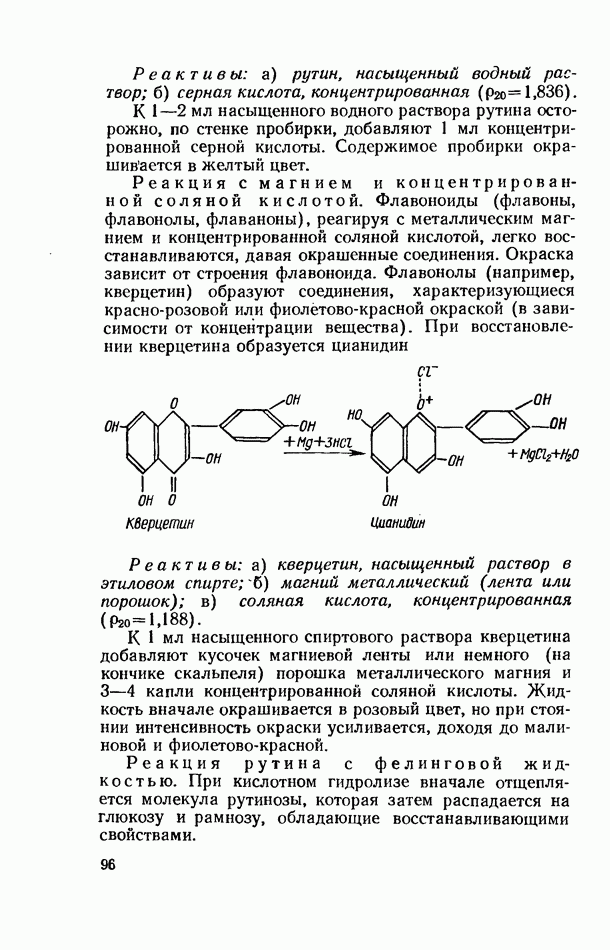
Пред.
След.
Макеты страниц
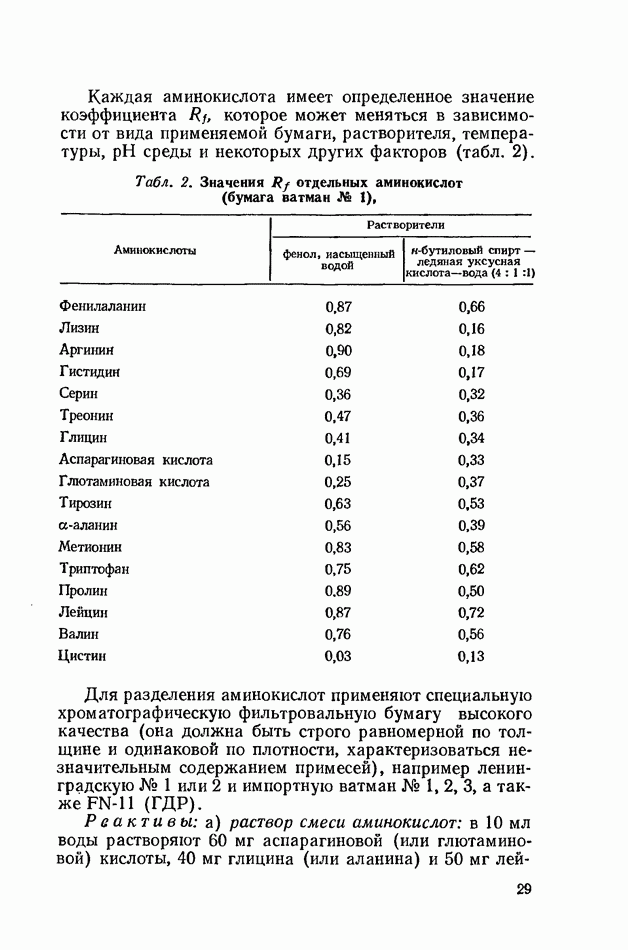
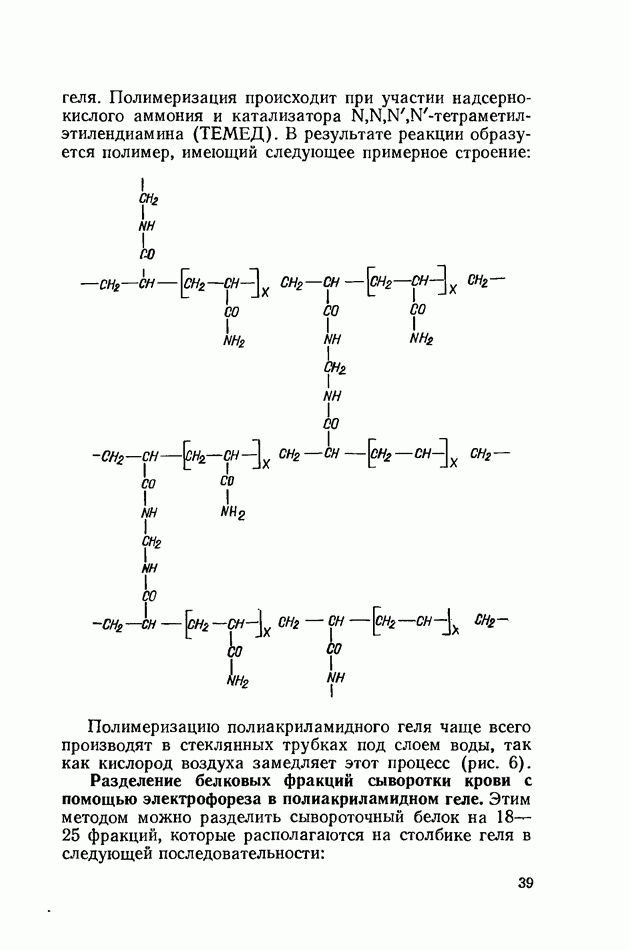
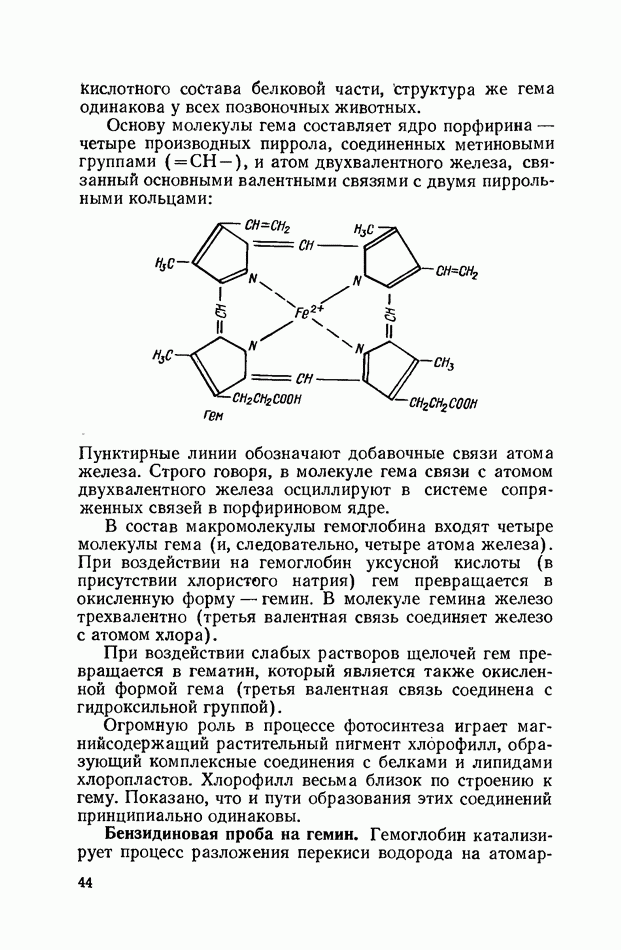
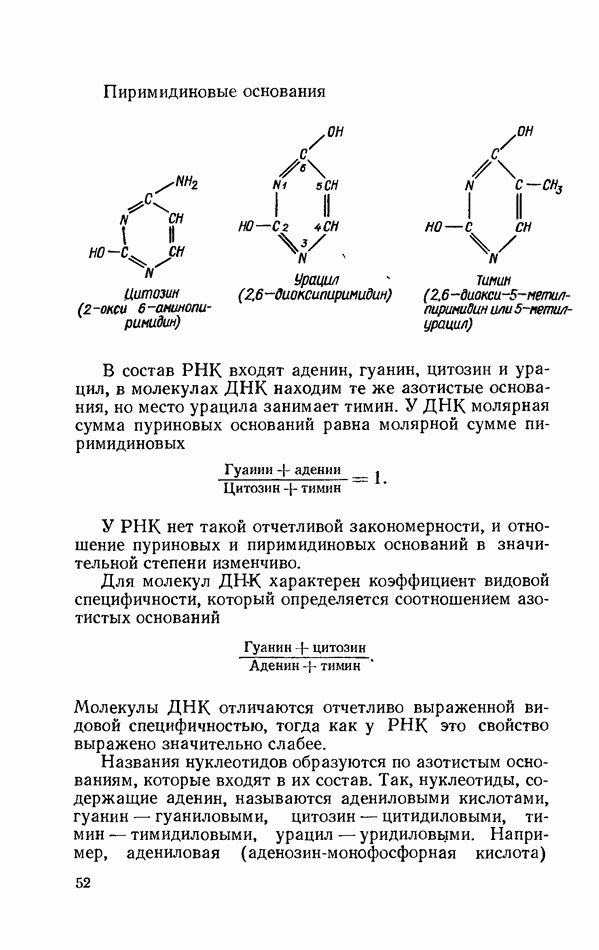
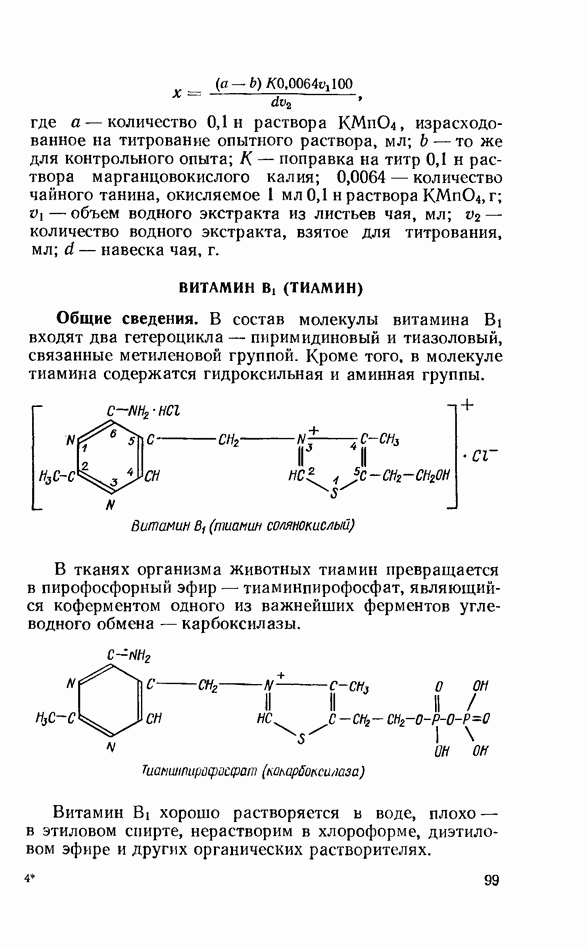
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике ДЛЯ СТУДЕНТОВ И ШКОЛЬНИКОВ ЕСТЬ
ZADANIA.TO
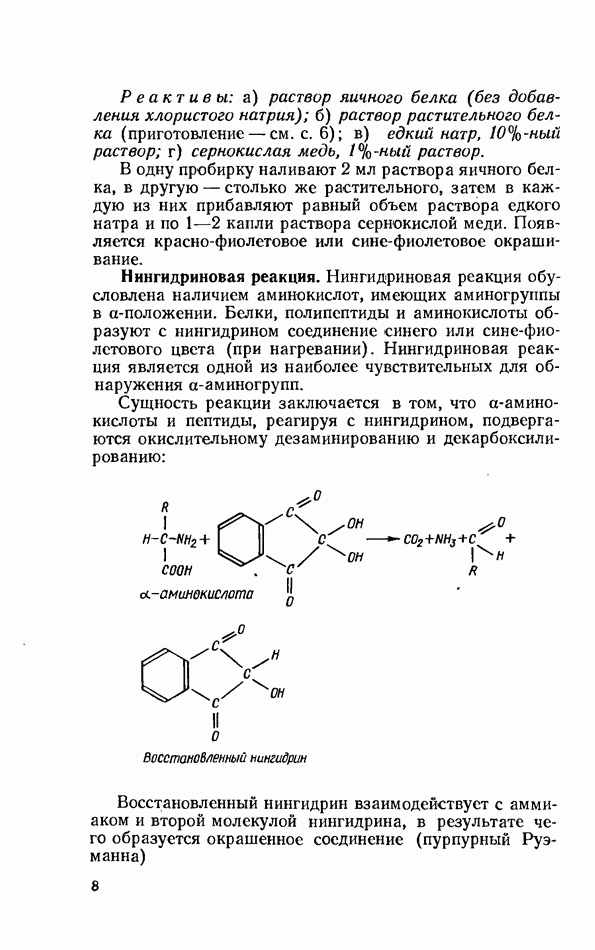
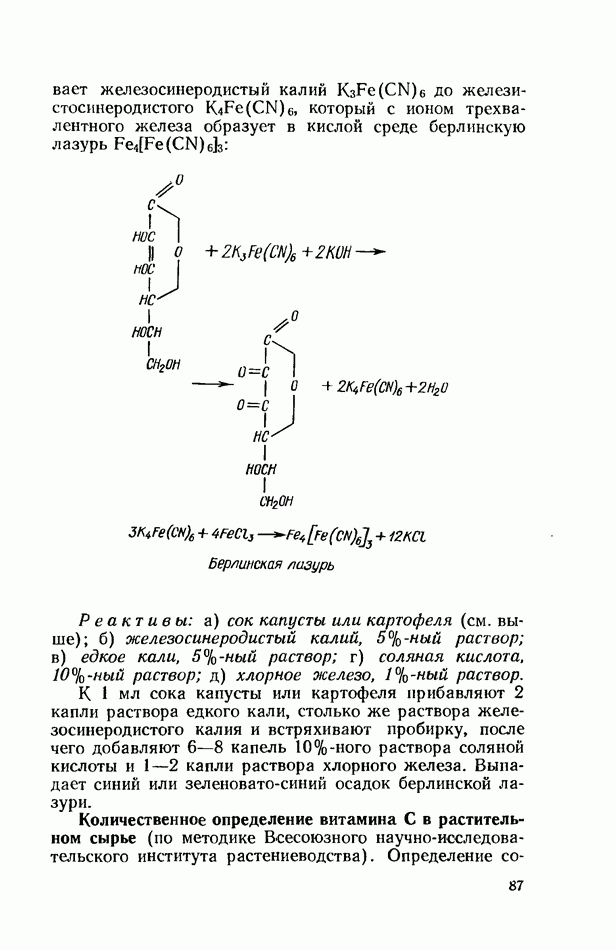
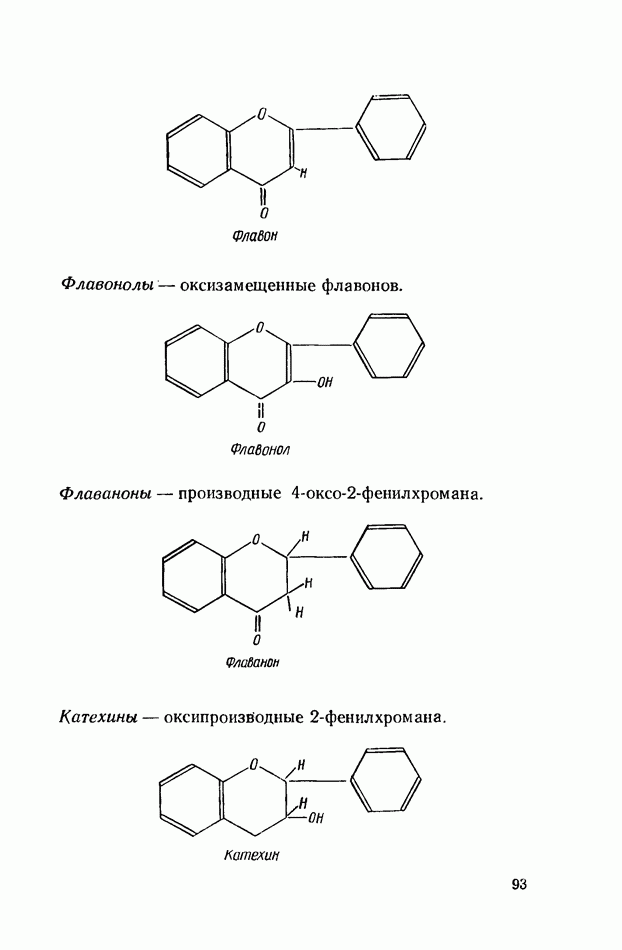
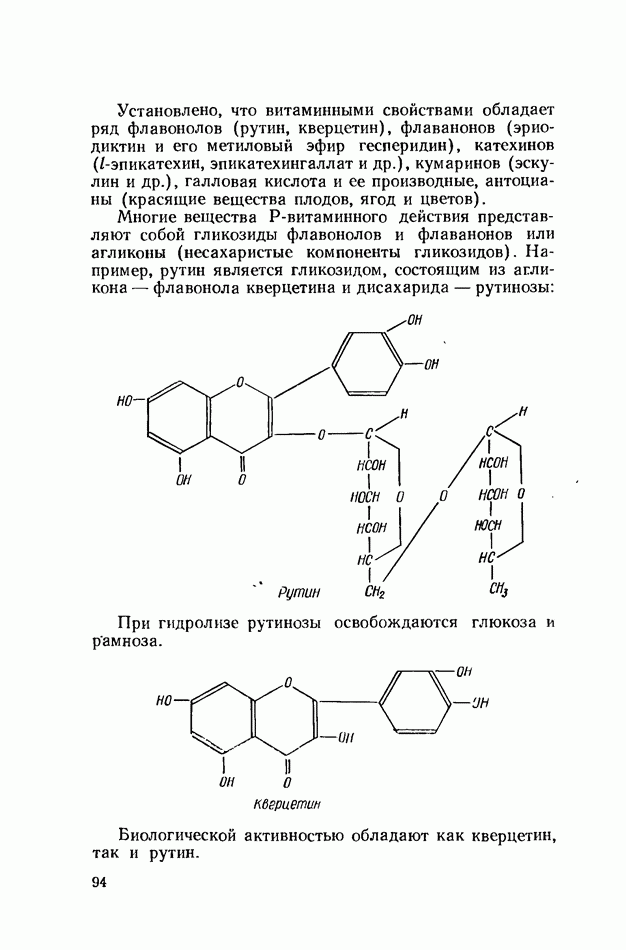
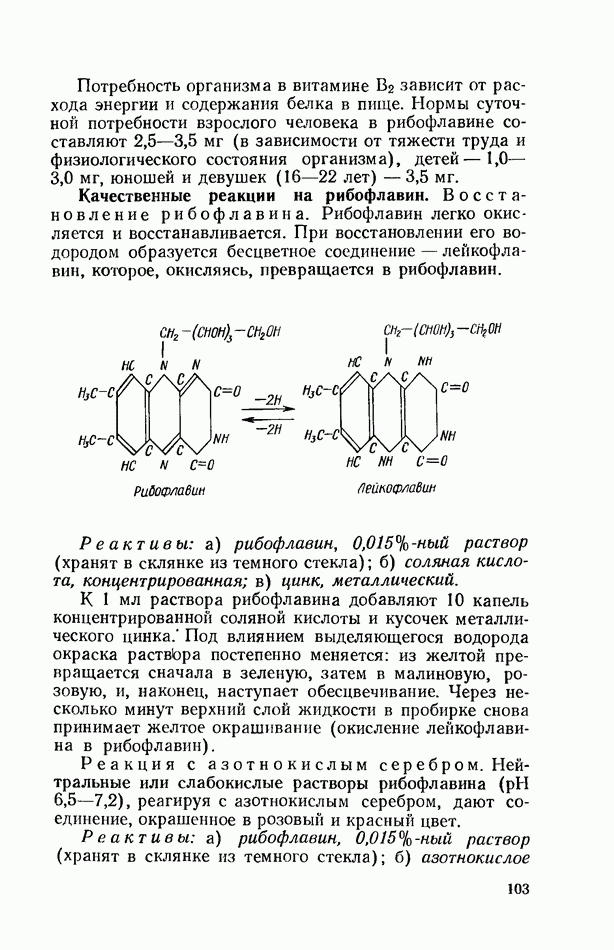
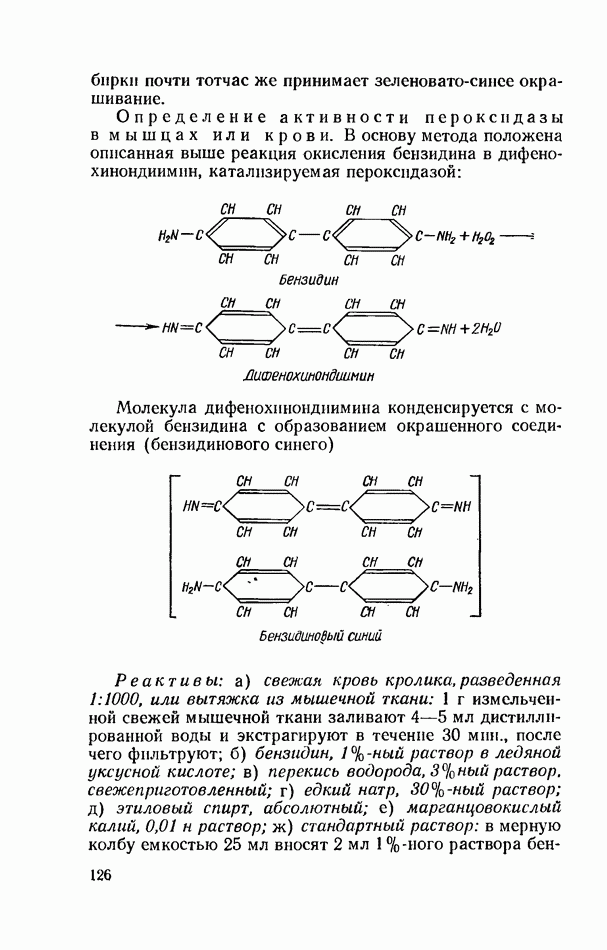
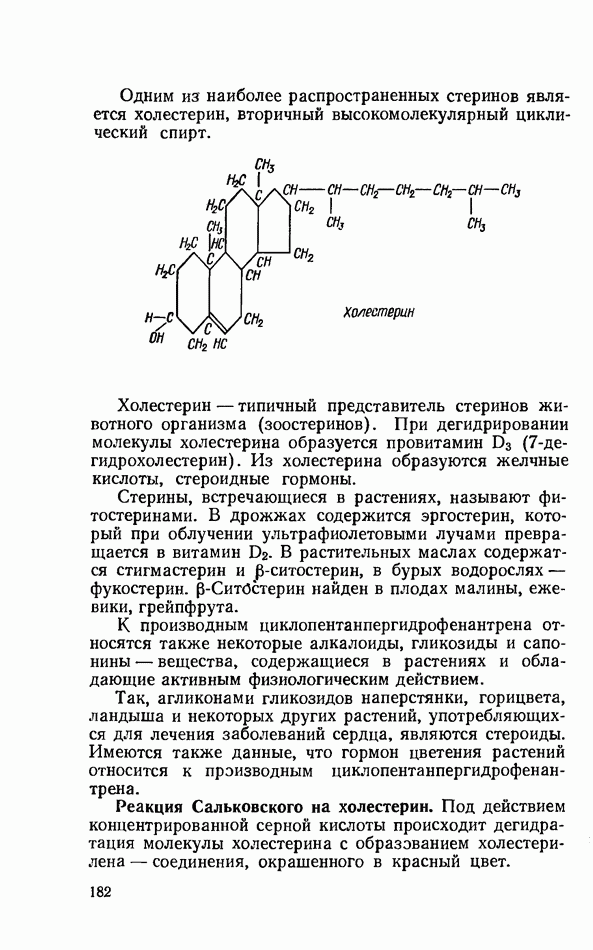
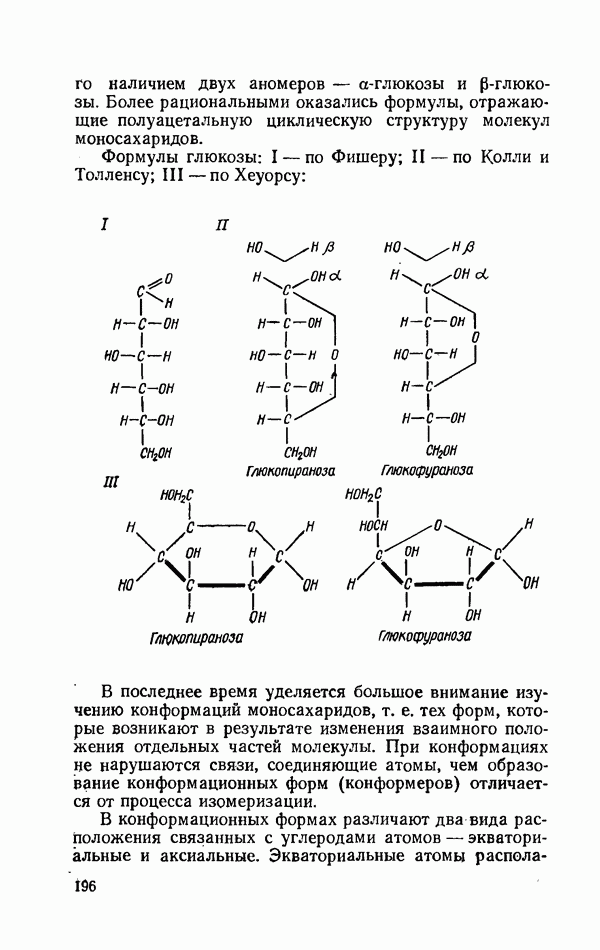
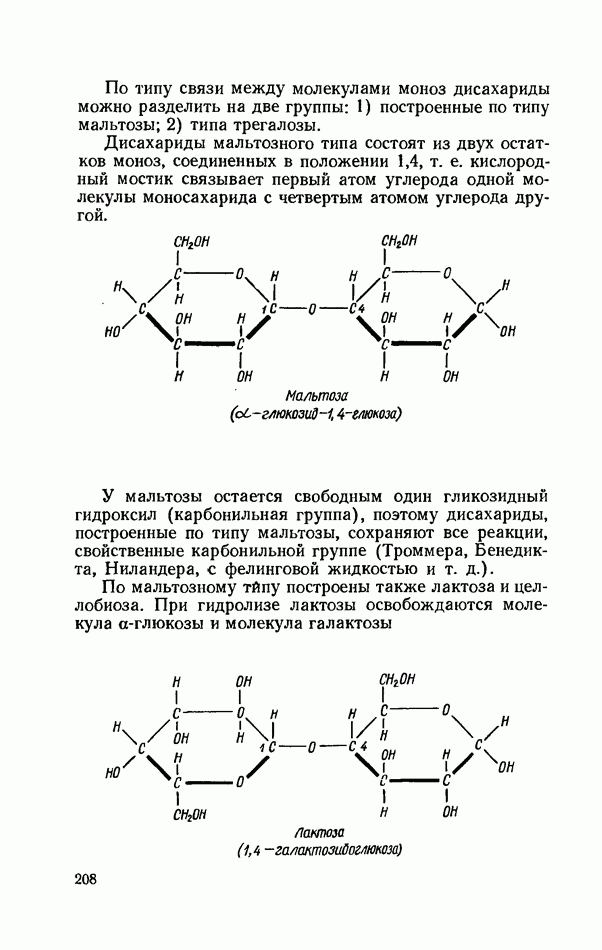
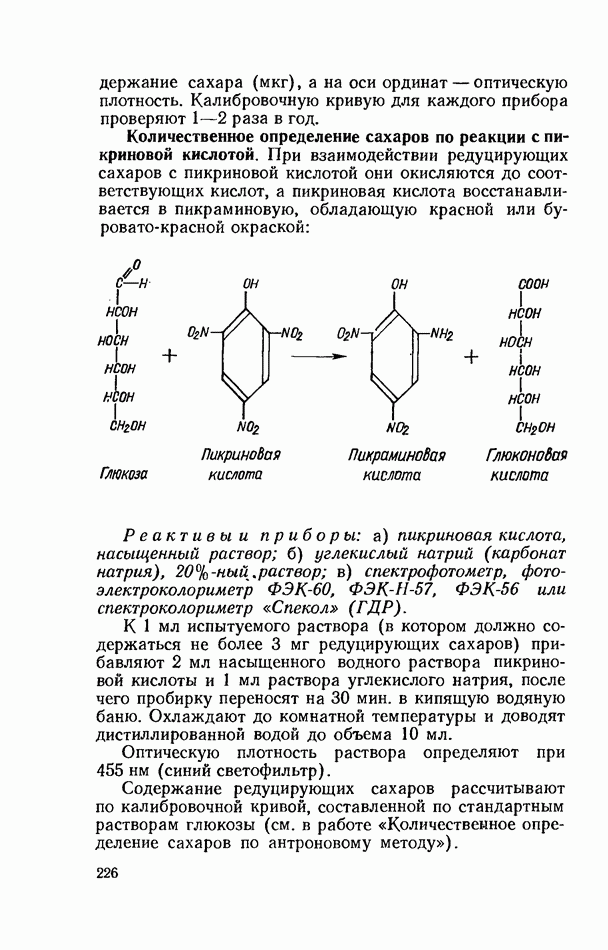
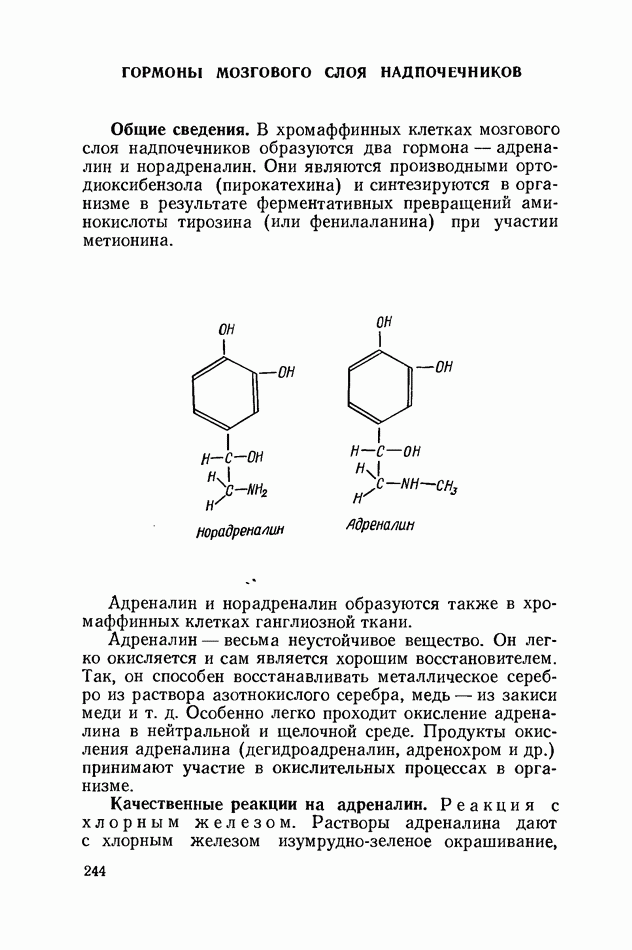
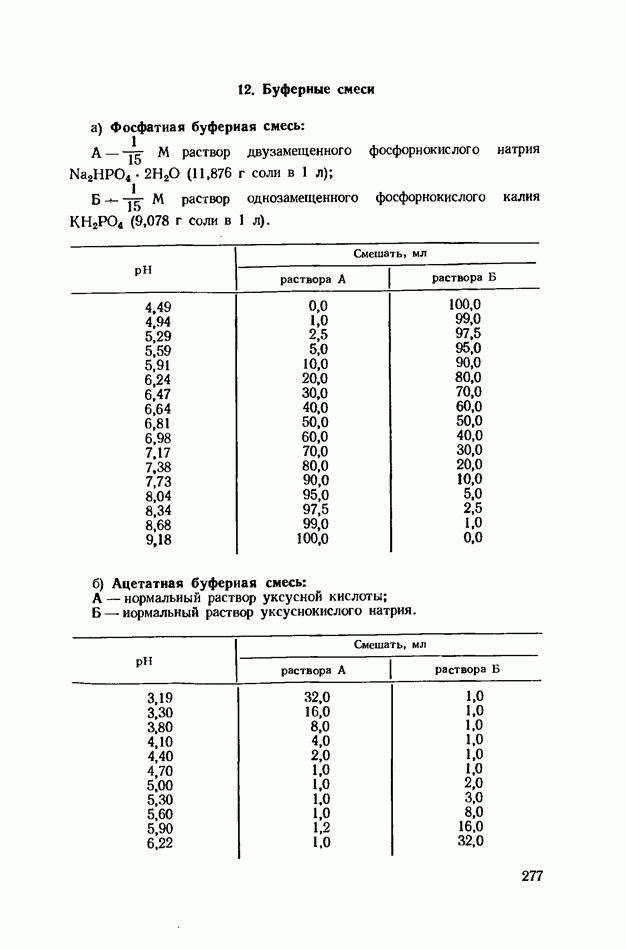
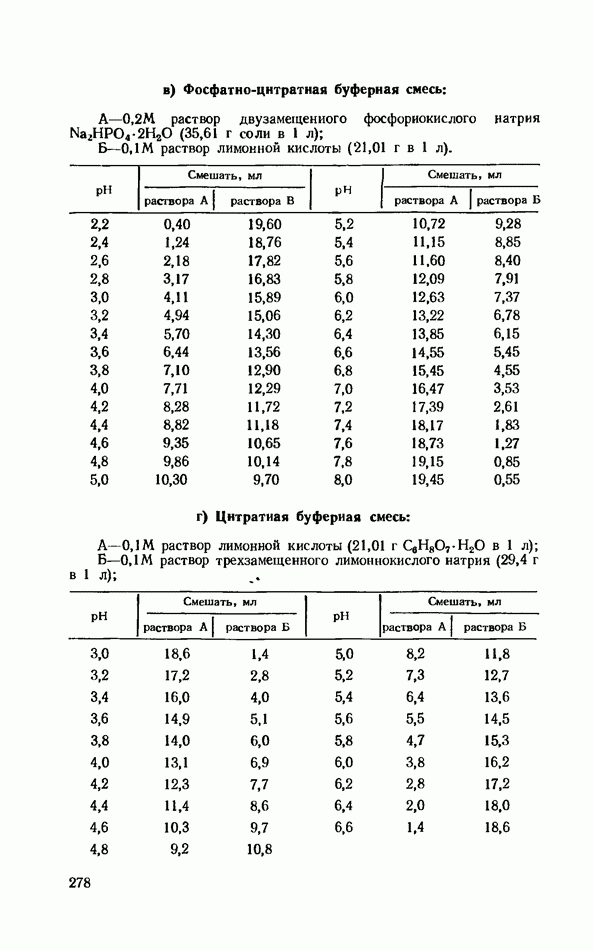
ФЕРМЕНТАТИВНЫЙ ГИДРОЛИЗ ЛИПИДОВОбщие сведения.Жиры и липоиды подвергаются гидролитическому расщеплению в пищеварительном тракте. Нейтральные жиры распадаются на глицерин и жирные кислоты; фосфатиды (лецитины, кефалины, серинфосфатиды и др.) расщепляются на глицерин, жирные кислоты, фосфорную кислоту и азотистые основания — холин, коламин (этаноламин), серин и т. д. При гидролизе стеридов освобождаются холестерин или эргостерин и жирные кислоты. Гидролитический распад жиров катализируется ферментами липазами, которые содержатся в соке желудка, поджелудочной железы и тонкого кишечника. Роль желудочной липазы у взрослого человека весьма невелика, так как фермент катализирует расщепление лишь тонкодиспергированных, предварительно эмульгированных жиров (например, молочного). Значительная роль в переваривании жиров принадлежит липазе поджелудочной железы. Расщепление жиров происходит главным образом в тонком кишечнике. Липаза поджелудочной железы выделяется в малоактивной форме и активируется желчными кислотами. Значение желчных кислот в переваривании жира очень велико. Они являются не только активаторами липазы. Будучи поверхностно-активными веществами, желчные кислоты способствуют эмульгированию жиров, что увеличивает во много раз их поверхность соприкосновения с водным раствором липазы. Липазы содержатся также в растительных объектах (семенах злаков, масличных растений) и микроорганизмах. При их участии происходит порча круп, муки и других продуктов при хранении. Гидролитическое расщепление жиров протекает в несколько стадий. Липаза действует главным образом на внешние (а) эфирные связи молекулы триглицерида. Вначале отщепляются жирные кислоты, связанные с глицерином в
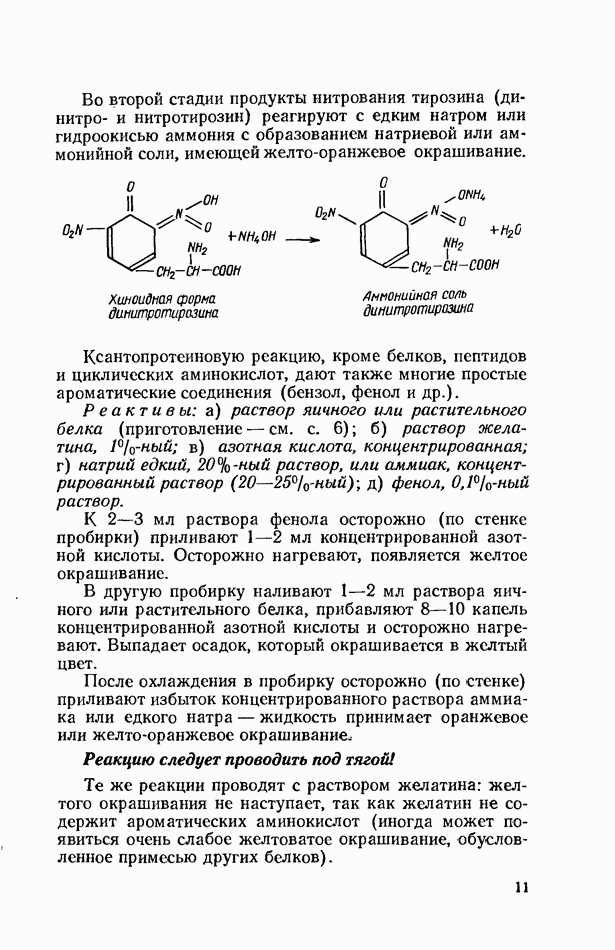
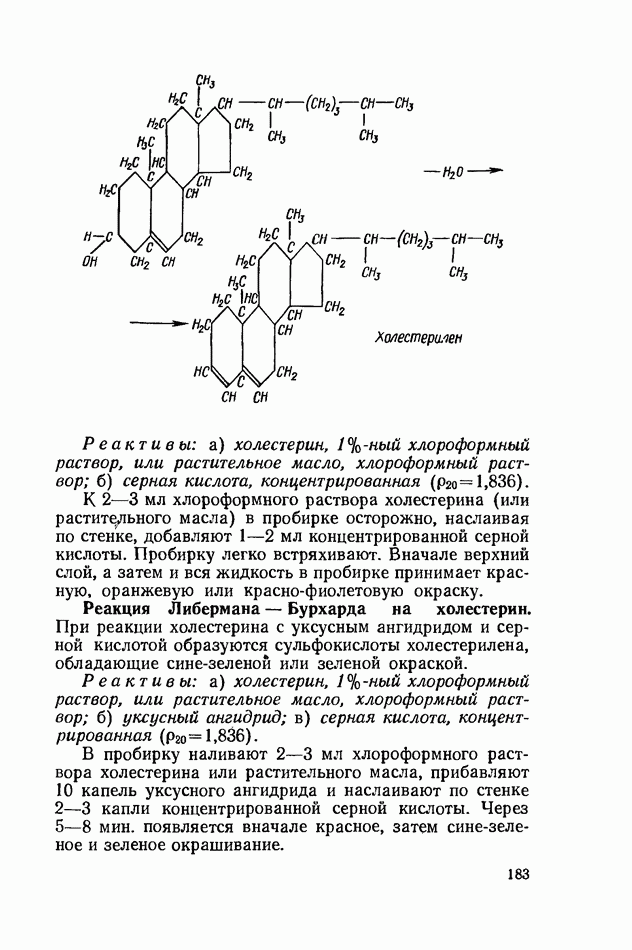
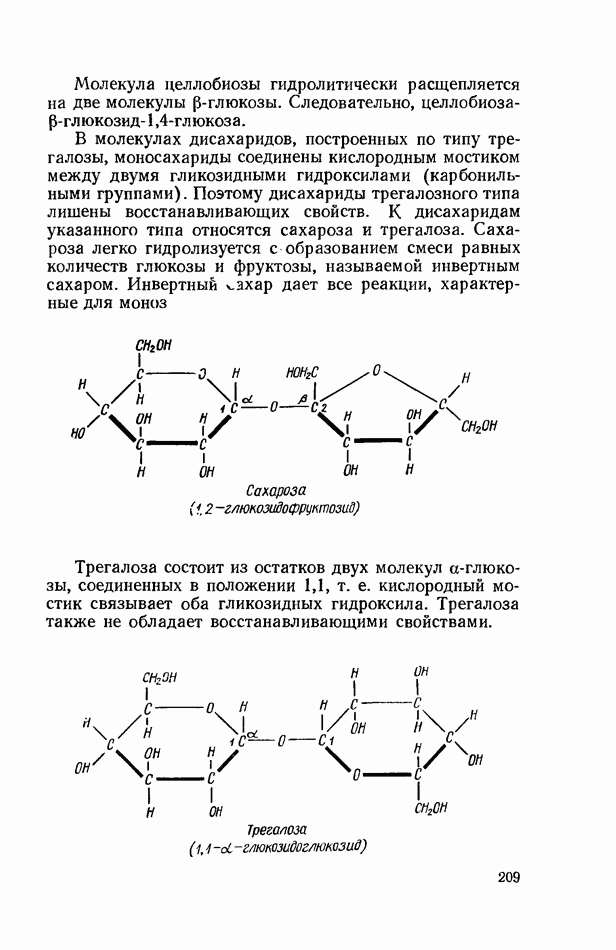
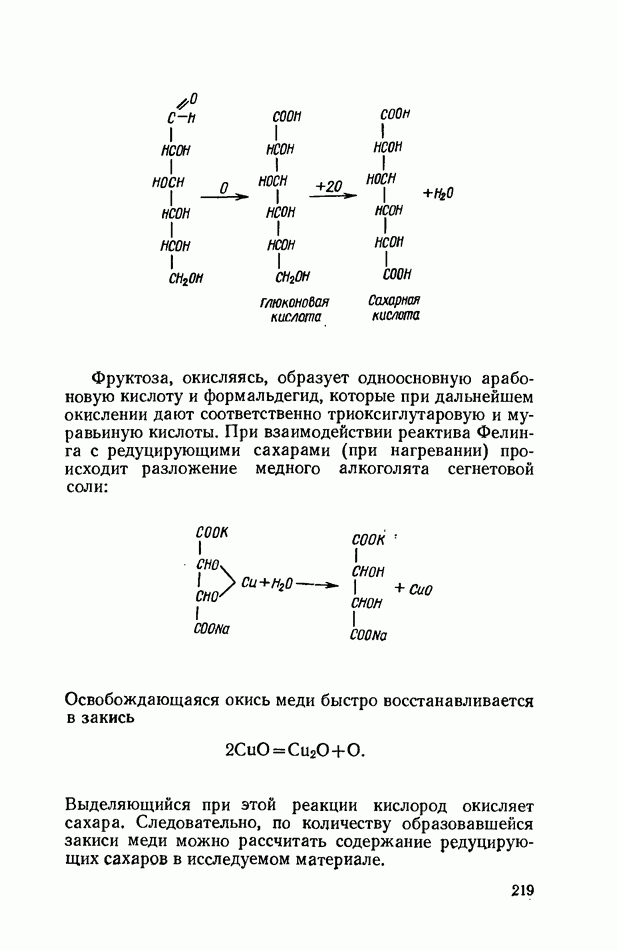
Продукты гидролитического расщепления жиров всасываются в тонком кишечнике. Глицерин растворим в воде и всасывается легко. Жирные кислоты образуют растворимые комплексные соединения с желчными кислотами (так называемые холеиновые кислоты), которые также всасываются в кишечнике. Холеиновые кислоты затем расщепляются на свои компоненты в клетках эпителия кишечных ворсинок. Освободившиеся желчные кислоты всасываются в кровь и через систему воротной вены снова поступают в печень. Жирные же кислоты вступают в сложноэфирную связь с глицерином, образуя жир, свойственный уже данному виду животного. Гидролиз фосфолипидов катализируется ферментами фосфолипазами (А, В, С, D), расщепление холестеридов происходит под влиянием холестеролэстеразы. Качественная реакция на желчные кислоты.Желчные кислоты по своему строению близки к холестерину и являются производными холановой кислоты.
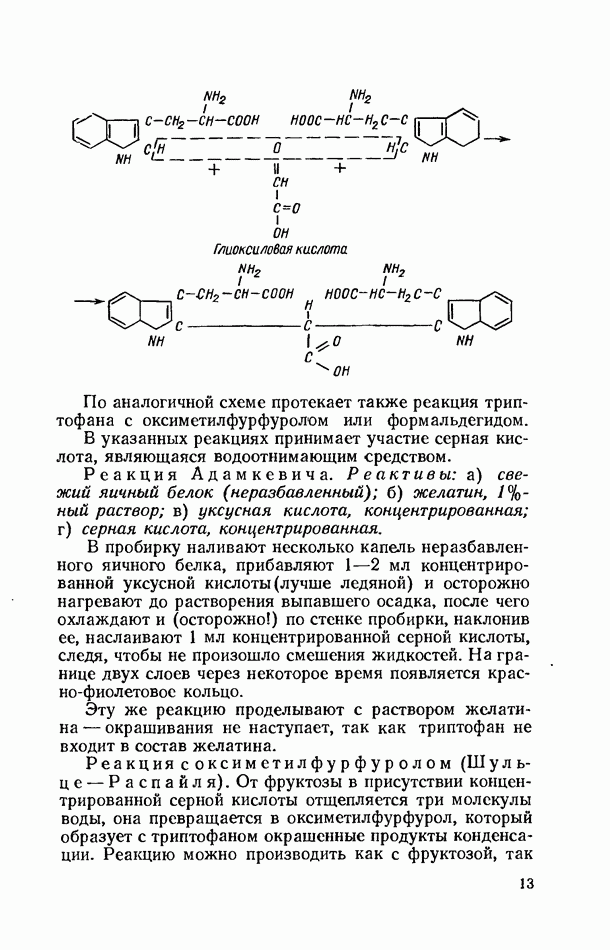
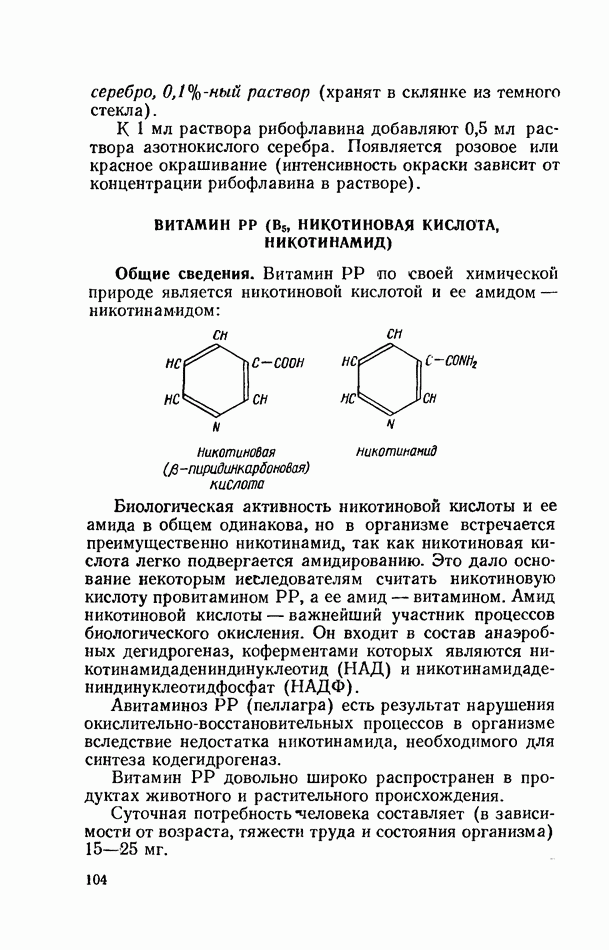
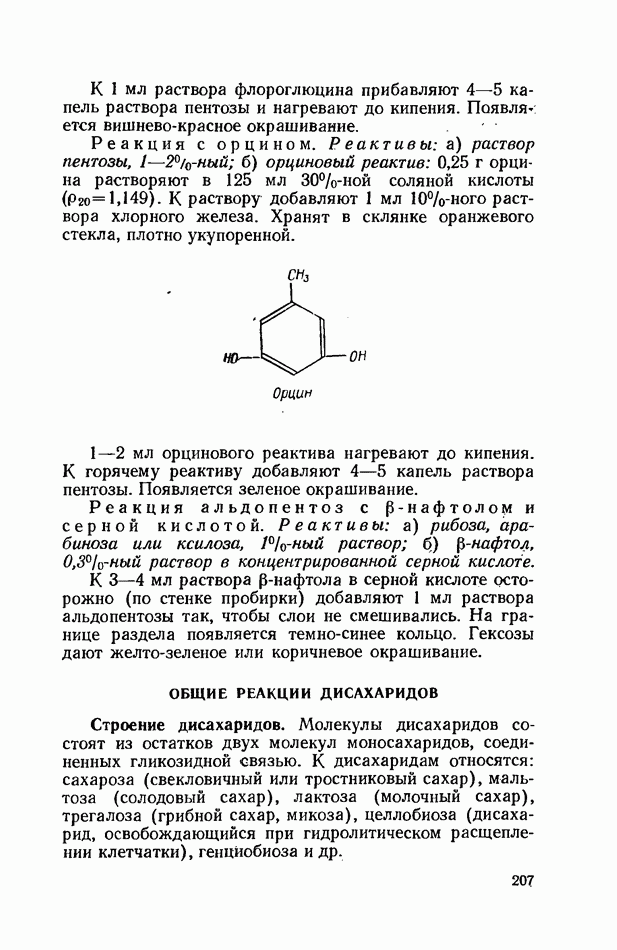
Желчные кислоты (холевая, дезоксихолевая, литохолевая) входят в состав желчи как в чистом виде, так и в виде парных соединений с гликоколлом (глицином) и таурином (с которыми они соединяются посредством пептидной ковалентной связи). Для открытия желчных кислот используют их способность давать красное окрашивание с оксиметилфурфуролом (реакция Петтенкофера). Оксиметилфурфурол образуется при реакции фруктозы с концентрированной соляной или серной кислотой. Реактивы: а) желчь, водный раствор (1:2); б) сахароза, 5%-ный раствор, или фруктоза, 3%-ный раствор; в) серная кислота, концентрированная. В сухую пробирку наливают 10 капель разведенной желчи, добавляют 1—2 капли раствора сахарозы (или фруктозы) и, наклонив пробирку, осторожно (по стенке) наслаивают равный объем концентрированной серной кислоты. На границе слоев образуется пурпурное кольцо, которое затем принимает красно-фиолетовое окрашивание. Исследование действия эмульгаторов на жир.В тонком кишечнике гидролизу подвергаются только эмульгированные жиры. Основной эмульгатор — желчные кислоты, но определенными эмульгирующими свойствами обладают и бикарбонаты кишечного и панкреатического соков, белковые вещества и в сравнительно небольшой степени мыла, содержащиеся в полости кишечника. Реактивы: а) желчь; б) мыло, 2%-ный раствор; в) яичный белок, 2%-ный раствор; г) двууглекислый натрий, 2%-ный раствор; д) растительное масло. В штатив ставят 5 пробирок. В первую из них наливают 20 капель желчи, во вторую — 20 капель раствора мыла, в третью, четвертую и пятую соответственно по 20 капель раствора яичного белка, двууглекислого натрия и дистиллированной воды. Затем во все пробирки вносят по 3 капли растительного масла, содержимое взбалтывают. Через 5 мин. учитывают результаты опыта. Исследование кинетики гидролитического действия липазы поджелудочной железы.Общие сведения. Скорость химической реакции определяется количеством субстрата, подвергшегося превращению в единицу времени. Единицей действия фермента называют то количество его, которое катализирует превращение одного микромоля субстрата в минуту при оптимальных условиях — соответствующей температуре, pH, концентрациях реагирующих веществ и фермента и т. д. Скорость ферментативной реакции зависит также от химической природы субстрата, присутствия активаторов или ингибиторов и некоторых других факторов. Одним из важнейших факторов, определяющих скорость ферментативной реакции, является концентрация субстратов. Фермент Е образует с субстратом S промежуточный комплекс ES, который затем распадается. При этом образуется продукт реакции Р, а фермент регенерирует
где Принимая, что фермент-субстратный комплекс способен к диссоциации, можем написать
т. е. константа диссоциации фермент-субстратного комплекса равна отношению констант скоростей обратной и прямой реакций. Если велико значение константы скорости обратной реакции В соответствии с законом действия масс можно написать
где Преобразуя данное уравнение, получим
или
Чем больше значение [ES], тем выше скорость ферментативной реакции. Наибольшая скорость реакции будет достигнута при Таким образом,
где v — скорость данной реакции; Но
Тогда уравнение (III) примет следующий вид: о
откуда
Уравнение (IV) называют уравнением Михаэлиса—Ментен. При более точных расчетах кинетики ферментативных реакций вместо константы диссоциации KS применяют так называемую константу Михаэлиса
В этом случае уравнение (IV) можно написать
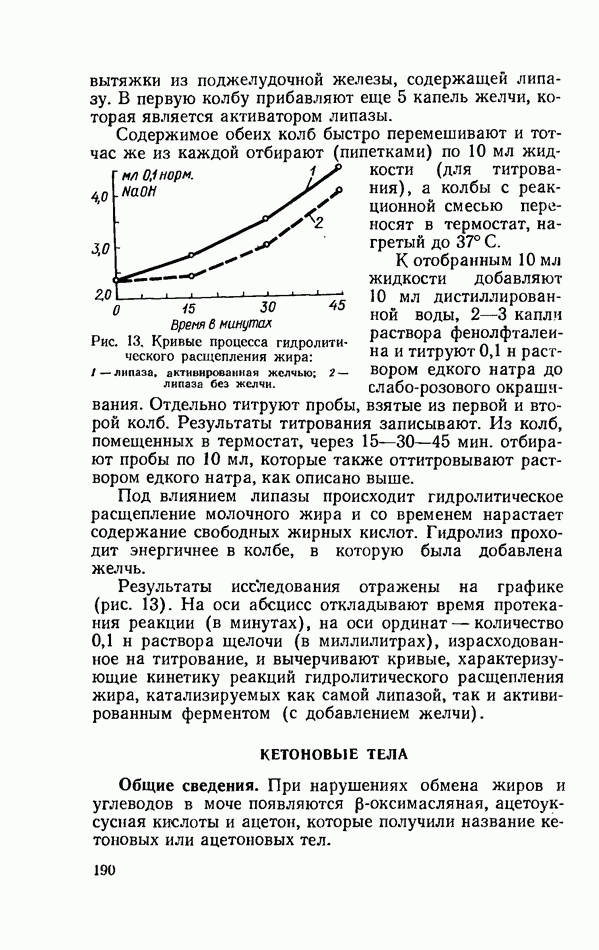
Уравнение (V) известно как уравнение Холдейна — Бриггса. Константа Михаэлиса соответствует той концентрации субстрата (в молях на литр), при которой скорость реакции равна половине максимальной. Кинетика реакции гидролитического расщепления жира. Исследуют скорость реакции расщепления молочного жира, катализируемой липазой поджелудочной железы. Реактивы: а) вытяжка из поджелудочной железы. Свежую поджелудочную железу (крупного рогатого скота, свиньи) тщательно очищают от жира, измельчают на мясорубке и растирают в ступке с водой до получения гомогенной массы (соотношение железы и воды 1:3), которую процеживают через марлю, сложенную в 2—3 слоя; б) молоко. Свежее молоко нагревают до кипения и охлаждают до 37—38° С; в) фенолфталеин, 0,1%-ный спиртовой раствор; г) едкий натр, 0,1 н раствор; д) желчь. В две конические колбы на 100 мл отмеривают по 50 мл кипяченого молока, после чего добавляют по 2 мл вытяжки из поджелудочной железы, содержащей липазу. В первую колбу прибавляют еще 5 капель желчи, которая является активатором липазы. Содержимое обеих колб быстро перемешивают и тотчас же из каждой отбирают (пипетками) по 10 мл жидкости (для титрования), а колбы с реакционной смесью переносят в термостат, нагретый до 37° С.
Рис. 13. Кривые процесса гидролитического расщепления жира: 1-липаза, активированная желчью; 2 — липаза без желчи. К отобранным 10 мл жидкости добавляют 10 мл дистиллированной воды, 2—3 капли раствора фенолфталеина и титруют 0,1 н раствором едкого натра до слабо-розового окрашивания. Отдельно титруют пробы, взятые из первой и второй колб. Результаты титрования записывают. Из колб, помещенных в термостат, через 15—30—45 мин. отбирают пробы по 10 мл, которые также оттитровывают раствором едкого натра, как описано выше. Под влиянием липазы происходит гидролитическое расщепление молочного жира и со временем нарастает содержание свободных жирных кислот. Гидролиз проходит энергичнее в колбе, в которую была добавлена желчь. Результаты исследования отражены на графике (рис. 13). На оси абсцисс откладывают время протекания реакции (в минутах), на оси ординат — количество 0,1 н раствора щелочи (в миллилитрах), израсходованное на титрование, и вычерчивают кривые, характеризующие кинетику реакций гидролитического расщепления жира, катализируемых как самой липазой, так и активированным ферментом (с добавлением желчи).
|
 |
Оглавление
|





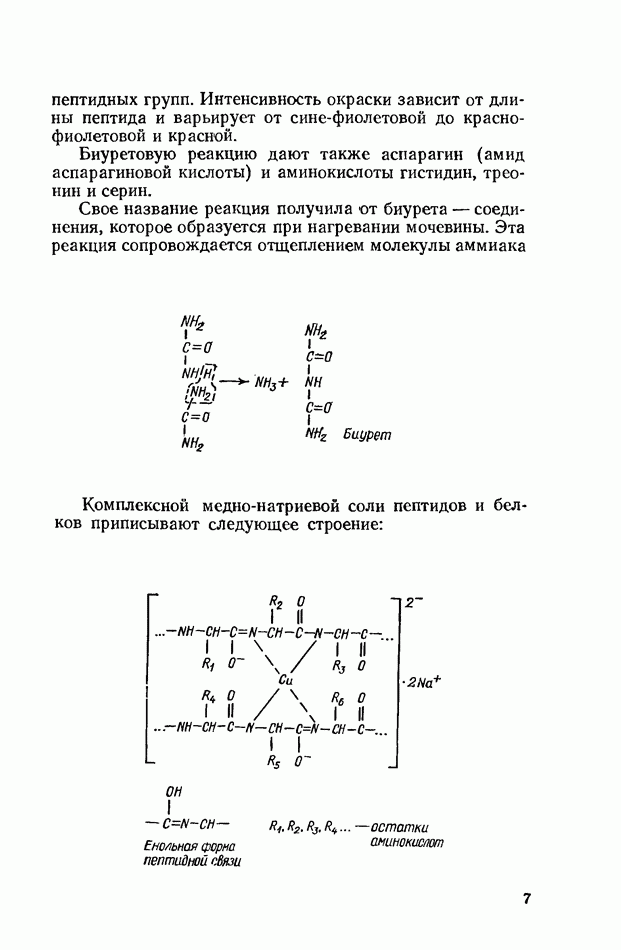








































































































































































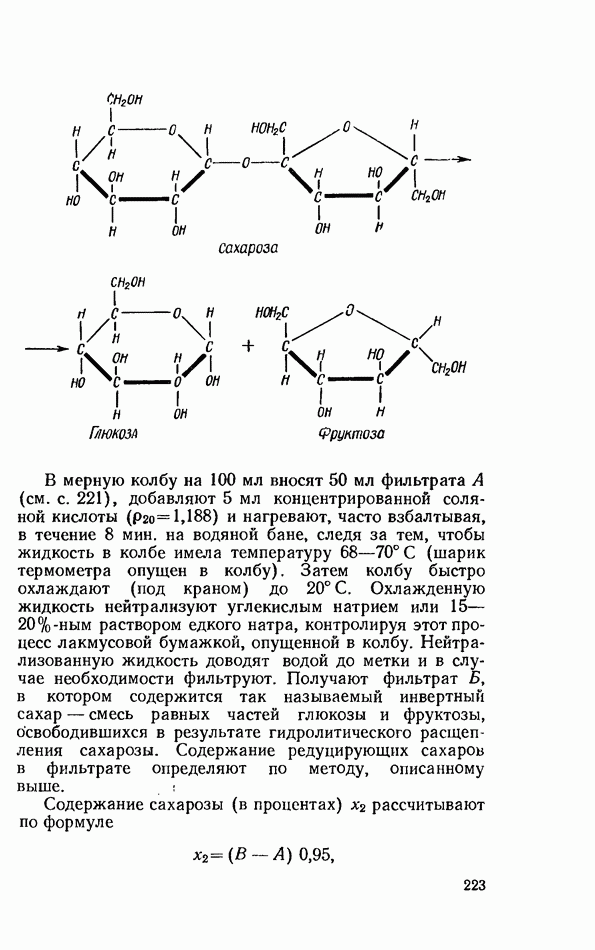








































 -положении, и образуется глицерин-2-жирная кислота, которая затем изомеризуется в
-положении, и образуется глицерин-2-жирная кислота, которая затем изомеризуется в  кислоту, подвергающуюся уже окончательному расщеплению.
кислоту, подвергающуюся уже окончательному расщеплению.



 константа скорости прямой реакции;
константа скорости прямой реакции;  константа скорости обратной реакции;
константа скорости обратной реакции;  константа скорости реакции расщепления фермент-субстратного комплекса;
константа скорости реакции расщепления фермент-субстратного комплекса;  скорости обратной реакции.
скорости обратной реакции.

 и мало значение
и мало значение  то константа диссоциации фермент-субстратного комплекса является высокой и комплекс легко подвергается распаду на исходные вещества. В этом случае реакция протекает с малой скоростью. Если же, наоборот, значение
то константа диссоциации фермент-субстратного комплекса является высокой и комплекс легко подвергается распаду на исходные вещества. В этом случае реакция протекает с малой скоростью. Если же, наоборот, значение  велико,
велико,  мало, то реакция идет с большой скоростью.
мало, то реакция идет с большой скоростью.

 — общая концентрация фермента в начале реакции;
— общая концентрация фермента в начале реакции;  — концентрация фермент-субстратного комплексного соединения;
— концентрация фермент-субстратного комплексного соединения;  — концентрация свободного фермента за вычетом той части, которая связана с субстратом.
— концентрация свободного фермента за вычетом той части, которая связана с субстратом.


 т. е. когда весь фермент соединится с субстратом.
т. е. когда весь фермент соединится с субстратом.

 — максимальная скорость реакции.
— максимальная скорость реакции.







