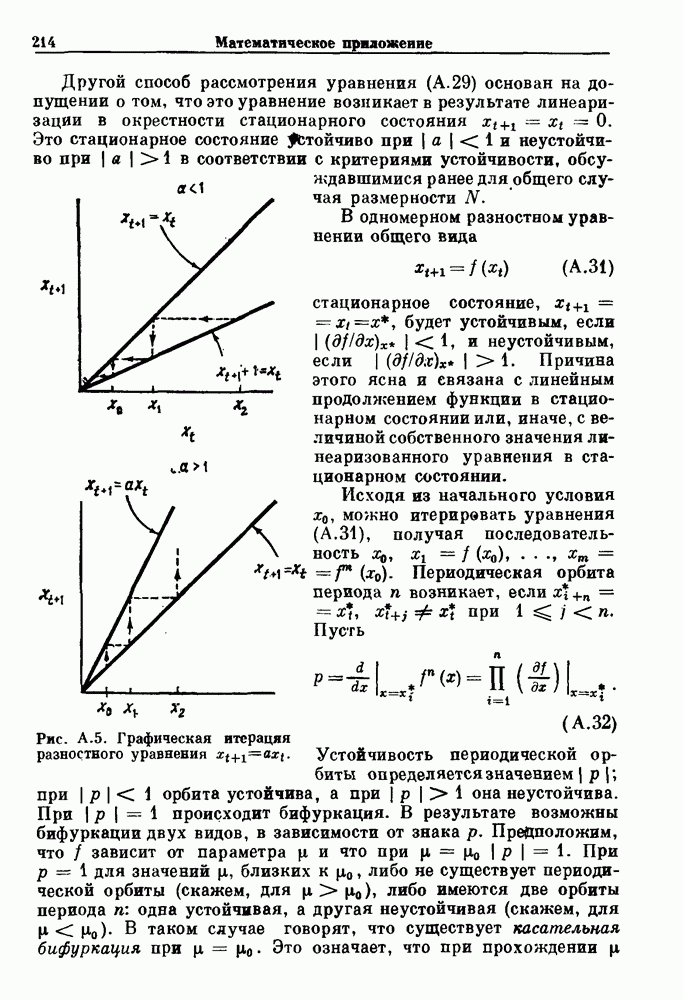
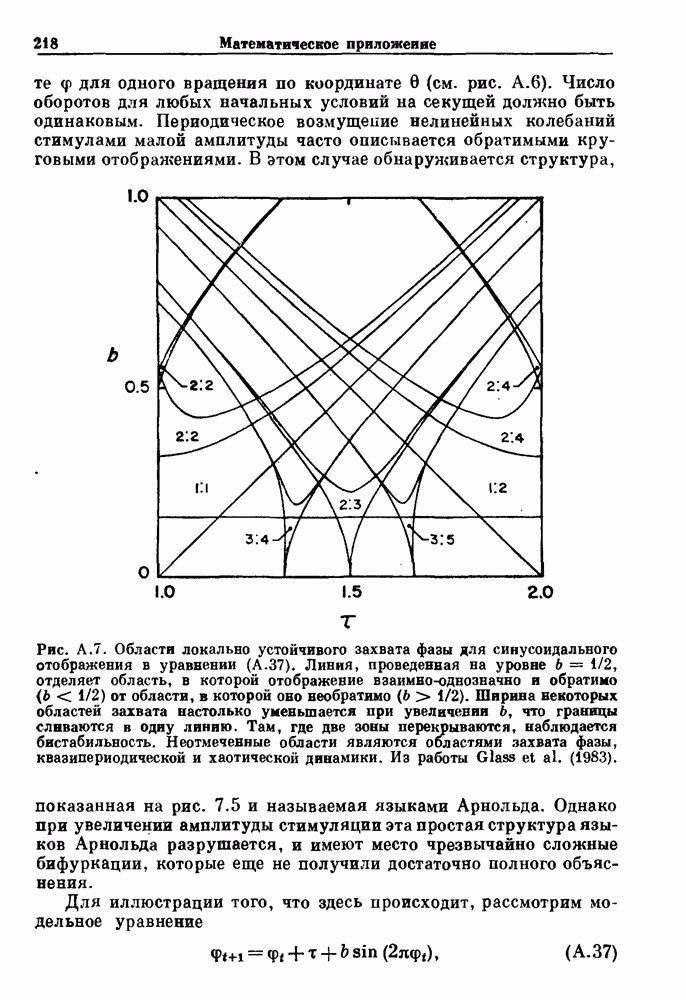
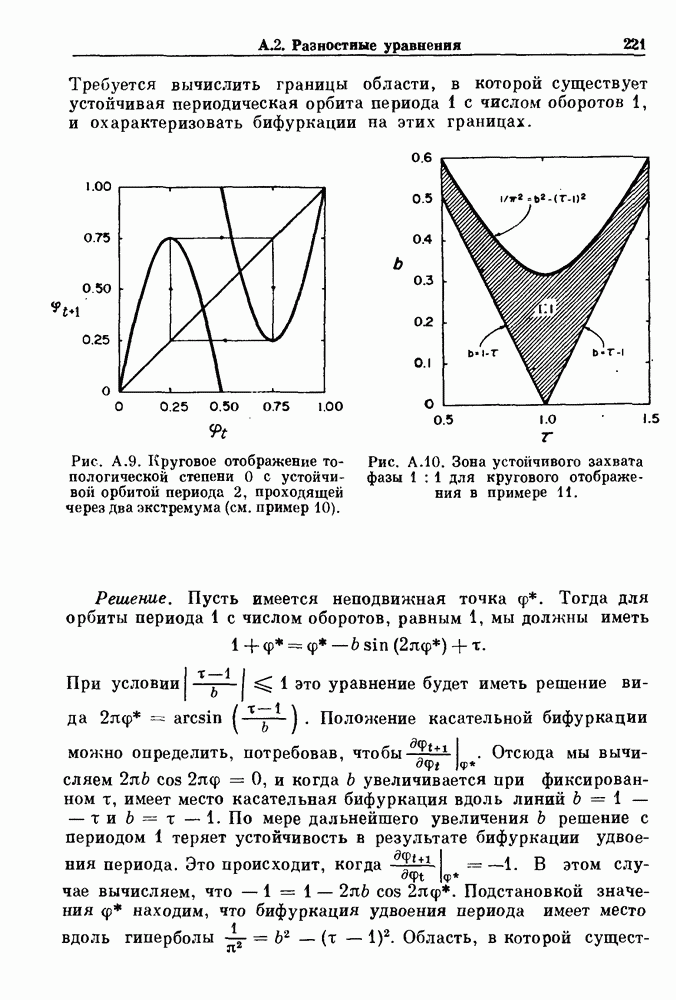
Пред.
След.
Макеты страниц
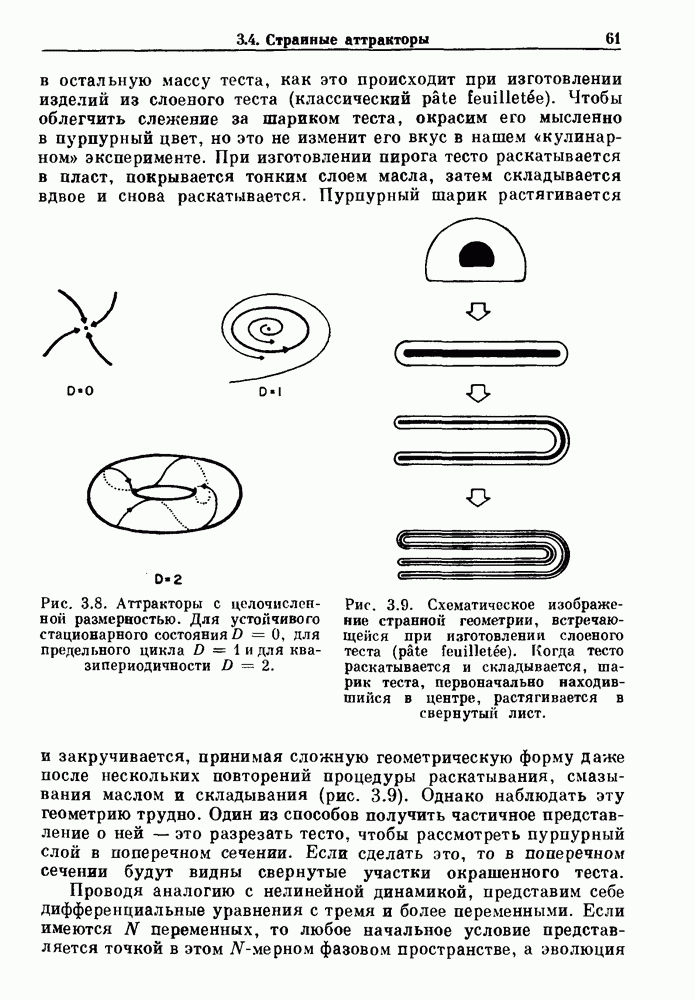
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
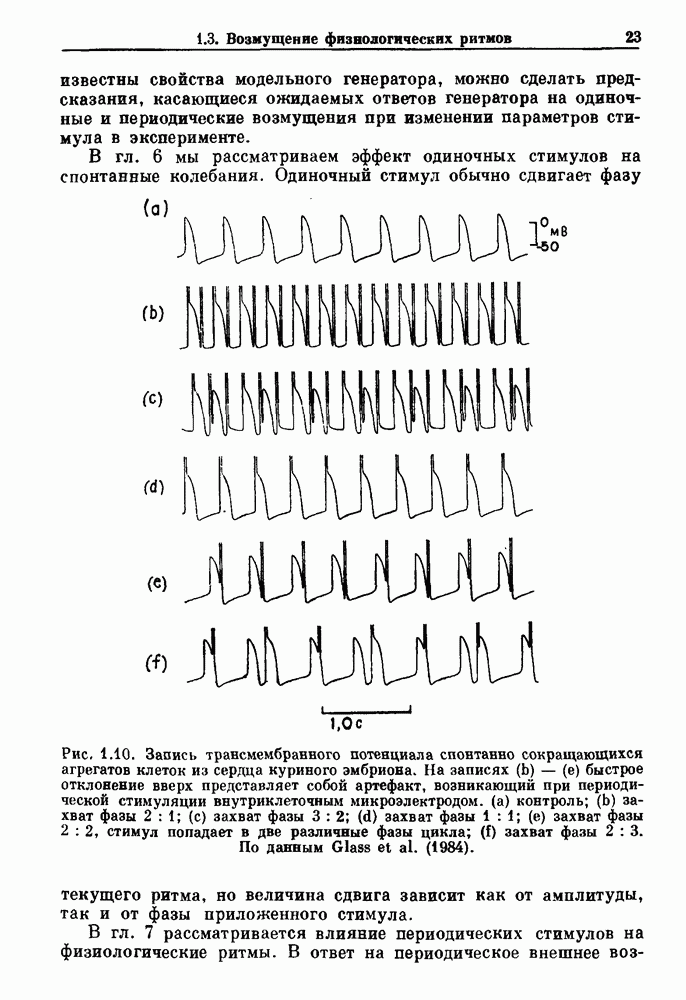
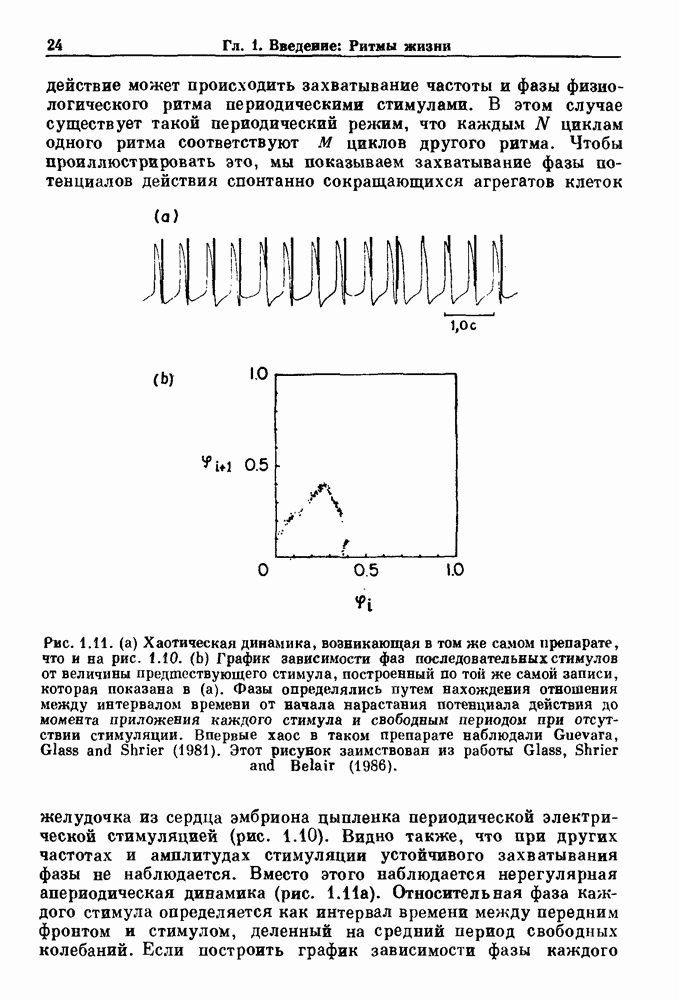
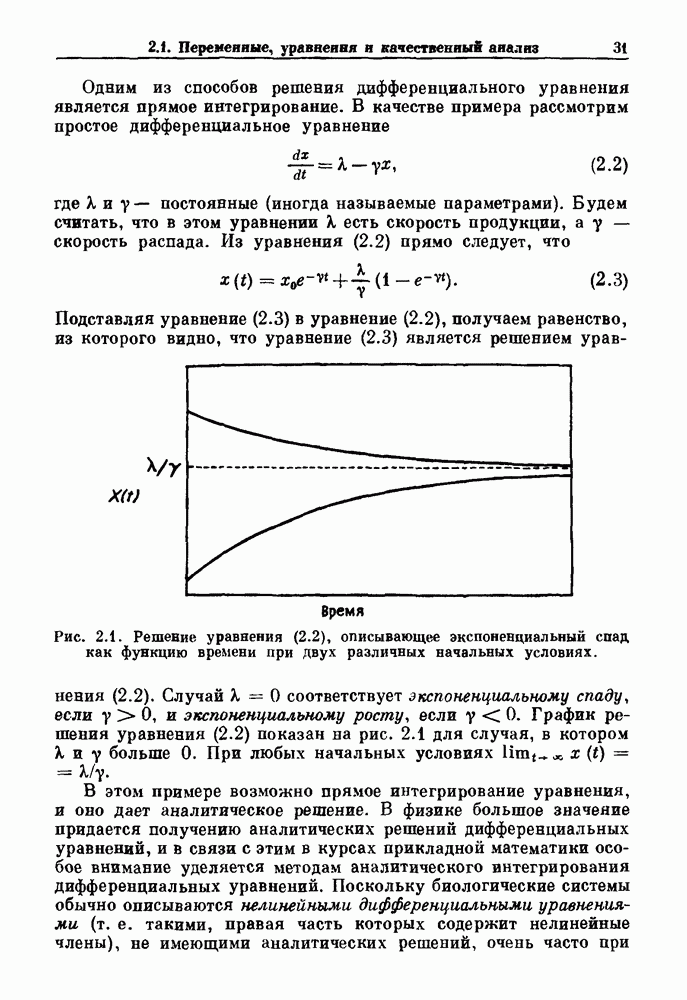
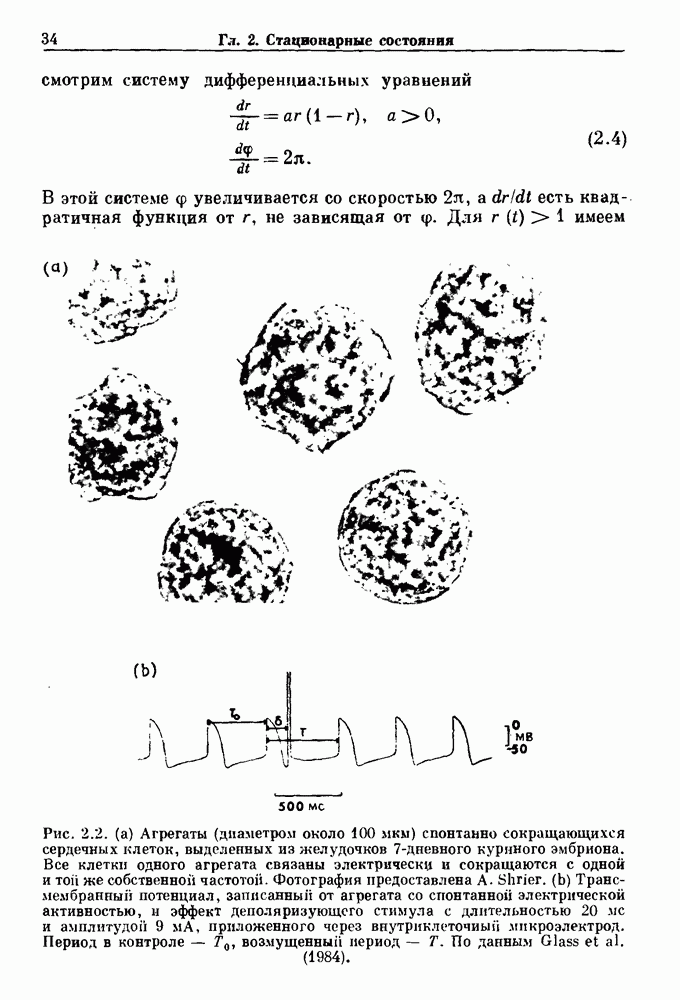
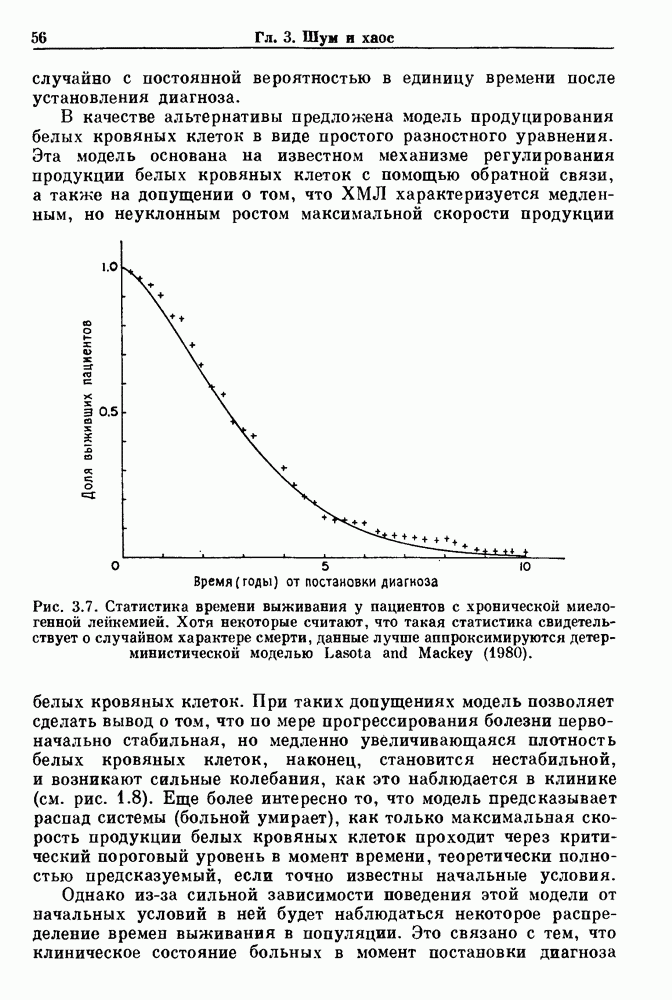
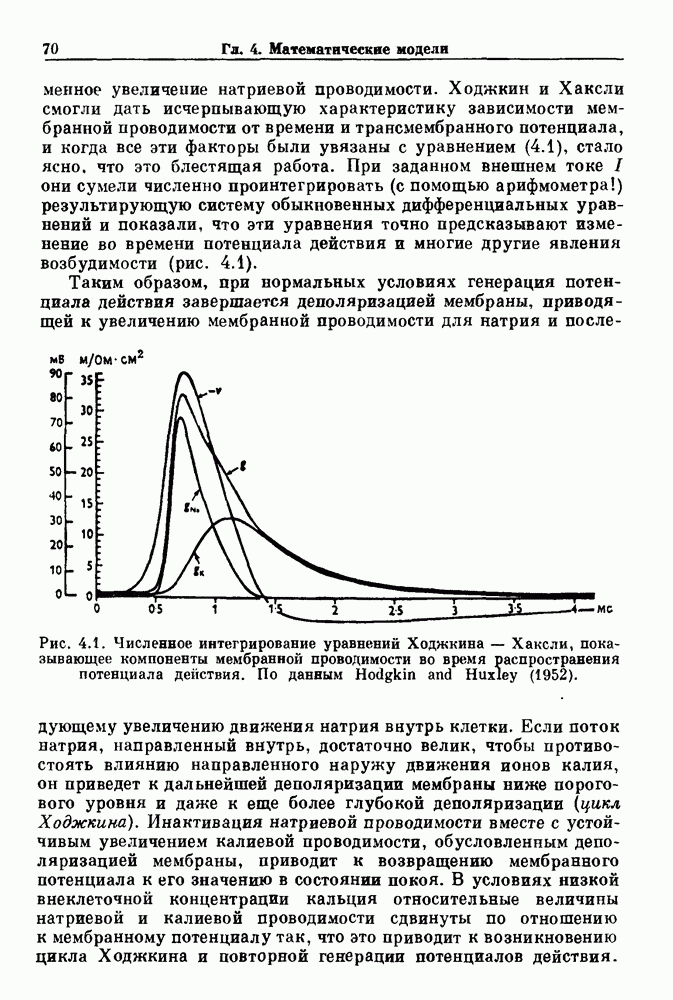
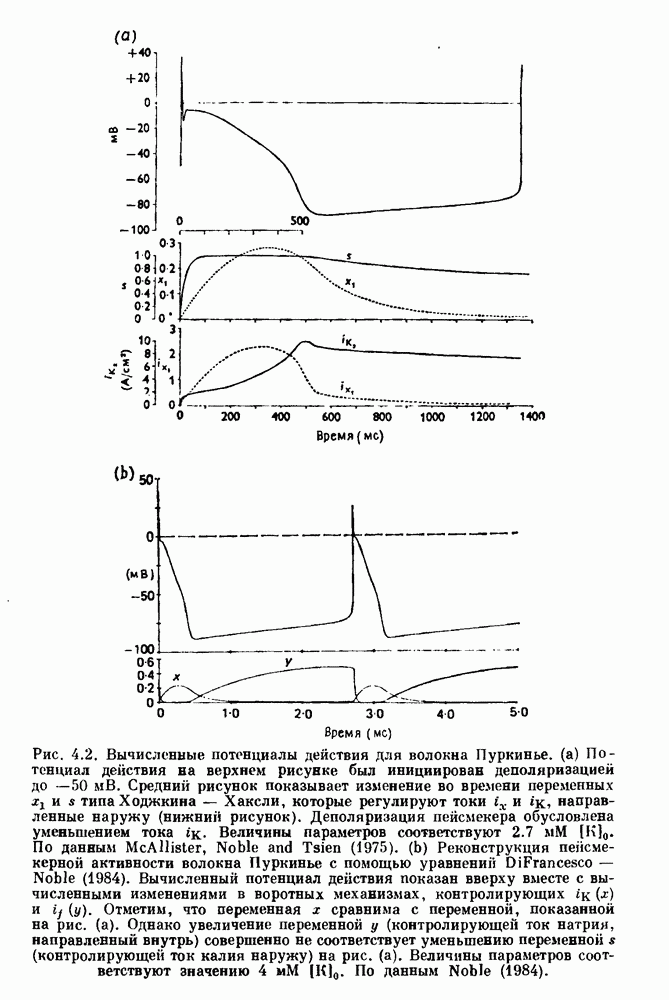
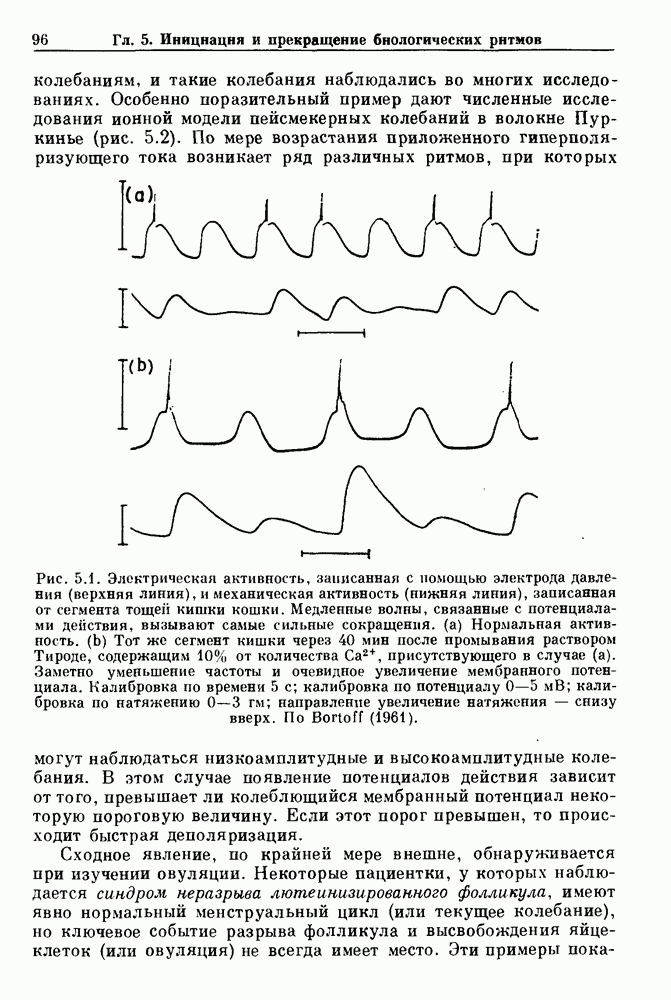
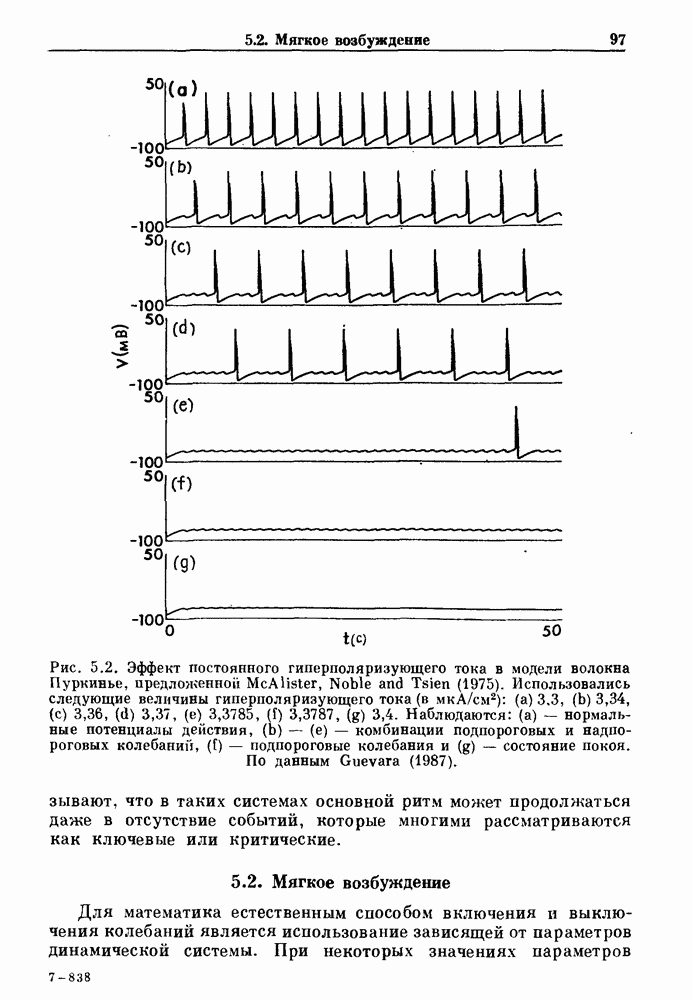
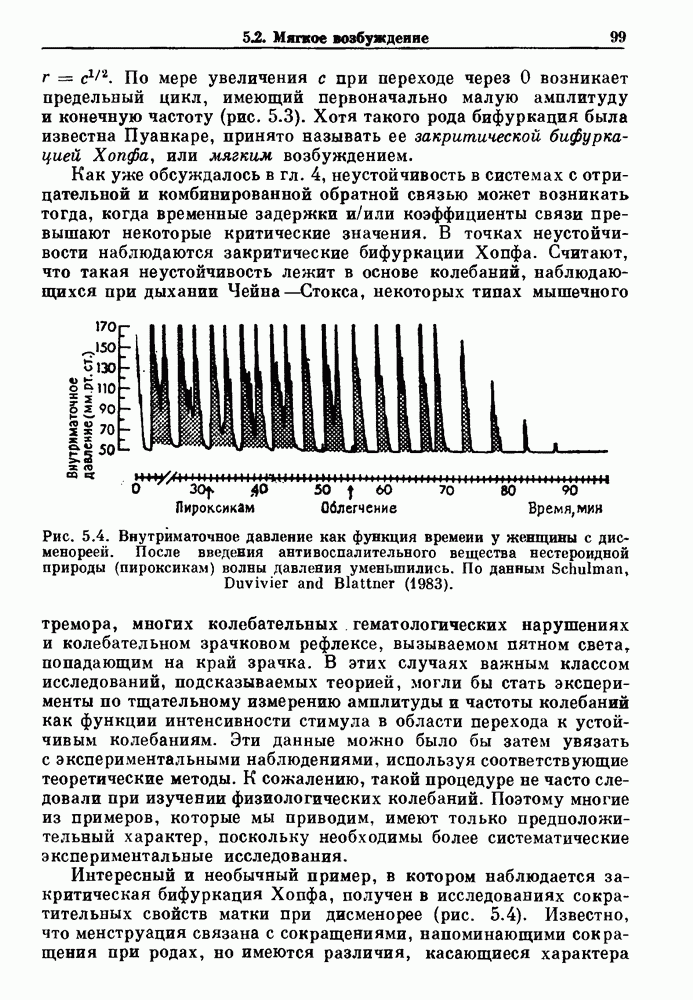
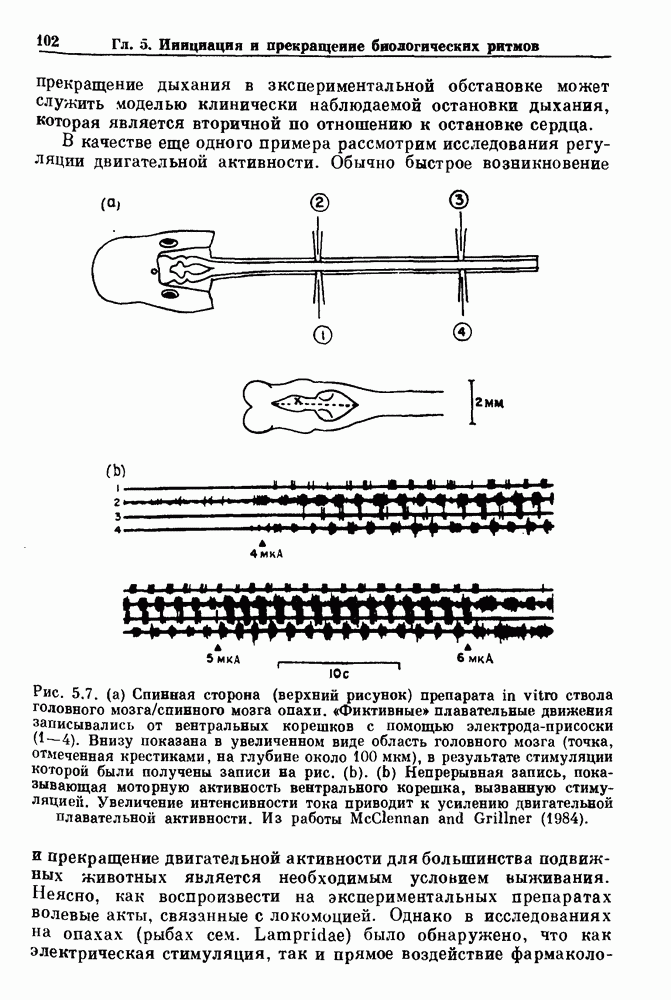
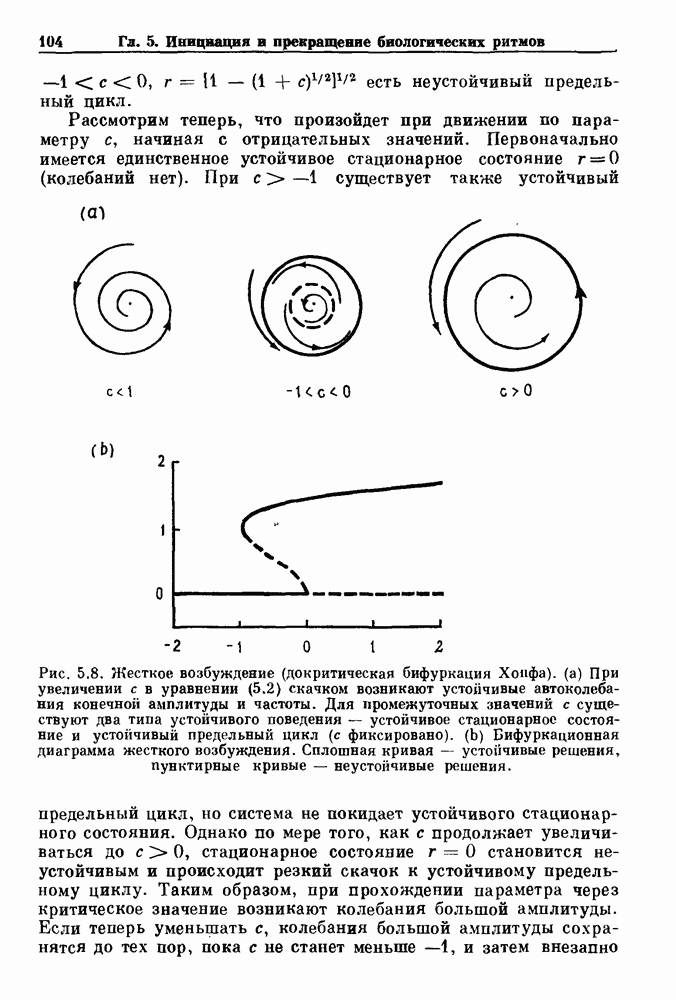
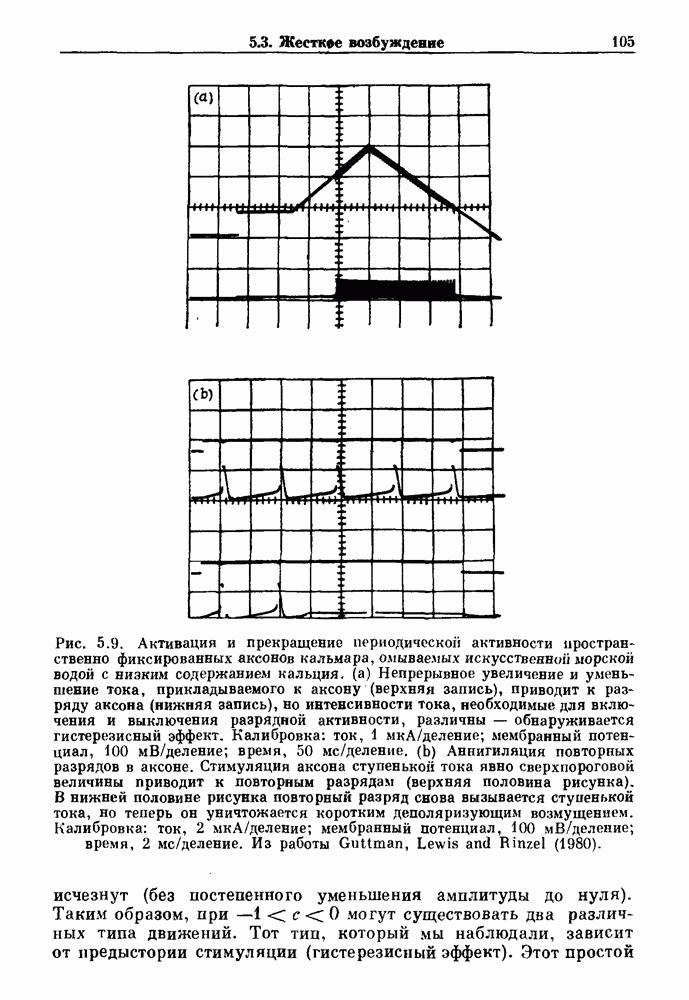
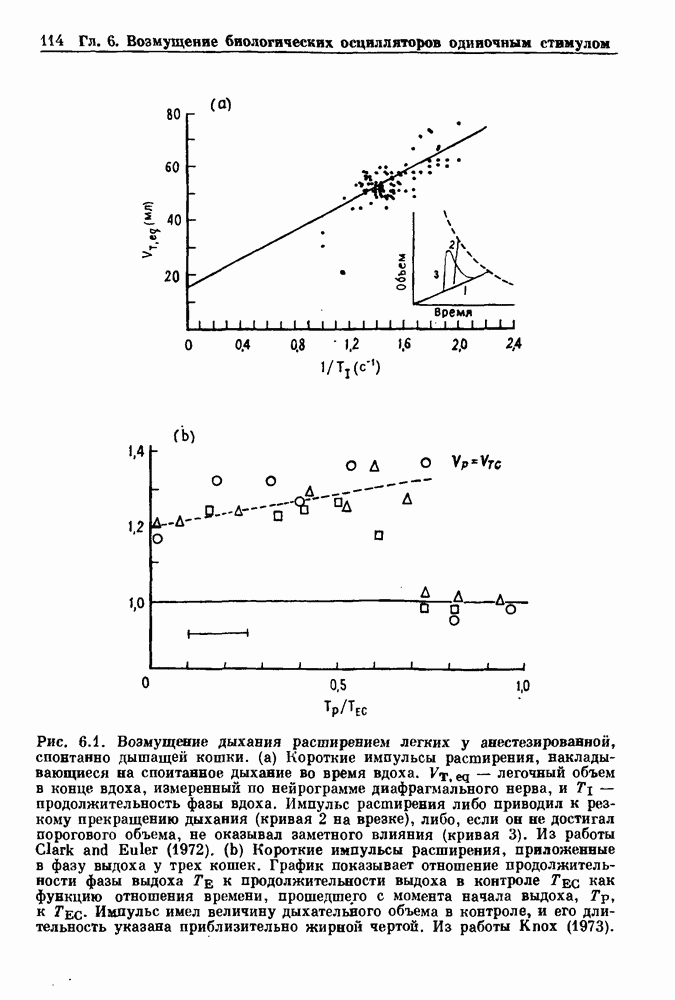
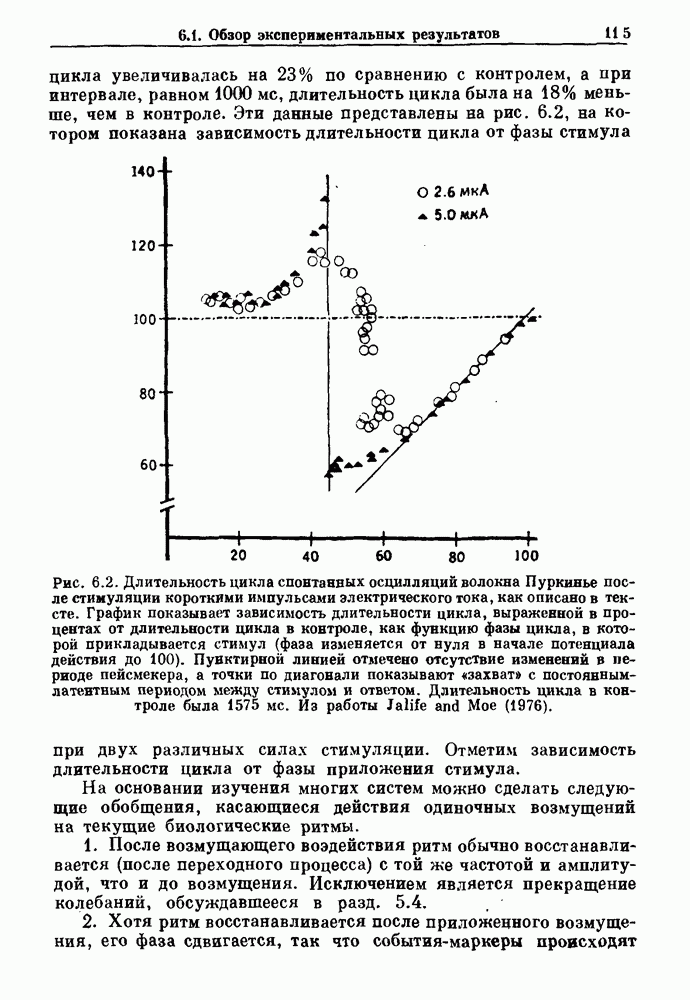
Глава 5. Инициация и прекращение биологических ритмовПереходы между колебательными и неколебательными состояниями обычны для физиологических систем. В этой главе мы обсуждаем экспериментально наблюдаемые переходы между колебательным и неколебательным динамическим поведением и предлагаем гипотезы для объяснения таких переходов. В разд. 5.1 мы показываем, что текущий ритм может постоянно присутствовать, но при этом его регистрируемые проявления могут иметь прерывистый характер. Два различных пути перехода от колебательных к неколебательным состояниям, известных в теории бифуркаций как мягкое и жесткое возбуждение, обсуждаются соответственно в разд. 5.2 и 5.3. В случае жесткого возбуждения для данного набора параметров могут быть два устойчивых динамических состояния — одно колебательное, а другое неколебательное. В зтом случае возможна аннигиляция колебаний при действии одиночного стимула. Наблюдение такой аннигиляции в экспериментальных системах обсуждается в разд. 5.4. 5.1. Вовлечение в текущие колебанияВозможное объяснение инициации и прекращения биологических ритмов заключается в том, что основной ритм поддерживается непрерывно, но организм может входить в этот ритм или выходить из него путем изменения регулирующего параметра. Примером могут служить электронные часы, которые всегда идут, но их показания могут быть либо включены, либо выключены. Низкоамплитудные колебания мембранного потенциала, иногда с накладывающимися на них потенциалами действия, наблюдались в различных системах. Важной физиологической системой, в которой обычно обнаруживается такое поведение, является кишечник. На рис. 5.1 показаны медленные колебания гладкой мышечной ткани. Потенциалы действия, заметные на некоторых медленных волнах, связаны с сокращениями гладкой мышцы (см. разд. 8.1). Возможность существования низкоамплитудных колебаний, которые могут быть названы медленными или подпороговыми колебаниями, является одним из свойств, присущим пейсмекерным колебаниям, и такие колебания наблюдались во многих исследованиях. Особенно поразительный пример дают численные исследования ионной модели пейсмекерных колебаний в волокне Пуркинье (рис. 5.2). По мере возрастания приложенного гиперполяризующего тока возникает ряд различных ритмов, при которых могут наблюдаться низкоамплитудные и высокоамплитудные колебания.
Рис. 5.1. Электрическая активность, записанная с помощью электрода давления (верхняя линия), и механическая активность (нижняя линия), записанная от сегмента тощей кишки кошки. Медленные волны, связанные с потенциалами действия, вызывают самые сильные сокращения, (а) Нормальная активность. (b) Тот же сегмент кишки через 40 мин после промывания раствором Тироде, содержащим 10% от количества В этом случае появление потенциалов действия зависит от того, превышает ли колеблющийся мембранный потенциал некоторую пороговую величину. Если этот порог превышен, то происходит быстрая деполяризация. Сходное явление, по крайней мере внешне, обнаруживается при изучении овуляции. Некоторые пациентки, у которых наблюдается синдром неразрыва лютеинизированного фолликула, имеют явно нормальный менструальный цикл (или текущее колебание), но ключевое событие разрыва фолликула и высвобождения яйцеклеток (или овуляция) не всегда имеет место. Эти примеры показывают,
Рис. 5.2. Эффект постоянного гиперполяризующего тока в модели волокна Пуркинье, предложенной McAlister, Noble and Tsien (1975). Использовались следующие величины гиперполяризующего тока что в таких системах основной ритм может продолжаться даже в отсутствие событий, которые многими рассматриваются как ключевые или критические.
|
 |
Оглавление
|























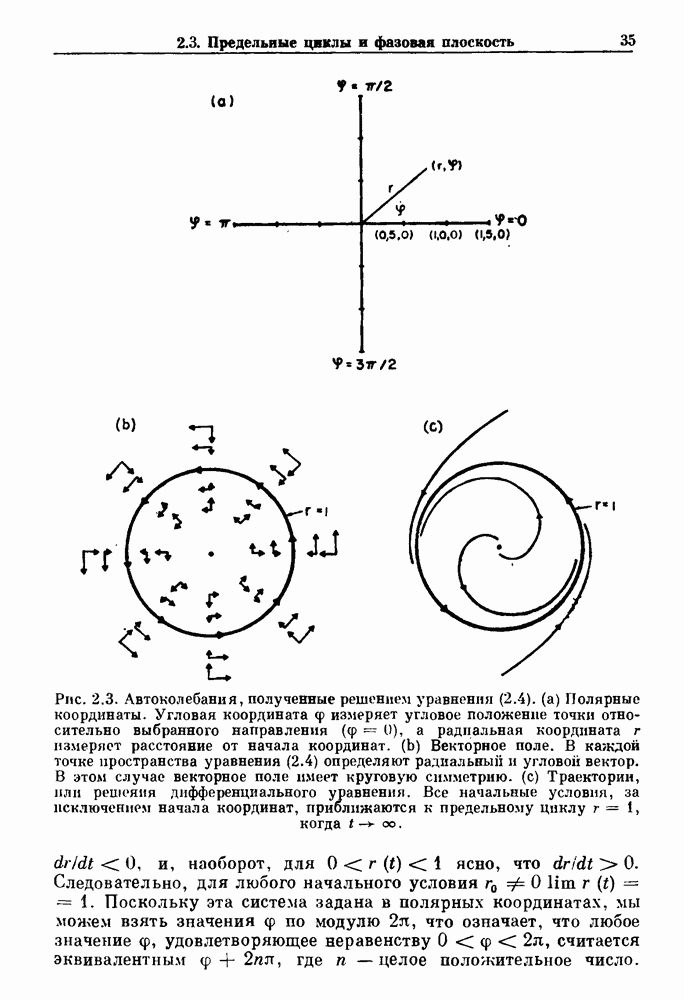







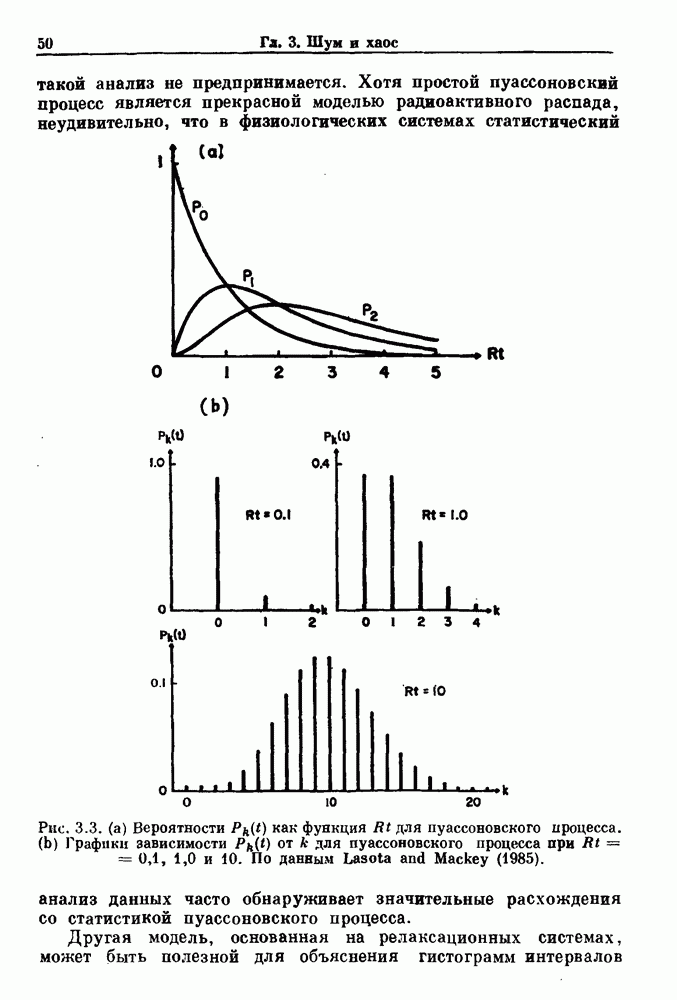















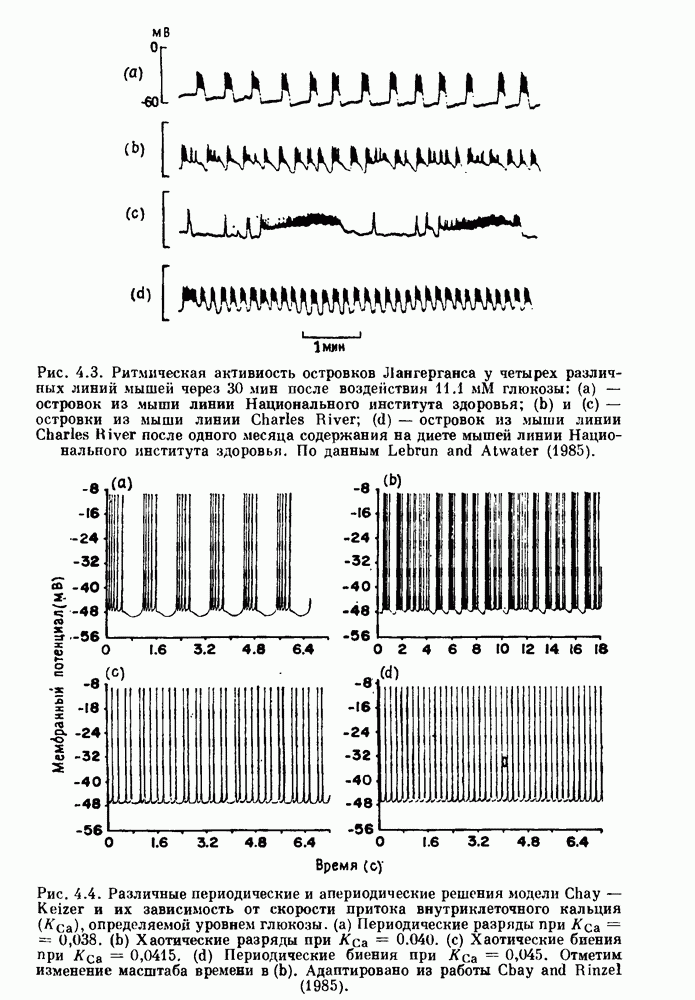





















































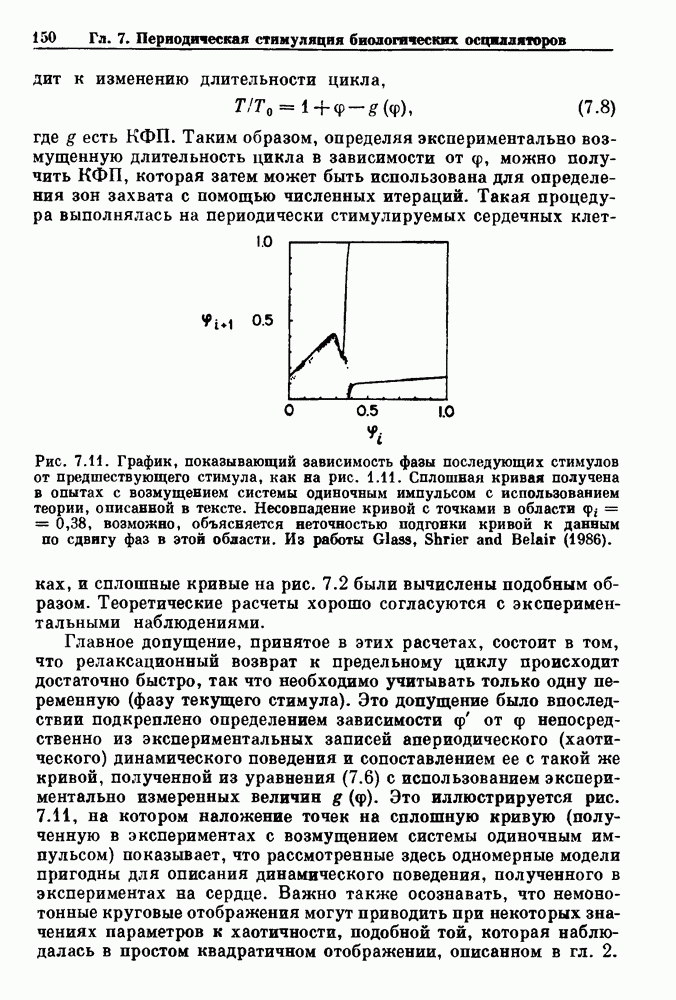


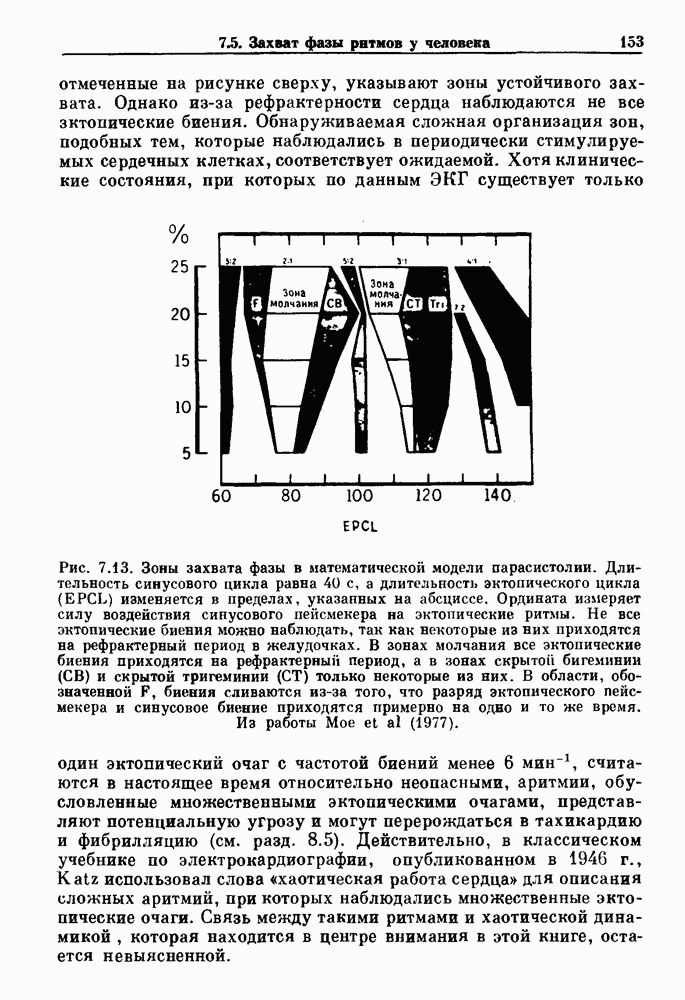
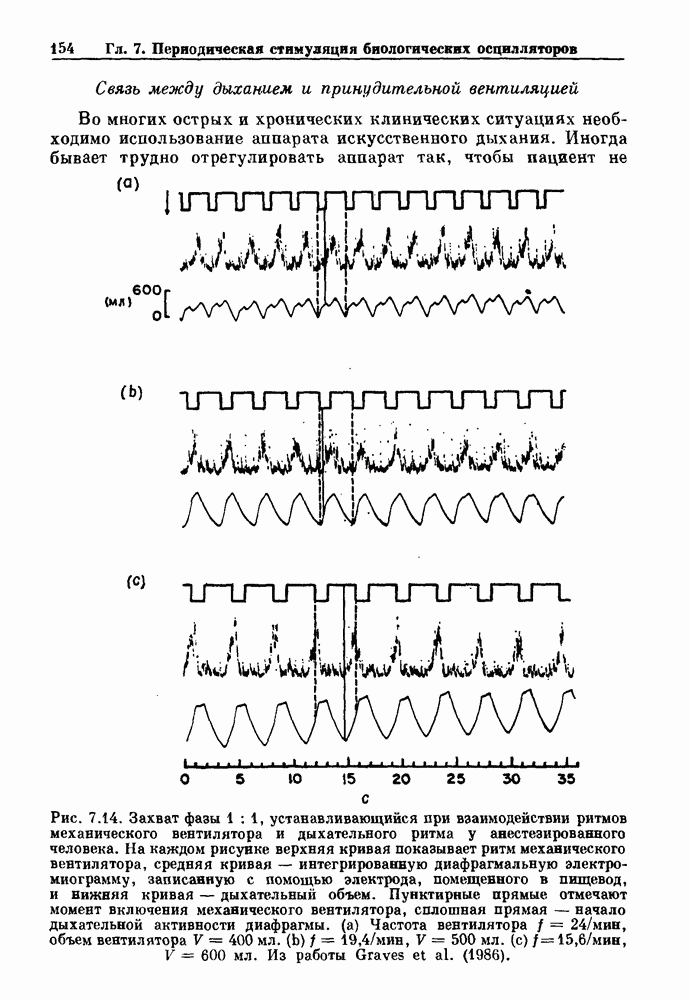
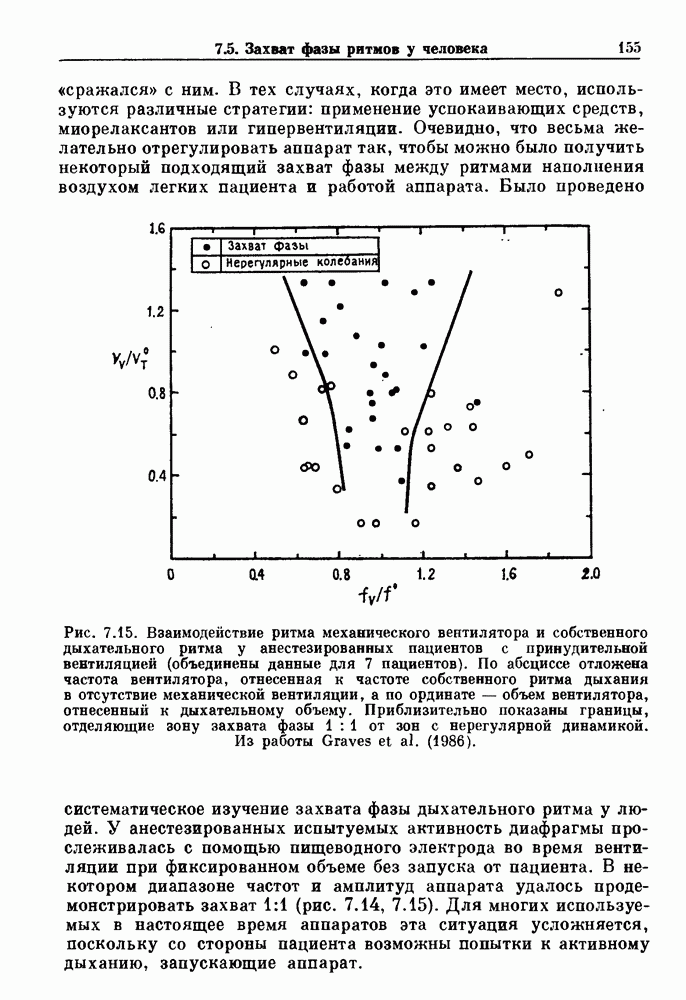







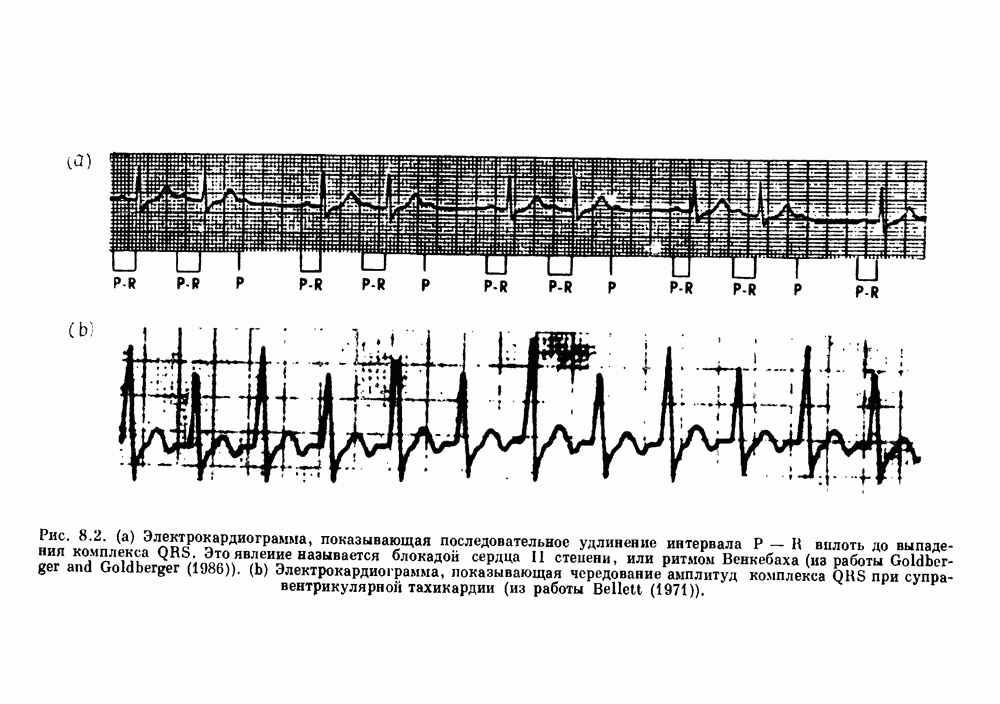


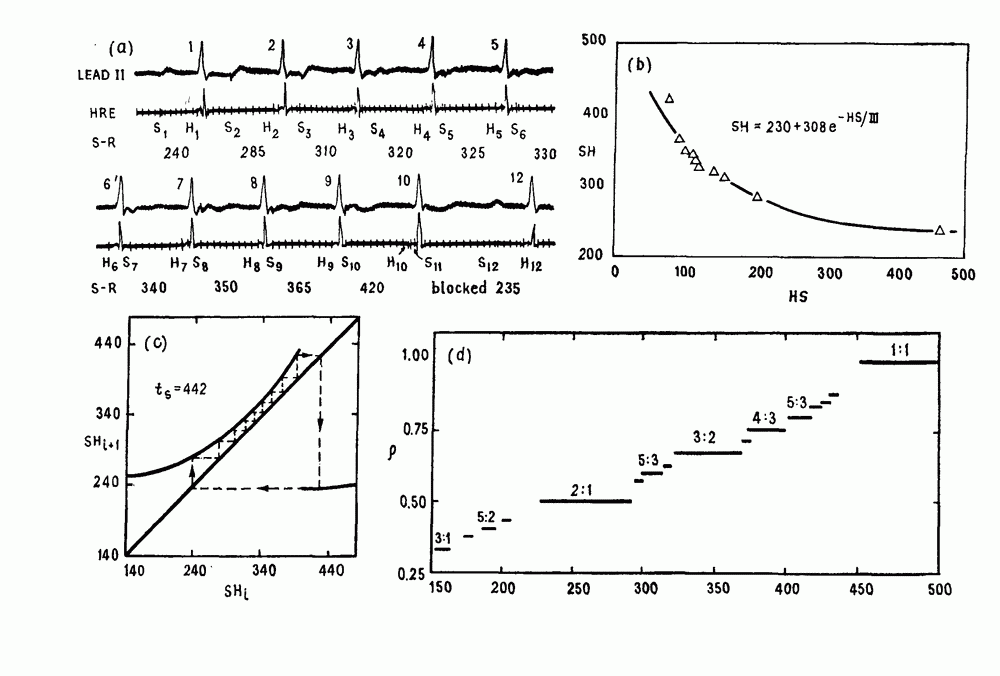

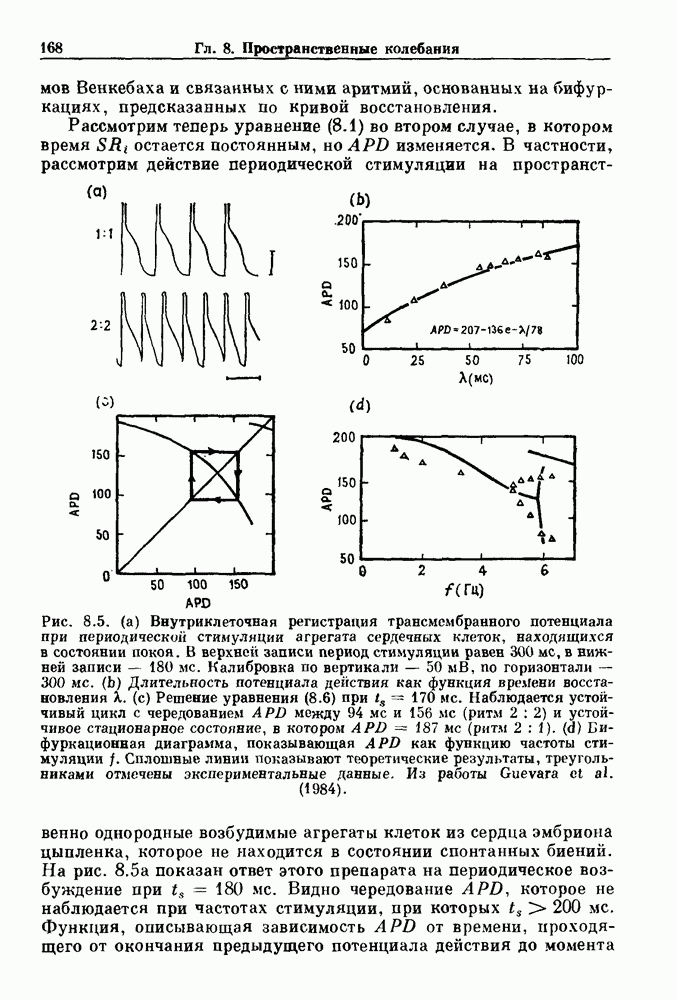

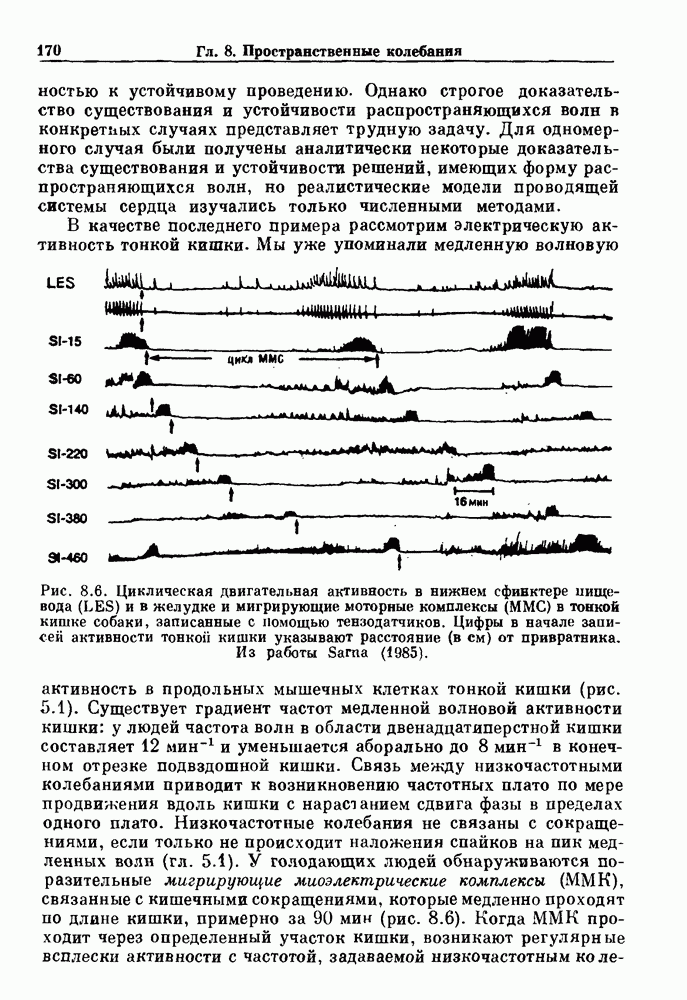







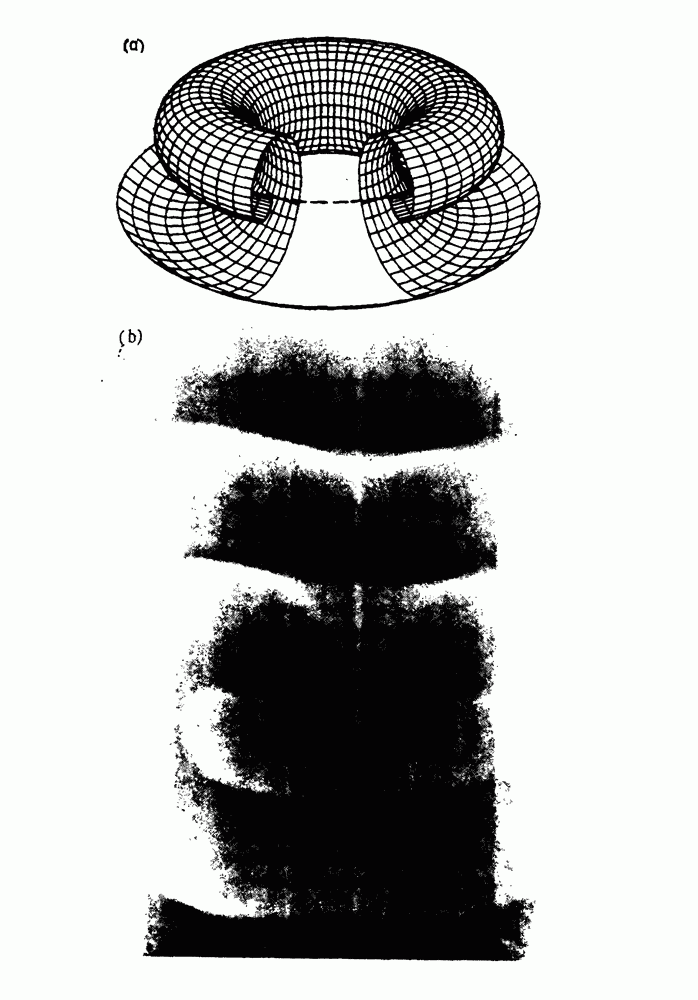































































 присутствующего в случае (а). Заметно уменьшение частоты и очевидное увеличение мембранного потенциала. Калибровка по времени 5 с; калибровка по потенциалу 0-5 мВ; калибровка по натяжению
присутствующего в случае (а). Заметно уменьшение частоты и очевидное увеличение мембранного потенциала. Калибровка по времени 5 с; калибровка по потенциалу 0-5 мВ; калибровка по натяжению  ; направление увеличение натяжения — снизу вверх. По Bortoff (1961).
; направление увеличение натяжения — снизу вверх. По Bortoff (1961).

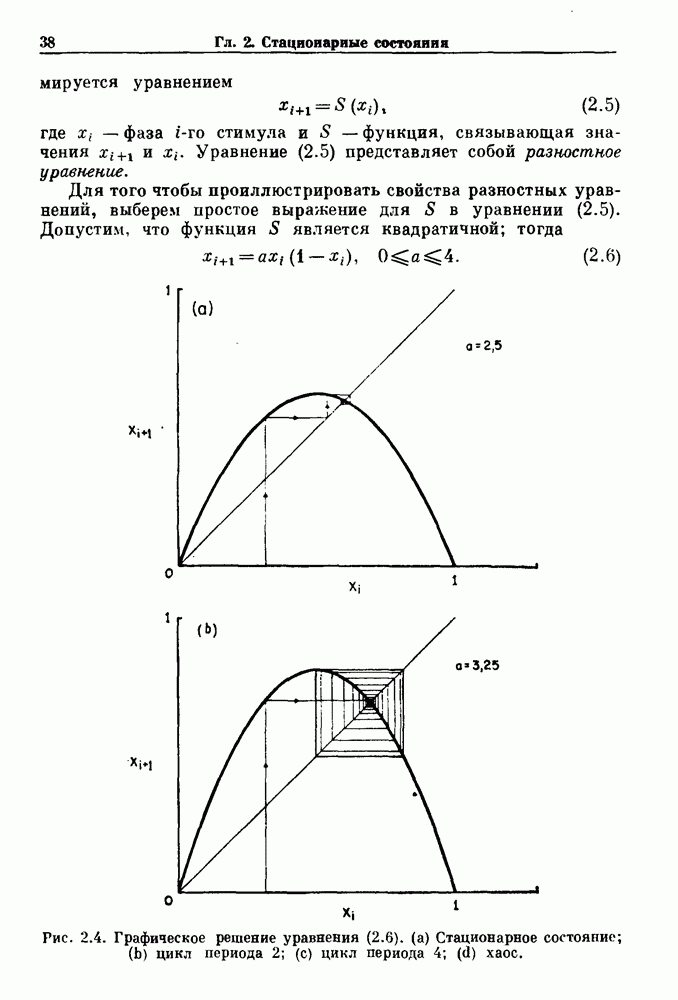
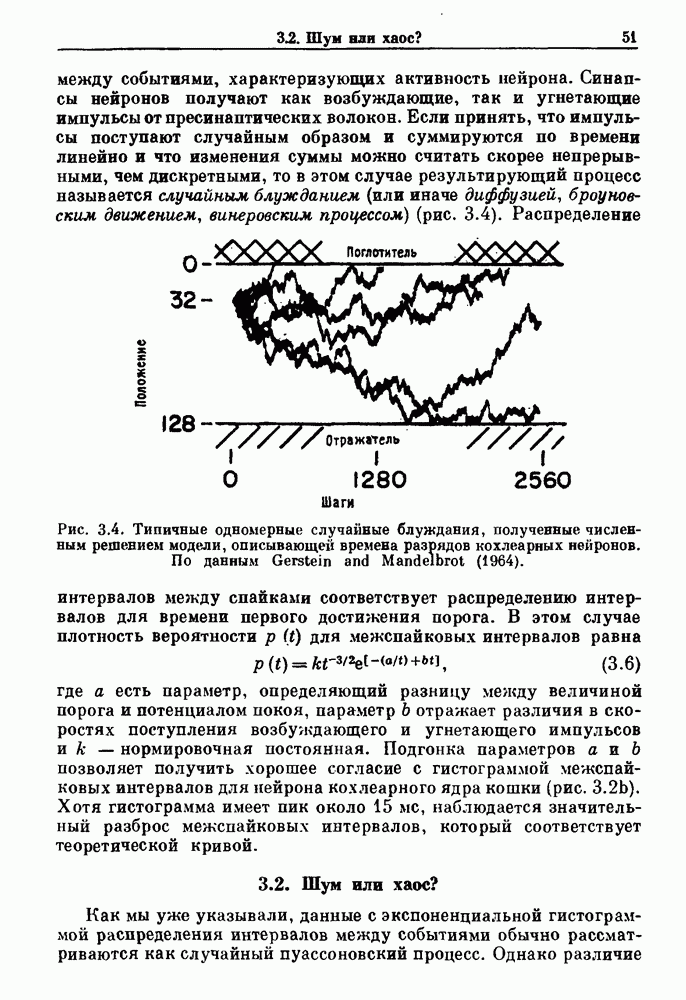
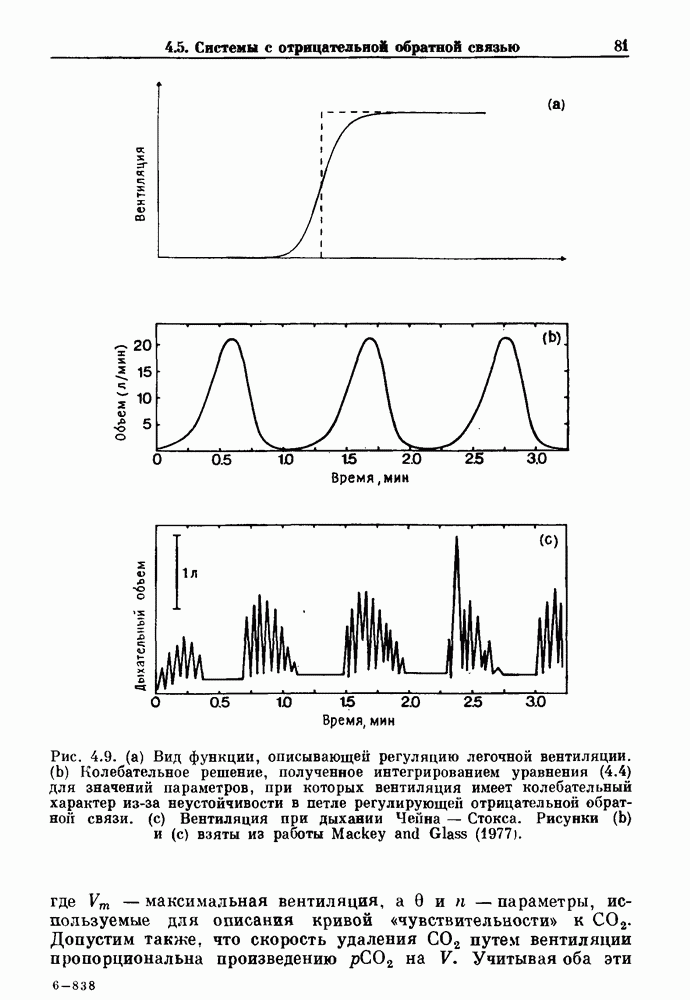
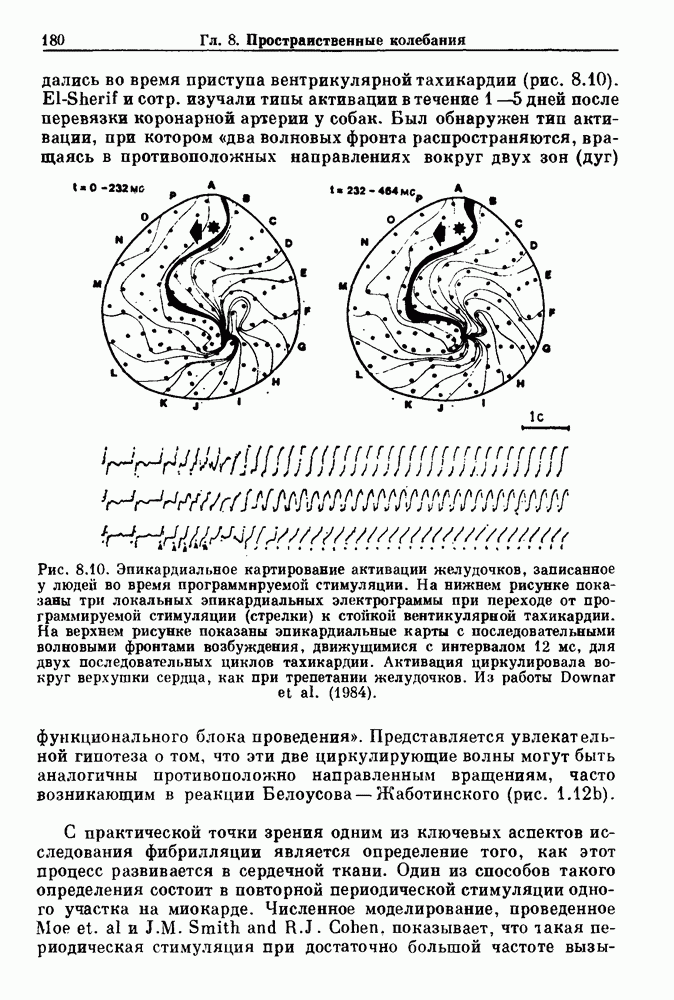
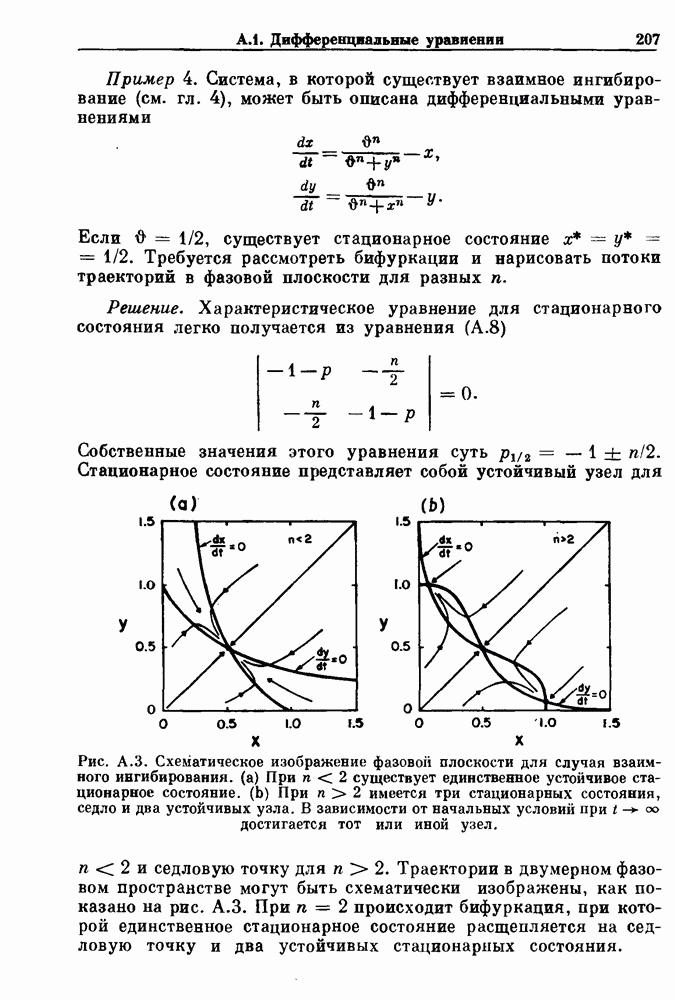
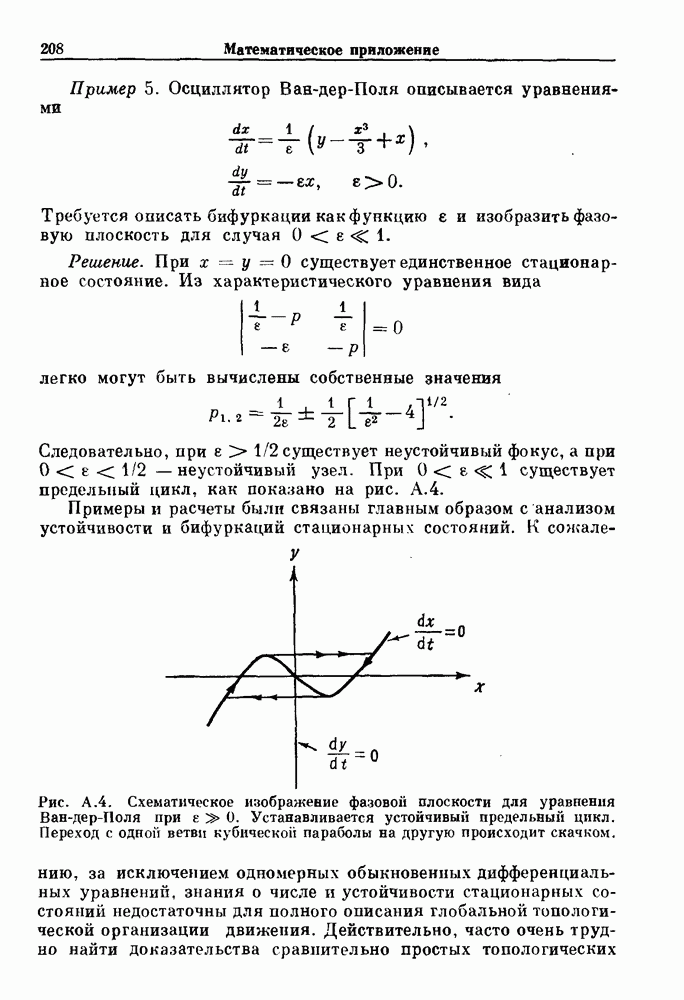
 . Наблюдаются: (а) — нормальные потенциалы действия, (b) — (е) — комбинации подпороговых и надпороговых колебаний, (f) — подпороговые колебания и (g) - состояние покоя. По данным Guevara (1987).
. Наблюдаются: (а) — нормальные потенциалы действия, (b) — (е) — комбинации подпороговых и надпороговых колебаний, (f) — подпороговые колебания и (g) - состояние покоя. По данным Guevara (1987).