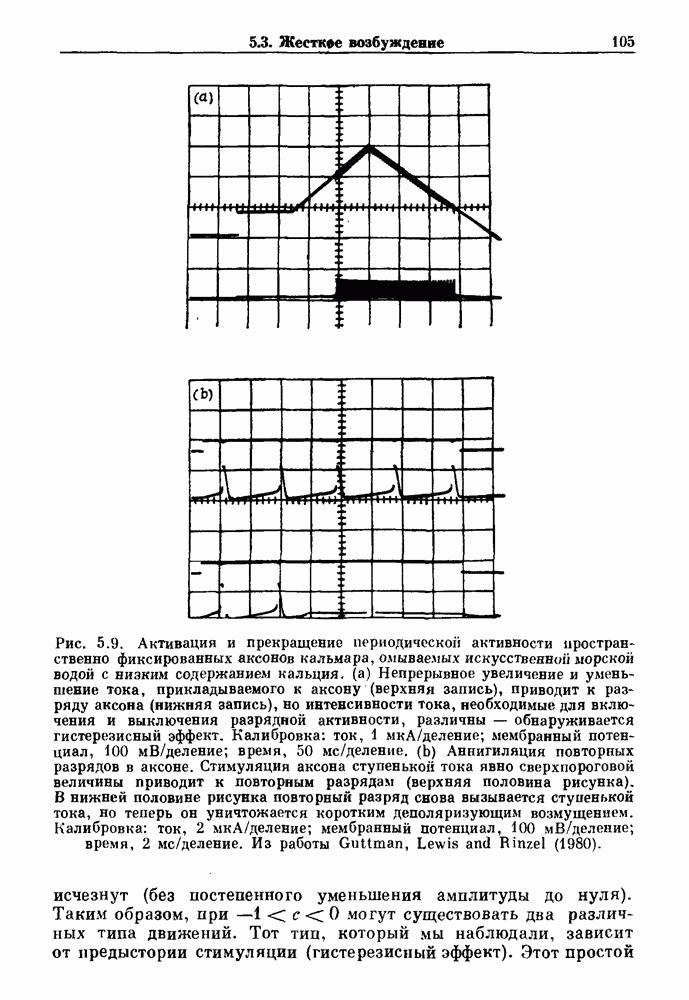
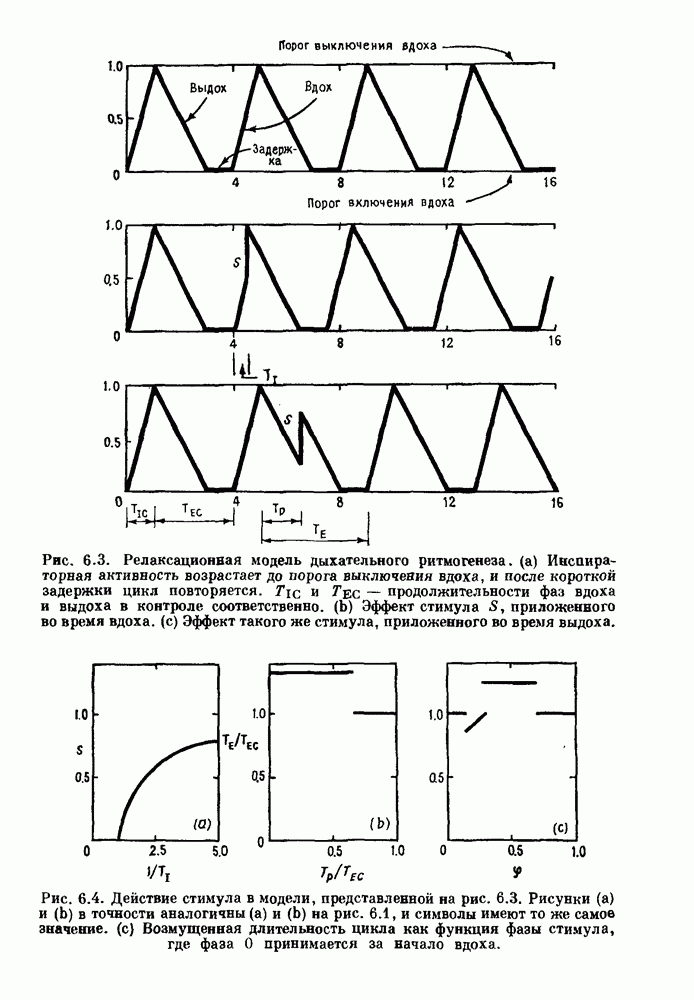
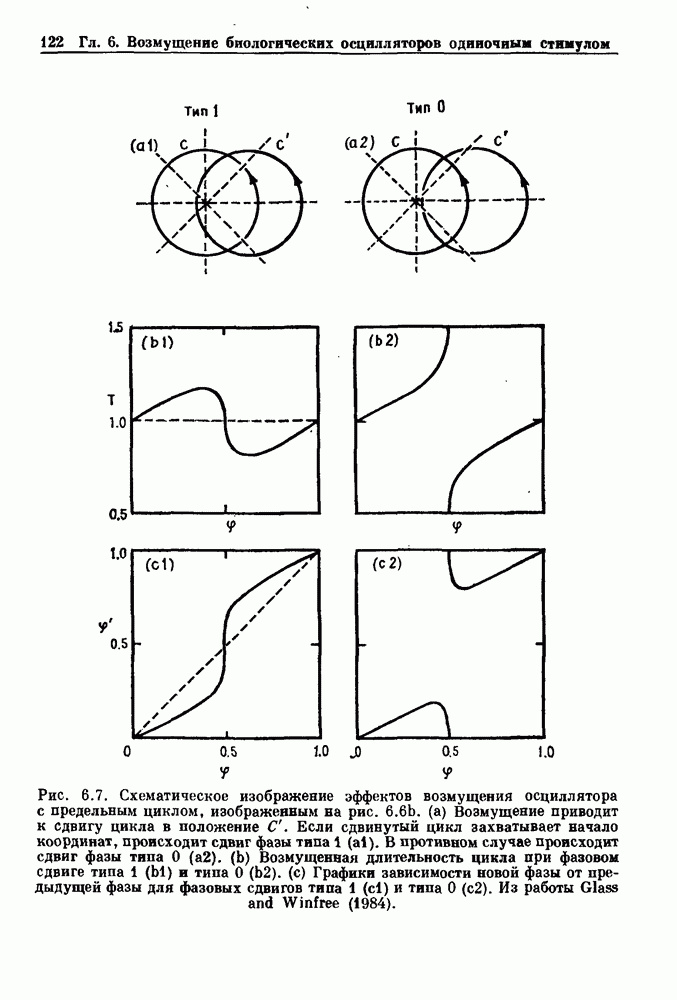
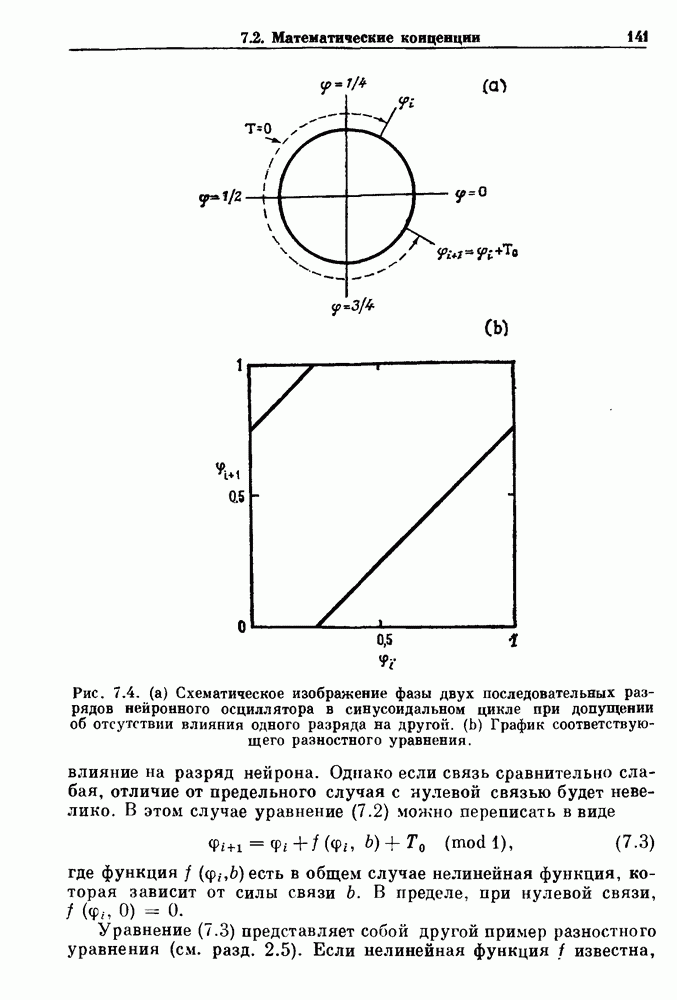
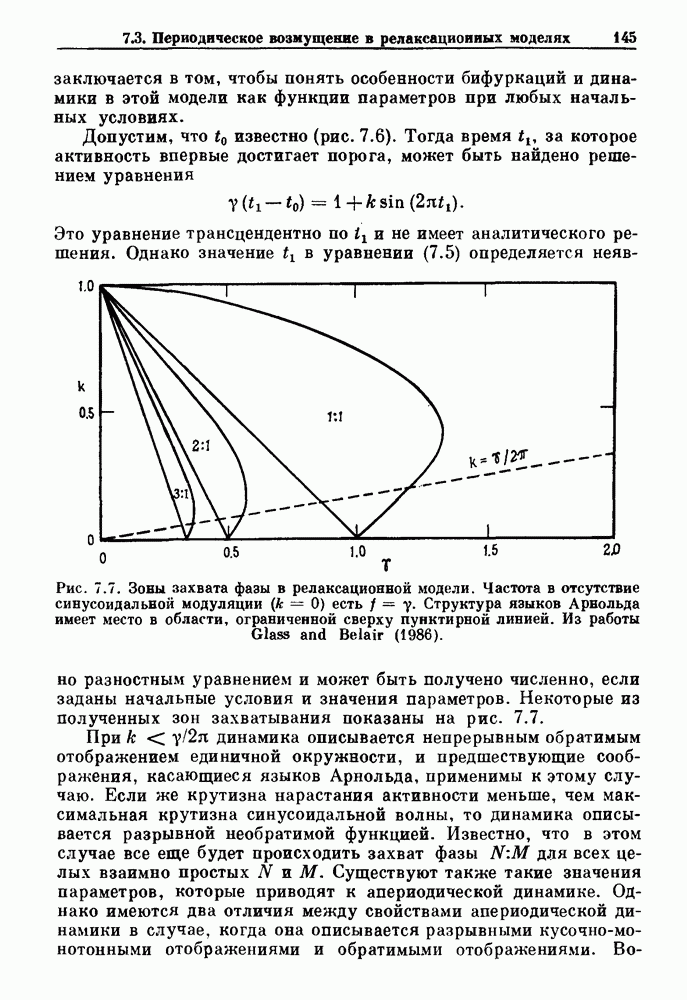
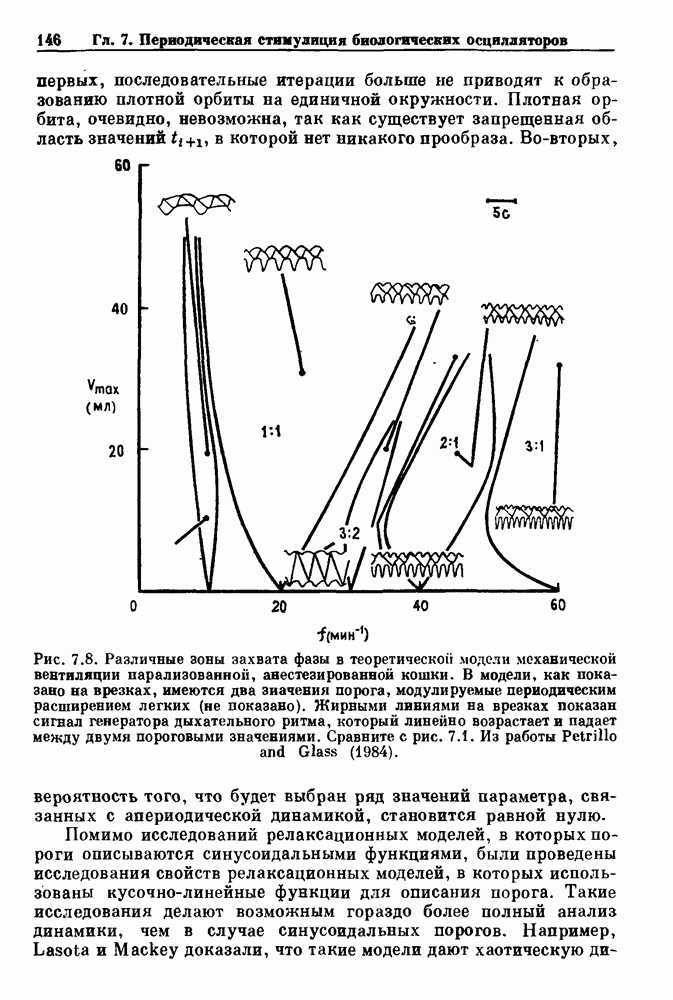
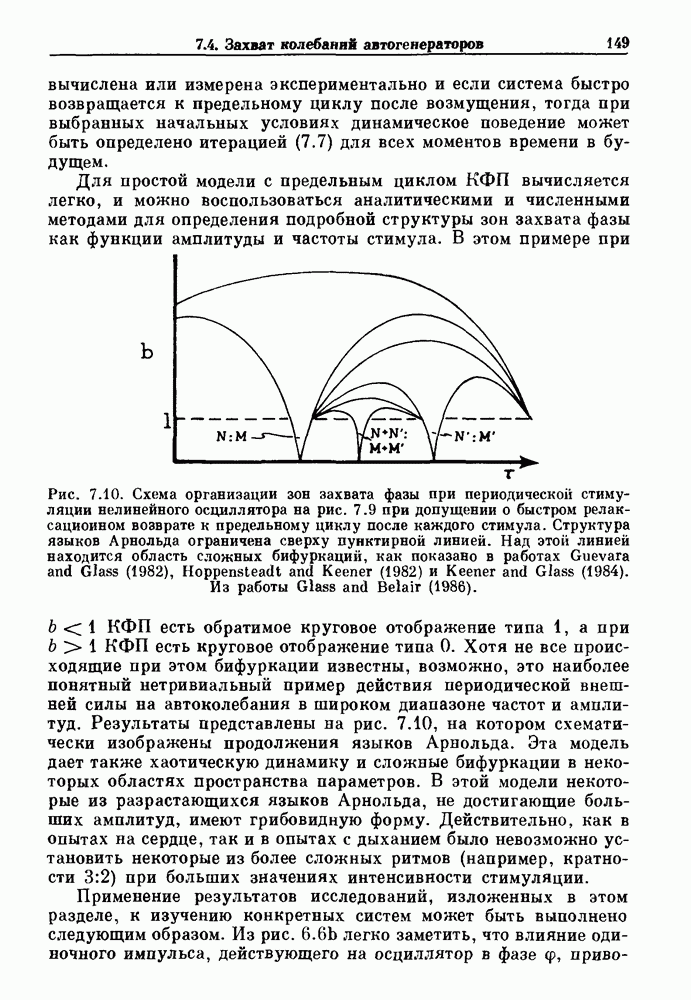
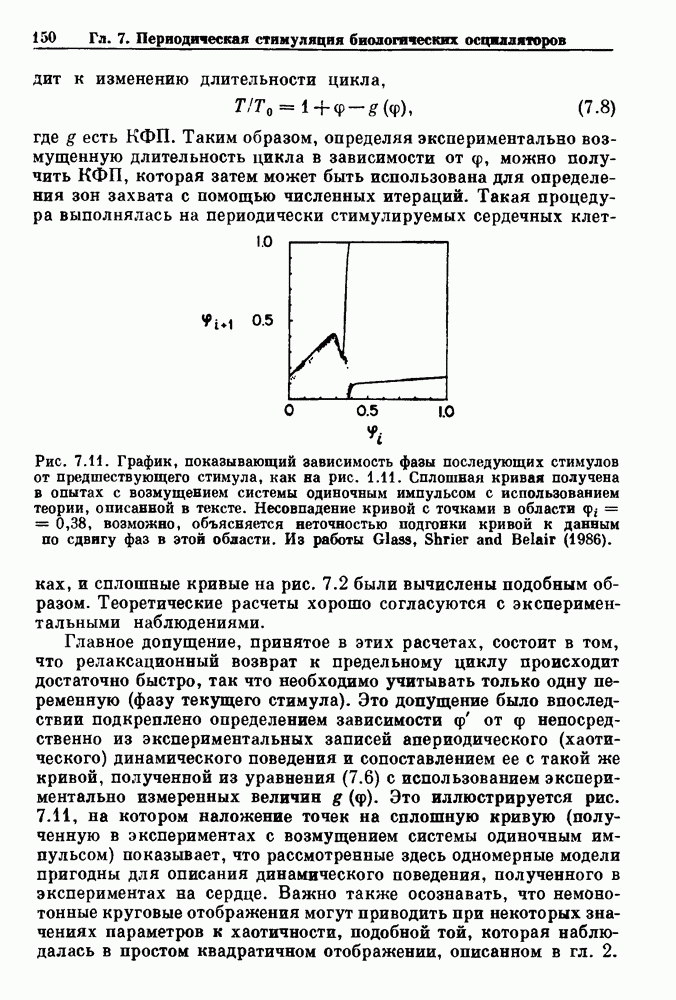
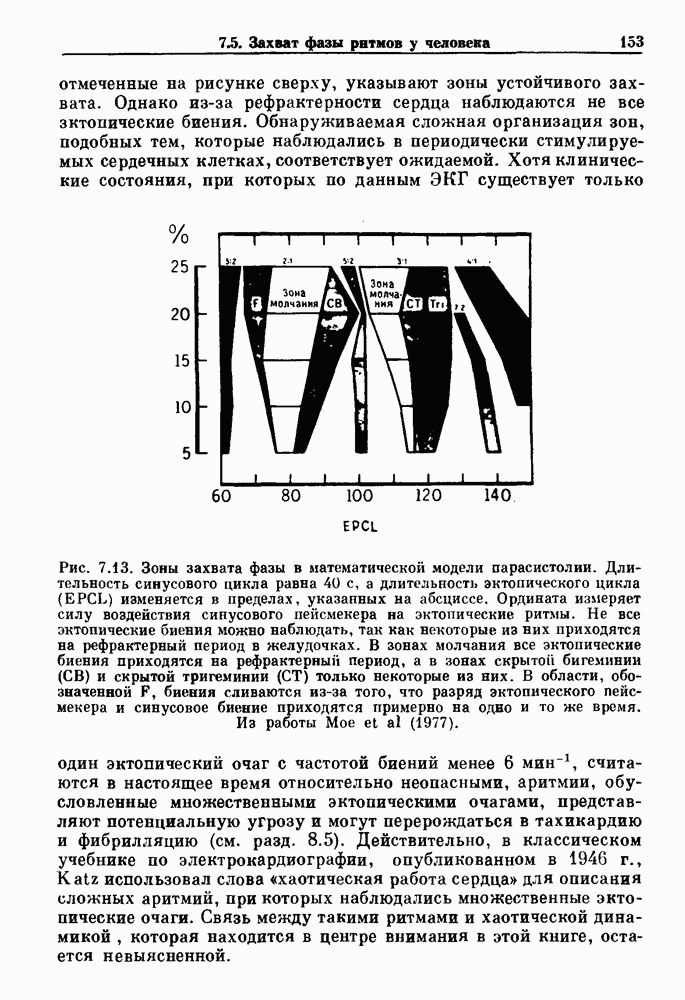
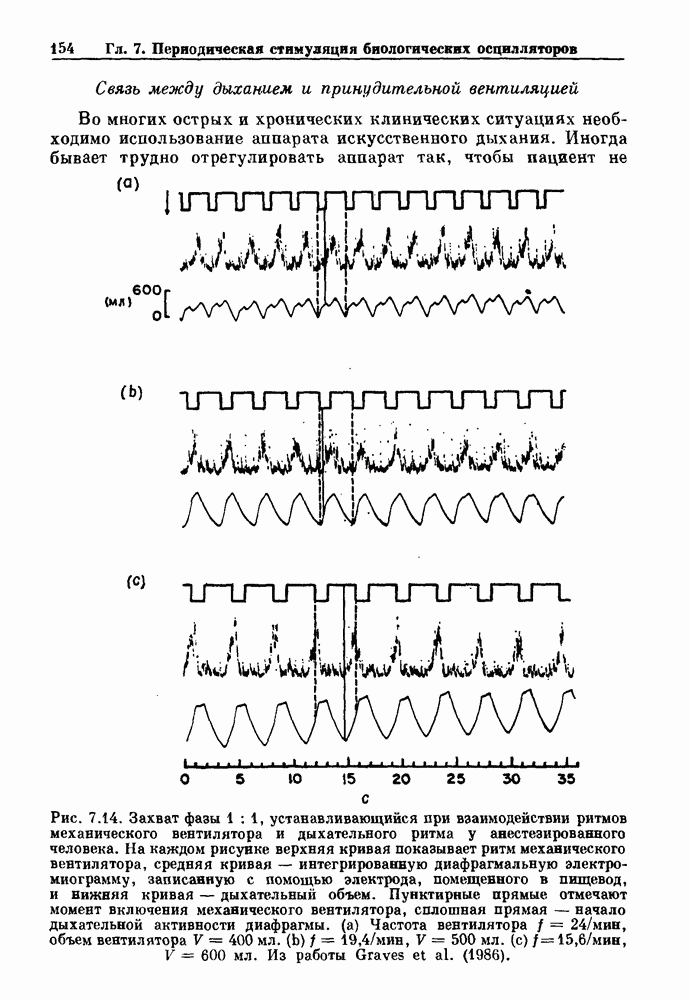
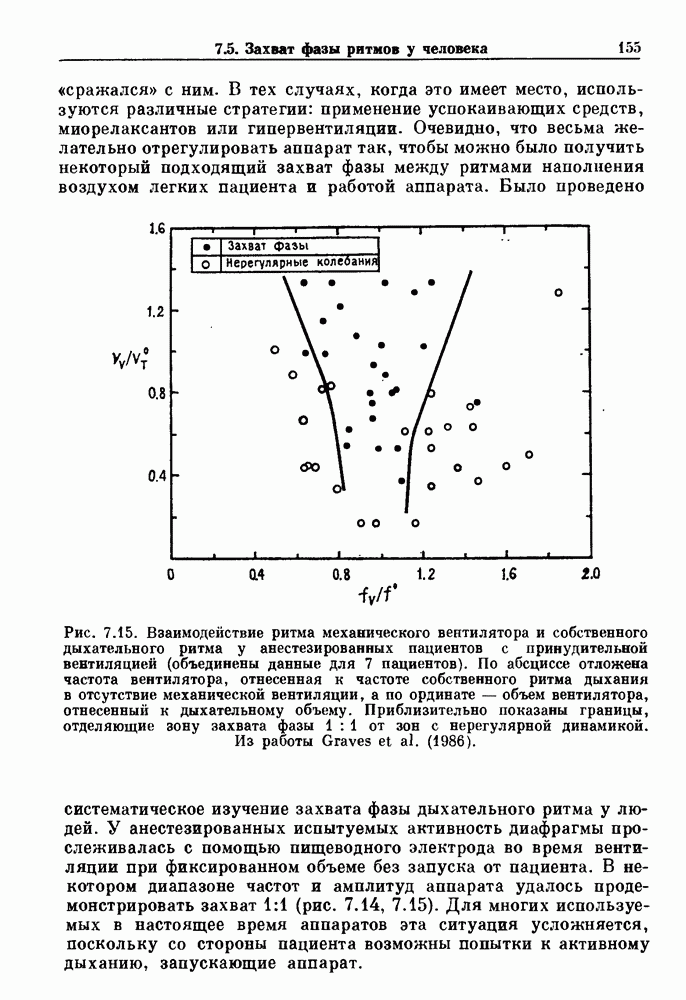
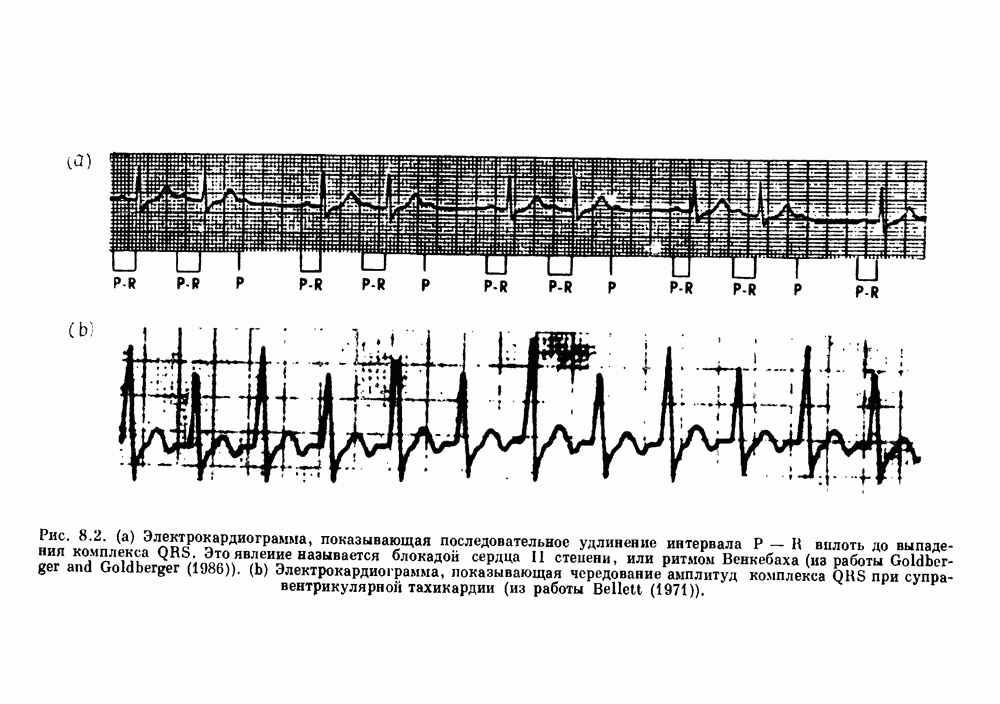
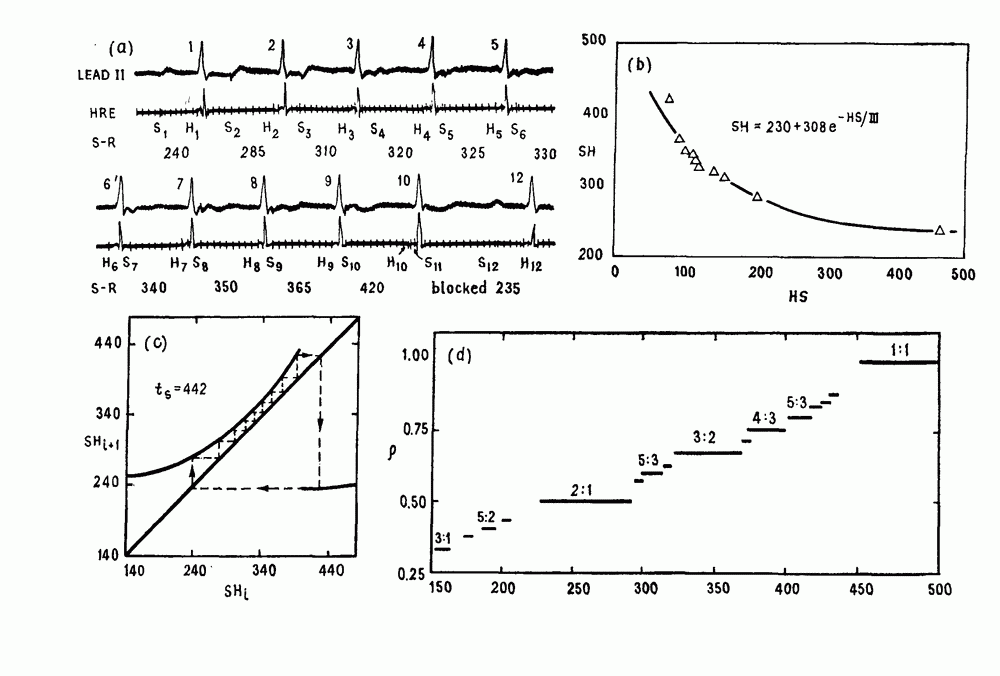
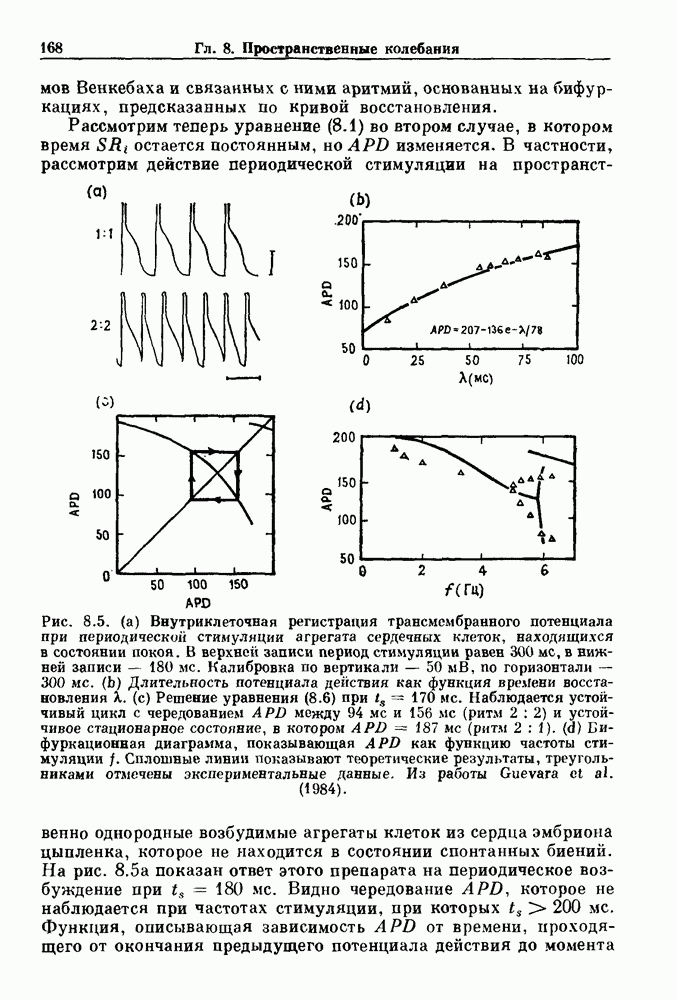
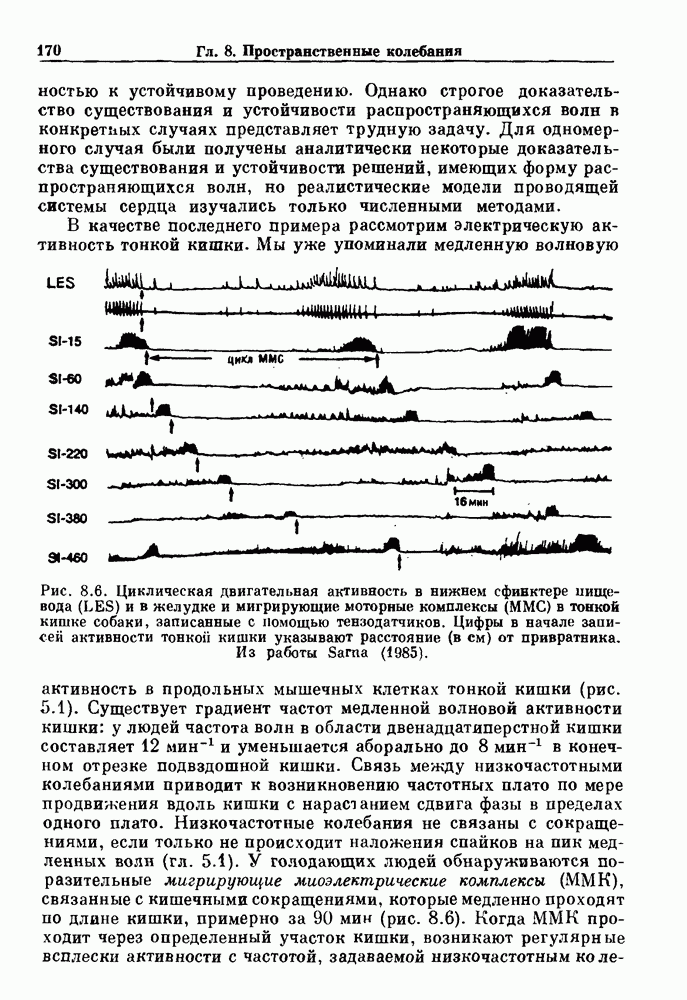
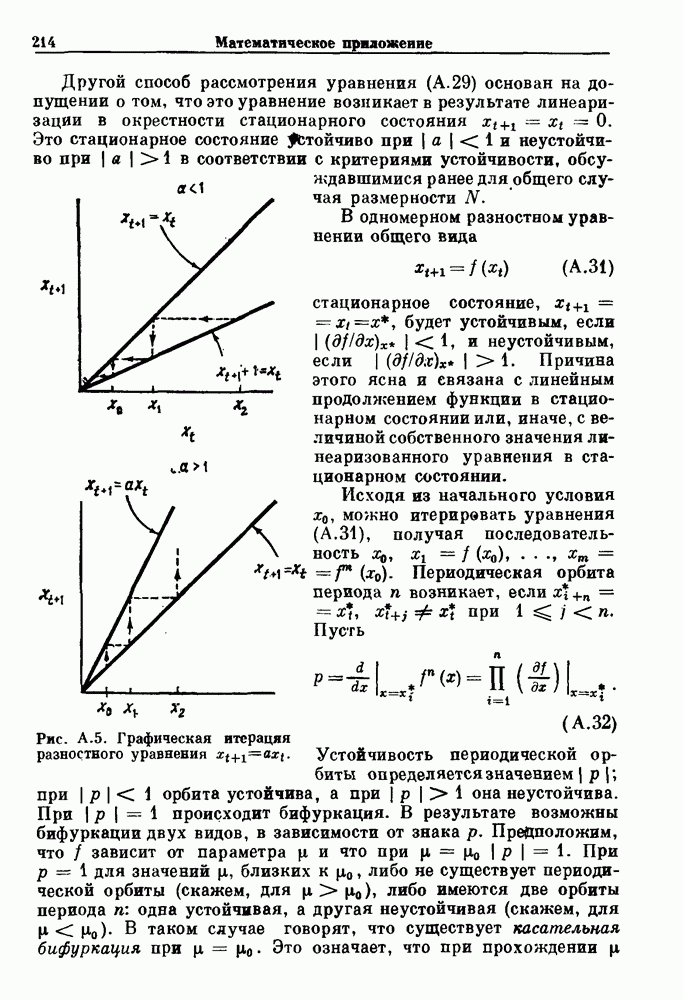
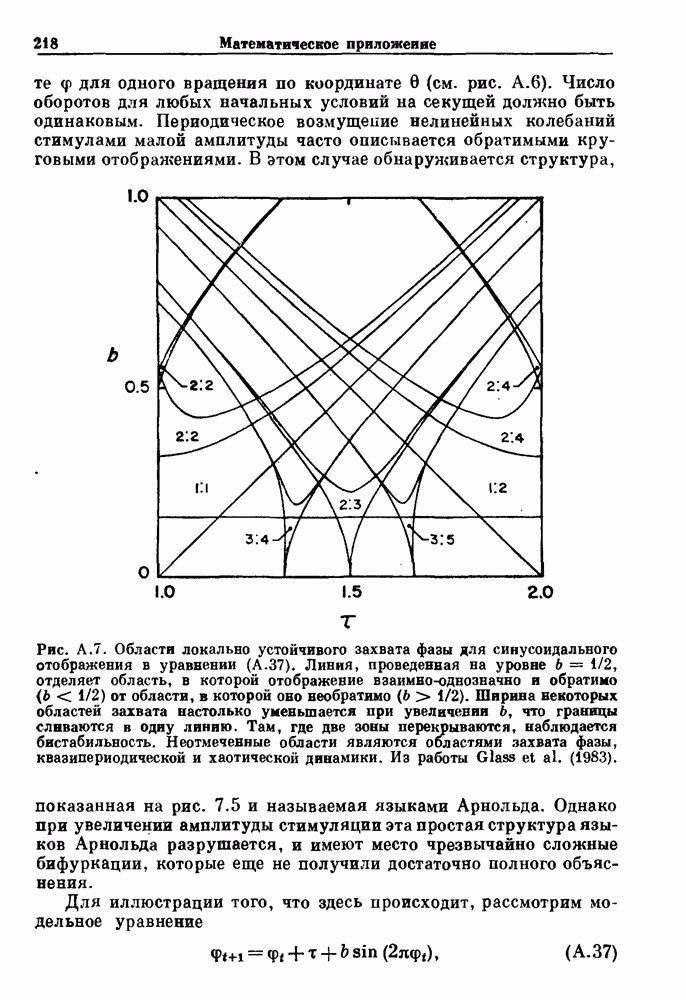
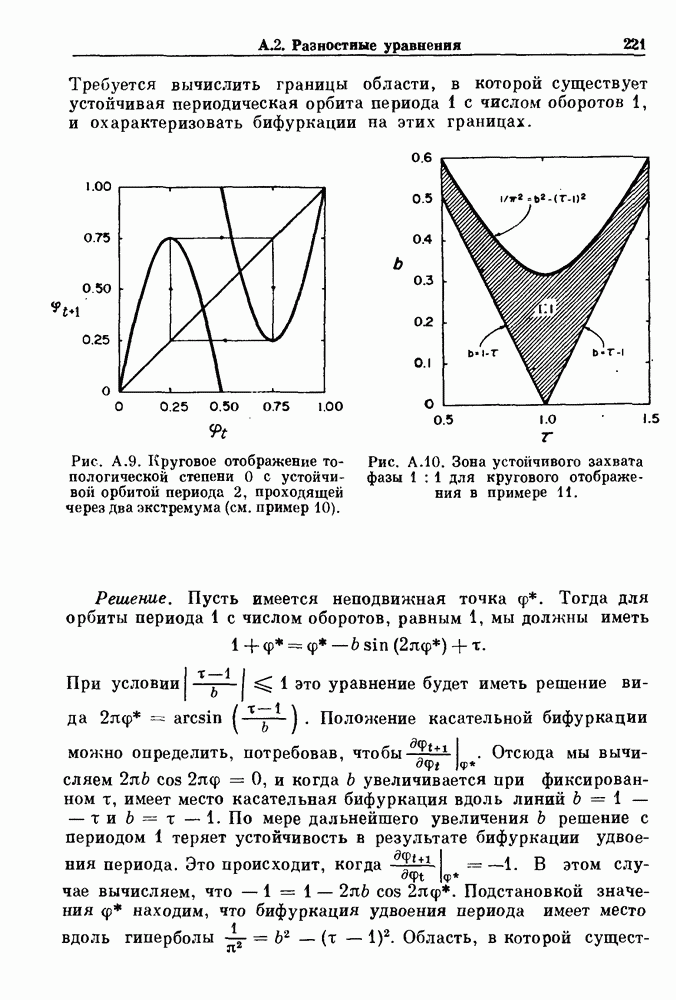
Глава 4. Математические модели биологических колебанийСуществует обширная литература, предлагающая множество различных моделей генерации физиологических ритмов. В этой главе с помощью соответствующих примеров рассматриваются основные классы механизмов действия биологических генераторов. В разд. 4.1 описываются принципы устройства генераторов пейсмекерных колебаний. Затем в разд. 4.2 мы обсуждаем генерацию двигательных ритмов генераторами центрального типа. Два из предложенных механизмов для таких генераторов — взаимное ингибирование и последовательное деингибирование — рассматриваются в разд. 4.3 и 4.4. Один из основных механизмов регулирования ритмов в организме связан с отрицательной обратной связью. В разд. 4.5 мы показываем, что такие системы могут терять устойчивость, в результате чего возникают устойчивые колебания. Системы с комбинированной положительной и отрицательной обратной связью рассматриваются в разд. 4.6. Такие системы могут обладать как колебательными, так и хаотическими динамическими свойствами. 4.1. Пейсмекерные колебанияМногие физиологические ритмы генерируются одиночной клеткой или электрически связанными изопотенциальньши клетками, способными генерировать колебания автономно или в присутствии постоянного сигнала. Мы называем такие клетки или группы клеток пейсмекерами. Полагают, что пейсмекерные колебания связаны с организацией колебательного поведения сердца, гладкой мышцы, многих гормональных систем и нейронов. Несмотря на то, что нам хотелось бы выделить из обширной литературы по пейсмекерным колебаниям несколько простых механизмов или принципов, это невозможно, так как, по-видимому, в основе ритмогенеза в различных системах лежат различные механизмы. Остается неясным, действительно ли это так или это всего лишь результат предварительных оценок, которые будут изменяться в ходе будущей экспериментальной работы. Проиллюстрируем вывод моделей пейсмекерных колебаний, рассмотрев в общих чертах математическую модель Ходжкина — Хаксли, описывающую периодическую генерацию потенциалов действия в гигантском аксоне кальмара при низкой концентрации кальция во внеклеточной среде. Как было показано Ходжкином и Хаксли в 1952 г., потенциал действия генерируется в результате изменения зависящей от времени и трансмембранного потенциала проводимости мембраны для натрия и калия. В гигантском аксоне кальмара, как и в других клетках, наблюдается относительно высокая концентрация натрия и низкая концентрация калия во внеклеточной жидкости по сравнению с внутриклеточной средой. Если бы мембрана была проницаема только для одного вида ионов, то трансмембранный потенциал отражал бы результирующий поток этого единственного вида, и результирующий равновесный потенциал для этого иона можно было бы вычислить по уравнению Нёрнста. Например, в случае натрия результирующий поток ионов был бы направлен внутрь клетки до тех пор, пока электрический потенциал, установившийся благодаря притоку ионов натрия, не уравновесил бы диффузию натрия в клетку, обусловленную его концентрационным градиентом. При равновесном потенциале Нёрнста сохраняется диффузия через мембрану, но ионные потоки, направленные внутрь, равны потокам, направленным наружу. По условию измерения всех потенциалов относительно внеклеточной жидкости, равновесный потенциал натрия положителен, тогда как равновесный потенциал калия отрицателен. Обозначив через 1 приложенный к мембране ток, Ходжкин и Хаксли предположили, что
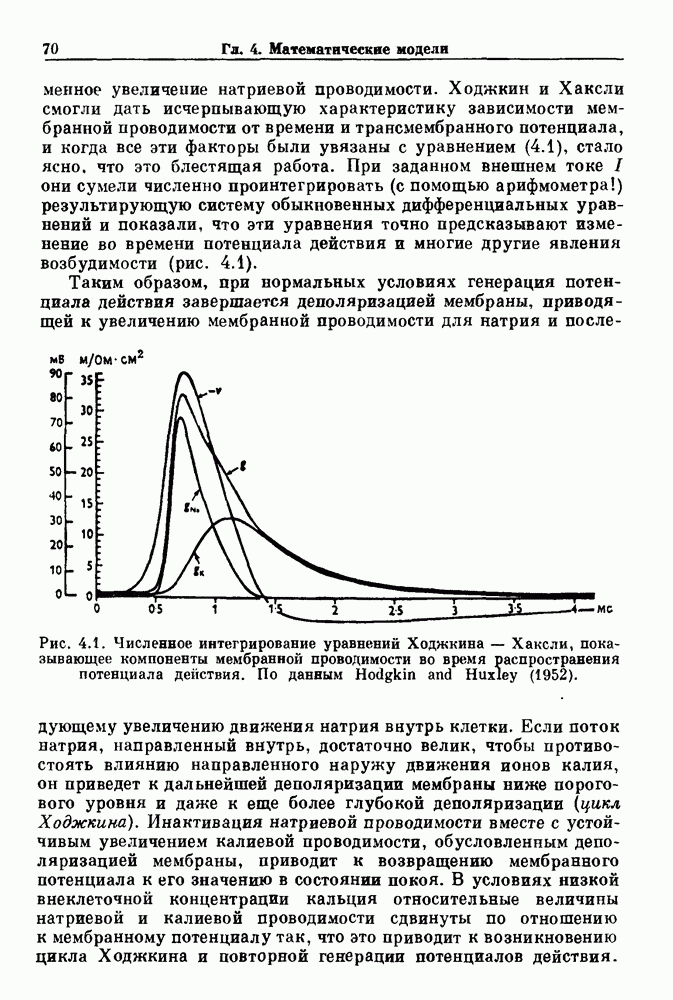
где С — емкость мембраны, V — мембранный потенциал, Для характеристики этой нелинейности Ходжкин и Хаксли использовали метод фиксации потенциала, при котором на мембрану подается постоянное напряжение для поддержания трансмембранного потенциала на заданной постоянной величине. При фиксированном потенциале временное увеличение натриевой проводимости. Ходжкин и Хаксли смогли дать исчерпывающую характеристику зависимости мембранной проводимости от времени и трансмембранного потенциала, и когда все эти факторы были увязаны с уравнением (4.1), стало ясно, что это блестящая работа. При заданном внешнем токе I они сумели численно проинтегрировать (с помощью арифмометра!) результирующую систему обыкновенных дифференциальных уравнений и показали, что эти уравнения точно предсказывают изменение во времени потенциала действия и многие другие явления возбудимости (рис. 4.1). Таким образом, при нормальных условиях генерация потенциала действия завершается деполяризацией мембраны, приводящей к увеличению мембранной проводимости для натрия и последующему увеличению движения натрия внутрь клетки.
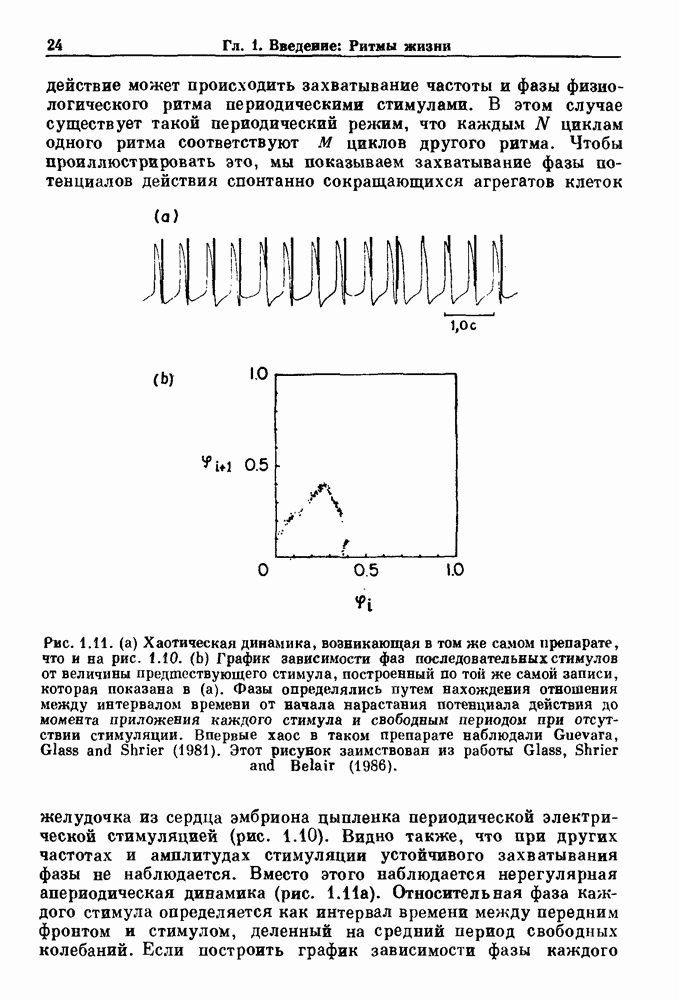
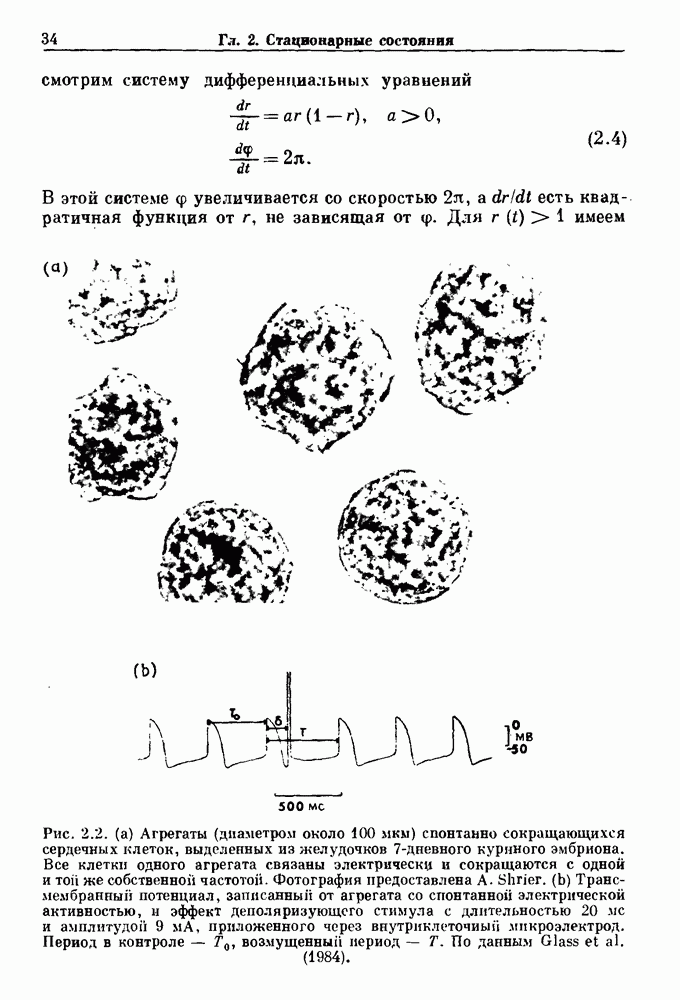
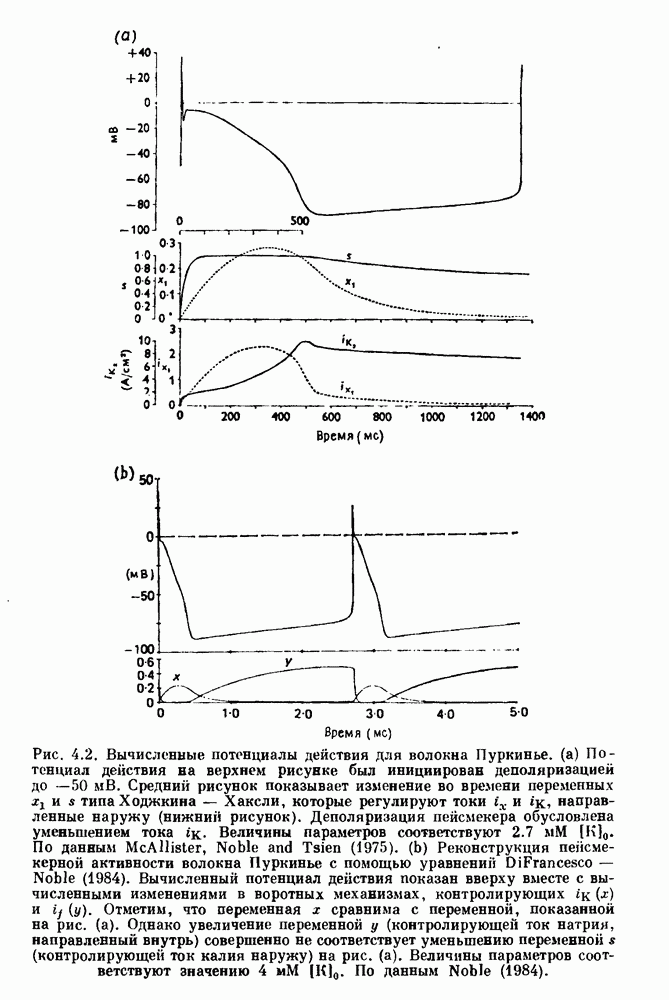
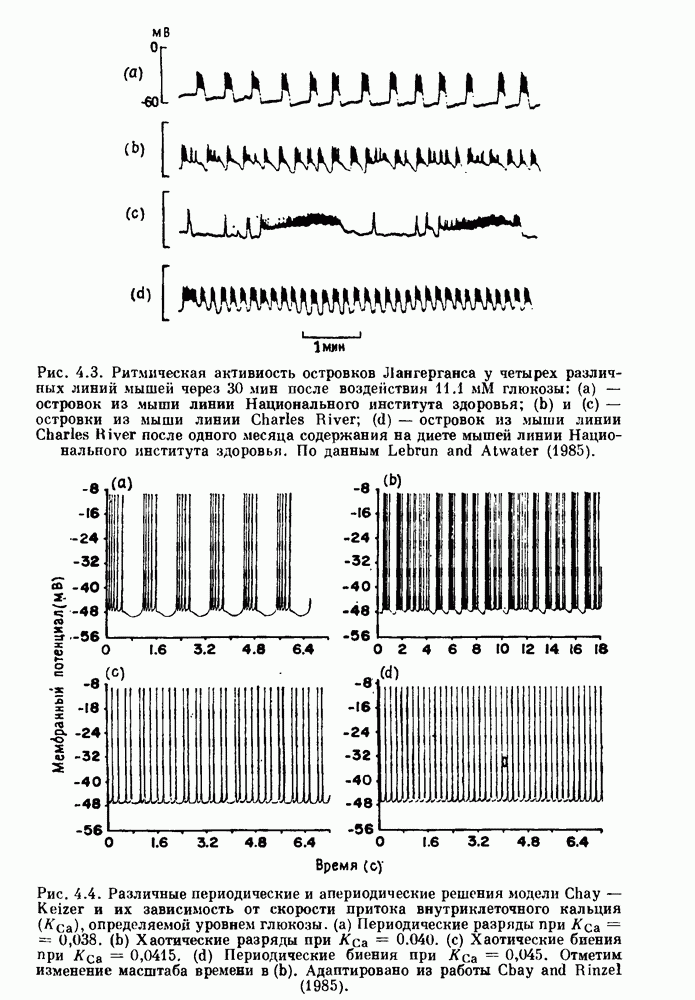
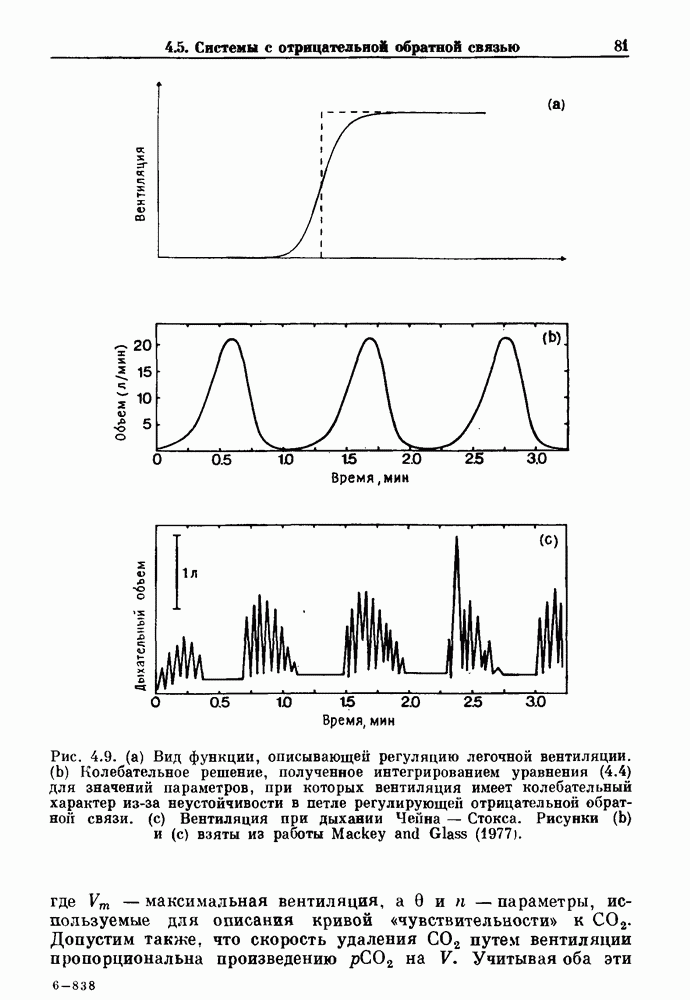
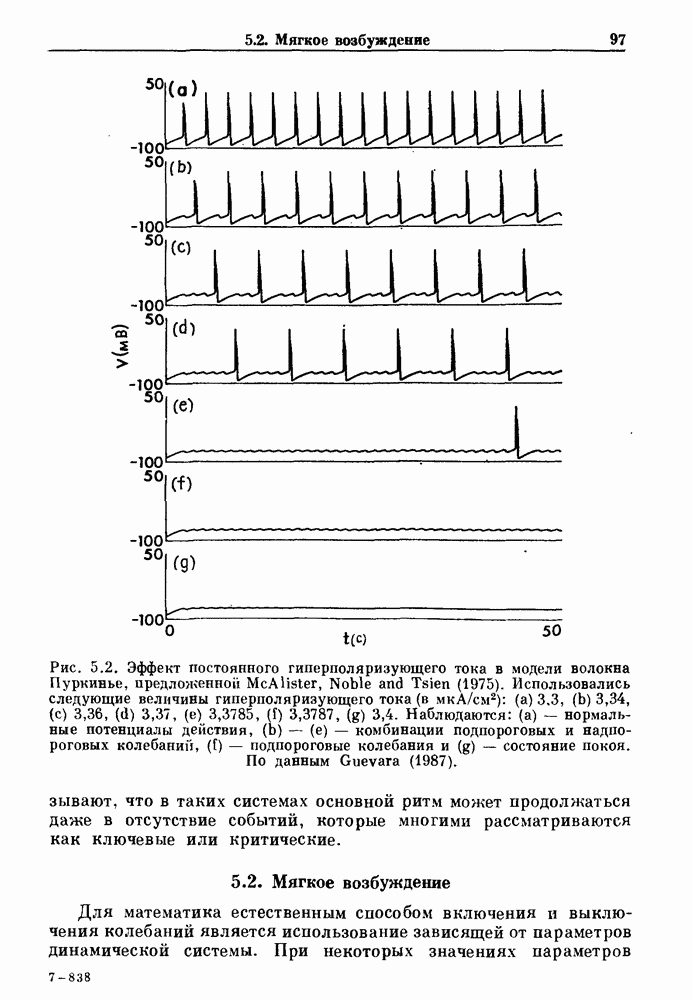
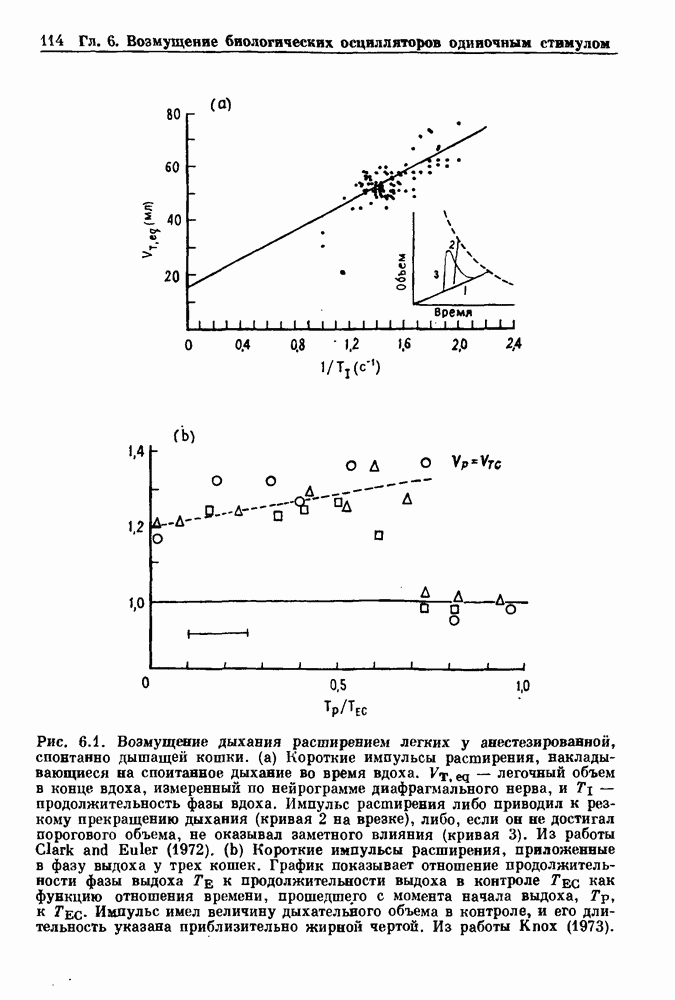
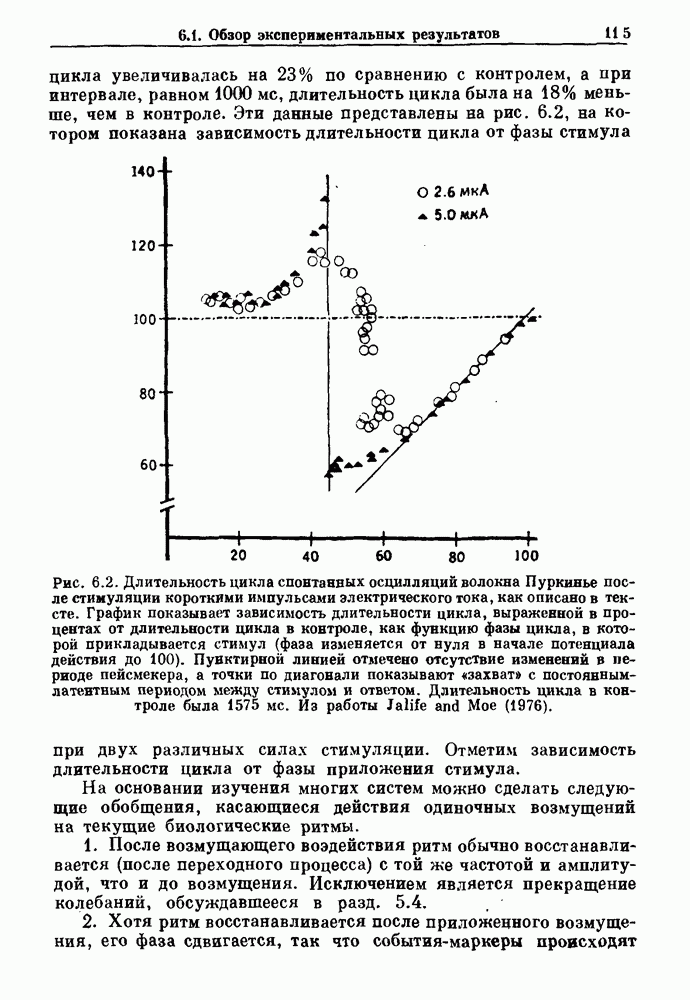
Рис. 4.1. Численное интегрирование уравнений Ходжкина — Хаксли, показывающее компоненты мембранной проводимости во время распространения потенциала действия. По данным Hodgkin and Huxley (1952). Если поток натрия, направленный внутрь, достаточно велик, чтобы противостоять влиянию направленного наружу движения ионов калия, он приведет к дальнейшей деполяризации мембраны ниже порогового уровня и даже к еще более глубокой деполяризации (цикл Ходжкина). Инактивация натриевой проводимости вместе с устойчивым увеличением калиевой проводимости, обусловленным деполяризацией мембраны, приводит к возвращению мембранного потенциала к его значению в состоянии покоя. В условиях низкой внеклеточной концентрации кальция относительные величины натриевой и калиевой проводимости сдвинуты по отношению к мембранному потенциалу так, что это приводит к возникновению цикла Ходжкина и повторной генерации потенциалов действия. Метод, разработанный Ходжкином и Хаксли, был использован для изучения механизмов ритмогенеза во многих других системах, особенно механизмов сердечного ритмогенеза. В препаратах сердца меньший размер клеток, а также трудности в получении хорошо перфузируемых препаратов интактных клеток усложняют опыты. Главные проблемы связаны с определением механизмов, лежащих в основе деполяризующего перехода при так называемых пейсмекерных потенциалах. В принципе по крайней мере два механизма могут привести к такому переходу: 1) уменьшение проводимости для потока калия, направленного наружу; 2) увеличение проводимости для потока натрия, направленного внутрь. В 1975 г. McAllister, Noble и Tsien высказали предположение, что главный вклад в потенциал сердечного пейсмекера связан с первым механизмом (рис. 4.2а), но современные данные, полученные Di Francesco и Noble, по-видимому, свидетельствуют о том, что главный вклад в потенциал пейсмекера обусловлен током натрия или какого-нибудь другого иона, направленного внутрь клетки (рис. 4.2 Ь). Трудности, связанные с применением метода фиксации потенциала к сердечным клеткам, частично возникают из-за большого числа различных ионных токов, существующих в сердечных пейсмекерах. В то время как в аксоне кальмара существуют два главных тока (натриевый и калиевый), которые могут быть разделены, в сердечных клетках намного больше токов (более десяти), и экспериментальные и теоретические трудности пропорционально возрастают. Даже в случае нерва имеющиеся в настоящее время данные свидетельствуют об одновременном действии многих ионных механизмов. За прошедшие несколько лет стало ясно как из экспериментальных, так и теоретических работ, что многие пейсмекеры, способные к генерации регулярных периодических колебаний, могут обнаруживать также нерегулярное динамическое поведение при изменении физиологических параметров или параметров математических моделей. Чтобы проиллюстрировать это интересное поведение, рассмотрим разряды клеток поджелудочной железы. Эти клетки связаны с секрецией инсулина. Разрядную активность клеток можно проследить с помощью внутриклеточных электродов. Lebrun и Atwater нашли, что разрядная активность может быть нерегулярной (рис. 4.3). Были предприняты попытки разработать математические модели типа уравнений Ходжкина — Хаксли для Р-клеток поджелудочной железы. Chay и Rinzel показали путем численного решения этих уравнений, что в некотором диапазоне величин параметров регулярные периодические разряды не обнаруживались: вместо них наблюдалась нерегулярная, апериодическая динамика (рис. 4.4). Поскольку она возникала в детерминированной математической модели, было сделано заключение, что эта модель обнаруживает хаотическое поведение. (см. скан) Рис. 4.2. Вычисленные потенциалы действия для волокна Пуркинье. (а) Потенциал действия на верхнем рисунке был инициирован деполяризацией до
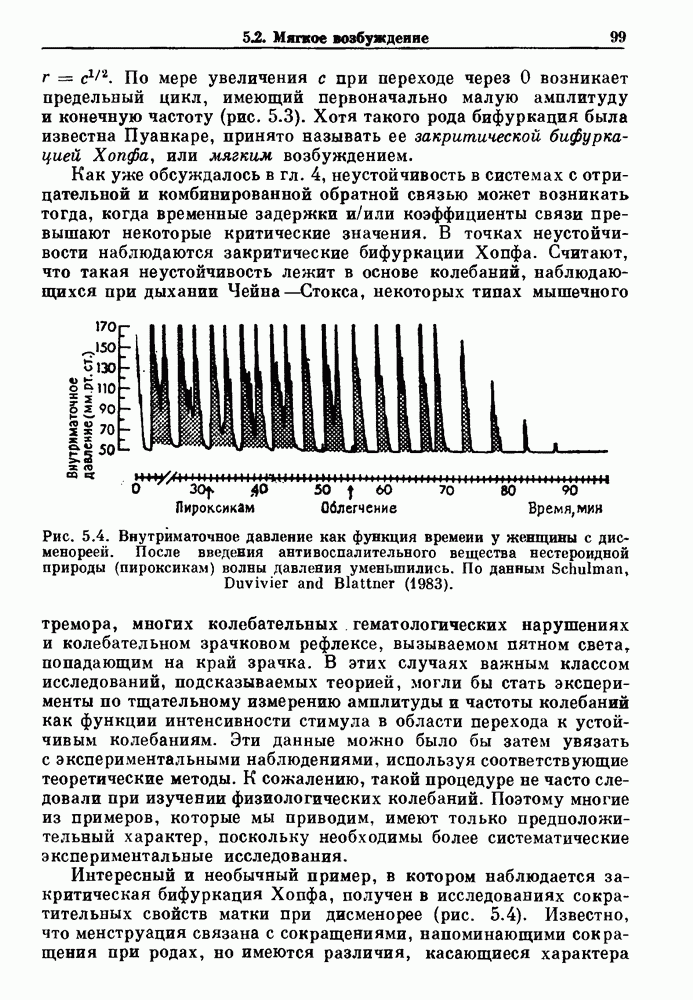
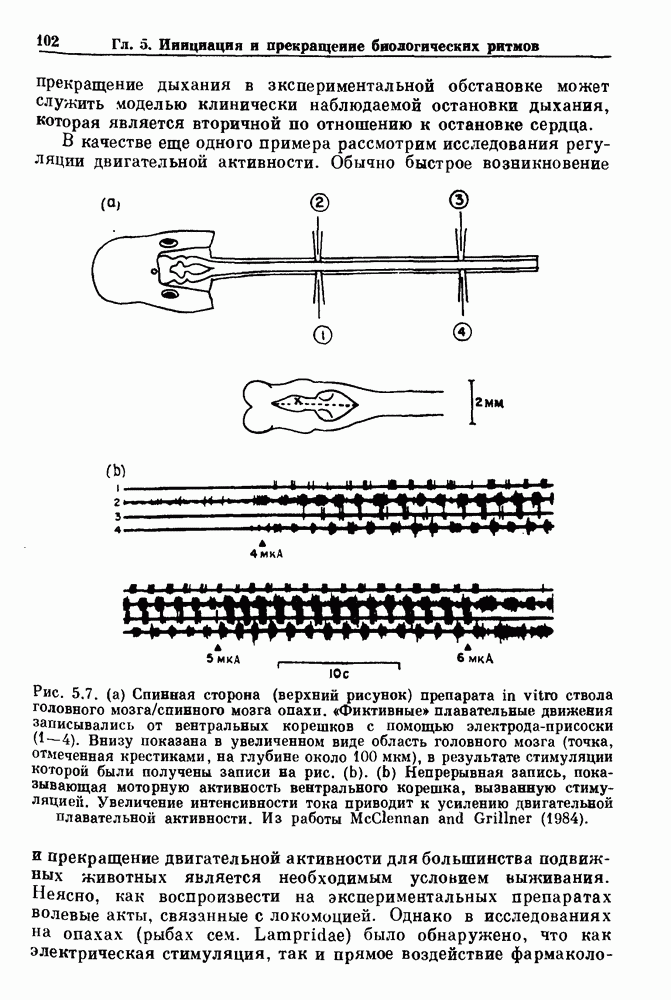
Рис. 4.3. Ритмическая активность островков Лангерганса у четырех различных линий мышей через 30 мин после воздействия (см. скан) Рис. 4.4. Различные периодические и апериодические решения модели Chay — Keizer и их зависимость от скорости притока внутриклеточного кальция Эти наблюдения поднимают важные и увлекательные вопросы. Нелинейные дифференциальные уравнения, предложенные для описания пейсмекерных колебаний в различных тканях, в действительности могут иметь хаотическое решение при изменениях параметров, связанных с изменениями в окружающей среде. Действительно, в сборнике статей под редакцией Chalaontis и Boisson, появившегося в 1978 г., было показано, что фармакологические воздействия на пейсмекерные клетки приводят к возникновению сложных ритмов. Таким образом, сложные хаотические ритмы, генерируемые пейсмекерной тканью, могут, по-видимому, оказаться более распространенными, чем принято считать. Существующие в настоящее время математические модели пейсмекерных колебаний, основанные на реальных ионных механизмах, настолько сложны, что единственный способ определения динамических свойств модели — это использование численного интегрирования. Кроме того, математические модели позволяли получить великолепную реконструкцию формы потенциала действия даже в тех случаях, когда моделируемые ионные механизмы, как впоследствии выяснилось, оказывались ошибочными. Таким образом, несмотря на то, что определение ионной основы пейсмекерных колебаний представляет большой научный интерес, математическое моделирование этих процессов является трудной задачей, потому что результирующие модели слишком сложны и возможны различные интерпретации одних и тех же данных.
|
 |
Оглавление
|





















































































































































































 — проводимости мембраны для натрия, калия и ионов утечки соответственно,
— проводимости мембраны для натрия, калия и ионов утечки соответственно,  — соответствующие равновесные потенциалы. Сложность задачи определяется сильно нелинейной зависимостью проводимостей от времени и трансмембранного потенциала.
— соответствующие равновесные потенциалы. Сложность задачи определяется сильно нелинейной зависимостью проводимостей от времени и трансмембранного потенциала.
 можно регистрировать отдельные ионные токи, используя различные фармакологические вещества. Эти авторы показали, что при поддерживаемом уровне деполяризации (положительное отклонение мембранного потенциала от потенциала покоя мембраны) гигантского аксона кальмара существует устойчивый подъем в проводимости калия и
можно регистрировать отдельные ионные токи, используя различные фармакологические вещества. Эти авторы показали, что при поддерживаемом уровне деполяризации (положительное отклонение мембранного потенциала от потенциала покоя мембраны) гигантского аксона кальмара существует устойчивый подъем в проводимости калия и

 Средний рисунок показывает изменение во времени переменных
Средний рисунок показывает изменение во времени переменных  и s типа Ходжкина — Хаксли, которые регулируют токи
и s типа Ходжкина — Хаксли, которые регулируют токи  направленные наружу (нижний рисунок). Деполяризация пейсмекера обусловлена уменьшением тока
направленные наружу (нижний рисунок). Деполяризация пейсмекера обусловлена уменьшением тока  Величины параметров соответствуют
Величины параметров соответствуют  По данным McAllister, Noble and Tsien (1975). (b) Реконструкция пейсме-керной активности волокна Пуркинье с помощью уравнений DiFrancesco - Noble (1984). Вычисленный потенциал действия показан вверху вместе с вычисленными изменениями в воротных механизмах, контролирующих
По данным McAllister, Noble and Tsien (1975). (b) Реконструкция пейсме-керной активности волокна Пуркинье с помощью уравнений DiFrancesco - Noble (1984). Вычисленный потенциал действия показан вверху вместе с вычисленными изменениями в воротных механизмах, контролирующих  Отметим, что переменная х сравнима с переменной, показанной на рис. (а). Однако увеличение переменной у (контролирующей ток натрия, направленный внутрь) совершенно не соответствует уменьшению переменной s (контролирующей ток калия наружу) на рис. (а). Величины параметров соответствуют значению
Отметим, что переменная х сравнима с переменной, показанной на рис. (а). Однако увеличение переменной у (контролирующей ток натрия, направленный внутрь) совершенно не соответствует уменьшению переменной s (контролирующей ток калия наружу) на рис. (а). Величины параметров соответствуют значению  По данным Noble (1984).
По данным Noble (1984).

 глюкозы: (а) — островок из мыши линии Национального института здоровья;
глюкозы: (а) — островок из мыши линии Национального института здоровья;  островки из мыши линии Charles River; (d) - островок из мыши линии Charles Hiver после одного месяца содержания на диете мышей линии Национального института здоровья. По данным Lebrun and Atwater (1985).
островки из мыши линии Charles River; (d) - островок из мыши линии Charles Hiver после одного месяца содержания на диете мышей линии Национального института здоровья. По данным Lebrun and Atwater (1985).
 определяемой уровнем глюкозы, (а) Периодические разряды при
определяемой уровнем глюкозы, (а) Периодические разряды при  ) Хаотические разряды при
) Хаотические разряды при  Хаотические биения при
Хаотические биения при  Периодические биения при
Периодические биения при  Отметим; изменение масштаба времени в (b). Адаптировано из работы Cbay and Rinzel (1985).
Отметим; изменение масштаба времени в (b). Адаптировано из работы Cbay and Rinzel (1985).