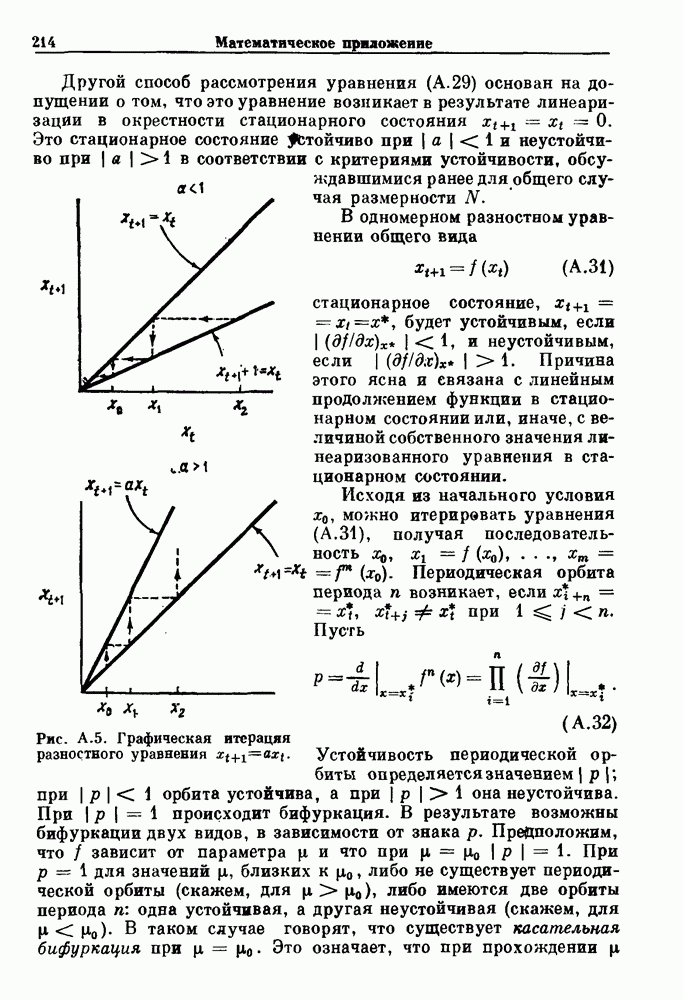
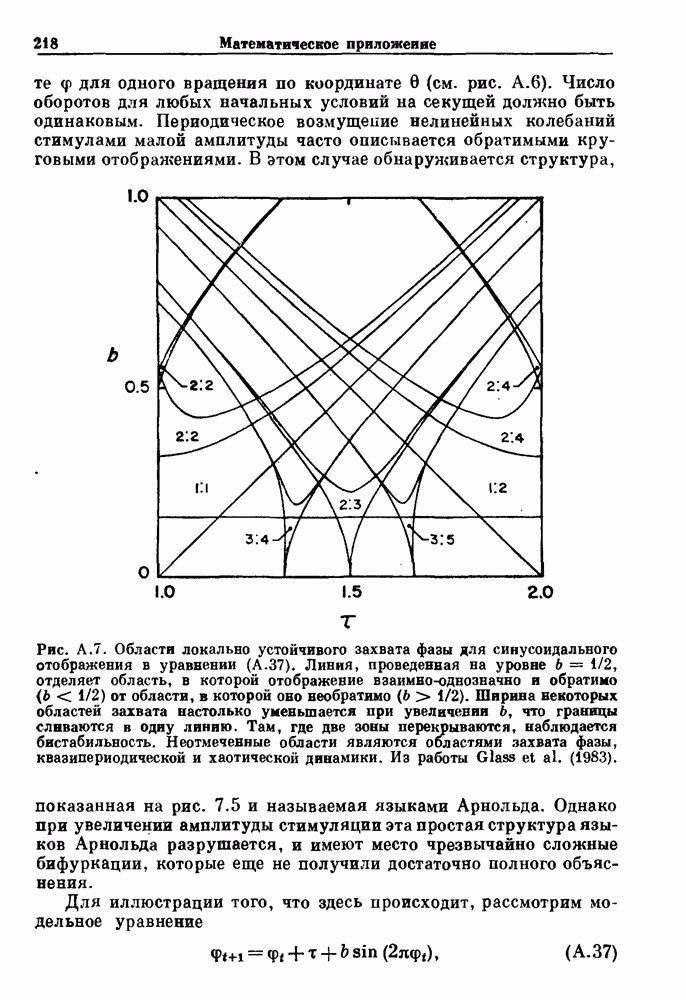
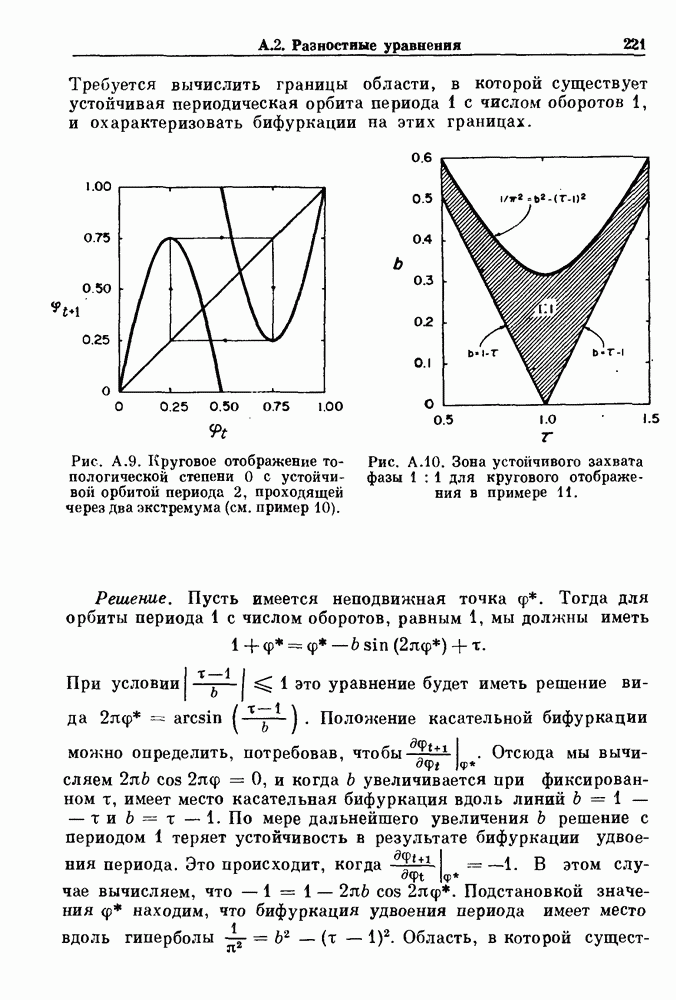
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
4.3. Взаимное ингибированиеРитмическое движение сустава обычно совершается периодической активацией противодействующих групп мышц: сгибателей и разгибателей. Самые первые представления о возможных механизмах генерации такой ритмической активности, относящиеся к началу века, связаны с предположением о существовании двух групп нейронов. Взаимодействия внутри каждой группы являются возбуждающими, а взаимодействия между группами — угнетающими. Таким образом, возникает взаимное угнетение, представленное в виде диаграммы на рис. 4.5. Такой механизм был назван Брауном (Brown) в 1914 г. half-center model, но этот термин в настоящее время редко используется. Предположим, что два взаимно угнетающих нейрона получают тонический сигнал. Тогда в зависимости от силы сигнала и силы угнетающего взаимодействия реализуется один из двух возможных типов качественного динамического поведения. Оба нейрона могут сохранить свою тоническую активность, но при более низкой частоте, чем в отсутствие угнетения. В другом случае частота разряда одного из нейронов высокая, а частота разряда другого — низкая.
Рис. 4.5. Модель нейронной сети с взаимным ингибированием. Предполагается, что каждый нейрон получает возбуждающий импульс и обладает тонической активностью при отсутствии угнетающего синапса. Такая система может давать колебания только при наличии некоторых дополнительных факторов, таких как утомление или возбуждение после торможения, которые модулируют активность при постоянной стимуляции. Этот последний случай аналогичен явлению, называемому в экологии конкурентным вытеснением, при котором один из двух конкурирующих видов побеждает в борьбе за пищу. Такие взаимно исключающие взаимодействия могут также служить основой для нейрофизиологического триггерного переключения. Для возникновения колебаний в системе двух нейронов с взаимным угнетением необходимы дополнительные физиологические факторы (такие как утомление, адаптация или возбуждение вслед за угнетением) для создания механизма переключения, чтобы сначала один нейрон, а затем другой становился активным. По мере того как клетка продолжает разряжаться с высокой частотой, происходит истощение метаболитов и накопление продуктов распада, соответствующее утомлению клетки. Это утомление приводит к уменьшению клеточной активации. При включении утомления в теоретическую модель можно получить колебательное поведение в системе с взаимным ингибированием. Адаптация также может приводить к уменьшению клеточной активности и колебаниям. Другим важным физиологическим механизмом считается возбуждение, часто наблюдаемое после сильного угнетающего воздействия на нейрон. Perkel и Mulloney показали в 1974 г., что в случае такого возбуждения вслед за угнетением можно получить колебательное поведение в системе из двух взаимно угнетающих нейронов при условии, что это возбуждение является достаточно сильным. Такие системы могут находиться также в устойчивом стационарном состоянии, при котором в обоих нейронах наблюдается низкий уровень тонической активности. Переход в колебательное состояние осуществляется путем сильного угнетающего воздействия на одну из клеток. Эффект возбуждения вслед за угнетением в этой клетке затем приводит к возникновению разряда высокой частоты, в результате чего происходит сильное угнетение другой клетки, и возникают устойчивые колебания. Конкретным примером колебаний в системе с взаимным угнетением являются данные, полученные при изучении стоматога-стрального ганглия у рака. Можно изолировать этот ганглий и изучать свойства клеток и сетей, лежащие в основе рит могенеза.
Рис. 4.6. Чередование разрядной активности, наблюдавшееся в LP и PD нейронах из стоматогастрального ганглия рака. Эти два нейрона являются взаимно угнетающими. Такая активность послужила первой четкой демонстрацией наличия полуцентральных колебаний в биологической системе. По данным Selverston, Miller and Wadepuhl (1983). Путем внутриклеточного введения светочувствительного красителя и облучения интенсивным светом можно избирательно убивать клетки стоматогастрального ганглия. Этот изящный метод можно применить для изоляции двух клеток, обозначаемых PD и LP, которые играют важную роль в ритмогенезе. Если правильно подобрать экспериментальные условия, то можно получить колебания в системе из двух клеток, даже если ни одна из них сама по себе не генерирует спонтанных колебаний (рис. 4.6). Хотя клеточные пары с взаимным угнетением были обнаружены также в других ГЦТ, существование таких взаимодействий не обязательно означает, что пара с взаимным угнетением представляет собой ГЦТ. Она может быть лишь частью ГЦТ.
|
 |
Оглавление
|