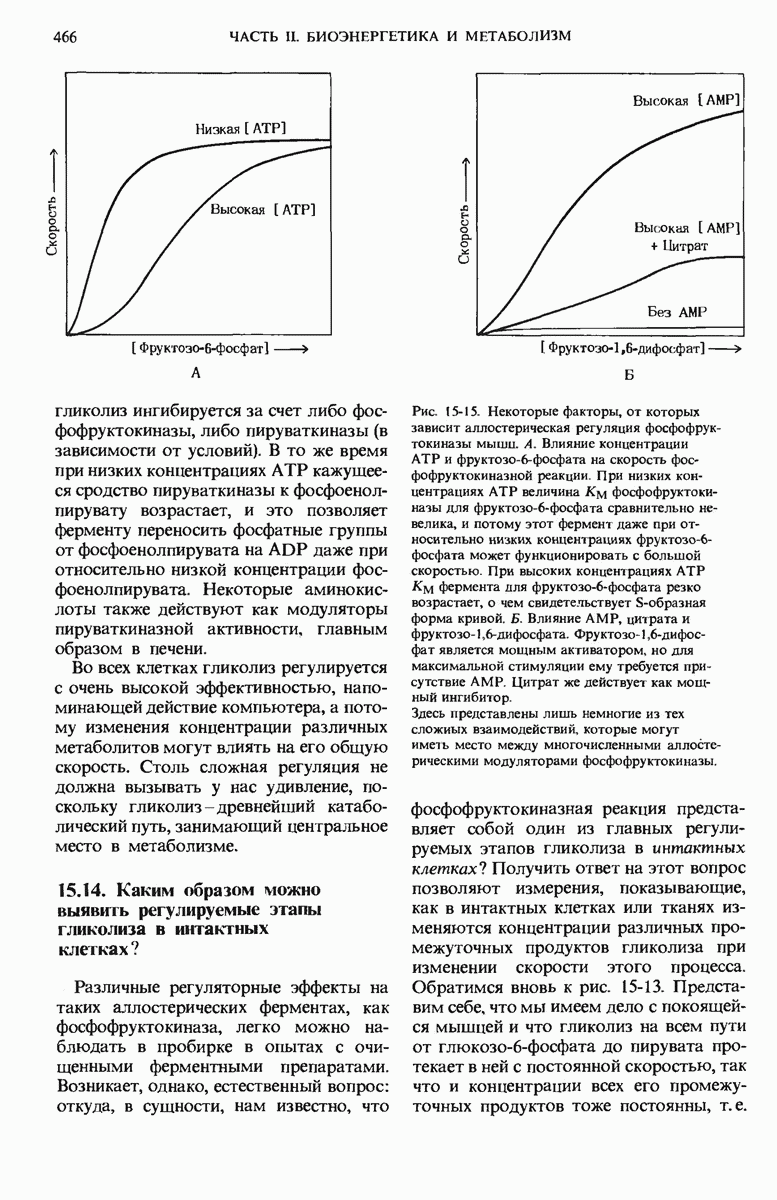
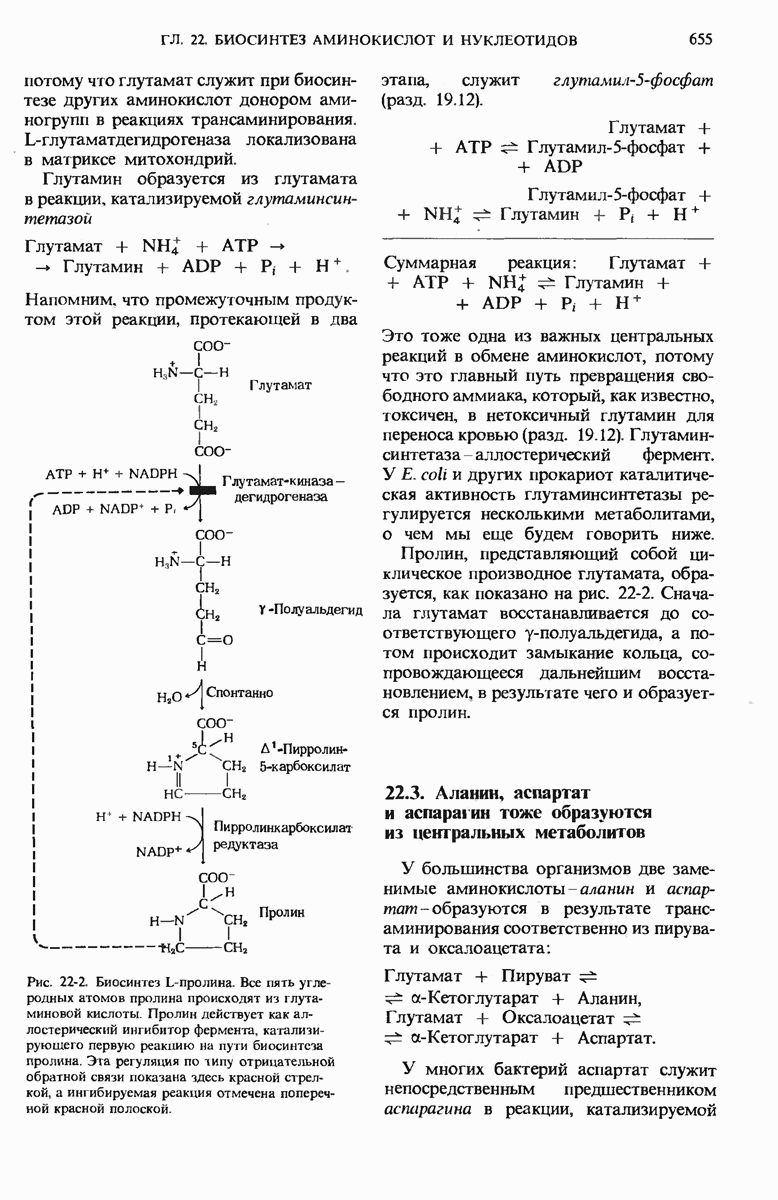
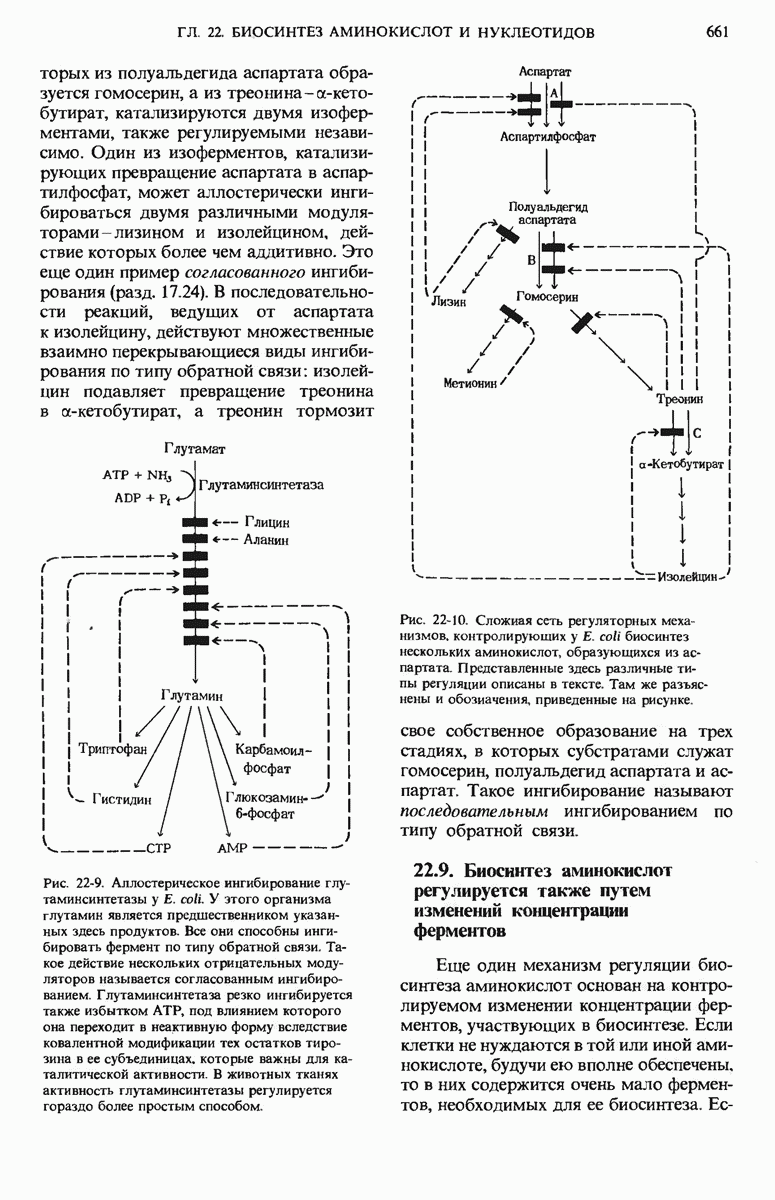
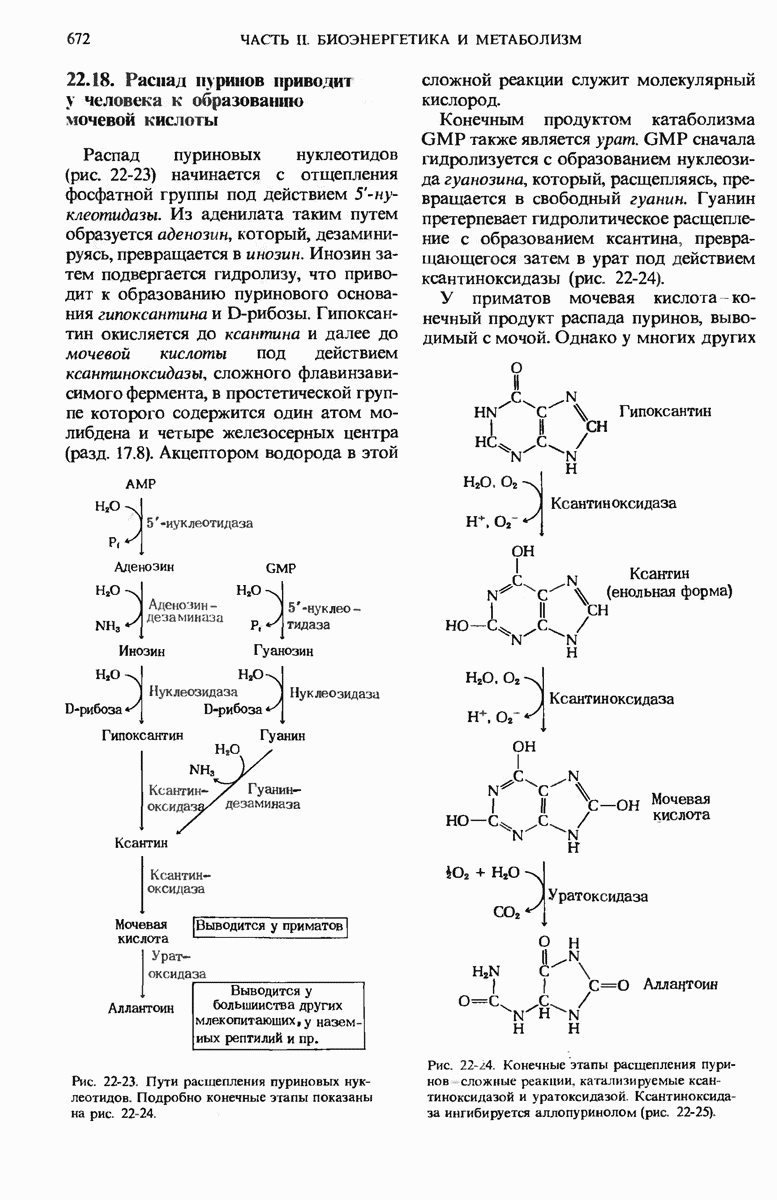
21.5. Присоединение каждого двухуглеродного фрагмента происходит в четыре этапа
а. Конденсация
На первом из четырех этапов удлинения углеродной цепи жирной кислоты ацетильная и малонильная группы, ковалентно связанные с SH-группами синтазы, подвергаются конденсации с образованием ацетоацетильной группы, ковалентно связанной с SH-группой фосфопантетеина: одновременно с этим происходит выделение молекулы  Эту реакцию катализирует 3-кетоацил-АПБ—синтаза:
Эту реакцию катализирует 3-кетоацил-АПБ—синтаза:

Обратите внимание на то, что ацетильная группа переносится с SH-группы цистеина на малонильную группу, которая связана с SH-группой фосфопантетеина; таким образом, она становится концевым двухуглеродным звеном вновь образованной ацетоацетильной группы (рис. 21-9). При этом ацетильная группа вытесняет свободную карбоксильную группу остатка малоната в виде  . В ходе этой реакции образуется та самая
. В ходе этой реакции образуется та самая  которая исходно включилась в молекулу малонил-СоА путем описанной выше ацетил-СоА-карбоксилазной реакции. Таким образом, в ходе биосинтеза жирной кислоты двуокись углерода не используется для построения ковалентного остова молекулы жирной кислоты, а играет роль катализатора, поскольку каждый раз при присоединении новой двухуглеродной единицы она регенири-рует, отщепляясь от растущей цепи жирной кислоты.
которая исходно включилась в молекулу малонил-СоА путем описанной выше ацетил-СоА-карбоксилазной реакции. Таким образом, в ходе биосинтеза жирной кислоты двуокись углерода не используется для построения ковалентного остова молекулы жирной кислоты, а играет роль катализатора, поскольку каждый раз при присоединении новой двухуглеродной единицы она регенири-рует, отщепляясь от растущей цепи жирной кислоты.

Рис. 21-9. Реакция конденсации в биосинтезе жирных кислот. Отщепление  от малонильной группы (показана красным цветом) создает движущую силу для переноса ацетильной группы (на сером фоне) от Cys—SH на 2-й углеродный атом малонильной группы. Исходная ацетильная группа по завершении синтеза жирной кислоты становится последней двухуглеродной единицей метального конца молекулы жирной кислоты.
от малонильной группы (показана красным цветом) создает движущую силу для переноса ацетильной группы (на сером фоне) от Cys—SH на 2-й углеродный атом малонильной группы. Исходная ацетильная группа по завершении синтеза жирной кислоты становится последней двухуглеродной единицей метального конца молекулы жирной кислоты.
Зачем клеткам понадобилось создавать малонильную группу из ацетильной, присоединяя  которая затем вновь отщепляется при образовании ацетоацетата? Ответ на этот вопрос заключается в том, что при отщеплении
которая затем вновь отщепляется при образовании ацетоацетата? Ответ на этот вопрос заключается в том, что при отщеплении  от малонильной группы резко возрастает реакционная способность оставшегося двухуглеродного фрагмента и благодаря этому он может быстро взаимодействовать с ацетильной группой (рис. 21-9).
от малонильной группы резко возрастает реакционная способность оставшегося двухуглеродного фрагмента и благодаря этому он может быстро взаимодействовать с ацетильной группой (рис. 21-9).
б. 3-кетовосстановление
Ацетоацетил-8-АПБ подвергается далее восстановлению по карбонильной группе с образованием D-3-гидроксибутирил-S-AnB.
В этой реакции, катализируемой 3-кетоацил-АПБ—редуктазой, в качестве донора электронов используется NADPH (рис. 21-10).

Отметим, что D-3-гидроксибутирильная группа стерически не идентична L-3-гидроксиацильной группе промежуточного продукта, образующегося при окислении жирных кислот (разд. 18.4,б).
в. Дегидратация
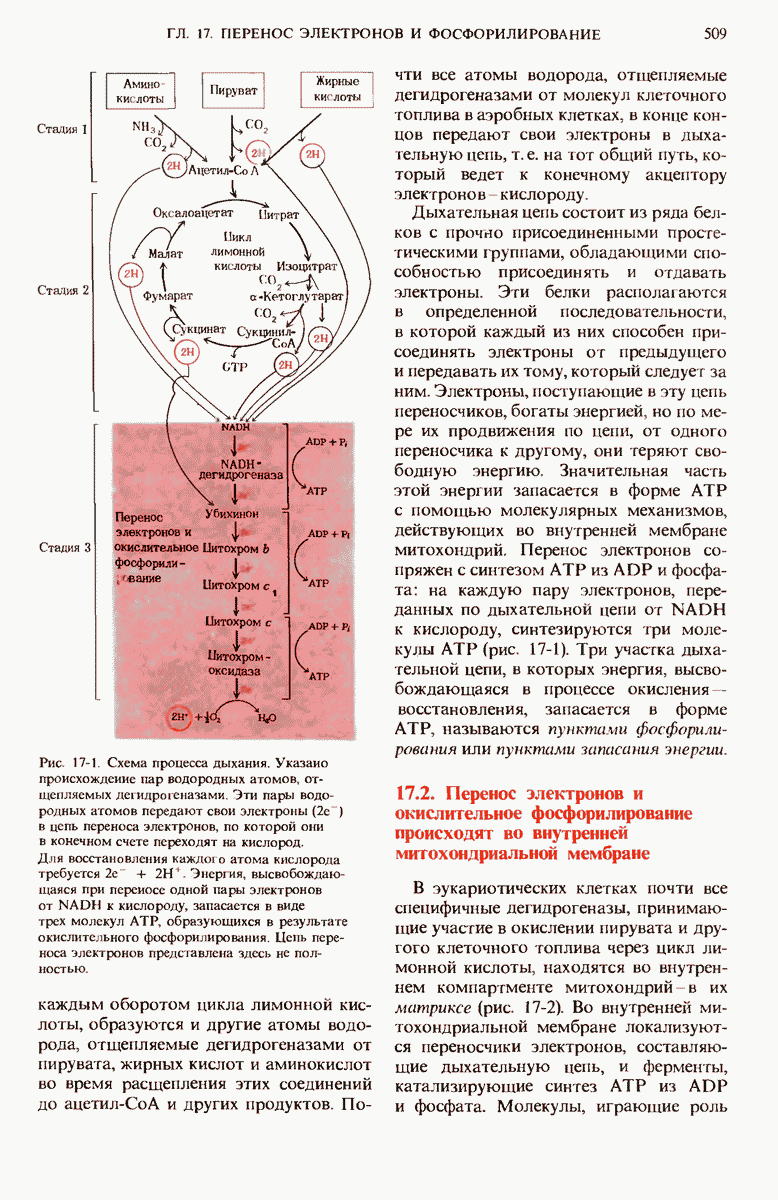
В ходе третьего этапа цикла синтеза жирной кислоты D-3-гидроксибутирил-S-АПБ дегидратируется под действием 3-гидроксиацил-АПБ— дегидратазы с образованием транс- -бутеноил-S-АПБ (рис. 21-10).
-бутеноил-S-АПБ (рис. 21-10).

г. Насыщение
На четвертом этапе, завершающем один цикл реакций, осуществляемых синтазным комплексом для жирных кислот, двойная связь транс- -бутеноил-S-АПБ восстанавливается, или насыщается, под действием еноил-АПБ—редуктазы с образованием бутирил-S-АПБ (рис. 21-10).
-бутеноил-S-АПБ восстанавливается, или насыщается, под действием еноил-АПБ—редуктазы с образованием бутирил-S-АПБ (рис. 21-10).

Рис. 21-10. Три последовательных этапа цикла синтеза жирной кислоты. По окончании последней реакции бутирильная группа переносится на SH-группу цнстеина. После этого Фп—SH-группа опять готова соединиться со следующей вступающей в реакцию малонильной группой малонил-СоА.
Процесс наращивания цепи заканчивается на 16-м углеродном атоме, после чего под действием гидролитического фермента молекула пальмитиновой кислоты отщепляется от молекулы АПБ.

Отметим, что для синтеза пальмитиновой кислоты необходимы два вида химической энергии энергия фосфатной группы АТР и восстановительный потенциал NADPH. АТР нужен для образования тиоэфирной связи в ацетил-СоА и построения малонил-СоА путем присоединения  к ацетил-СоА, тогда как NADP используется для восстановления двойных связей.
к ацетил-СоА, тогда как NADP используется для восстановления двойных связей.
NADPH, который необходим для восстановительных реакций, протекающих в ходе биосинтеза жирной кислоты, в разных клетках поступает из двух различных источников В печени NADPH образуется главным образом в реакциях пентозофосфатного пути (разд. 16.13), в основном под действием глюкозо-6-фосфат—дегидрогеназы

В жировых клетках (адипоцитах) NADPH образуется преимущественно в результате действия малат-дегидрогеназы

Обе эти реакции, приводящие к образованию NADPH, протекают в цитозоле, в котором молярное отношение NADPH/NADP+ очень велико (около 75), что обеспечивает высокий восстановительный потенциал, необходимый для синтеза жирных кислот. Напротив, молярное отношение NADH/NAD+ в цитозоле значительно меньше (оно составляет приблизительно 8-10 4). NADPH в цитозоле прекрасно приспособлен для роли первичного донора водородных атомов, используемых в ходе восстановительных биосинтетических реакции.
Суммируя все изложенное выше, можно сказать, что ферментативный биосинтез пальмитиновой кислоты отличается от ее ферментативного окисления 1) внутриклеточной локализацией; 2) природой переносчика ацильных групп; 3) формой, в которой двухуглеродные единицы присоединяются к цепи жирной кислоты или отщепляются от нее; 4) стереоконфигурацией промежуточного  -гидроксиацильного соединения; 5) типом пиридинового нуклеотида, используемого в восстановительных реакциях, и 6) участием
-гидроксиацильного соединения; 5) типом пиридинового нуклеотида, используемого в восстановительных реакциях, и 6) участием  Перечисленные различия (табл. 21-1) показывают, что эти два противоположно направленных процесса характеризуются разными физическими и химическими параметрами.
Перечисленные различия (табл. 21-1) показывают, что эти два противоположно направленных процесса характеризуются разными физическими и химическими параметрами.
Таблица 21-1. Различия между ферментативным биосинтезом и ферментативным окислением пальмитиновой кислоты





























































































































































































































































































































































 в малонильной группе на
в малонильной группе на  -АПБ. В результате получается шестиуглеродная ацильная группа, ковалентно связанная с —
-АПБ. В результате получается шестиуглеродная ацильная группа, ковалентно связанная с — -группой фосфопантетеина. В ходе трех следующих этапов синтазного цикла
-группой фосфопантетеина. В ходе трех следующих этапов синтазного цикла  -кетогруппа образовавшейся ацильной группы восстанавливается и образуется шестиуглеродная насыщенная ацильная группа подобно тому, как это происходит в предыдущем цикле реакций.
-кетогруппа образовавшейся ацильной группы восстанавливается и образуется шестиуглеродная насыщенная ацильная группа подобно тому, как это происходит в предыдущем цикле реакций.

 -группой цистеина. Вступающая в реакцию малонильная группа связывается с
-группой цистеина. Вступающая в реакцию малонильная группа связывается с  -группой. На этапе конденсации бутирильная группа, соединенная с
-группой. На этапе конденсации бутирильная группа, соединенная с  -группой, обменивается на свободную карбоксильную группу малонильного остатка, которая при этом отщепляется в виде
-группой, обменивается на свободную карбоксильную группу малонильного остатка, которая при этом отщепляется в виде  (на красном фоне). В образовавшемся продукте - шестиуглеродной
(на красном фоне). В образовавшемся продукте - шестиуглеродной  -кетоацильной группе-четыре атома углерода происходят из
-кетоацильной группе-четыре атома углерода происходят из  а остальные два - из
а остальные два - из  с которого начиналась реакция.
с которого начиналась реакция.
 -группы фосфопантетеина на
-группы фосфопантетеина на  -группу цистеина.
-группу цистеина.
 -АПБ.
-АПБ.
