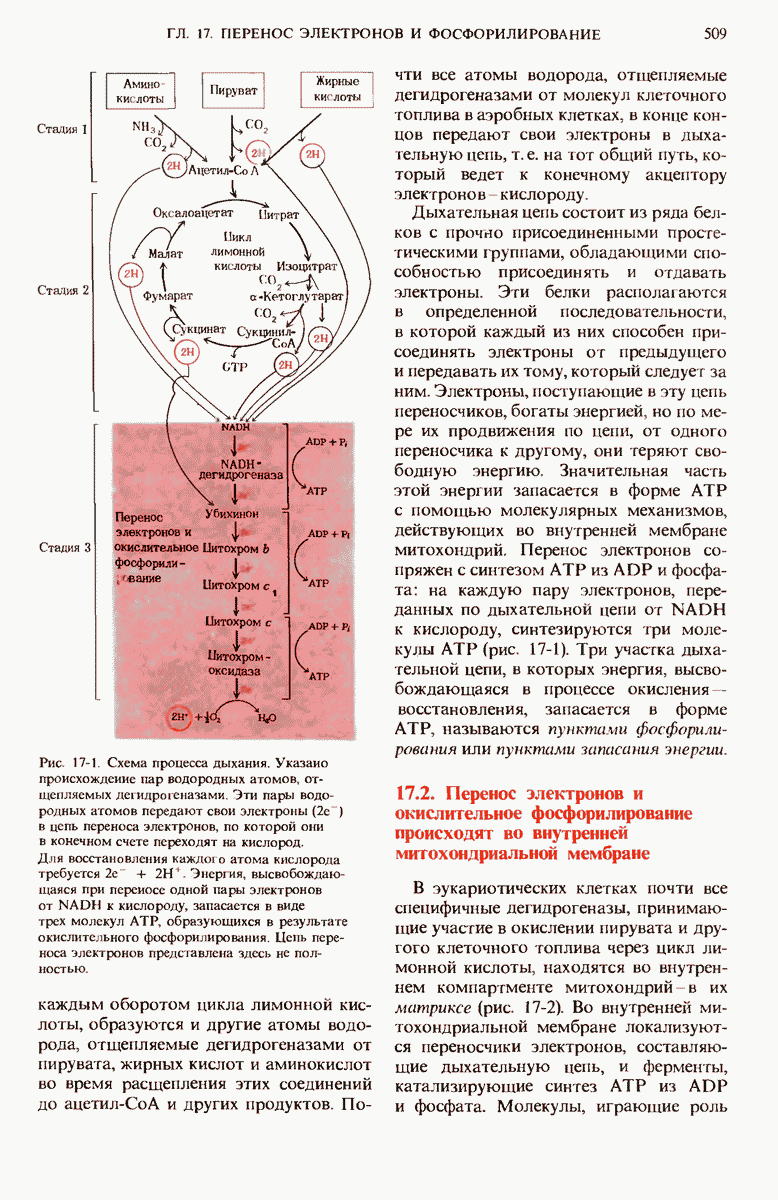
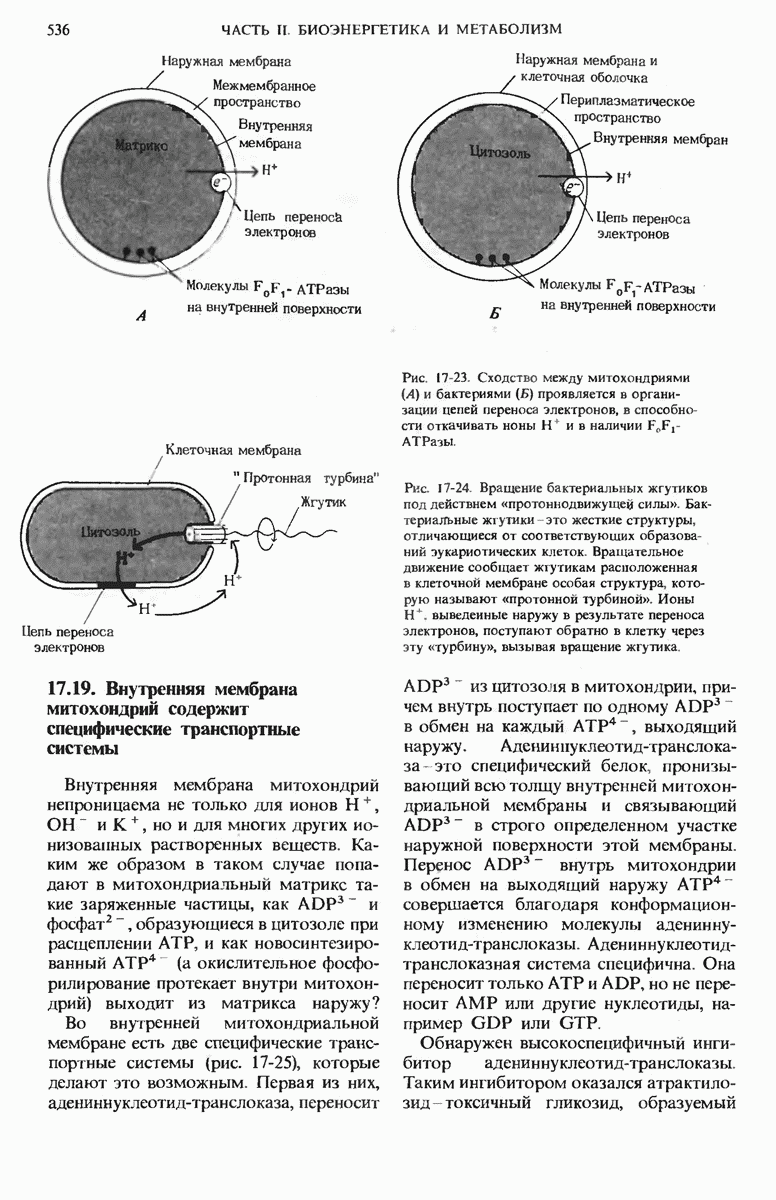
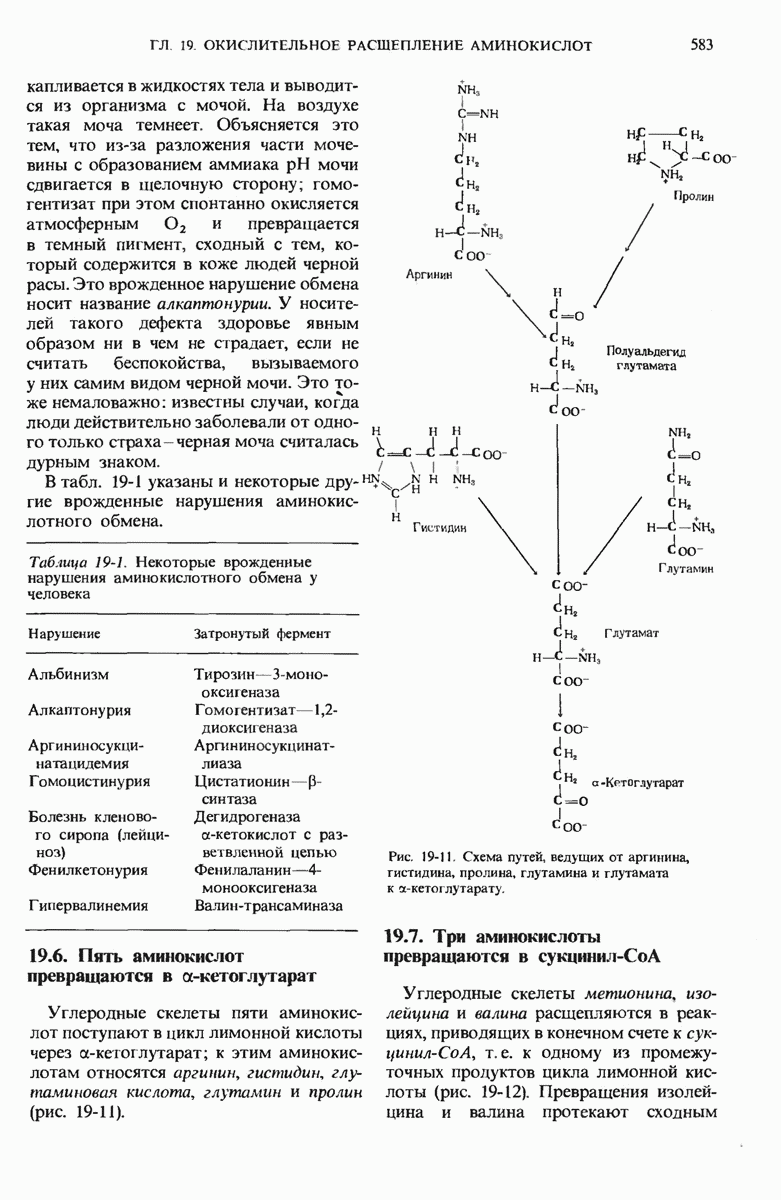
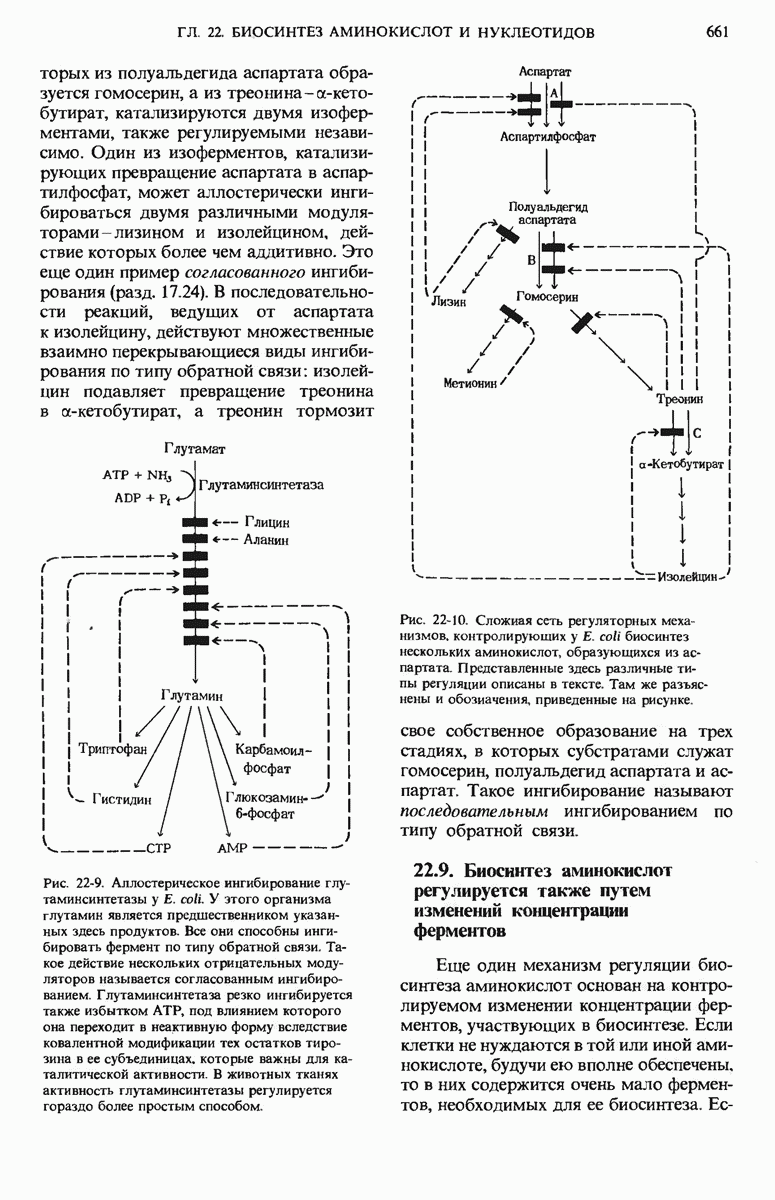
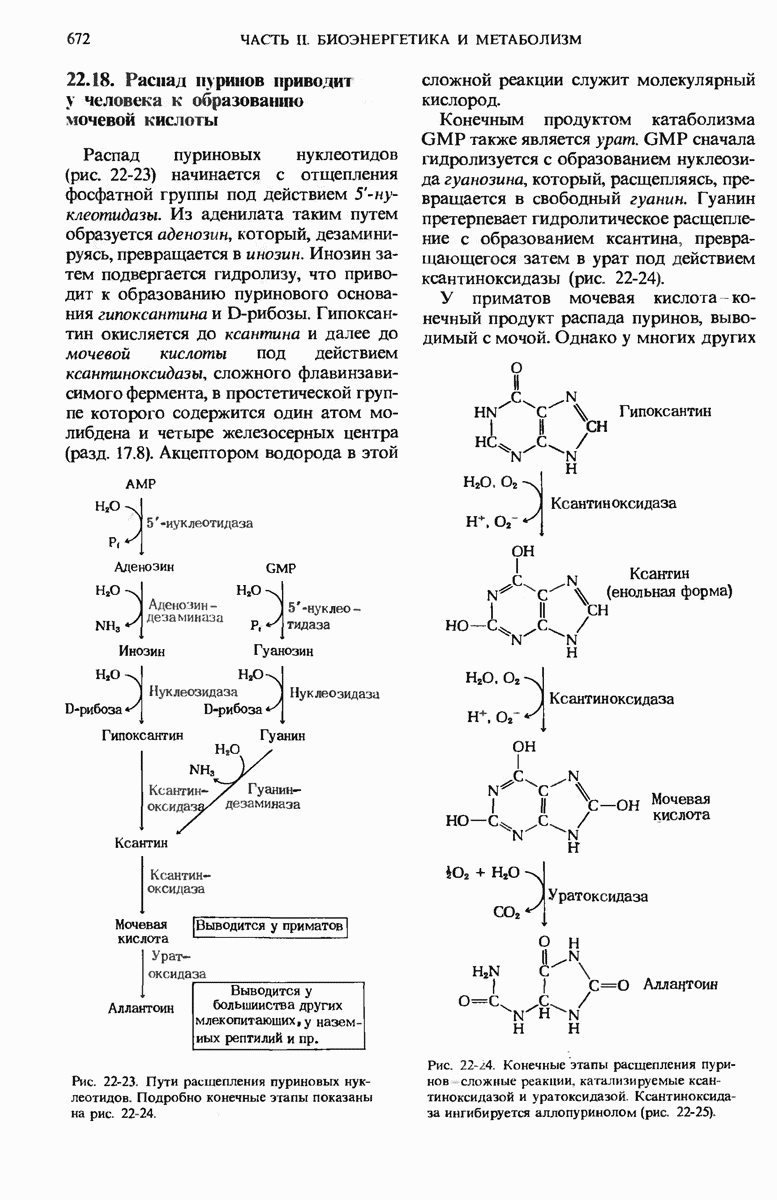
16.9. Превращение пирувата в ацетил-СоА регулируется
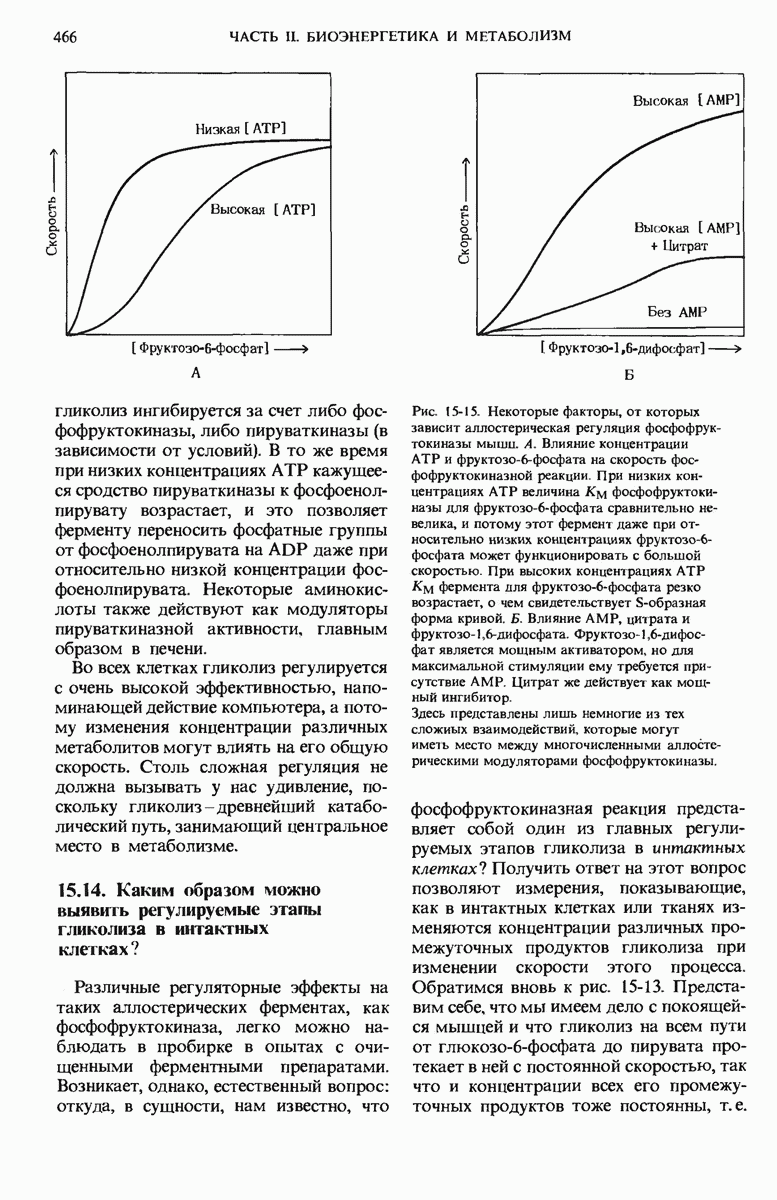
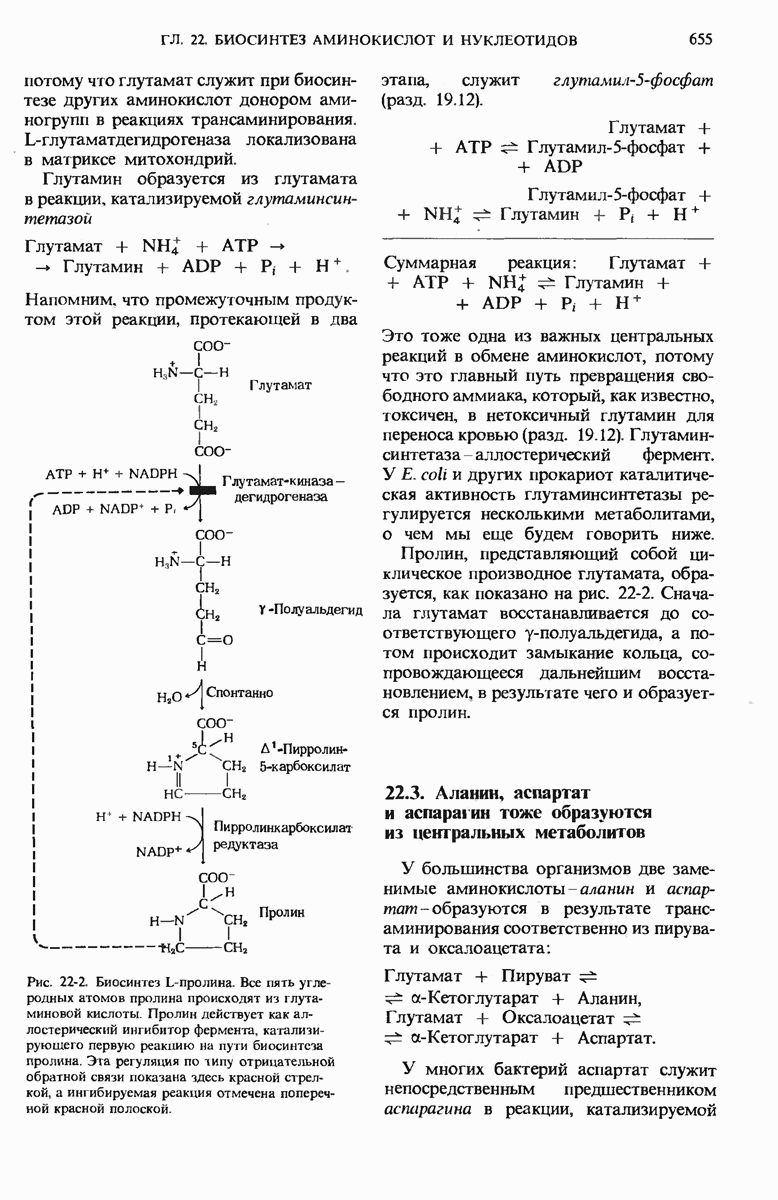
Из гл. 15 мы знаем, что скорость гликолиза регулируется на двух уровнях. В первую очередь регулируется, как мы видели, сама подача «топлива» для гликолиза. В этой регуляции участвуют два регуляторных фермента, контролирующих вхождение глюкозы в последовательность гликолитических реакций, — гексокиназа, катализирующая фосфорилирование D-глюкозы с образованием глюкозо-6-фосфата, и гликогенфосфорилаза, катализирующая первый этап образования глюкозо-6-фосфата из гликогена. Скорость цикла лимонной кислоты регулируется также прежде всего за счет скорости образования «топлива» для этого цикла. Этим «топливом» является ацетил-СоА, образующийся при окислении пирувата и жирных кислот (гл. 18). На рис. 16-14 показано, как реакция образования ацетил-СоА, катализируемая пируватдегидрогеназным комплексом. регулируется в животных тканях при помощи ковалентной модификации этого комплекса (разд. 9.22). Когда концентрация АТР в митохондриях относительно велика и когда ацетил-СоА, а также промежуточные продукты цикла Кребса имеются в достаточном количестве, обеспечивающем удовлетворение энергетических нужд клетки, дальнейшее образование ацетил-СоА приостанавливается.
В этих условиях, которые служат сигналом для такой приостановки, АТР является положительным модулятором, активирующим вспомогательный фермент - киназу пируватдегидрогеназы. Этот фермент использует АТР для фосфорилирования остатка серина в активном центре молекулы пируватдегидрогеназы, в результате чего образуется неактивная форма фермента - фосфопируватдегидрогеназа (рис. 16-14). Если, однако, потребность в АТР возрастает и уровень АТР соответственно снижается, то неактивная, фосфорилированная, форма пируватдегидрогеназы может быть вновь активирована. Это происходит в результате гидролитического отщепления от молекулы пируватдегидрогеназы ингибирующей фосфатной группы. Катализирует эту реакцию другой фермент - фосфатаза фосфопируватдегидрогеназы. Стимулирующее действие на этот фермент оказывает повышение концентрации ионов  играющих роль важного метаболического посредника; концентрация ионов
играющих роль важного метаболического посредника; концентрация ионов  увеличивается всякий раз, когда возникает потребность в АТР. Киназа пируватдегидрогеназы и фосфатаза фосфопируватдегидрогеназы присутствуют в пируватдегидрогеназном комплексе. Этот комплекс, следовательно, представляет собой очень сложную, независимую и саморегулирующуюся систему.
увеличивается всякий раз, когда возникает потребность в АТР. Киназа пируватдегидрогеназы и фосфатаза фосфопируватдегидрогеназы присутствуют в пируватдегидрогеназном комплексе. Этот комплекс, следовательно, представляет собой очень сложную, независимую и саморегулирующуюся систему.
Пируватдегидрогеназный комплекс регулируется также путем аллостерической модуляции.

Рис. 16-14. Регуляция пируватдегидрогеназной реакции путем ковалентного взаимопревращения активной и неактивной форм фермента. Ионы  способствуют образованию активной формы фермента, стимулируя фосфатазу фосфопируватдегидрогеназы.
способствуют образованию активной формы фермента, стимулируя фосфатазу фосфопируватдегидрогеназы.
Сильное ингибирующее действие оказывают на него (помимо АТР) ацетил-СоА и NADH, которые являются продуктами пируватдегидрогеназной реакции и в то же время играют роль аллостерических ингибиторов этой системы. Аллостерическое ингибирование окисления пирувата резко усиливается в присутствии высокомолекулярных жирных кислот; позже мы узнаем (гл. 18), что жирные кислоты тоже служат источником ацетил-СоА. Таким образом, каталитическая активность пируватдегидрогеназного комплекса выключается в тех случаях, когда в клетках имеется достаточно топлива в виде жирных кислот и ацетил-СоА или когда в них повышаются концентрация АТР и отношение NADH/NAD+.