Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
§ 3. Математические модели в микробиологииВ этом параграфе мы исследуем несколько простых моделей роста микроорганизмов, не претендуя на полноту изложения вопроса. Модель
(или, по современной терминологии, экономический коэффициент);
и нашел ее аналитическое решение. В безразмерных переменных
зависимость
(рис. 3.6, кривая I).
Рис. 3.6. Рост биомассы:
Рис. 3.7. Фазовый портрет системы (3.16). На фазовой плоскости х, у решение представляется в виде прямых система стремится в конечное состояние Таким образом, уже в 40-х годах была построена математическая модель роста культуры бактерий в замкнутом сосуде. Прекращение роста биомассы в этой модели имеет явную физическую причину — истощение питательной среды, и в этом, с нашей точки зрения, заключается преимущество модели Моно (3.8) перед другими, например, перед моделью Ферхюльста (3.12). Приведем здесь для сравнения получающееся из последней уравнение логистической кривой (рис. 3.6, кривая
Отметим еще раз, что равновесие в модели (3.12) имеет несколько иной смысл, чем в модели Моно (3.16). Можно, например, считать, что ресурсы пищи неограниченны и клетки сначала (при малых х) размножаются с постоянной скоростью, однако, начиная с некоторого момента, прирост биомассы уравновешивается убылью, происходящей из-за взаимодействия клеток. Модель II. Следующим этапом в математической микробиологии можно считать разработку моделей роста микроорганизмов в проточных условиях. Проточное, или непрерывное, культивирование издавна имеет широкое применение в промышленном производстве, так как обеспечивает непрерывный вывод из установки однородной массы клеток. Теория непрерывного культиватора — хемостата — была предложена в 50-х годах Моно [31] и параллельно в США Новиком и Сциллардом [32]. Уравнения баланса биомассы и лимитирующего субстрата в хемостате (с использованием формулы Моно (3.8)) были записаны в виде системы уравнений
где
Здесь введены параметры
Система (3.21) имеет две особые точки:
Точка (3.236) лежит в положительном квадранте в ограниченной области изменения параметра
Граничное значение скорости протока Определим характер устойчивости особых точек. Для точки (3.23а) корни характеристического уравнения: Для состояния равновесия На рис. 3.9 изображен фазовый портрет системы (3.21). В случае (а) все траектории сходятся к стационарной точке 2, в случае Исследование поведения системы (3.20) при наличии малого параметра
Рис. 3.8. Зависимость сационарных концентраций
Рис. 3.9. Фазовый портрет модели II (Моно). А — изоклина вертикальных касательных: Введя безразмерные переменные по формулам
Таким образом, по крайней мере для больших значений х, система разбивается на быстрое уравнение для Итак, в проточном культиваторе при лимитировании роста микроорганизмов субстратом может существовать устойчивое состояние равновесия с ненулевой биомассой. Устойчивость системы обеспечивается как бы «отрицательной обратной связью» через концентрацию лимитирующего субстрата [33]. Важное свойство хемостата — «автостабилизация» лимитирующего фактора — было отмечено в работе Печуркина с соавторами [34]. Из формул (3.17), (3.23) следует, что в стационарном состоянии концентрация субстрата
Если при заданном
Рис. 3.10. Автостабилизация уровня лимитирующего субстрата. Отметим здесь еще одно свойство хемостата, впервые исследованное Новиком и Сциллардом Модель III. Обратимся теперь к модели, учитывающей субстратное угнетение (уменьшение скорости роста описывающая рост биомассы в хемостате при наличии субстратного угнетения, будет
Одна особая точка системы (3.25) по-прежнему имеет координаты
которые сливаются, когда
Таким образом, если Исследуем теперь устойчивость особых точек. Для первой точки
Из формулы (3.27) следует, что точка, соответствующая нижнему участку кривой Фазовый портрет системы (3.25) в случае, когда одновременно существуют все три особые точки, приведен на рис. 3.12. Изоклина горизонтальных касательных В трижды пересекает прямую стационарных значений Модель IV. Рассмотрим теперь практически важный случай, когда скорость роста клетки зависит не только от концентрации субстрата, но также и от выделяемых в процессе жизнедеятельности клетки метаболитов. При ингибирующем действии продуктов метаболизма скорость роста может быть выражена известным законом Моно — Иерусалимского (формула (3.11)). Замкнутая модель, описывающая рост культуры, должна включать в себя уравнение для изменения концентрации продукта
Рис. 3.11. Зависимость стационарных концентраций х и у от 6 (модель III) при
Рис. 3.12. Фазовый портрет модели III. Изоклины вертикальных касательных Запишем его в виде (см. [36])
При этом мы предполагаем, что увеличение концентрации продукта пропорционально приросту биомассы. Заметим, что коэффициент пропорциональности а в (3.28) ограничен сверху ввиду закона сохранения вещества; если все концентрации Введем новую безразмерную переменную учете ингибирующего продукта, будет иметь вид
Стационарные значения концентраций определяются из алгебраической системы:
Одна особая точка снова соответствует нулевой биомассе, другая определяется из соотношения
Рис. 3.13. Влияние константы ингибирования на стационарные концентрации биомассы в модели Таким образом, во всех рассмотренных моделях проточного культиватора Модель V. Учтем в модели Моно вероятность спонтанной гибели клеток; это приведет к появлению дополнительного «оттока» биомассы:
(член — Особые точки системы (3.31):
Скорость вымывания, когда Стационарные кривые Исследование устойчивости показывает, что, как обычно, точка 1 — это узел при
может быть устойчивым фокусом (при противоположном знаке неравенства — устойчивым узлом). Система обладает очень малой добротностью и поведение ее мало отличается от апериодического. Модель VI. До сих пор мы считали, что субстрат расходуется исключительно на прирост биомассы клеток с коэффициентом пропорциональности
откуда для удельной скорости роста имеем
Далее встает вопрос, какой вид имеют зависимости
при больших значениях х и малых у становится бессмысленным. Действительно, при
Обозначив
полностью совпадающую по форме с моделью V, построенной при учете гибели клеток. Нетрудно убедиться, что система (3.34) во всем положительном квадранте вполне разумно описывает динамику культуры в хемостате. Отметим здесь еще раз, что стационарное состояние с отличной от нуля биомассой существует в системе (3.34) при скоростях протока
что, в свою очередь, накладывает ограничение на параметр
или
Это означает, что при Модель VII. В качестве примера автоколебательной микробиологической системы рассмотрим случай, когда в проточном культиваторе обитают два вида, х и у. Субстратом для развития вида у (хищника) является биомасса вида х (жертвы); прирост биомассы хищника представляется функцией Моно. Для развития жертвы выберем модель Ферхюльста с самоограничением (3.1). Тогда можно записать безразмерную систему:
имеющую три стационарные точки:
Вторая точка существует в положительном квадранте при Исследуем устойчивость особых точек. Нулевая точка — устойчивый узел при больших скоростях протока Для особой точки 2 корни характеристического уравнения: жертвы стремится к конечному значению Можно показать, что точка 3 при
Рис. 3.14. Параметрический портрет модели VII при При увеличении коэффициента репродукции хищника
Рис. 3.15. Варианты фазовых портретов модели VII. В — изоклина горизонтальных касательных и тогда вокруг нее должен появиться по крайней мере один устойчивый предельный цикл, соответствующий автоколебаниям в системе хищник — жертва. На рис. 3.14 изображены четыре области в плоскости параметров Мы не затронули модели, описывающие работу других видов проточного культиватора: турбидостата, плотностата, рН-стата, в которых с помощью систем автоматического слежения поддерживаются заданные значения плотности биомассы или других параметров, а также модели полупроточной системы, в которую непрерывно или периодически добавляется свежий питательный субстрат, а жидкость и биомасса не изымаются из культиватора до окончания процесса, систем с газовым питанием и многих других. Эти модели и ссылки на них можно найти в монографиях
|
 |
Оглавление
|













































































































































































































































































































 Исторически математическая микробиология как самостоятельная наука ведет свое начало от фундаментальной работы Моно [22]. Изучая рост культуры микроорганизмов на лимитирующих субстратах, Моно показал, что при постоянстве внешних условий сохраняется постоянным отношение
Исторически математическая микробиология как самостоятельная наука ведет свое начало от фундаментальной работы Моно [22]. Изучая рост культуры микроорганизмов на лимитирующих субстратах, Моно показал, что при постоянстве внешних условий сохраняется постоянным отношение

 соответственно текущая и начальная концентрации биомассы и субстрата. Далее Моно в этой же работе записал замкнутую систему уравнений, описывающих рост культуры, засеянной в некотором сосуде:
соответственно текущая и начальная концентрации биомассы и субстрата. Далее Моно в этой же работе записал замкнутую систему уравнений, описывающих рост культуры, засеянной в некотором сосуде:


 выражается в виде суммы логарифмов:
выражается в виде суммы логарифмов:


 -модель Моно (3.18),
-модель Моно (3.18),  -модель Ферхюльста
-модель Ферхюльста 

 (рис. 3.7). Из начальной точки
(рис. 3.7). Из начальной точки  при
при 
 вся прямая
вся прямая  является прямой стационарных состояний.
является прямой стационарных состояний.



 скорость протока, или разбавления, имеющая размерность
скорость протока, или разбавления, имеющая размерность  концентрация субстрата, поступающего в культиватор,
концентрация субстрата, поступающего в культиватор,  экономический коэффициент. В безразмерных переменных (3.17) имеем
экономический коэффициент. В безразмерных переменных (3.17) имеем




 называется скоростью вымывания. Действительно, при
называется скоростью вымывания. Действительно, при  прирост биомассы не может уже компенсировать ее отток и культура полностью «вымывается» из культиватора. Стационарные концентрации х и у (3.23) в зависимости от
прирост биомассы не может уже компенсировать ее отток и культура полностью «вымывается» из культиватора. Стационарные концентрации х и у (3.23) в зависимости от  при
при  представлены на рис. 3.8 сплошными линиями.
представлены на рис. 3.8 сплошными линиями.
 Следовательно, в условиях вымывания, когда
Следовательно, в условиях вымывания, когда  оба корня отрицательны и особая точка с нулевой концентрацией биомассы — устойчивый узел. При
оба корня отрицательны и особая точка с нулевой концентрацией биомассы — устойчивый узел. При  эта особая точка становится седлом.
эта особая точка становится седлом.
 В силу условия (3.24) оба корня всегда отрицательны и особая точка (3.236) всегда — устойчивый узел.
В силу условия (3.24) оба корня всегда отрицательны и особая точка (3.236) всегда — устойчивый узел.
 к точке 1 (происходит вымывание биомассы).
к точке 1 (происходит вымывание биомассы).
 проведено в работе [33].
проведено в работе [33].

 от 6 при
от 6 при  Сплошные линии — модель II, пунктир—модель
Сплошные линии — модель II, пунктир—модель 

 В — горизонтальных:
В — горизонтальных: 
 и обозначив
и обозначив  можно привести систему (3.20) к виду
можно привести систему (3.20) к виду

 и медленное — для х.
и медленное — для х.
 в культиваторе не зависит от о о — входной концентрации; от
в культиваторе не зависит от о о — входной концентрации; от  зависит лишь стационарная концентрация биомассы X:
зависит лишь стационарная концентрация биомассы X:

 увеличивать
увеличивать  то в области
то в области  (рис. 3.10) осуществляется режим вымывания
(рис. 3.10) осуществляется режим вымывания  при
при  биомасса X растет линеино вместе с
биомасса X растет линеино вместе с  а концентрация субстрата будет стабилизирована на уровне
а концентрация субстрата будет стабилизирована на уровне  Существенно, что это справедливо только до тех пор, пока данный субстрат действительно лимитирует рост; когда же в игру вступает другой лимитирующий фактор (при
Существенно, что это справедливо только до тех пор, пока данный субстрат действительно лимитирует рост; когда же в игру вступает другой лимитирующий фактор (при  на рис. 3.10), автостабилизация уровня
на рис. 3.10), автостабилизация уровня  прекращается и он начинает расти линейно с
прекращается и он начинает расти линейно с 

 автоселекцию мутантов: если в результате мутации образуется новый штамм, лучше утилизирующий данный субстрат, то с течением времени он вытесняет исходный штамм из хемостата. Иначе говоря, два конкурирующих за один субстрат штамма не могут устойчиво сосуществовать в проточном культиваторе (см., например,
автоселекцию мутантов: если в результате мутации образуется новый штамм, лучше утилизирующий данный субстрат, то с течением времени он вытесняет исходный штамм из хемостата. Иначе говоря, два конкурирующих за один субстрат штамма не могут устойчиво сосуществовать в проточном культиваторе (см., например,  Это, собственно, частный случай закона перекрывающихся ниш Гаузе, о котором шла речь в § 1.
Это, собственно, частный случай закона перекрывающихся ниш Гаузе, о котором шла речь в § 1.
 при больших концентрациях субстрата). В этом случае безразмерная скорость роста в зависимости от безразмерной концентрации субстрата дается выражением (3.9). Функция
при больших концентрациях субстрата). В этом случае безразмерная скорость роста в зависимости от безразмерной концентрации субстрата дается выражением (3.9). Функция  достигает максимального значения
достигает максимального значения  при
при  (см. рис. 3.4). Система уравнений,
(см. рис. 3.4). Система уравнений,

 но кроме нее могут существовать еще две точки:
но кроме нее могут существовать еще две точки:

 иначе говоря, когда скорость протока равна максимальному значению скорости роста. При
иначе говоря, когда скорость протока равна максимальному значению скорости роста. При  могут быть два действительных положительных значения
могут быть два действительных положительных значения  з, но они оба не всегда имеют смысл, так как для большего из них может нарушиться условие
з, но они оба не всегда имеют смысл, так как для большего из них может нарушиться условие  (условие положительности
(условие положительности  Это условие накладывает ограничение на параметр
Это условие накладывает ограничение на параметр  аналогичное условию вымывания в предыдущих моделях:
аналогичное условию вымывания в предыдущих моделях:

 То существует лишь одно положительное стационарное значение у и поведение системы практически не отличается от поведения модели Моно (в этом случае мы не заходим за максимум кривой
То существует лишь одно положительное стационарное значение у и поведение системы практически не отличается от поведения модели Моно (в этом случае мы не заходим за максимум кривой  и влияние субстратного подавления не сказывается). При увеличении
и влияние субстратного подавления не сказывается). При увеличении  в интервале скоростей разбавления
в интервале скоростей разбавления  будут существовать оба корня
будут существовать оба корня  из (3.26).
из (3.26).
 корни характеристического уравнения
корни характеристического уравнения  т. е. эта точка устойчива при
т. е. эта точка устойчива при  Для двух других особых точек имеем
Для двух других особых точек имеем

 устойчивым узлом, а верхнему
устойчивым узлом, а верхнему  -седлом. При
-седлом. При  будет наблюдаться катастрофа типа складки — появление двух точек (устойчивой и неустойчивой). В интервале значений параметров
будет наблюдаться катастрофа типа складки — появление двух точек (устойчивой и неустойчивой). В интервале значений параметров  существуют одновременно две устойчивые особые точки и, следовательно, при плавном изменении параметра
существуют одновременно две устойчивые особые точки и, следовательно, при плавном изменении параметра  будет наблюдаться гистерезис. Например, при увеличении
будет наблюдаться гистерезис. Например, при увеличении  режим вымывания наступает не при
режим вымывания наступает не при  а позднее при
а позднее при  Концентрации х и у на границах области гистерезиса меняются скачком (рис. 3.11).
Концентрации х и у на границах области гистерезиса меняются скачком (рис. 3.11).
 На фазовой плоскости существует сепаратриса, разделяющая зоны притяжения устойчивых точек 1 и 3.
На фазовой плоскости существует сепаратриса, разделяющая зоны притяжения устойчивых точек 1 и 3.


 Сплошные линии —
Сплошные линии —  пунктир —
пунктир — 

 горизонтальных — кривая В
горизонтальных — кривая В  -Пунктир-сепаратриса седловой точки 2.
-Пунктир-сепаратриса седловой точки 2.

 выражены в одних и тех же единицах (например, молях углерода), то
выражены в одних и тех же единицах (например, молях углерода), то 
 и коэффициент
и коэффициент  Тогда система безразмерных уравнений, описывающая рост биомассы в проточном культиваторе при
Тогда система безразмерных уравнений, описывающая рост биомассы в проточном культиваторе при


 Скорость вымывания в модели та же, что и в модели Моно:
Скорость вымывания в модели та же, что и в модели Моно:  величина параметра ингибирование сказывается лишь на форме зависимости
величина параметра ингибирование сказывается лишь на форме зависимости  при
при  На рис. 3.13 приведено семейство кривых (8) при разных значениях х. Видно, что ингибирующее влияние продукта ведет к значительному уменьшению стационарных концентраций биомассы при всех скоростях протока. Исследование устойчивости показывает, что нулевая (по биомассе) особая точка, как обычно, устойчива при
На рис. 3.13 приведено семейство кривых (8) при разных значениях х. Видно, что ингибирующее влияние продукта ведет к значительному уменьшению стационарных концентраций биомассы при всех скоростях протока. Исследование устойчивости показывает, что нулевая (по биомассе) особая точка, как обычно, устойчива при  и седло — при
и седло — при  Другая точка (когда она существует) — устойчивый узел.
Другая точка (когда она существует) — устойчивый узел.


 состояние равновесия с ненулевой биомассой представляет собой устойчивый узел, т. е. процессы установления в системе носят апериодический характер. В следующей простой модели (см. [37, 38]) может существовать устойчивый фокус, т. е. в принципе могут быть затухающие колебания концентраций вблизи стационарного значения.
состояние равновесия с ненулевой биомассой представляет собой устойчивый узел, т. е. процессы установления в системе носят апериодический характер. В следующей простой модели (см. [37, 38]) может существовать устойчивый фокус, т. е. в принципе могут быть затухающие колебания концентраций вблизи стационарного значения.

 как раз и описывает гибель клеток).
как раз и описывает гибель клеток).

 обращается в нуль, в данной модели меньше, чем в модели Моно:
обращается в нуль, в данной модели меньше, чем в модели Моно: 
 при
при  изображены пунктиром на рис. 3.8. В отличие от модели Моно (представленной на этом же рисунке сплошными линиями) при уменьшении скорости протока концентрация биомассы снижается, стремясь к нулю при
изображены пунктиром на рис. 3.8. В отличие от модели Моно (представленной на этом же рисунке сплошными линиями) при уменьшении скорости протока концентрация биомассы снижается, стремясь к нулю при  Это объясняется тем, что при малых скоростях протока все. большую роль начинает играть спонтанная гибель клеток.
Это объясняется тем, что при малых скоростях протока все. большую роль начинает играть спонтанная гибель клеток.
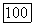 и седло при
и седло при 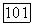 точка же 2 при некотором соотношении параметров, определяемом из неравенства
точка же 2 при некотором соотношении параметров, определяемом из неравенства

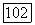 Однако хорошо известно, что многие, так называемые «энергетические», субстраты расходуются также на поддержание жизненных функций клеток [П 38, 39]. В этом случае суммарный расход субстрата будет состоять из двух членов:
Однако хорошо известно, что многие, так называемые «энергетические», субстраты расходуются также на поддержание жизненных функций клеток [П 38, 39]. В этом случае суммарный расход субстрата будет состоять из двух членов:


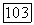 В ранних работах Перта [39] предполагалось, что и в этом случае
В ранних работах Перта [39] предполагалось, что и в этом случае 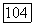 можно выразить формулой Моно:
можно выразить формулой Моно: 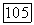 Тогда безразмерное уравнение для субстрата
Тогда безразмерное уравнение для субстрата

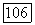 имеем
имеем 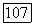 т. е. фазовые траектории уходят в область отрицательных у. В последних работах Перта (см. [П 38]), было сделано более естественное предположение о том, что при
т. е. фазовые траектории уходят в область отрицательных у. В последних работах Перта (см. [П 38]), было сделано более естественное предположение о том, что при 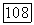 стремится к нулю полное потребление субстрата единицей биомассы, т. е.
стремится к нулю полное потребление субстрата единицей биомассы, т. е. 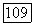 Тогда, используя гиперболический закон Моно для выражения зависимости
Тогда, используя гиперболический закон Моно для выражения зависимости 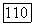 имеем
имеем

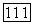 получим для культивирования в хемостате на этот раз следующую безразмерную систему:
получим для культивирования в хемостате на этот раз следующую безразмерную систему:

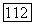

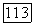


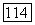 поглощаемого биомассой субстрата нехватает даже на удовлетворение энергетических нужд клеток и они должны погибнуть. Для клеток остается, правда, возможность изменить свой метаболизм и использовать другие источники энергии, однако это обстоятельство в рассматриваемой модели Перта не учитывается.
поглощаемого биомассой субстрата нехватает даже на удовлетворение энергетических нужд клеток и они должны погибнуть. Для клеток остается, правда, возможность изменить свой метаболизм и использовать другие источники энергии, однако это обстоятельство в рассматриваемой модели Перта не учитывается.


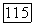 третья — при
третья — при 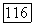
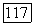 что соответствует случаю полного вымывания обеих популяций. При скоростях протока, меньших коэффициента репродукции жертвы
что соответствует случаю полного вымывания обеих популяций. При скоростях протока, меньших коэффициента репродукции жертвы 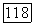 нулевая точка становится неустойчивой (седло).
нулевая точка становится неустойчивой (седло).
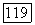 т. е. условием ее устойчивости будет условие малости коэффициента репродукции хищника
т. е. условием ее устойчивости будет условие малости коэффициента репродукции хищника 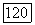 в этом случае популяция хищника вымирает, а величина популяции
в этом случае популяция хищника вымирает, а величина популяции
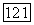 При
При 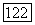 точка 2 тоже становится седлом.
точка 2 тоже становится седлом.
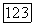 устойчива, т. е. в этой области изменения параметров возможно устойчивое сосуществование хищника и жертвы в проточном культиваторе.
устойчива, т. е. в этой области изменения параметров возможно устойчивое сосуществование хищника и жертвы в проточном культиваторе.

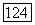 Области определяются условиями: о)
Области определяются условиями: о) 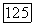
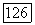 и
и 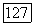 условии не слишком сильного самоограничения жертвы
условии не слишком сильного самоограничения жертвы 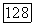 точка 3 становится неустойчивым узлом или фокусом
точка 3 становится неустойчивым узлом или фокусом

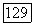 , А — вертикальных
, А — вертикальных 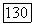 Жирной линией проведен предельный цикл. Случаи а) — г) соответствуют областям параметров на рис. 3.14.
Жирной линией проведен предельный цикл. Случаи а) — г) соответствуют областям параметров на рис. 3.14.
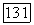 в которых решения системы (3.35) имеют существенно различный характер. Соответствующие фазовые портреты приведены на рис. 3.15.
в которых решения системы (3.35) имеют существенно различный характер. Соответствующие фазовые портреты приведены на рис. 3.15.
 Отметим также интересную работу [40], где наряду с динамикой биомассы и субстрата рассматривается изменение концентрации промежуточного метаболита.
Отметим также интересную работу [40], где наряду с динамикой биомассы и субстрата рассматривается изменение концентрации промежуточного метаболита.