Пред.
След.
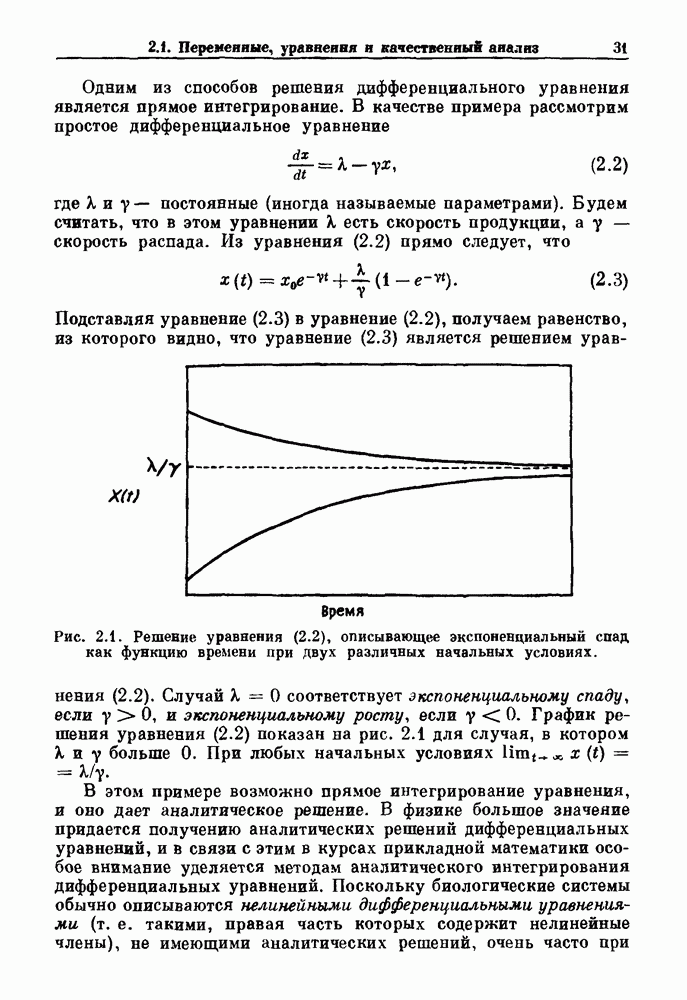
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
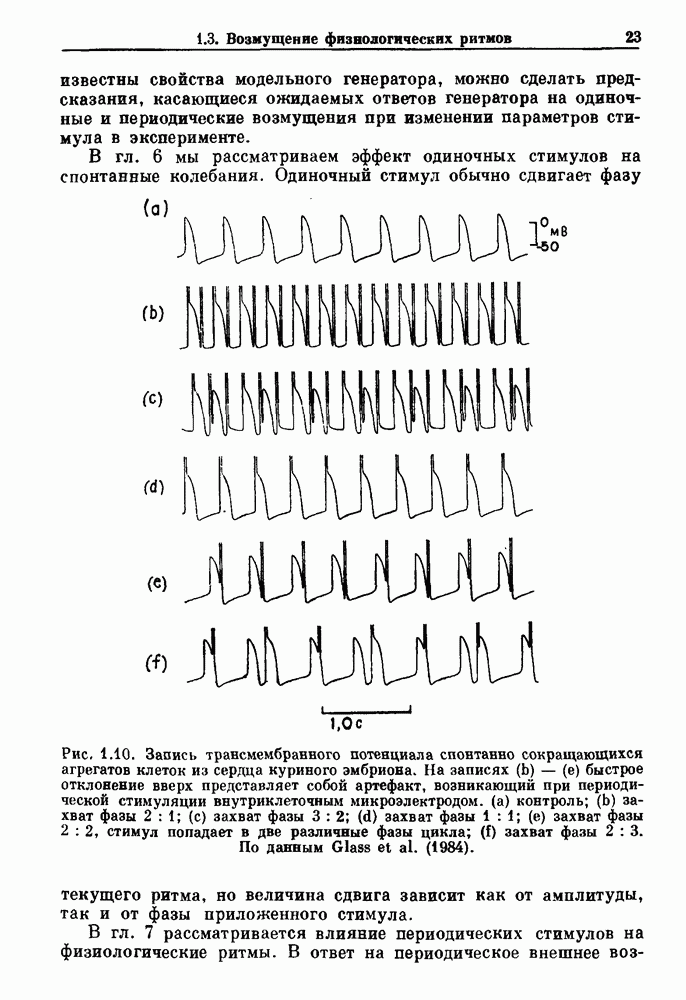
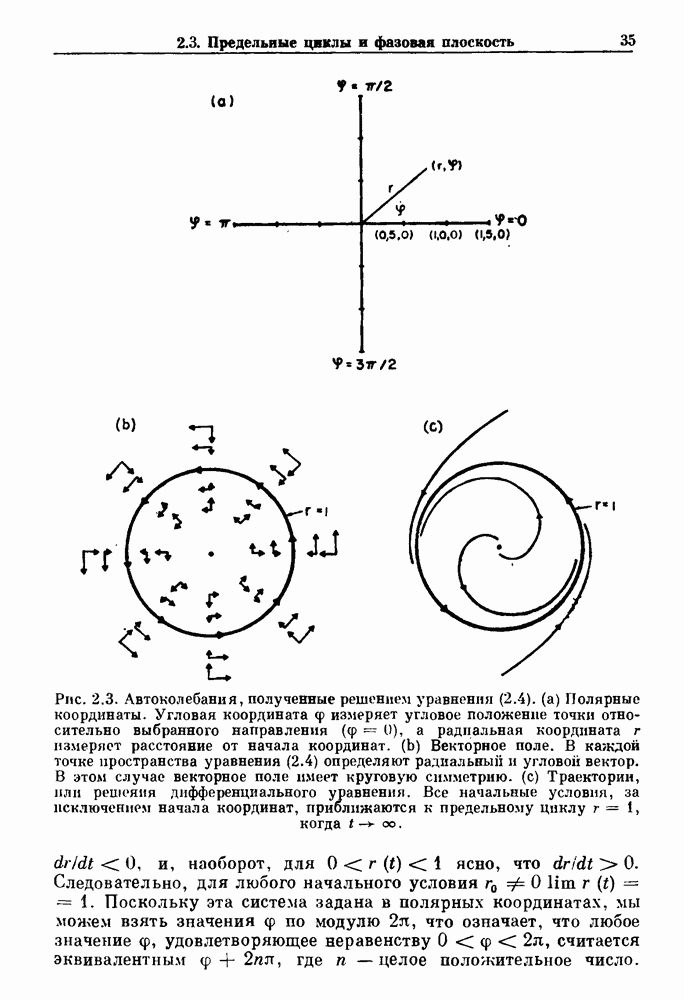
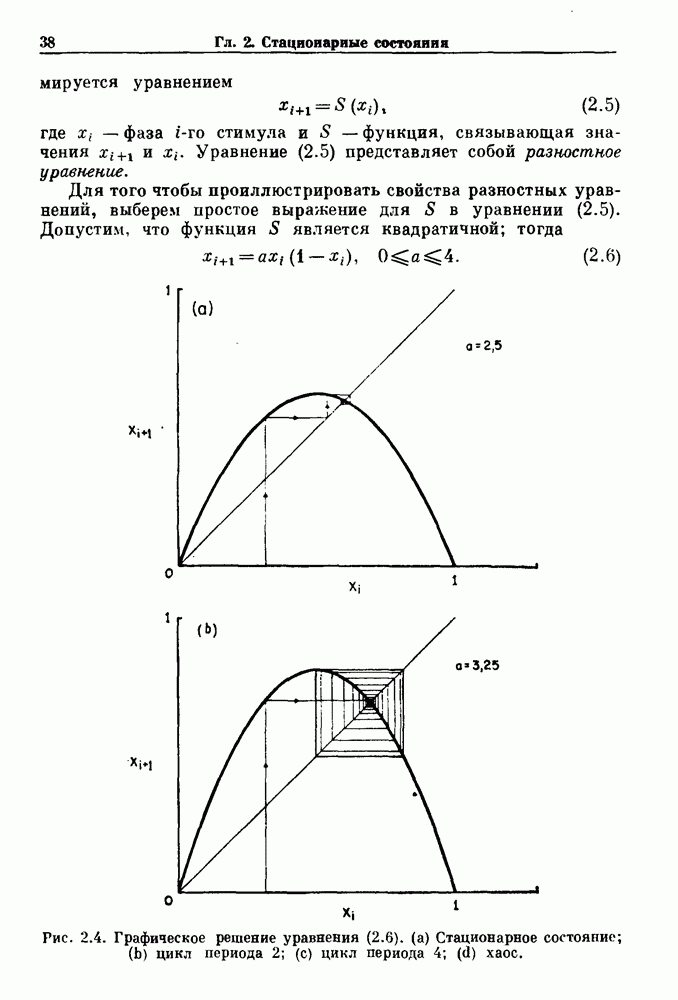
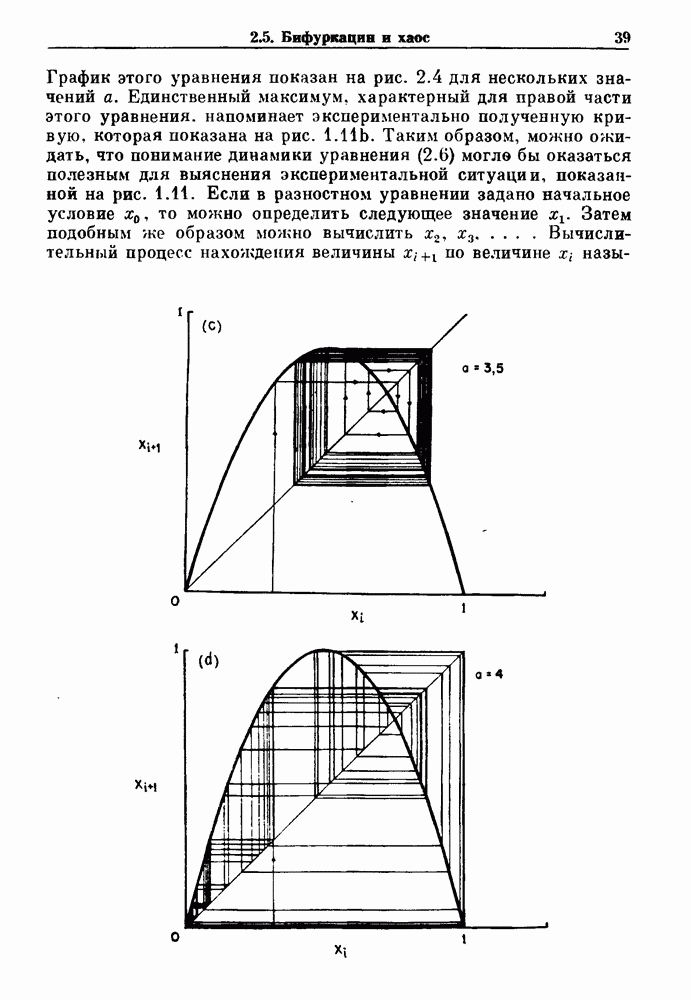
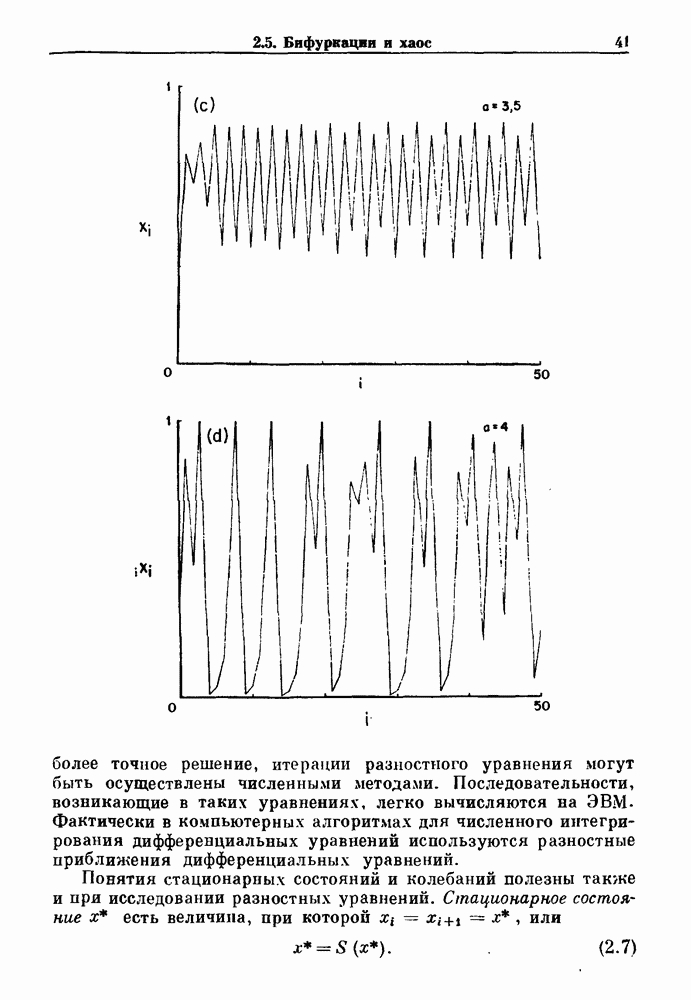
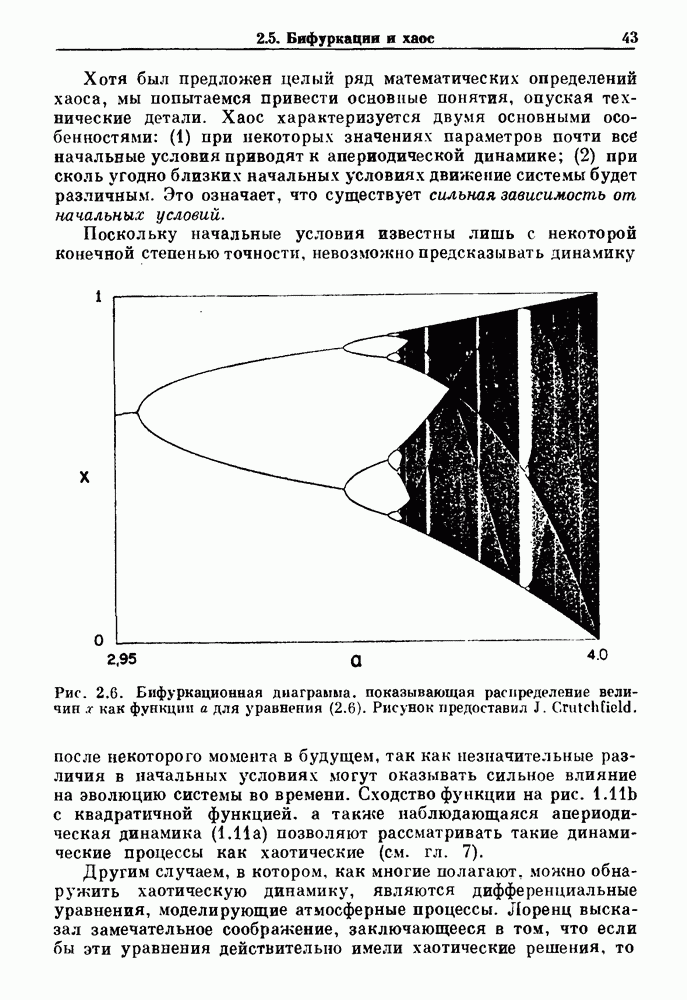
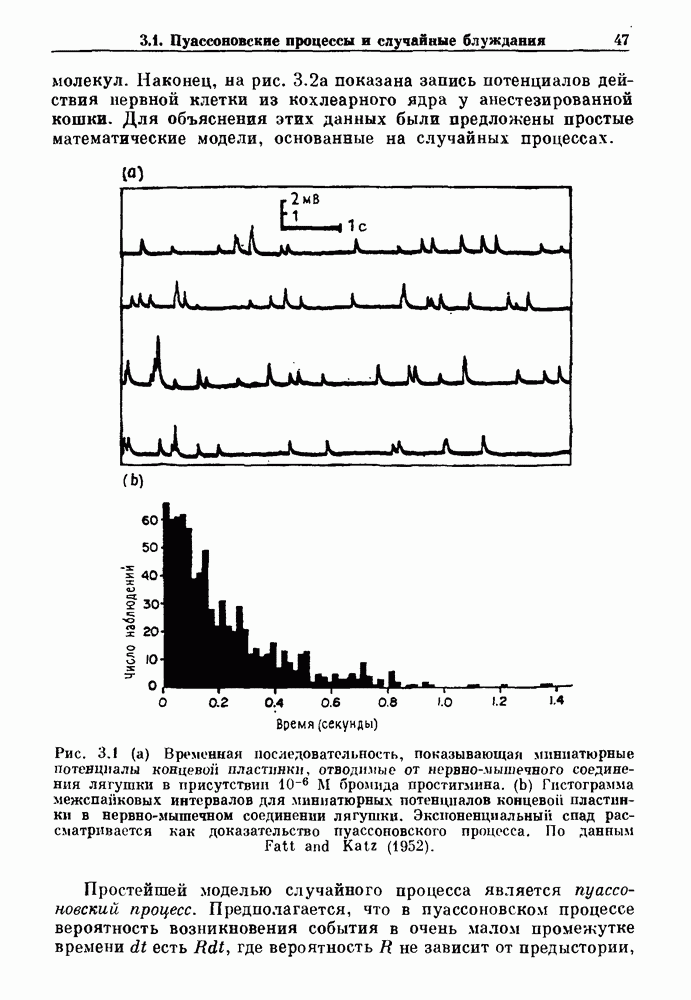
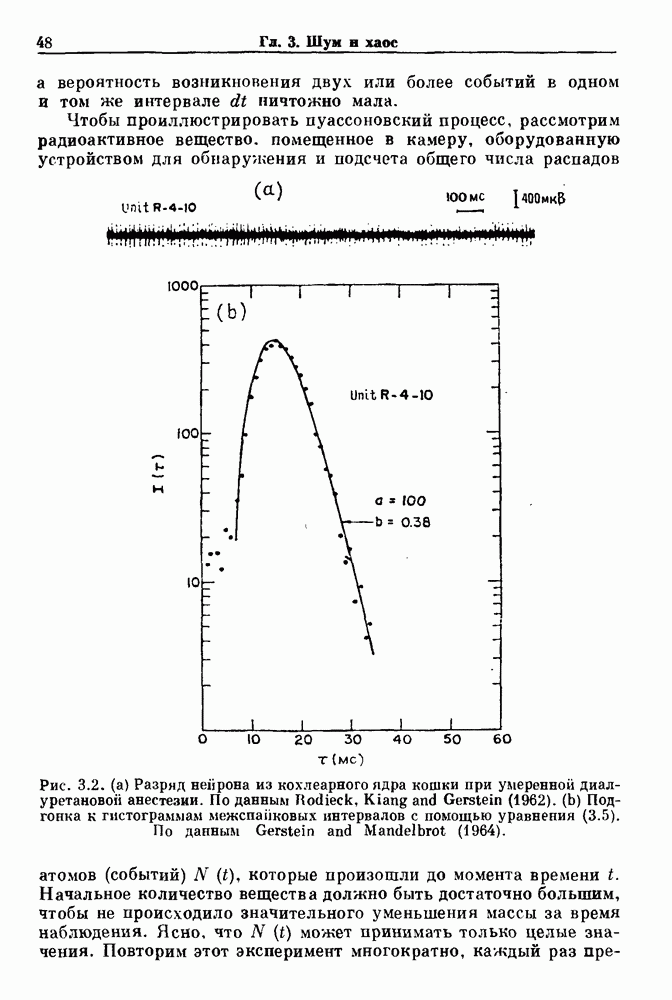
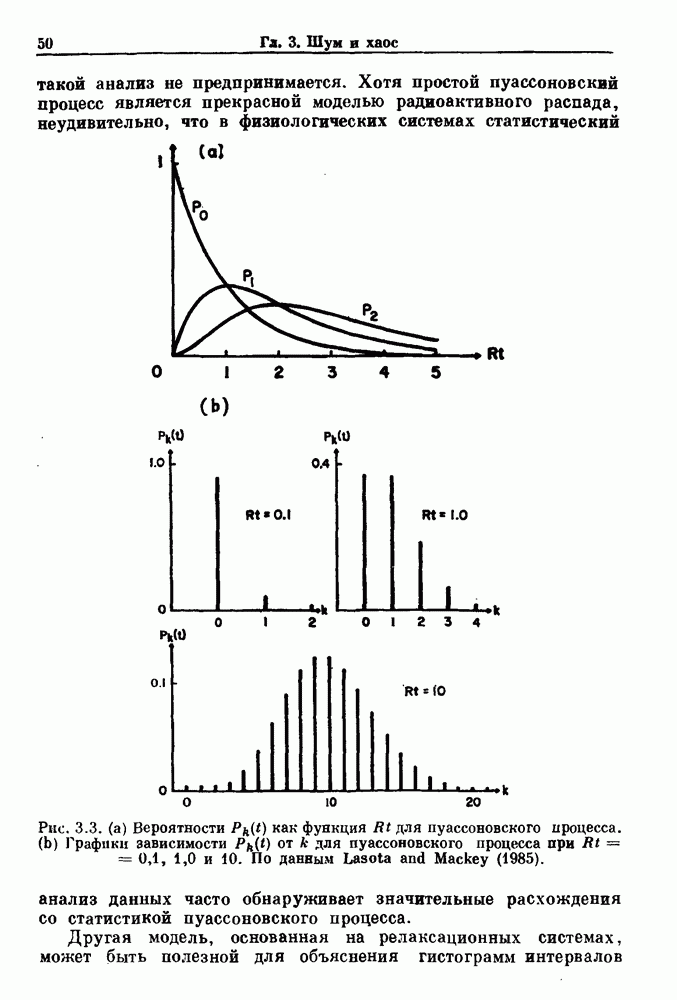
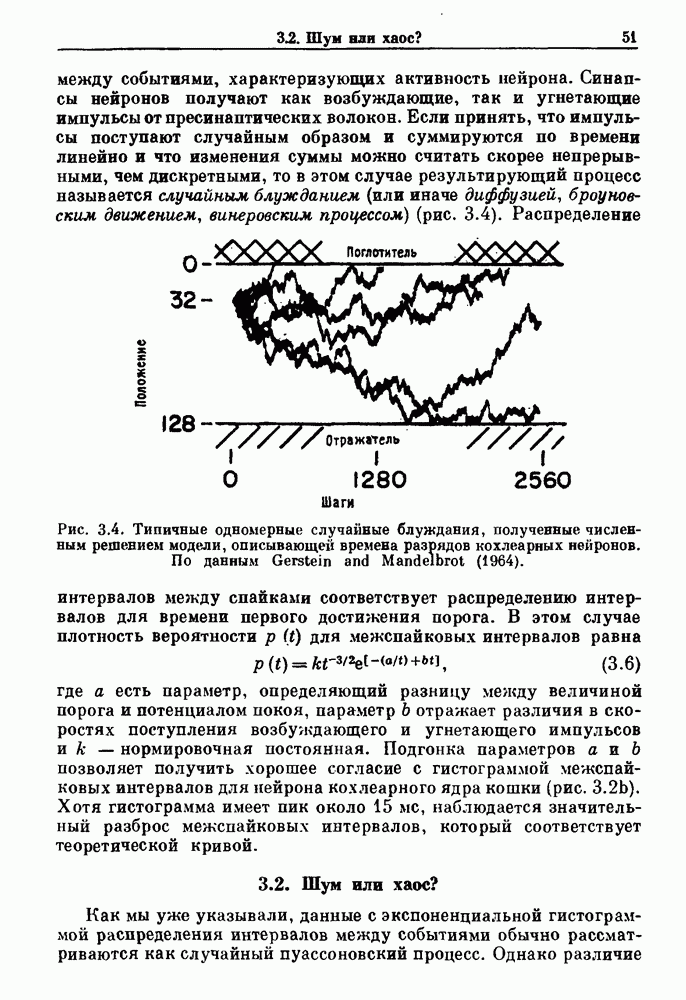
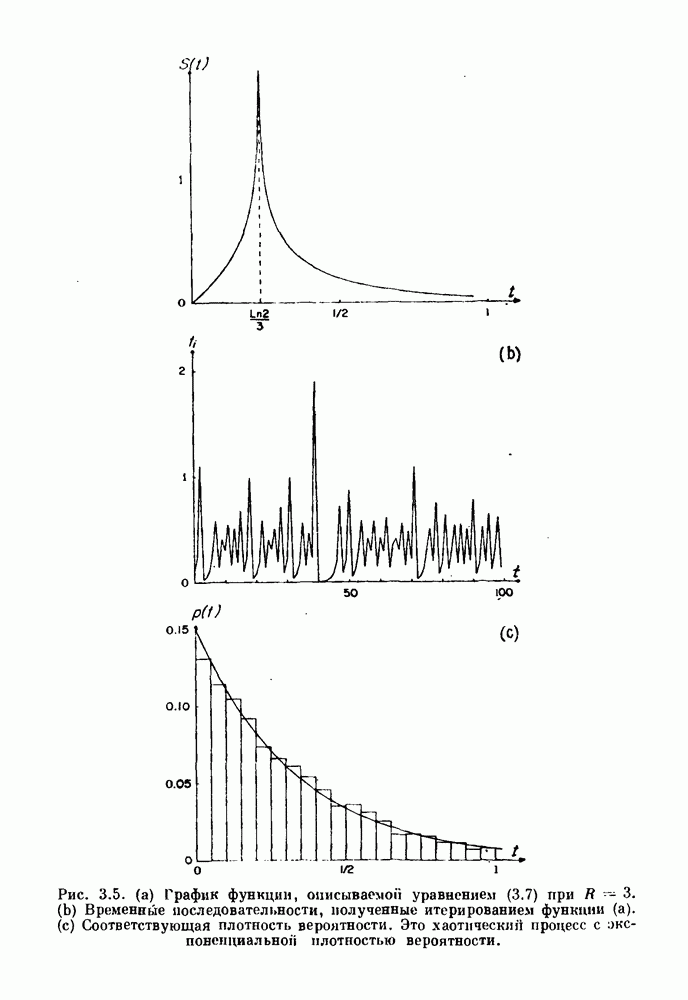
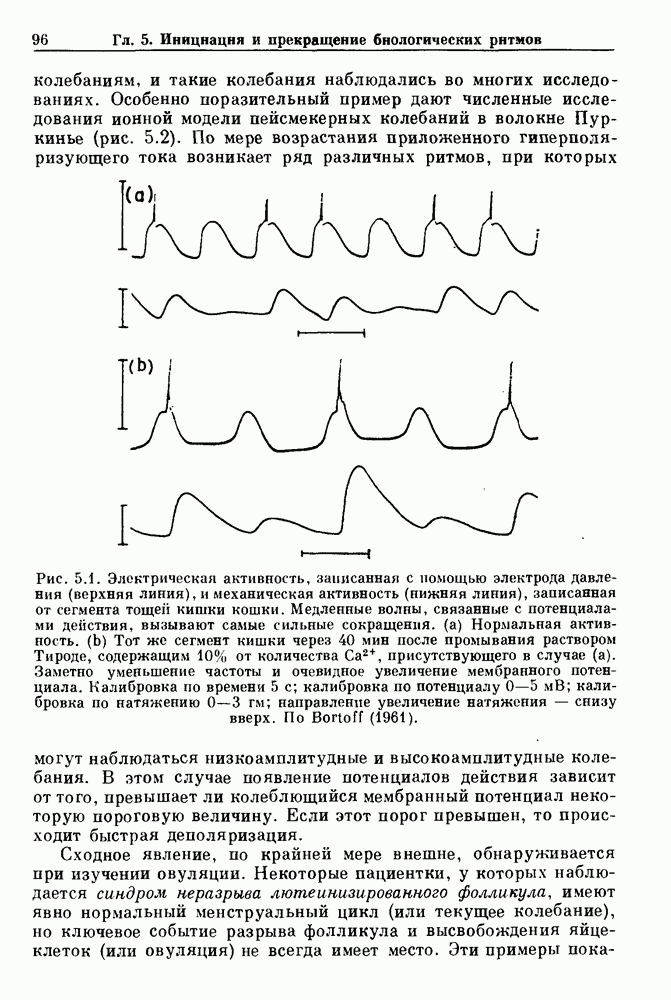
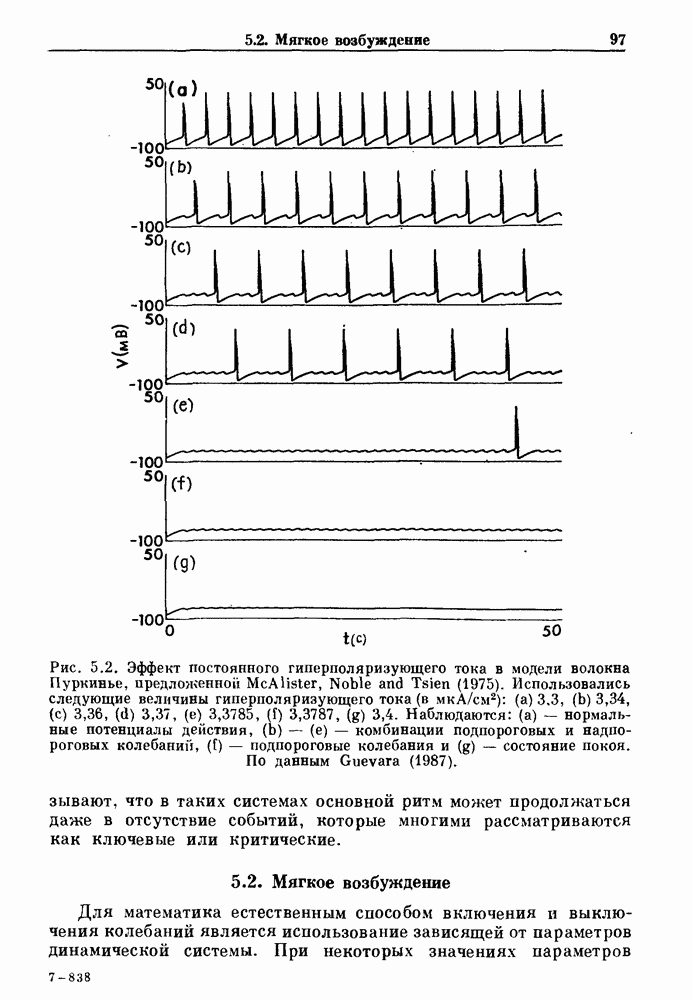
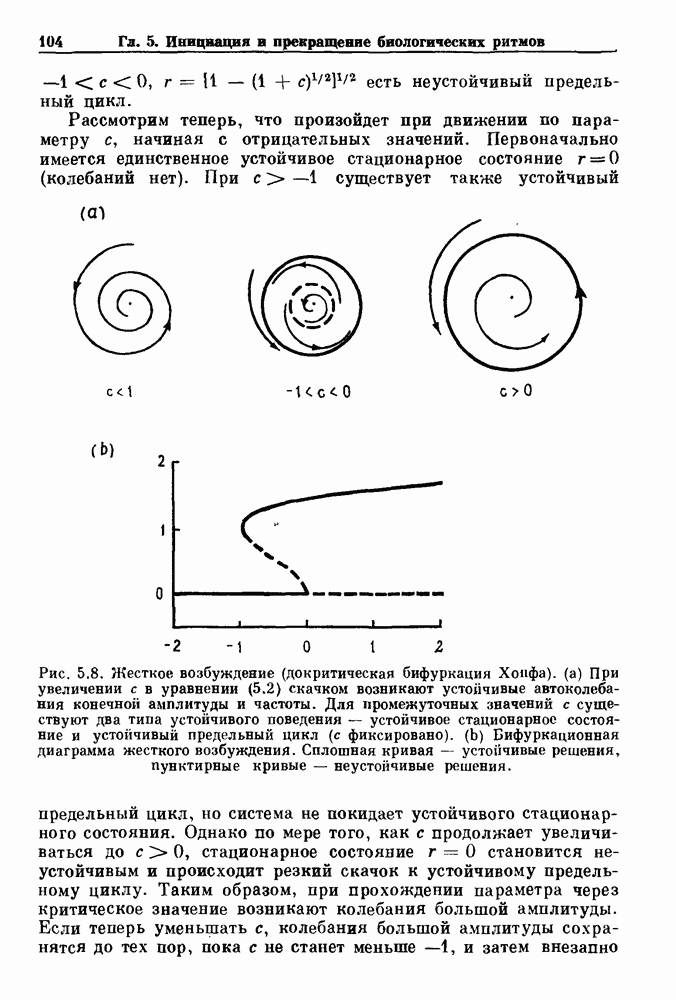
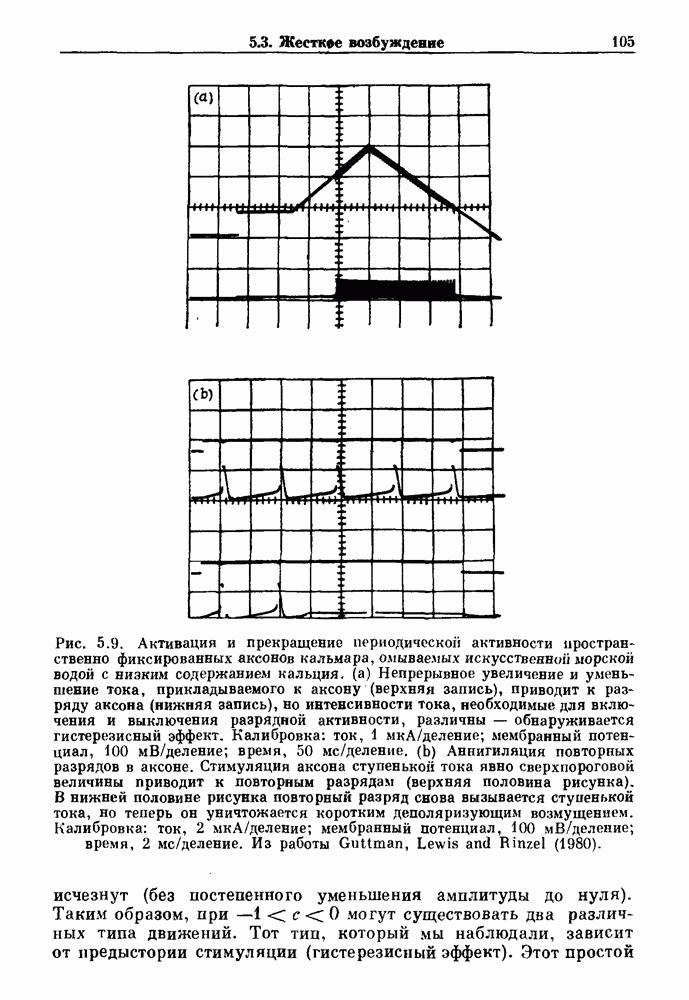
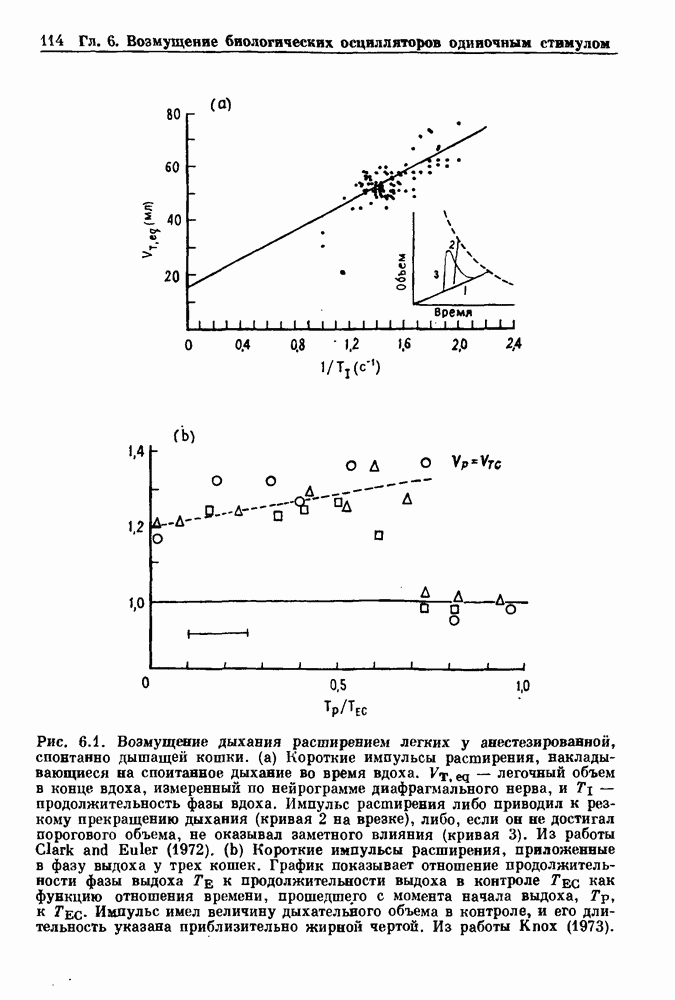
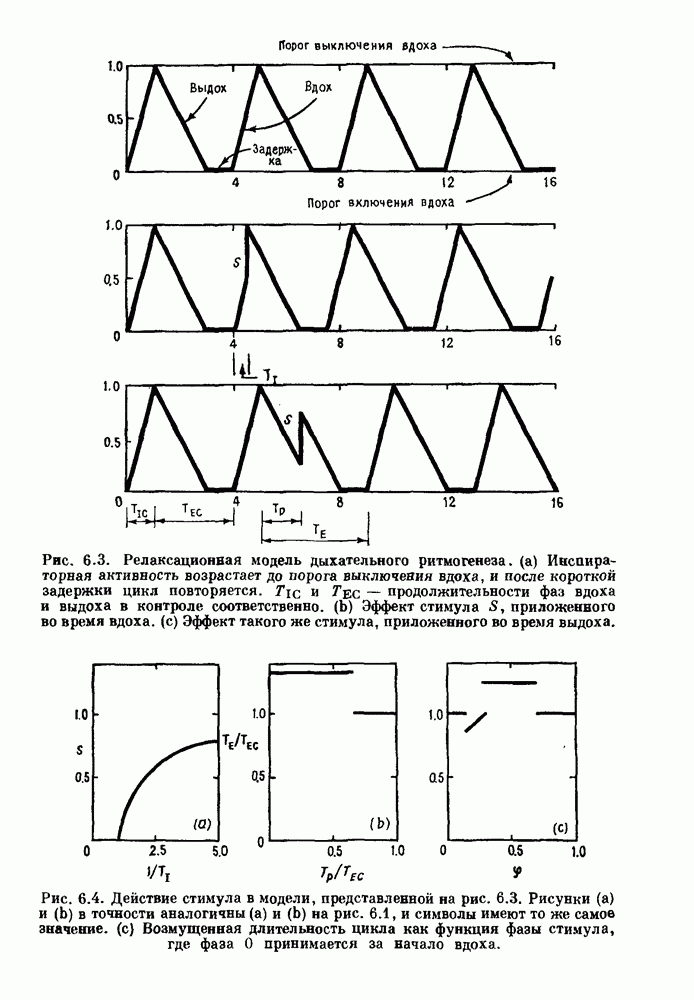
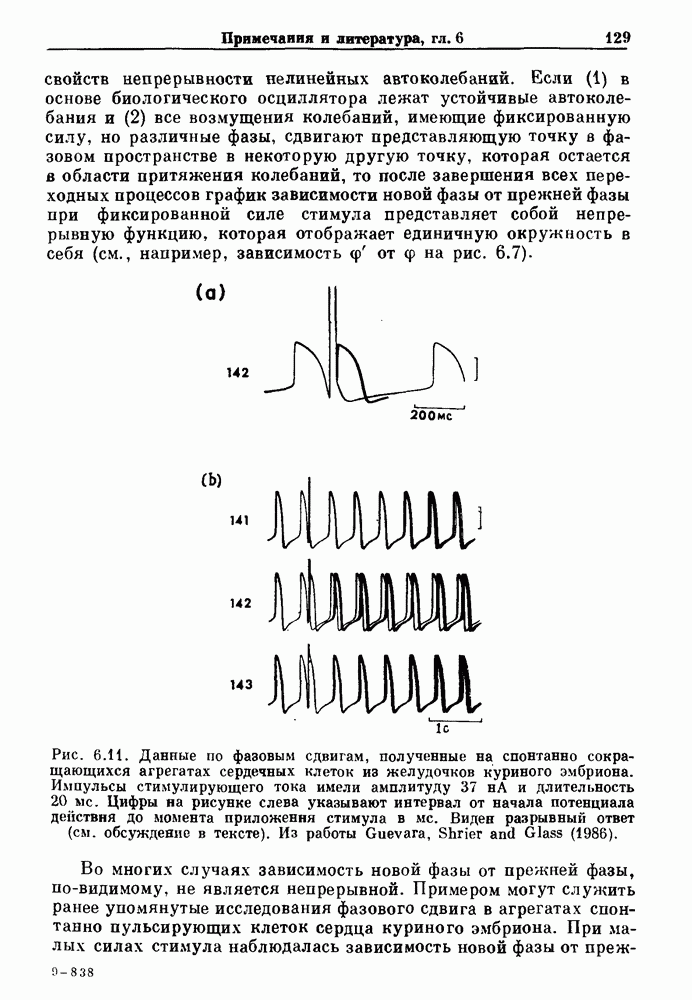
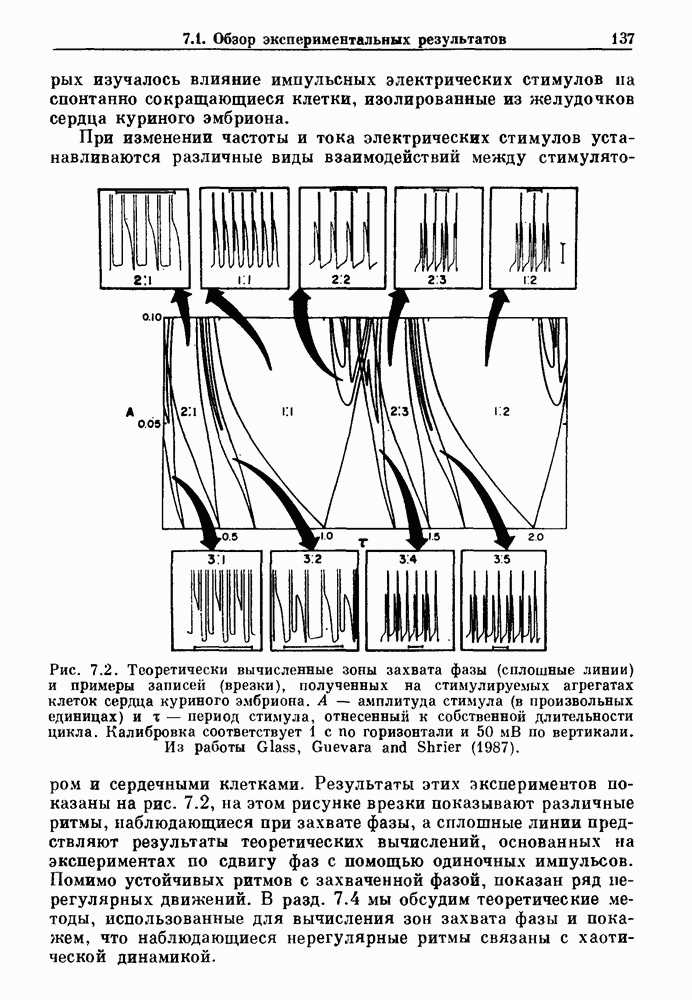
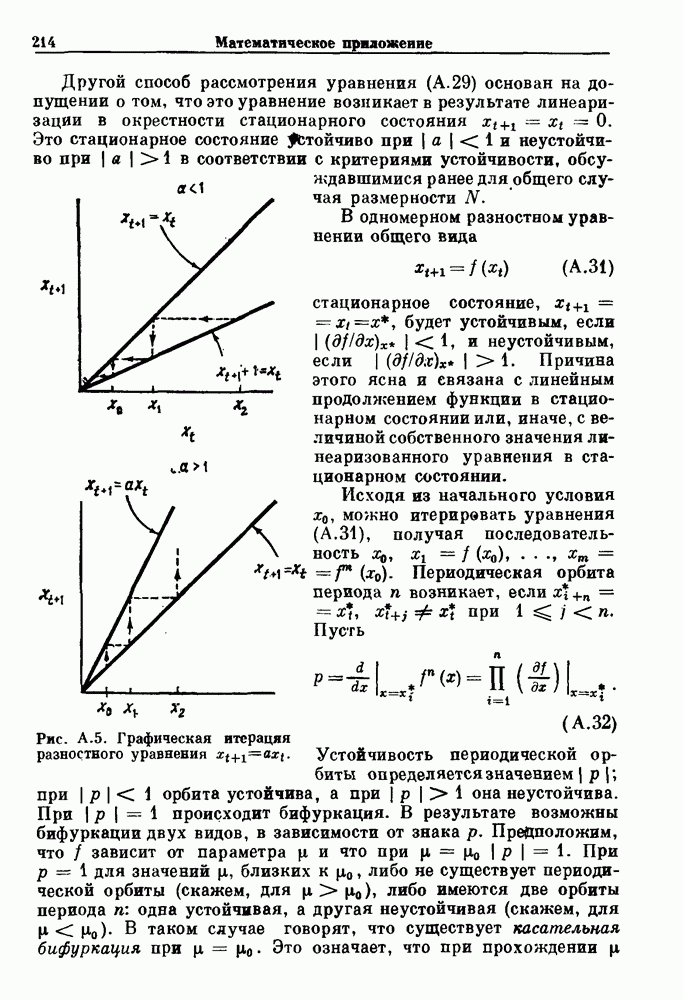
3.2. Шум или хаос?Как мы уже указывали, данные с экспоненциальной гистограммой распределения интервалов между событиями обычно рассматриваются как случайный пуассоновский процесс. Однако различие между шумом и хаосом не всегда очевидно, а иногда его просто невозможно установить. Действительно, практически для любой одномерной плотности вероятности можно построить бесконечное число (детерминированных) разностных уравнений, решения которых хаотичны и имеют данную плотность. Это означает, что на основании одной только плотности распределения нельзя судить о динамике, лежащей в основе рассматриваемых временных последовательностей. В качестве простого примера рассмотрим разностное уравнение, специально сконструированное для получения экспоненциальной плотности вероятности. Можно строго показать, что последовательные решения разностного уравнения (см. рис. 3.5 а)
имеют постоянную плотность
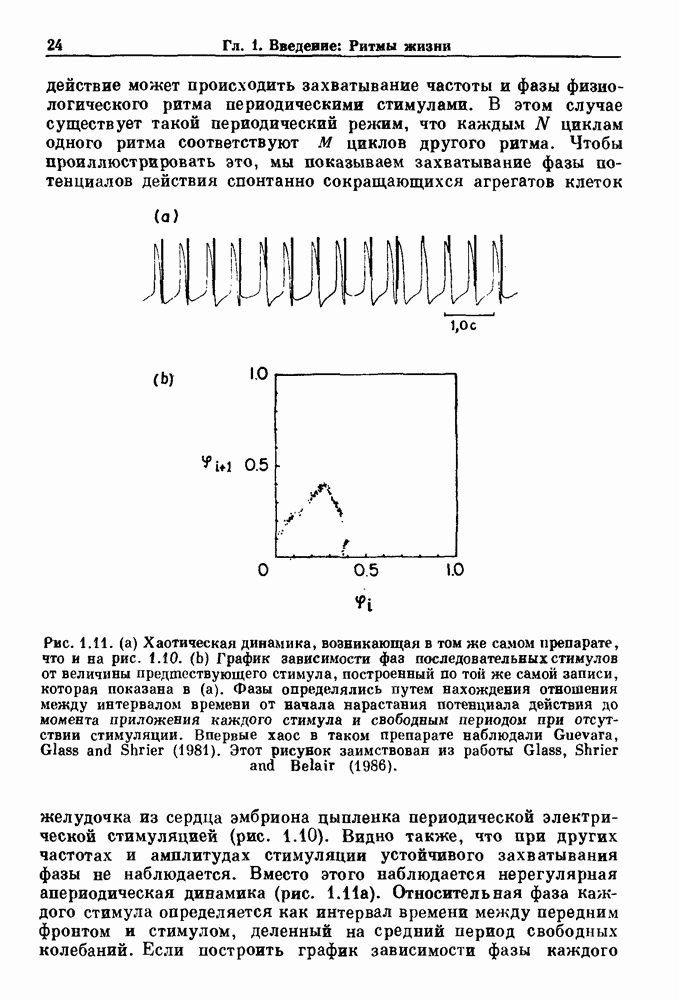
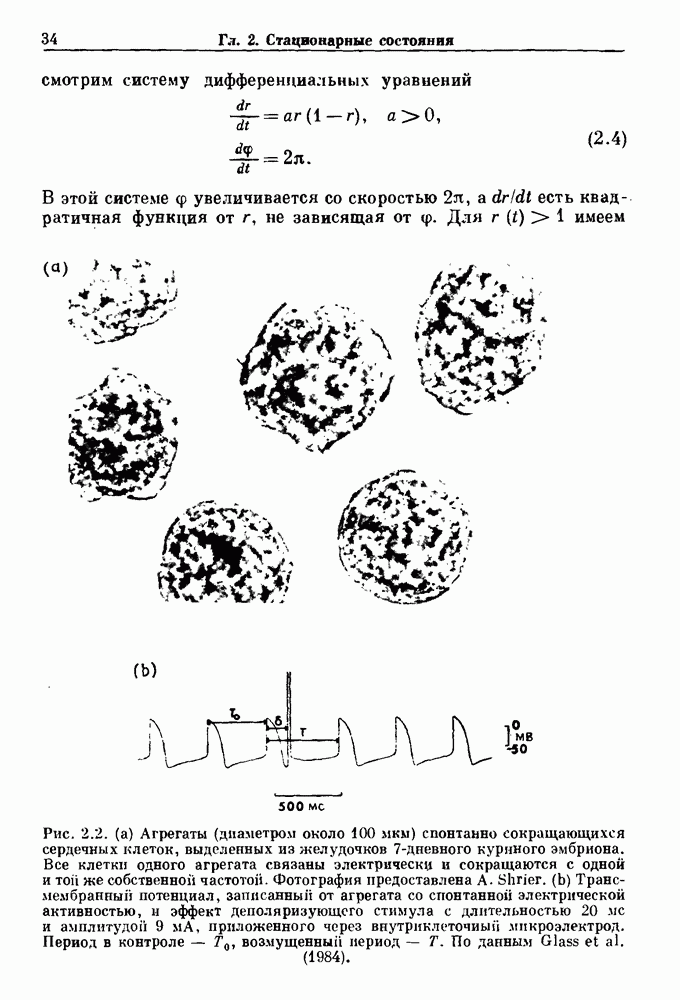
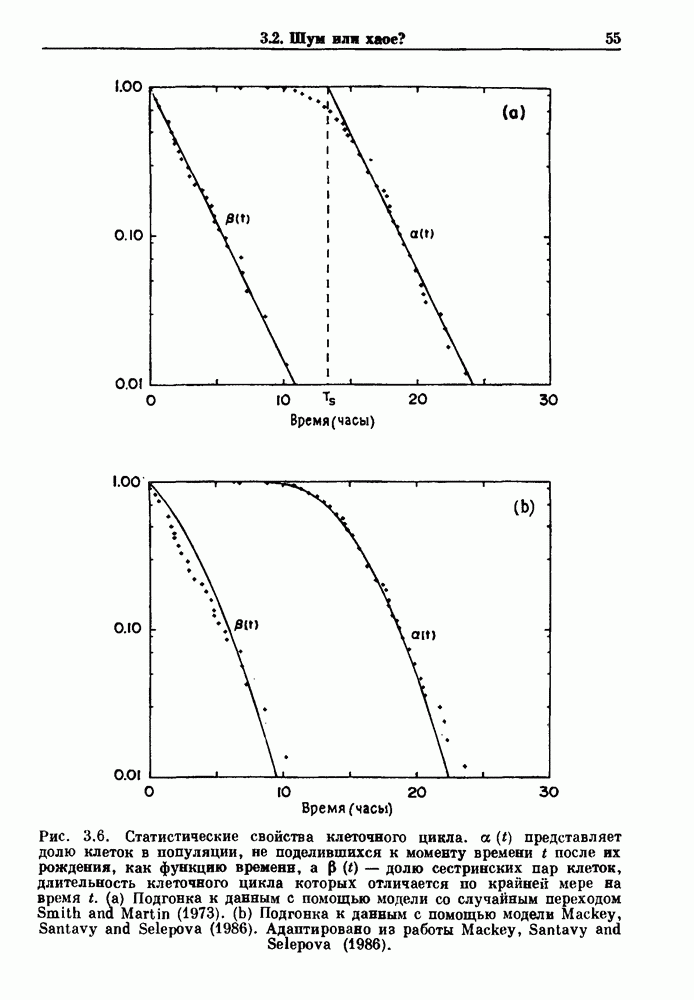
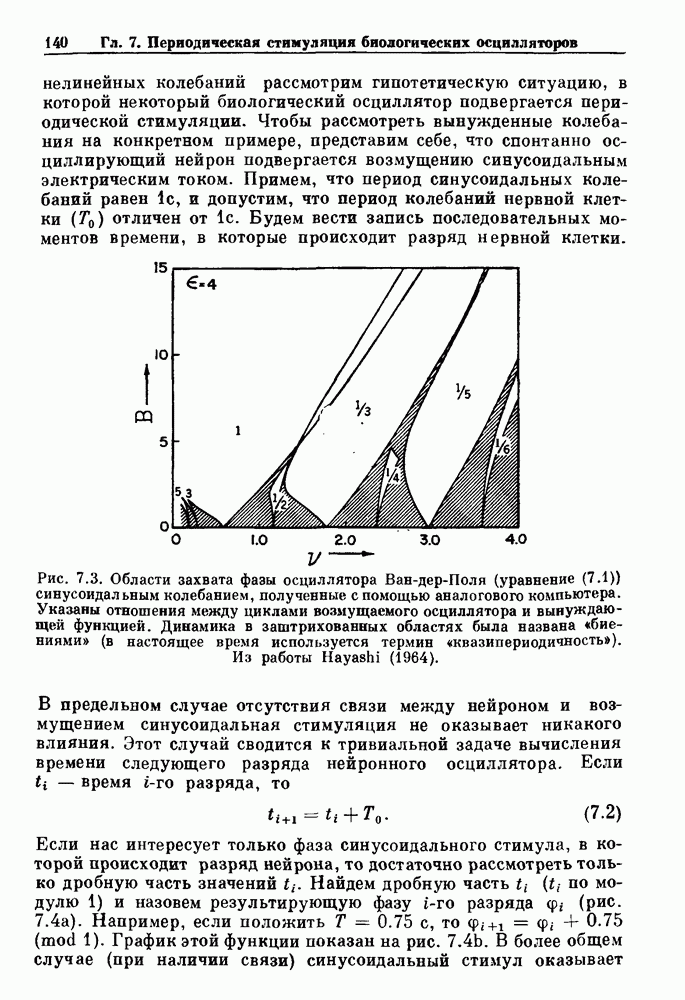
совпадающую с плотностью вероятности пуассоновского процесса (3.5) для Два примера из биологии иллюстрируют проблемы, связанные с различением шума и хаоса в случае последовательностей, имеющих неэкспоненциальные плотности. Один из них касается длительности клеточного цикла, а другой — выживания больных лейкемией. Данные, полученные на культивируемых клетках, позволили предположить, что спустя некоторое время после рождения в каждой клетке происходит случайное событие, необходимое для того, чтобы начался митоз. В этой модели клеточного цикла со случайным переходом принимается, что, раз случайное событие произошло, клеточное деление будет иметь место некоторое время (см. скан) Рис. 3.5. (а) График функции, описываемой уравнением (3.7) при большой популяции, поделилась в момент времени t после рождения, т. е.
Таким образом, доля клеток в популяции
откуда мы вычисляем
что довольно хорошо согласуется с полученными данными (см. рис. 3.6 а), за исключением узкого интервала времени около
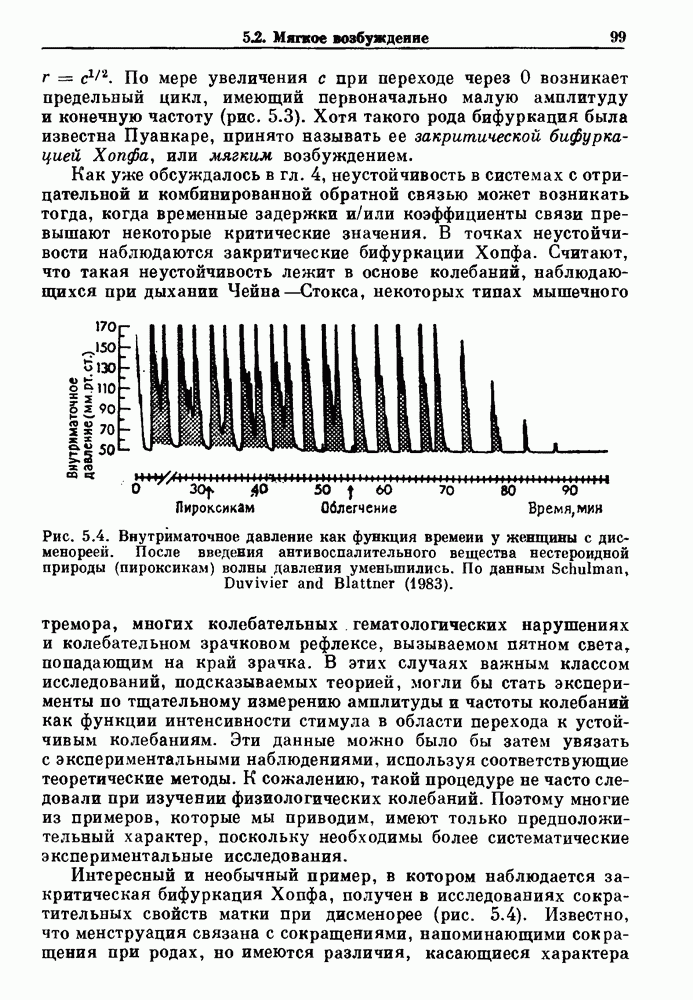
что также соответствует данным на рис. 3.6 а. Альтернативная гипотеза предполагает существование внутриклеточного генератора хаотичных (но детерминированных) митотических ритмов, который организует клеточный цикл во времени. С помощью простой специальной модели, основанной на этой гипотезе и содержащей только два параметра (напомним, что модель со случайным переходом также содержит два параметра, R и В качестве второго примера, в котором данные одинаково хорошо интерпретируются как в рамках случайного, так и детерминированного механизма, рассмотрим статистику выживания больных с хронической миелогенной лейкемией (ХМЛ) после установления диагноза (рис. 3.7). Такие статистические данные обычно описываются экспоненциальной функцией типа уравнения (3.3) или линейной суммой экспонент, даже если имеется горб или плечо в области малых времен выживания после постановки диагноза. Эти экспоненциальные подгонки к данным по выживанию рассматриваются затем как указание на то, что больные умирают (см. скан) Рис. 3.6. Статистические свойства клеточного цикла, а случайно с постоянной вероятностью в единицу времени после установления диагноза. В качестве альтернативы предложена модель продуцирования белых кровяных клеток в виде простого разностного уравнения. Эта модель основана на известном механизме регулирования продукции белых кровяных клеток с помощью обратной связи, а также на допущении о том, что XMЛ характеризуется медленным, но неуклонным ростом максимальной скорости продукции белых кровяных клеток.
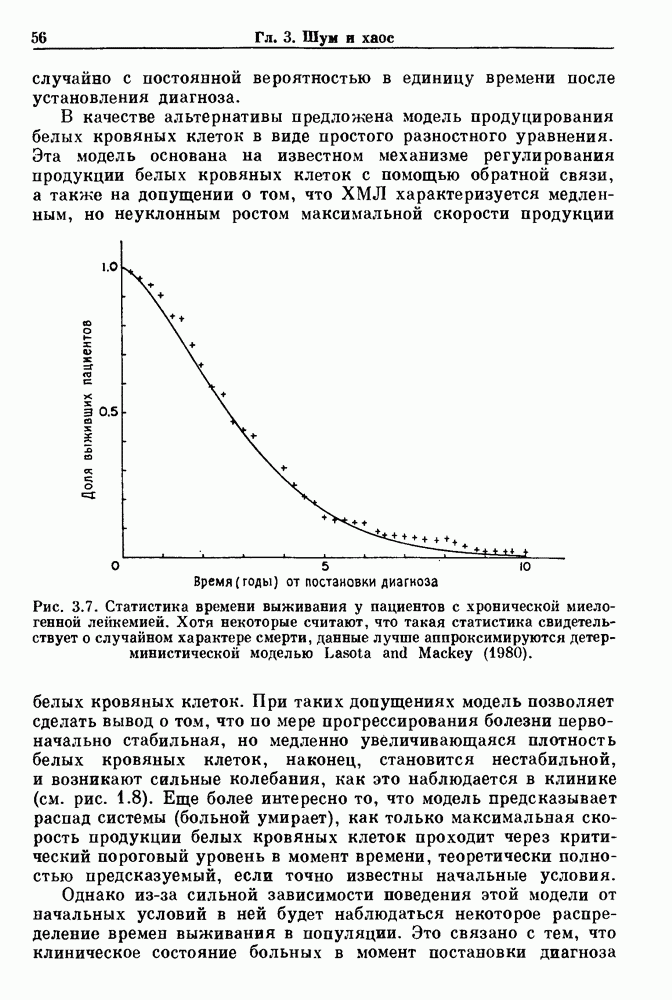
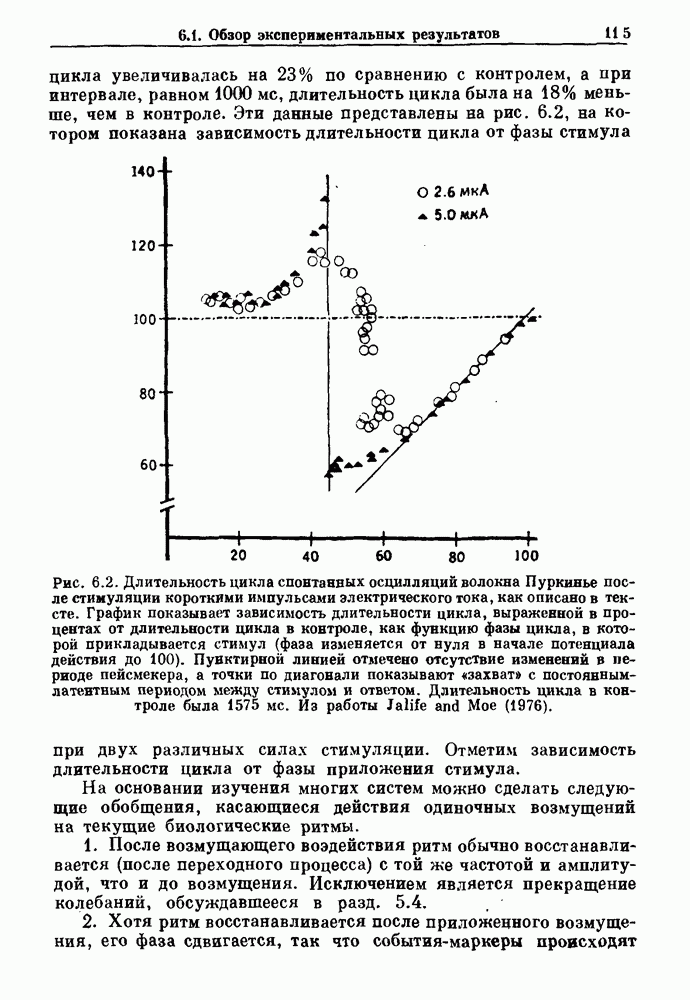
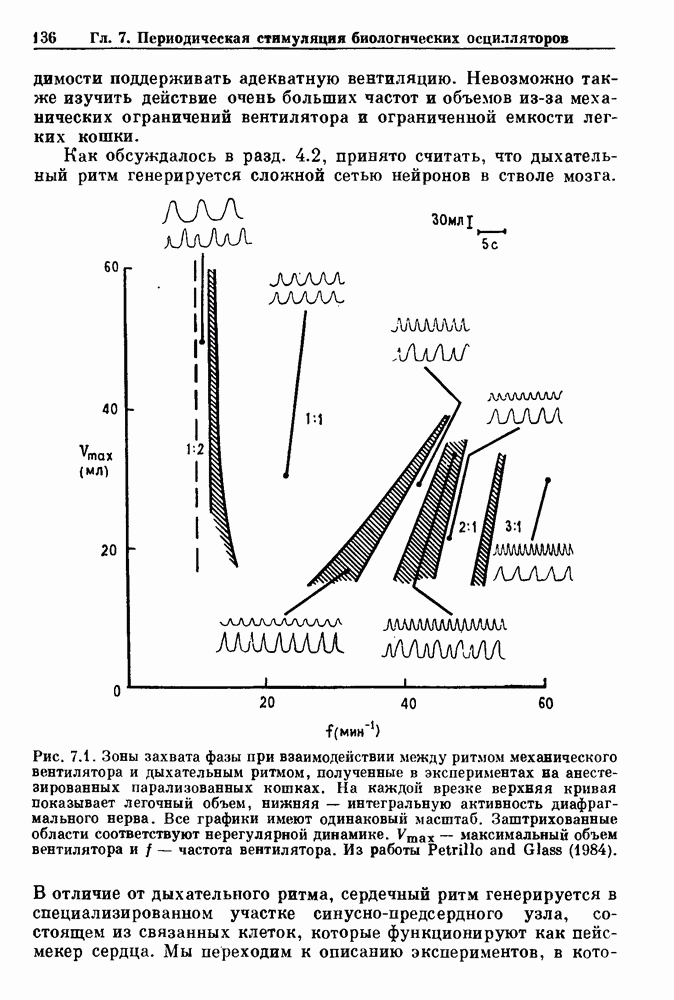
Рис. 3.7. Статистика времени выживания у пациентов с хронической миелогенной лейкемией. Хотя некоторые считают, что такая статистика свидетельствует о случайном характере смерти, данные лучше аппроксимируются детерминистической моделью Lasota and Mackey (1980). При таких допущениях модель позволяет сделать вывод о том, что по мере прогрессирования болезни первоначально стабильная, но медленно увеличивающаяся плотность белых кровяных клеток, наконец, становится нестабильной, и возникают сильные колебания, как это наблюдается в клинике (см. рис. 1.8). Еще более интересно то, что модель предсказывает распад системы (больной умирает), как только максимальная скорость продукции белых кровяных клеток проходит через критический пороговый уровень в момент времени, теоретически полностью предсказуемый, если точно известны начальные условия. Однако из-за сильной зависимости поведения этой модели от начальных условий в ней будет наблюдаться некоторое распределение времен выживания в популяции. Это связано с тем, что клиническое состояние больных в момент постановки диагноза неодинаково. Модель предсказывает, что часть популяции больных с XMЛ, переживающих время t после установления диагноза, равна
Более того, в больших популяциях доля выживающих больных не зависит от распределения начальных состояний. Это предсказание имеет несколько интересных особенностей. Во-первых, оно основывается на полностью детерминированной модели динамики образования белых кровяных клеток. Во-вторых, оно дает гораздо более точное описание имеющихся статистических данных по выживанию, чем допущение о чисто случайной смерти при постоянной вероятности в единицу времени. Однако наиболее интересный аспект связан с тем фактом, что кривые выживания больных с ХМЛ относительно мало меняются при использовании различных терапевтических средств, включая хемо- и/или радиотерапию. Так как эти цитотоксические средства вызывают возвращение к исходному уровню численности пролиферирующих клеточных популяций, отсутствие их влияния на статистику выживания популяции получает простую интерпретацию в контексте этой детерминированной модели. А именно, статистические свойства времен выживания популяции нечувствительны к распределению начальных состояний, несмотря на тот факт, что использование этих цитотоксических агентов с целью повлиять на начальные условия может оказывать сильное воздействие на время выживания отдельного больного. В частности, применение различных видов терапии к одному и тому же больному может резко сократить или продлить его жизнь. В заключение отметим, что при наличии некоторого динамического процесса — будь то детерминированный хаос или стохастический процесс, подобный пуассоновскому процессу или случайному блужданию — часто оказывается возможным вычисление соответствующей плотности вероятности. Однако обратная операция нахождения динамического процесса, приводящего к данной плотности вероятности, не имеет однозначного решения. Таким образом, экспоненциальная плотность вероятности для интервалов между событиями не может служить критерием пуассоновского процесса без тщательного рассмотрения других статистических свойств динамической системы.
|
 |
Оглавление
|




































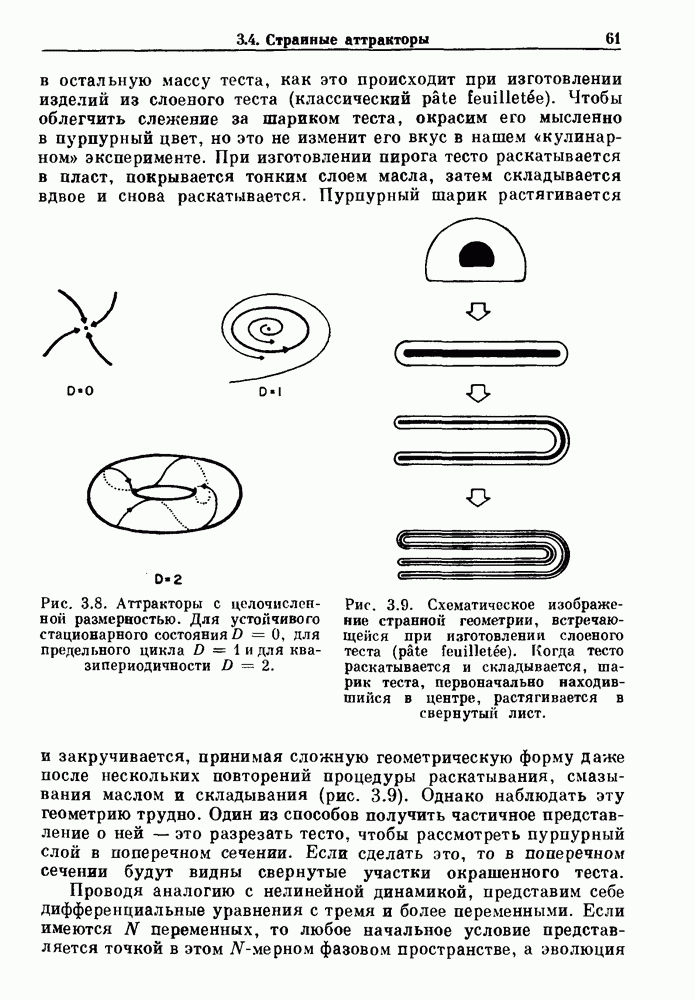








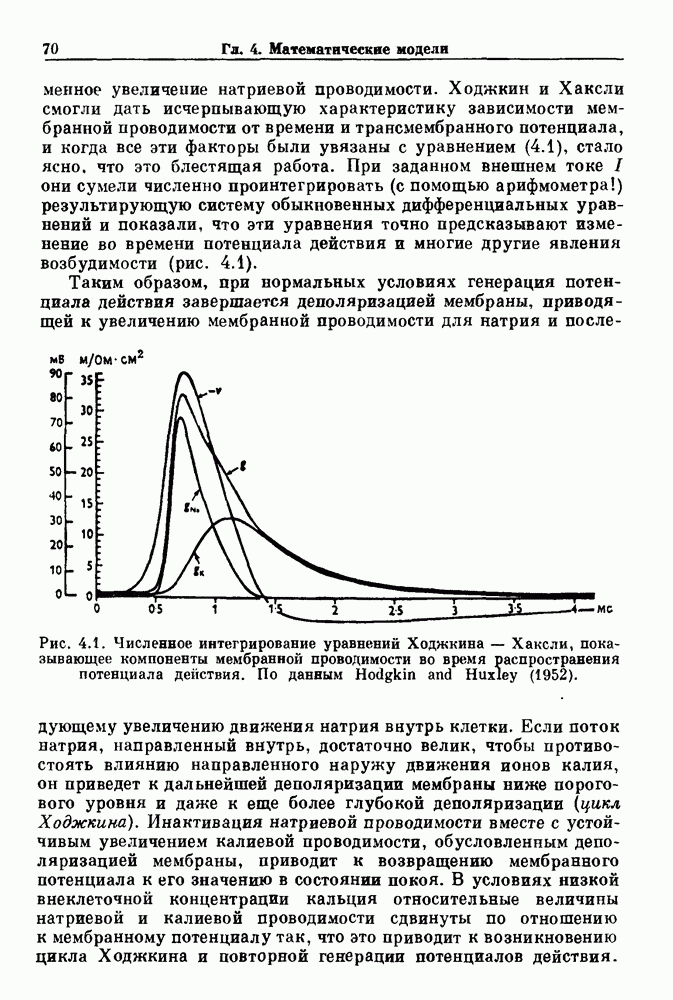

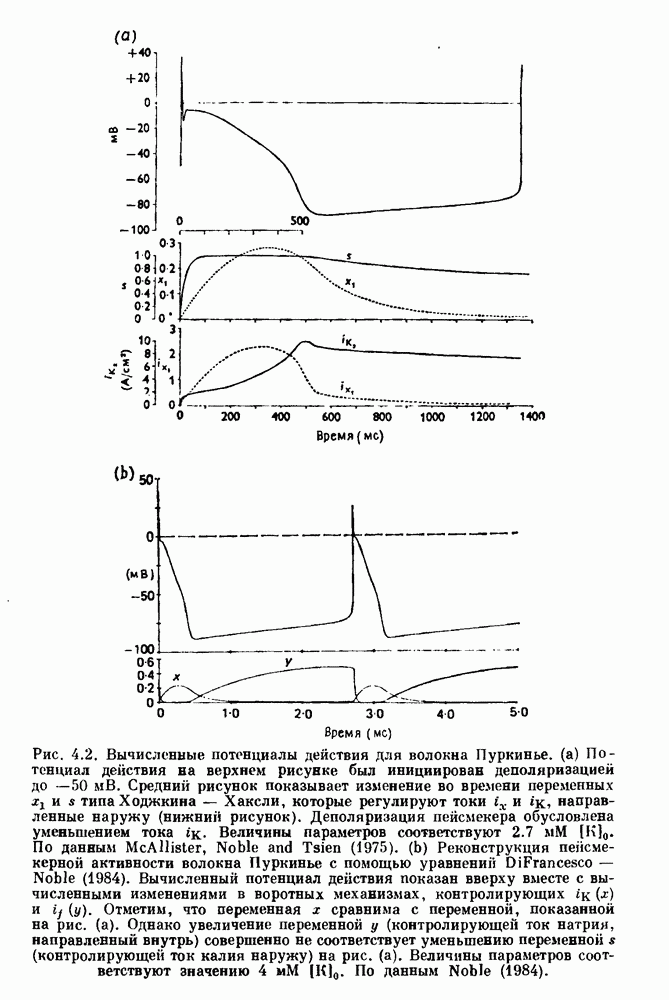
















































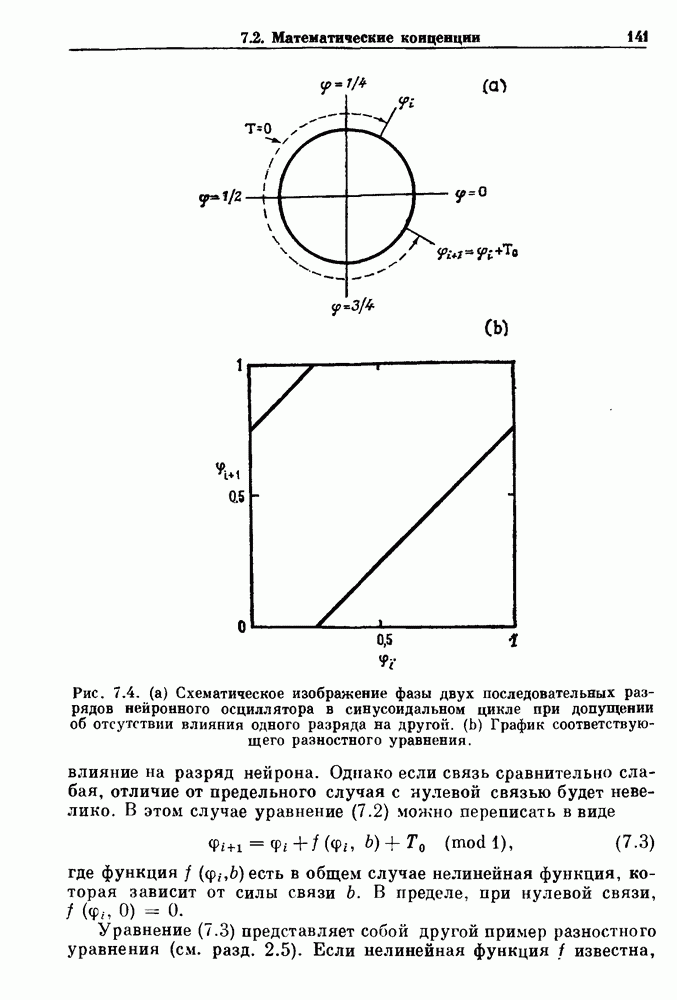



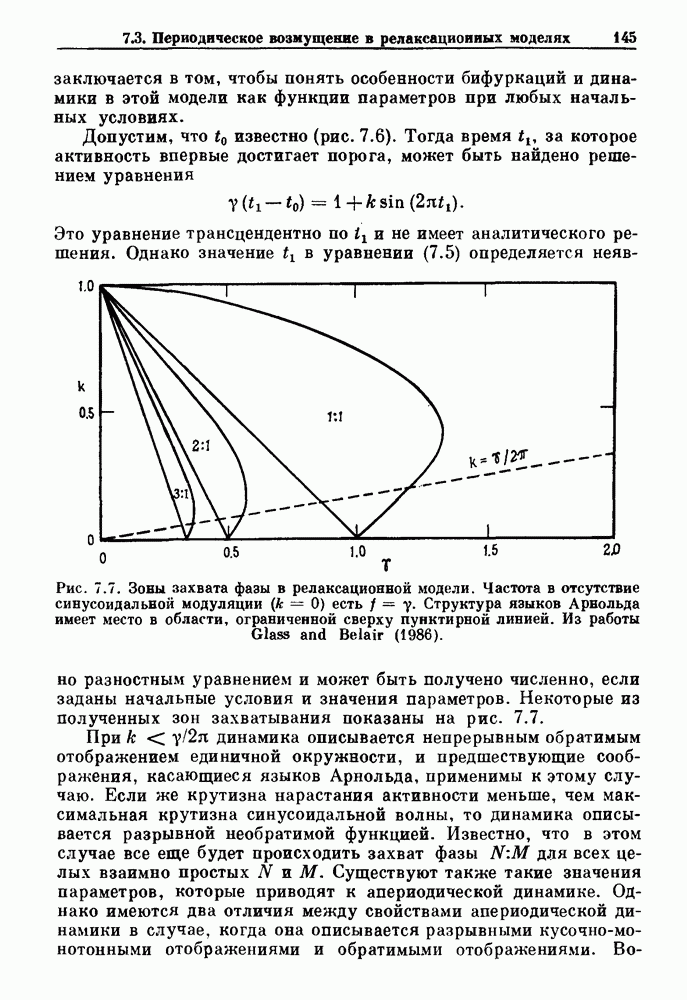
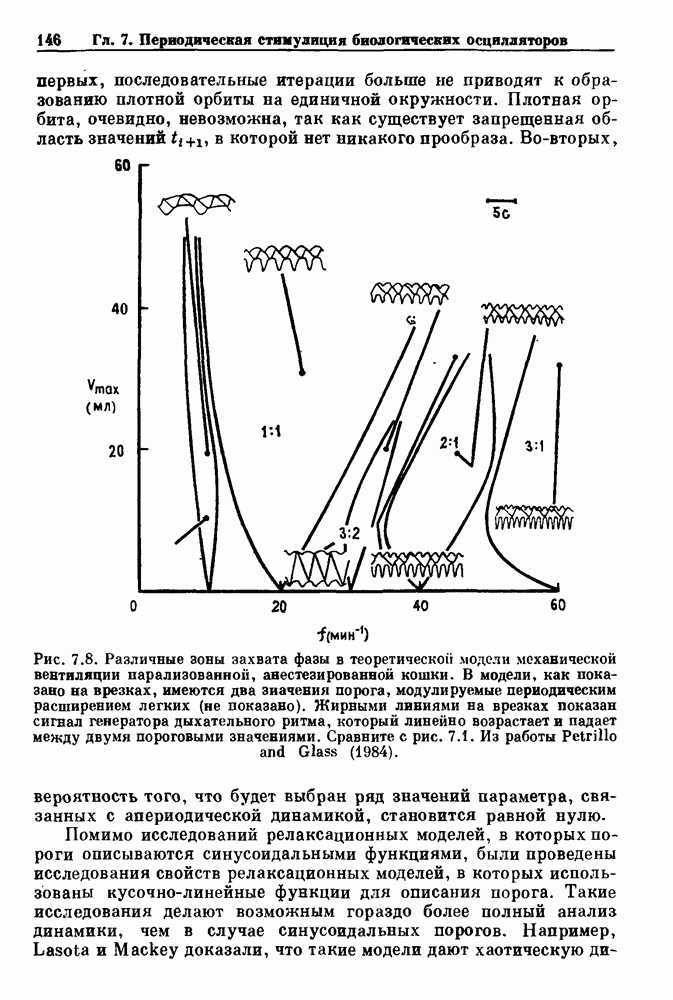


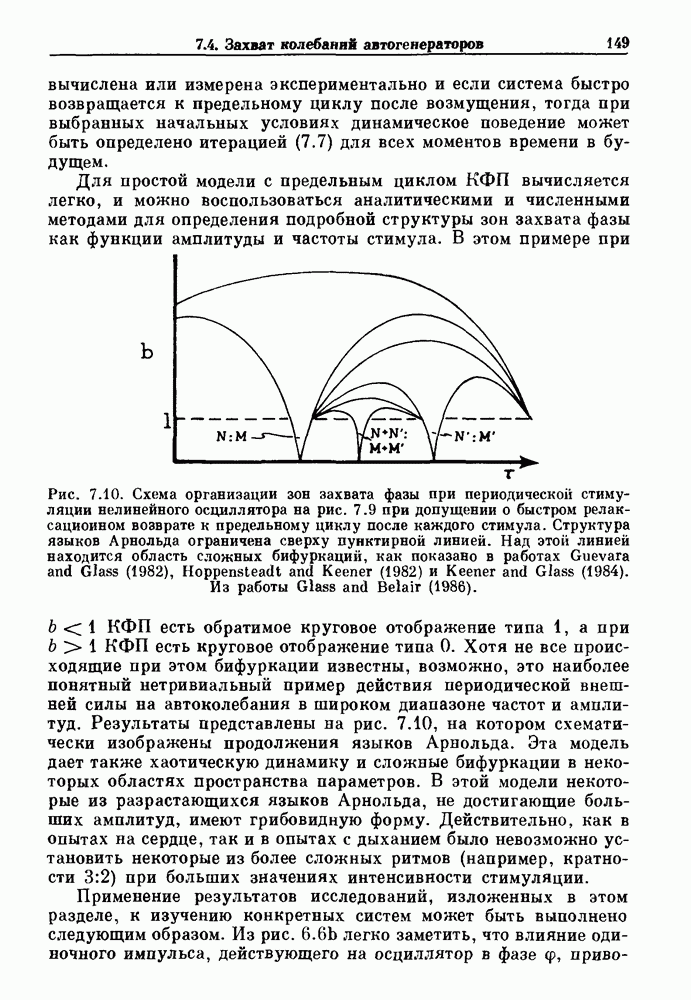























































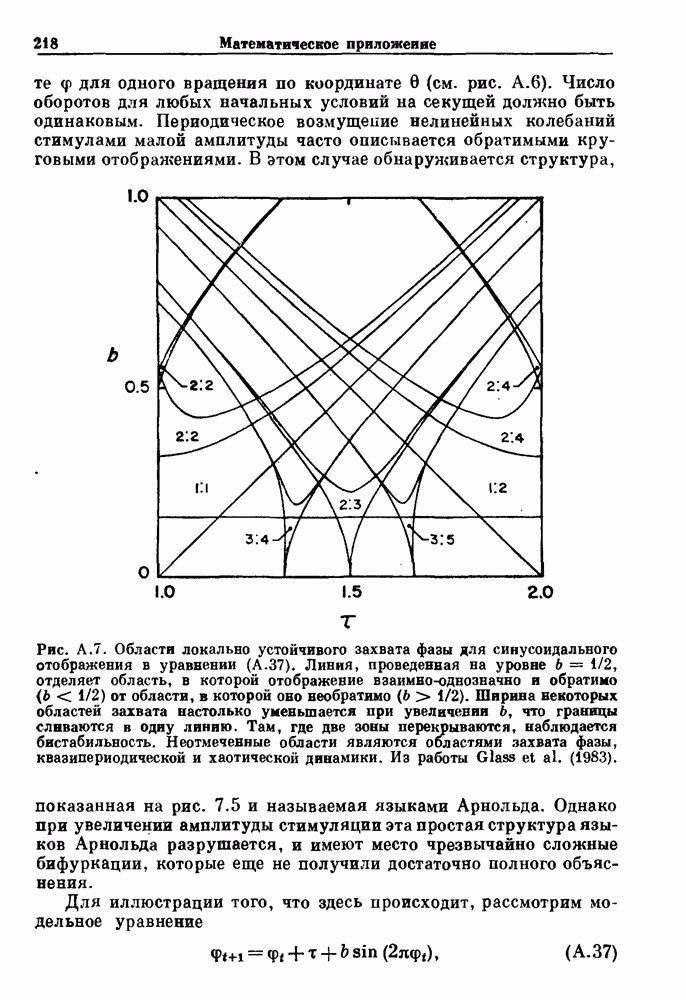


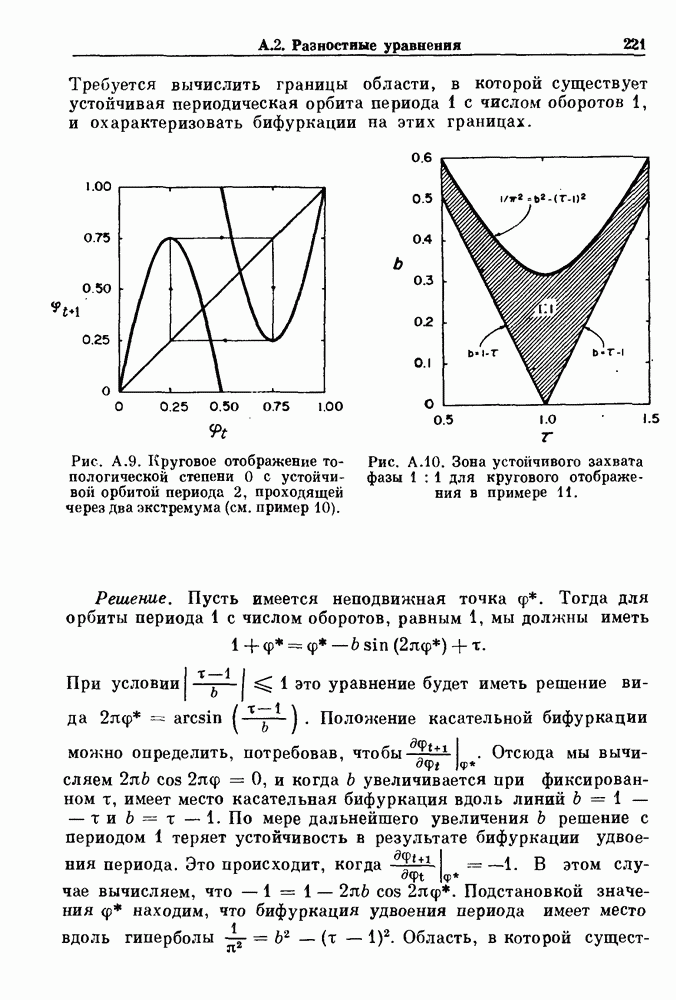


























 (3.8)
(3.8)
 . На рис. 3.5b показана временная последовательность, полученная решением уравнения (3.7) при
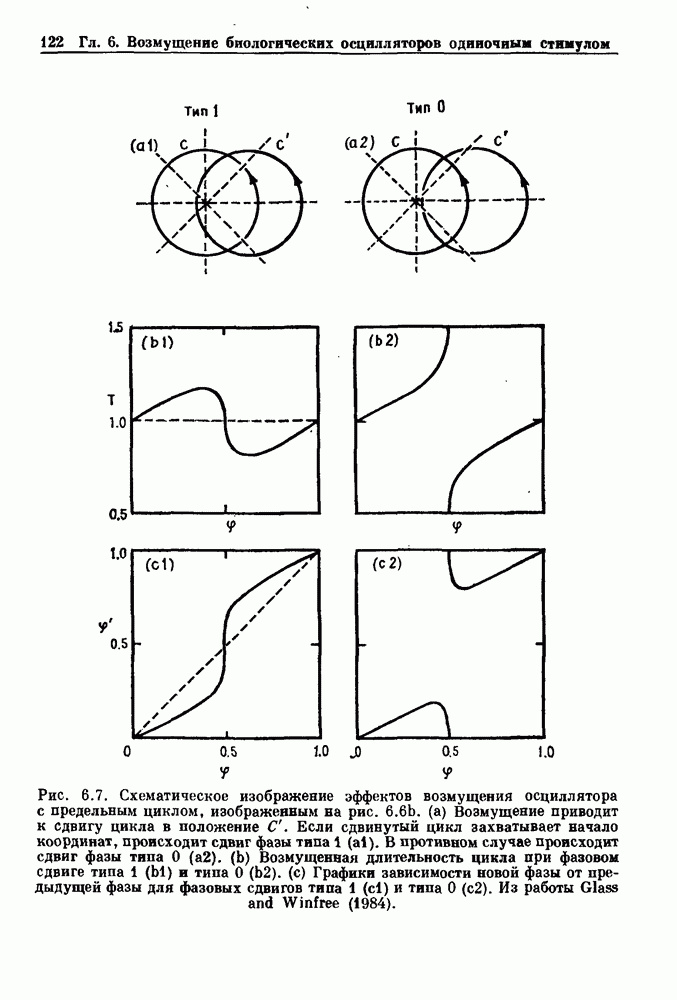
. На рис. 3.5b показана временная последовательность, полученная решением уравнения (3.7) при  Чтобы показать, что такие временные последовательности действительно имеют экспоненциальную плотность, определяемую уравнением (3.7), на рис. 3.5b представлена численно определенная плотность вероятности для этой временной последовательности. Следовательно, одного только факта, что плотность распределения интервалов является экспоненциальной, недостаточно для того, чтобы считать процесс пуассоновским. На основании графика зависимости данной величины от ее предшествующего значения можно судить о структуре процесса, если этот процесс генерируется одномерным разностным уравнением, подобным уравнению (3.7). Однако идентифицировать хаос в детерминированных системах более высокого порядка трудно, простых методов для этого не существует.
Чтобы показать, что такие временные последовательности действительно имеют экспоненциальную плотность, определяемую уравнением (3.7), на рис. 3.5b представлена численно определенная плотность вероятности для этой временной последовательности. Следовательно, одного только факта, что плотность распределения интервалов является экспоненциальной, недостаточно для того, чтобы считать процесс пуассоновским. На основании графика зависимости данной величины от ее предшествующего значения можно судить о структуре процесса, если этот процесс генерируется одномерным разностным уравнением, подобным уравнению (3.7). Однако идентифицировать хаос в детерминированных системах более высокого порядка трудно, простых методов для этого не существует.
 спустя. Вероятность того, что клетка, выбранная случайным образом из
спустя. Вероятность того, что клетка, выбранная случайным образом из
 . (b) Временные последовательности, полученные итерированием функции (а). (c) Соответствующая плотность вероятности. Это хаотический процесс с экспотенциальной плотностью вероятности.
. (b) Временные последовательности, полученные итерированием функции (а). (c) Соответствующая плотность вероятности. Это хаотический процесс с экспотенциальной плотностью вероятности.
 есть просто
есть просто

 , которые не поделились к моменту времени t после их рождения, равна
, которые не поделились к моменту времени t после их рождения, равна


 Кроме того, эти данные позволяют проверить другую статистику,
Кроме того, эти данные позволяют проверить другую статистику,  , представляющую собой долю сестринских пар клеток, клеточный цикл которых отличается по крайней мере на время t. Модель со случайным переходом предсказывает, что
, представляющую собой долю сестринских пар клеток, клеточный цикл которых отличается по крайней мере на время t. Модель со случайным переходом предсказывает, что

 , определялась длительность клеточного цикла в различных клеточных популяциях. Во всех случаях эта детерминированная хаотическая модель дает такое же хорошее согласие с данными, показанными на рис. 3.6b, что и модель со случайным переходом. Что это, шум или хаос?
, определялась длительность клеточного цикла в различных клеточных популяциях. Во всех случаях эта детерминированная хаотическая модель дает такое же хорошее согласие с данными, показанными на рис. 3.6b, что и модель со случайным переходом. Что это, шум или хаос?
 представляет долю клеток в популяции, не поделившихся к моменту времени t после
представляет долю клеток в популяции, не поделившихся к моменту времени t после  рождения, как функцию времени, а
рождения, как функцию времени, а  долю сестринских пар клеток, длительность клеточного цикла которых отличается по крайней мере на время t. (а) Подгонка к данным с помощью модели со случайным переходом Smith and Martin (1973). (b) Подгонка к данным с помощью модели Mackey, Santavy and Selepova (1986). Адаптировано из работы Mackey, Santavy and Selepova (1986).
долю сестринских пар клеток, длительность клеточного цикла которых отличается по крайней мере на время t. (а) Подгонка к данным с помощью модели со случайным переходом Smith and Martin (1973). (b) Подгонка к данным с помощью модели Mackey, Santavy and Selepova (1986). Адаптировано из работы Mackey, Santavy and Selepova (1986).

 , где
, где  — параметр, зависящий от модели. Таким образом, предсказания модели очень хорошо согласуются с данными по выживанию, представленными на рис. 3.7, которые аппроксимировались (сплошная линия) функцией такого же вида при
— параметр, зависящий от модели. Таким образом, предсказания модели очень хорошо согласуются с данными по выживанию, представленными на рис. 3.7, которые аппроксимировались (сплошная линия) функцией такого же вида при
