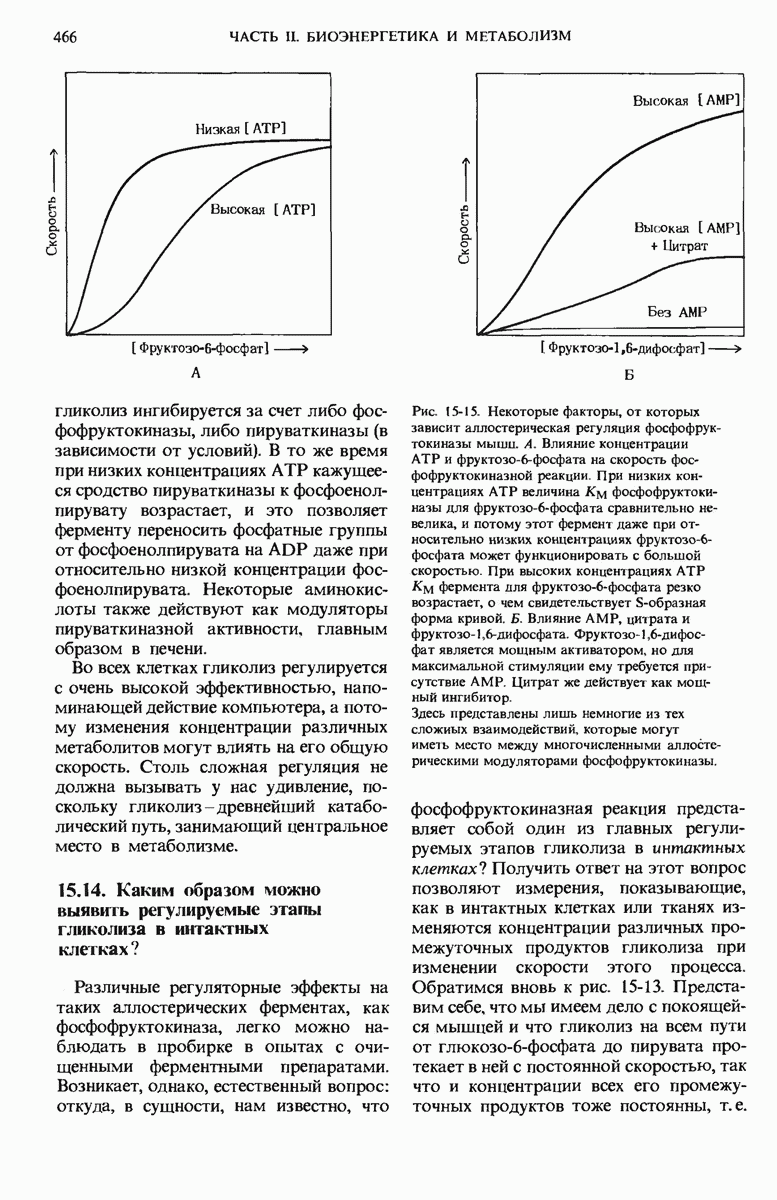
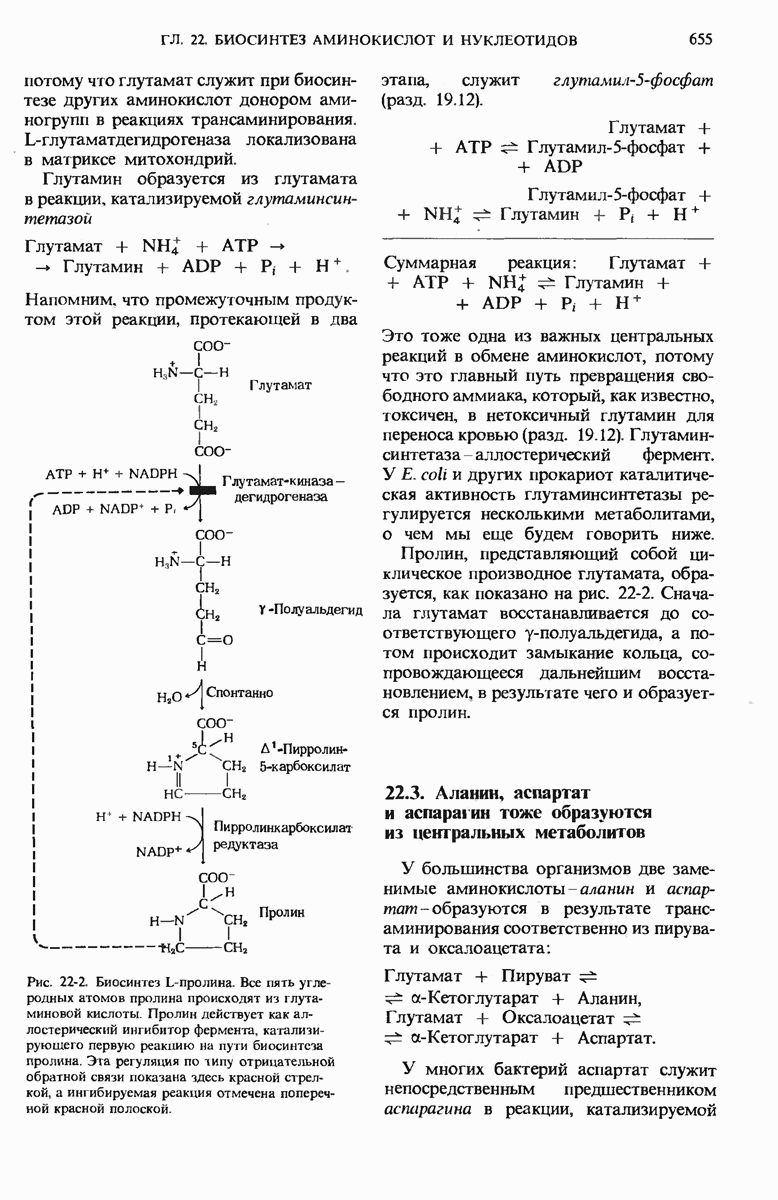
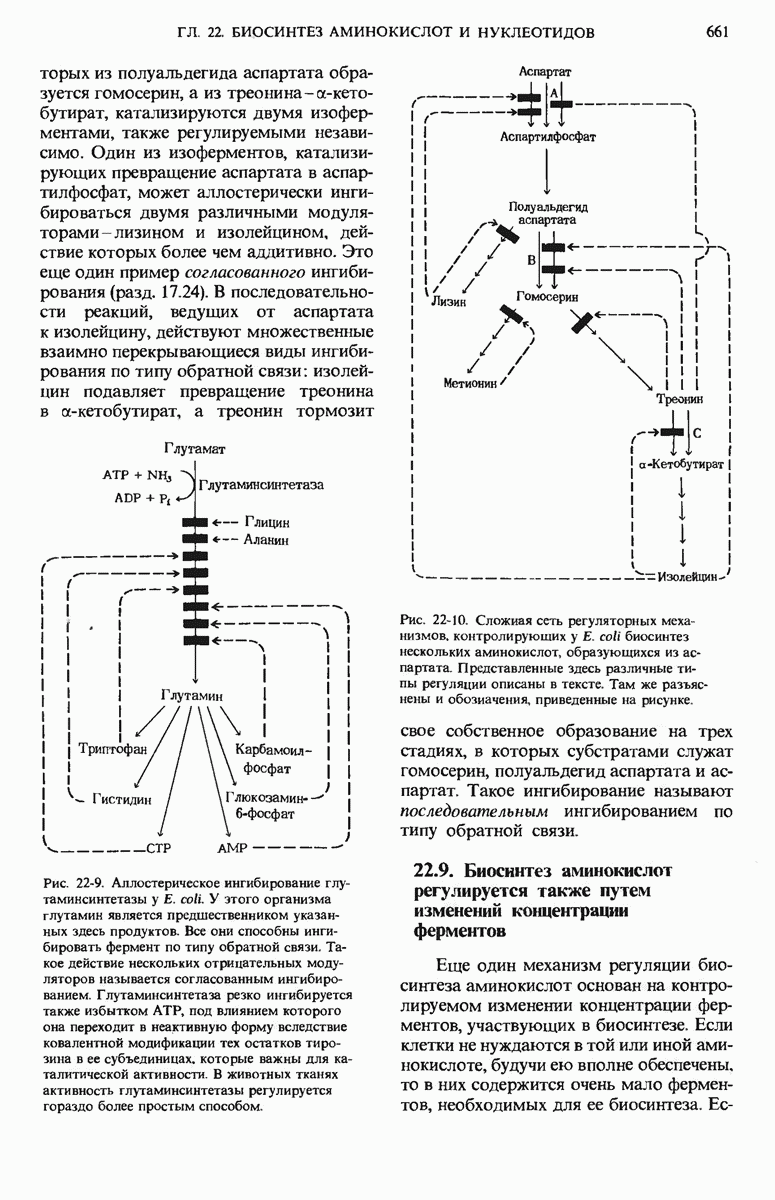
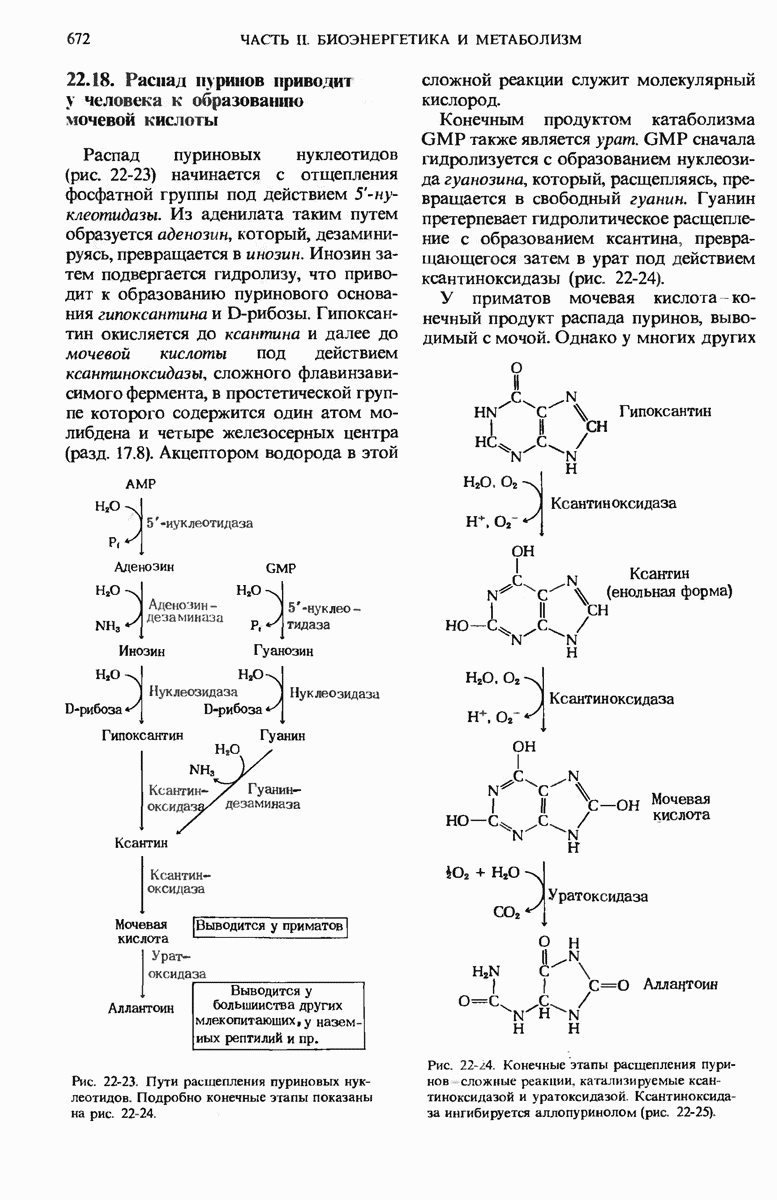
22.15. Пиримидиновые нуклеотиды синтезируются из аспартата и рибозофосфата
Обычные пиримидиновые нуклеотиды - это цитидин-5-монофосфат ( СМР), или цитидилат, и уридин-5-монофосфат (UMP), или уридилат; в их состав входят пиримидиновые основания - соответственно цитозин и урацил. Биосинтез пиримидиновых нуклеотидов (рис. 22-19) несколько отличается от синтеза пуриновых нуклеотидов; в этом случае сначала образуется шестичленное пиримидиновое кольцо, а затем к нему присоединяется рибозофосфат. Для этого процесса требуется карбамоилфосфат, играющий роль промежуточного продукта также и в цикле мочевины (разд. 19.7). Однако карбамоилфосфат, необходимый для синтеза мочевины, образуется в митохондриях при участии митохондриального фермента карбамоилфосфат-синтетазы I, а карбамоилфосфат, участвующий в биосинтезе пиримидинов, образуется в цитозоле под действием другой формы того же фермента - карбамоилфосфат-синтетазы II. Цитозольный карбамоилфосфат взаимодействует с аспартатом, в результате чего получается N-карбамоиласпартат. Эта реакция катализируется аспартат-транс-карбамоилазой, одним из наиболее тщательно изученных аллостерических ферментов (дополнение 9-5); о ней мы еще будем говорить ниже. При отщеплении молекулы воды от N-карбамоиласпарта-та под действием дигидрооротазы пиримидиновое кольцо замыкается; в результате возникает L-дигидрооротат, который затем окисляется с образованием оротовой кислоты, представляющей собой производное пиримидина.
Рис. 22-19. (см. скан) Путь биосинтеза UTP и СТР (через образование оротидиловой кислоты). PRPP-5-фосфорибозил-1-пирофосфат.
Конечным акцептором электронов в этой реакции служит NAD+.
На этой стадии D-рибозо-5-фосфатная боковая цепь, поставляемая 5-фосфорибозил-1-пирофосфатом, присоединяется к оротату, в результате чего образуется оротидиловая кислота. Оротидилат декарбоксилируется и образовавшийся уридилат, фосфорилируясь, превращается в UTP; последний присоединяет аминогруппу от глутамина и превращается в цитидинтрифосфат (СТР) (рис. 22-19).