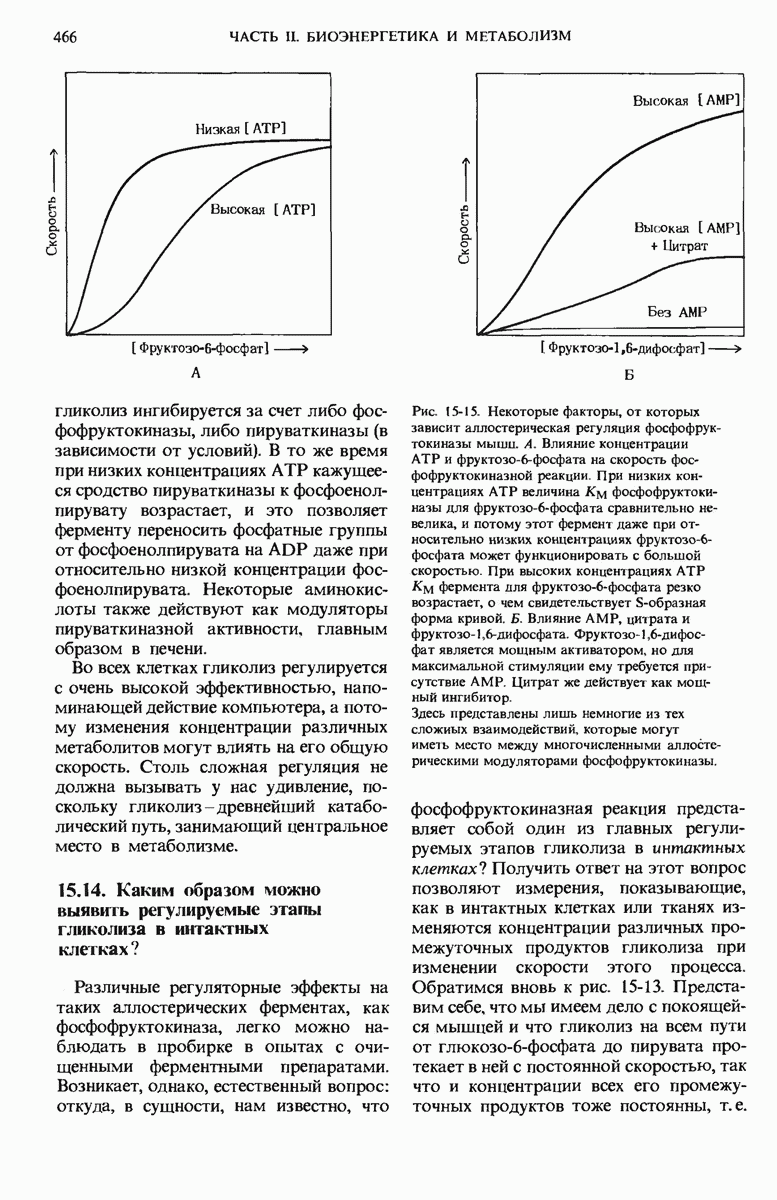
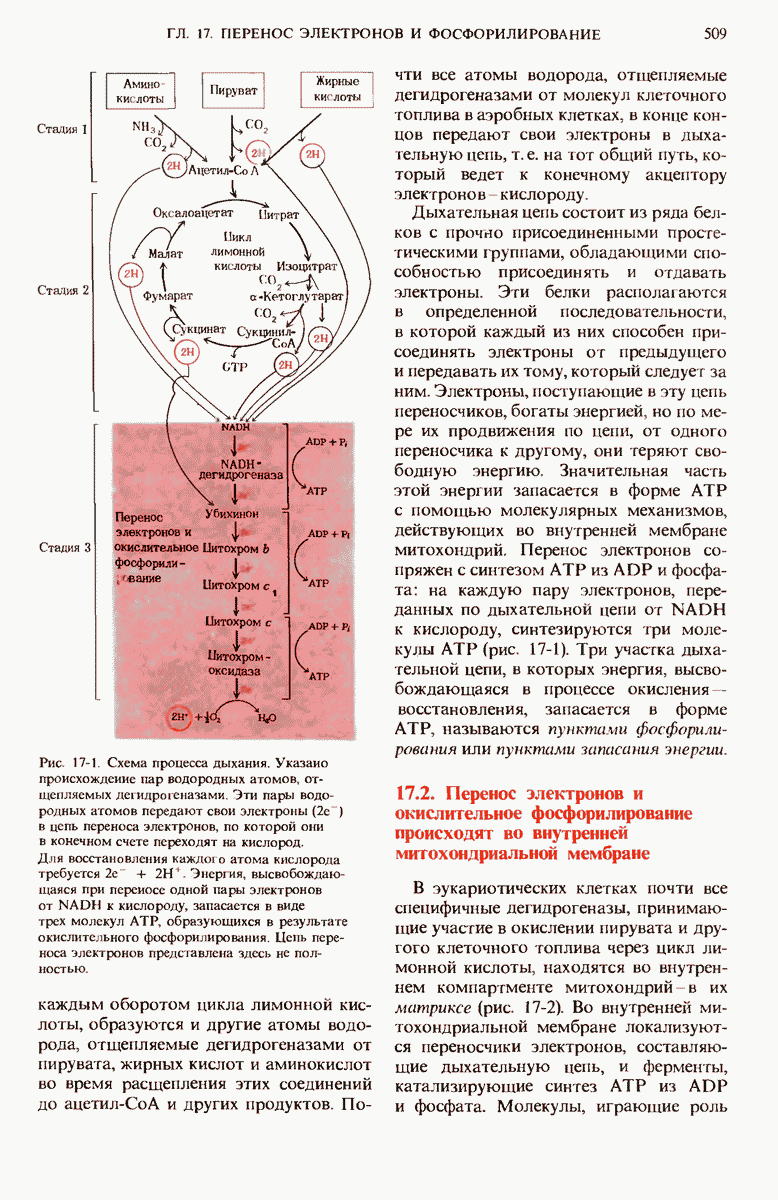
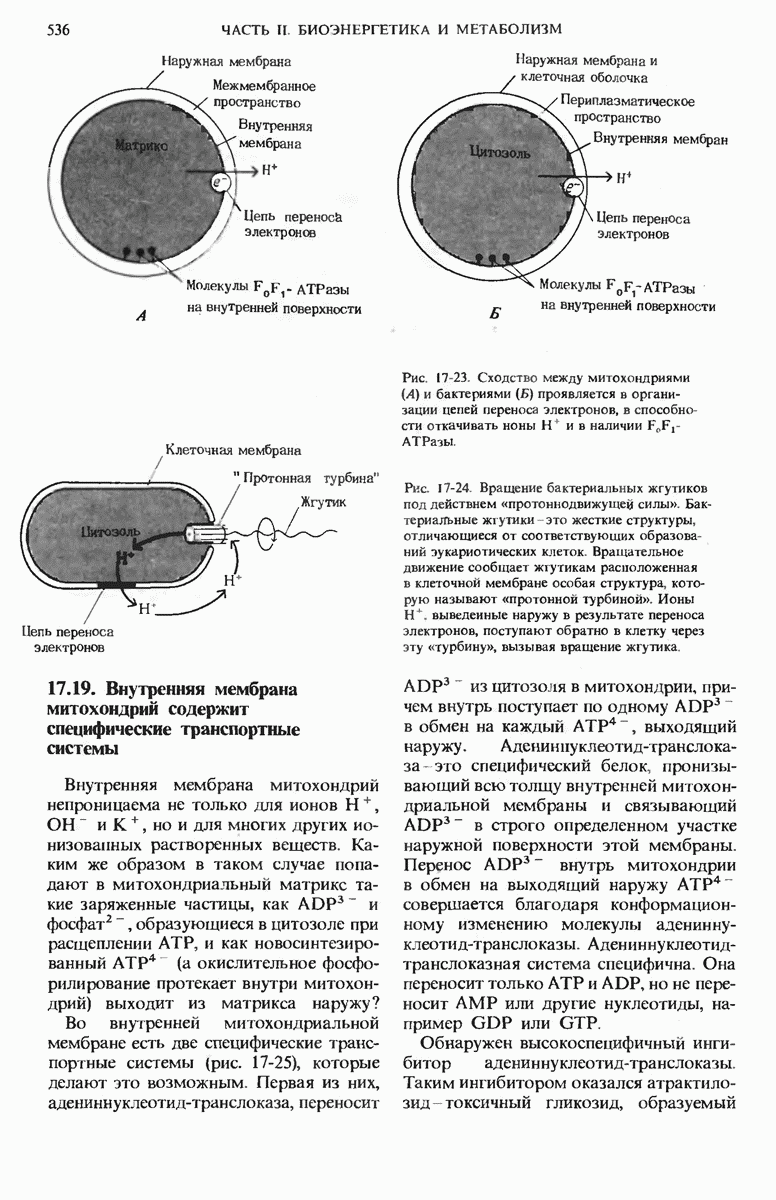
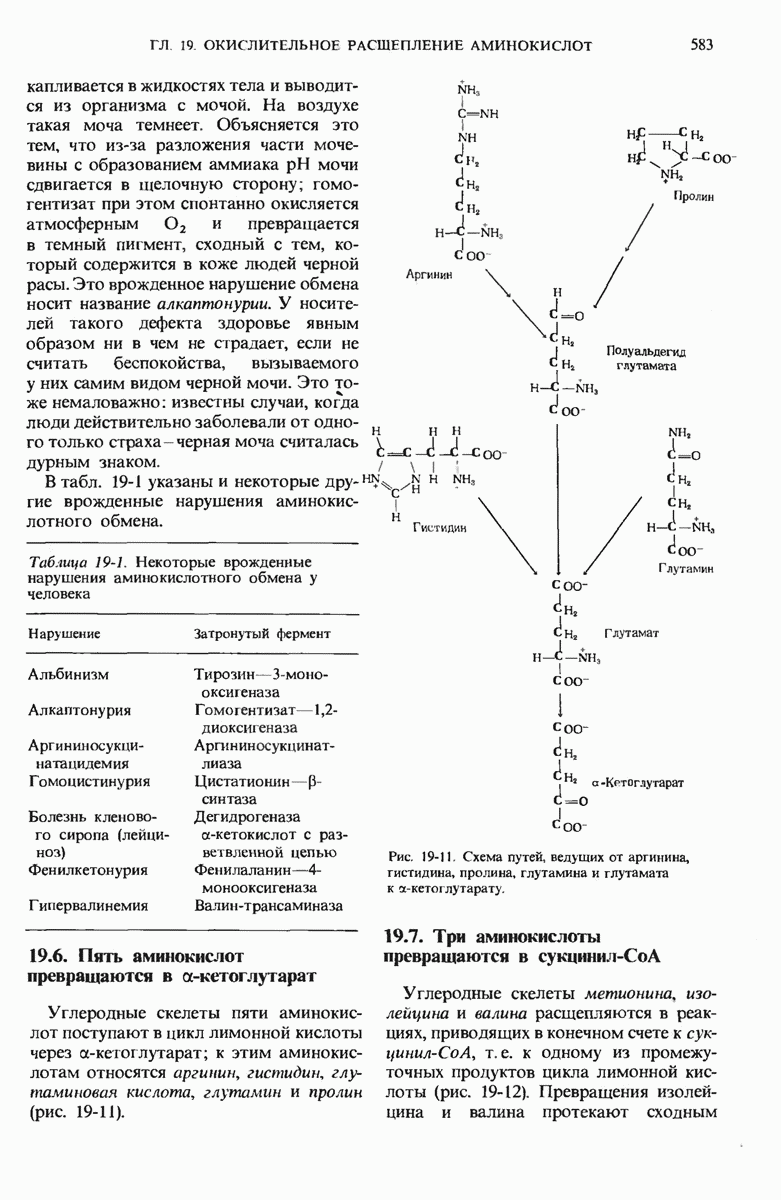
20.13. Путь биосинтеза гликогена отличается от пути его расщепления
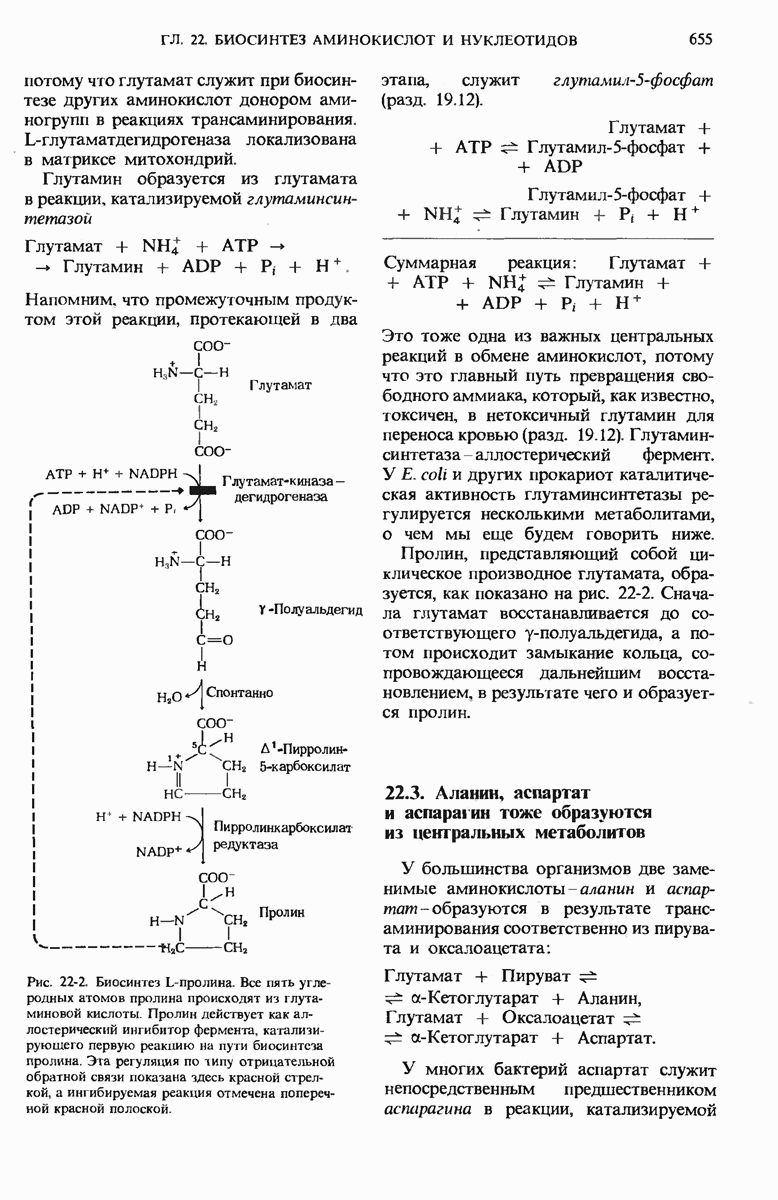
Теперь, когда мы уже знаем, каким образом из простых предшественников синтезируется глюкоза, мы можем заняться рассмотрением биосинтетического пути, ведущего к превращению остатков глюкозы в гликоген. У животных гликоген синтезируется практически во всех тканях, но особенно активны в этом отношении печень и скелетные мышцы. Начинается синтез гликогена из свободной глюкозы с гексокиназной реакции, т.е. с фосфорилирования глюкозы, в результате которого образуется глюкозо-6-фосфат

На следующем этапе глюкозо-6-фосфат обратимо превращается в глюкозо-1-фосфат в реакции, катализируемой фосфоглюкомутазой (разд. 15.8).

Далее следует ключевая реакция биосинтеза гликогена, отсутствующая в процессе его расщепления. Эта реакция представляет собой образование уридиндифосфатглюкозы ( -глюкозы) (рис. 20-8), катализируемое глюкозо-1-фосфат-уридилтрансферазой
-глюкозы) (рис. 20-8), катализируемое глюкозо-1-фосфат-уридилтрансферазой

Реакция вынуждена идти слева направо под действием пирофосфатазы, гидролизующей пирофосфат  до ортофосфата
до ортофосфата  . Ранее мы видели, что UDP-глюкоза играет роль промежуточного продукта в процессе превращения D-галактозы в D-глюкозу (рис. 15-12).
. Ранее мы видели, что UDP-глюкоза играет роль промежуточного продукта в процессе превращения D-галактозы в D-глюкозу (рис. 15-12).

Рис. 20-8. Уридиндифосфатглюкоза ( -глюкоза), играющая роль донора глюкозильных групп в гликогенсинтазной реакции.
-глюкоза), играющая роль донора глюкозильных групп в гликогенсинтазной реакции.
Она же служит и непосредственным донором глюкозильных групп при ферментативном образовании гликогена; перенос глюкозильных групп от UDP-глюкозы на нередуцирующий конец разветвленной молекулы гликогена катализируется ферментом, который носит название гликоген-синтазы (рис. 20-9). В этой реакции образуется новая  -
- -связь между 1-м углеродным атомом добавляемого остатка глюкозы и 4-м углеродным атомом концевого остатка глюкозы данной боковой цепи гликогена
-связь между 1-м углеродным атомом добавляемого остатка глюкозы и 4-м углеродным атомом концевого остатка глюкозы данной боковой цепи гликогена

Удлиненная боковая цепь гликогена

Рис. 20-9. Удлинение одной из боковых цепей гликогена, катализируемое гликоген-синтазой. D-глюкозилъная группа UDP-D-глюкозы переносная на нередуцирующий конец боковой цепи гликогена с образованием новой  связи.
связи.
Общее равновесие этих трех реакций сильно сдвинуто в сторону синтеза гликогена. Гликоген-синтазе требуется в качестве затравки а(1-4)-полиглюкозная цепь, или ветвь молекулы гликогена, состоящая не менее чем из четырех глюкозных остатков, к которым фермент последовательно присоединяет глюкозильные группы с нередуцирующего конца.
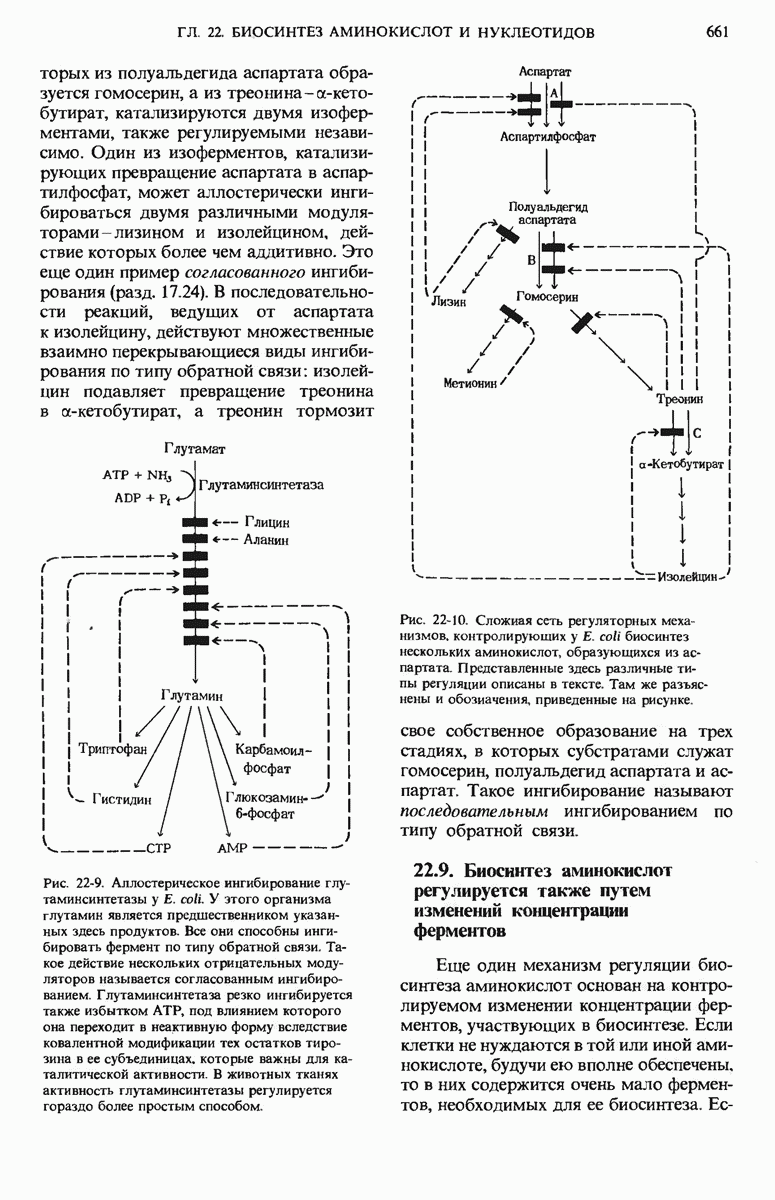
Роль UTP- и UDP-глюкозы в биосинтезе гликогена и многих других углеводов выяснил аргентинский биохимик Луис Лелуар. За эти работы он был удостоен в 1970 г. Нобелевской премии. Выше мы уже познакомились с другими примерами биосинтеза углеводов и их производных, в которых промежуточными продуктами тоже служили нуклеозиддифосфатсахара (разд. 15.9).
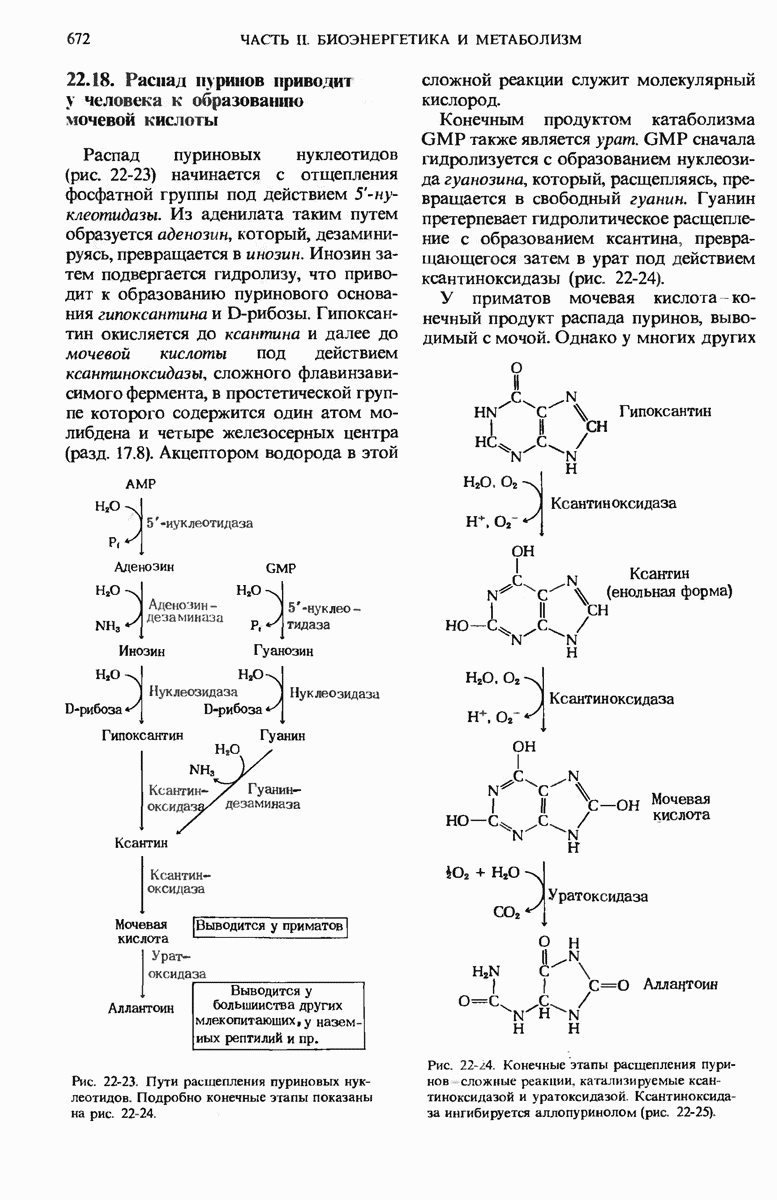
Гликоген-синтаза неспособна катализировать образование  -связей. находящихся в точках ветвления цепей гликогена. Существует специальный «ветвящий» фермент -
-связей. находящихся в точках ветвления цепей гликогена. Существует специальный «ветвящий» фермент -  -трансфераза. Он катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксильную группу остатка глюкозы той же или другой цепи гликогена, расположенного ближе к внутренней части молекулы, в результате чего образуется новая боковая цепь (рис. 20-10).
-трансфераза. Он катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксильную группу остатка глюкозы той же или другой цепи гликогена, расположенного ближе к внутренней части молекулы, в результате чего образуется новая боковая цепь (рис. 20-10).
После этого гликоген-синтаза может добавлять к этой боковой цепи новые остатки глюкозы. Биологический смысл ветвления заключается в повышении растворимости гликогена и в увеличении числа нередуцирующих концов у его молекул, что делает гликоген более доступным для действия гликоген-фосфорилазы и гликоген-синтазы.
Крахмал в растениях синтезируется таким же путем, но донором глюкозильных групп служит при этом не  -глюкоза, а
-глюкоза, а  -глюкоза.
-глюкоза.