Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
§ 3. Функции распределения клеток по размерамНаряду с распределением микроорганизмов по возрастам исследователи большое внимание уделяют функциям распределения по размерам. Фактически индивидуальный возраст клетки не поддается непосредственному измерению (если не иметь дела с синхронной культурой), тогда как размеры клеток определяются достаточно точно. Существует несколько различных методик определения размеров клеток, простейшей из которых является измерение линейных размеров по микрофотографиям, снятым со специально" приготовленных мазков [15]. Используются также более совершенные методы, дающие возможность построения практически непрерывных кривых распределения. Например, метод Коултера основан на измерении проводимости раствора в сосуде, разделенном на две части перегородкой с микроотверстием; при прохождении единичной живой клетки через отверстие проводимость раствора изменяется пропорционально объему этой клетки (подробнее см. монографию [16], а также [17, 18]). В настоящее время созданы автоматизированные устройства для комплексного статистического анализа клеток, соединенные с ЭВМ. Таким образом, получение экспериментальных кривых распределения клеток по размерам не встречает в настоящее время существенных трудностей. Более сложным является установление связи формы кривых распределения с физиологическим состоянием клеток и условиями их выращивания. Этому вопросу посвящены теоретические работы, содержащие модели роста клеточных популяций, распределенных по размерам. Наиболее последовательная теория ведет свое начало от работы Икмана и др. [19] и целой серии статей Белла и Андерсена [20], где из закона сохранения числа клеток выводится уравнение неразрывности для плотности распределения клеток по массам или по возрастам и объемам. На основании перечисленных выше работ запишем уравнение неразрывности для функции распределения клеток по объемам (см. [11]). Пусть
Полная производная по времени от
Первый член в левой части уравнения (4.33) представляет изменение функции
Убыль клеток в единицу времени из данного размера складывается из выноса их со скоростью протока
Если при делении каждой родительской клетки получается
Для бактерий в большинстве случаев можно считать, что родительская клетка делится на две дочерние
Для почкующихся клеток можно предположить, что почка отделяется от материнской клетки тогда, когда отношение ее размера к суммарному достигает некоторого предельного значения а. В этом случае функция
т. е. клетки после деления прибывают в разные размеры: дочерняя — в размер Введем далее нормированную функцию распределения
Учитывая (4.32), имеем условие нормировки для функции
Кроме полного числа клеток
Вместо уравнения (4.33) запишем систему двух уравнений, описывающих динамику числа клеток и изменение нормированной функции распределения клеток по размерам. Будем считать, что клетки делятся на две дочерние, т. е.
Запишем также соответствующее уравнение для концентрации биомассы:
Условие нормировки (4.37) заменяет граничное условие для функции распределения
Задача определения законов распределения клеток по размерам будет полностью определена, если задать начальное число клеток в культиваторе Будем решать стационарную задачу, т. е. предположим, что параметры системы не зависят от времени и культура в хемостате находится в равновесии: обыкновенное дифференциальное уравнение:
Условие сохранения числа клеток в культиваторе при
где слева стоит прибыль клеток в единицу времени за счет деления, а справа — убыль за счет вымывания. Условие сохранения массы клеток
Отметим, что для определения решения Для решения задачи необходимо задать 1. Клетки растут экспоненциально:
2. Объем клеток в промежутках между делениями увеличивается с постоянной скоростью:
Далее, нам надо выбрать модель клеточного деления для определения функции
Подчеркнем здесь еще раз, что под размера Связь между
Таким образом,
Рассмотрим сначала идеальный случай: все клетки делятся пополам, достигнув определенного размера
а скорость деления, как следует из (4.48), равна нулю при
Рис. 4.8. Кривые распределения числа клеток по объемам в случае деления при фиксированном размере В случае линейного роста клетки (4.47) аналогичные вычисления дают
Таким образом, нормированное распределение Перейдем теперь к более реалистической ситуации, когда размер деления является случайной функцией. При этом определение стационарного нормированого распределения «упреждающей» точке: Приближенное решение задачи в предположении гауссова распределения размеров деления и размеров дочерних клеток проведено в работе [21]. Используя несколько вариантов законов роста индивидуальной клетки (экспоненциальный, линейный, инверсный, кусочно-линейный), авторы рассчитывали теоретические кривые распределения клеток по массам Отметим здесь также направление моделирования, где элемент случайности вводится не в закон деления клетки, а в закон роста. В работе [22] предполагается, что размер I клетки возраста Мы в работах [23—25] решали аналогичную задачу в более простом варианте — деления бактерий-палочек точно пополам.
Рис. 4.9. Плотность распределения вероятностей деления клетки: а) условной, б) безусловной.
Рис. 4.10. Удельная скорость деления клетки. Сплошная линия — экспоненциальный закон роста клетки, пунктир — линейный. Предполагая, что все клетки имеют одинаковый диаметр и растут только в длину, в силу линейности уравнений мы можем вместо распределения по объемам Выберем в качестве закона распределения размеров деления ступенчатую функцию
т. е. предположим, что клетка не может делиться, пока не достигнет размера
Удельная скорость деления
Для линейно растущих клеток будет, соответственно,
Запишем теперь уравнение, которому удовлетворяет нормированная функция распределения клеток по длинам
Для случая экспоненциального роста, подставив
Для случая линейного роста клетки уравнение для функции распределения
где А — средний размер клетки:
Точно решить уравнения (4.57), (4.58) не представляется возможным, так как в правую часть уравнения для
Рис. 4.11. Распределение клеток по длинам при экспоненциальном законе роста. По осям отложены безразмерные величины Используя его в качестве функции источника, можно машинными методами найти решение на предыдущем участке Сравнение теоретических зависимостей хорошего совпадения теории с экспериментом как в предположении экспоненциального роста, так и линейного. Пример такого подбора приведен на рис. 4.13.
Рис. 4.12. Распределение клеток по длинам при линейном законе роста. Сплошные линии — Однако оптимальные значения параметров А соответствующие разным теориям, при этом оказались несколько различными, поэтому ответ на вопрос: по какому закону растут клетки? — может быть дан лишь в том случае, когда функция распределения «размеров деления»
Рис. 4.13. Сравнение экспериментального распределения тифозных бактерий-палочек по длинам (ломаная линия) [12] с теоретическим, вычисленным по линейной теории (плавная кривая): Итак, мы изложили в этом параграфе модели, в которых исследуется распределение микроорганизмов по размерам. Экспериментальные функции распределения нашли широкое применение для управления микробиологическим производством, в частности, для контроля стабильности культуры [26]. Сочетание модели с экспериментом позволит изучить более полно законы роста индивидуальных клеток, найти связь физиологических характеристик клеток с условиями роста культуры в целом. Внедрение машинных методов в обработку экспериментальных данных, о чем мы упоминали в начале параграфа, дает основание надеяться на успехи в этом направлении.
|
 |
Оглавление
|













































































































































































































































































































 число клеток, имеющих в момент времени
число клеток, имеющих в момент времени  объем в интервале
объем в интервале  Общее число клеток в проточном культиваторе обозначим через
Общее число клеток в проточном культиваторе обозначим через  Тогда
Тогда

 должна быть равна сумме «источников» и «стоков»:
должна быть равна сумме «источников» и «стоков»:

 со временем, второй — изменение, связанное с ростом клеток. Обозначим через
со временем, второй — изменение, связанное с ростом клеток. Обозначим через  скорость прироста объема индивидуальной клетки
скорость прироста объема индивидуальной клетки 

 гибели со скоростью
гибели со скоростью  и деления на дочерние клетки с удельной скоростью
и деления на дочерние клетки с удельной скоростью  Будем считать, что гибелью клеток можно пренебречь, так как
Будем считать, что гибелью клеток можно пренебречь, так как  Последний член в (4.33), обозначенный
Последний член в (4.33), обозначенный  функция источника, описывающая прибыль клеток за единицу времени в данный размер за счет деления родительских:
функция источника, описывающая прибыль клеток за единицу времени в данный размер за счет деления родительских:

 объем родительской клетки; произведение и
объем родительской клетки; произведение и  есть число клеток в интервале
есть число клеток в интервале  делящихся за единицу времени; функция
делящихся за единицу времени; функция  описывает долю дочерних клеток размера
описывает долю дочерних клеток размера  полученных из родительской
полученных из родительской  Суммарная прибыль клеток в размер
Суммарная прибыль клеток в размер  за счет деления родительских выражается интегралом по всем клеткам, для которых
за счет деления родительских выражается интегралом по всем клеткам, для которых 
 -дочерних, мы должны записать:
-дочерних, мы должны записать:

 и при этом точно пополам, тогда
и при этом точно пополам, тогда  выражается через
выражается через  -функцию:
-функцию:

 будет иметь вид
будет иметь вид

 а материнская — в размер
а материнская — в размер 
 поделив
поделив  на полное число клеток:
на полное число клеток:



 нас будет также интересовать другая макроскопическая характеристика культуры —
нас будет также интересовать другая макроскопическая характеристика культуры —  концентрация биомассы в культиваторе объема
концентрация биомассы в культиваторе объема  Введем у — плотность вещества клетки, тогда масса клетки объема
Введем у — плотность вещества клетки, тогда масса клетки объема  будет
будет  и
и

 Тогда
Тогда


 Функцию источника для бактерий, делящихся точно пополам, вычислим на основании выражений (4.35) и (4.36):
Функцию источника для бактерий, делящихся точно пополам, вычислим на основании выражений (4.35) и (4.36):

 начальное распределение по размерам
начальное распределение по размерам  и считать известными функции и
и считать известными функции и  и
и 
 Для стационарного распределения
Для стационарного распределения  получим из (4.40) (положив
получим из (4.40) (положив  )
)

 дает соотношение
дает соотношение

 приводит к известному соотношению между скоростью протока
приводит к известному соотношению между скоростью протока  и удельной скоростью роста
и удельной скоростью роста  в стационарном состоянии:
в стационарном состоянии:

 достаточно использовать любые два условия из трех - (4.37), (4.44), (4.45), третье условие должно приводить к тождеству.
достаточно использовать любые два условия из трех - (4.37), (4.44), (4.45), третье условие должно приводить к тождеству.
 скорость роста индивидуальной клетки и
скорость роста индивидуальной клетки и  скорость деления. В литературе имеются противоречивые данные о законах роста клеток; по-видимому, разнообразие экспериментальных данных свидетельствует о том, что различные виды микроорганизмов в разных условиях могут расти по-разному. Поэтому рассмотрим два частных случая.
скорость деления. В литературе имеются противоречивые данные о законах роста клеток; по-видимому, разнообразие экспериментальных данных свидетельствует о том, что различные виды микроорганизмов в разных условиях могут расти по-разному. Поэтому рассмотрим два частных случая.


 удельной скорости деления. По аналогии с возрастными моделями (см. формулы (4.17)-(4.20) § 3) введем функцию распределения размеров делящихся клеток
удельной скорости деления. По аналогии с возрастными моделями (см. формулы (4.17)-(4.20) § 3) введем функцию распределения размеров делящихся клеток  которая имеет смысл плотности безусловной вероятности деления клетки, достигшей размера
которая имеет смысл плотности безусловной вероятности деления клетки, достигшей размера  Связь функции
Связь функции  с плотностью вероятности деления клетки
с плотностью вероятности деления клетки  при условии, что клетка достигла размера
при условии, что клетка достигла размера  не разделившись, будет
не разделившись, будет

 мы имеем в виду априорную вероятность для клетки разделиться по достижении
мы имеем в виду априорную вероятность для клетки разделиться по достижении
 эта вероятность не должна зависеть от других причин убыли клеток — вымывания, гибели и т. п.
эта вероятность не должна зависеть от других причин убыли клеток — вымывания, гибели и т. п.
 и скоростью деления и
и скоростью деления и  в случае однозначной зависимости
в случае однозначной зависимости  т. е. при заданном законе роста клетки можно получить из соотношения
т. е. при заданном законе роста клетки можно получить из соотношения


 Распределение размеров деления в этом случае представляется в виде
Распределение размеров деления в этом случае представляется в виде

 и становится бесконечно большой при
и становится бесконечно большой при  Решение уравнения (4.43) в предположении экспоненциального роста индивидуальной клетки
Решение уравнения (4.43) в предположении экспоненциального роста индивидуальной клетки


 . По осям отложены безразмерные величины
. По осям отложены безразмерные величины 

 в обоих случаях определяется лишь максимальным объемом клетки
в обоих случаях определяется лишь максимальным объемом клетки  и не зависит от параметра
и не зависит от параметра  Характер функции
Характер функции  несколько меняется в зависимости от выбора закона роста отдельной клетки. На рис. 4.8 видно, что при экспоненциальном законе роста (кривая 1) функция распределения имеет более крутой спад, чем при линейном (кривая 2).
несколько меняется в зависимости от выбора закона роста отдельной клетки. На рис. 4.8 видно, что при экспоненциальном законе роста (кривая 1) функция распределения имеет более крутой спад, чем при линейном (кривая 2).
 усложняется, так как даже в случае деления клеток точно пополам в правой части уравнения (4.43) для
усложняется, так как даже в случае деления клеток точно пополам в правой части уравнения (4.43) для  стоит функция источника в
стоит функция источника в
 а в случае, когда дочерние клетки неравнозначны, функция источника будет выражаться интегралом (4.35).
а в случае, когда дочерние клетки неравнозначны, функция источника будет выражаться интегралом (4.35).
 для шести значений скорости протока
для шести значений скорости протока  Параметры распределений и скоростей
Параметры распределений и скоростей  определялись (по методу наименьших квадратов) при сравнении с экспериментальными кривыми распределения.
определялись (по методу наименьших квадратов) при сравнении с экспериментальными кривыми распределения.
 есть случайная величина, распределенная по нормальному закону с коэффициентом вариации, не зависящим от возраста. Тогда, предполагая линейный рост клетки, а также детерминированный размер материнской и дочерней клеток в момент почкования, авторы получают в интегральной форме закон распределенная клеток по размерам, в котором три параметра остаются неизвестными и подбираются при сравнении с экспериментом.
есть случайная величина, распределенная по нормальному закону с коэффициентом вариации, не зависящим от возраста. Тогда, предполагая линейный рост клетки, а также детерминированный размер материнской и дочерней клеток в момент почкования, авторы получают в интегральной форме закон распределенная клеток по размерам, в котором три параметра остаются неизвестными и подбираются при сравнении с экспериментом.


 записать
записать  аналогичное распределение по длинам
аналогичное распределение по длинам  Учтем только, что в (4.38) у — это масса, приходящаяся на единицу длины клетки.
Учтем только, что в (4.38) у — это масса, приходящаяся на единицу длины клетки.

 для больших же размеров
для больших же размеров  условная вероятность деления (при условии, что клетка не разделилась раньше) не зависит от длины (рис. 4.9, а). Тогда безусловная вероятность (или функция распределения размеров деления) будет согласно (4.48) (рис. 4.9, б)
условная вероятность деления (при условии, что клетка не разделилась раньше) не зависит от длины (рис. 4.9, а). Тогда безусловная вероятность (или функция распределения размеров деления) будет согласно (4.48) (рис. 4.9, б)

 (рис. 4.10) будет связана с функцией
(рис. 4.10) будет связана с функцией  через скорость роста клеток
через скорость роста клеток  В случае клеток, экспоненциально растущих в длину, имеем аналогично (4.49)
В случае клеток, экспоненциально растущих в длину, имеем аналогично (4.49)


 в предположении деления клеток точно пополам. Из (4.43) получим при замене
в предположении деления клеток точно пополам. Из (4.43) получим при замене  на I:
на I:

 и учитывая соотношение
и учитывая соотношение  получим
получим

 будет
будет

 а скорость роста
а скорость роста  связана со скоростью протока и параметрами функции деления соотношениями
связана со скоростью протока и параметрами функции деления соотношениями

 входит неизвестная функция в «упреждающей» точке
входит неизвестная функция в «упреждающей» точке  Для приближенного расчета можно ввести максимальную возможную длину клеток
Для приближенного расчета можно ввести максимальную возможную длину клеток  так что
так что  при
при  (удобно выбрать
(удобно выбрать 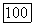 На участке
На участке 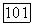 в уравнениях (4.57), (4.58) функция источника отсутствует и можно найти соответствующее аналитическое решение.
в уравнениях (4.57), (4.58) функция источника отсутствует и можно найти соответствующее аналитическое решение.

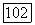 Расчет проводился при
Расчет проводился при 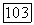
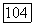 и далее последовательно на всех остальных участках вплоть до
и далее последовательно на всех остальных участках вплоть до 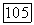 На рис. 4.11 приведено семейство вычисленных по подобной методике безразмерных кривых распределения для ряда значений безразмерного параметра
На рис. 4.11 приведено семейство вычисленных по подобной методике безразмерных кривых распределения для ряда значений безразмерного параметра 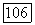 при экспоненциальном законе роста, а на рис. 4.12 — при линейном. На последнем рисунке сплошными линиями проведены кривые, рассчитанные в предположении
при экспоненциальном законе роста, а на рис. 4.12 — при линейном. На последнем рисунке сплошными линиями проведены кривые, рассчитанные в предположении 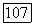 пунктиром — при
пунктиром — при 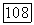 Как видно из рисунка, увеличение
Как видно из рисунка, увеличение 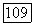 практически не привело к изменению приближенного решения.
практически не привело к изменению приближенного решения.
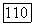 с экспериментальными кривыми распределения тифозных бактерий-палочек по размерам [15, 23—25] показало, что подбором безразмерного параметра
с экспериментальными кривыми распределения тифозных бактерий-палочек по размерам [15, 23—25] показало, что подбором безразмерного параметра 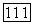 (или двух размерных параметров
(или двух размерных параметров 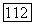 можно добиться
можно добиться

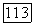 пунктир
пунктир 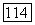
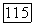 будет определена из независимых экспериментов.
будет определена из независимых экспериментов.