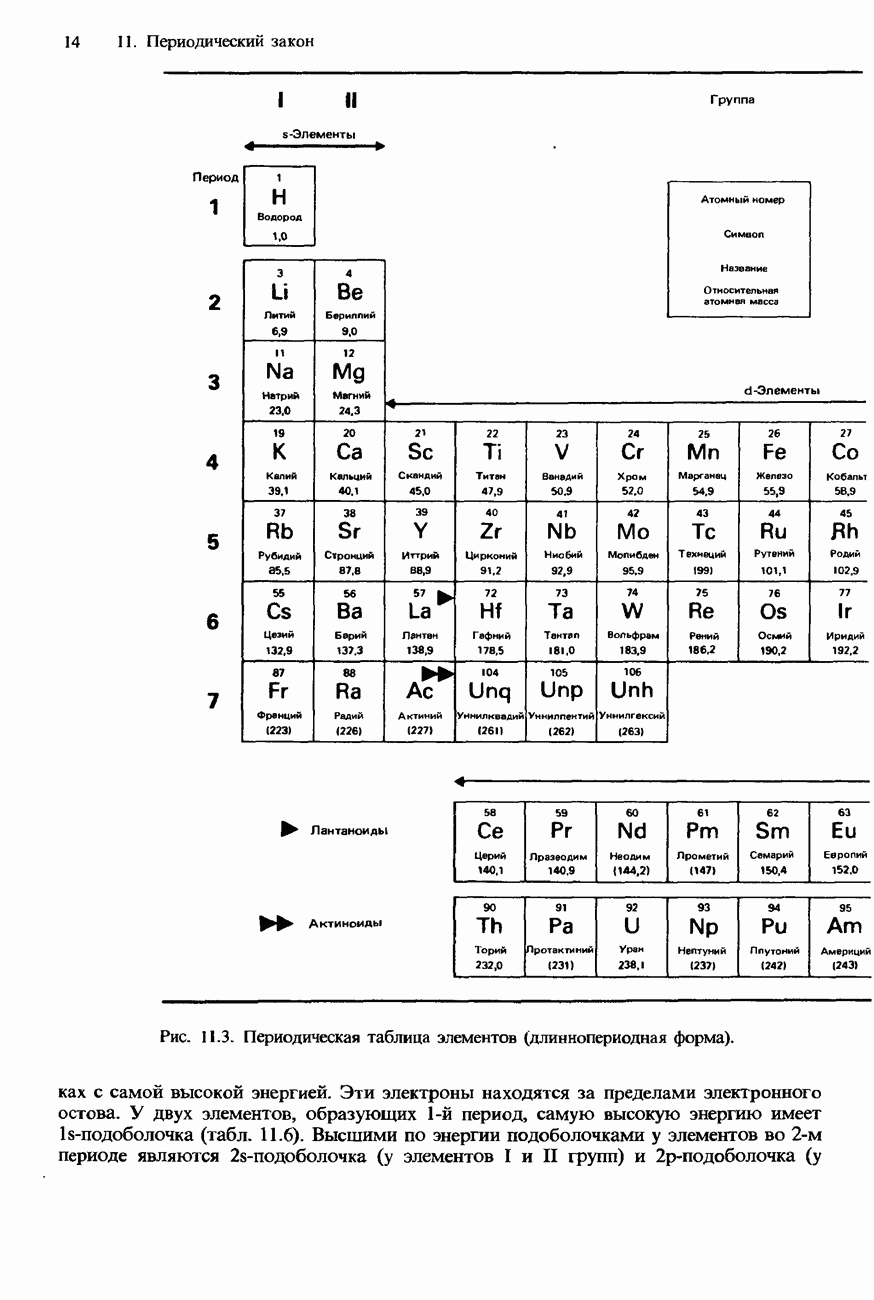
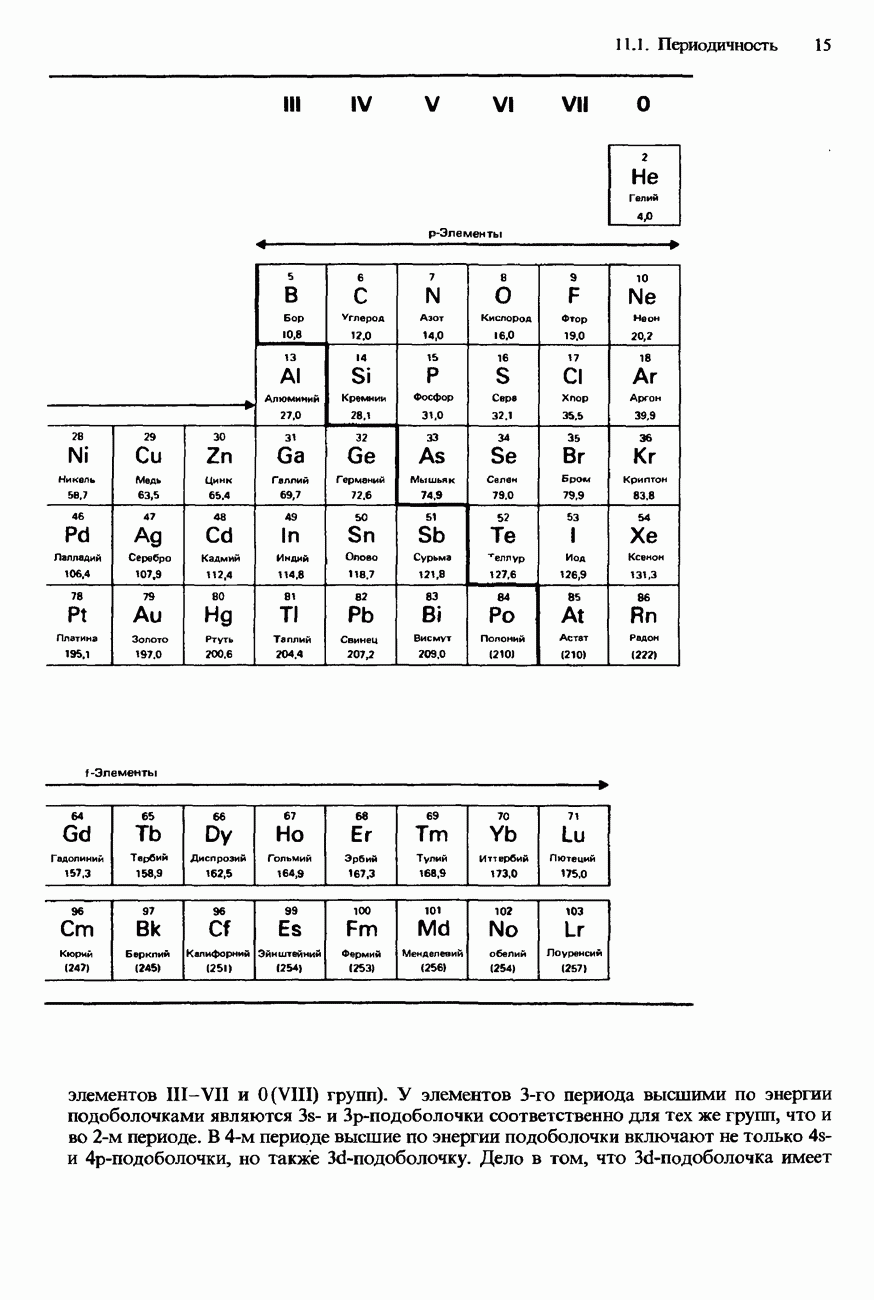
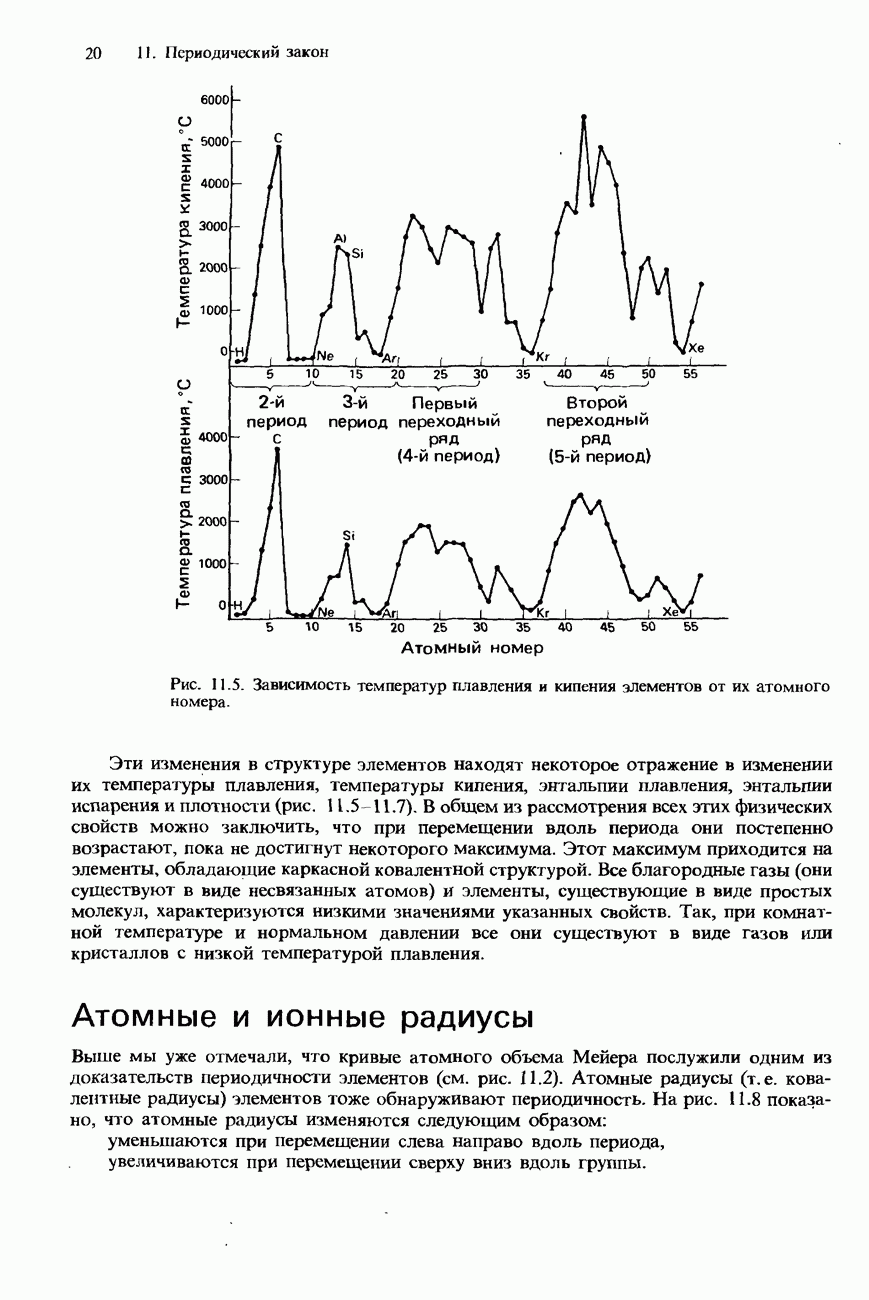
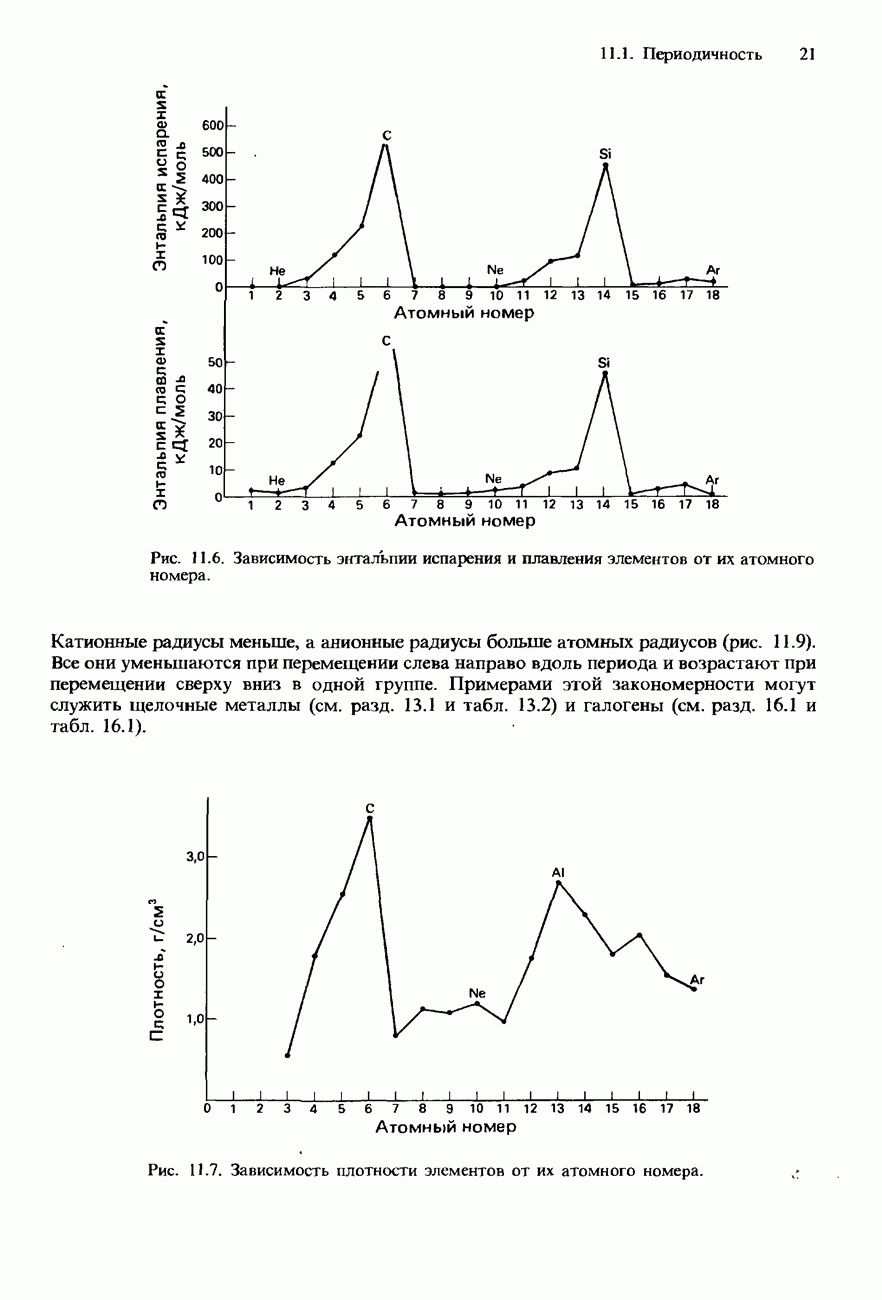
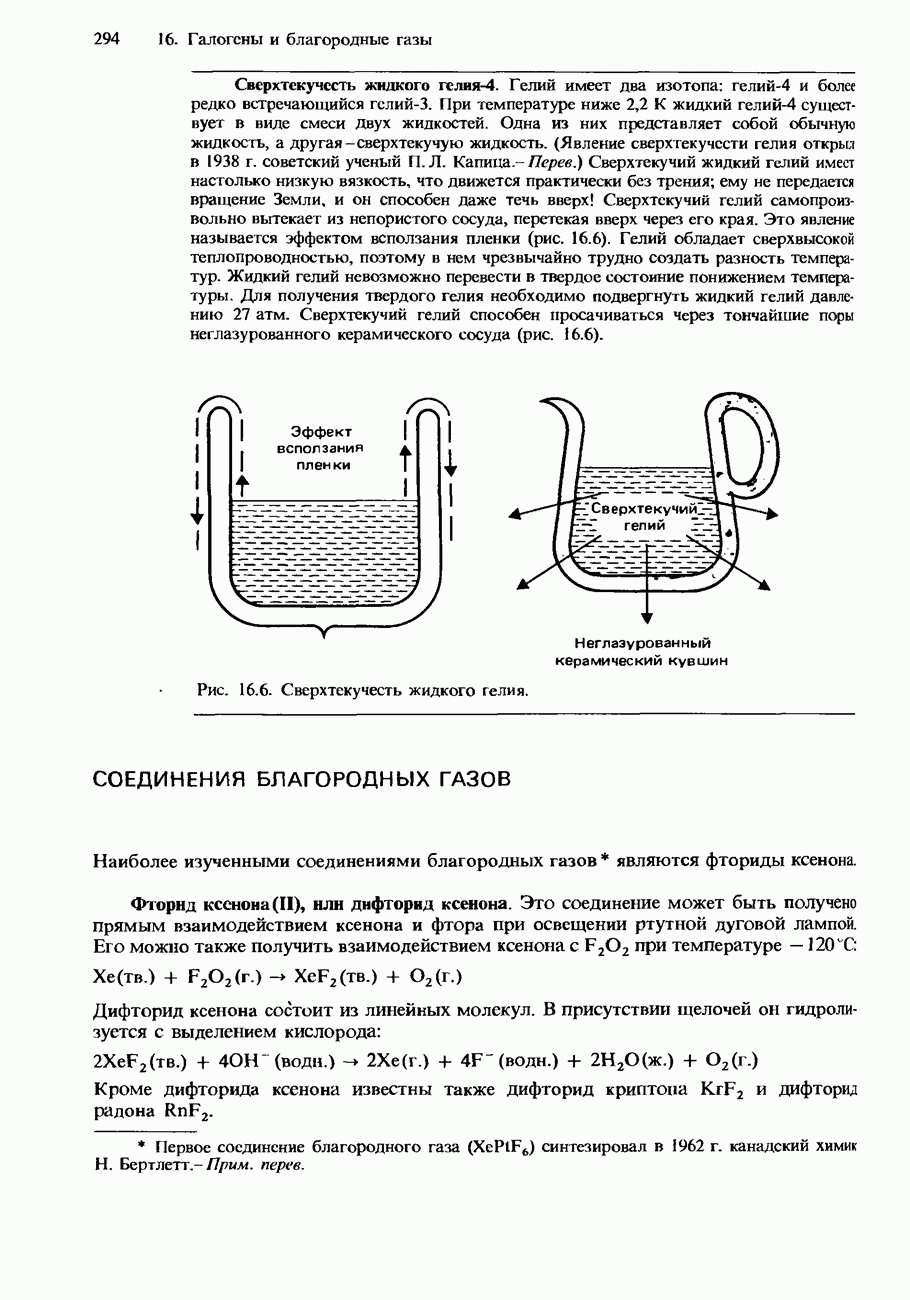
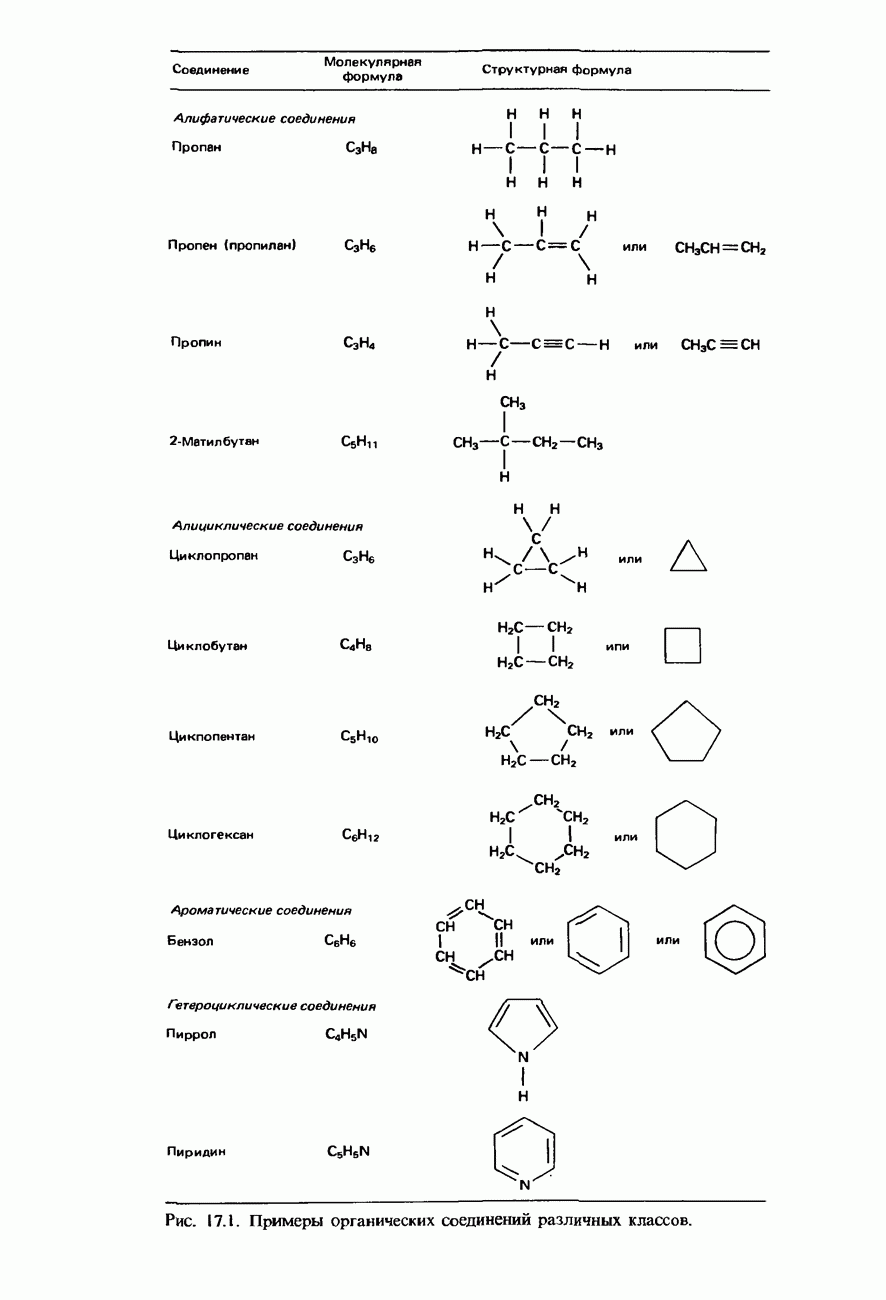
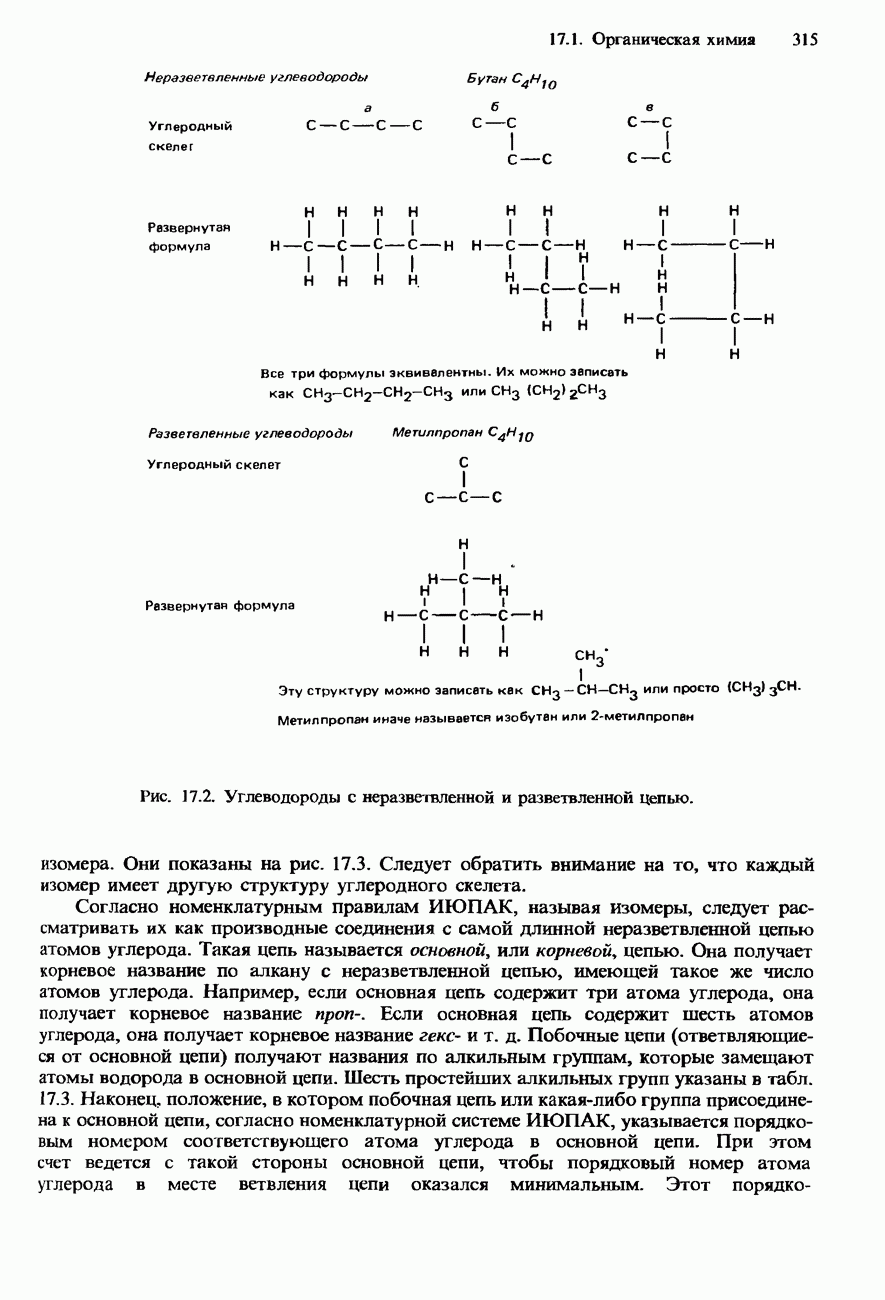
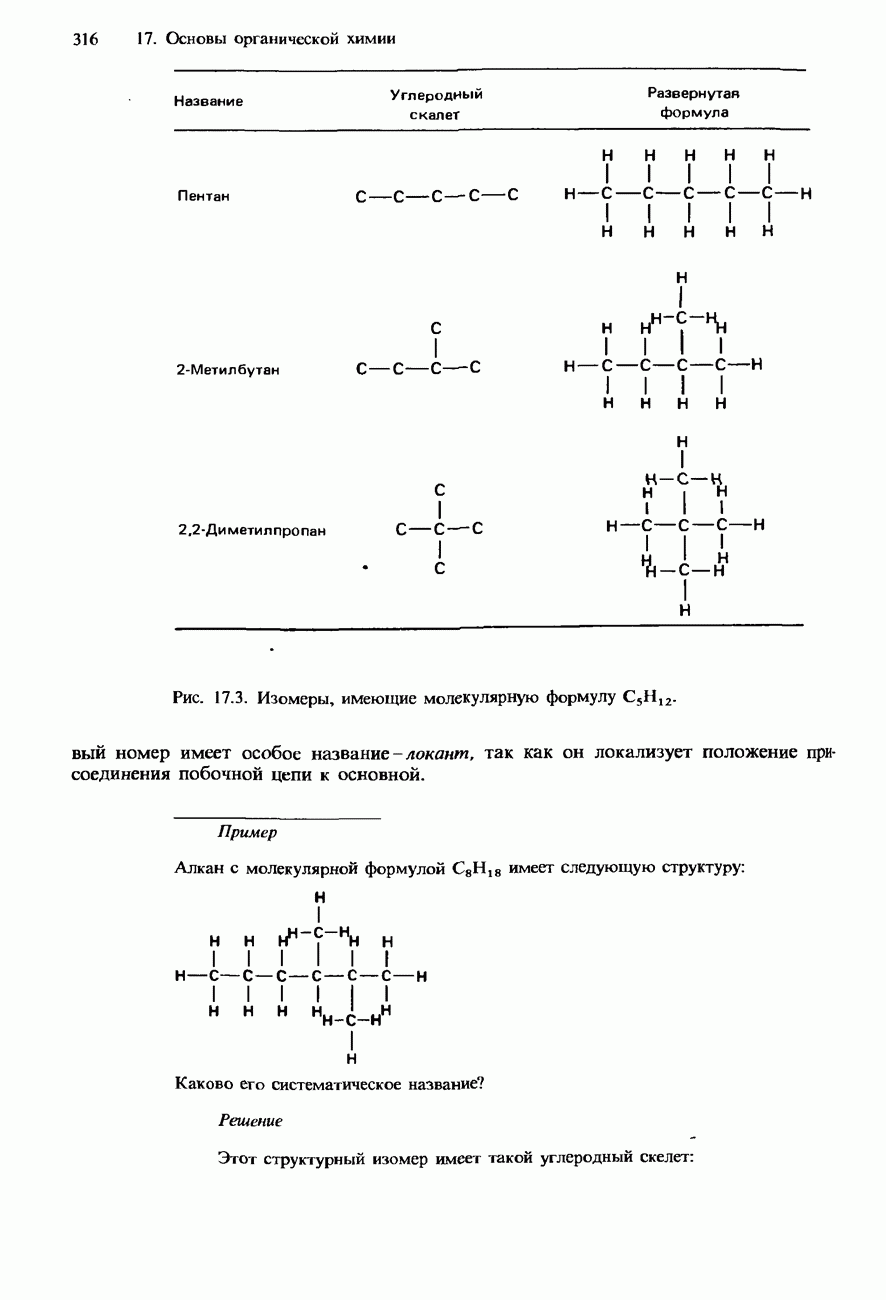
Пред.
След.
Макеты страниц
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
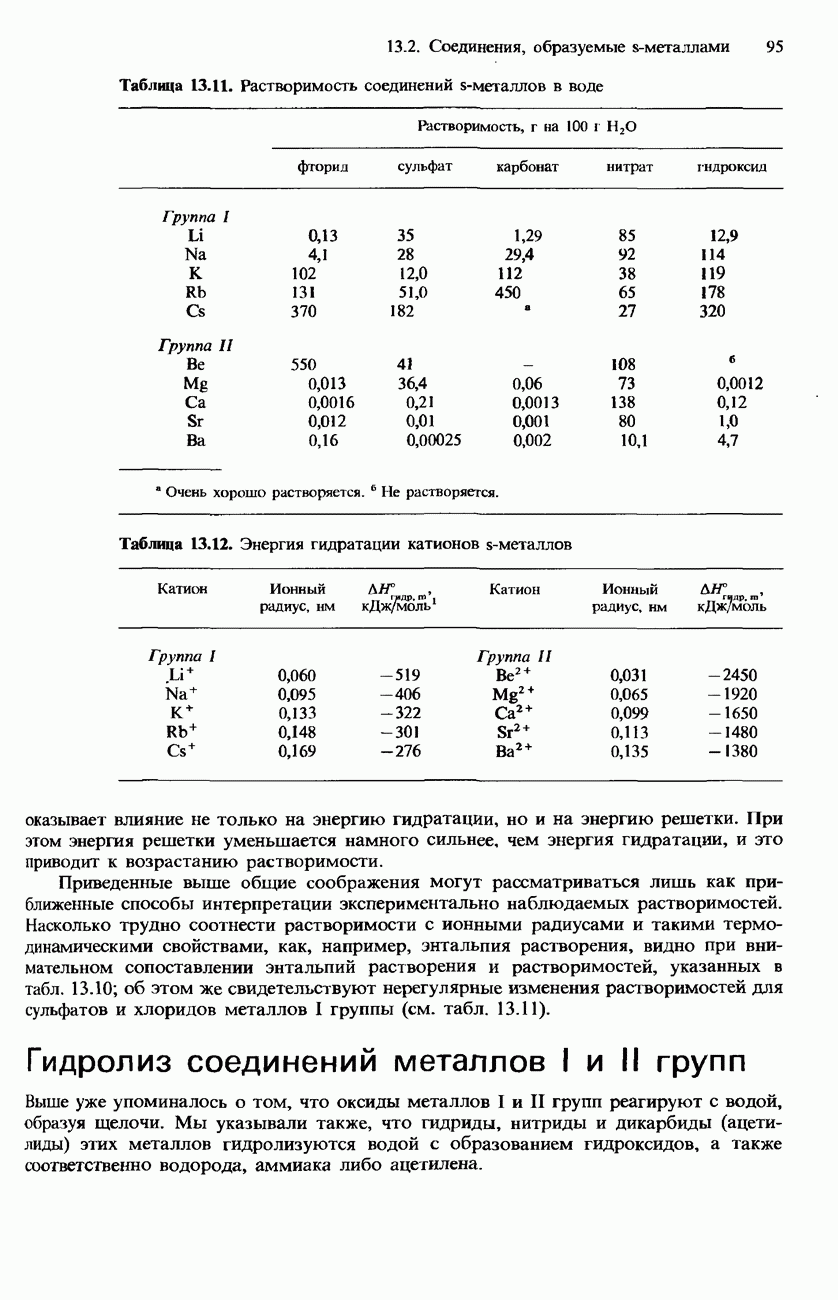
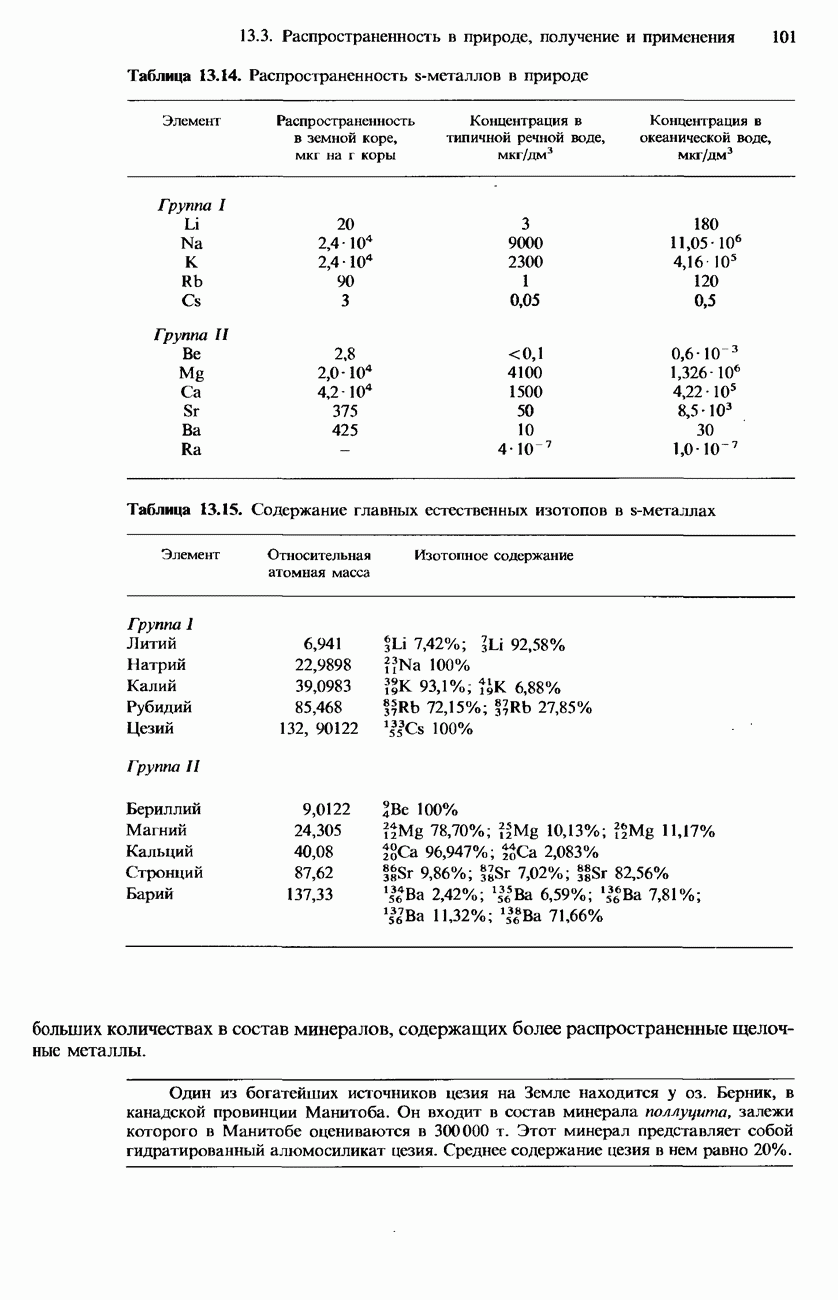
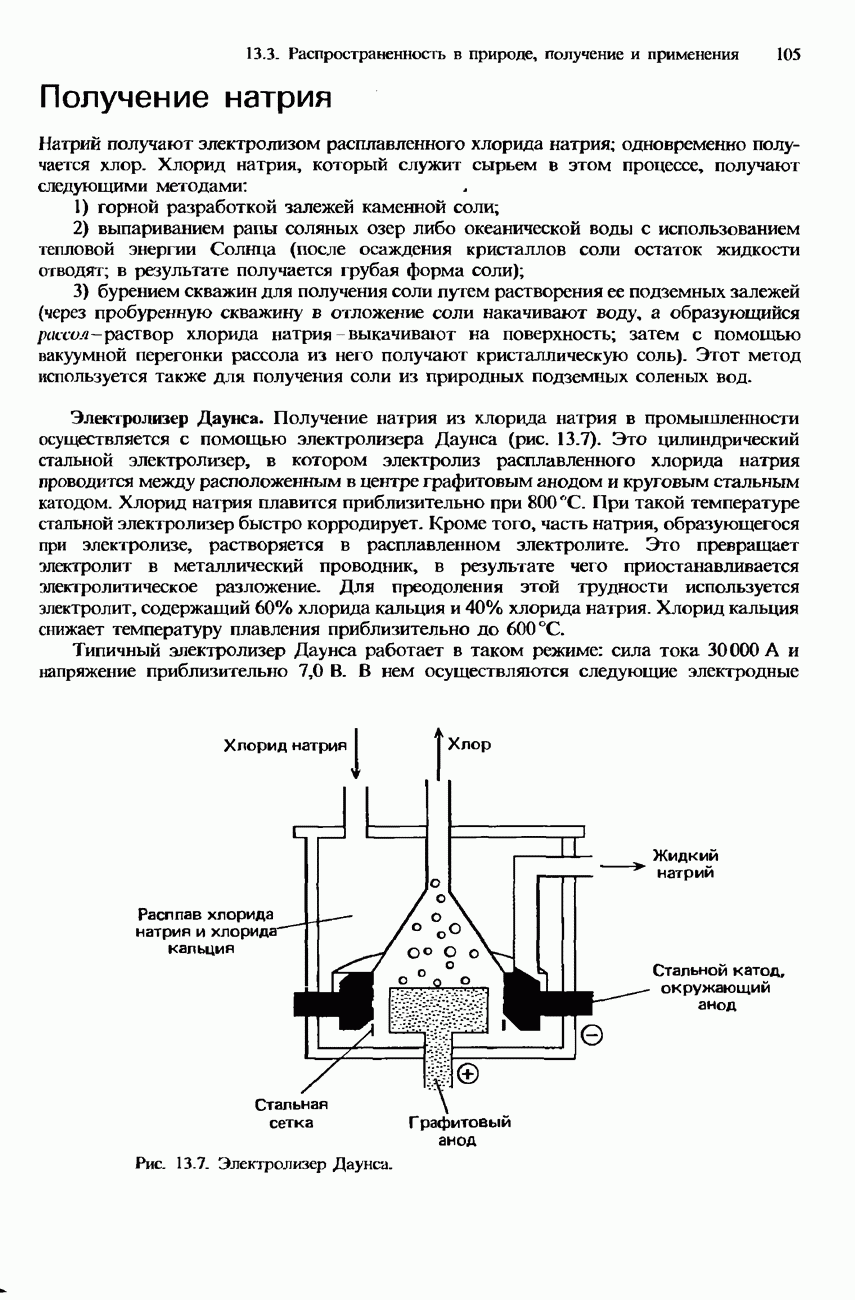
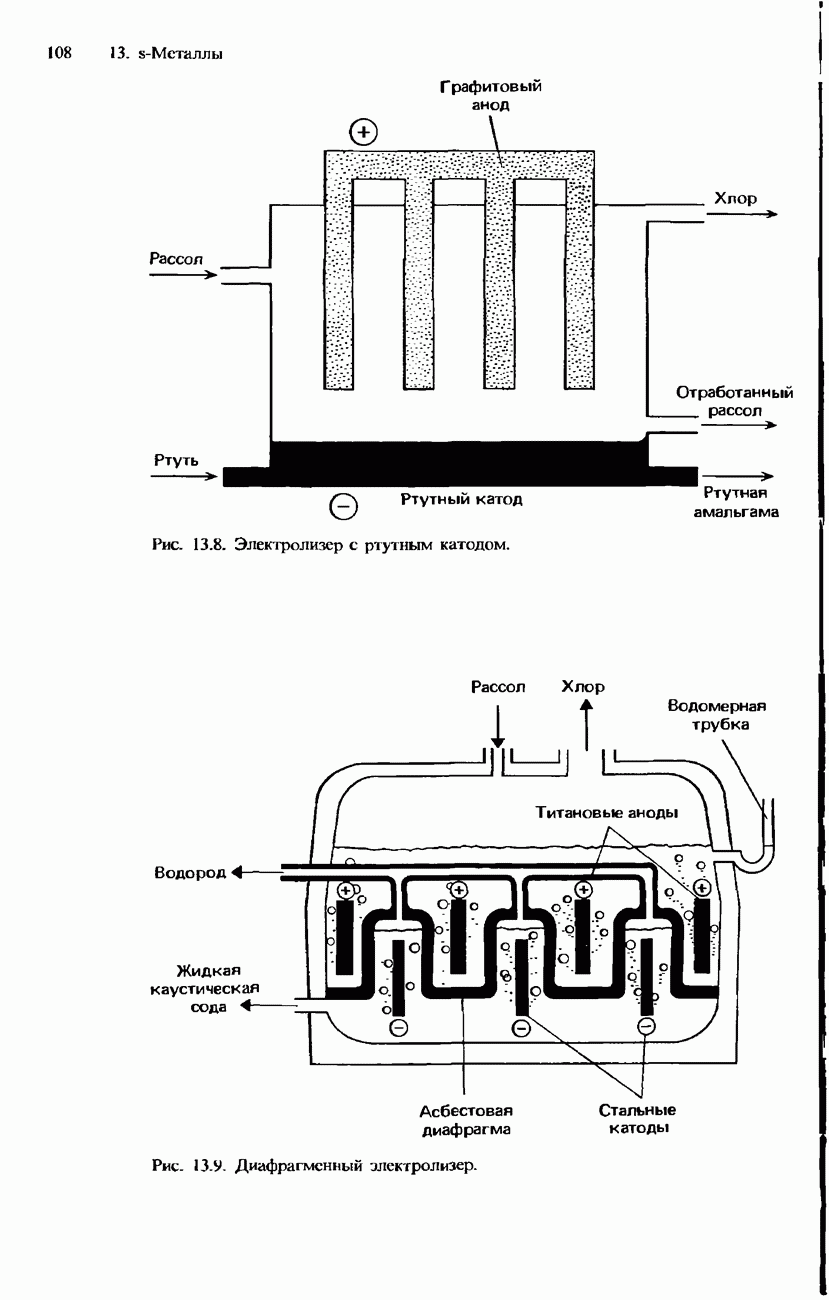
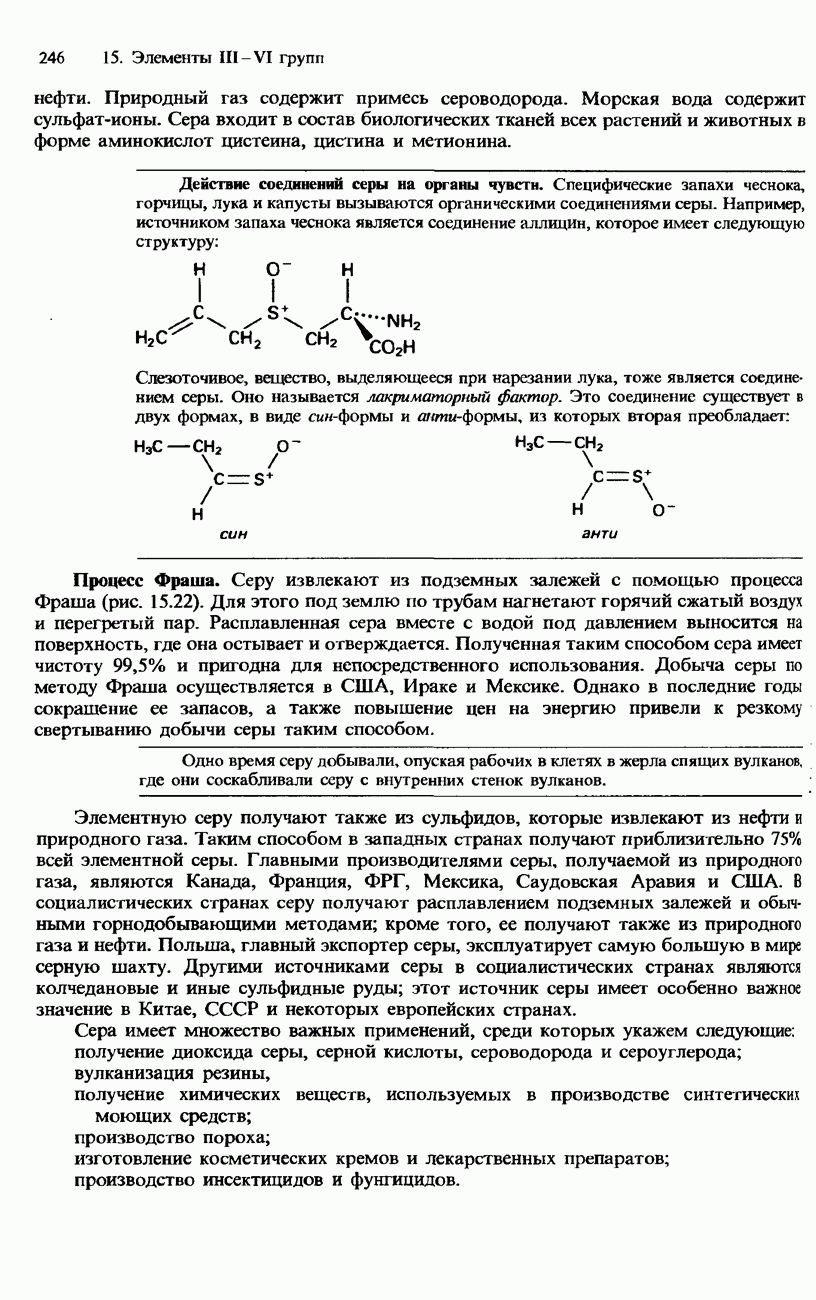
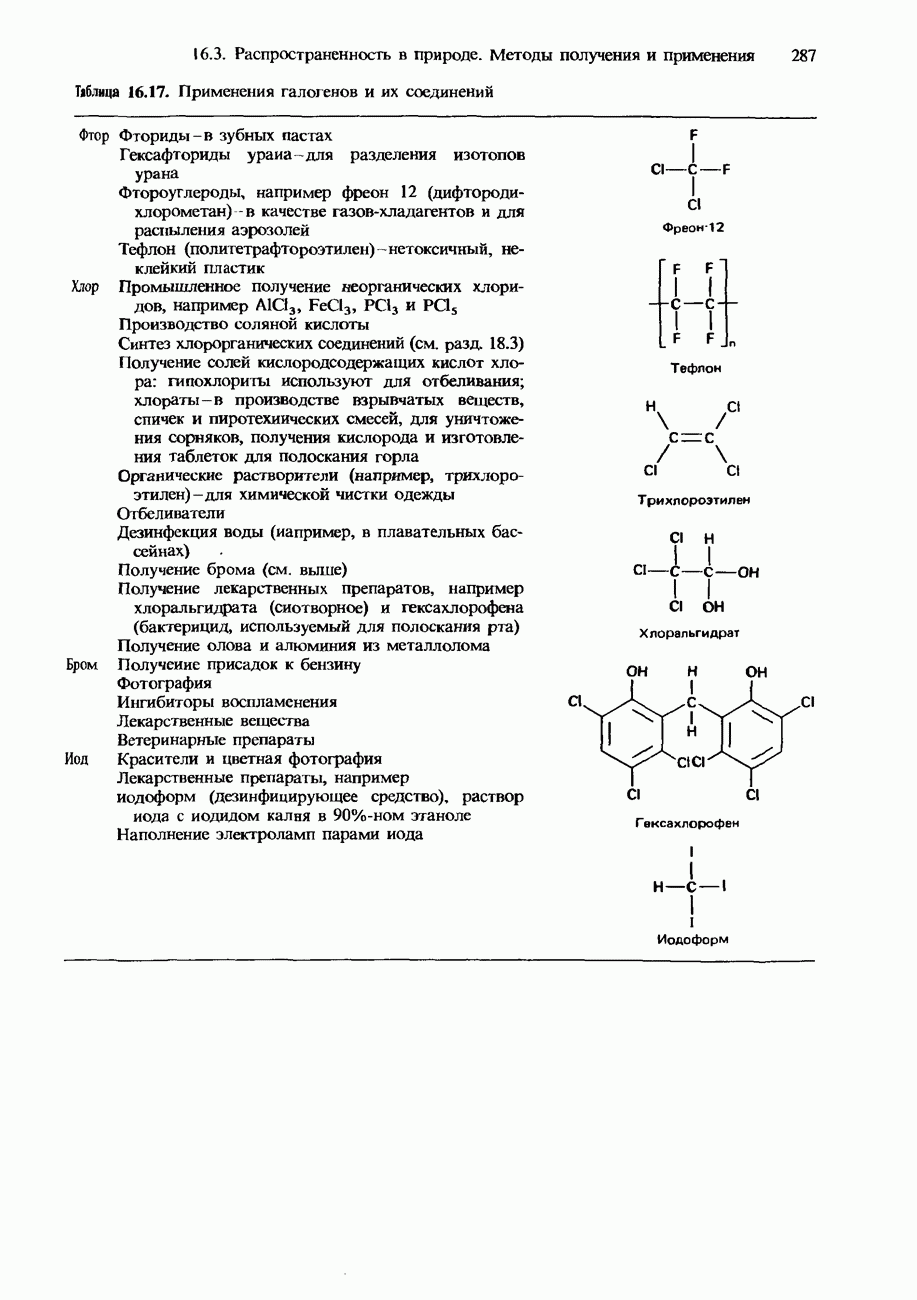
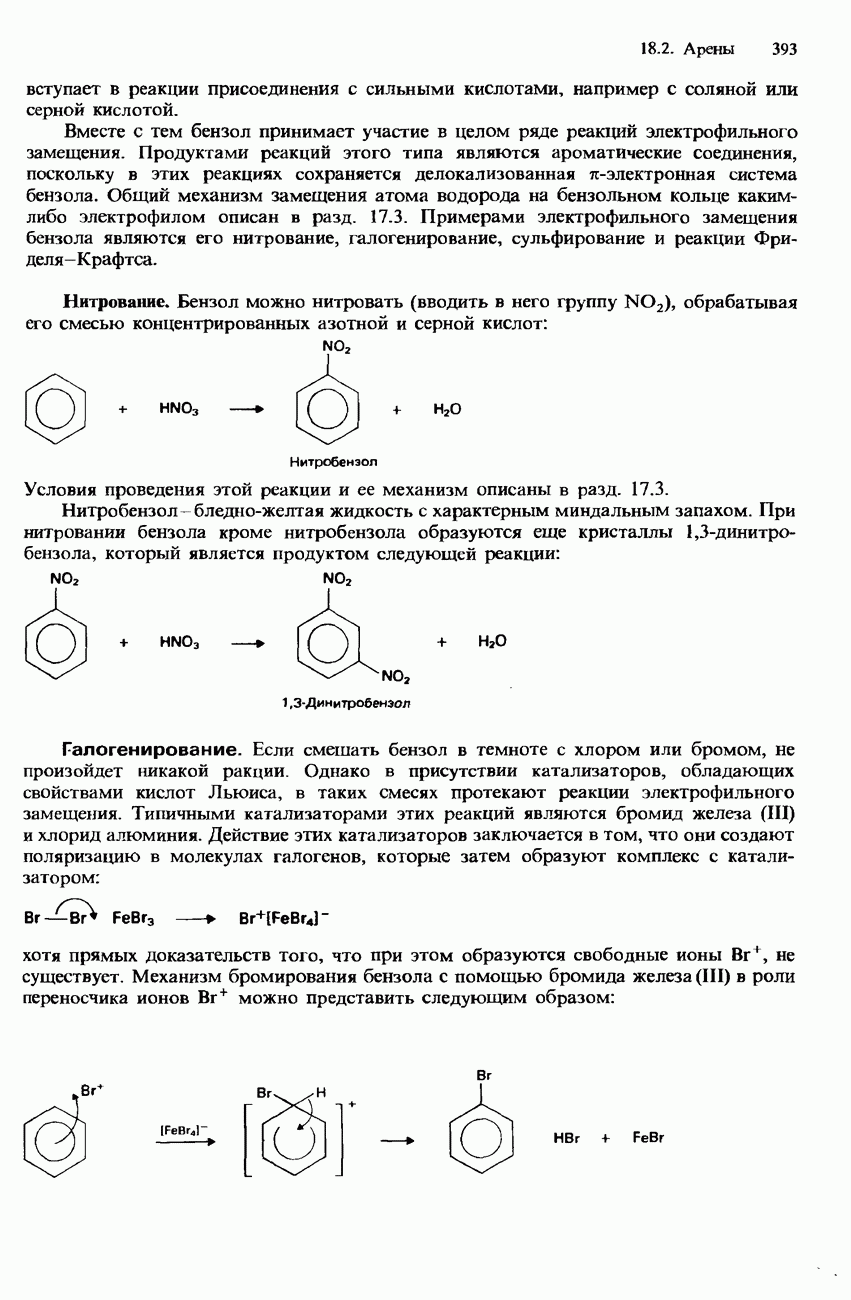
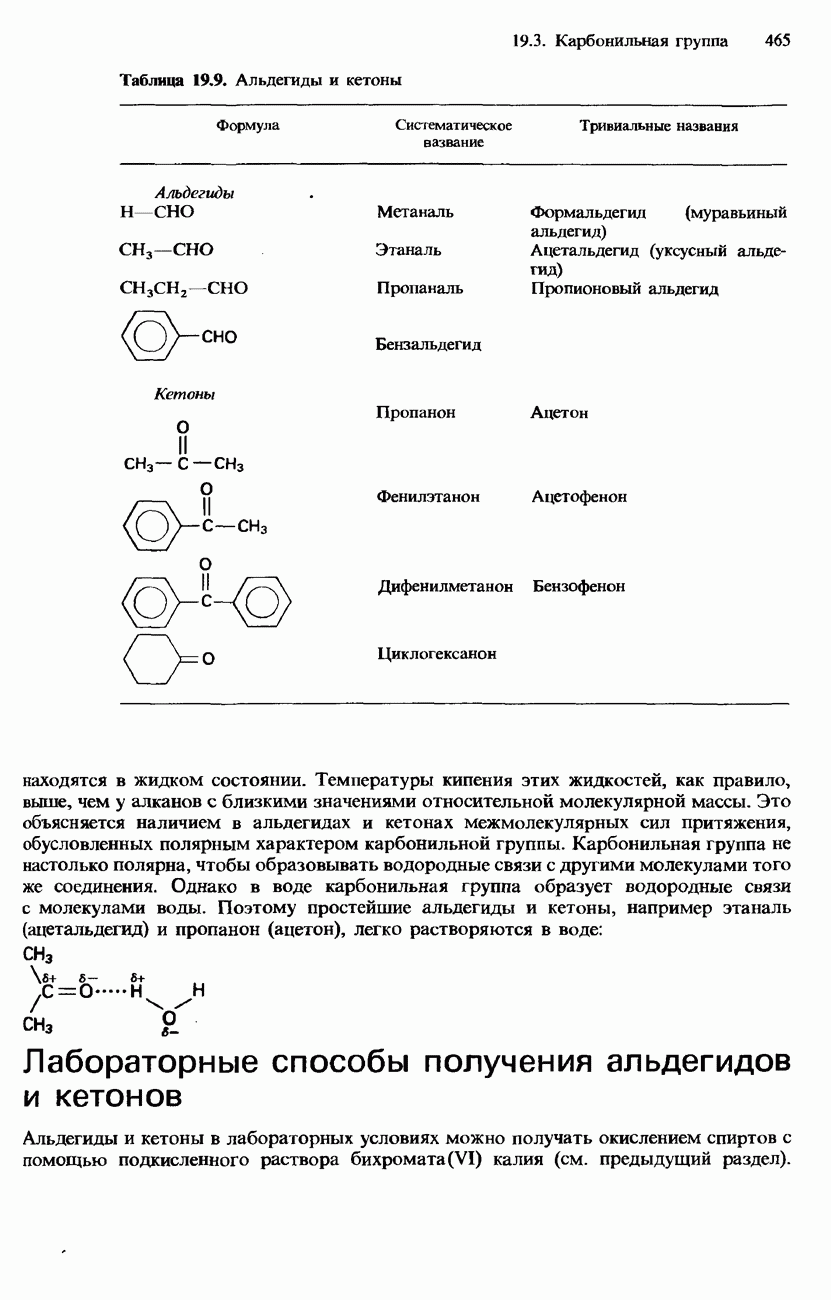
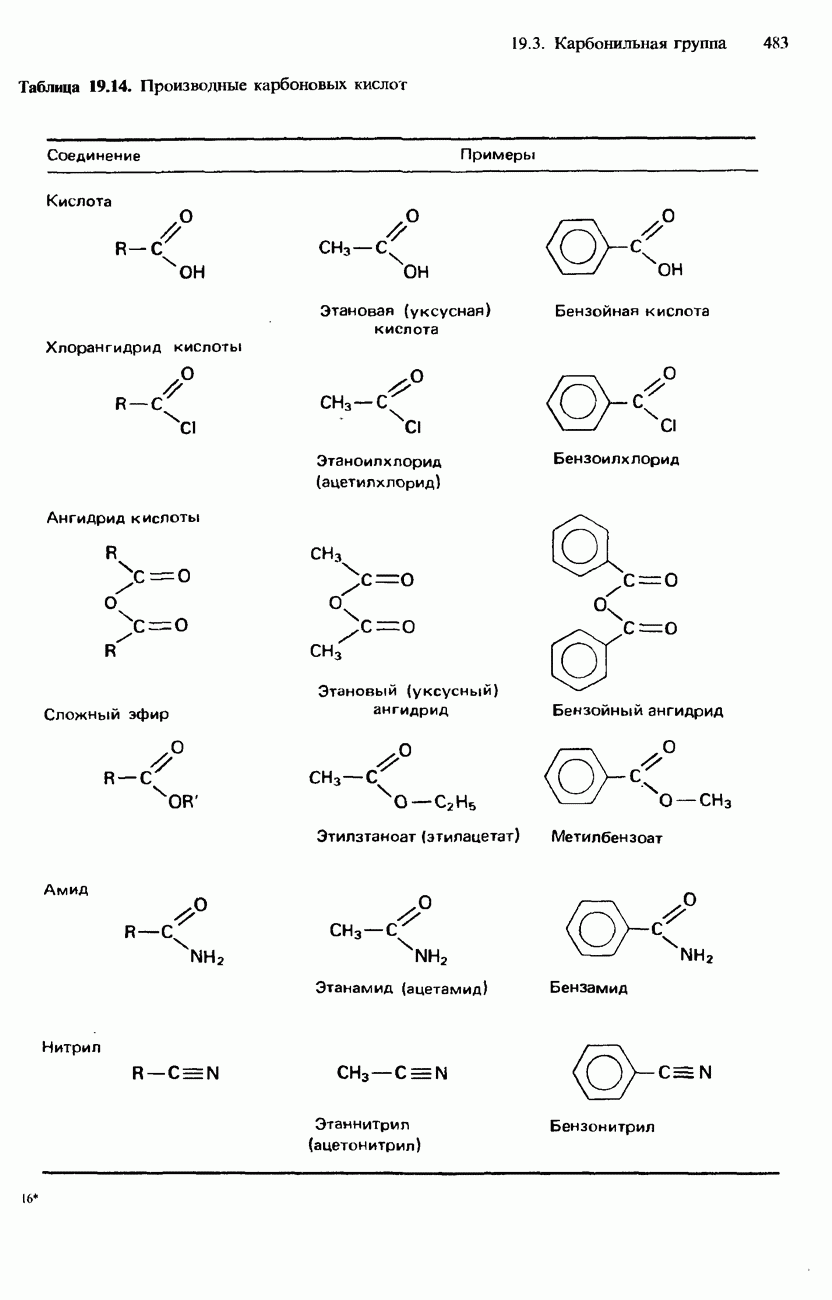
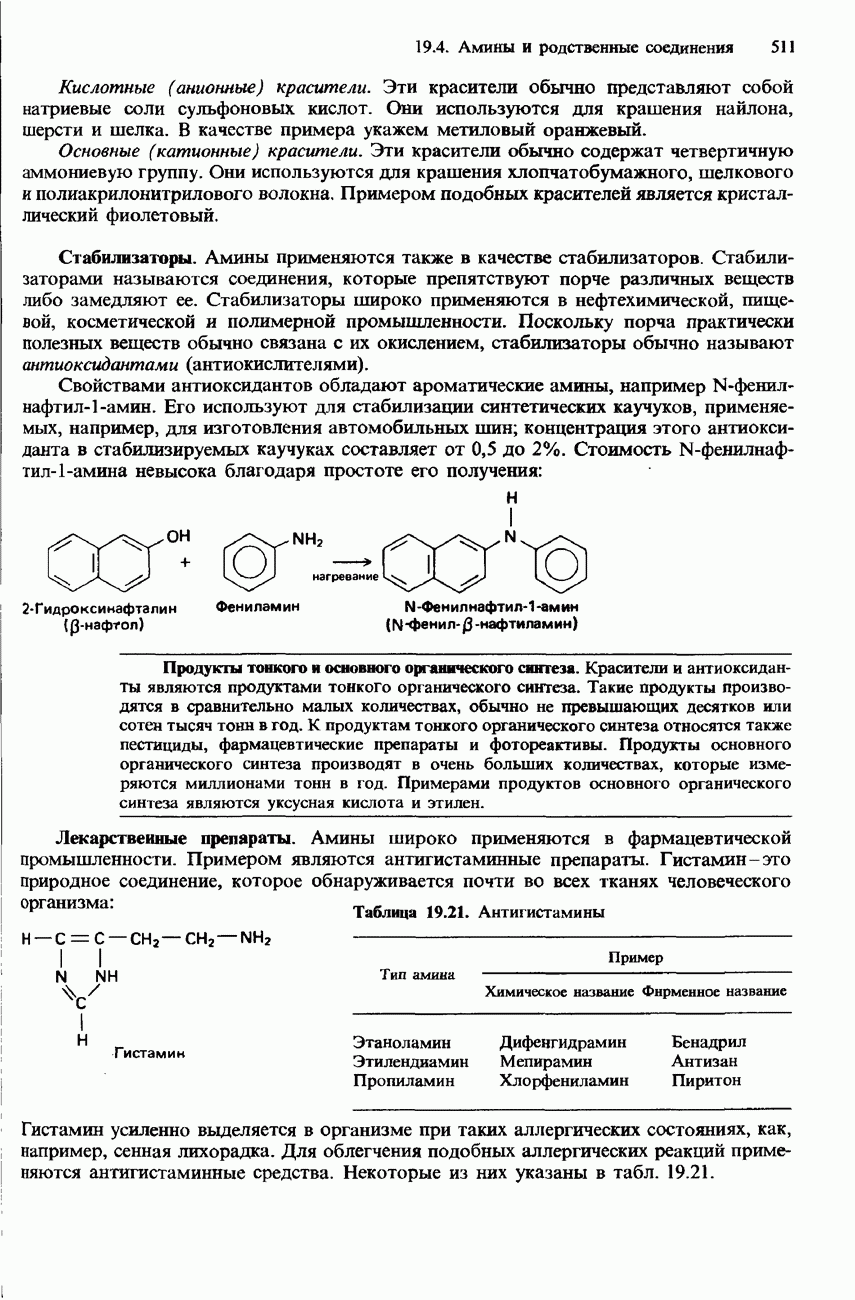
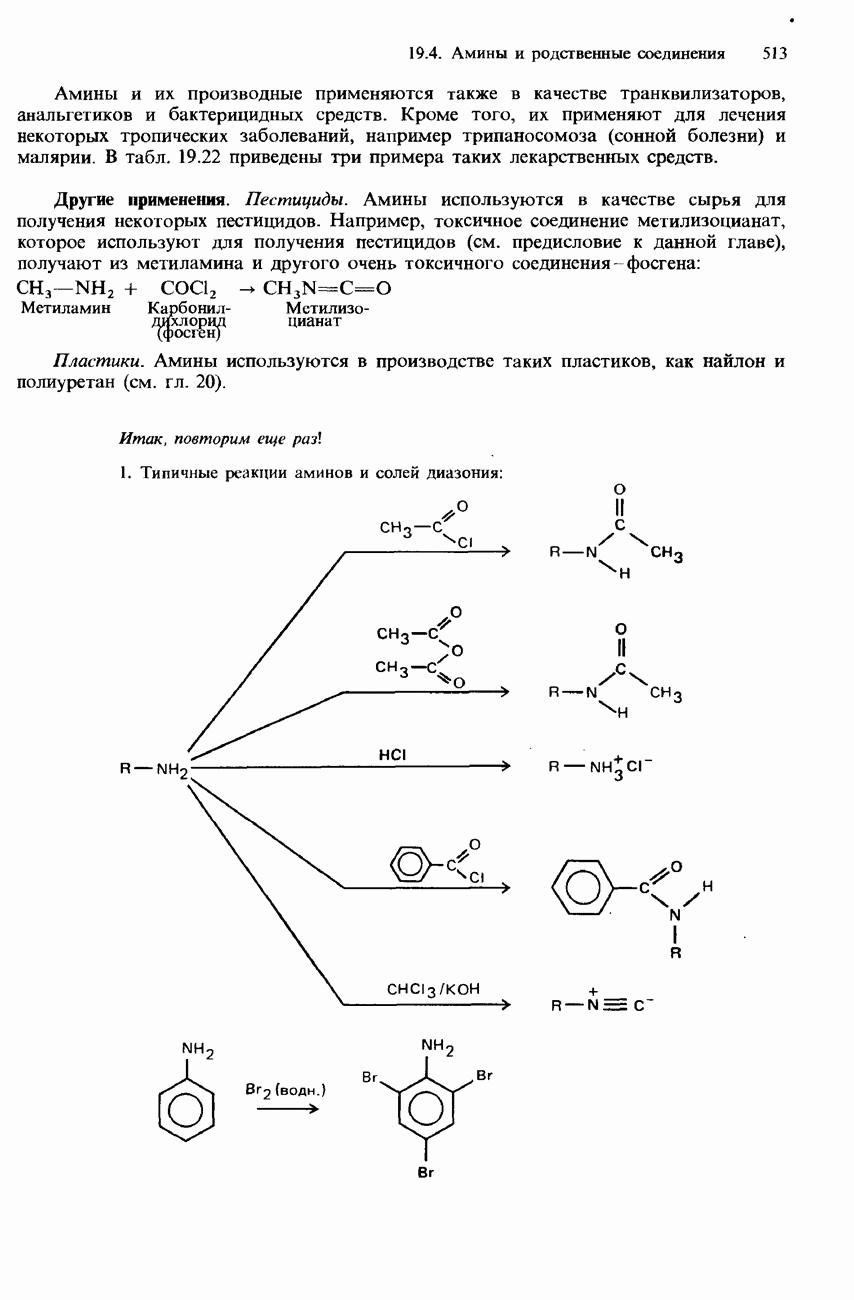
БЕЛКИБелки-это природные полипептиды с высокими значениями относительной молекулярной массы. Эти значения могут колебаться от 10000 до нескольких миллионов. Белки, фрагментами которых являются исключительно Глобулярные белки - это простые белки, растворимые в воде либо образующие в ней коллоидные растворы. Все они растворимы в разбавленных водных растворах кислот, оснований и солей, но осаждаются в концентрированных растворах солей и кислот, а также в органических растворителях. Например, глобулярный белок альбумин, содержащийся в яичном белке, растворим в воде. Белковые гормоны, например инсулин, белки крови и ферменты, тоже представляют собой глобулярные белки. Таблица 20.3. Простетические группы
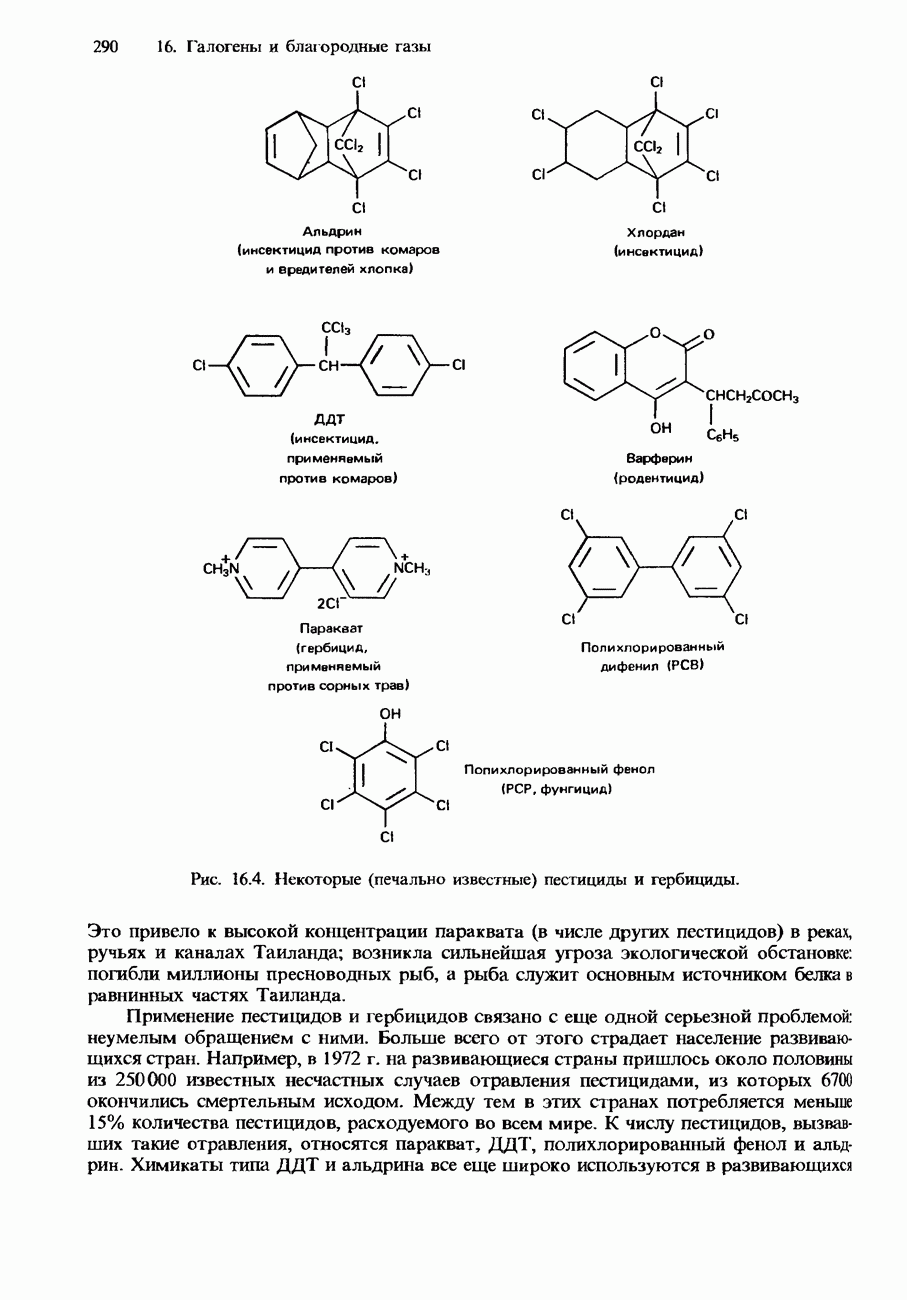
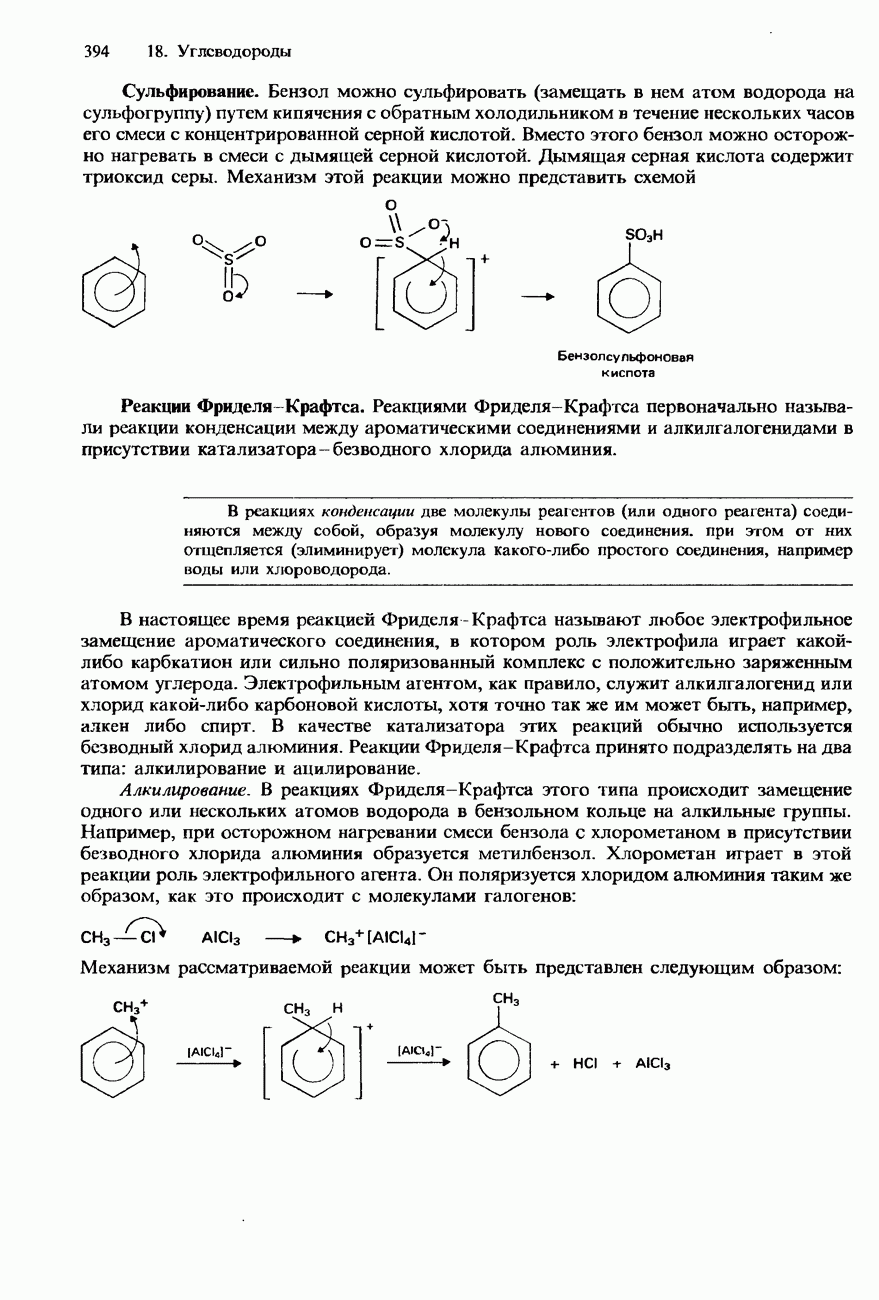
Растворимость и биологическая активность глобулярных белков снижается при повышении температуры и Фибриллярные белки -это простые белки, нерастворимые в воде. Такие белки содержатся в коже, волосах и мускульных тканях и поэтому иногда называются структурными белками. К числу фибриллярных белков относятся, например: кератины-белки, содержащиеся в шерсти, волосах и ногтях; коллагены-белки, содержащиеся в коже и соединительных тканях; фиброин-белок, содержащийся в шелке; эластины-белки, содержащиеся в легких, артериях и некоторых связках. Некоторые белки содержат не только аминокислоты, но и другие группы. Такие неаминокислотные группы называются простетическими (небелковыми) группами. Примеры подобных групп приведены в табл. 20.3. Разделение, аналитическое обнаружение и физические свойства белковБелки обычно существуют в виде смесей. Для их разделения может использоваться методика фракционного осаждения. Эта методика основана на использовании различной растворимости белков в водных растворах солей или в органических растворителях, например в пропаноне (ацетоне) или этаноле. При осторожном добавлении одного из таких растворителей происходит осаждение определенных белков, тогда как другие белки остаются в растворе. Регулируя Для разделения белков применимы не все разновидности хроматографии. Например, хроматография на бумаге вызывает денатурацию белков. Однако для их разделения применимы колоночная хроматография и ионообменная хроматография. В лабораторных исследованиях для этой цели применяется также электрофорез. Белки не характеризуются резко выраженными температурами плавления, Таблица 20.4. Изоэлекгрические точки белков
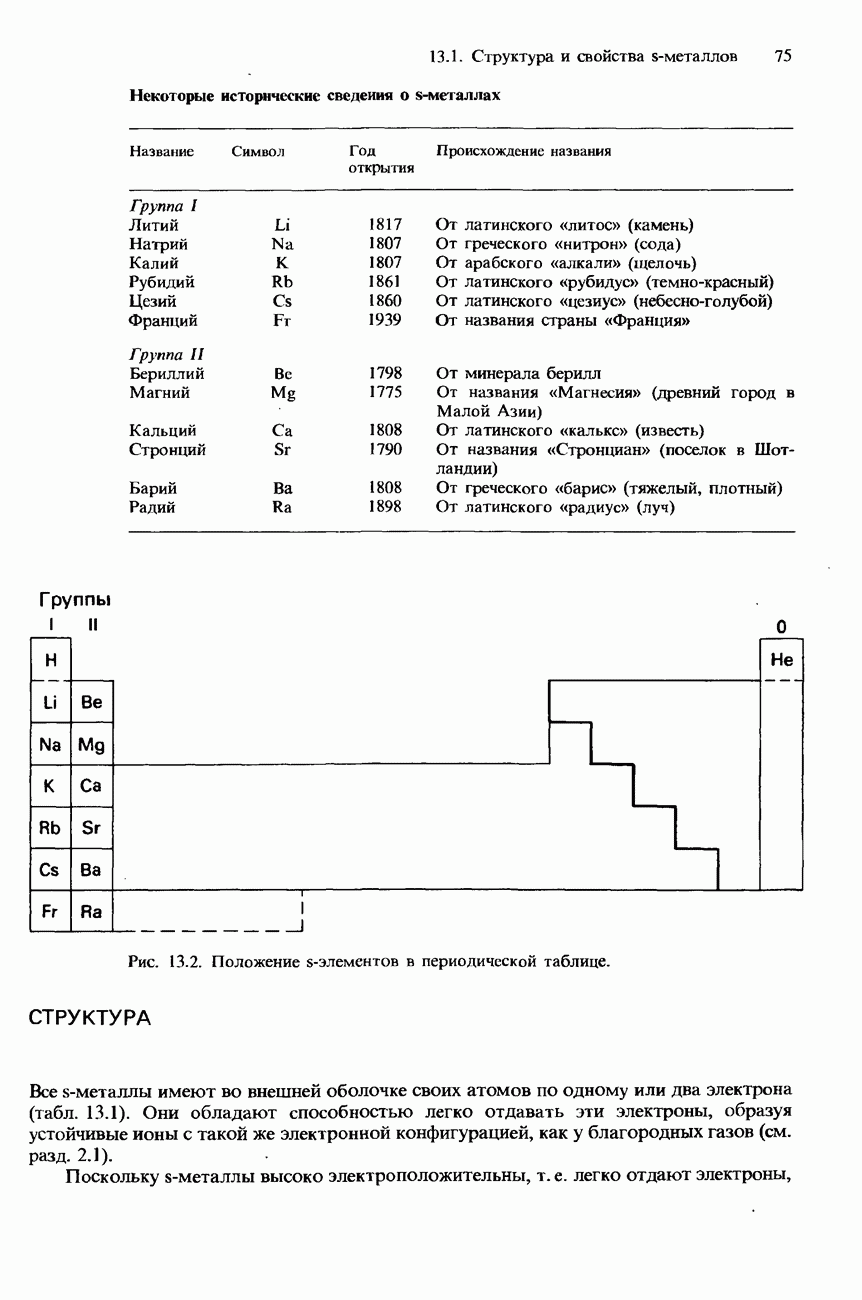
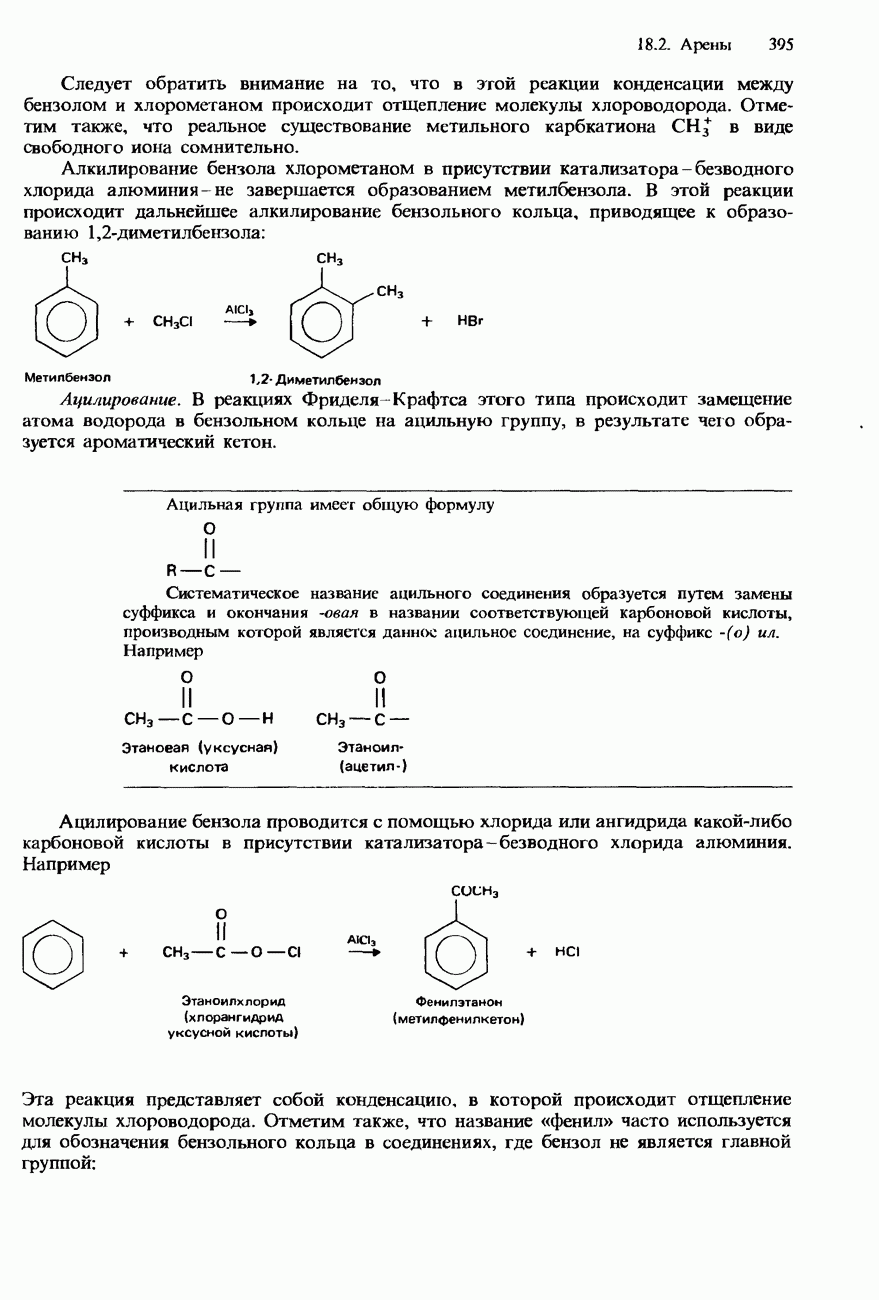
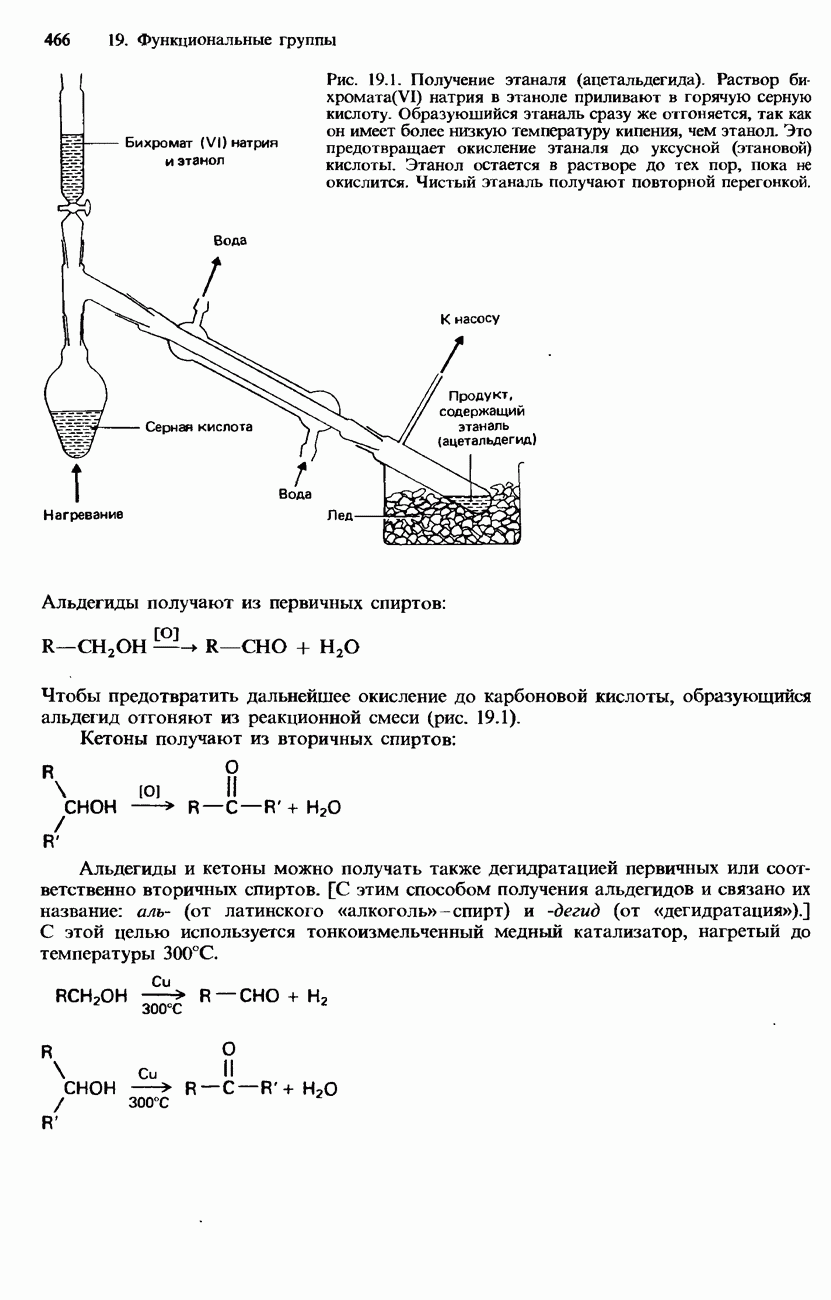
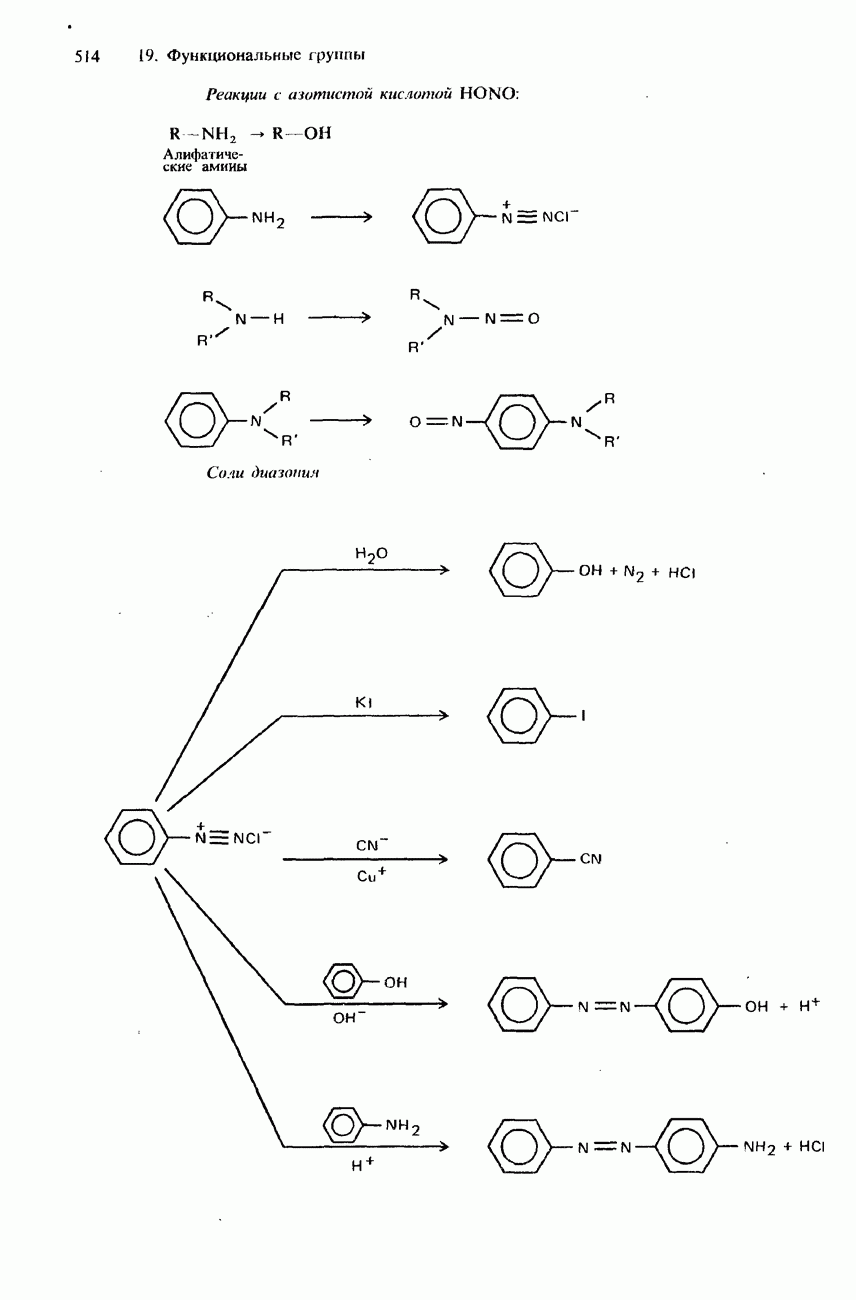
поскольку нагревание вызывает их разложение. Для определения степени чистоты белка и определения его относительной молекулярной массы применяется методика ультрацентрифугирования. Ультрацентрифуги вращают образцы белковых растворов с очень большой скоростью. Скорость перемещения белков к концу пробирки и образования осадка зависит от формы, плотности и относительной молекулярной массы белков. Скорость осаждения чистых белков отличается от скорости осаждения неочищенных белков. Для аналитического обнаружения белков в растворе может использоваться биуретовая проба (см. разд. 19.3). При проведении этой пробы к испытуемому раствору приливают щелочный раствор, содержащий ионы меди(II). Присутствие белков в испытуемом растворе обнаруживается по появлению фиолетовой окраски. Биуретовая реакция может использоваться также для определения концентрации белков в растворе. Интенсивность фиолетовой окраски, определяемая количественно с помощью УФ-спектроскопии (см. разд. 17.1), пропорциональна концентрации белка. Поскольку белки содержат большое число способных диссоциировать аминогрупп и карбоксигрупп, они являются электролитами. При низких значениях pH белкового раствора в нем преобладают положительно заряженные аминогруппы и, следовательно, белки находятся в катионной форме. При высоких значениях pH белкового раствора белки находятся в анионной форме. Подобно аминокислотам, белки тоже имеют свои характеристические изоэлектрические точки (табл. 20.4). В своей изоэлектрической точке каждый белок имеет минимальную растворимость. Показатели кислотности Структура белковБелки обладают очень сложной структурой. Поэтому принято рассматривать четыре различных элемента структуры белков: первичную, вторичную, третичную и четвертичную структуры. Первичная структура. Первичная структура белка определяется последовательностью ковалентно связанных аминокислотных остатков в полипептидной цепи. Например, бычий инсулин состоит из двух полипептидных цепей, одна из которых содержит 21 аминокислотный остаток, а другая - 30 (рис. 20.1). Число аминокислот, содержащихся в белке или полипептиде, а также их конкретную природу можно определить при помощи гидролиза. С этой целью белки или полипептиды нагревают в разбавленном растворе какой-либо кислоты, например соляной. Частичный гидролиз белка или полипептида приводит к образованию смеси
Рис. 20.1. Бычий инсулин состоит из двух полипептидных цепей, в одной из которых содержится 21 аминокислотный остаток, а в другой - 30. Эти две цепи связаны между собой дисульфидными мостиковыми связями. пептидов и аминокислот. При гидролизе белков образуется максимум 20 аминокислот. Их можно разделить при помощи хроматографии либо электрофореза. Затем для идентификации аминокислот используют надлежащие окрашивающие реактивы, например нингидрин. Разумеется, некоторые аминокислоты могут встречаться в аминокислотной последовательности полипептидной цепи по нескольку раз. Количественное содержание каждой аминокислоты в пептидной цепи можно определить по интенсивности окраски, для измерения которой используется УФ-спектроскопия (см. разд. 17.1). Для определения конкретной последовательности аминокислот в полипептидной цепи используется метод анализа концевых аминогрупп. С этой целью метят аминогруппу на одном конце полипептидной цепи с помощью 2,4-динитрофторобензола:
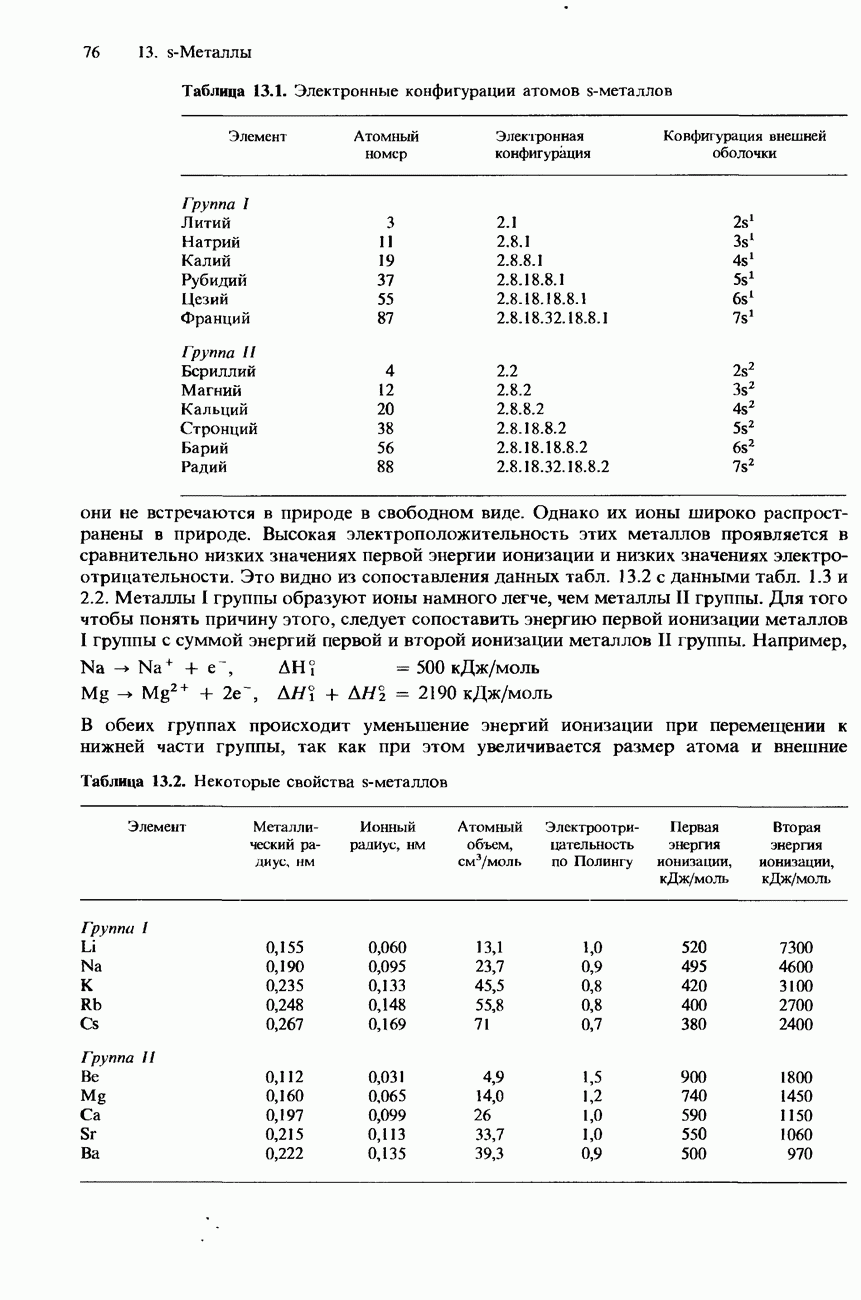
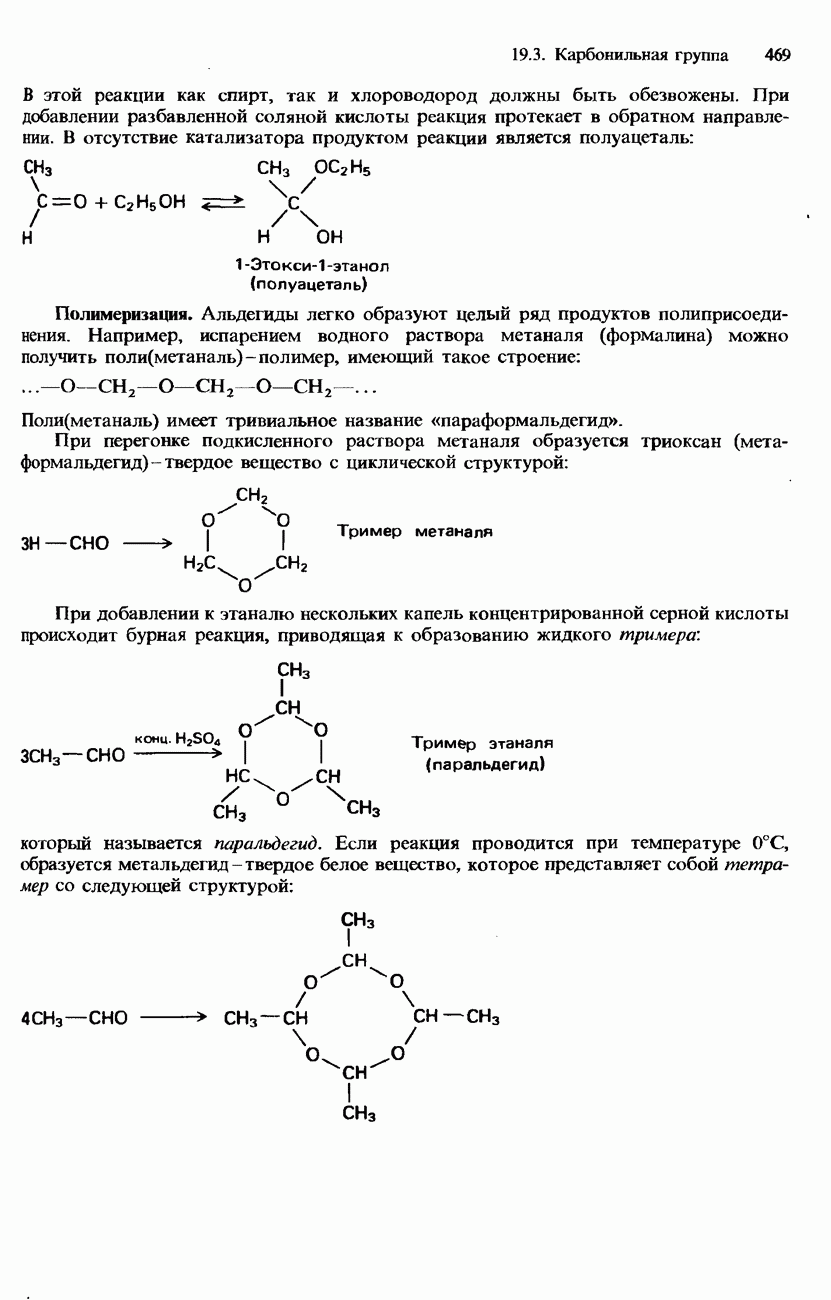
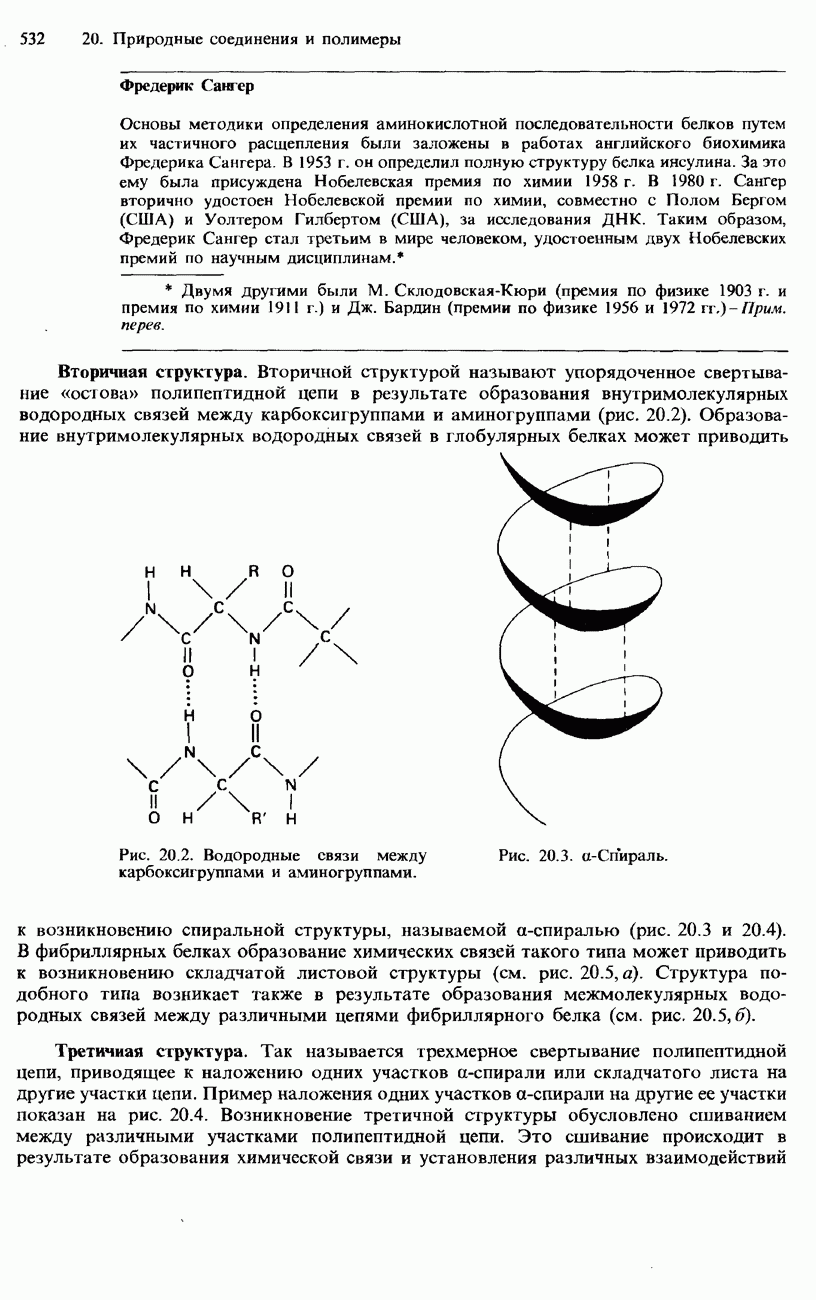
После гидролиза белка меченую аминокислоту можно обнаружить с помощью хроматографии на бумаге. Меченая аминокислота дает на хроматограмме желтое пятно. Используя определенные селективные химические реактивы и ферменты, можно удалять по одной аминокислоте с меченого конца цепи. Однако по мере укорочения цепи такая процедура все более усложняется и становится все менее точной. Поэтому большее распространение получила методика, называемая картированием пептидов. При использовании этой методики полипептидную цепь разрушают при помощи какого-либо расщепляющего реактива, что позволяет разделить фрагменты пептидной цепи. Для таких фрагментов легче установить аминокислотную последовательность. Используя два или несколько расщепляющих реактивов, удается составить полную карту полипептидной цепи. Для расщепления полипептидной цепи используется «химический» гидролиз либо применяется какой-либо фермент, например трипсин. Фредерик Сангер Основы методики определения аминокислотной последовательности белков путем их частичного расщепления были заложены в работах английского биохимика Фредерика Сангера. В 1953 г. он определил полную структуру белка инсулина. За это ему была присуждена Нобелевская премия по химии 1958 г. В 1980 г. Сангер вторично удостоен Нобелевской премии по химии, совместно с Полом Бергом (США) и Уолтером Гилбертом (США), за исследования ДНК. Таким образом, Фредерик Сангер стал третьим в мире человеком, удостоенным двух Нобелевских премий по научным дисциплинам. Вторичная структура. Вторичной структурой называют упорядоченное свертывание «остова» полипептидной цепи в результате образования внутримолекулярных водородных связей между карбоксигруппами и аминогруппами (рис. 20.2). Образование внутримолекулярных водородных связей в глобулярных белках может приводить к возникновению спиральной структуры, называемой
Рис. 20.2. Водородные связи между карбоксигруппами и аминогруппами.
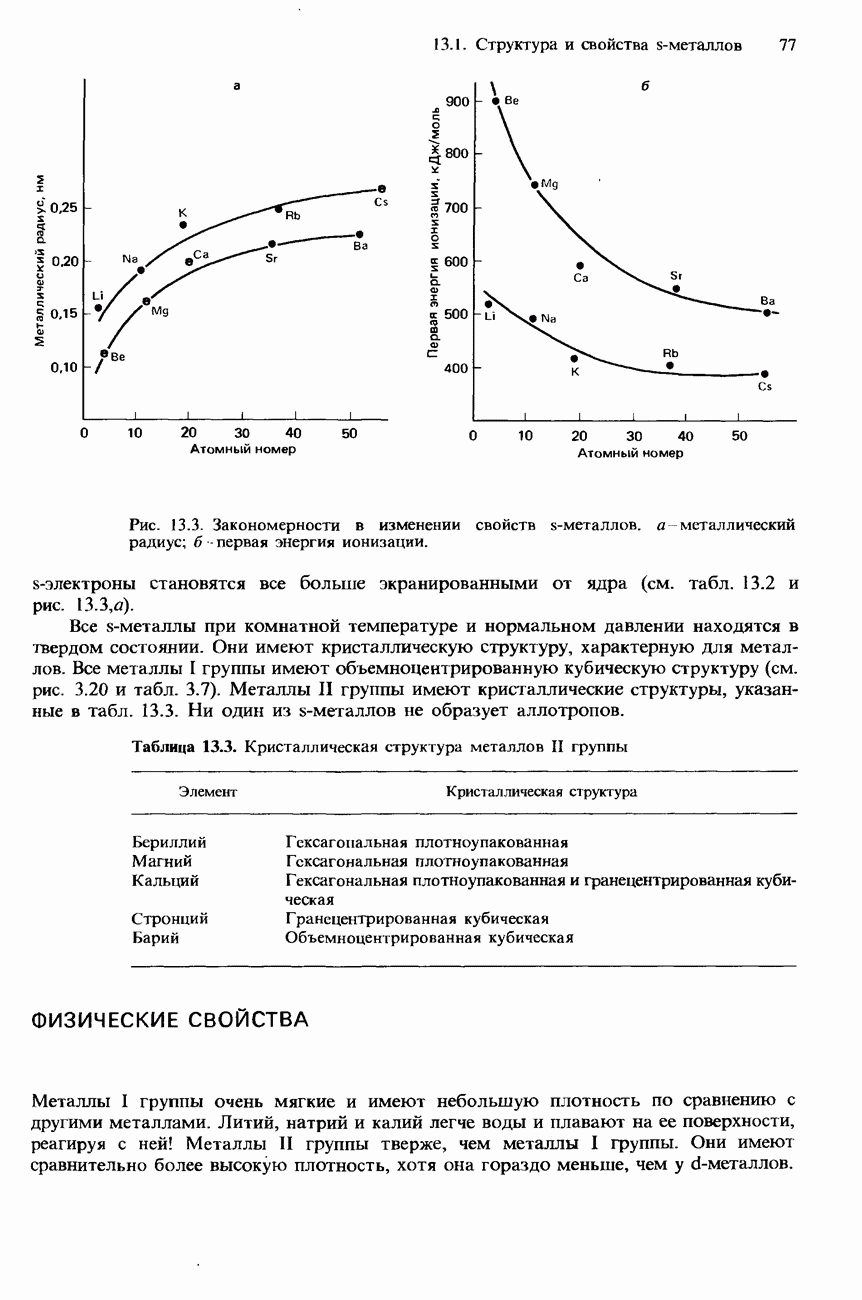
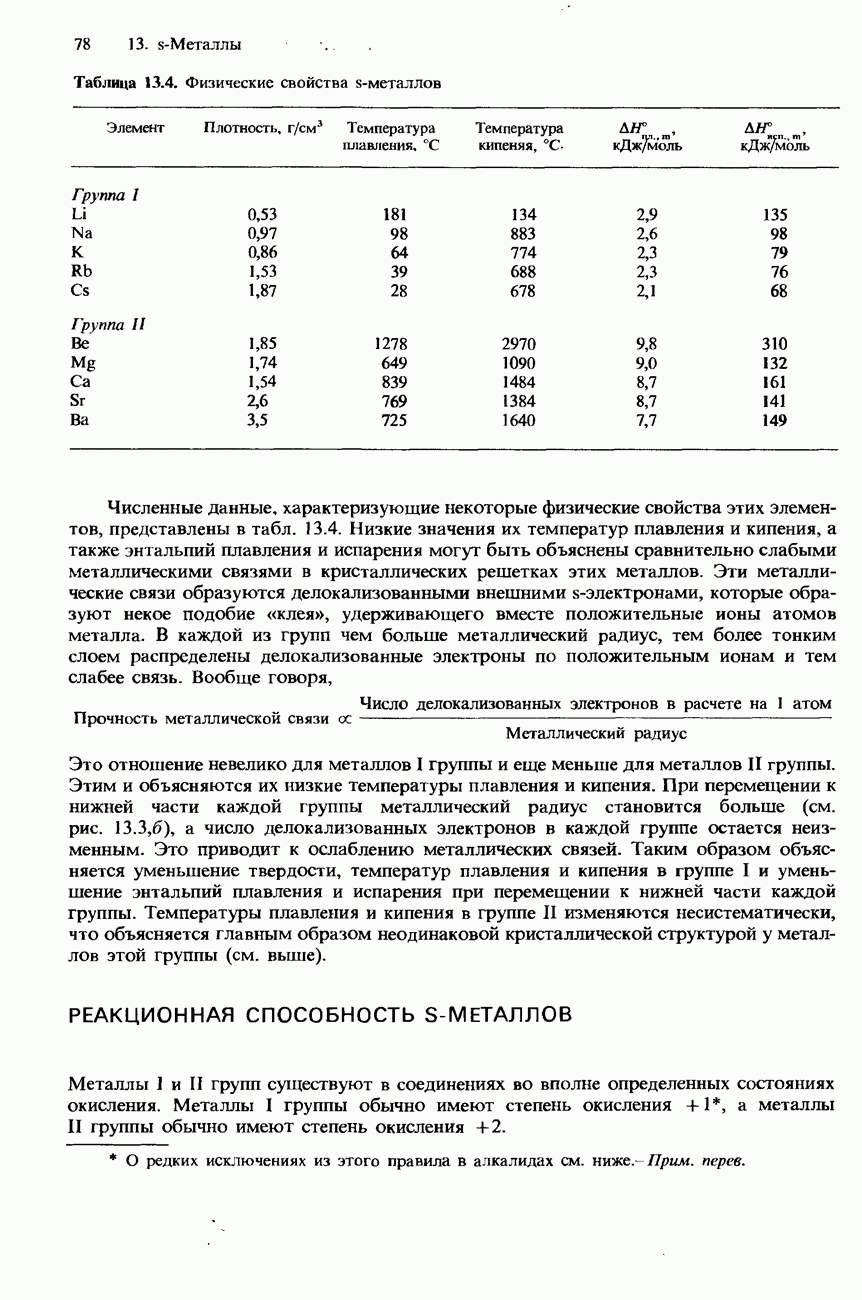
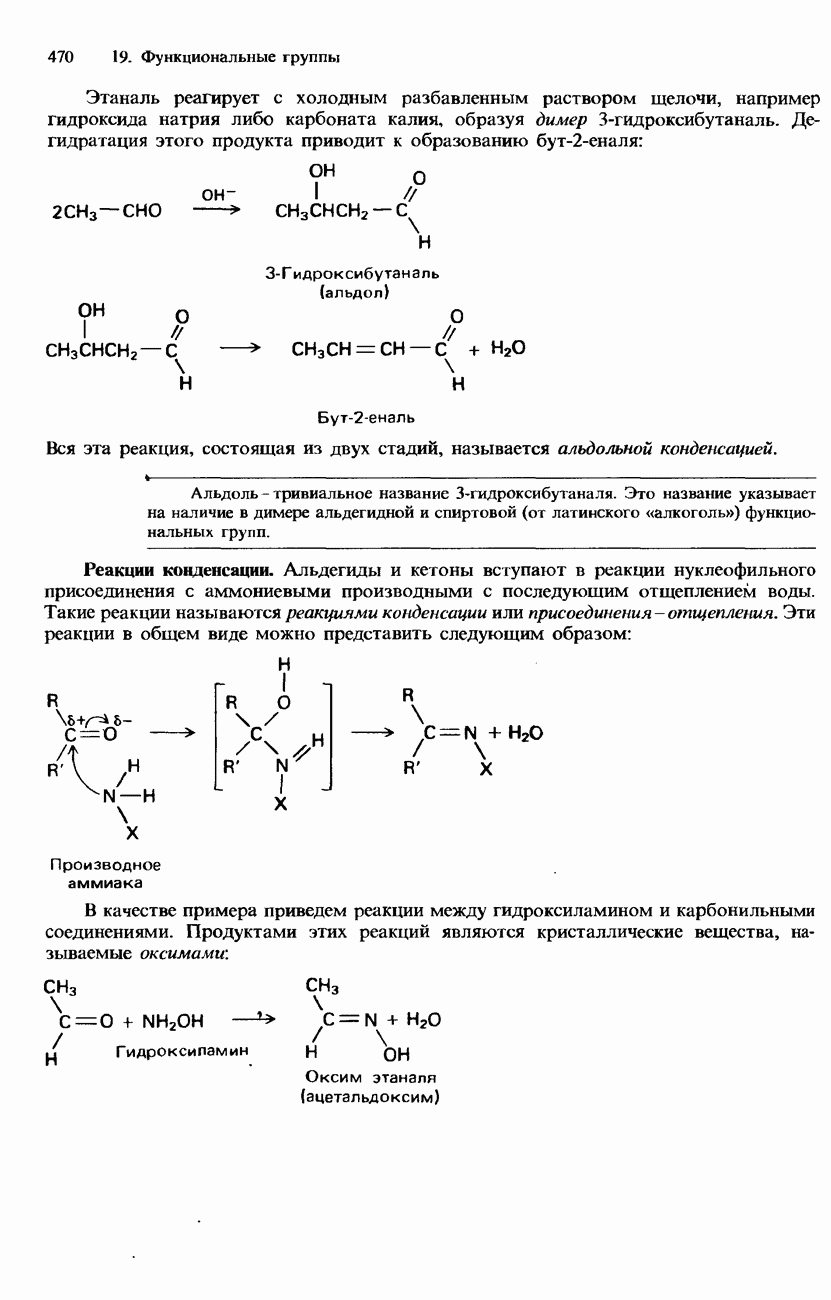
Рис. 20.3. а-Спираль. В фибриллярных белках образование химических связей такого типа может приводить к возникновению складчатой листовой структуры (см. рис. 20.5, а). Структура подобного типа возникает также в результате образования межмолекулярных водородных связей между различными цепями фибриллярного белка (см. рис. 20.5,б). Третичная структура. Так называется трехмерное свертывание полипептидной цепи, приводящее к наложению одних участков
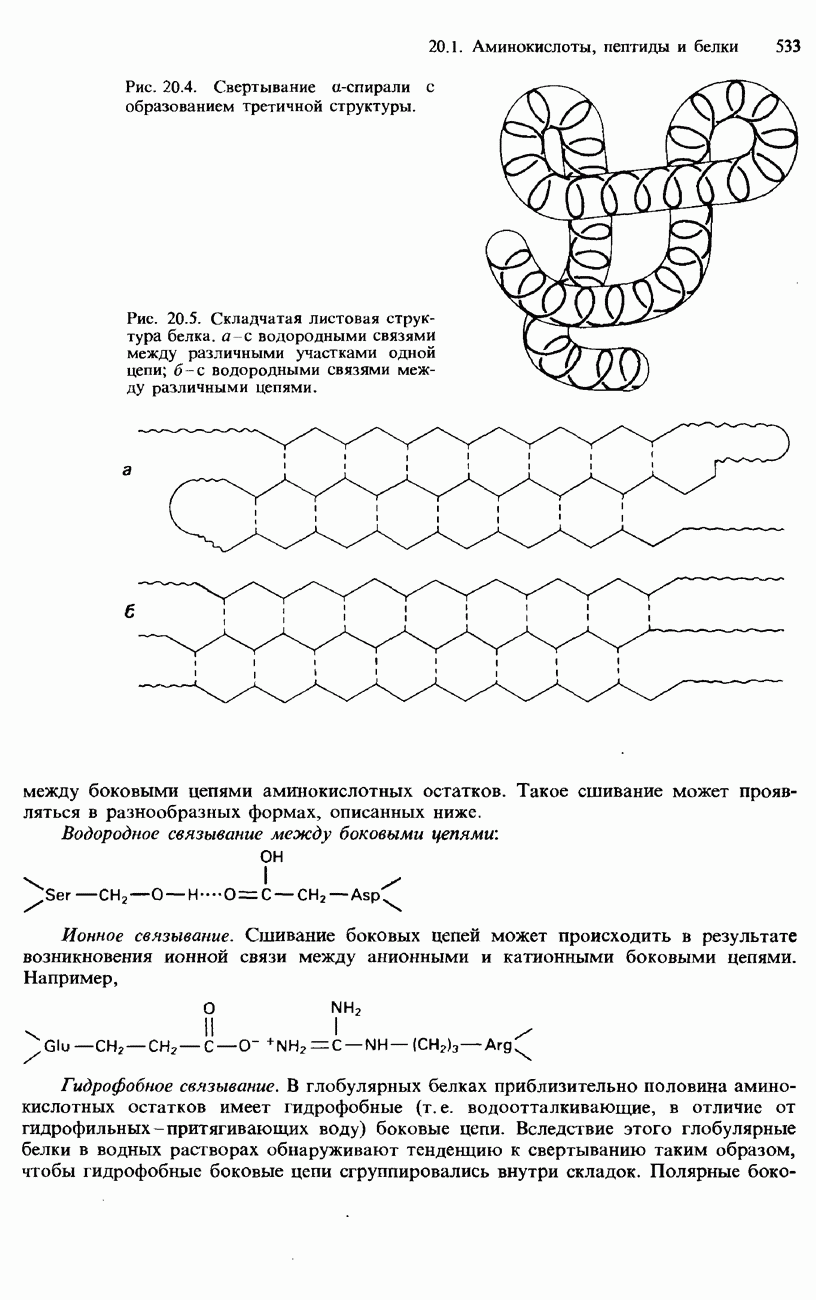
Рис. 20.4. Свертывание
Рис. 20.5. Складчатая листовая структура белка, а с водородными связями между различными участками одной цепи; между боковыми цепями аминокислотных остатков. Такое сшивание может проявляться в разнообразных формах, описанных ниже. Водородное связывание между боковыми цепями:
Ионное связывание. Сшивание боковых цепей может происходить в результате возникновения ионной связи между анионными и катионными боковыми цепями. Например,
Гидрофобное связывание. В глобулярных белках приблизительно половина аминокислотных остатков имеет гидрофобные (т. е. водоотталкивающие, в отличие от гидрофильных - притягивающих воду) боковые цепи. Вследствие этого глобулярные белки в водных растворах обнаруживают тенденцию к свертыванию таким образом, чтобы гидрофобные боковые цепи сгруппировались внутри складок. Полярные боковые цепи, которые, разумеется, являются гидрофильными, оказываются, наоборот, на внешней поверхности молекулы. В результате происходит выталкивание воды из внутренней области молекулы белка. Ковалентное связывание. Наиболее распространенной формой сшивания между цепями является образование дисульфидной связи между атомами серы двух цистеиновых остатков. Например, бычий инсулин состоит из двух полипептидных цепей, связанных между собой дисульфидными мостиками (см. рис. 20.1).
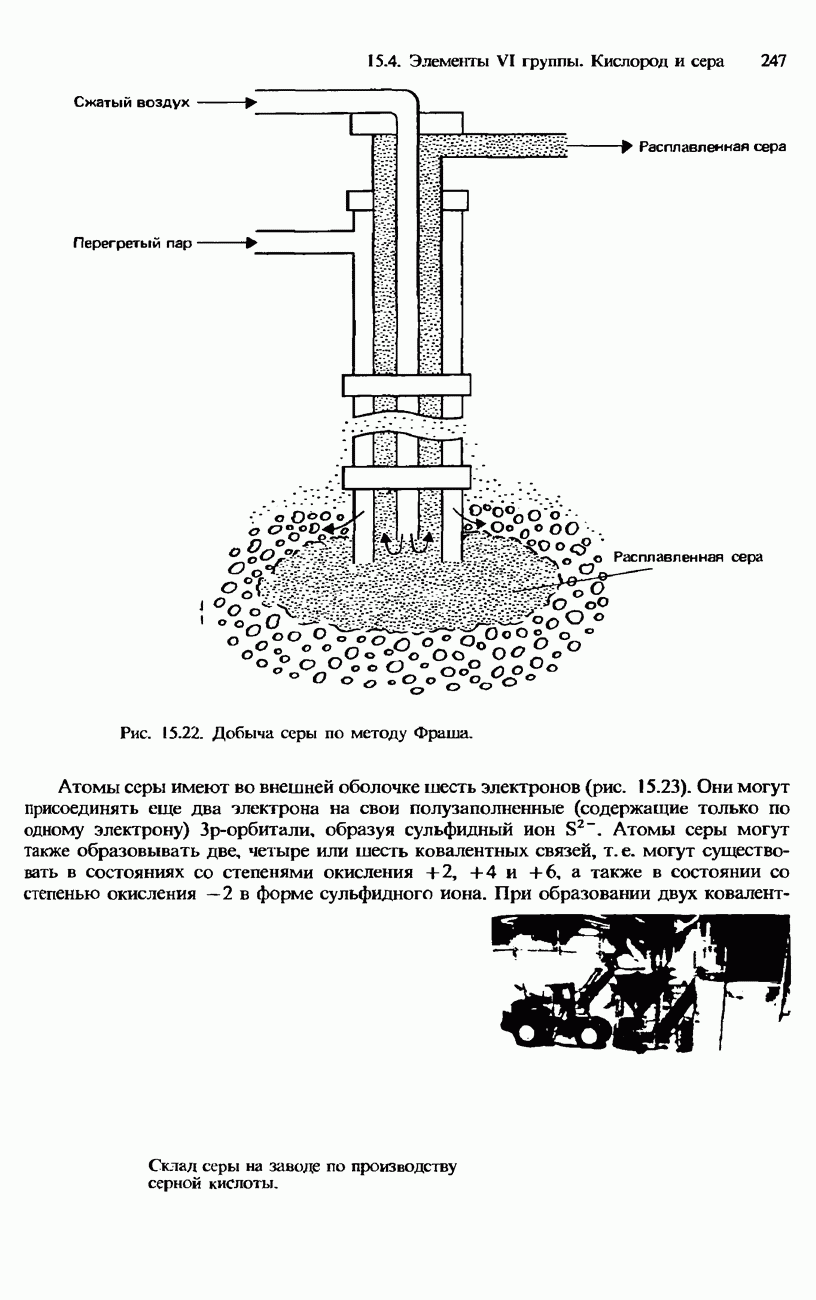
Цистеин и цистин. Цистеин это аминокислота. Цистин состоит из двух цистеиновых фрагментов, связанных между собой дисульфидным мостиком. Ковалентное связывание обнаруживается также в коллагене. Существует несколько типов коллагена, однако все они состоят из трех полипептидных цепей, скрученных вместе в тройную спираль (рис. 20.6). Ковалентное сшивание обнаруживается также между лизиновыми цепями как внутри такой тройной спирали, так и между соседними тройными спиралями. Четвертичная структура. Некоторые белки, например гемоглобин, обладают еще четвертичной структурой. Четвертичная структура образуется несколькими субъединицами
Рис. 20.6. Скручивание трех полипептидных цепей коллагена в тройную спираль. Коллаген составляет до 25% всего белкового вещества в организмах млекопитающих. Он содержится в подкожном слое, зубных и костных тканях, сухожилиях и хрящах.
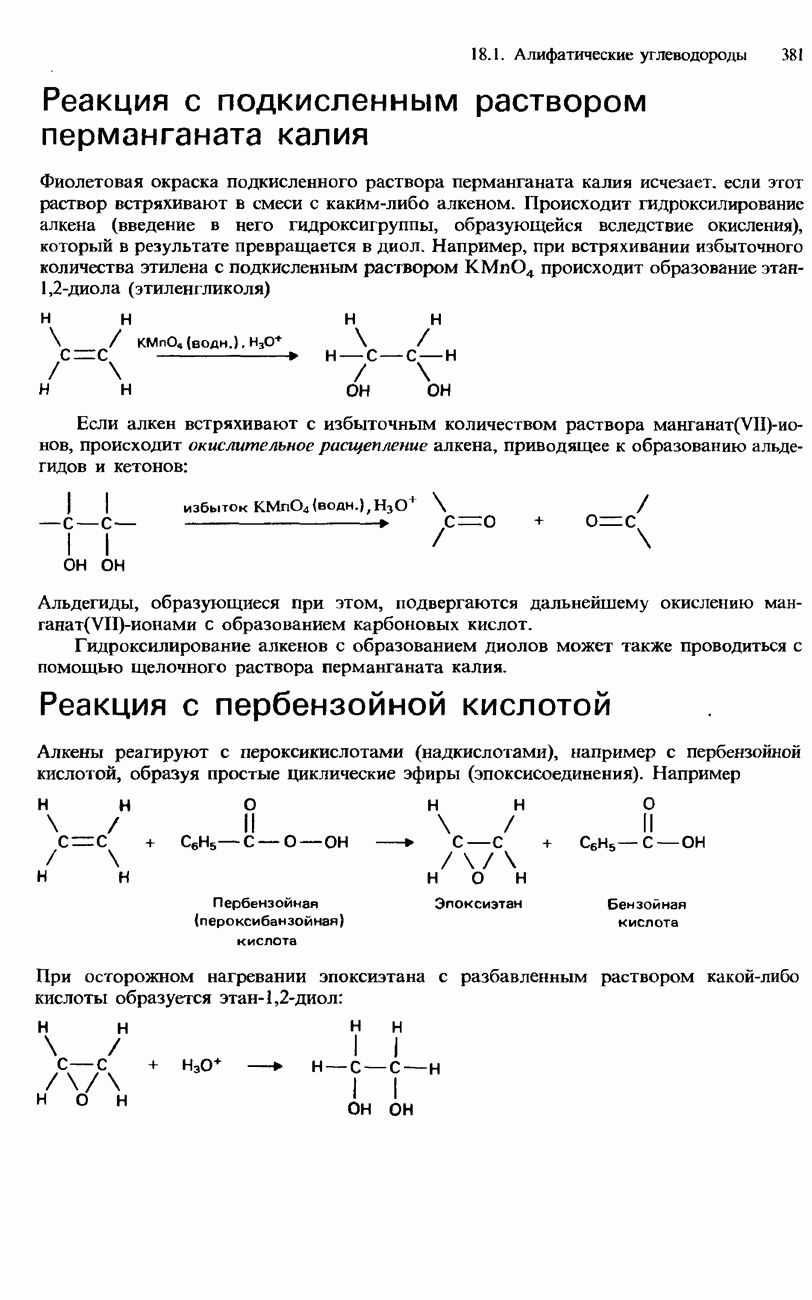
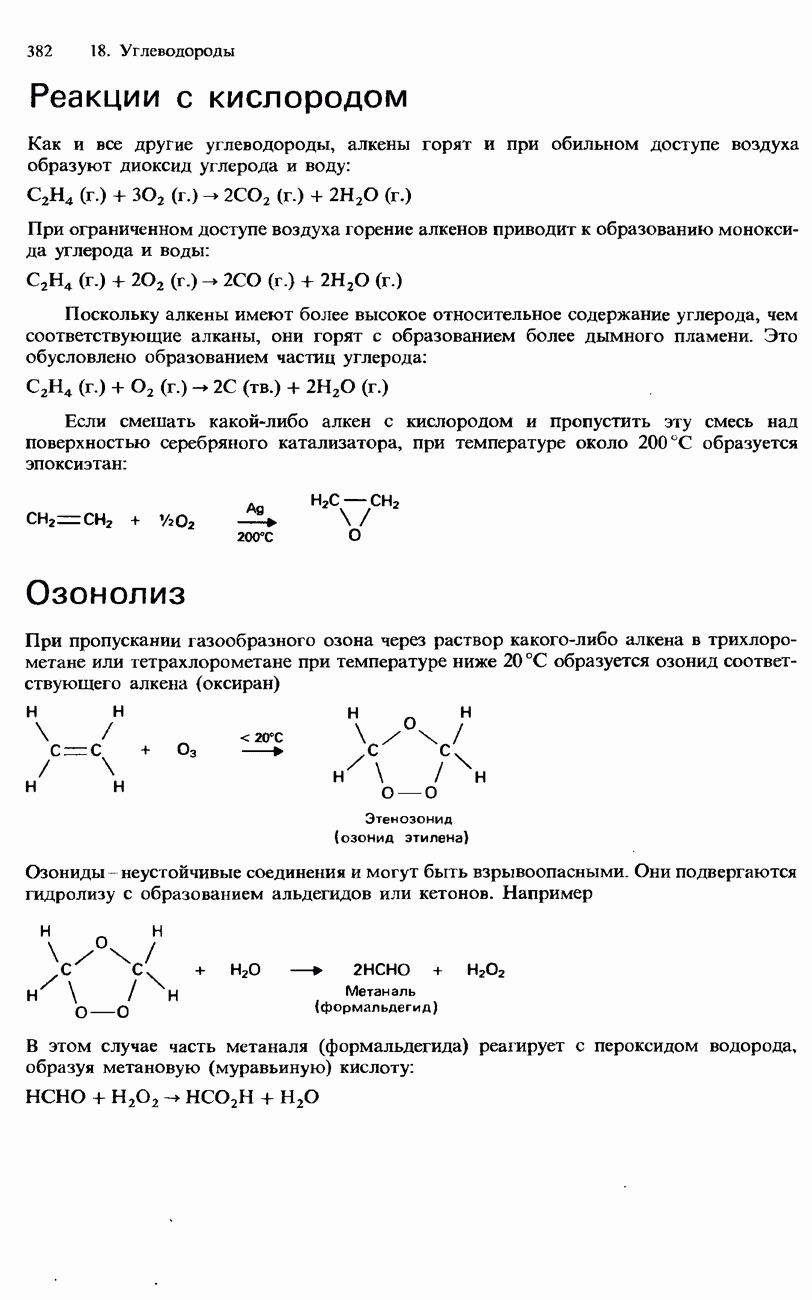
Рис. 20.7. Четвертичная структура белка. Одинаковые или сходные субъединицы соединяются в единое целое с симметричным пространственным расположением. белка. Каждая субъединица имеет отдельную полипептидную цепь. В некоторых белках такие субъединицы одинаковы или имеют сходное строение, а другие белки состоят из субъединиц с цепями разных типов. Эти субъединицы образуют друг с другом единое целое, с симметричным пространственным расположением (рис. 20.7). Четвертичная структура удерживается как единое целое благодаря нековалентным взаимодействиям между боковыми цепями, как в третичной структуре. Синтез белковСинтез пептидов из аминокислот позволил получить за последние десятилетия много сведений о структуре белков. В настоящее время достигнута возможность синтеза пептидов, обладающих биологической активностью. Синтез пептидов приобрел огромное значение в генной инженерии и биотехнологии (см. предисловие к данной главе). Как было указано выше, аминокислоты имеют две разные функциональные группы - карбоксигруппу и аминогруппу. Они называются соответственно С-концевой и N-концевой группами. Чтобы синтезировать дипептид из двух аминокислот, необходимо «заблокировать» С-концевую группу одной аминокислоты и N-концевую группу другой аминокислоты с помощью каких-либо защитных групп. Такие защитные группы предотвращают протекание реакции на рассматриваемых концевых группах. В результате происходит образование пептидной связи между двумя аминокислотами, после чего осуществляют деблокирование концевых групп. Таким образом, синтез пептидов включает три стадии. 1. Блокирование
Здесь X и Y - защитные группы. 2. Образование пептида
3. Деблокирование
Этот процесс может повторяться со следующими аминокислотами, в результате чего осуществляется синтез трипептидов и более сложных пептидов:
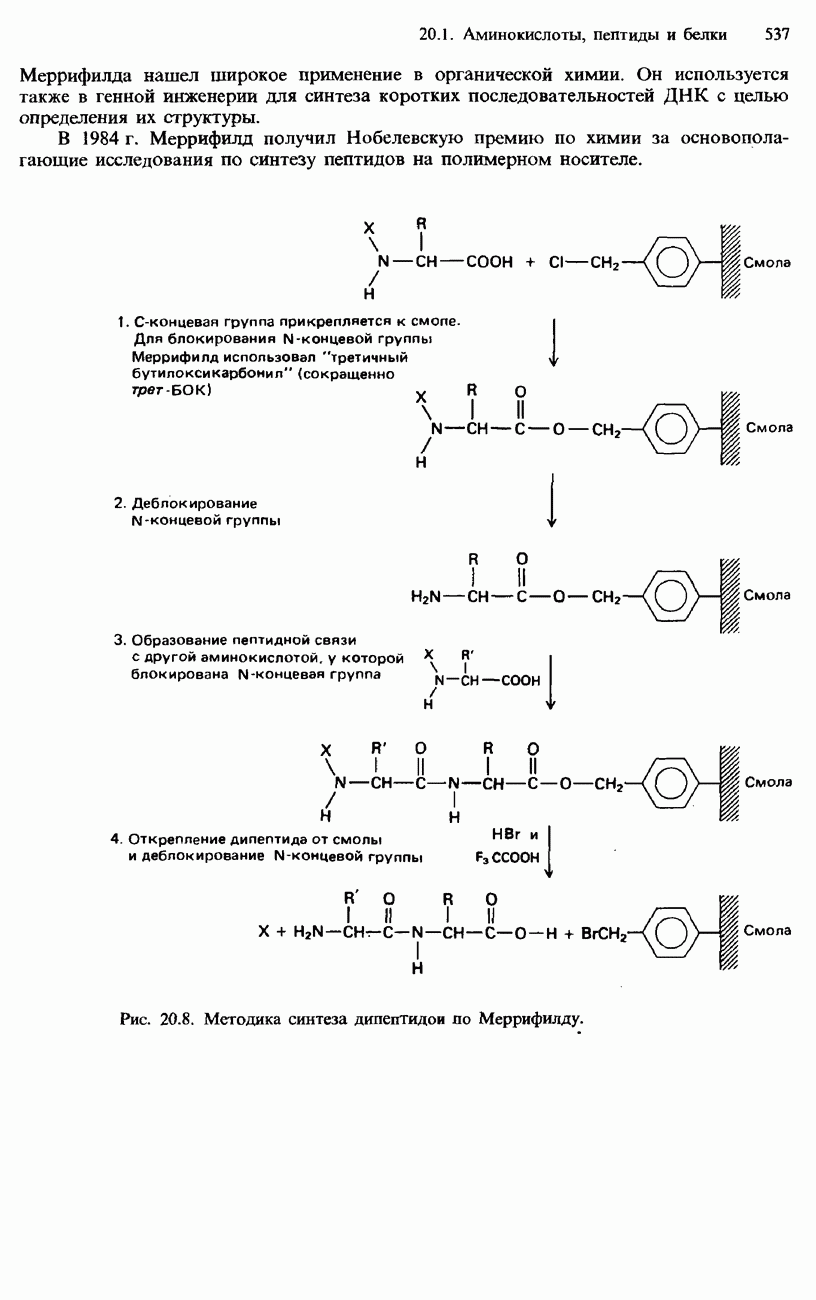
Для блокирования С-концевых и N-концевых групп подобрано много разных защитных групп, а для их последующего удаления разработаны различные методы. До конца 1950-х и начала 1960-х гг. синтез пептидов представлял собой медленный и очень трудоемкий процесс. Бурный прогресс в этой области начался с исследований американского химика Брюса Меррифилда. В 1963 г. он опубликовал метод твердофазного синтеза пептида, который приобрел известность как метод Меррифилда. Этот метод основан на присоединении блокируемой аминогруппы к носителю из синтетической смолы - хлорометилполистирола. Такая смола представляет собой нерастворимое желеобразное вещество. Методика синтеза дипептидов по Меррифилду состоит из нескольких стадий, изображенных на рис. 20.8. С помощью своей методики Меррифилд в 1964 г. синтезировал гормон брадикинин, который содержит девять аминокислотных остатков. С этого времени метод
Брюс Меррифилд, лауреат Нобелевской премии по химии 1984 г. Меррифилда нашел широкое применение в органической химии. Он используется также в генной инженерии для синтеза коротких последовательностей ДНК с целью определения их структуры. В 1984 г. Меррифилд получил Нобелевскую премию по химии за основополагающие исследования по синтезу пептидов на полимерном носителе.
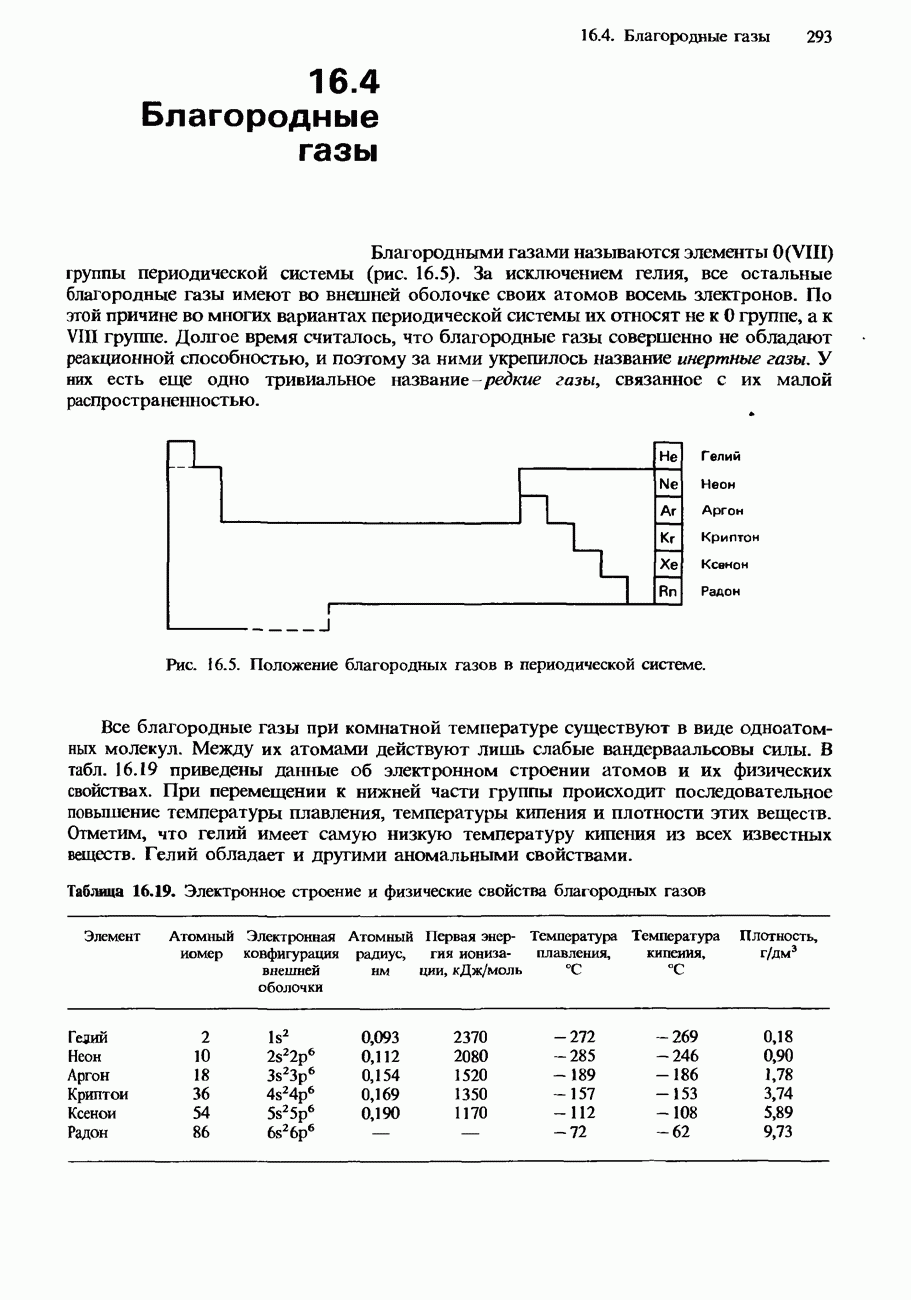
Рис. 20.8. Методика синтеза дипептидои по Меррифилду. 1. Белки состоят из аминокислот, соединенных друг с другом пептидными связями. 2. Аминокислоты обнаруживают оптическую изомерию. 3. Незаменимые аминокислоты не могут синтезироваться в человеческом организме. Они обязательно должны входить в нашу диету. 4. Цвиттер-ион содержит в одной молекуле отрицательный заряд и положительный заряд. 5. Полипептиды содержат несколько связанных между собой аминокислот. 6. Глобулярные белки растворимы в воде либо образуют в ней коллоидные растворы. 7. Фибриллярные белки нерастворимы в воде. 8. Для разделения белков используются фракционное осаждение, колоночная хроматография, ионообменная хроматография и электрофорез. 9. Для определения относительной молекулярной массы белков используется ультрацентрифугирование. 10. Последовательность аминокислот в полипептидной цепи белка называется его первичной структурой. 11. Упорядоченное свертывание белка или полипептидной цепи называется вторичной структурой. Она возникает в результате образования водородных связей между карбоксильными группами и аминогруппами. Примером вторичной структуры является винтовая 12. Третичная структура белка обусловлена сшиванием между боковыми цепями аминокислотных остатков. 13. Четвертичная структура белка состоит из нескольких субъединиц, каждая из которых содержит отдельную полипептидную цепь. 14. Для синтеза пептидов широко используется метод Меррифилда. При этом N-концевая группа аминокислоты блокируется защитной группой, а С-концевая группа аминокислоты связывается с носителем из полимерной смолы. Затем производится деблокирование N-концевой группы и образуется пептидная связь.
|
 |
Оглавление
|




































































































































































































































































































































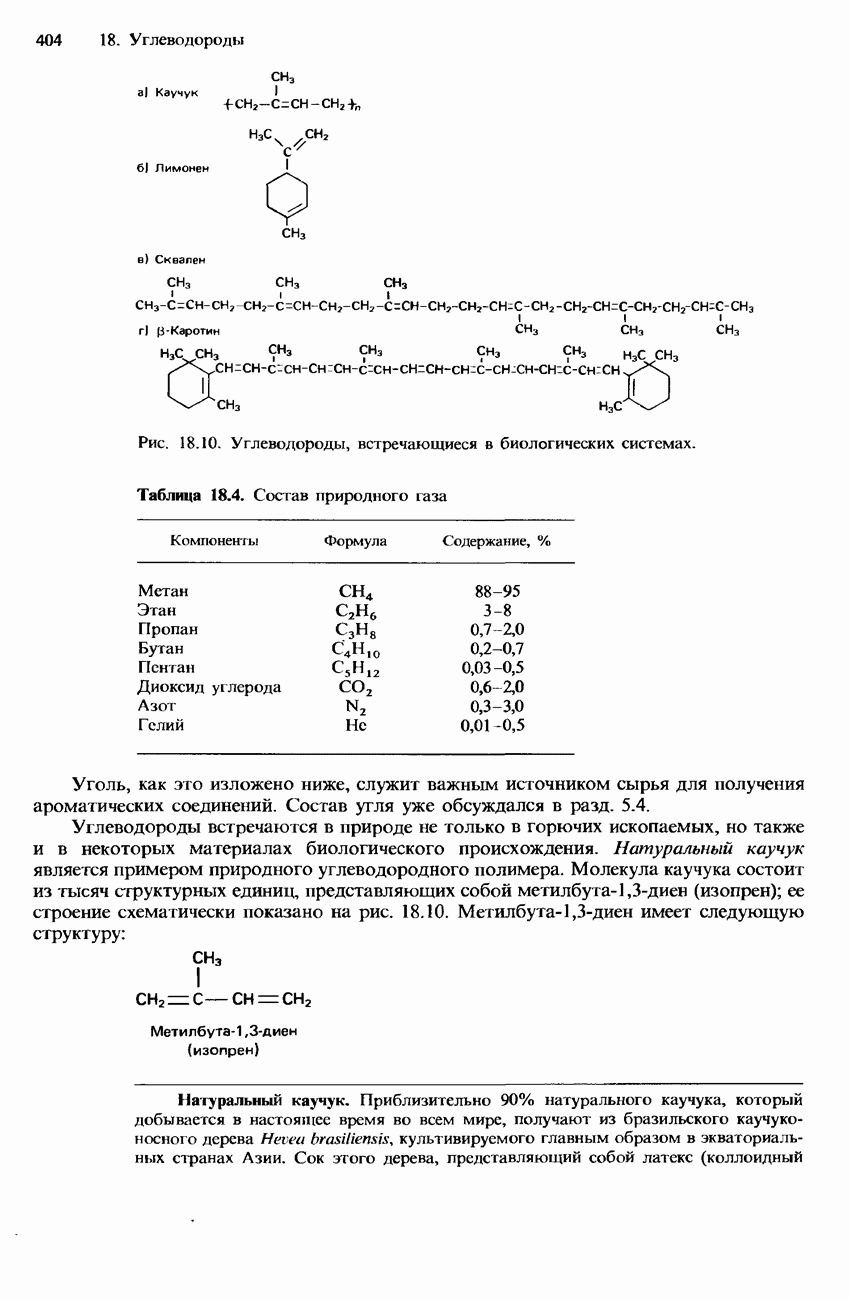








































































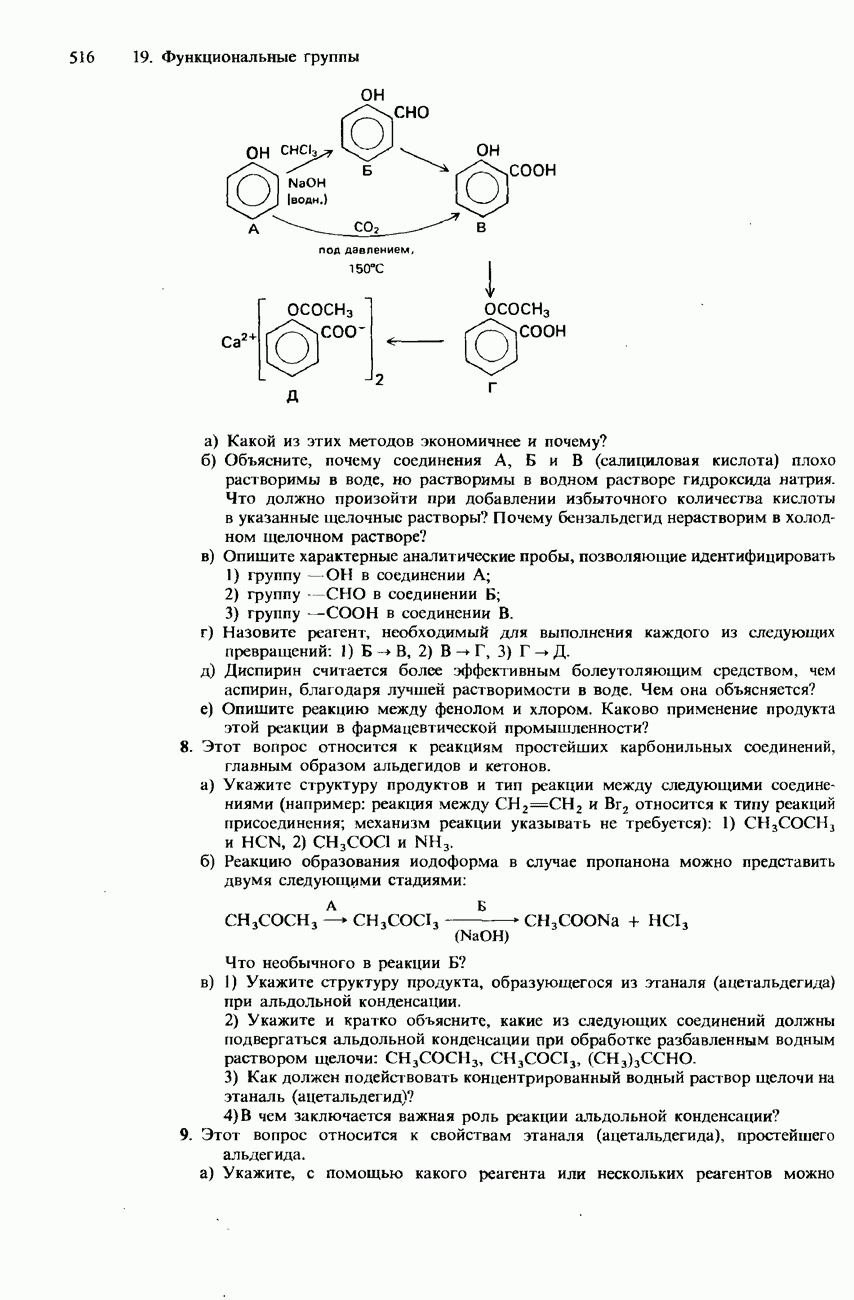





















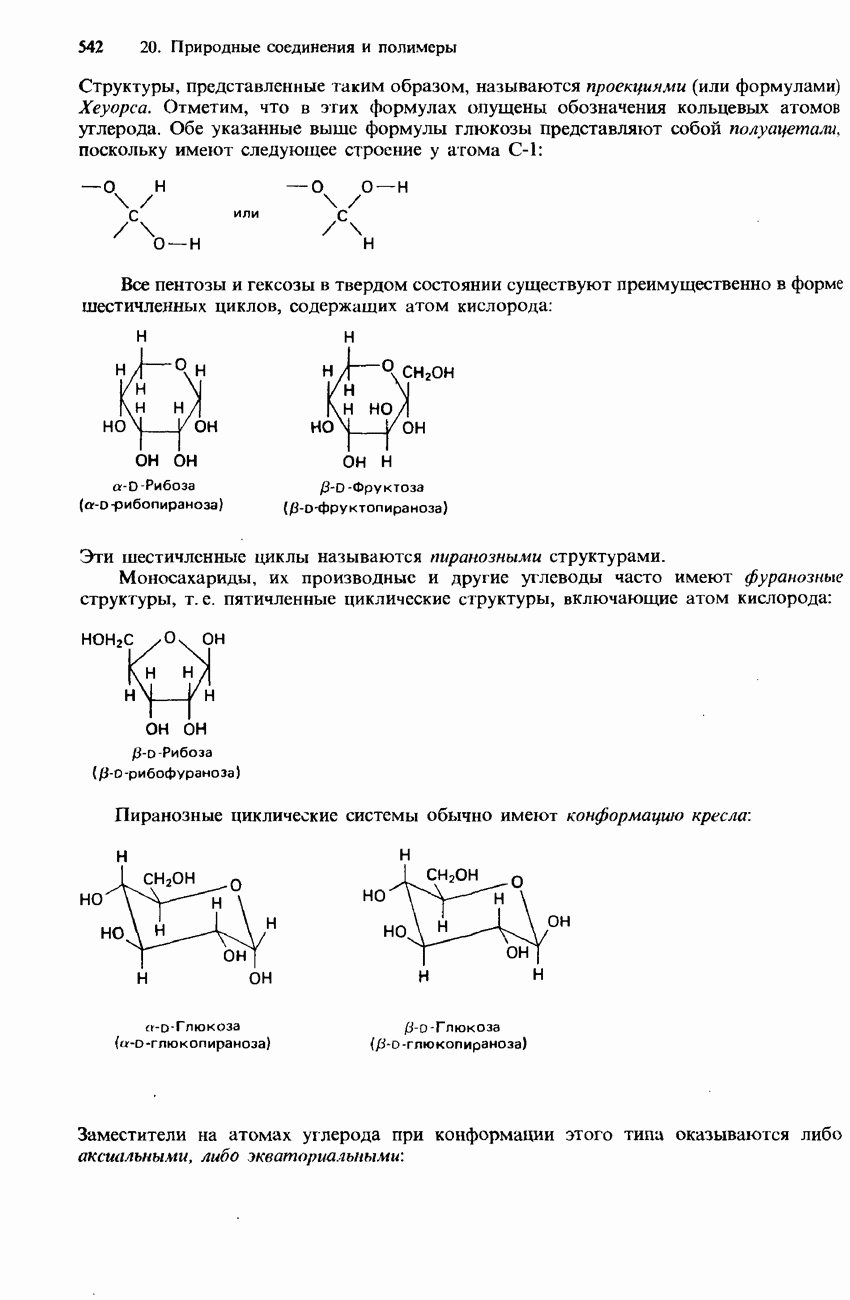
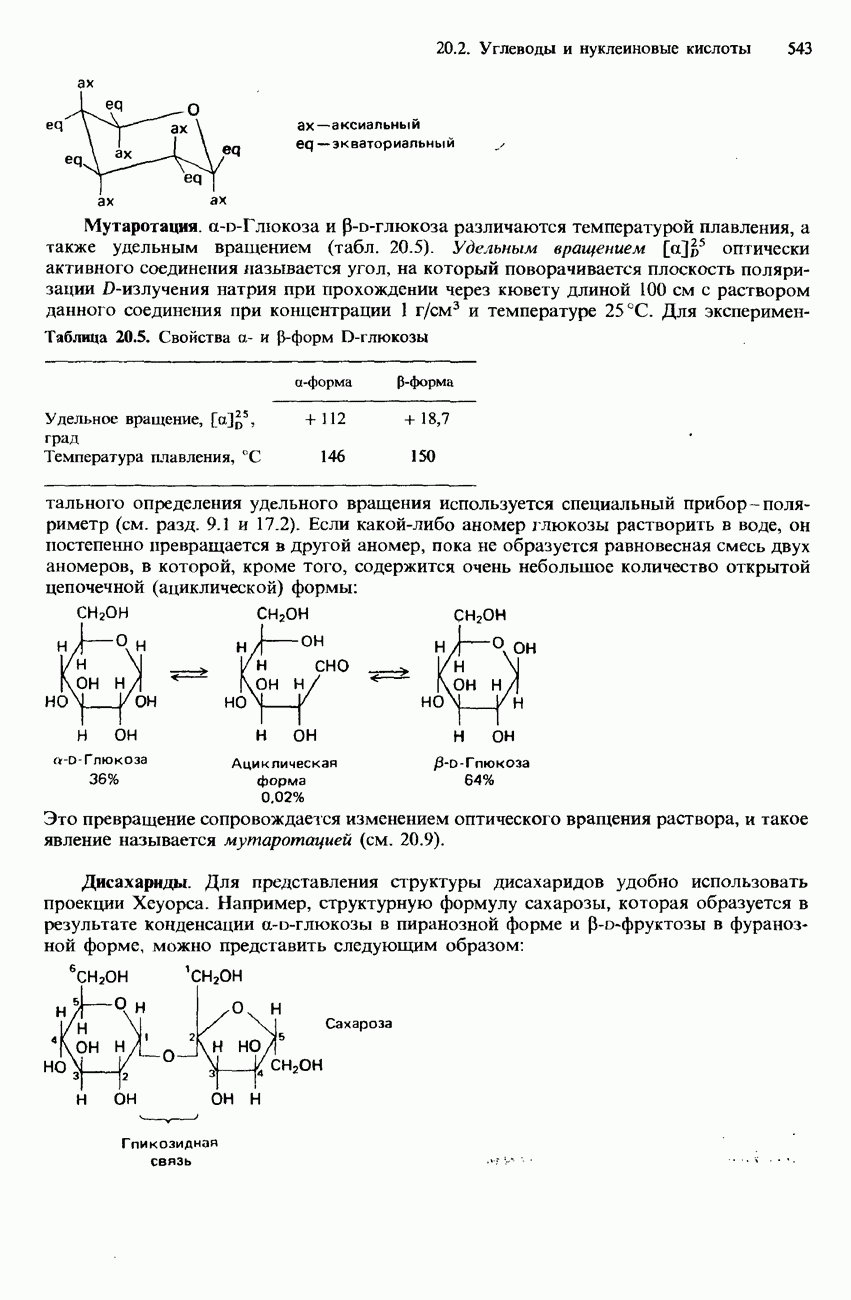
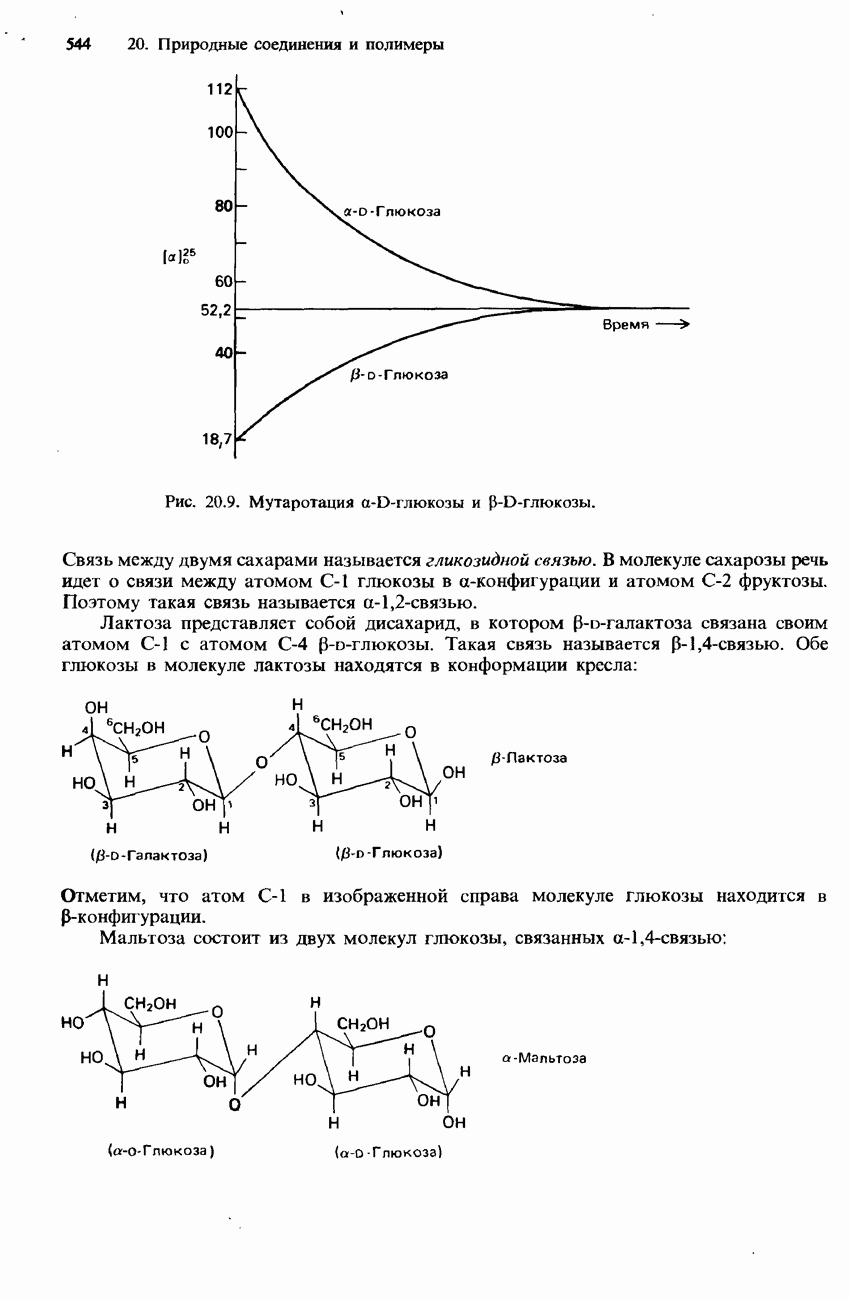





































 -аминокислоты, называются простыми белками. Такие белки принято подразделять по их растворимости в воде на глобулярные и фибриллярные.
-аминокислоты, называются простыми белками. Такие белки принято подразделять по их растворимости в воде на глобулярные и фибриллярные.

 средах с предельными значениями pH. Это явление называется денатурацией. В процессе денатурации происходит изменение упорядоченной свернутой структуры белка (см. ниже) на неупорядоченную, хаотическую структуру. Денатурация белка происходит, например, при варке яиц. Хотя денатурация обычно представляет собой необратимый процесс, он не сопровождается разрушением полипептидной цепи.
средах с предельными значениями pH. Это явление называется денатурацией. В процессе денатурации происходит изменение упорядоченной свернутой структуры белка (см. ниже) на неупорядоченную, хаотическую структуру. Денатурация белка происходит, например, при варке яиц. Хотя денатурация обычно представляет собой необратимый процесс, он не сопровождается разрушением полипептидной цепи.
 водного раствора белков, можно тоже добиваться селективного осаждения одного или нескольких из них.
водного раствора белков, можно тоже добиваться селективного осаждения одного или нескольких из них.

 аминокислот при вхождении их в пептидную либо белковую цепь могут изменяться. Дело в том, что на диссоциацию аминокислоты, находящейся в цепи, могут оказывать влияние соседние заряженные группы и другие факторы, обусловленные ее ближайшим окружением.
аминокислот при вхождении их в пептидную либо белковую цепь могут изменяться. Дело в том, что на диссоциацию аминокислоты, находящейся в цепи, могут оказывать влияние соседние заряженные группы и другие факторы, обусловленные ее ближайшим окружением.


 -спиралью (рис. 20.3 и 20.4).
-спиралью (рис. 20.3 и 20.4).


 -спирали или складчатого листа на другие участки цепи. Пример наложения одних участков
-спирали или складчатого листа на другие участки цепи. Пример наложения одних участков  -спирали на другие ее участки показан на рис. 20.4. Возникновение третичной структуры обусловлено сшиванием между различными участками полипептидной цепи. Это сшивание происходит в результате образования химической связи и установления различных взаимодействий
-спирали на другие ее участки показан на рис. 20.4. Возникновение третичной структуры обусловлено сшиванием между различными участками полипептидной цепи. Это сшивание происходит в результате образования химической связи и установления различных взаимодействий

 -спирали с образованием третичной структуры.
-спирали с образованием третичной структуры.

 водородными связями между различными цепями.
водородными связями между различными цепями.











 -спираль.
-спираль.