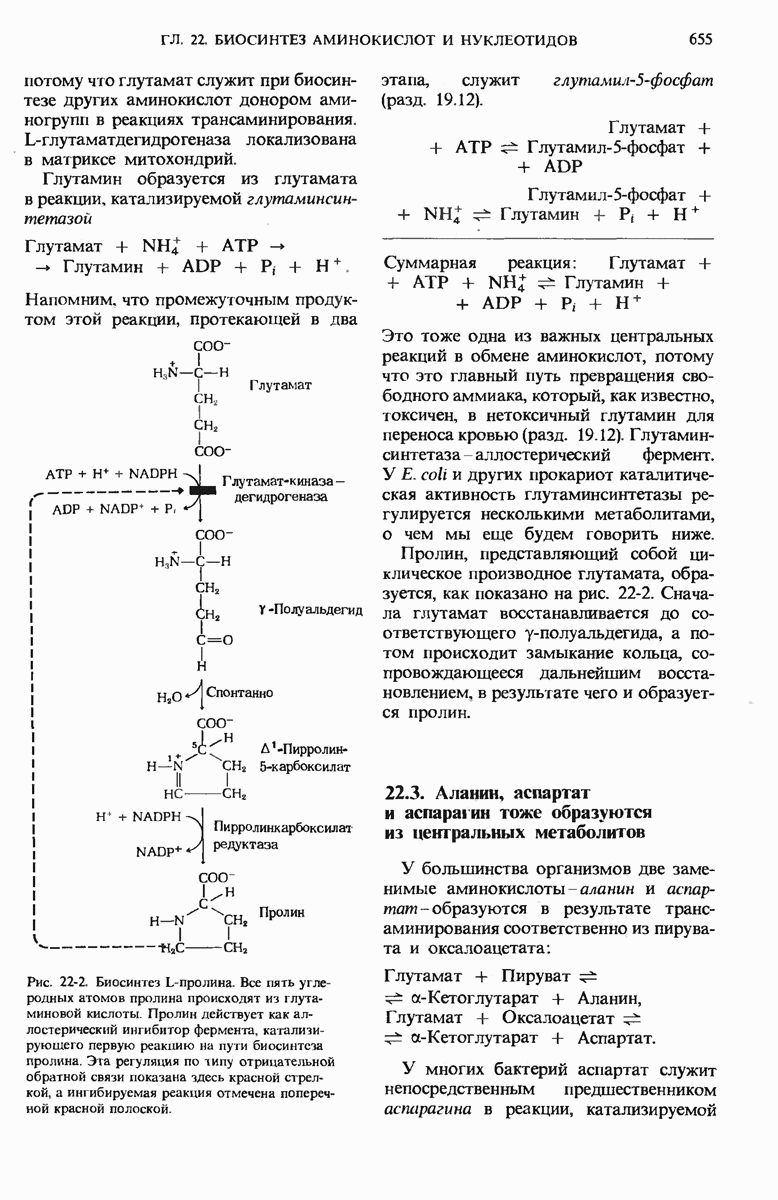
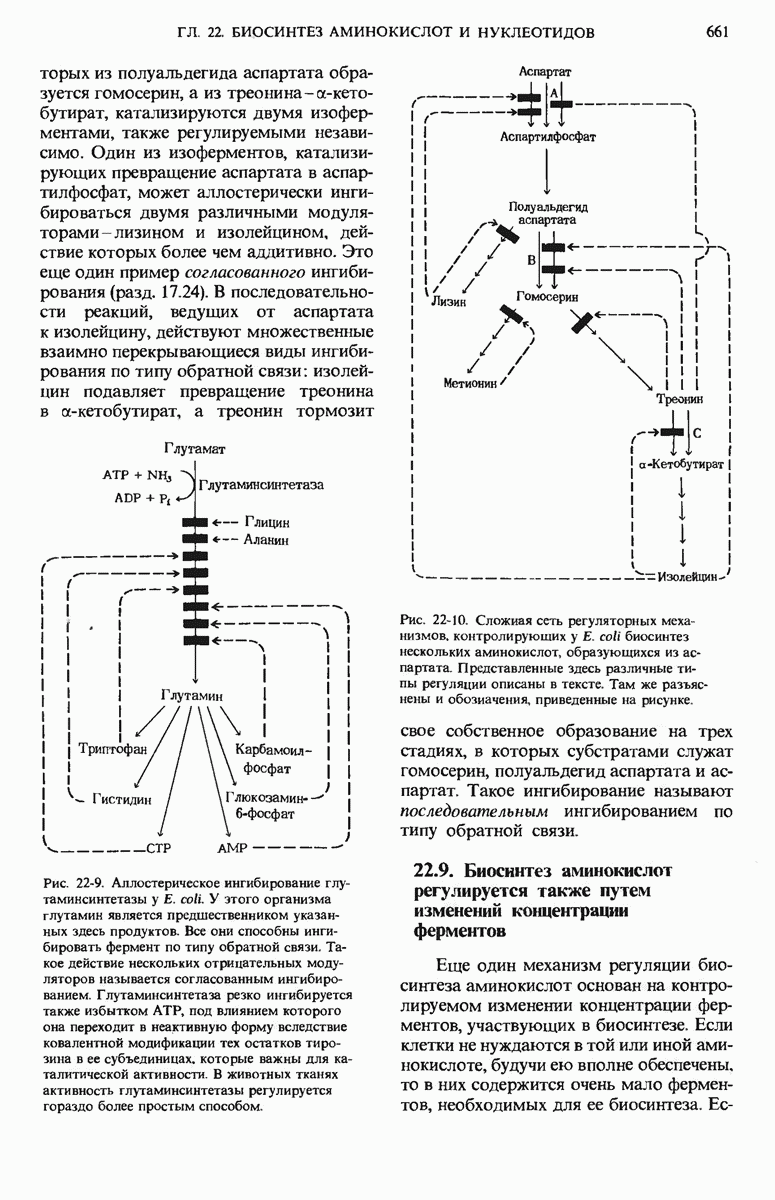
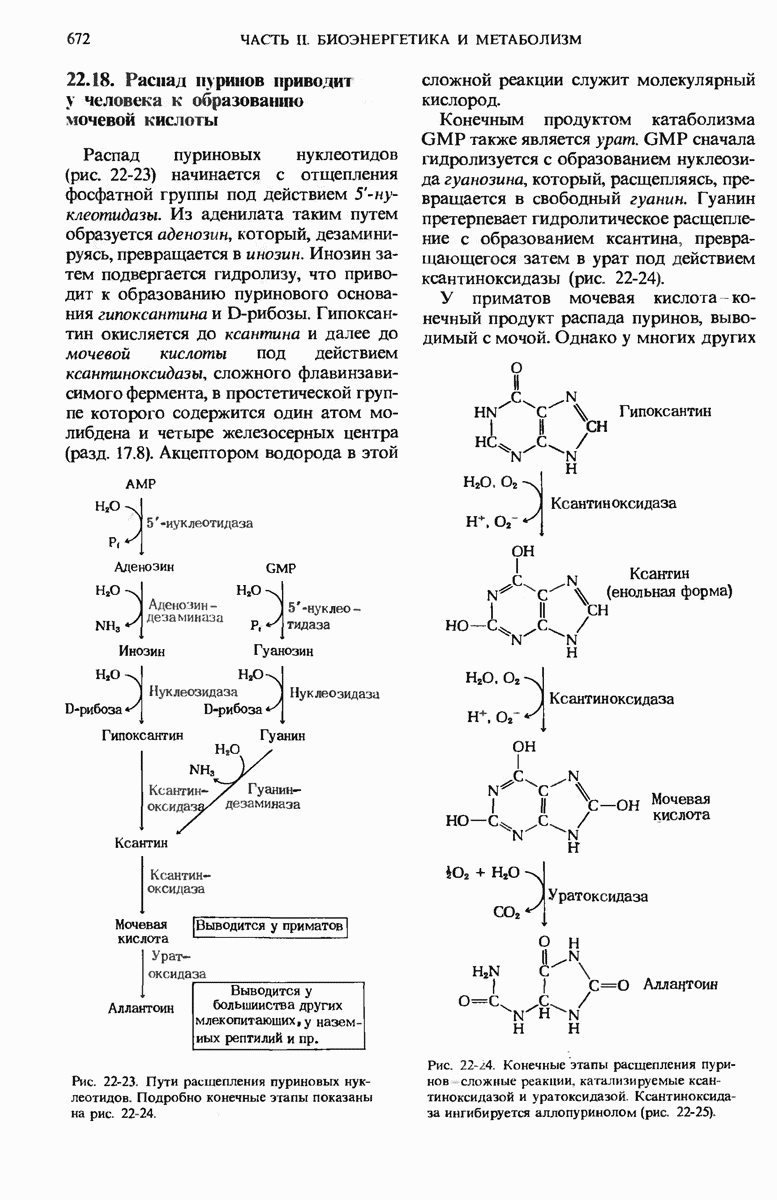
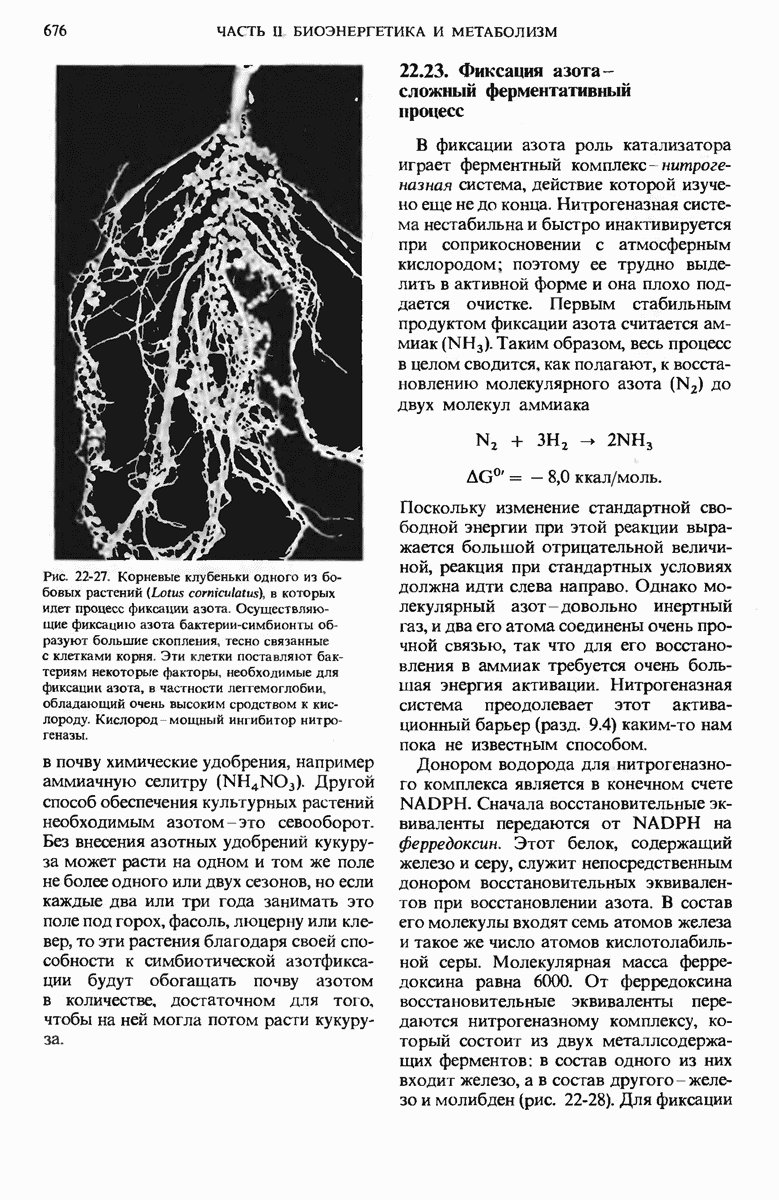
17.24. Регуляторные механизмы гликолиза, цикла лимонной кислоты и окислительного фосфорилирования взаимосвязаны
Три стадии катаболизма углеводов обеспечивают получение энергии: гликолиз (гл. 15), цикл лимонной кислоты (гл. 16) и окислительное фосфорилирование. Каждая из этих стадий регулируется при помощи своих собственных регуляторных механизмов с таким расчетом, чтобы ее скорость была достаточной для удовлетворения сиюминутной потребности клетки в продуктах, образующихся на этой стадии. Более того, эти три стадии так согласованы друг с другом, что все они функционируют в едином экономичном и саморегулируемом режиме, подобно хорошо отлаженной механической системе. Именно так вырабатывается АТР-конечный продукт катаболизма, снабжающего клетку энергией, а также некоторые специфические промежуточные продукты, такие, как пируват и цитрат, используемые в качестве предшественников в процессах биосинтеза других клеточных компонентов. Интеграция этих трех стадий оказывается возможной благодаря взаимосвязи их регуляторных механизмов. На рис. 17-29 видно, что относительные концентрации АТР и ADP (иными словами, отношение действующих масс ATP-системы) определяют не только скорости переноса электронов и окислительного фосфорилирования, но и скорости цикла лимонной кислоты, окисления пирувата и процесса гликолиза. Всякий раз, как увеличивается расходование АТР, т. е. снижается концентрация АТР, а концентрации ADP и Р; возрастают, сразу же вслед за этим возрастают скорости переноса электронов и окислительного фосфорилирования. Одновременно повышается и скорость окисления пирувата через цикл лимонной кислоты, т. е. усиливается приток электронов в дыхательную цепь.
Эти события в свою очередь приводят к увеличению скорости гликолиза, обеспечивая тем самым усиленное образование пирувата. Затем наступает момент, когда отношение  возвращается к своему обычному высокому уровню. Теперь перенос электронов и окислительное фосфорилирование замедляются, поскольку концентрация ADP устанавливается на низком уровне, соответствующем состоянию покоя. Цикл лимонной кислоты и гликолиз при этом также замедляются, потому что АТР действует как аллостерический ингибитор гликолиза и окисления пирувата.
возвращается к своему обычному высокому уровню. Теперь перенос электронов и окислительное фосфорилирование замедляются, поскольку концентрация ADP устанавливается на низком уровне, соответствующем состоянию покоя. Цикл лимонной кислоты и гликолиз при этом также замедляются, потому что АТР действует как аллостерический ингибитор гликолиза и окисления пирувата.
Согласованно действуют также регуляторные ферменты гликолиза и цикла лимонной кислоты. Когда АТР (образующийся в результате окислительного фосфорилирования) и цитрат (первый промежуточный продукт цикла лимонной кислоты) накапливаются в количествах, превышающих их обычный уровень, они, действуя согласованно, вызывают аллостерическое ингибирование фосфофруктокиназы (рис. 17-29), причем эффект от такого двойного ингибирования оказывается большим, чем сумма индивидуальных эффектов.
Рис. 17-29. Взаимозависимая регуляция гликолиза, окисления пирувата, цикла лимонной кислоты и окислительного фосфорилирования, определяемая относи гельными концентрациями АТР, ADP и АМР. Регуляторные воздействия, ингибирующие и стимулирующие, обозначены здесь красными полосками и стрелками. При высокой концентрации АТР и соответственно при низких концентрациях ADP и АМР скорости гликолиза, окисления пирувата, цикла лимонной кислоты и окислительного фосфори-лировання минимальны. Если расходование АТР в клетке резко усиливаются и, значит, концентрации ADP, АМР и возрастают, то все эти четыре процесса ускоряются. Взаимосвязь гликолиза и цикла лимонной кислоты, осуществляемая через цитрат (она также показана на этой схеме), дополняет регуляторное действие аденилатной системы. Кроме того, при повышении концентраций NADH и  подавляется процесс окисления пирувата до ацетил-СоА.
подавляется процесс окисления пирувата до ацетил-СоА.  -глюкозо-6-фосфат; Ф6Ф-фруктозо-6-фосфат; ФДФ фруктозодифосфат; ГЗФ-глицеральдегид-З-фосфат; ЗФГ
-глюкозо-6-фосфат; Ф6Ф-фруктозо-6-фосфат; ФДФ фруктозодифосфат; ГЗФ-глицеральдегид-З-фосфат; ЗФГ  -фосфоглицерат;
-фосфоглицерат;  -фосфоглицерат; ФЕП фосфоенолпируват;
-фосфоглицерат; ФЕП фосфоенолпируват;  -КГ
-КГ  -кетоглутарат.
-кетоглутарат.
Таким образом, гликолиз контролируется целой сетью взаимосвязанных регуляторных механизмов, благодаря чему пируват образуется лишь с той скоростью, с какой он потребляется в цикле лимонной кислоты, который служит поставщиком электронов для процесса окислительного фосфорилирования.
В раковых клетках эта координация регуляторных влияний, по-видимому, нарушена: гликолиз протекает в них со значительно большей скоростью, чем это требуется для обеспечения топливом цикла лимонной кислоты. Поэтому аэробные раковые клетки потребляют гораздо больше глюкозы из крови, чем нормальные, но оказываются не в состоянии окислить весь пируват, образовавшийся в процессе гликолиза. Большая его часть окисляется в них до лактата, который уносится кровью.