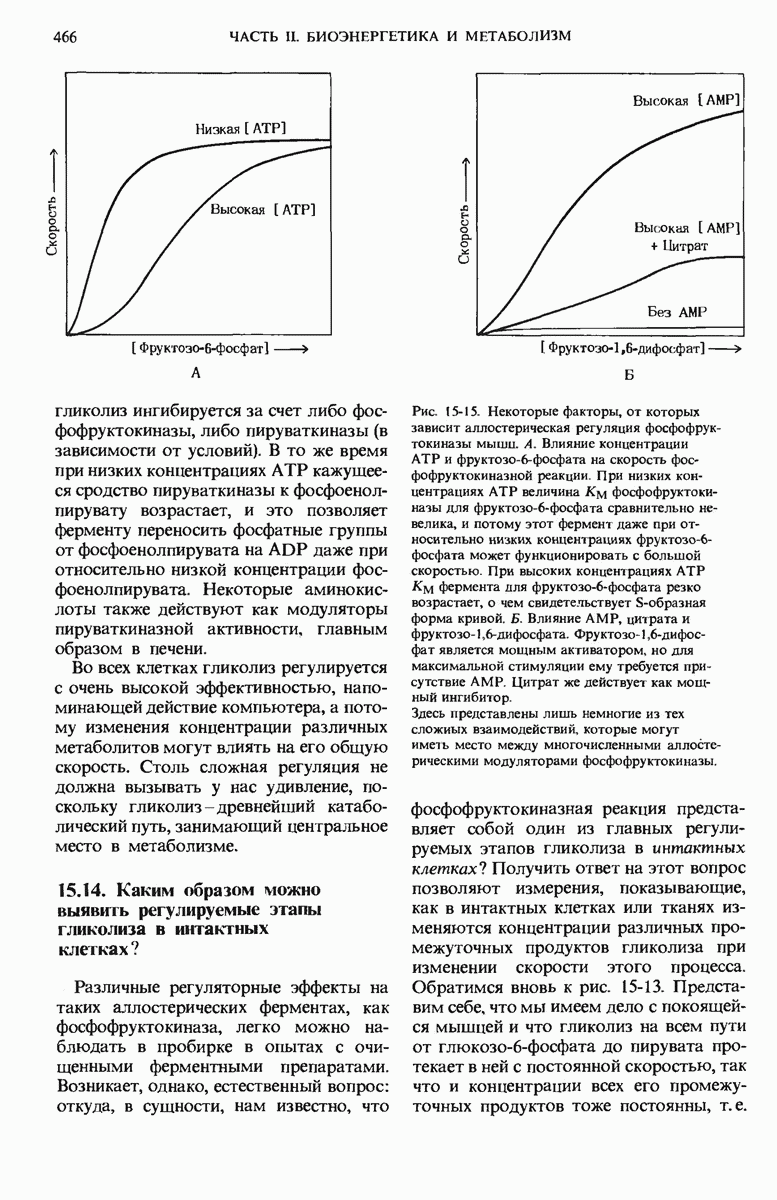
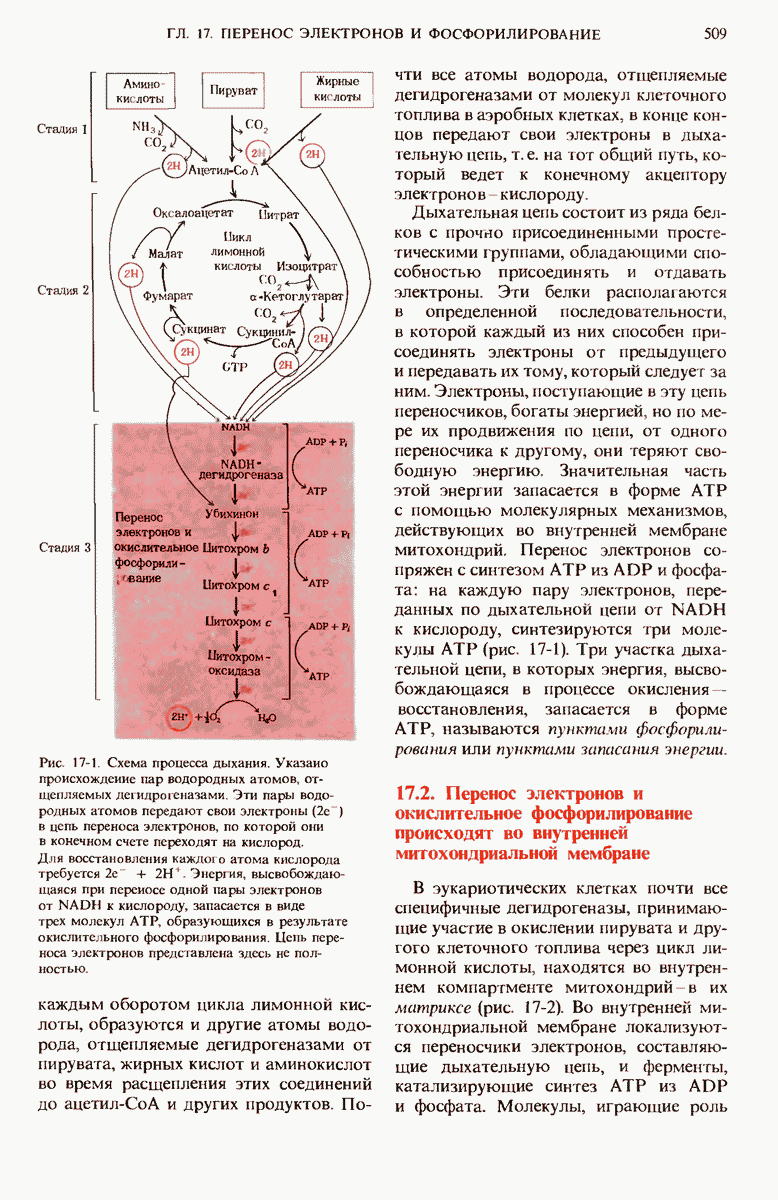
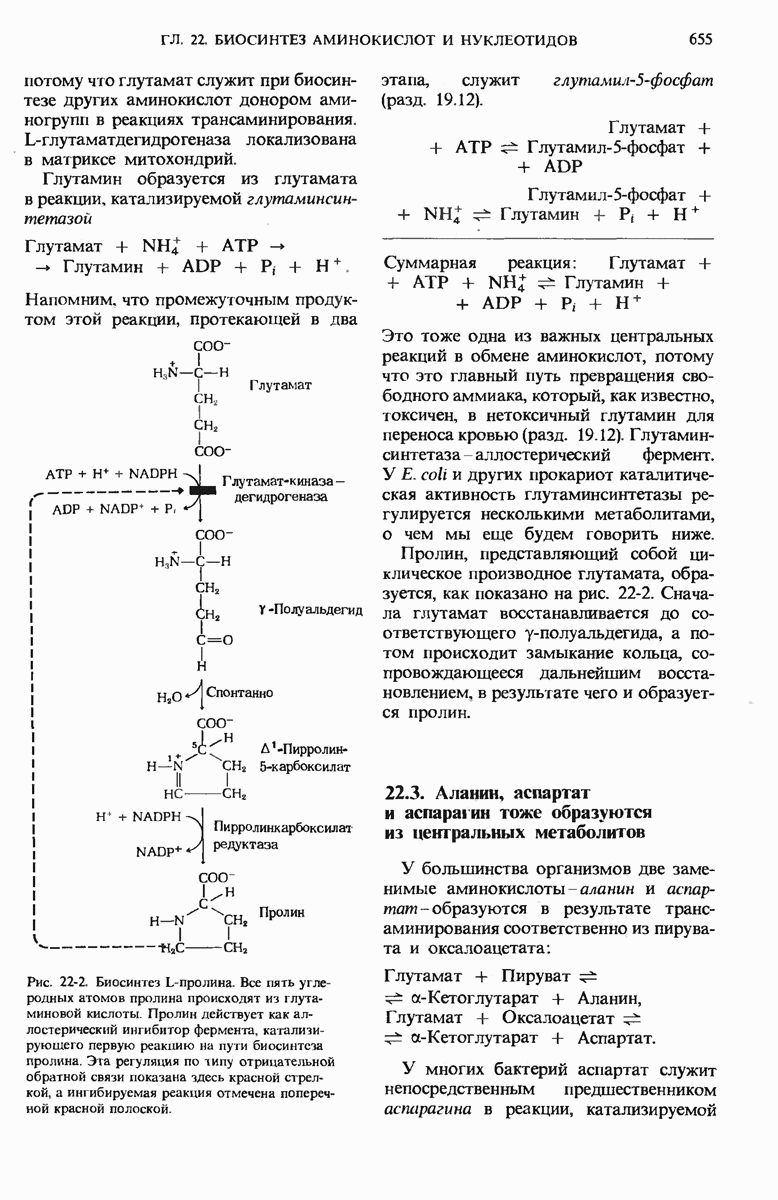
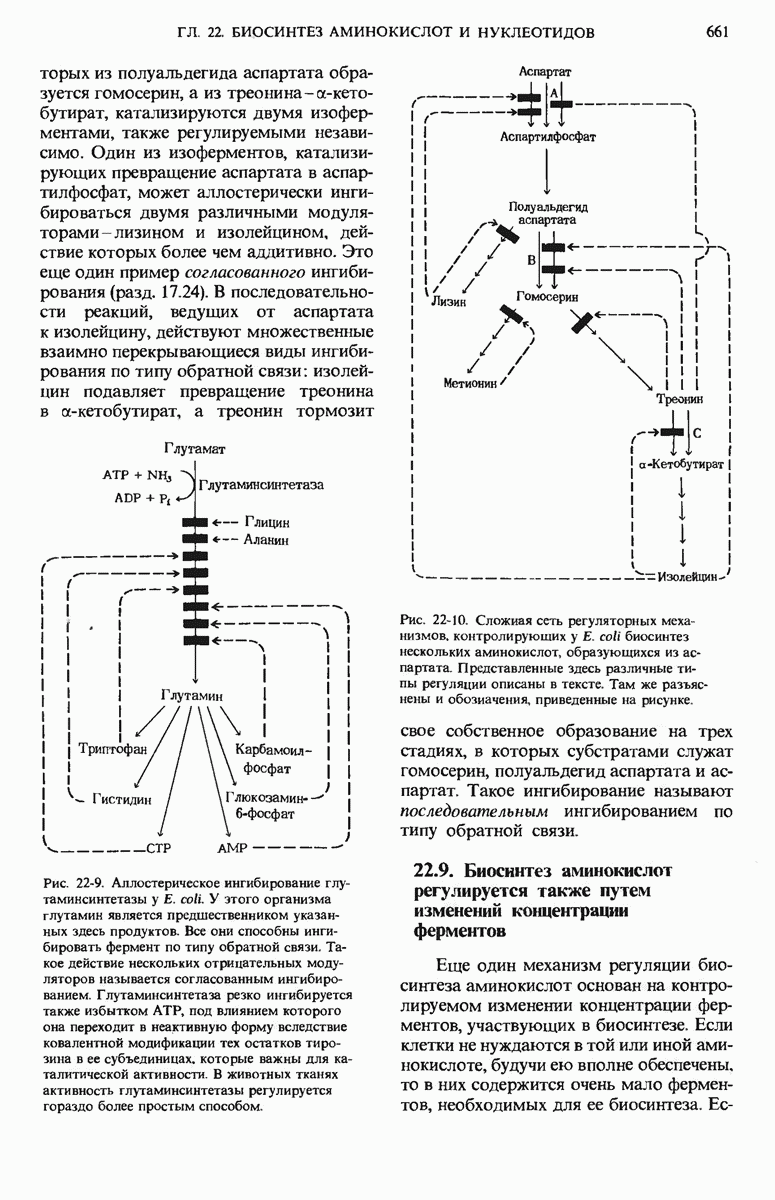
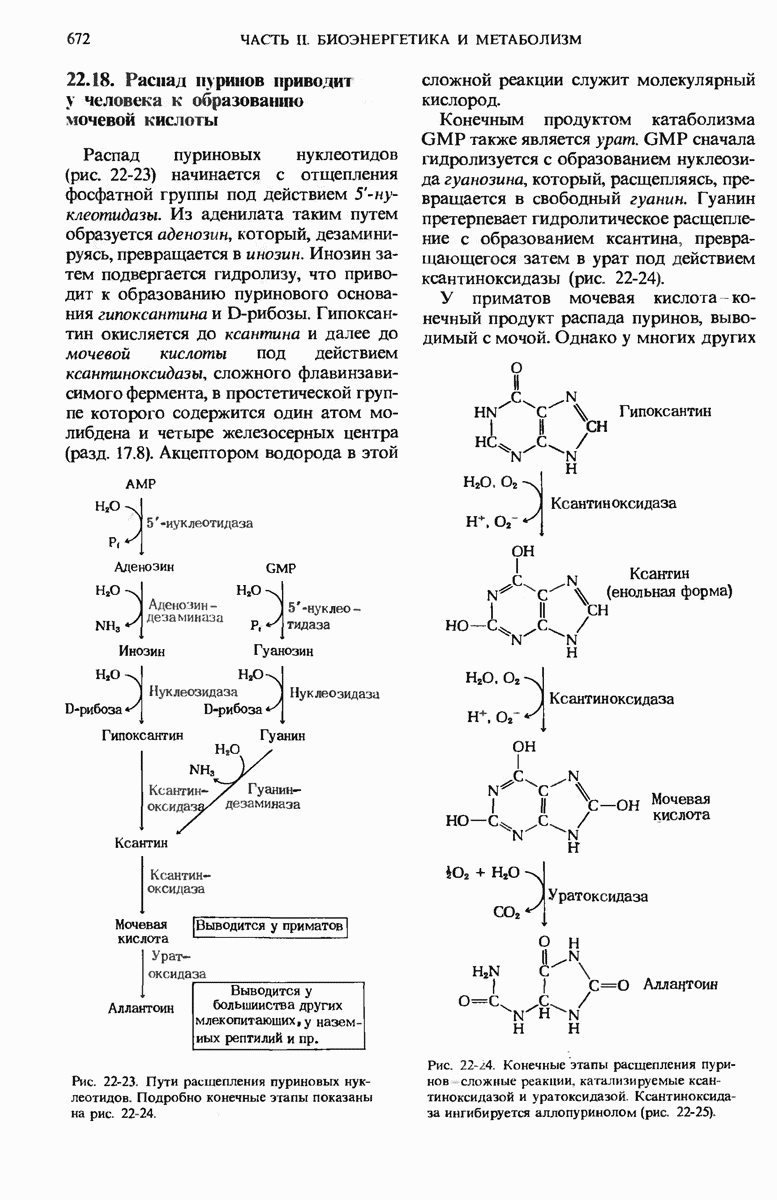
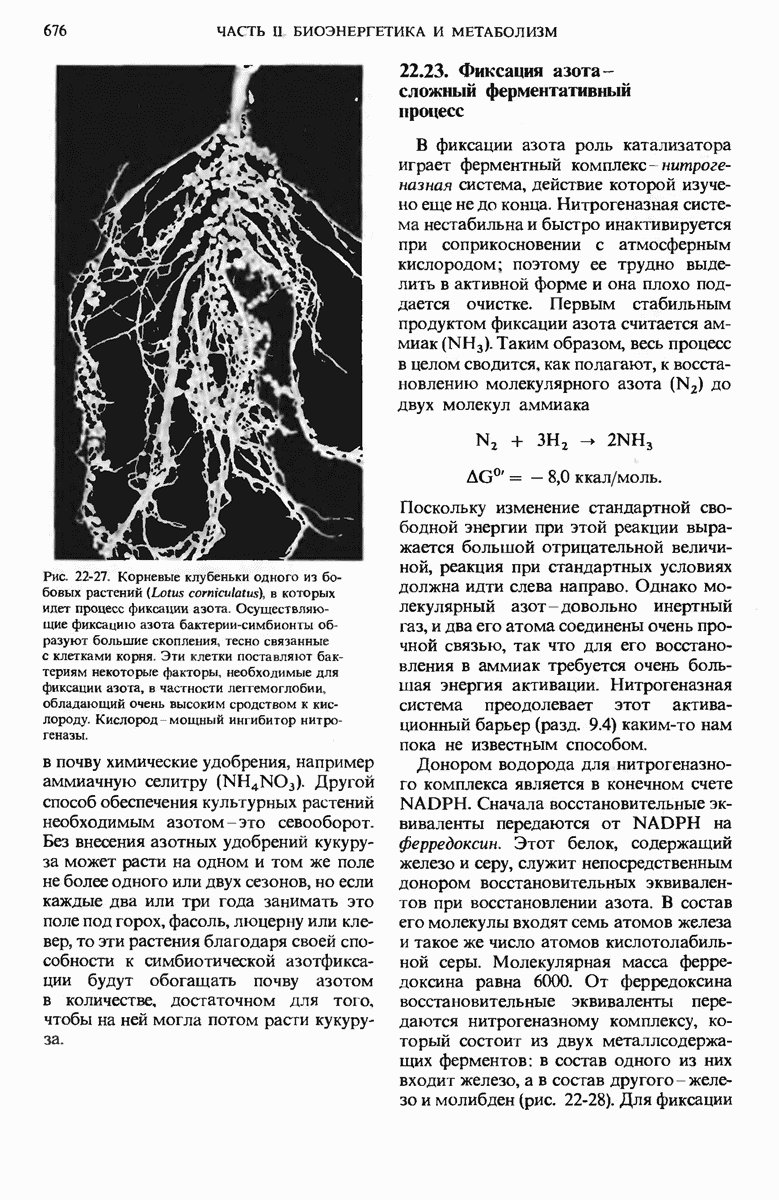
17.19. Внутренняя мембрана митохондрий содержит специфические транспортные системы
Внутренняя мембрана митохондрий непроницаема не только для ионов  но и для многих других ионизованных растворенных веществ. Каким же образом в таком случае попадают в митохондриальный матрикс такие заряженные частицы, как
но и для многих других ионизованных растворенных веществ. Каким же образом в таком случае попадают в митохондриальный матрикс такие заряженные частицы, как  и фосфат
и фосфат  образующиеся в цитозоле при расщеплении АТР, и как новосинтезированный
образующиеся в цитозоле при расщеплении АТР, и как новосинтезированный  (а окислительное фосфорилирование протекает внутри митохондрий) выходит из матрикса наружу?
(а окислительное фосфорилирование протекает внутри митохондрий) выходит из матрикса наружу?
Во внутренней митохондриальной мембране есть две специфические транспортные системы (рис. 17-25), которые делают это возможным.

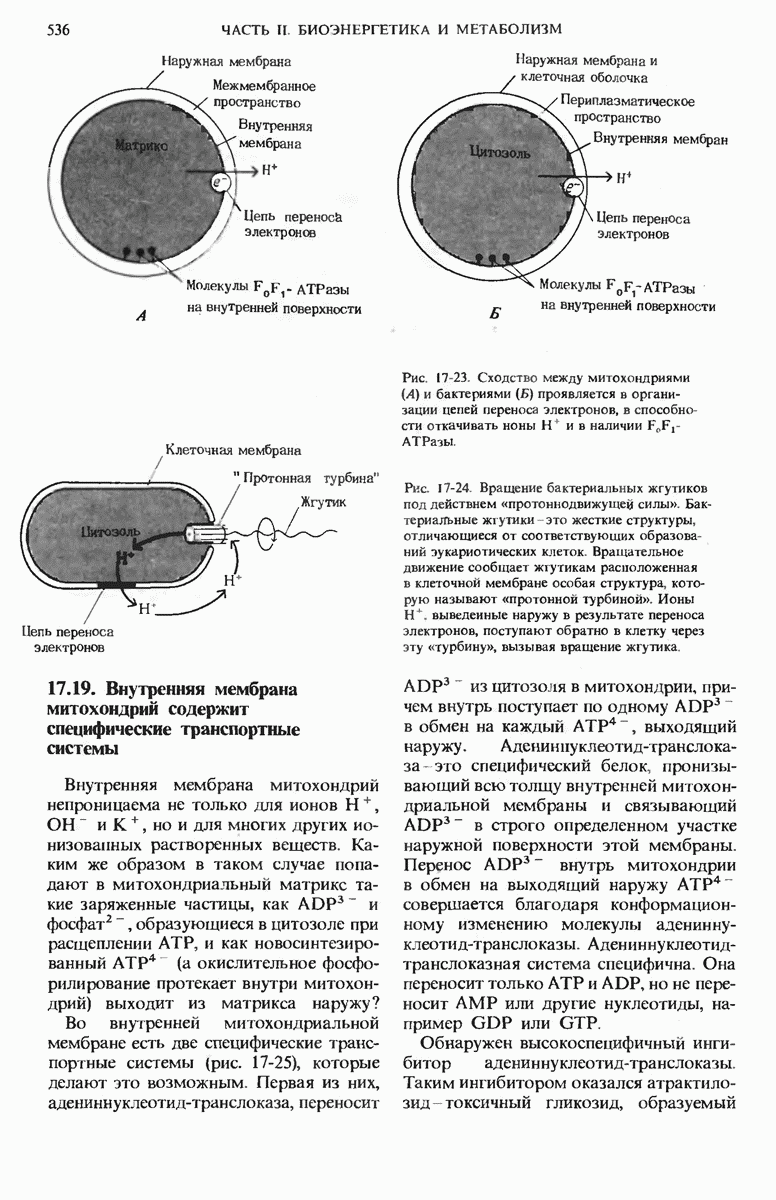
Рис. 17-23. Сходство между митохондриями (А) и бактериями (Б) проявляется в организации цепей переноса электронов, в способности откачивать ионы  и в наличии
и в наличии 

Рис. 17-24. Вращение бактериальных жгутиков под действием «протоннодвижущей силы». Бактериальные жгутики - это жесткие структуры, отличающиеся от соответствующих образований эукариотических клеток. Вращательное движение сообщает жгутикам расположенная в клеточной мембране особая структура, которую называют «протонной турбиной». Ионы  выведенные наружу в результате переноса электронов, поступают обратно в клетку через эту «турбину», вызывая вращение жгутика.
выведенные наружу в результате переноса электронов, поступают обратно в клетку через эту «турбину», вызывая вращение жгутика.
Первая из них, адениннуклеотид-транслоказа, переносит  из цитозоля в митохондрии, причем внутрь поступает по одному
из цитозоля в митохондрии, причем внутрь поступает по одному  - в обмен на каждый
- в обмен на каждый  выходящий наружу. Аденинпу клеотид-транслоказа - это специфический белок, пронизывающий всю толщу внутренней митохондриальной мембраны и связывающий
выходящий наружу. Аденинпу клеотид-транслоказа - это специфический белок, пронизывающий всю толщу внутренней митохондриальной мембраны и связывающий  - в строго определенном участке наружной поверхности этой мембраны. Перенос
- в строго определенном участке наружной поверхности этой мембраны. Перенос  - внутрь митохондрии в обмен на выходящий наружу
- внутрь митохондрии в обмен на выходящий наружу  - совершается благодаря конформационному изменению молекулы адениннуклеотид-транслоказы. Адениннуклеотид-транслоказная система специфична. Она переносит только АТР и ADP, но не переносит АМР или другие нуклеотиды, например GDP или
- совершается благодаря конформационному изменению молекулы адениннуклеотид-транслоказы. Адениннуклеотид-транслоказная система специфична. Она переносит только АТР и ADP, но не переносит АМР или другие нуклеотиды, например GDP или 
Обнаружен высокоспецифичный ингибитор адениннуклеотид-транслоказы. Таким ингибитором оказался атрактилозид - токсичный гликозид, образуемый одним видом чертополоха, произрастающим в некоторых районах Средиземноморья.
Местным жителям с незапамятных времен известно, что скот может отравиться, если поест этого растения в определенное время года. Выделение этого фактора в чистом виде и выяснение его роли, т. е. того, что он действует как ингибитор переноса адениновых нуклеотидов, явилось результатом ряда блестящих исследований, проведенных итальянскими, французскими, немецкими и американскими биохимиками. Ясно, что если в клетках нарушен транспорт адениновых нуклеотидов и ADP не поступает в митохондрии, а АТР не выходит из них, то регенерация цитозольного АТР и ADP оказывается невозможной.
Вторая транспортная система мембран, участвующая в окислительном фосфорилировании, переносит из цитозоля внутрь митохондрий ион  которому сопутствует ион
которому сопутствует ион  (рис. 17-25).
(рис. 17-25).

Рис. 17-25. Транспортные системы внутренней митохондриальной мембраны, переносящие ADP и фосфат из цитозоля в матрикс, а новосинтезированный АТР - из матрикса в цитозоль.
Эта ферментная система, названная фосфат-транслоказой, специфична в отношении фосфата; она также ингибируется некоторыми химическими агентами. В результате совместного действия фосфат-транслоказы и адениннуклеотид-транслоказы фосфат и ADP получают возможность проникнуть в митохондриальный матрикс, а АТР-выйти из митохондрий в цитозоль, туда, где протекает большая часть клеточных процессов, требующих затраты энергии.
В митохондриях печени внутренняя мембрана тоже содержит специфичные транспортные системы. Это системы для переноса пирувата, поступающего в митохондриальный матрикс из цитозоля, в котором он образуется; для дикарбоксилатов, таких, как малат и сукцинат, и, наконец, для трикарбоксилатов - цитрата и изоцитрата. В митохондриях есть также транспортные системы, специфичные в отношении аспартата и глутамата.