Млекопитающие не способны синтезировать линолевую кислоту и должны получать ее с пищей растительного происхождения; экзогенную линолевую кислоту они могут превращать в арахидоновую кислоту, которая служит предшественником простагландинов.
Триацилглицеролы синтезируются путем взаимодействия двух молекул СоА - производных жирных кислот с глицерол-3-фосфатом; при этом образуется фосфатидная кислота, которая затем дефосфорилируется до диацилглицерола. Последний ацилируется третьей молекулой СоА-производного жирной кислоты с образованием триацилглицерола. Диацилглицеролы являются также главными предшественниками фосфоглицеролов. Голова молекулы фосфатидилэтаноламина сначала формируется в виде цитидиндифосфатэтаноламина путем взаимодействия цитидинтрифосфата (СТР) с фосфоэтаноламином. Далее фосфоэтаноламинная группа переносится на диацилглицерол, что приводит к образованию фосфатидилэтаноламина. Фосфатидилхолин образуется путем метилирования фосфатидилэтаноламина или при взаимодействии диацилглицерола с цитидиндифосфатхолином. Холестерол также синтезируется из ацетил-СоА в результате очень сложной последовательности реакций, в которых образуются важные промежуточные соединения - гидроксиметилглутарил-СоА, мевалонат и линейный углеводород сквален. Последний циклизуется с образованием циклической стероидной системы конденсированных колец с боковой цепью. Биосинтез холестерола ингибируется поступающим с пищей холестеролом.
ЛИТЕРАТУРА
Биосинтез жирных кислот Cunningham Е.В. Biochemistry: Mechanisms of Metabolism, McGraw-Hill, New York, 1978. В гл. 12 приведено много дополнительных сведений, касающихся энзимологии и механизма синтеза жирных кислот.
Jeffcoat R. The Biosynthesis of Unsaturated Fatty Acids and Its Control in Mammalian Liver, Essays Biochem., 15, 1-36 (1979).
Биосинтез липидов
Snyder F. (ed.). Lipid Metabolism in Mammals, vols. 1 and 2, Plenum, New York, 1977.
Обмен холестерола
Bloch К. S. The Biological Synthesis of Cholesterol, Science, 150, 19-28 (1965).
Brown M. S., Goldstein J. L. Receptor-Mediated Control of Cholesterol Metabolism, Science, 191, 150-154 (1976).
Общие вопросы
Clarke M. R. The Head of the Sperm Whale, Sci. Am., 240, 128-141, January 1979. Более подробные сведения о биологической функции спермацета у кашалотов.
Nelson R. A. Protein and Fat Metabolism in Hibernating Bears, Fed. Proc., 39, 2955-2958 (1980).
Генетические нарушения липидного обмена
Allison А. С. Lysosomes. In: J. J. Head (ed.). Carlina Biology Readers, Carolina Biological Supply Company, Burlington, N. C., 1977.
Brown M. S., Goldstein J. L. Familial Hypercholesterolemia: Defective Binding of Lipoproteins to Cultured Fibroblasts Associated with Impaired Regulation of 3-Hydroxy-3-methylglutaryl Coenzyme
A Reductase Activity, Proc. Natl. Acad. Sci. USA, 71, 788-792 (1974).
Dietschy J. M, Gotto A. M, Jr., Ontko J. A. (eds.). Disturbances in Lipid and Lipoprotein Metabolism, American Physiological Society, 1978.
Stanbury J. B., Wyngaarden J. B., Fredrickson D. S. (eds.). The Metabolic Besis of Inherited Disease, 4th ed, McGraw-Hill, New York, 1978.























































































































































































































































































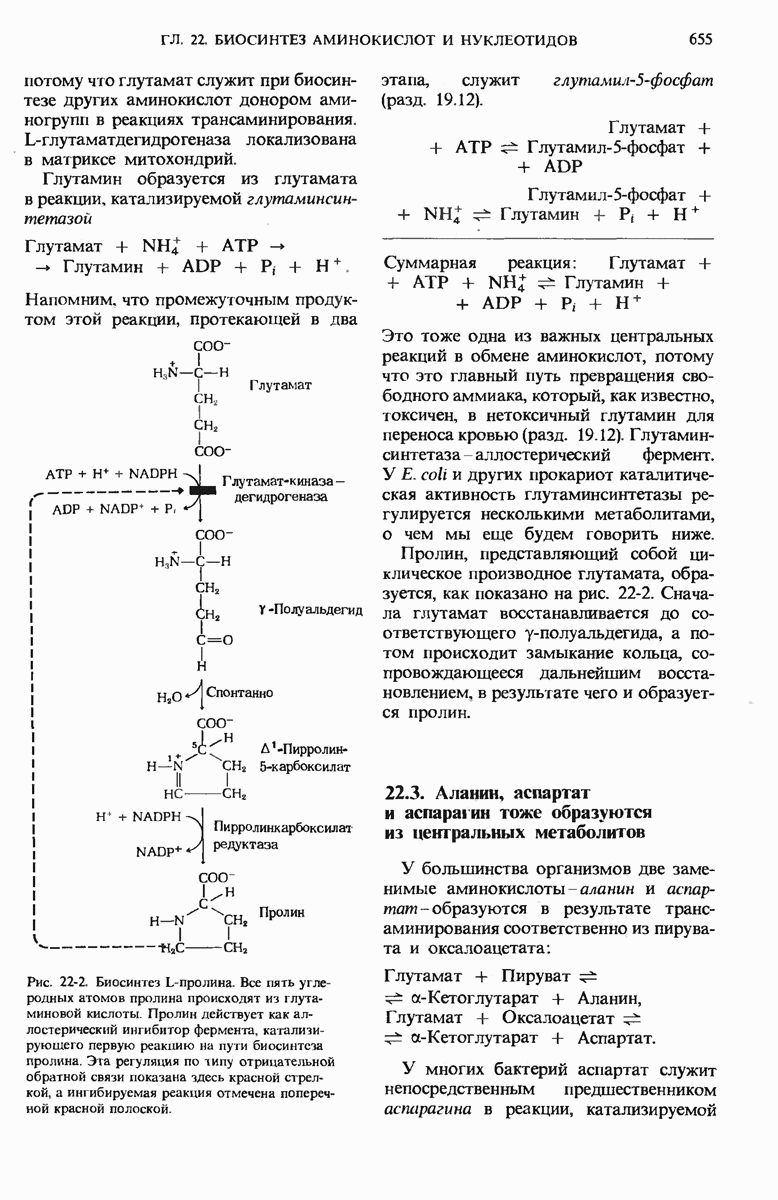





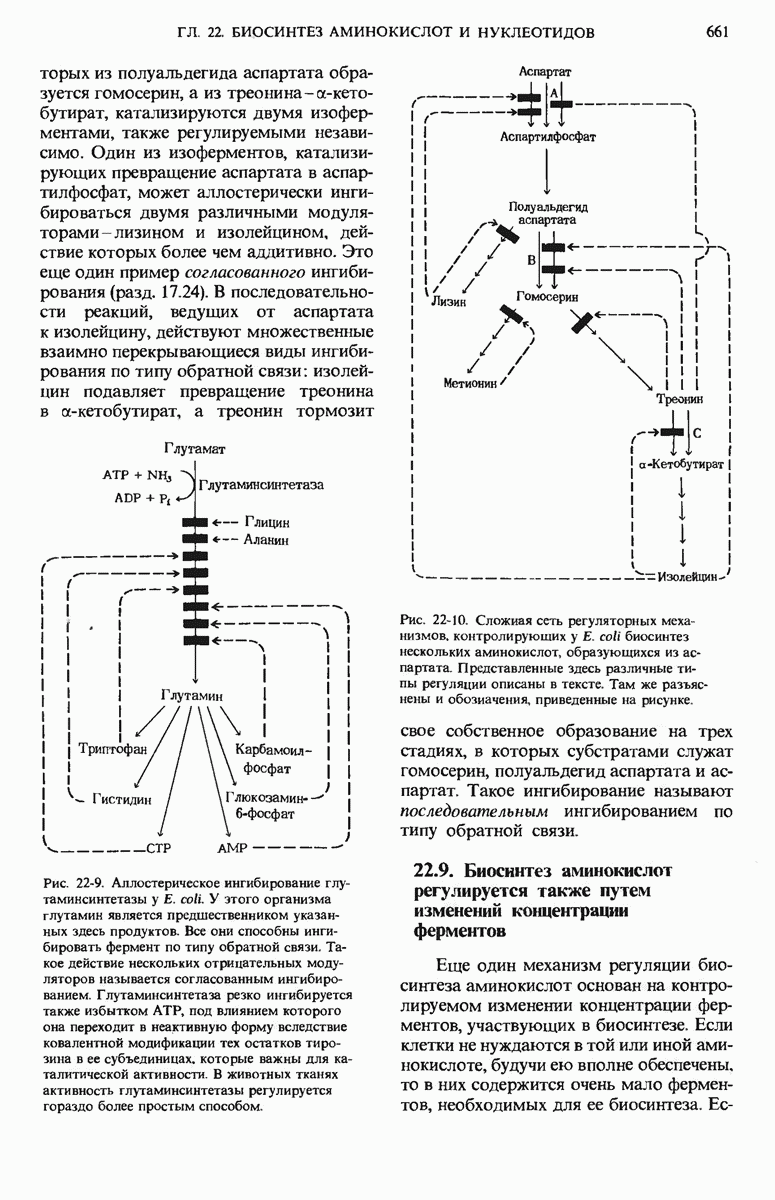










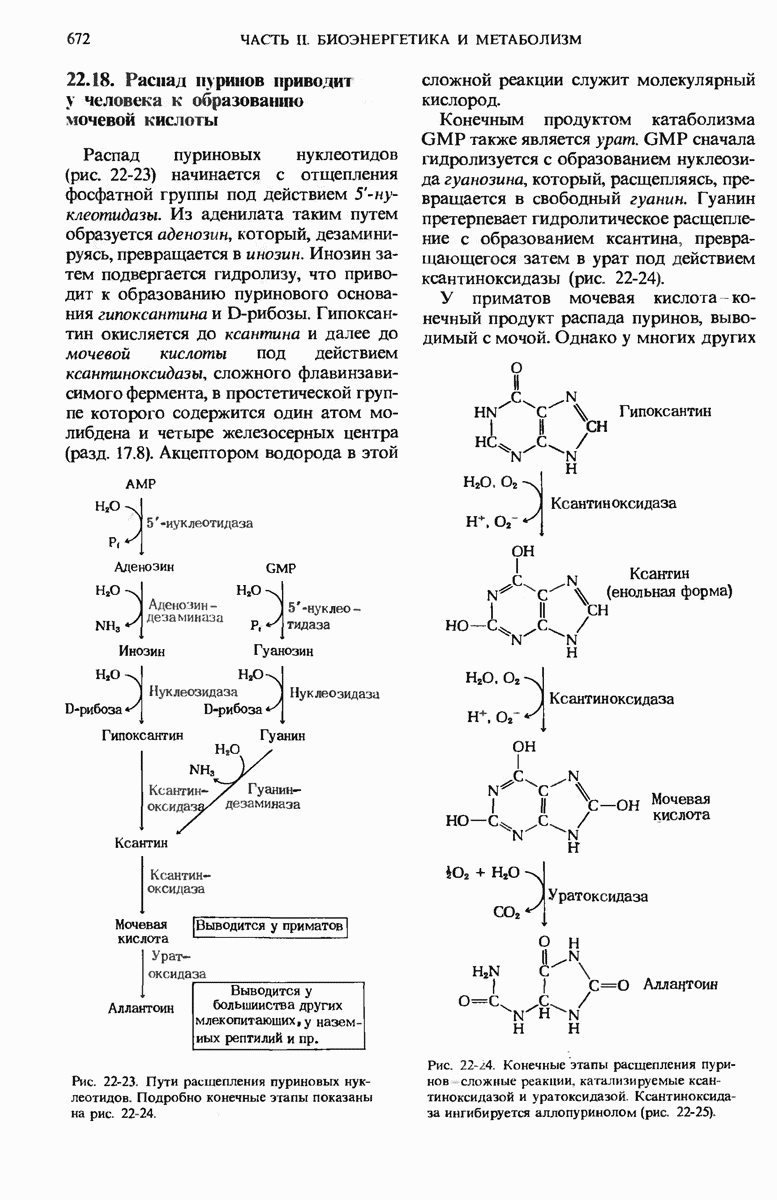



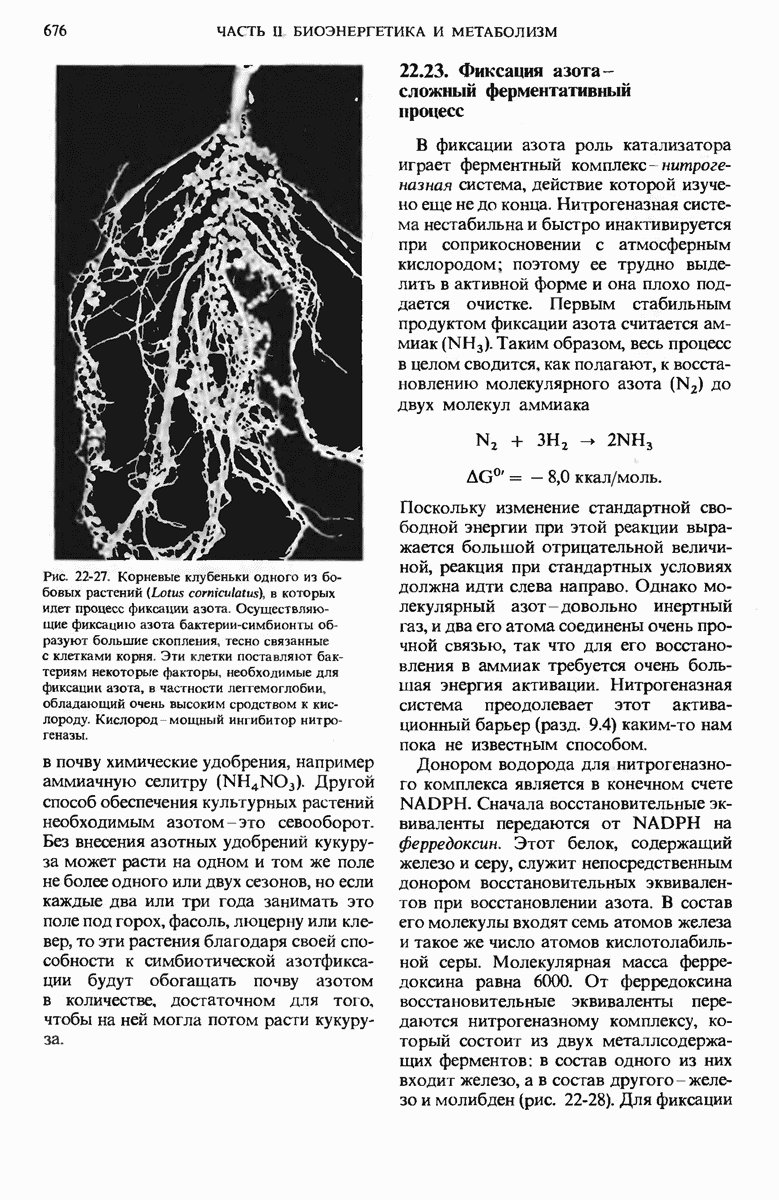

















































 а другая - остатку цистеина (Cys). АПБ функционирует как переносчик промежуточных продуктов синтеза жирных кислот. Образующийся из ацетил-СоА ацетил-S-Cys-AnE реагирует с малонил-S-АПБ, образовавшимся из малонил-СоА, в результате чего получается ацетоацетил-S-AnE и выделяется
а другая - остатку цистеина (Cys). АПБ функционирует как переносчик промежуточных продуктов синтеза жирных кислот. Образующийся из ацетил-СоА ацетил-S-Cys-AnE реагирует с малонил-S-АПБ, образовавшимся из малонил-СоА, в результате чего получается ацетоацетил-S-AnE и выделяется  . Вслед за этим происходит восстановление ацетоацетил-Б-АПБ в D-3-гидроксипроизвод-ное, которое затем гидратируется до транс-
. Вслед за этим происходит восстановление ацетоацетил-Б-АПБ в D-3-гидроксипроизвод-ное, которое затем гидратируется до транс- -бутеноил-8-АПБ; двойная связь в ацильной группе этого соединения восстанавливается и насыщается за счет NADPH с образованием бутирил-S-АПБ. Далее к карбоксильному концу растущей цепи жирной кислоты последовательно присоединяются еще шесть молекул малонил-S-АПБ и в результате получается пальмитоил-S—Cys-АПБ — конечный продукт реакций, катализируемых синтазным комплексом для образования жирных кислот. Затем происходит гидролитическое отщепление свободной пальмитиновой кислоты. При удлинении пальмитиновой кислоты возникает 18-углеродная стеариновая кислота. Под действием оксигеназ со смешанной функцией в молекулах пальмитиновой и стеариновой кислот возникают двойные связи, что приводит к образованию соответственно пальмитолеиновой и олеиновой кислот.
-бутеноил-8-АПБ; двойная связь в ацильной группе этого соединения восстанавливается и насыщается за счет NADPH с образованием бутирил-S-АПБ. Далее к карбоксильному концу растущей цепи жирной кислоты последовательно присоединяются еще шесть молекул малонил-S-АПБ и в результате получается пальмитоил-S—Cys-АПБ — конечный продукт реакций, катализируемых синтазным комплексом для образования жирных кислот. Затем происходит гидролитическое отщепление свободной пальмитиновой кислоты. При удлинении пальмитиновой кислоты возникает 18-углеродная стеариновая кислота. Под действием оксигеназ со смешанной функцией в молекулах пальмитиновой и стеариновой кислот возникают двойные связи, что приводит к образованию соответственно пальмитолеиновой и олеиновой кислот.
