Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
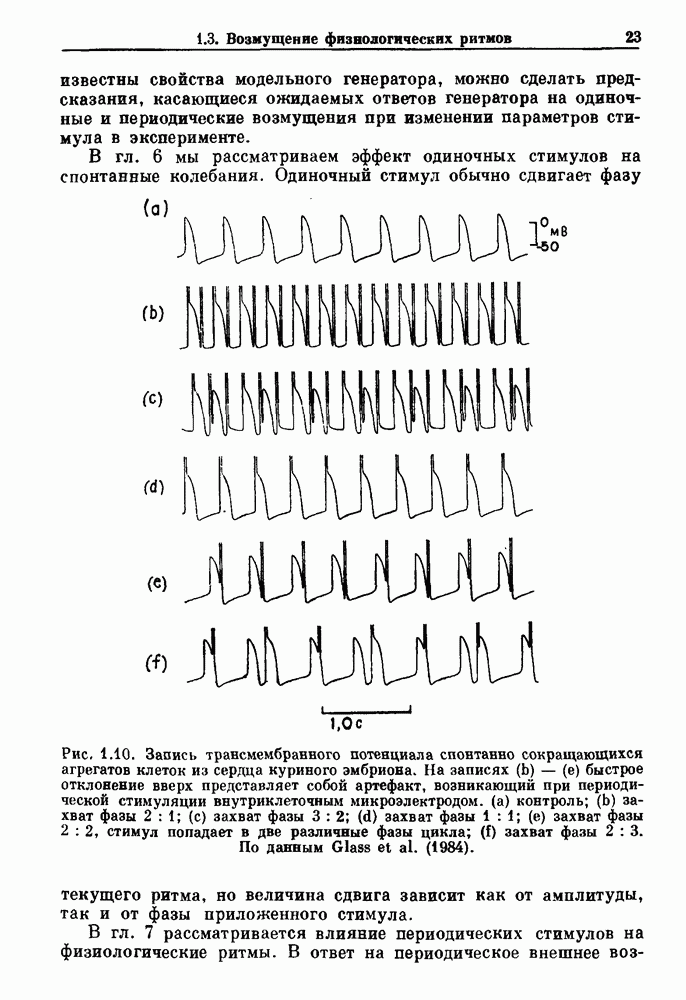
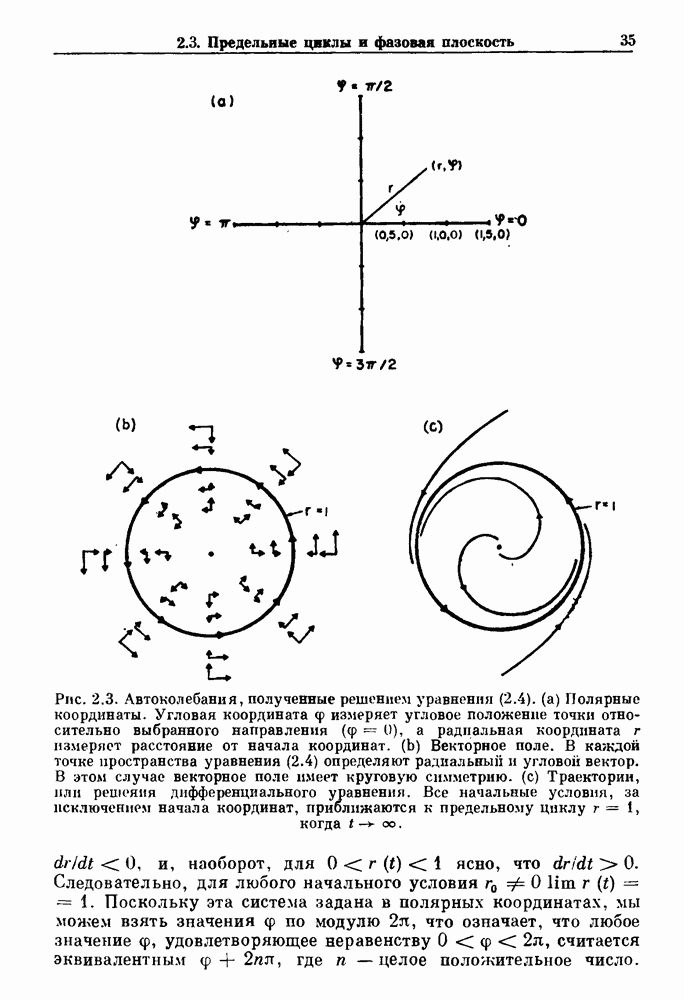
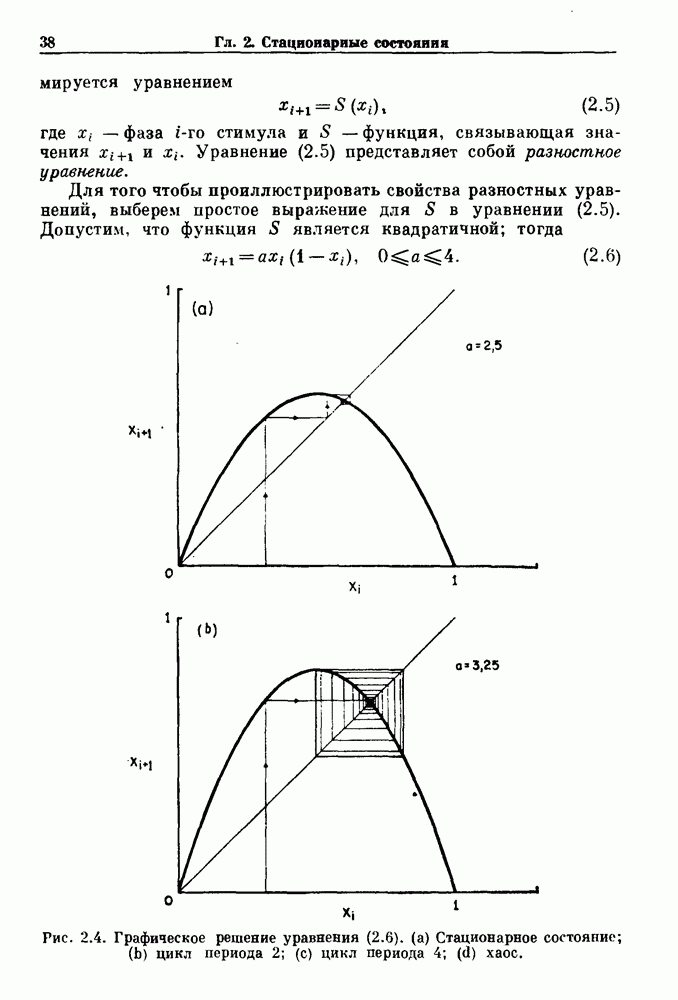
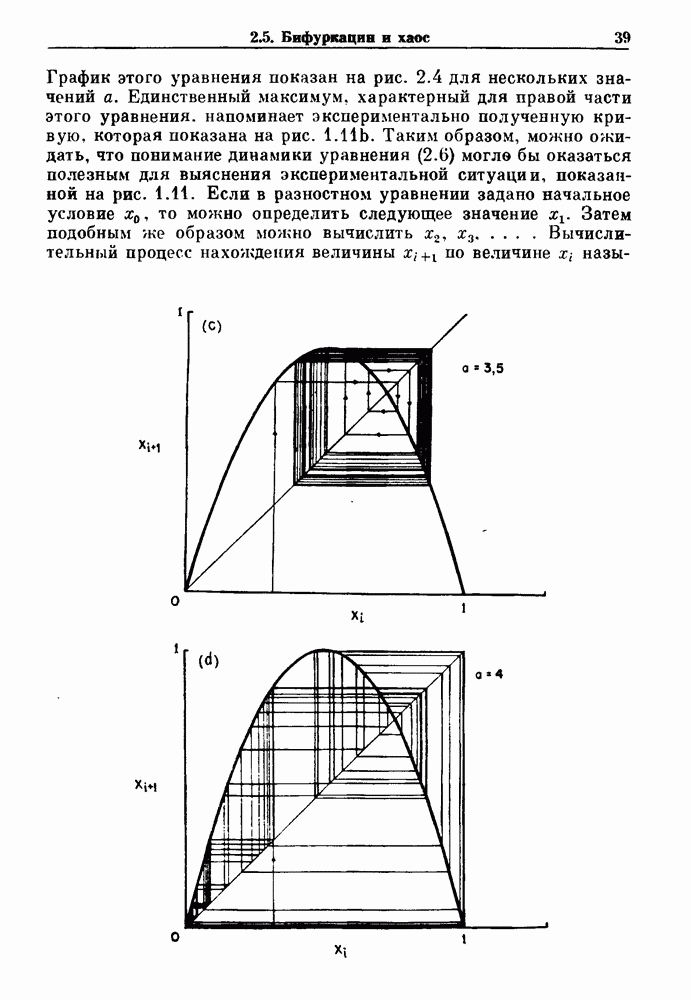
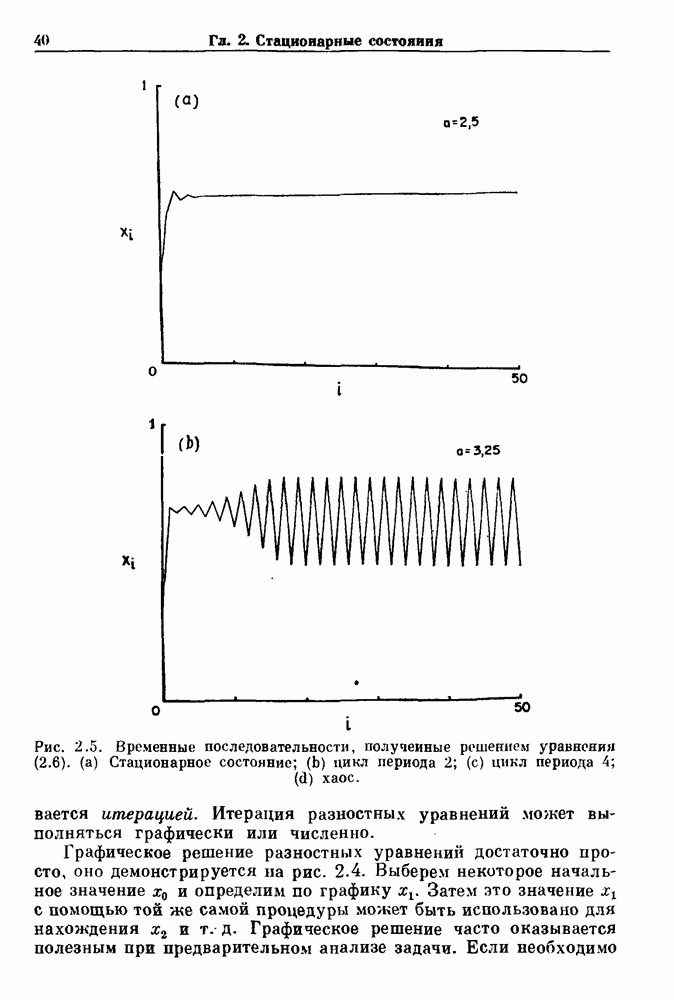
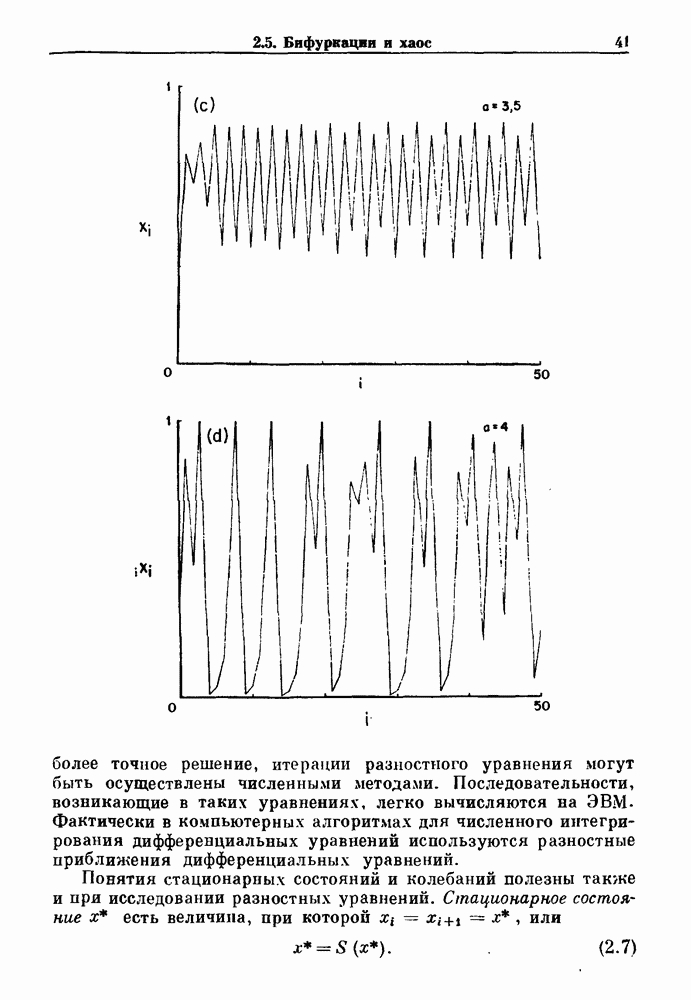
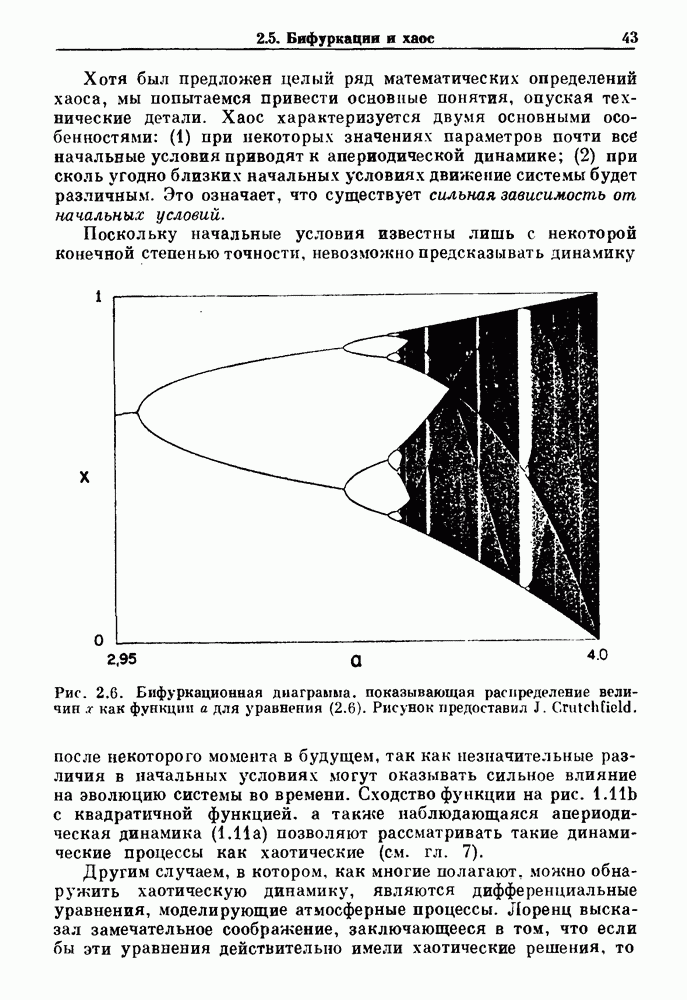
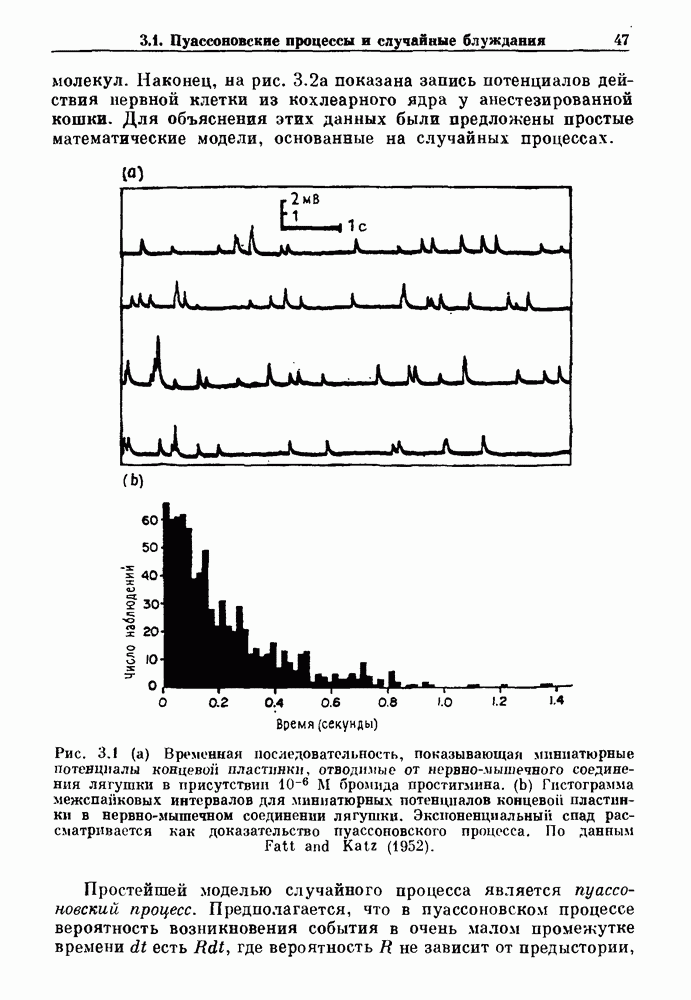
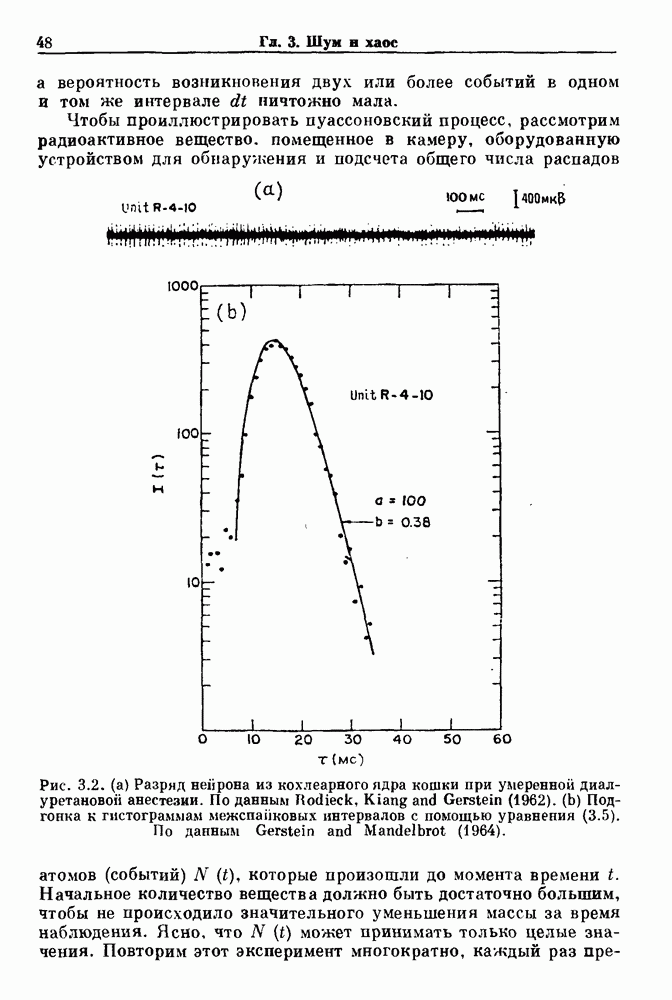
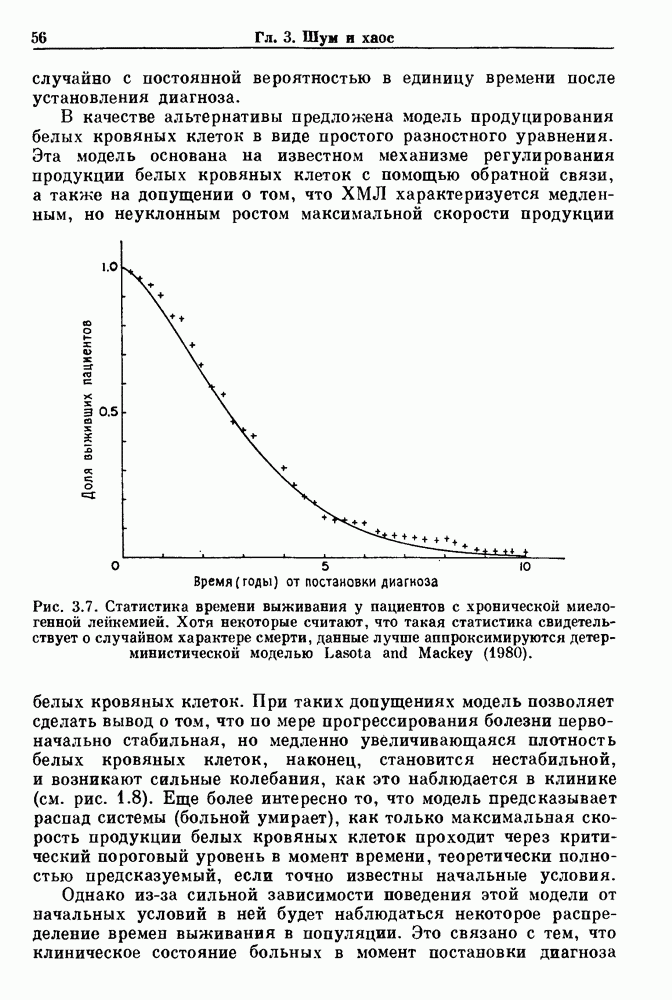
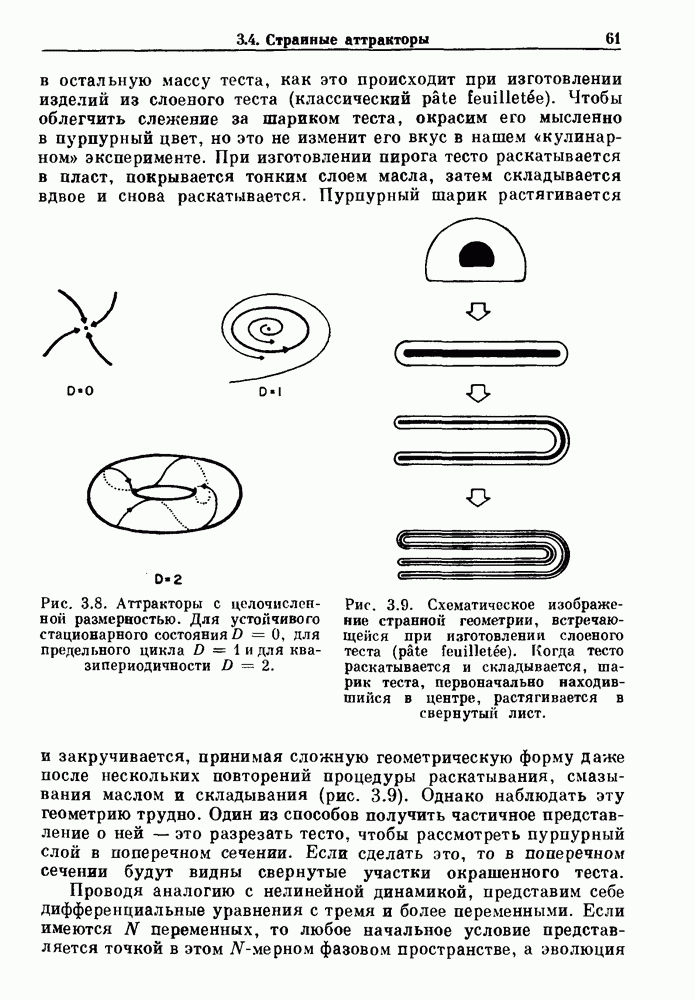
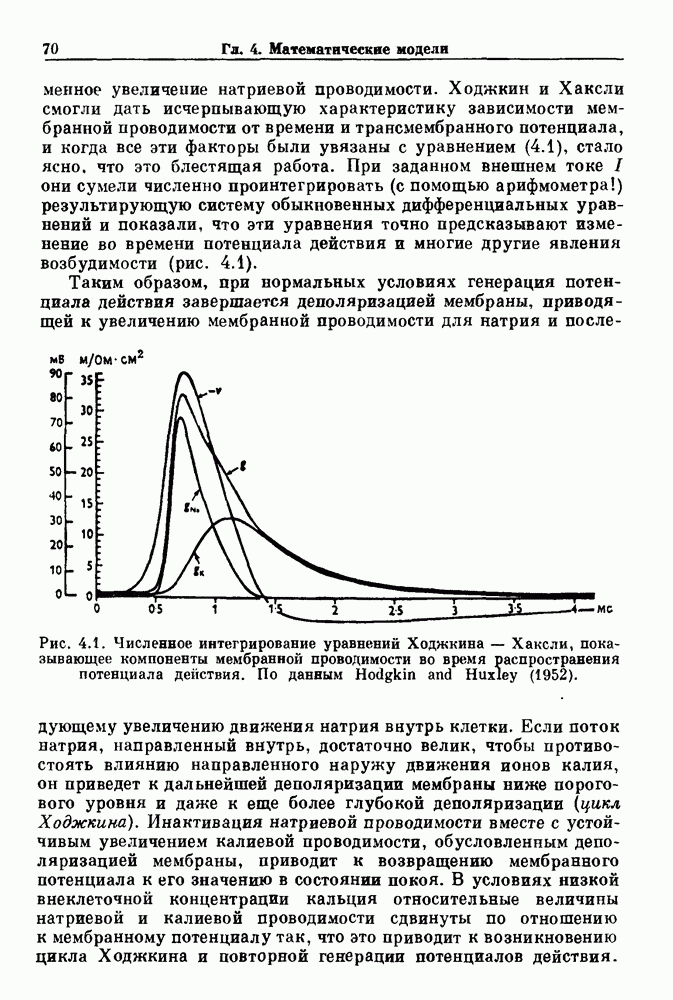
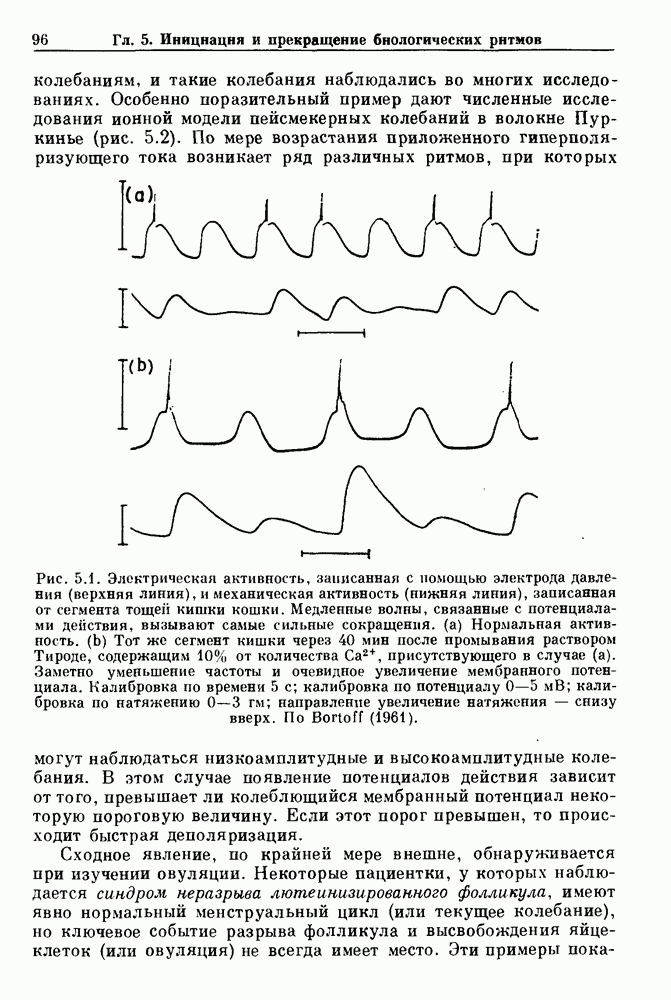
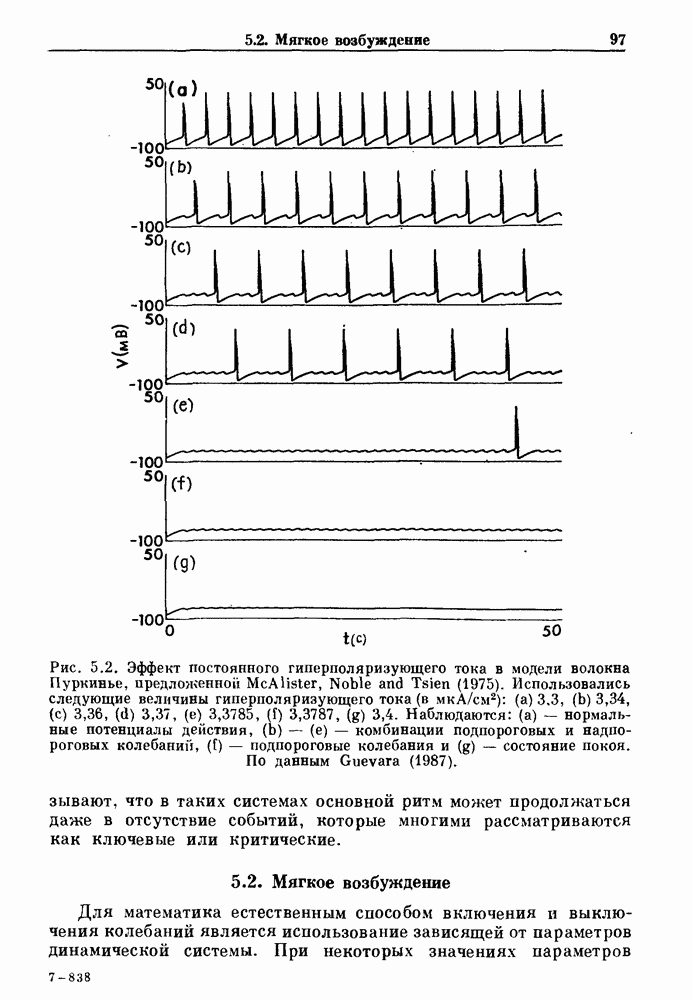
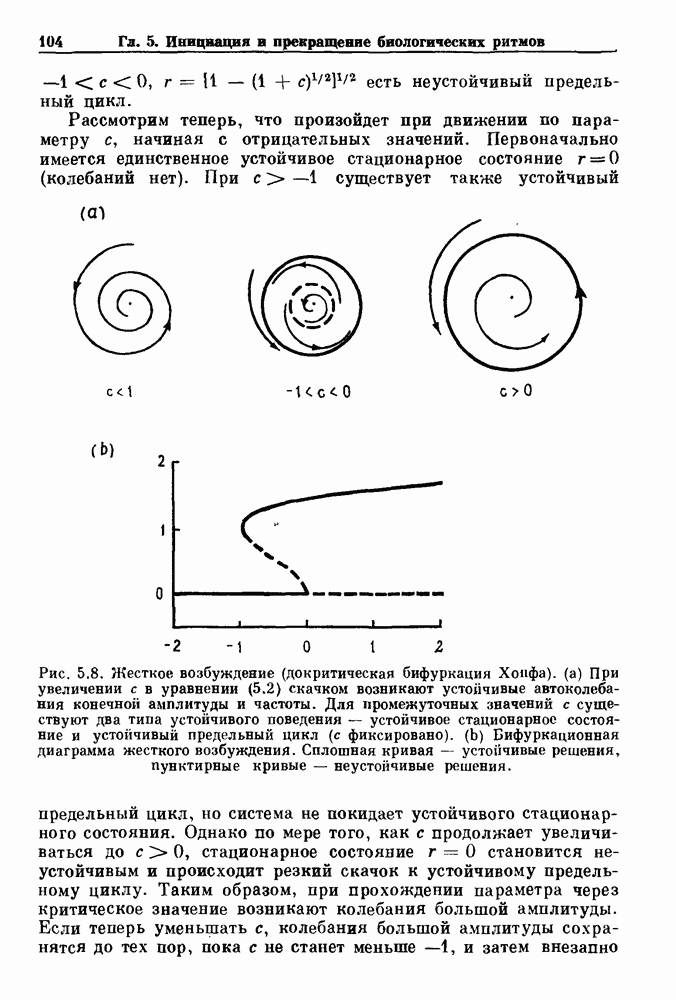
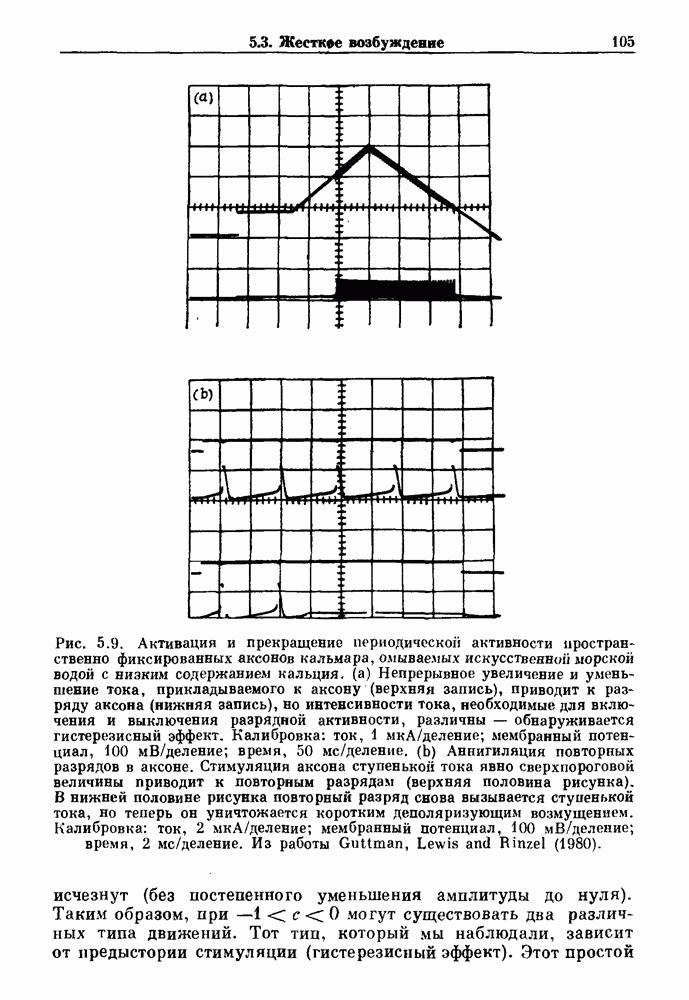
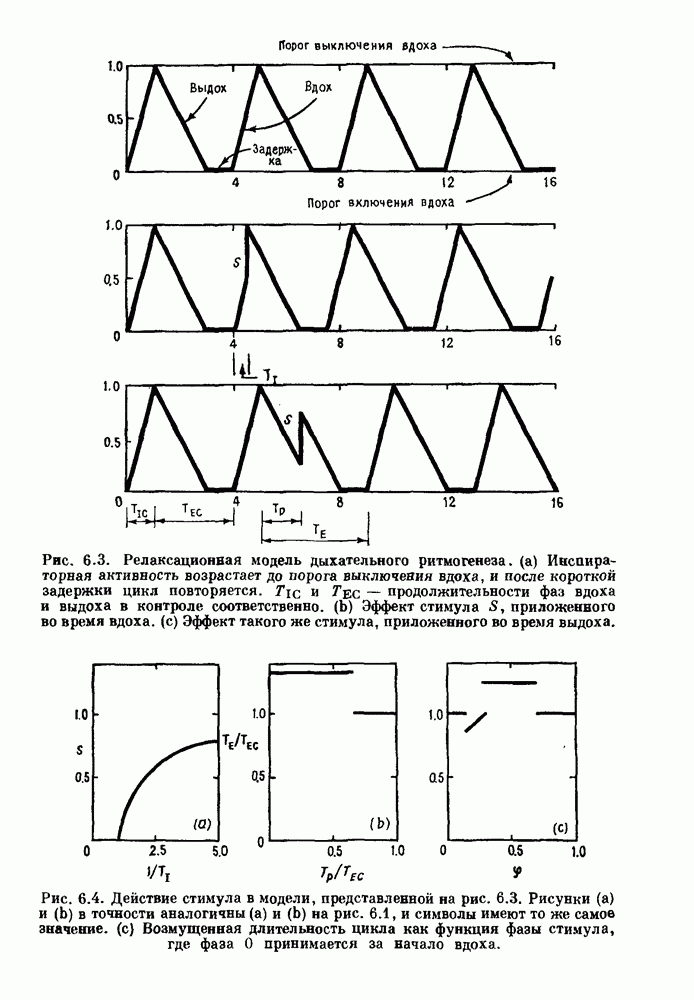
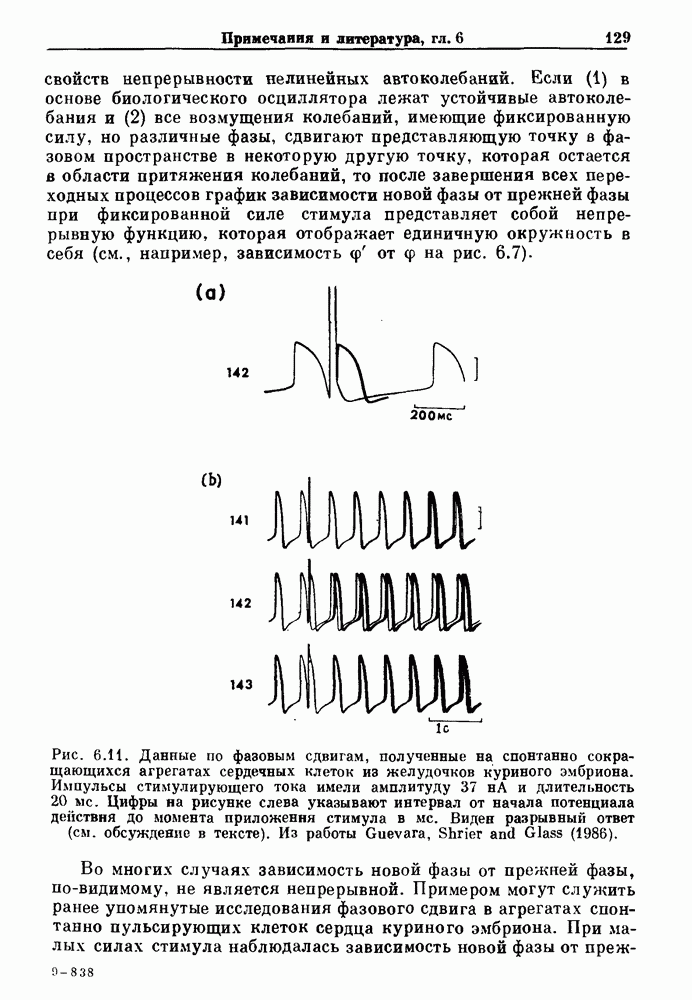
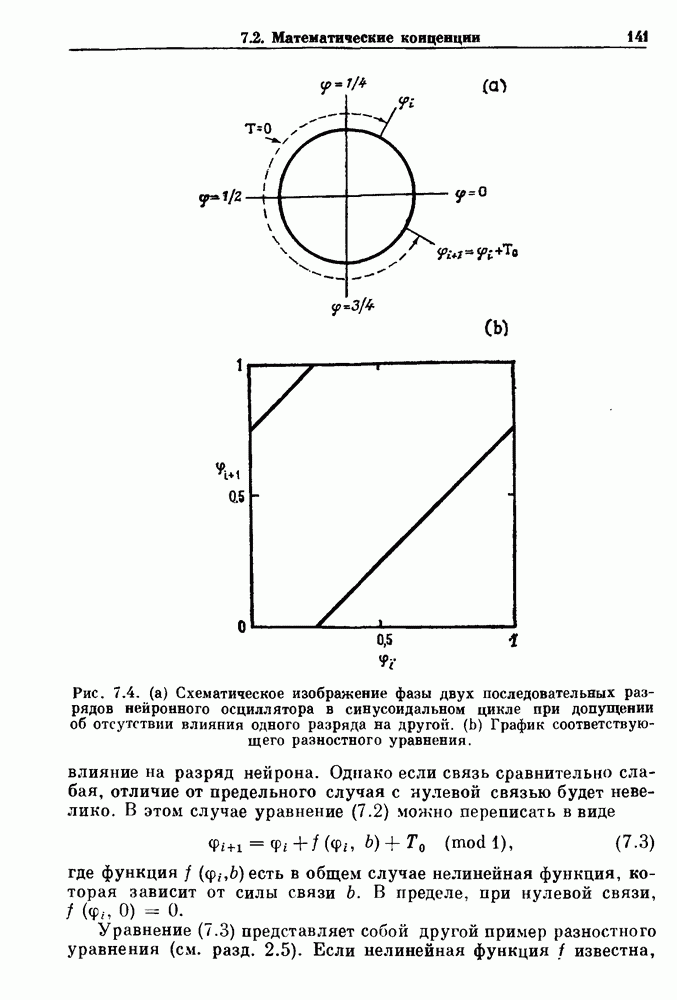
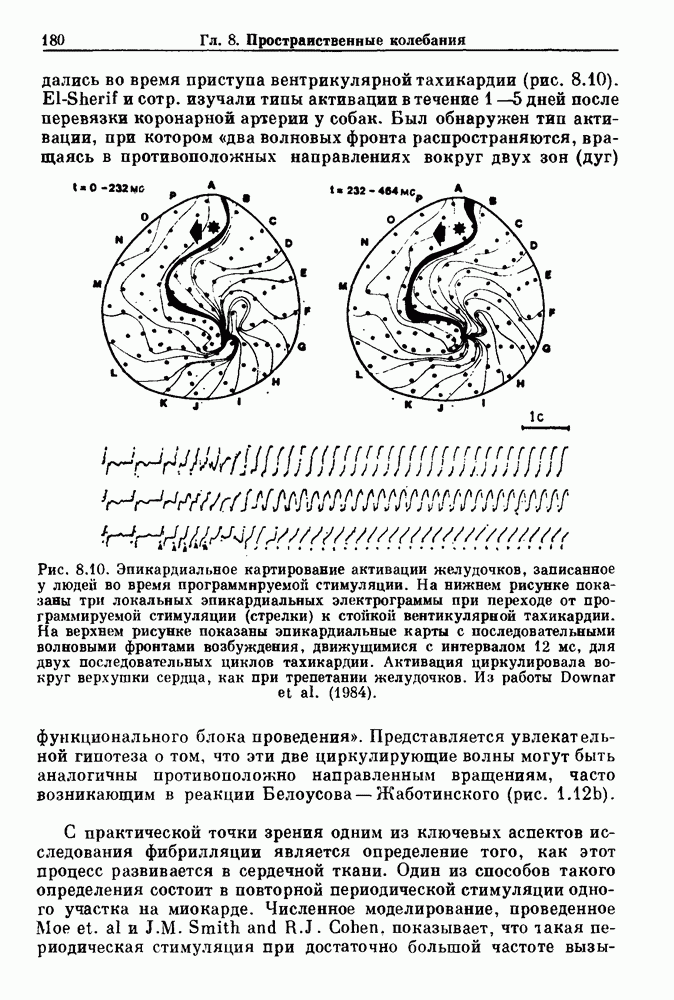
Глава 8. Пространственные колебанияФизиологические ритмы упорядочены как в пространстве, так и во времени. Обычно колебания обнаруживают способность к простому волнообразному распространению, начинающемуся из области пейсмекера. Однако во многих случаях вместо периодического распространения волн, исходящих из точечного источника, наблюдаются другие типы пространственной организации. В этой главе мы обсуждаем подобные необычные режимы распространения волн. Распространение волн в одномерной полоске ткани рассматривается в разд. 8.1, а в одномерном кольце ткани — в разд. 8.2. В разд. 8.3 мы обсуждаем распространение волн в двумерной системе, а в разд. 8.4 — в трехмерной системе. Полагают, что фибрилляция связана с аномальным риентри, или круговым распространением возбуждения в двух- и трехмерной среде. Теоретические и экспериментальные исследования фибрилляции обсуждаются в разд. 8.5. 8.1. Одномерное распространение волнХорошим примером физиологической системы, в которой происходит одномерное распространение волн, является мочеточник. Мочеточники транспортируют мочу от почек к мочевому пузырю с помощью перистальтических волн, начинающихся в ограниченной области около места соединения мочеточника с почечной лоханкой, называемой почечно-лоханочным пейсмекером. Распространение волн в мочеточнике можно изучать либо путем измерения спонтанной активности, либо путем электрической стимуляции одного конца мочеточника и измерения электрической активности на противоположном конце. В опытах на интактных собаках (рис. 8.1 а) наблюдалась передача возбуждения 1:1 при периоде стимула, равном 4 с (верхняя запись) и 5:4 при периоде, равном 3.38 с (нижняя запись). При соотношении между частотой стимуляции и собственной частотой 5:4 длительность латентного периода между стимулом и следующей волной возбуждения увеличивается до тех пор. пока, наконец, не произойдет выпадение одного из биений. Аналогичный эффект возникает при проведении импульса в сердце человека. Несмотря на то, что сердце представляет собой сложную трехмерную структуру, для наших целей мы будем считать его одномерным, причем возбуждение передается по следующему пути: синоатриальный узел — предсердия — атриовентрикулярный узел — пучок Гиса — волокна Пуркинье — желудочки. Сокращения желудочков обычно следуют за сокращениями предсердий спустя 0.12-0.20 с (рис. 1.2).
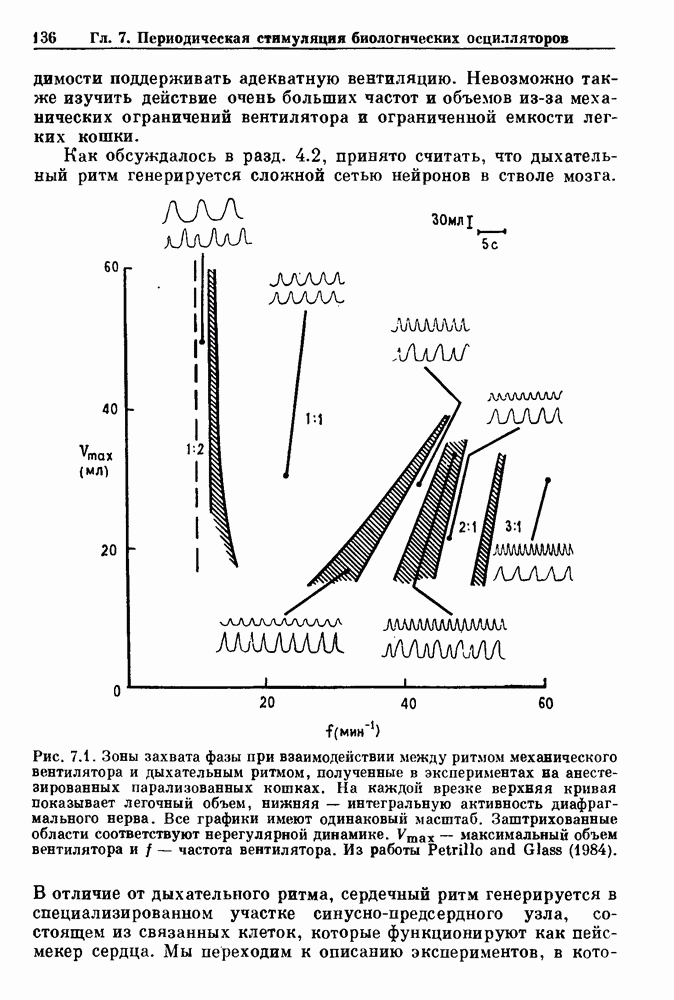
Рис. 8.1. Ответ мочеточника на периодическую электрическую стимуляцию, (а) Запись, полученная от анестезированной собаки. Короткими штрихами отмечены моменты приложения стимула, сплошная кривая показывает электрическую активность, отводимую расположенным вне мочеточника биполярным электродом. Верхняя запись показывает проведение 1:1, когда период стимула был равен 4 с, на нижней записи виден ритм Венкебаха при периоде стимуляции, равном 3.38 с (из работы Weiss, Wagner and Hoffmann (1968)). (b) Отведение электрической активности от мочеточника крысы in vitro. Видно чередование высокоамплитудных и низкоамплитудных ответов при периодической стимуляции (из работы Prosser, Smith and Melton (1955)). Однако атриовентрикулярная блокада сердца характеризуется аномальной координацией между предсердными и желудочковыми ритмами, приводящей к удлинению интервала между предсердными и желудочковыми сокращениями (АВ-блокада I степени), к увеличению числа предсердных сокращений по сравнению с числом желудочковых сокращений из-за блокированного проведения некоторых из предсердных биений (АВ-блокада II степени) или к полному отсутствию координации между предсердными и желудочковыми ритмами (АВ-блокада III степени). Блокада II степени, при которой происходит увеличение интервала между сокращениями предсердий и желудочков (интервал Р — R), приводящее к выпадению биения, показан на рис. 8.2а. Ритмы с увеличивающимся латентным периодом между (см. скан) Рис. 8.2. (а) Электрокардиограмма, показывающая последовательное удлинение интервала стимулом и возбуждением и возможным выпадением биения известны теперь под названием ритмов Венкебаха. Другое явление, возникающее как в мочеточнике, так и в сердце, заключается в чередовании ответов на периодическую последовательность импульсов. На рис. 8.1b показано чередование ответов, которое можно наблюдать при воздействии на изолированный препарат мочеточника быстрых периодических стимулов. Подобные явления обнаруживаются в кардиологии, в электрограммах возникают чередования импульсов разной амплитуды или электрических комплексов (альтернанс) (рис. 8.2 в). Альтернанс часто обнаруживается при больших скоростях сердечных сокращений и может считаться нормальным ответом, но его появление при более низких скоростях часто рассматривается как признак дисфункции. Несмотря на то, что ответ одномерной возбудимой ткани на периодическую стимуляцию высокой частоты, имеющий характер альтернанса, встречается не так часто, как ритмы Венкебаха, такой ответ все же является достаточно общим, чтобы поставить вопрос о его возможном механизме. В заключение отметим, что периодическая стимуляция возбудимой одномерной ткани при низких частотах приводит к возникновению периодических распространяющихся волн возбуждения равной амплитуды, которые следуют за периодическим стимулом в соотношении Физиологическую основу для понимания такой динамики составляют следующие три наблюдения, сделанные Майнсом в начале века: 1) по мере увеличения частоты стимуляции скорость распространения волны уменьшается (зависимость скорости распространения от частоты называется дисперсионным отношением; 2) по мере увеличения частоты стимуляции длительность возбуждения уменьшается; 3) после возбуждения следует временной интервал, называемый рефрактерным периодом (в), во время которого следующий стимул не может привести к возбуждению. Включение этих свойств в математическую модель позволяет частично понять описанную выше динамику. Схематическое изображение физиологической ситуации представлено на рис. 8.3. Стимул (S вызывает возбуждение после некоторой задержки (APD) при возбуждении, вызываемом стимулом
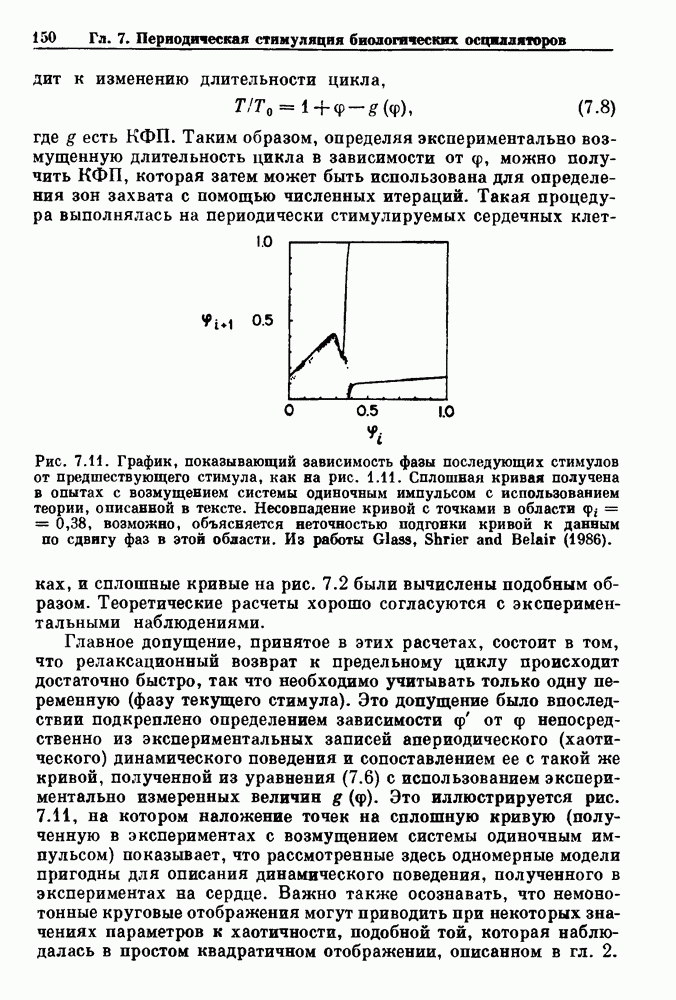
Рис. 8.3. Схематическая диаграмма, показывающая периодическую стимуляцию возбудимой ткани с интервалом времени Обозначая временной интервал между стимулами через
где F и (7 — искомые функции. Хотя уравнения (8.1а,б) выписаны для распространения волн в распределенных системах, они применимы также к пространственно гомогенным системам, в которых стимул и ответ на него измеряются в одной и той же точке. Насколько нам известно, общий анализ уравнений (8.1а,б) не проводился, однако, были рассмотрены два специальных случая. В первом случае мы полагаем, что длительность возбуждения является постоянной и что, следовательно,
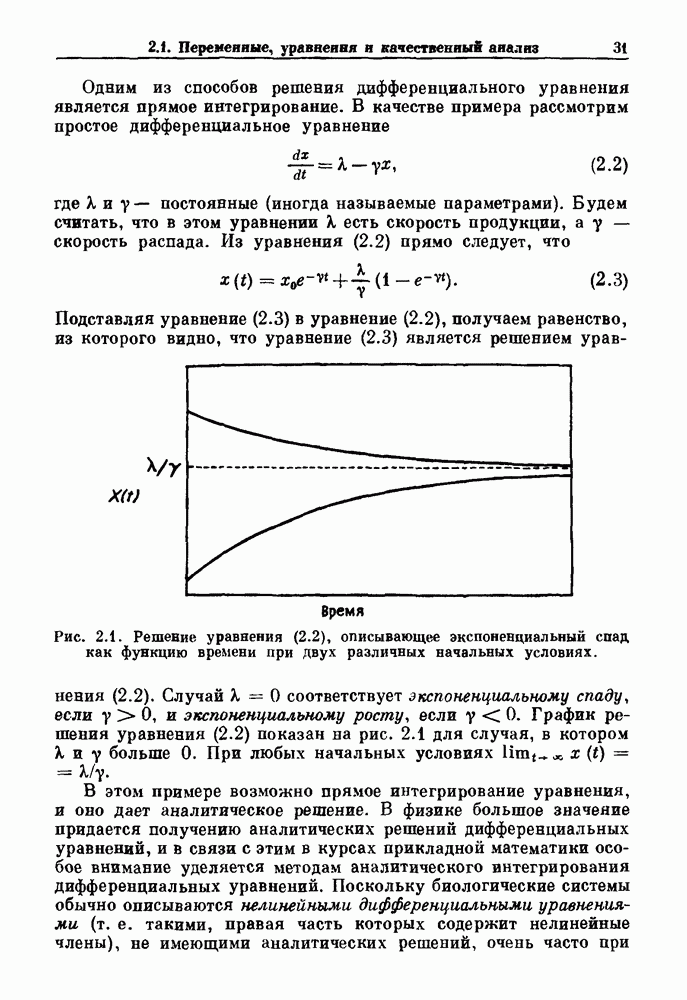
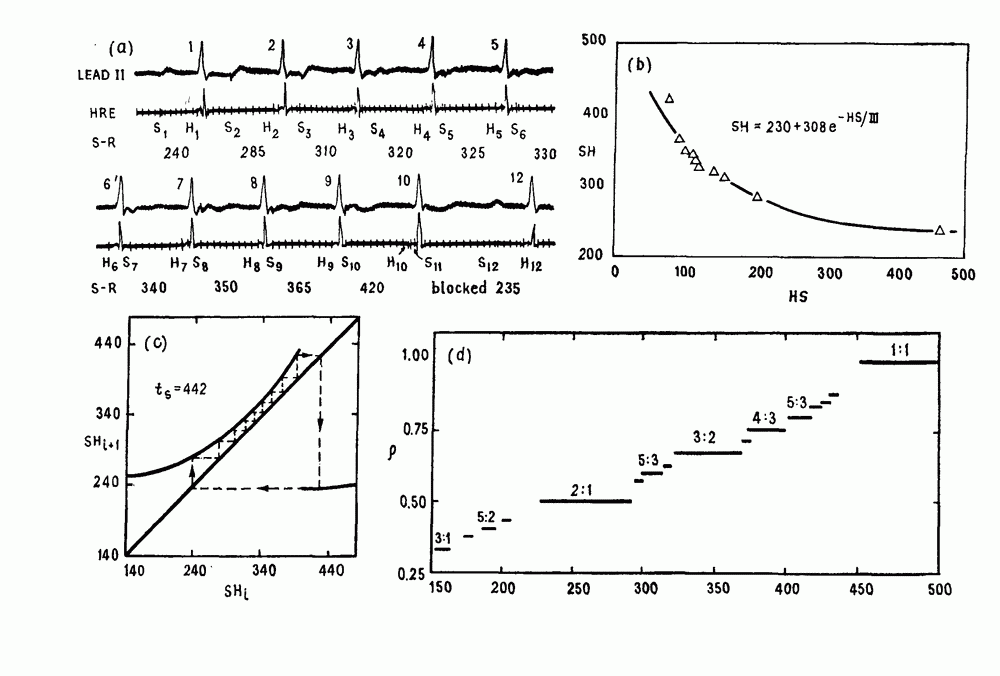
В электрофизиологии сердца функция F изучалась в связи с распространением потенциалов действия через АВ узел. Функция F называется кривой восстановления (recovery curve). В этом случае интервал Чтобы проиллюстрировать использование кривой восстановления в практических ситуациях, рассмотрим следующий случай. (см. скан) Рис. 8.4. (а) Электрокардиографическое отведение II и электрограмма пучка Гиса (НВЕ), записанная у пациента во время стимуляции предсердий при интервале S — S, равном Внутрипредсердный электрод используется для периодической стимуляции сердца человека и записывается активность в пучке Гиса. Интервал между стимулом и началом активности в пучке Гиса обозначается как интервал S—Н. При периоде стимуляции равном
где
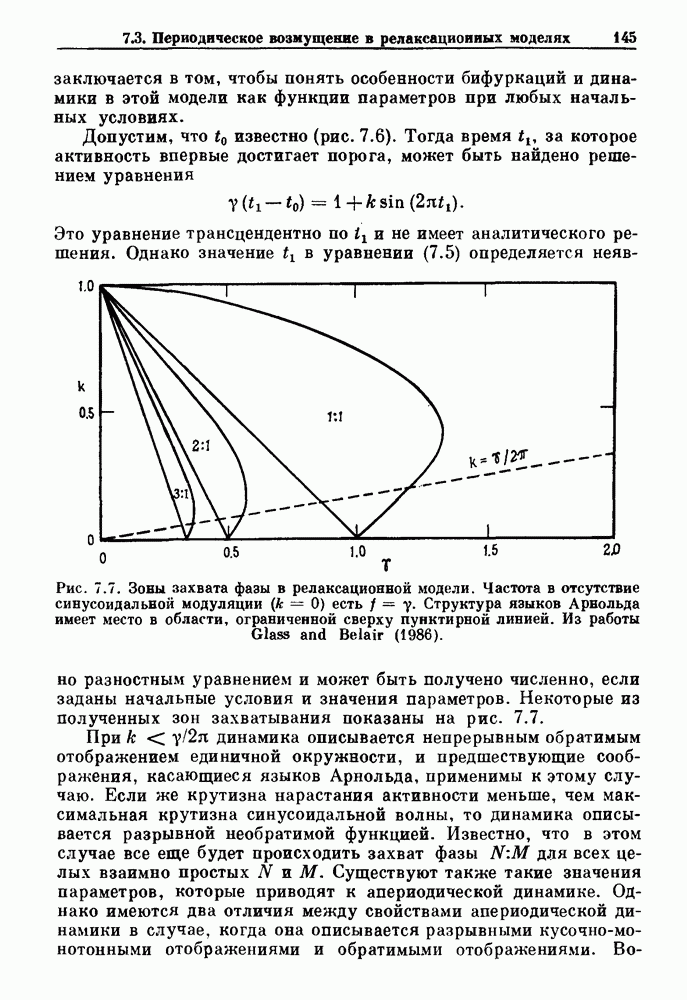
где N есть наименьшее целое число, такое, что Итерируя уравнение (8.4), можно определить динамику для любого значения Результаты, представленные на рис. ритмов Венкебаха и связанных с ними аритмий, основанных на бифуркациях, предсказанных по кривой восстановления. Рассмотрим теперь уравнение (8.1) во втором случае, в котором время
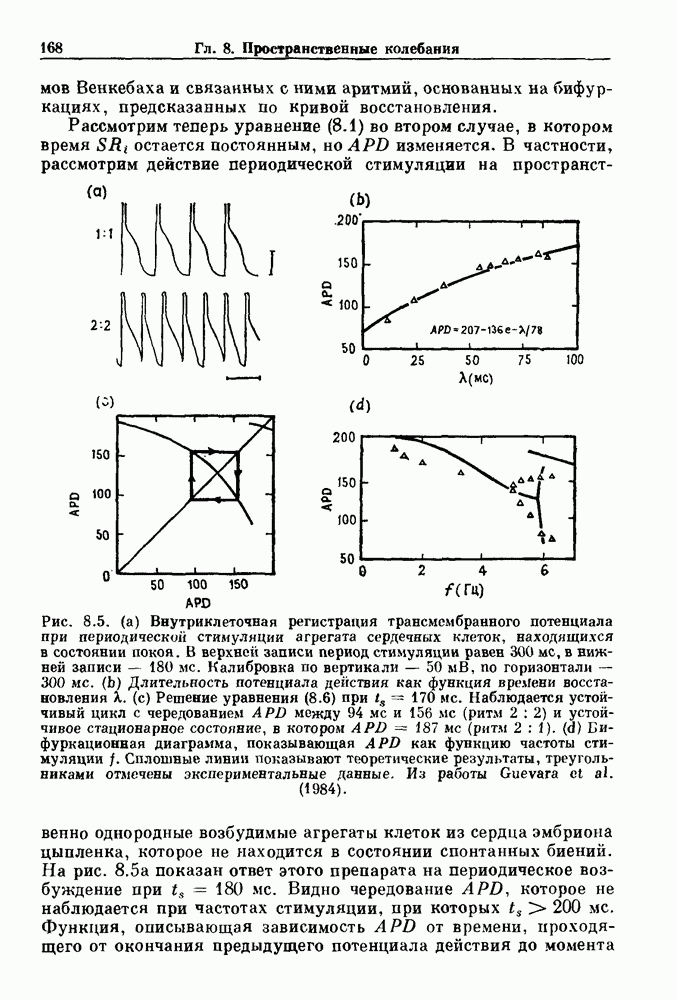
Рис. 8.5. (а) Внутриклеточная регистрация трансмомбранного потенциала при периодической стимуляции агрегата сердечных клеток, находящихся в состоянии покоя. В верхней записи период стимуляции равен На рис. 8.5 а показан ответ этого препарата на периодическое возбуждение при приложения стимула, называется кривой восстановления электрического состояния (electrical restitution curve). Эта кривая может аппроксимироваться экспоненциальной функцией
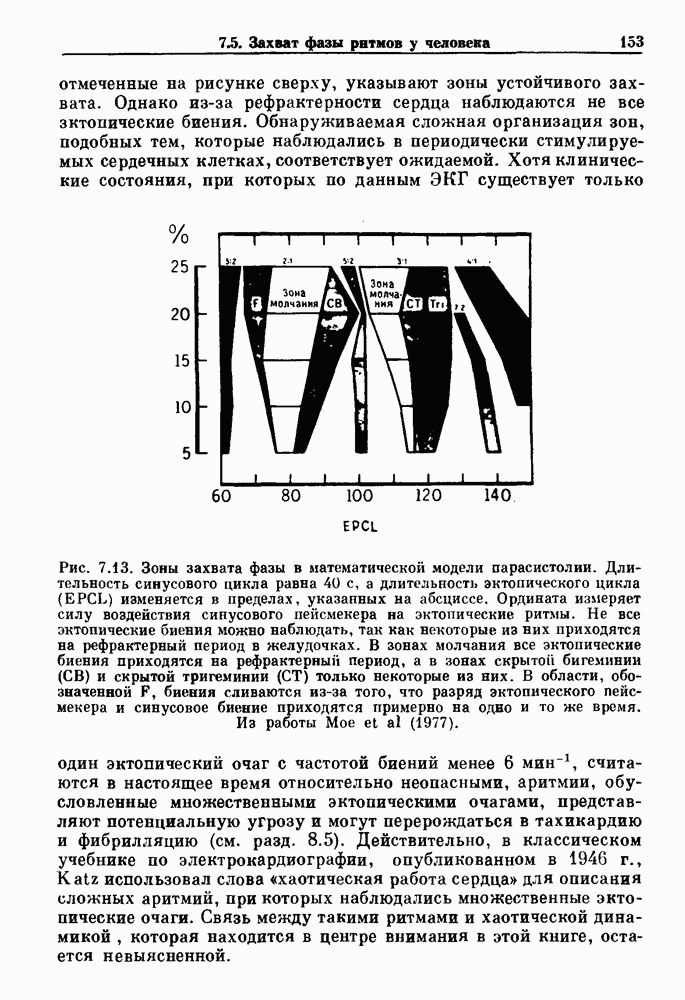
где
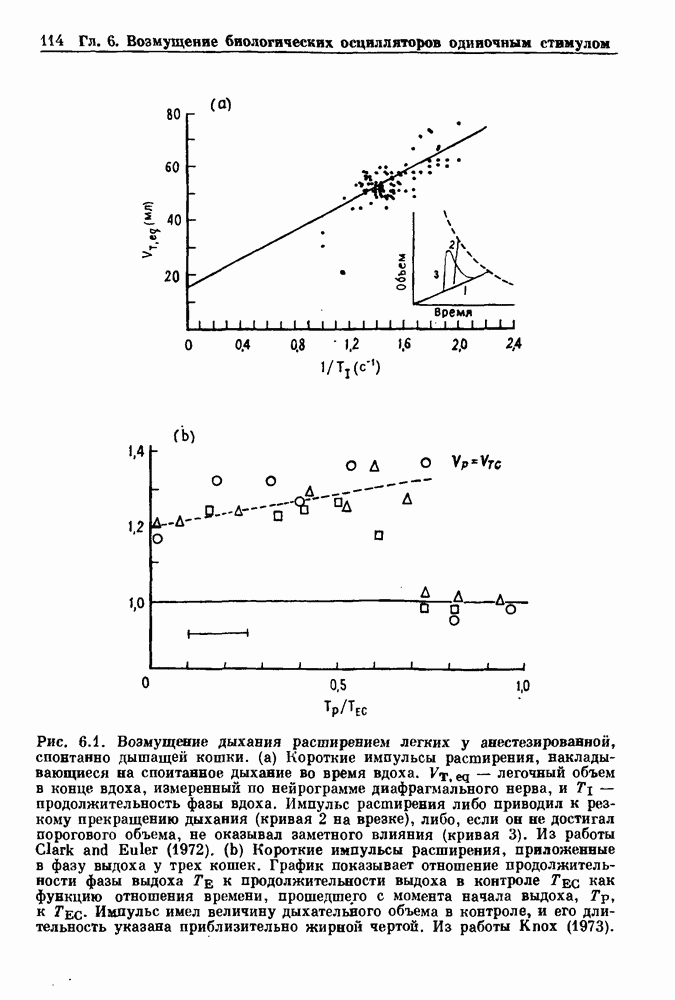
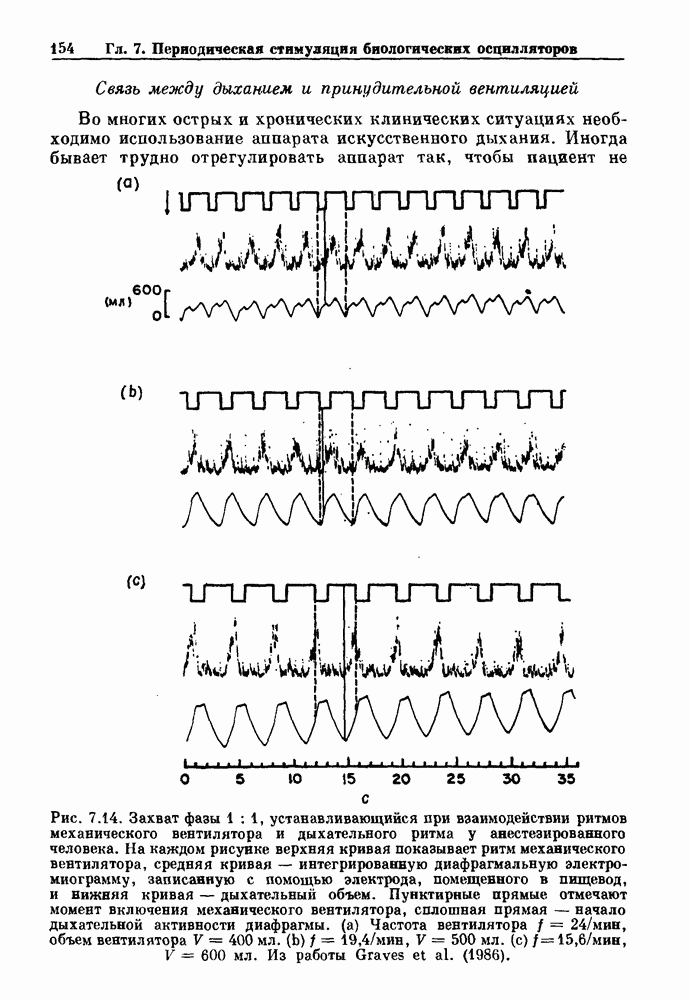
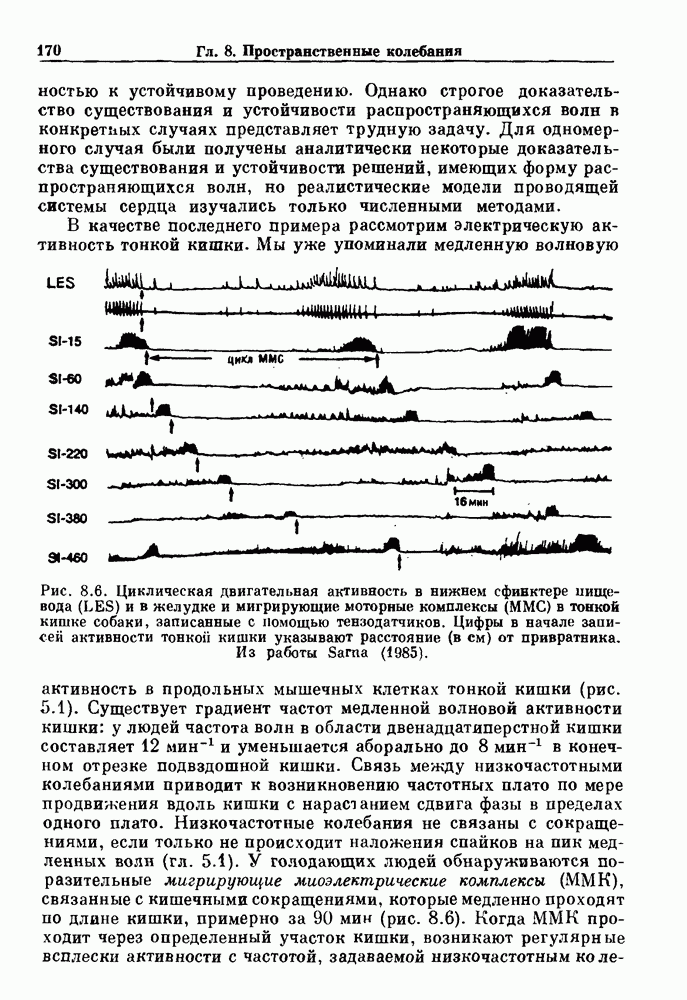
где N — наименьшее целое число, такое, что До сих пор мы рассматривали одномерное распространение возбуждения, которое позволяет использовать разностное уравнение для анализа динамических свойств. Однако, физиологически важные и математически более сложные ситуации также были изучены. Мы кратко прокомментируем две из них — распространение потенциалов действия в нерве и других возбудимых системах и электрическую активность тонкой кишки. Существует большое количество работ по распространению электрической активности в нерве, использующих модель Ходжкина — Хаксли или ее модификации. Так как нервные клетки передают возбуждение с помощью незатухающих потенциалов действия, модели нервных клеток, сформулированные в виде дифференциальных уравнений в частных производных, должны обладать способностью к устойчивому проведению. Однако строгое доказательство существования и устойчивости распространяющихся волн в конкретных случаях представляет трудную задачу. Для одномерного случая были получены аналитически некоторые доказательства существования и устойчивости решений, имеющих форму распространяющихся волн, но реалистические модели проводящей системы сердца изучались только численными методами. В качестве последнего примера рассмотрим электрическую активность тонкой кишки. Мы уже упоминали медленную волновую активность в продольных мышечных клетках тонкой кишки (рис. 5.1).
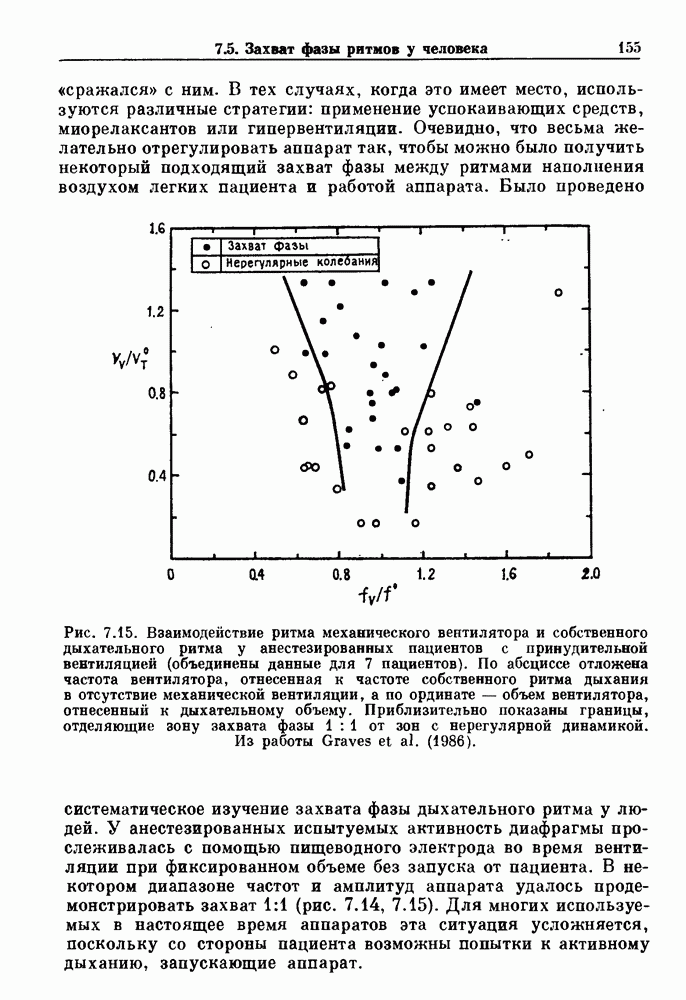
Рис. 8.6. Циклическая двигательная активность в нижнем сфинктере пищевода (LES) и в желудке и мигрирующие моторные комплексы (ММС) в тонкой кишке собаки, записанные с помощью тензодатчиков. Цифры в начале записей активности тонкой кишки указывают расстояние (в см) от привратника. Из работы Sarna (1985). Существует градиент частот медленной волновой активности кишки: у людей частота волн в области двенадцатиперстной кишки составляет Эта активность занимает примерно 20 мин из 90-минутного ММК-цикла. В остальное время ММК-цикла либо не происходит никаких сокращений, либо сокращения имеют нерегулярный характер. Прием еды устраняет ММ К-активность, и возникают два различных типа кишечных сокращений. Перистальтические волны проходят аборально вдоль тонкой кишки на короткие расстояния (1 —4 см) со скоростью, задаваемой кажущейся скоростью медленных волн. Сегментация представляет собой локализованные сокращения, которые не координируются с сокращениями, происходящими выше или ниже. Координация этих различных ритмических активностей является сложной проблемой, которая еще не получила полного объяснения.
|
 |
Оглавление
|















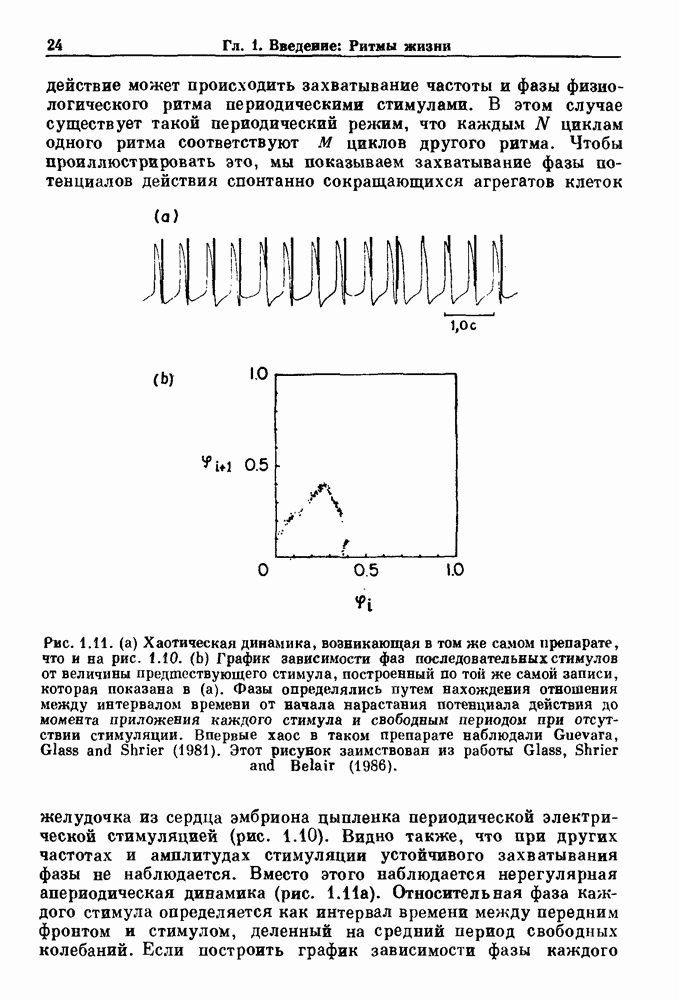






























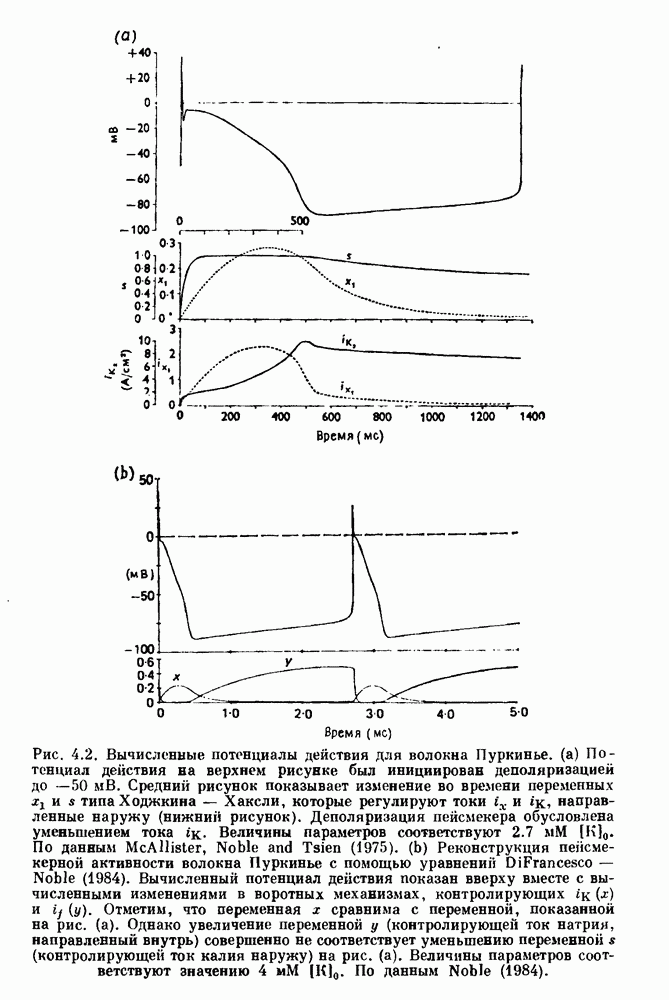




















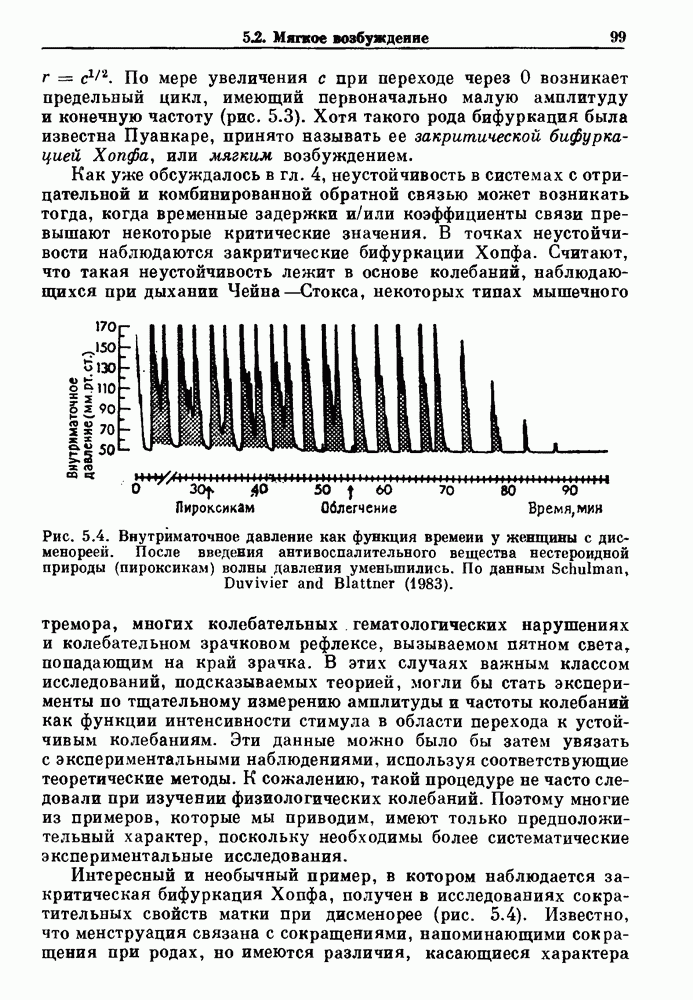


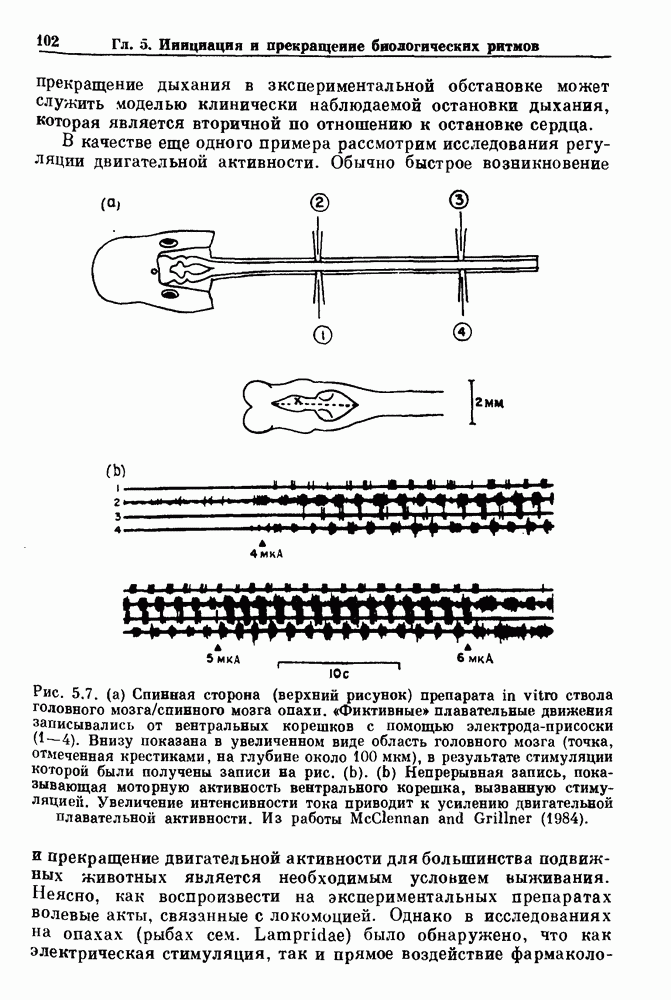














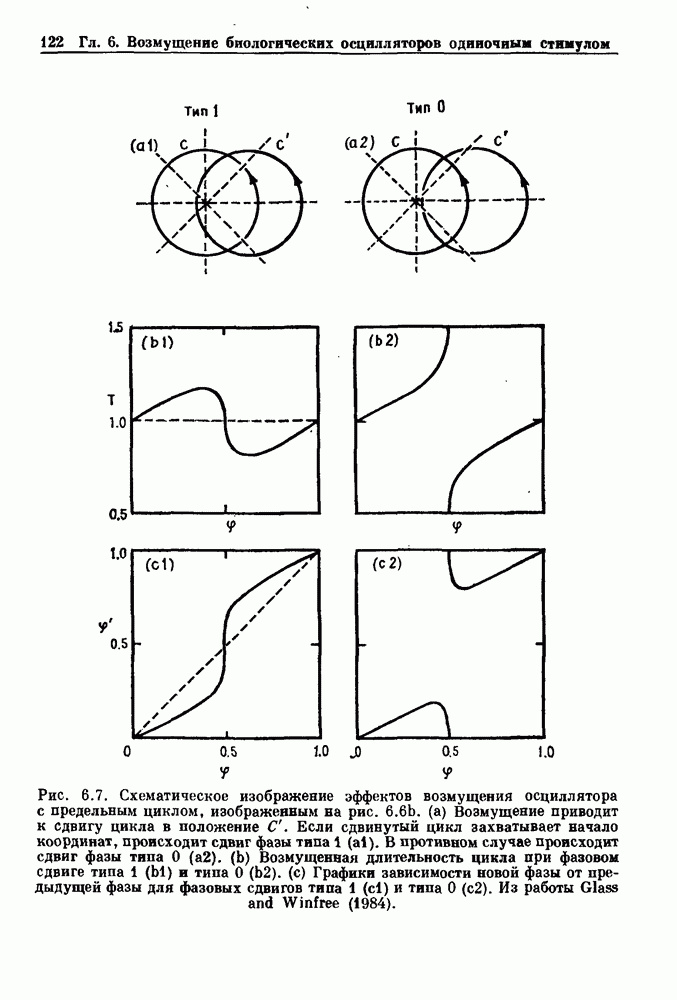


























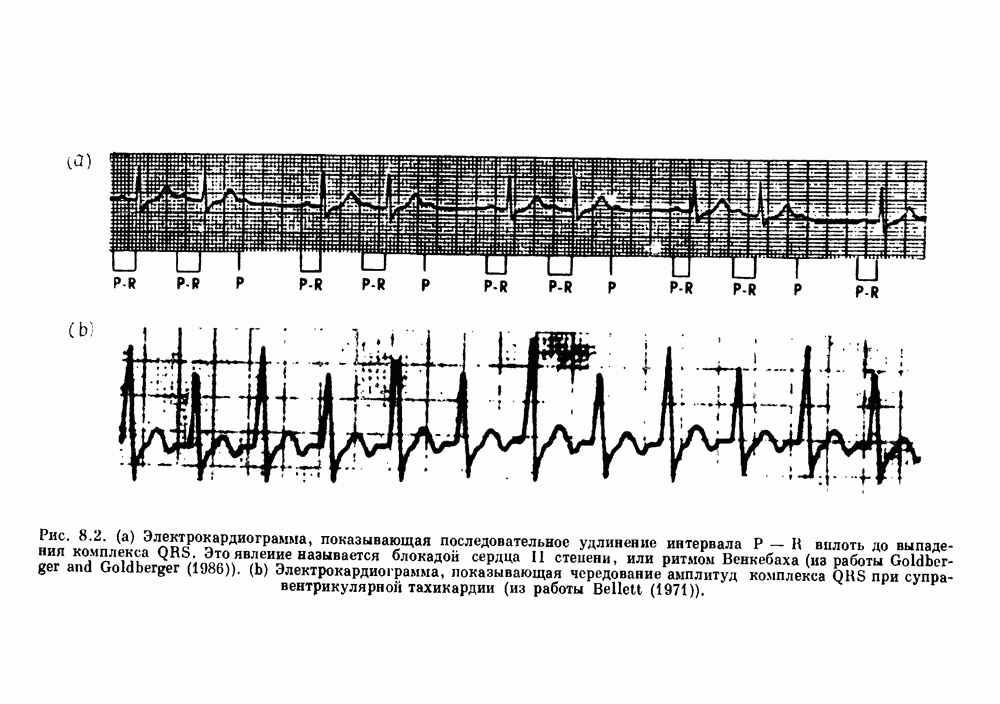










































































 вплоть до выпадения комплекса QRS. Это явление называется блокадой сердца II степени, или ритмом Венкебаха (из работы Goldberger and Goldberger (1986)). (b) Электрокардиограмма, показывающая чередование амплитуд комплекса QKS при суправентрикулярной тахикардии (из работы Bellett (1971)).
вплоть до выпадения комплекса QRS. Это явление называется блокадой сердца II степени, или ритмом Венкебаха (из работы Goldberger and Goldberger (1986)). (b) Электрокардиограмма, показывающая чередование амплитуд комплекса QKS при суправентрикулярной тахикардии (из работы Bellett (1971)).
 При увеличении частоты простое распространение волн равной амплитуды в отношении
При увеличении частоты простое распространение волн равной амплитуды в отношении  к стимулу не наблюдается. Одним из возможных объяснений является то, что каждый стимул по-прежнему приводит к возникновению волны возбуждения, но амплитуды последующих волн изменяются. Другая возможность состоит в том, что некоторые из волн блокируются и что обнаруживаются ритмы вида
к стимулу не наблюдается. Одним из возможных объяснений является то, что каждый стимул по-прежнему приводит к возникновению волны возбуждения, но амплитуды последующих волн изменяются. Другая возможность состоит в том, что некоторые из волн блокируются и что обнаруживаются ритмы вида  при
при  Хотя более сложные типы распространения волн также явно возможны, мы сосредоточим внимание на зтих двух главных типах динамического поведения.
Хотя более сложные типы распространения волн также явно возможны, мы сосредоточим внимание на зтих двух главных типах динамического поведения.
 . Длительность потенциала действия
. Длительность потенциала действия
 обозначается APD. Стимул и результирующая волна возбуждения часто измеряются в различных участках изучаемой системы. Хотя в реальных условиях
обозначается APD. Стимул и результирующая волна возбуждения часто измеряются в различных участках изучаемой системы. Хотя в реальных условиях  могут зависеть от состояния, предшествовавшего стимуляции, мы допускаем, что длительность этих интервалов является функцией времени от окончания предшествующего возбуждения до момента приложения стимула
могут зависеть от состояния, предшествовавшего стимуляции, мы допускаем, что длительность этих интервалов является функцией времени от окончания предшествующего возбуждения до момента приложения стимула 

 между последовательными стимулами.
между последовательными стимулами.  обозначает интервал от
обозначает интервал от  стимула до начала следующего потенциала действия, a APD — длительность
стимула до начала следующего потенциала действия, a APD — длительность  потенциала действия.
потенциала действия.
 имеем
имеем


 соответствует времени от начала зубца Р на ЭКГ до начала комплекса QRS (рис. 1.2). С начала этого века было принято считать, что интервал
соответствует времени от начала зубца Р на ЭКГ до начала комплекса QRS (рис. 1.2). С начала этого века было принято считать, что интервал  есть убывающая функция предшествующего интервала
есть убывающая функция предшествующего интервала  и что АВ-кривая восстановления может использоваться для вычисления эффектов периодической стимуляции как функции частоты периодических стимулов.
и что АВ-кривая восстановления может использоваться для вычисления эффектов периодической стимуляции как функции частоты периодических стимулов.
 обозначает артефакт стимуляции, Н — отклонения, обусловленные активностью пучка Гиса. Интервал времени от каждого стимула до следующего пика активности пучка Гиса указан в мс. (Из работы Levy et al. (1974а).) (b) Решение уравнения (8.4) при
обозначает артефакт стимуляции, Н — отклонения, обусловленные активностью пучка Гиса. Интервал времени от каждого стимула до следующего пика активности пучка Гиса указан в мс. (Из работы Levy et al. (1974а).) (b) Решение уравнения (8.4) при  Показан цикл, соответствующий А В блокаде
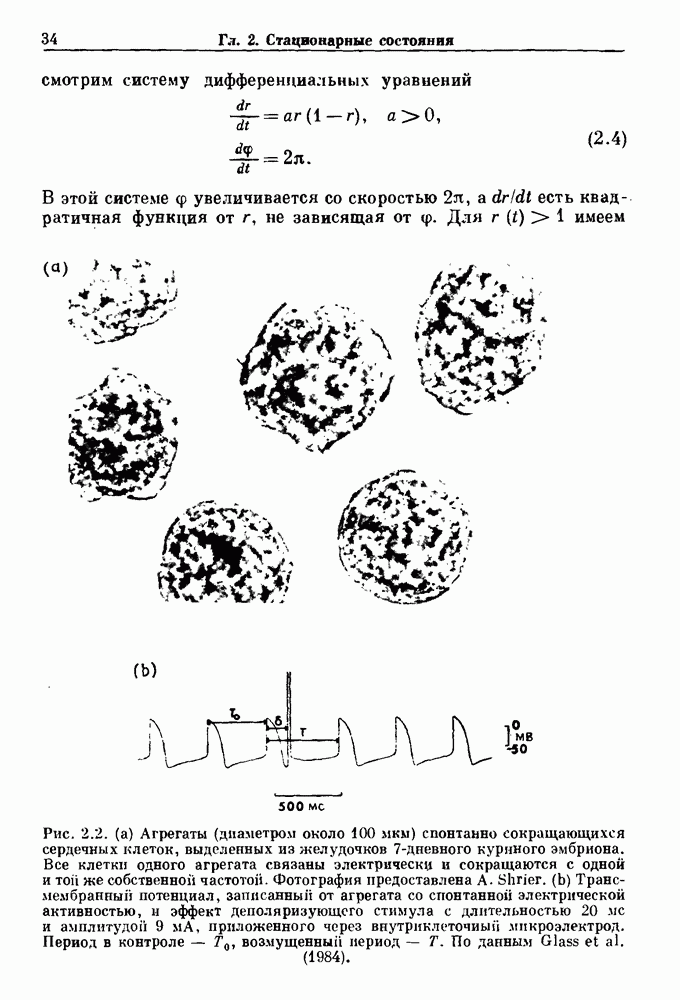
Показан цикл, соответствующий А В блокаде  Коэффициент проведения
Коэффициент проведения  (число сокращений желудочков, отнесенное к числу стимулов) как функция
(число сокращений желудочков, отнесенное к числу стимулов) как функция  вычислялся из уравнения (8.4) для значений параметров, указанных в (b). Эта кривая называется функцией Кантора (Mandelbrot (1977,1982)). Из работы Glass, Guevara and Shrier (1987a).
вычислялся из уравнения (8.4) для значений параметров, указанных в (b). Эта кривая называется функцией Кантора (Mandelbrot (1977,1982)). Из работы Glass, Guevara and Shrier (1987a).
 возникает блокада Венкебаха, при которой на 11 сокращений предсердий приходятся 10 сокращений желудочков (11:10 АВ-блокада) (рис. 8.4 а). Последовательные измерения интервала S—Н как функции его предшествующего значения показаны на рис. 8.4. Данные могут аппроксимироваться экспоненциальной функцией
возникает блокада Венкебаха, при которой на 11 сокращений предсердий приходятся 10 сокращений желудочков (11:10 АВ-блокада) (рис. 8.4 а). Последовательные измерения интервала S—Н как функции его предшествующего значения показаны на рис. 8.4. Данные могут аппроксимироваться экспоненциальной функцией
 (8.3)
(8.3)
 Используя кривую восстановления уравнения (8.3), мы получаем
Используя кривую восстановления уравнения (8.3), мы получаем
 (8.4)
(8.4)

 Графическая итерация для
Графическая итерация для  дает ритм Венкебаха
дает ритм Венкебаха  (рис. 8.4с). На рис.
(рис. 8.4с). На рис.  показано отношение между числом проводимых биений и числом стимулов,
показано отношение между числом проводимых биений и числом стимулов,  , как функция
, как функция  Рисунок представляет собой график функции Кантора. Поскольку число шагов, в основном невообразимо малого размера, бесконечно, график функции Кантора иногда называют дьявольской лестницей.
Рисунок представляет собой график функции Кантора. Поскольку число шагов, в основном невообразимо малого размера, бесконечно, график функции Кантора иногда называют дьявольской лестницей.
 имеют весьма общий характер. Для любой монотонно убывающей кривой восстановления, подобной той, которая показана на рис.
имеют весьма общий характер. Для любой монотонно убывающей кривой восстановления, подобной той, которая показана на рис.  проведение будет описываться функцией Кантора. Таким образом, мы наблюдаем классические ритмы Венкебаха вида N:
проведение будет описываться функцией Кантора. Таким образом, мы наблюдаем классические ритмы Венкебаха вида N:  также более сложные их варианты. Таким образом, итерация разностного уравнения служит для общего представления и классификации
также более сложные их варианты. Таким образом, итерация разностного уравнения служит для общего представления и классификации
 остается постоянным, но APD изменяется. В частности, рассмотрим действие периодической стимуляции на пространственно однородные возбудимые агрегаты клеток из сердца эмбриона цыпленка, которое не находится в состоянии спонтанных биений.
остается постоянным, но APD изменяется. В частности, рассмотрим действие периодической стимуляции на пространственно однородные возбудимые агрегаты клеток из сердца эмбриона цыпленка, которое не находится в состоянии спонтанных биений.

 в нижней записи —
в нижней записи —  Калибровка по вертикали —
Калибровка по вертикали —  по горизонтали — 300 мс. (b) Длительность потенциала действия как функция времени восстановления X.
по горизонтали — 300 мс. (b) Длительность потенциала действия как функция времени восстановления X.  Решение уравнения (8.6) при
Решение уравнения (8.6) при  Наблюдается устойчивый цикл с чередованием APD между
Наблюдается устойчивый цикл с чередованием APD между  (ритм
(ритм  и устойчивое стационарное состояние, в котором
и устойчивое стационарное состояние, в котором  (ритм 2 : 1). (d) Бифуркационная диаграмма, показывающая APD как функцию частоты стимуляции
(ритм 2 : 1). (d) Бифуркационная диаграмма, показывающая APD как функцию частоты стимуляции  Сплошные линии показывают теоретические результаты, треугольниками отмечены экспериментальные данные. Из работы Guevara
Сплошные линии показывают теоретические результаты, треугольниками отмечены экспериментальные данные. Из работы Guevara 
 Видно чередование APD, которое не наблюдается при частотах стимуляции, при которых
Видно чередование APD, которое не наблюдается при частотах стимуляции, при которых  Функция, описывающая зависимость APD от времени, проходящего от окончания предыдущего потенциала действия до момента
Функция, описывающая зависимость APD от времени, проходящего от окончания предыдущего потенциала действия до момента

 — максимальная
— максимальная  — время, прошедшее от окончания предшествующего потенциала действия до стимула, а
— время, прошедшее от окончания предшествующего потенциала действия до стимула, а  — положительные константы. График кривой восстановления электрического состояния показан на рис.
— положительные константы. График кривой восстановления электрического состояния показан на рис.  Подстановка уравнен
Подстановка уравнен  в уравнение (8.1) дает
в уравнение (8.1) дает

 В уравнении (8.6) возникает бифуркация удвоения периода при условии, что
В уравнении (8.6) возникает бифуркация удвоения периода при условии, что  при критической частоте стимуляции
при критической частоте стимуляции  где
где  ). При критической частоте стимуляции
). При критической частоте стимуляции  и время восстановления равно
и время восстановления равно  Таким образом, в этой экспериментальной системе альтернанс идентифицируется с бифуркацией удвоения периода. Графическое решение уравнения (8.6) показано на рис.
Таким образом, в этой экспериментальной системе альтернанс идентифицируется с бифуркацией удвоения периода. Графическое решение уравнения (8.6) показано на рис.  для
для  При таком периоде стимуляции возможны два типа устойчивого асимптотического поведения, соответствующие альтернансу, а также ритмы вида
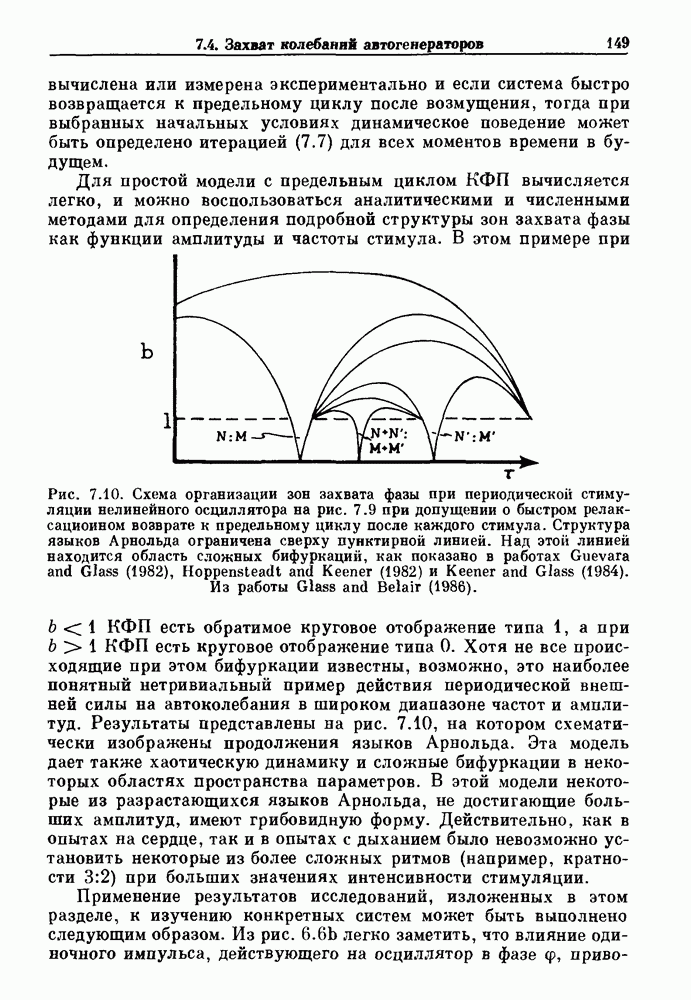
При таком периоде стимуляции возможны два типа устойчивого асимптотического поведения, соответствующие альтернансу, а также ритмы вида  Вычисленные значения APD как функции частоты стимуляции представлены на рис.
Вычисленные значения APD как функции частоты стимуляции представлены на рис.  Теоретические расчеты указывают на появление альтернанса, но не дают точного значения APD для диапазона частот стимуляции, в котором альгернанс наблюдался экспериментально. Теория предсказывает бистабильность для динамических состояний 1:1 и 2 : 1 и динамических состояний 2 : 2 и 2:1. Бистабильность в проведении через АВ-узел наблюдалась Майнсом, но нам неизвестны другие случаи этого явления в АВ проведении, хотя оно наблюдалось при периодической стимуляции спонтанно пульсирующих агрегатов клеток сердца куриного эмбриона.
Теоретические расчеты указывают на появление альтернанса, но не дают точного значения APD для диапазона частот стимуляции, в котором альгернанс наблюдался экспериментально. Теория предсказывает бистабильность для динамических состояний 1:1 и 2 : 1 и динамических состояний 2 : 2 и 2:1. Бистабильность в проведении через АВ-узел наблюдалась Майнсом, но нам неизвестны другие случаи этого явления в АВ проведении, хотя оно наблюдалось при периодической стимуляции спонтанно пульсирующих агрегатов клеток сердца куриного эмбриона.

 и уменьшается аборально до
и уменьшается аборально до  в конечном отрезке подвздошной кишки. Связь между низкочастотными колебаниями приводит к возникновению частотных плато по мере продвижения вдоль кишки с нарастанием сдвига фазы в пределах одного плато. Низкочастотные колебания не связаны с сокращениями, если только не происходит наложения спайков на пик медленных волн (гл. 5.1). У голодающих людей обнаруживаются поразительные мигрирующие миоэлектрические комплексы (ММК), связанные с кишечными сокращениями, которые медленно проходят по длине кишки, примерно за 90 мин (рис. 8.6). Когда ММК проходит через определенный участок кишки, возникают регулярные всплески активности с частотой, задаваемой низкочастотным колебанием.
в конечном отрезке подвздошной кишки. Связь между низкочастотными колебаниями приводит к возникновению частотных плато по мере продвижения вдоль кишки с нарастанием сдвига фазы в пределах одного плато. Низкочастотные колебания не связаны с сокращениями, если только не происходит наложения спайков на пик медленных волн (гл. 5.1). У голодающих людей обнаруживаются поразительные мигрирующие миоэлектрические комплексы (ММК), связанные с кишечными сокращениями, которые медленно проходят по длине кишки, примерно за 90 мин (рис. 8.6). Когда ММК проходит через определенный участок кишки, возникают регулярные всплески активности с частотой, задаваемой низкочастотным колебанием.