Креатинфосфат может передавать свою фосфатную группу на ADP в реакции, катализируемой креатинкиназой:

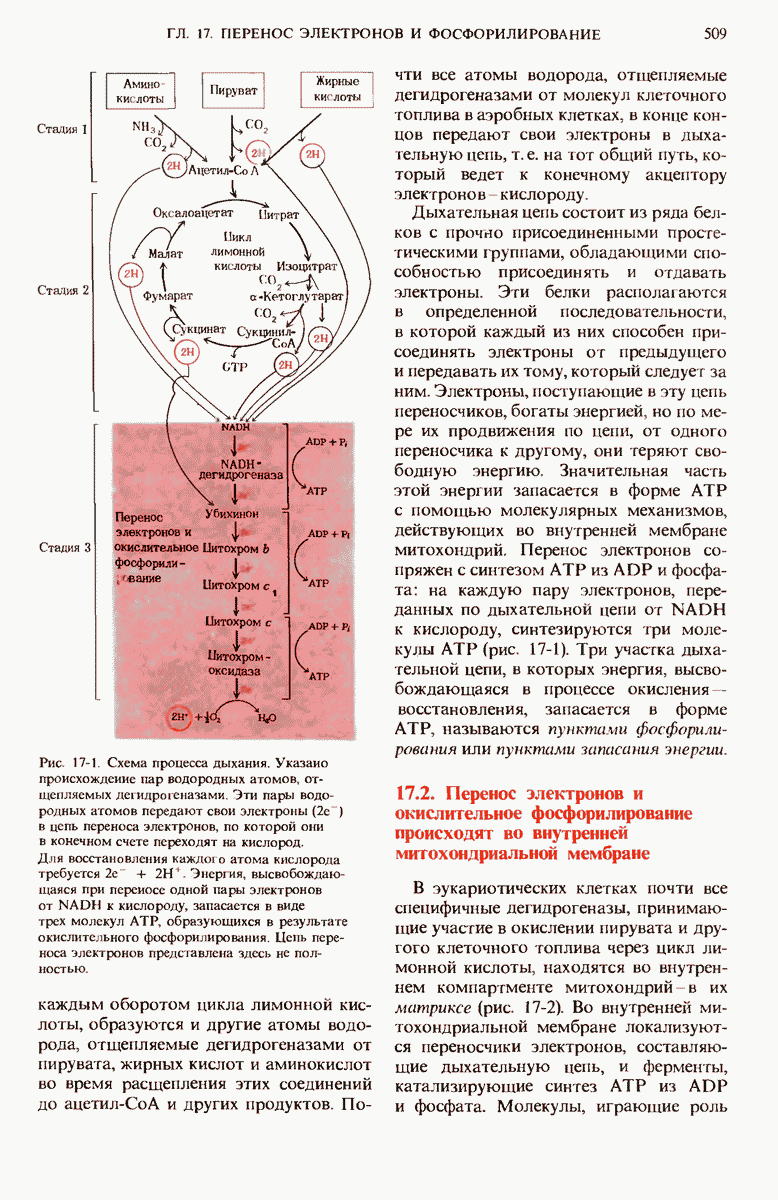
Благодаря креатинфосфату концентрация АТР в мышечных клетках поддерживается на постоянном и притом довольно высоком уровне. Особенно это существенно для скелетных мышц, работающих с перерывами, но иногда очень напряженно с большой скоростью. Всякий раз, когда часть АТР мышечной клетки расходуется на сокращение, в результате гидролиза АТР образуется ADP. Креатинфосфат при участии креатинкиназы быстро передает свою фосфатную группу молекулам ADP, и нормальный уровень АТР восстанавливается. Содержание креатинфосфата в мышцах в 3-4 раза превышает содержание АТР (табл. 14-4); поэтому в форме креатинфосфата может храниться достаточное количество фосфатных групп, полностью обеспечивающее поддержание постоянного уровня АТР в короткие периоды усиленной мышечной активности.

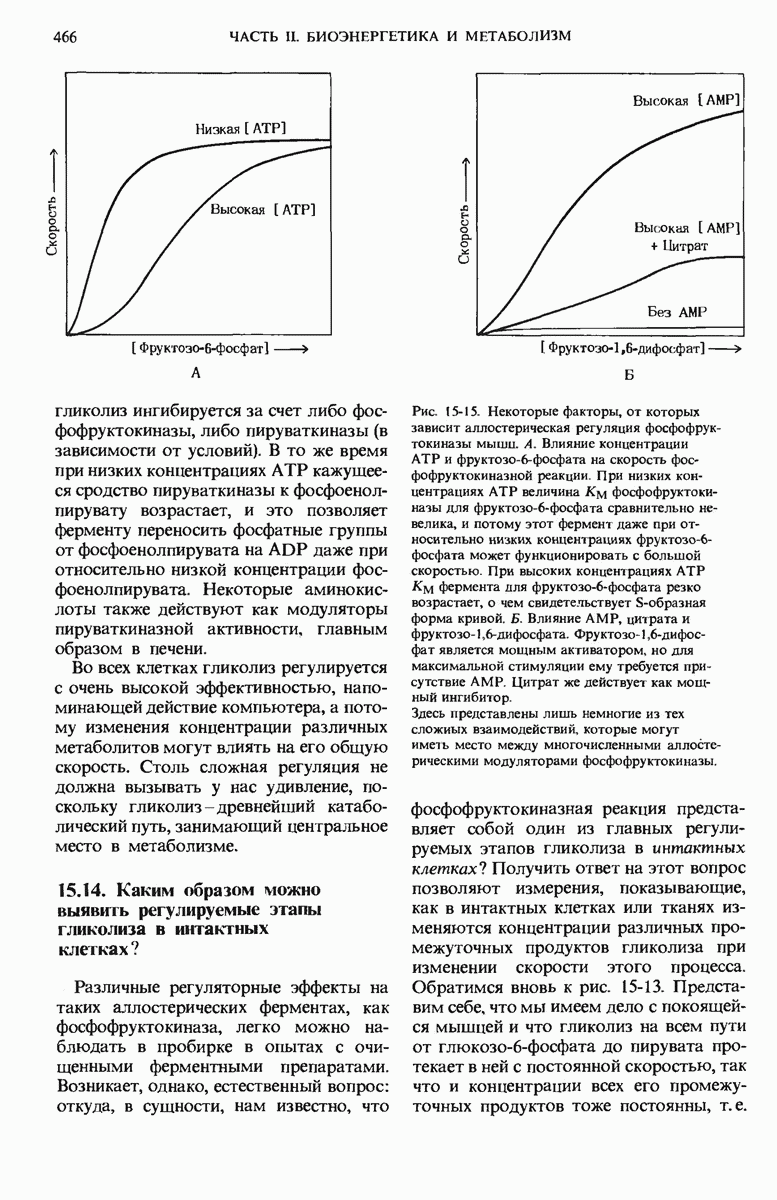
Рис. 14-12. В ресничках и жгутиках эукариотических клеток механическая сила развивается за счет использования АТР. А. Поперечный разрез реснички. Эти структуры состоят из девяти пар микротрубочек, образующих наружное кольцо, и двух одиночных центральных микротрубочек (расположение по типу «9 + 2»; разд. 2.16). Реснички окружены оболочкой, представляющей собой вырост клеточной мембраны. Энергию для характерных движений ресничек (волнообразного, скользящего или вращательного) поставляет гидролиз АТР. Эти движения осуществляются ресничками за счет скольжения или скручнвання парных микротрубочек, которое весьма напоминает наблюдаемое в скелетных мышцах АТР-зависимое скольжение толстых и тонких нитей друг относительно друга. От наружных (парных) микротрубочек отходят находящиеся на равном расстоянии друг от друга отростки, или выступы, напоминающие миозиновые головки в толстых нитях мышц. Эти выступы состоят из молекул динеина - довольно крупного белка, обладающего АТРазной активностью. Катализируемый динеином гилролиз АТР поставляет энергию для механического движения - скольжения или скручивания микротрубочек. Было высказано предположение, что центральные микротрубочки регулируют скорость движения ресничек. Б. Отдельные фазы биения реснички в жабрах морского червя, у которого реснички имеют длину около 30 мкм. Эти характерные движения сообщает ресничкам АТР-зависимое скольжение трубчатых нитей друг относительно друга.

Рис. 14-13. Креатинфосфат в мышцах играет роль запасного донора высокоэнергетических фосфатных групп. Он действует как своеобразный буфер, обеспечивающий постоянство концентрации АТР.
Благодаря обратимости креатинкиназной реакции накопившийся креатин в период восстановления вновь фосфорилируется за счет АТР до креатинфосфата. Поскольку другого метаболического пути для образования и расщепления креатинфосфата не существует, это соединение хорошо приспособлено для выполнения своей функции - резервуара фосфатных групп.
В мышцах многих беспозвоночных роль носителя резервной формы энергии выполняет не креатинфосфат, а аргипипфосфат. Соединения, служащие, подобно креатинфосфату и аргининфосфату, запасными источниками энергии, носят название фосфагенов.
14.16. АТР поставляет энергию также и для активного транспорта через мембраны
Химическая энергия АТР используется также и для выполнения осмотической работы, т.е. работы, необходимой для переноса каких-либо ионов или молекул через мембрану из одного компартмента в другой, в котором их концентрация выше. Мы можем рассчитать количество свободной энергии, необходимое для переноса 1 моль неионизованного растворенного вещества через мембрану, например из окружающей среды в клетку, если нам известны концентрации растворенного вещества в несвязанной форме в окружающей среде и в клетке (рис. 14-14). Для такого расчета воспользуемся общим уравнением

где  - молярная концентрация данного растворенного вещества в окружающей среде,
- молярная концентрация данного растворенного вещества в окружающей среде,  - его молярная концентрация в клетке, R - газовая постоянная и Т - абсолютная температура. Пользуясь этим уравнением, можно определить количество свободной энергии, необходимое для того, чтобы переместить 1 моль глюкозы против стократного градиента концентрации, например из среды с исходной концентрацией глюкозы
- его молярная концентрация в клетке, R - газовая постоянная и Т - абсолютная температура. Пользуясь этим уравнением, можно определить количество свободной энергии, необходимое для того, чтобы переместить 1 моль глюкозы против стократного градиента концентрации, например из среды с исходной концентрацией глюкозы  в компартмент, где ее конечная концентрация составит
в компартмент, где ее конечная концентрация составит  . Подставляя в уравнение соответствующие значения, получаем
. Подставляя в уравнение соответствующие значения, получаем


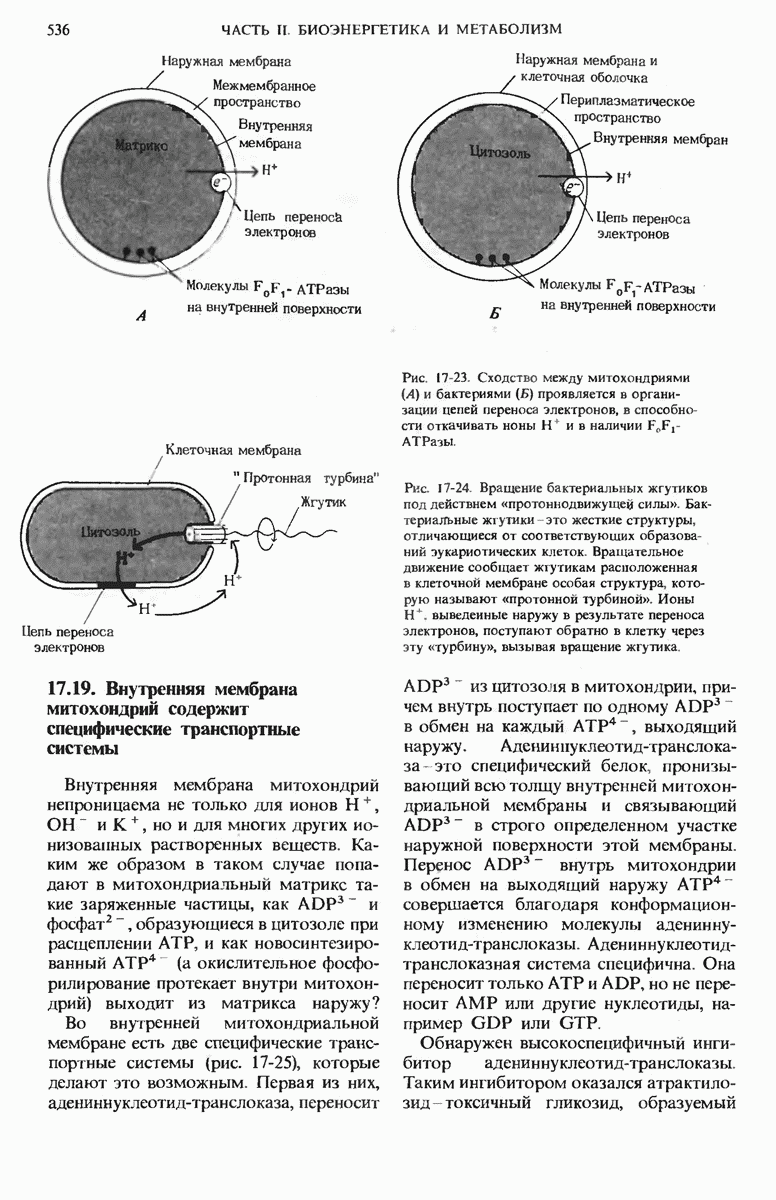
Рис. 14-14. Активный транспорт растворенного вещества против градиента концентрации. Начиная с момента равновесия, т.е. с того момента. когда концентрации данного растворенного вещества в обоих компартментах одинаковы, активный транспорт вещества из одного компартмента в другой обеспечивает его перемещение против градиента концентрации. Для создания и поддержания градиента концентрации какого-либо растворенною вещества между компартментами, находящимися по обе стороны мембраны, требуется затрата свободной энергии. Если энергия почему-либо перестает поступать, то вещество из компартмента с более высокой его концентрацией начинает диффундировать обратно, и диффузия продолжается до тех пор. пока снова не установится равновесие, т. е. пока концентрации вещества по обе стороны мембраны не сравняются.
Изменение свободной энергии выражается в этом случае положительной величиной, и это значит, что 2,72 ккал свободной энергии, которые требуются для переноса 1 моля глюкозы (или любого нейтрального вещества) против стократного градиента концентрации, должны быть переданы системе за счет какой-то сопряженной реакции, способной служить источником энергии.
Градиенты концентрации между двумя сторонами клеточных мембран (трансмембранные градиенты) варьируют очень сильно. Пожалуй, максимальный градиент концентрации в организме поддерживается плазматической мембраной обкладочных клеток слизистой оболочки желудка, секретирующих соляную кислоту в желудочный сок. Концентрация  в желудочном соке может достигать
в желудочном соке может достигать  тогда как концентрация ионов
тогда как концентрация ионов  в клетках составляет приблизительно
в клетках составляет приблизительно  . Это означает, что обкладочные клетки обладают способностью секретировать ионы водорода даже против градиента порядка
. Это означает, что обкладочные клетки обладают способностью секретировать ионы водорода даже против градиента порядка  . По-видимому, эти клетки имеют какие-то очень активные мембранные «насосы» для секреции ионов водорода, так как для поддержания столь высокого градиента концентрации требуется значительное количество энергии. Перенос веществ через мембраны против градиента концентрации называют активным транспортом. Образование желудочной
. По-видимому, эти клетки имеют какие-то очень активные мембранные «насосы» для секреции ионов водорода, так как для поддержания столь высокого градиента концентрации требуется значительное количество энергии. Перенос веществ через мембраны против градиента концентрации называют активным транспортом. Образование желудочной  стимулируется особым, связанным с мембраной ферментом - так называемой
стимулируется особым, связанным с мембраной ферментом - так называемой  -транспортирующей АТРазой. При образовании желудочного сока на каждую молекулу цитозольного АТР, гидролизованного до ADP и фосфата, из цитозоля наружу через плазматическую мембрану выводятся два иона
-транспортирующей АТРазой. При образовании желудочного сока на каждую молекулу цитозольного АТР, гидролизованного до ADP и фосфата, из цитозоля наружу через плазматическую мембрану выводятся два иона  .
.
Другим важным примером активного транспорта может служить перенос ионов  через плазматическую мембрану во всех животных клетках. Лучше всего изучен этот процесс в эритроцитах. Установлено, что концентрация
через плазматическую мембрану во всех животных клетках. Лучше всего изучен этот процесс в эритроцитах. Установлено, что концентрация  в цитозоле эритроцитов достигает примерно
в цитозоле эритроцитов достигает примерно  тогда как в плазме крови она составляет всего
тогда как в плазме крови она составляет всего  . В то же время концентрация
. В то же время концентрация  в плазме крови достигает
в плазме крови достигает  а в эритроцитах она равна приблизительно
а в эритроцитах она равна приблизительно  . Для поддержания столь высоких трансмембранных градиентов требуется энергия АТР. В мембране эритроцита содержится специализированный фермент, получивший название
. Для поддержания столь высоких трансмембранных градиентов требуется энергия АТР. В мембране эритроцита содержится специализированный фермент, получивший название  -транспортирующей АТРазы, который функционирует и как фермент, и как молекулярный насос. Эта АТРаза катализирует гидролитическое расщепление АТР до ADP и фосфата, а высвобождающуюся при этом свободную энергию использует для перекачивания ионов
-транспортирующей АТРазы, который функционирует и как фермент, и как молекулярный насос. Эта АТРаза катализирует гидролитическое расщепление АТР до ADP и фосфата, а высвобождающуюся при этом свободную энергию использует для перекачивания ионов  из окружающей среды внутрь клетки, а ионов
из окружающей среды внутрь клетки, а ионов  из клетки в окружающую среду (рис. 14-15). Стадией, на которой происходит передача энергии в этом процессе, является перенос концевой фосфатной группы АТР на молекулу
из клетки в окружающую среду (рис. 14-15). Стадией, на которой происходит передача энергии в этом процессе, является перенос концевой фосфатной группы АТР на молекулу  -АТРазы.
-АТРазы.

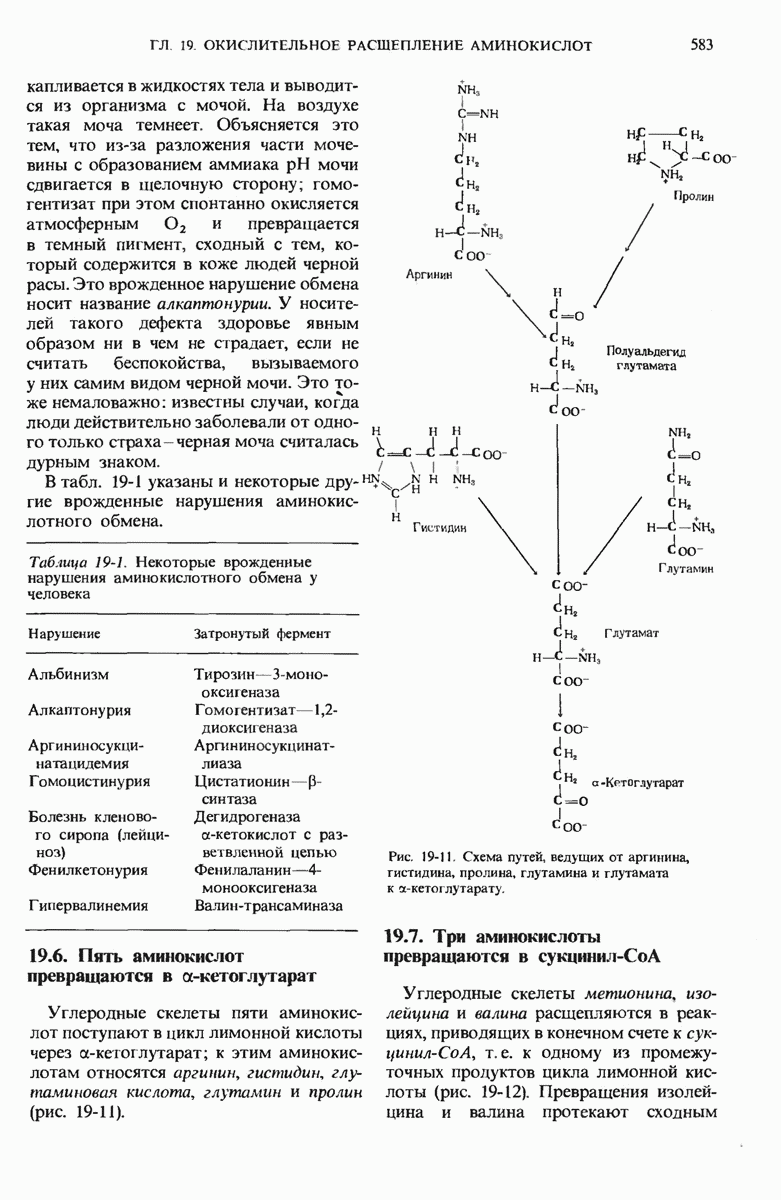
Рис. 14-15. Схема, поясняющая действие  -АТРазы. Для транспорта
-АТРазы. Для транспорта  в клетку (где его концентрация выше, чем в окружающей среде) и транспорта
в клетку (где его концентрация выше, чем в окружающей среде) и транспорта  из клетки в окружающую среду (где концеш рация этих ионов выше, чем в клетке) требуется свободная энергия. Источником ее служит гидролиз АТР. На каждую молекулу АТР, гидролизованного до ADP и
из клетки в окружающую среду (где концеш рация этих ионов выше, чем в клетке) требуется свободная энергия. Источником ее служит гидролиз АТР. На каждую молекулу АТР, гидролизованного до ADP и  из клетки выходят три иона
из клетки выходят три иона  и два иона
и два иона  поступают в нее из окружающей среды. Этот транспорт ионов включает два этапа. На первом этапе молекула АТРазы фосфорилируется под действием АТР. и это позволяет ей присоединить ион
поступают в нее из окружающей среды. Этот транспорт ионов включает два этапа. На первом этапе молекула АТРазы фосфорилируется под действием АТР. и это позволяет ей присоединить ион  На втором этапе присоединяется ион
На втором этапе присоединяется ион  следствием чего оказывается перенос
следствием чего оказывается перенос  и К через мембрану с отщеплением свободного фосфата, поступающего в цитозоль. АТР и продукты его гидролиза (ADP и
и К через мембрану с отщеплением свободного фосфата, поступающего в цитозоль. АТР и продукты его гидролиза (ADP и  ) остаются в клетке.
) остаются в клетке.
На следующей стадии связанная с ферментом фосфатная группа отщепляется и в виде неорганического фосфата переходит в цитозоль, обеспечивая энергией перенос  через мембрану в противоположных направлениях, но в обоих случаях против градиента концентрации. Продукты этого процесса, ADP и фосфат, вновь «заряжаются», т.е. образуют АТР, за счет энергии, выделяющейся при расщеплении глюкозы.
через мембрану в противоположных направлениях, но в обоих случаях против градиента концентрации. Продукты этого процесса, ADP и фосфат, вновь «заряжаются», т.е. образуют АТР, за счет энергии, выделяющейся при расщеплении глюкозы.  -АТРаза гидролизует АТР лишь в том случае, если в клетке имеются ионы
-АТРаза гидролизует АТР лишь в том случае, если в клетке имеются ионы  а в окружающей среде - ионы
а в окружающей среде - ионы  . Молекула
. Молекула  -АТРазы, состоящая из двух
-АТРазы, состоящая из двух  и двух
и двух  -субъединиц, насквозь пронизывает плазматическую мембрану. Во время переноса ионов она претерпевает конформационное изменение. В почках, где ионы
-субъединиц, насквозь пронизывает плазматическую мембрану. Во время переноса ионов она претерпевает конформационное изменение. В почках, где ионы  переходят в мочу, а ионы
переходят в мочу, а ионы  должны удерживаться в крови, на активный перенос этих ионов расходуется почти две трети всего АТР, образующегося в процессе дыхания. В различных клеточных мембранах важную роль играют АТРазы, транспортирующие другие ионы (табл. 14-6).
должны удерживаться в крови, на активный перенос этих ионов расходуется почти две трети всего АТР, образующегося в процессе дыхания. В различных клеточных мембранах важную роль играют АТРазы, транспортирующие другие ионы (табл. 14-6).


























































































































































































































































































































































 гидролиза креатинфосфата
гидролиза креатинфосфата  несколько превышает
несколько превышает  гидролиза АТР.
гидролиза АТР.
