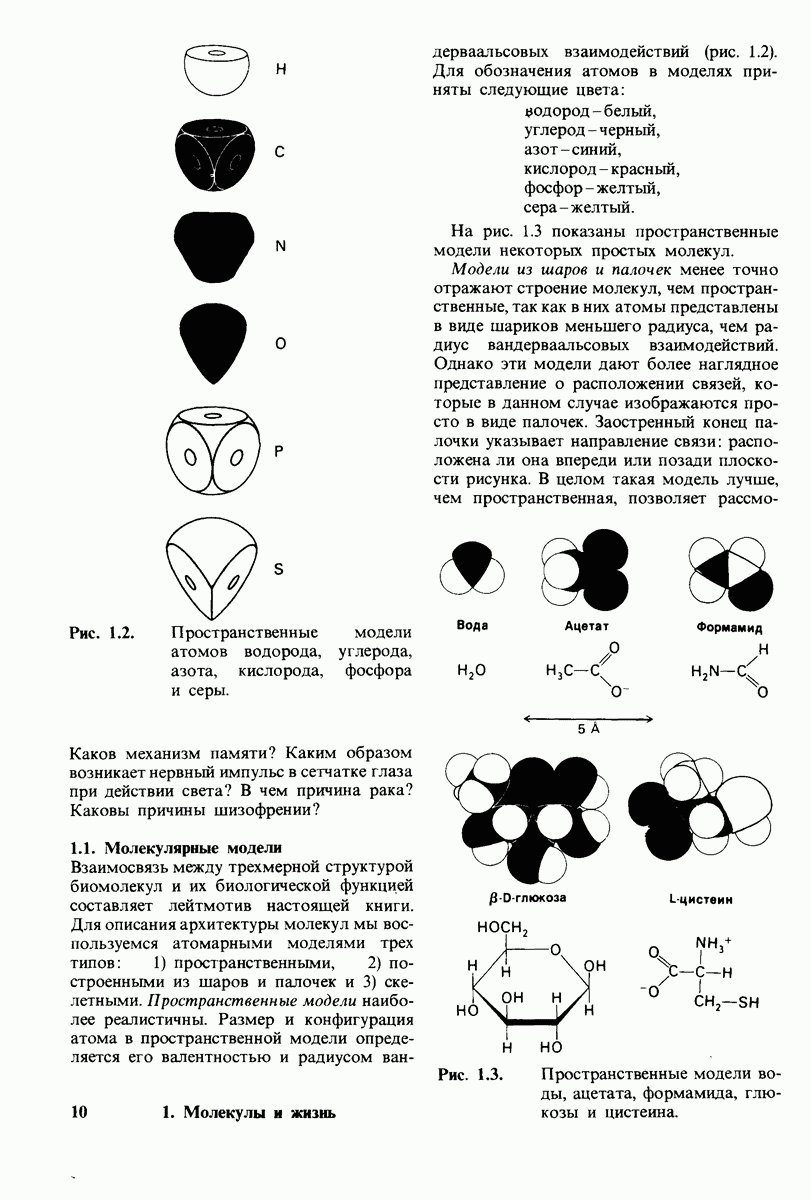
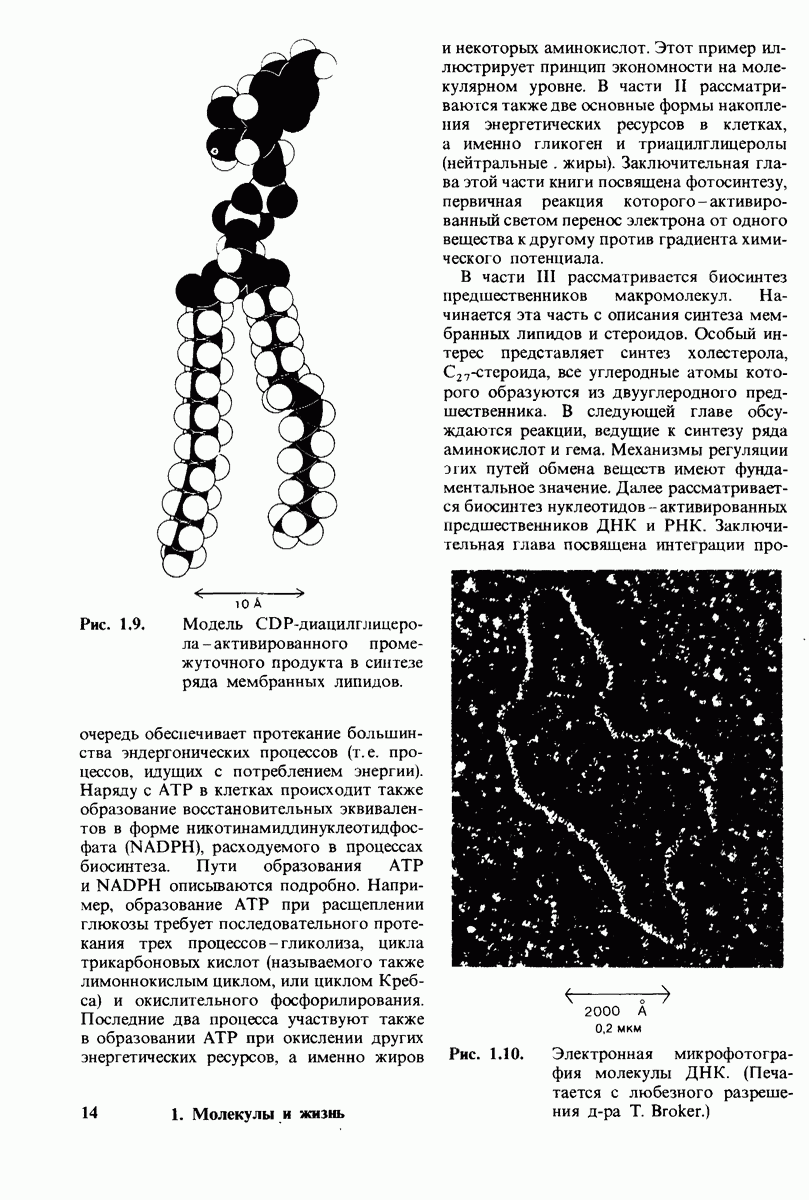
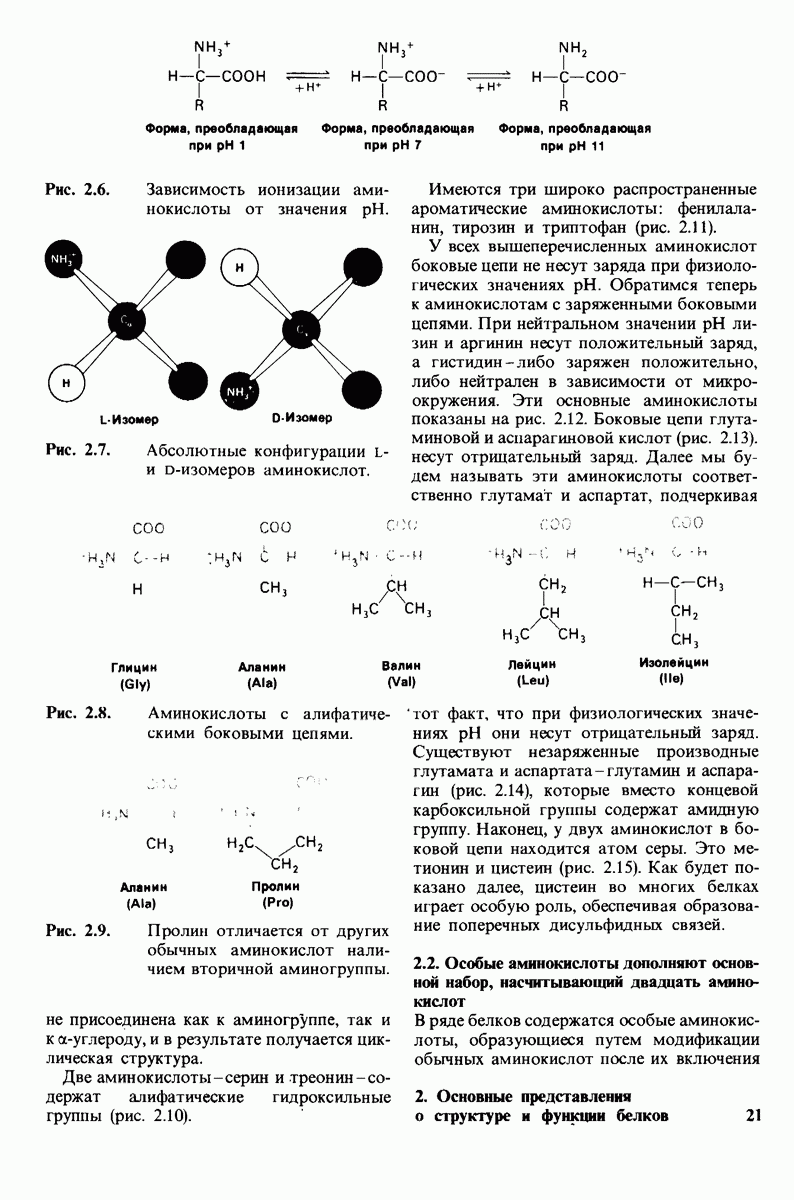
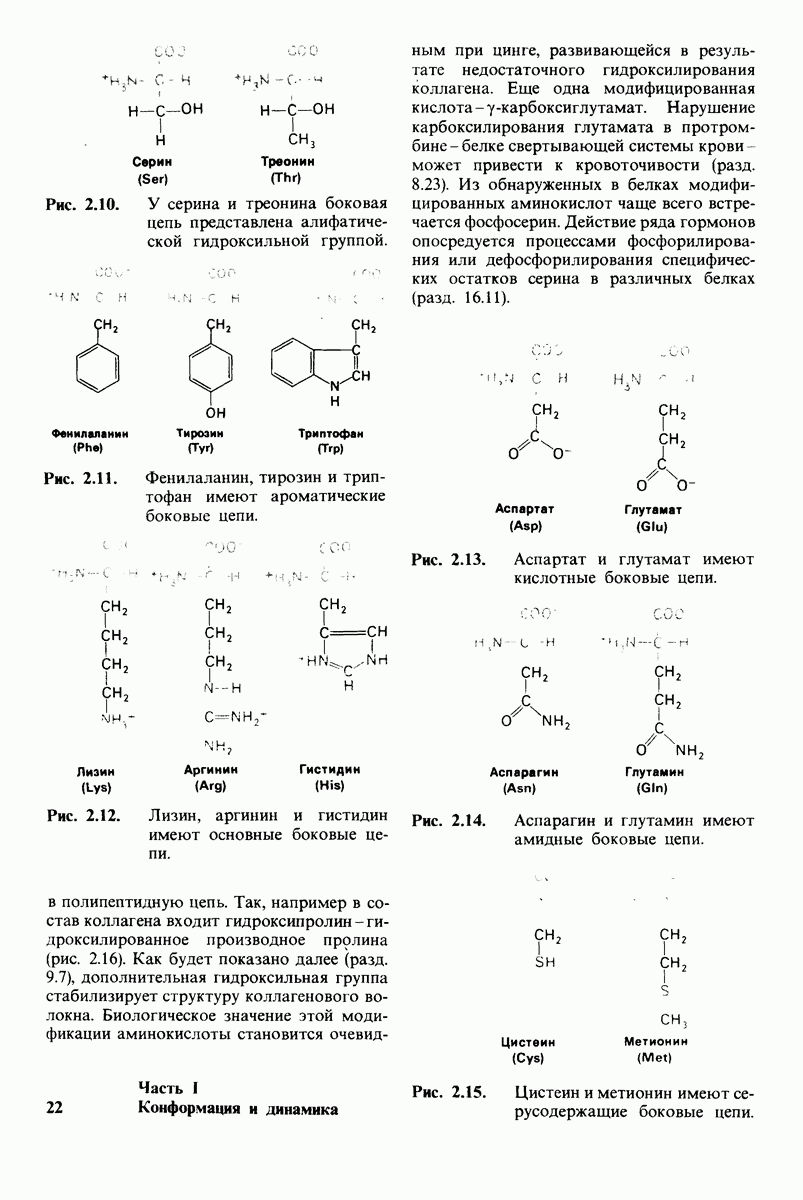
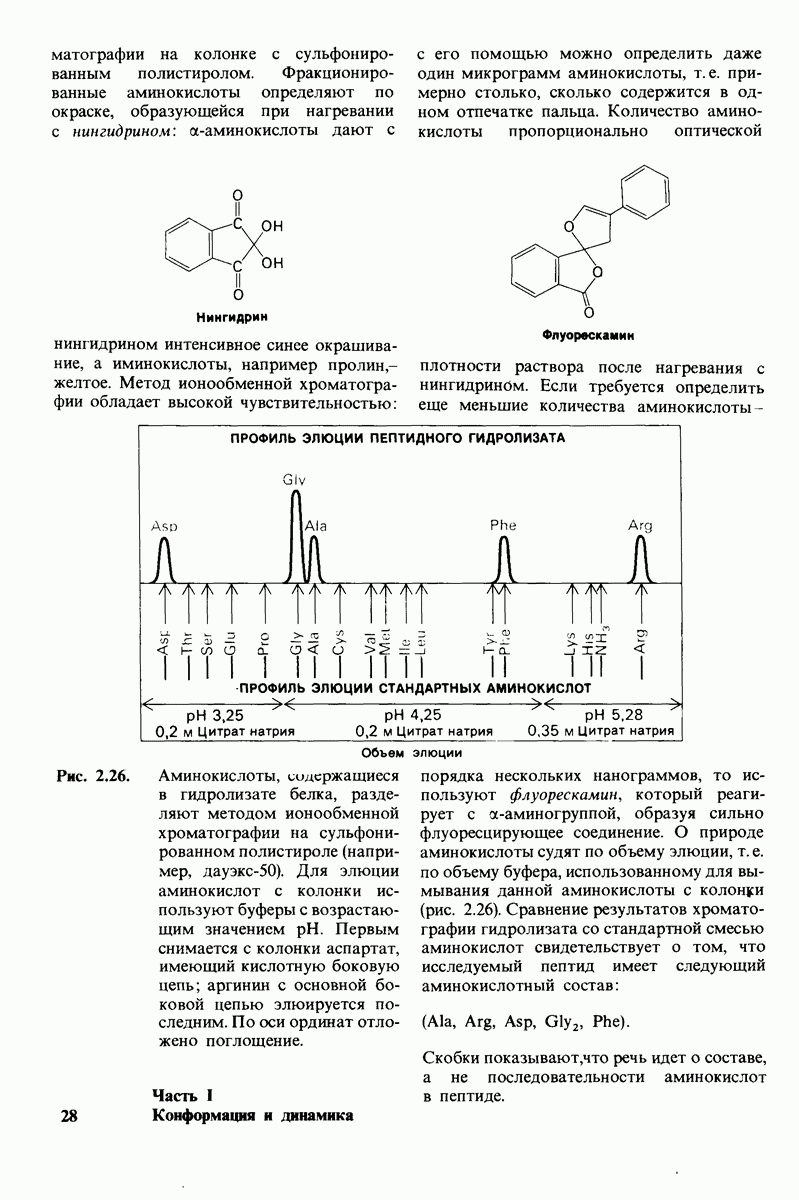
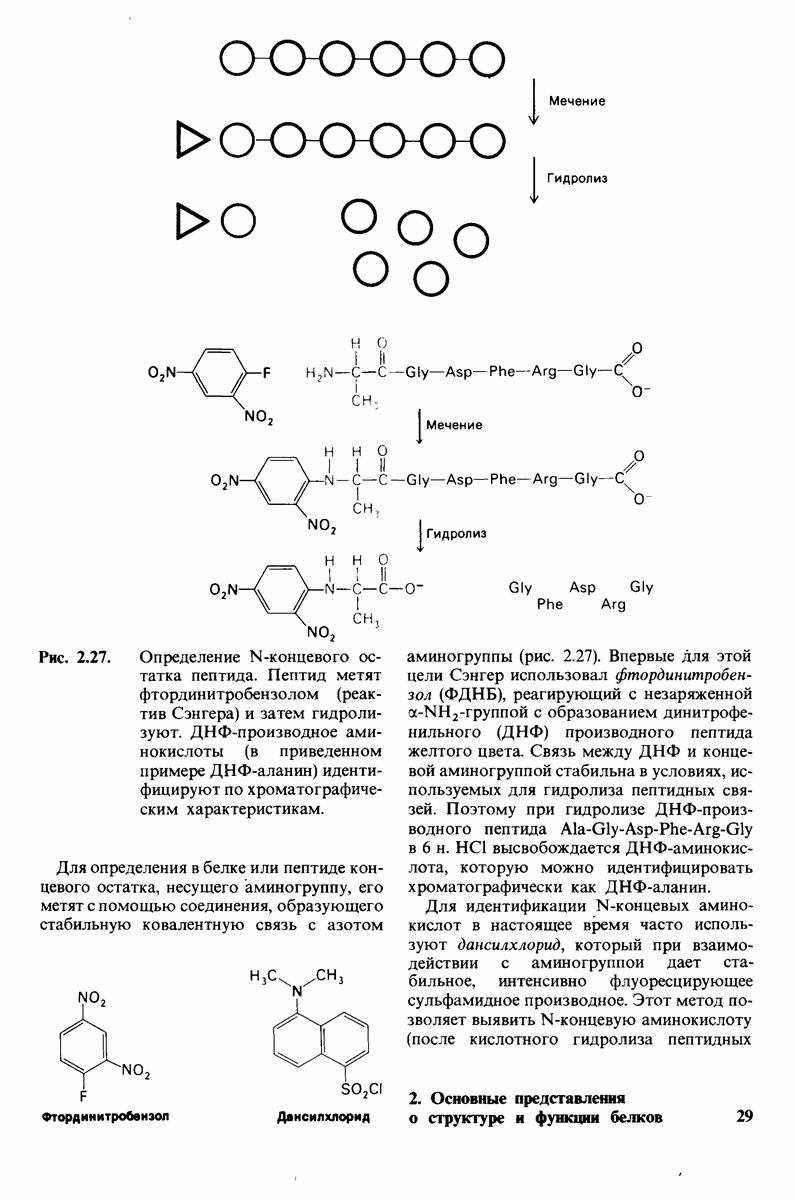
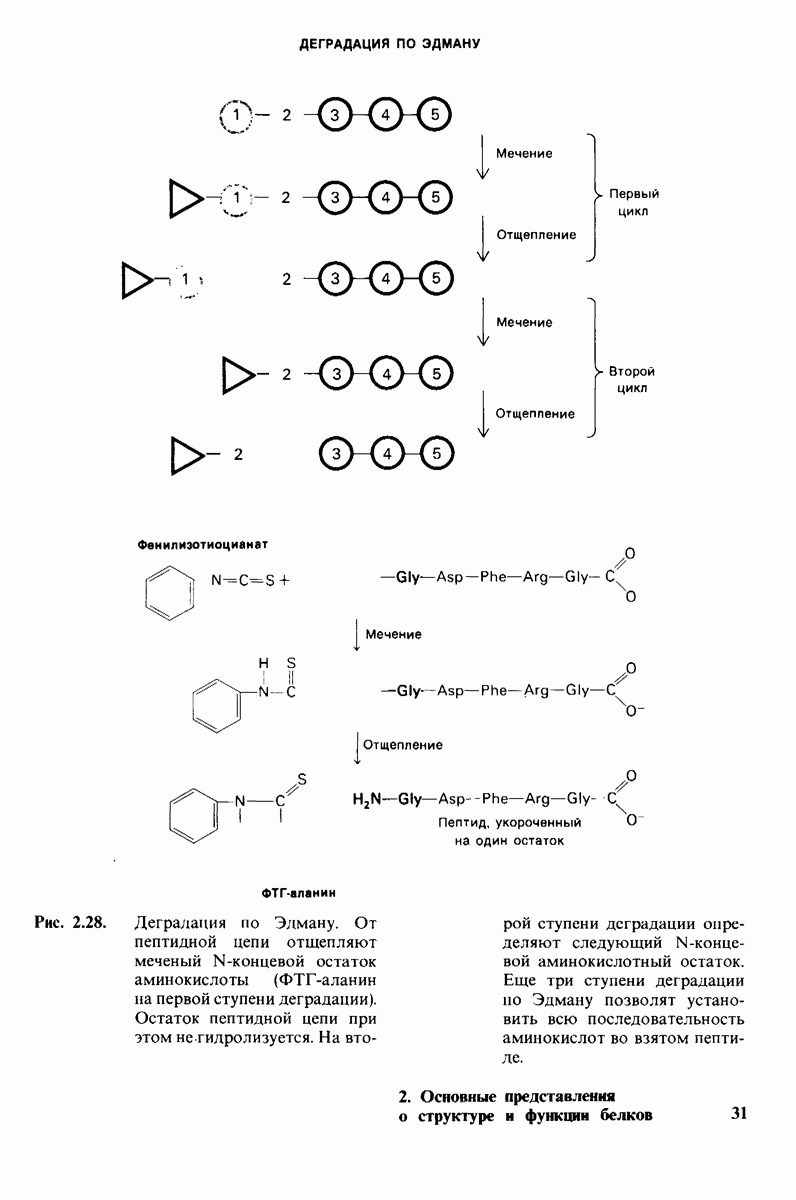
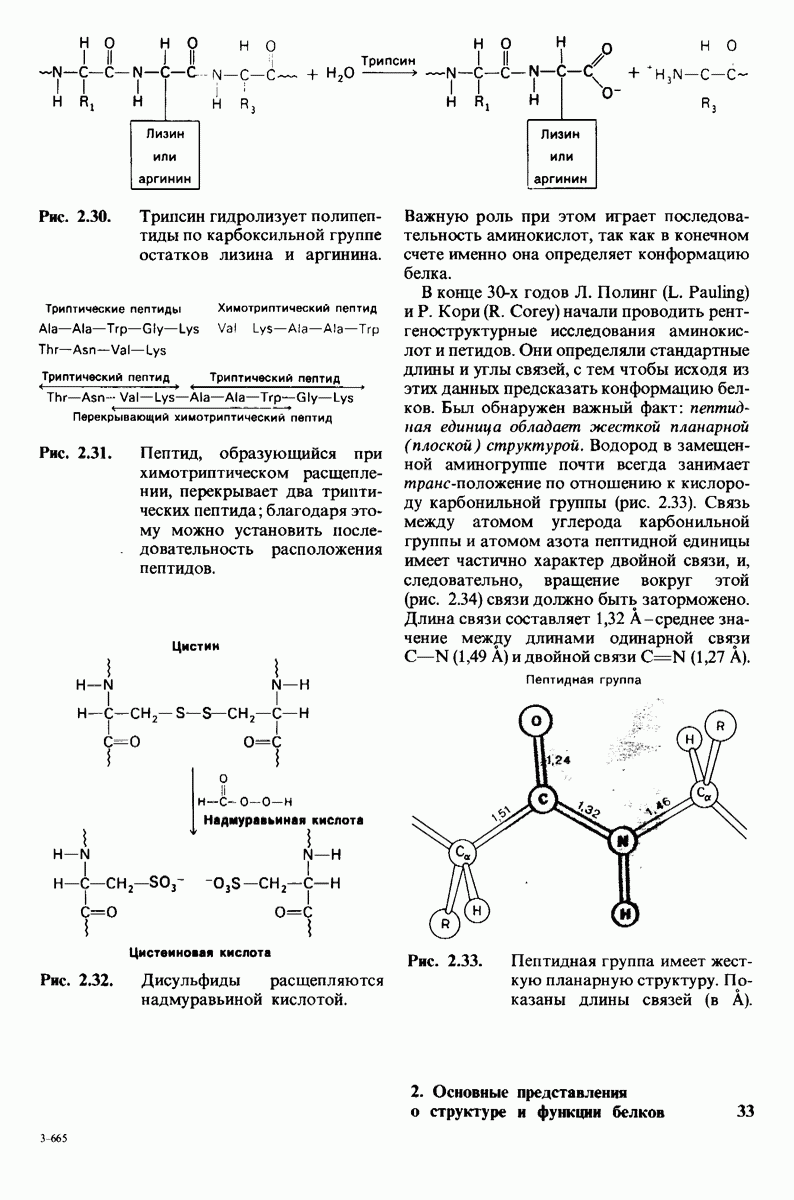
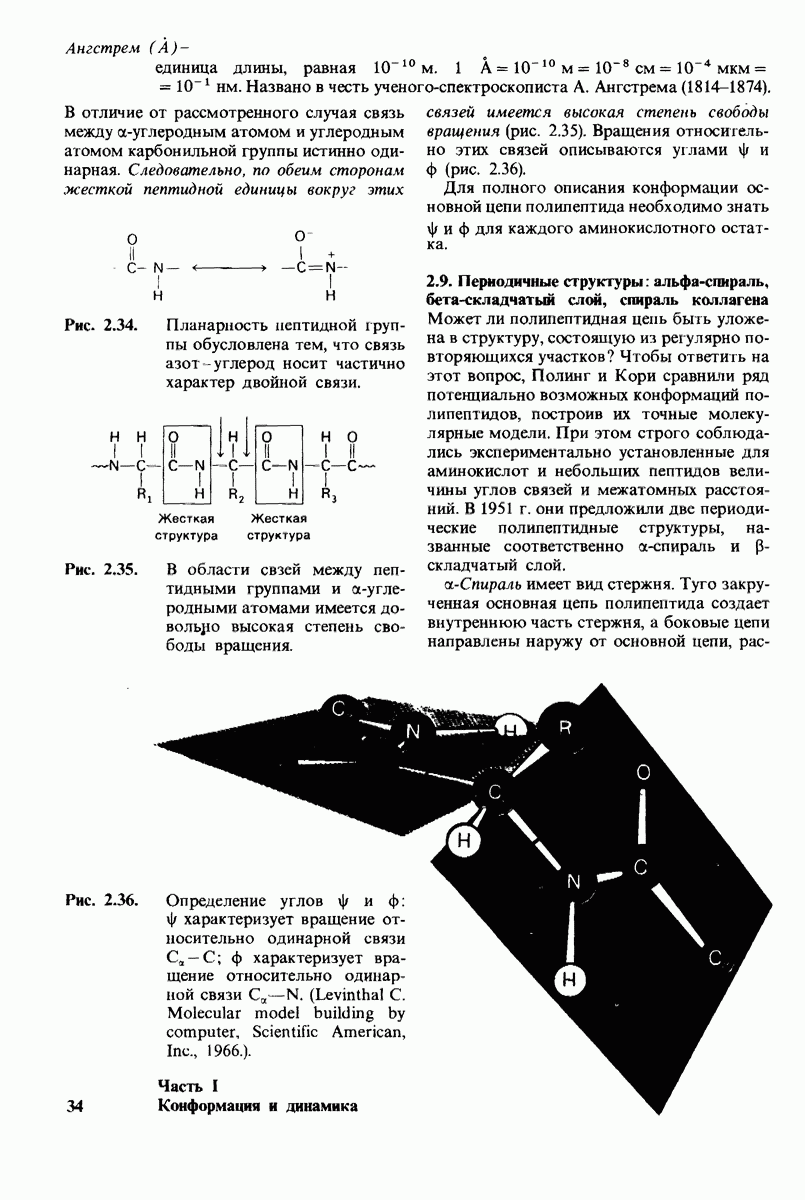
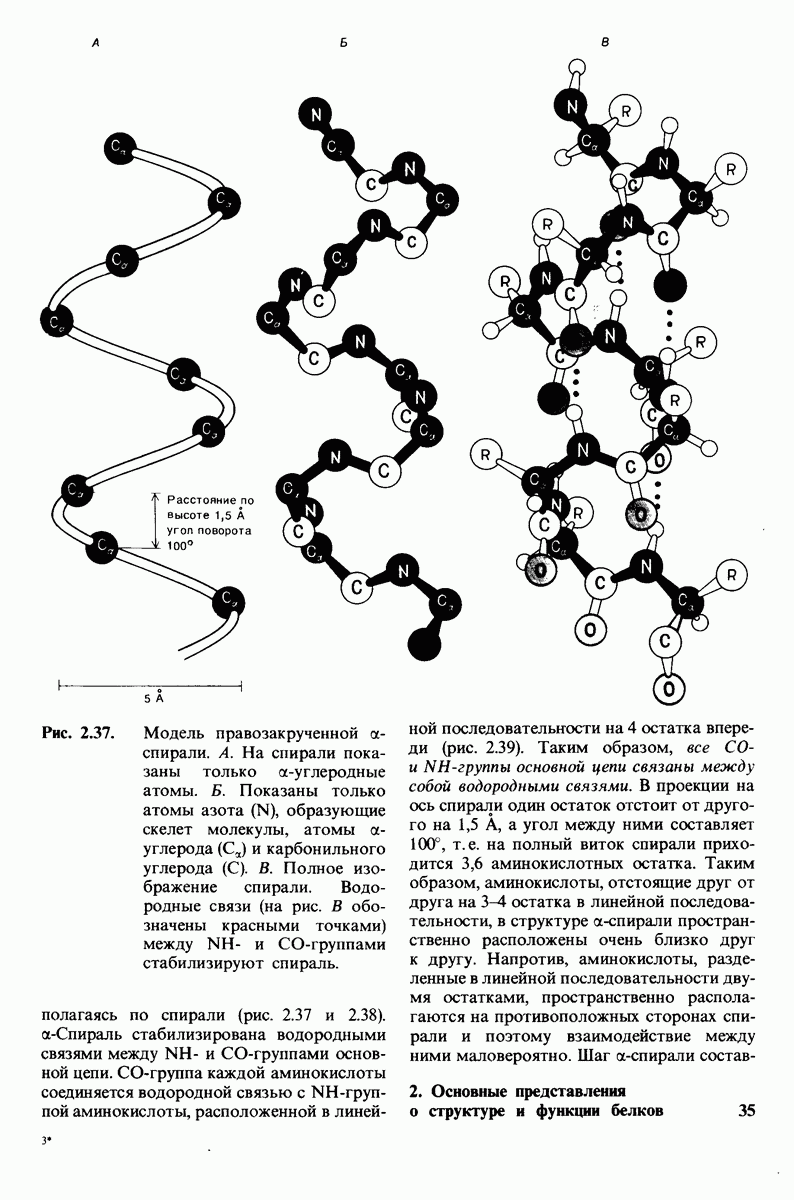
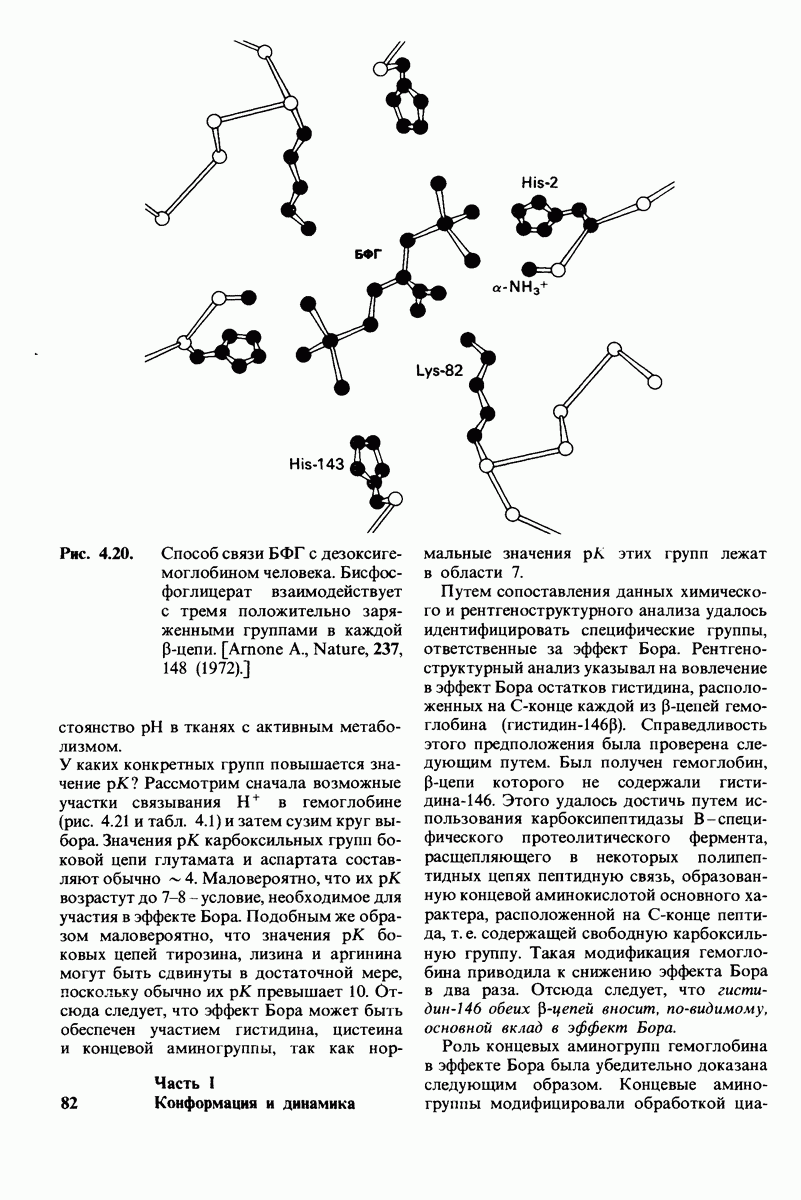
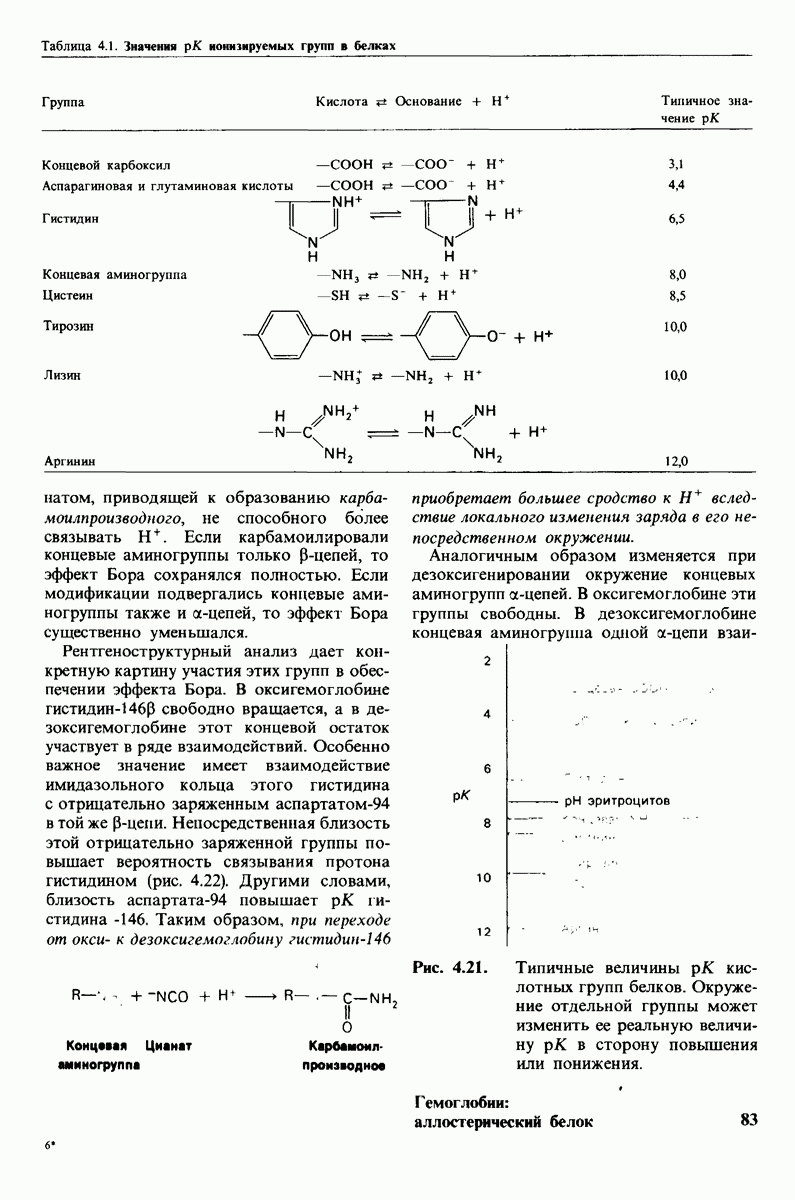
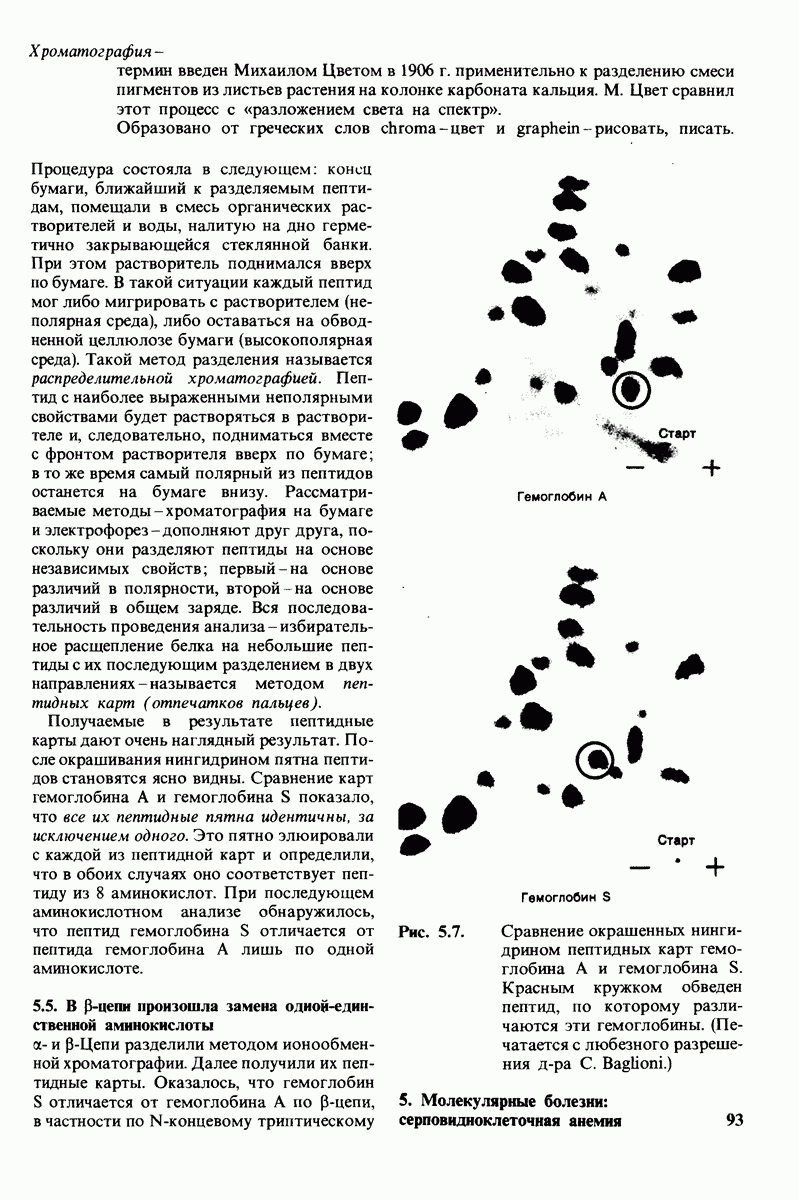
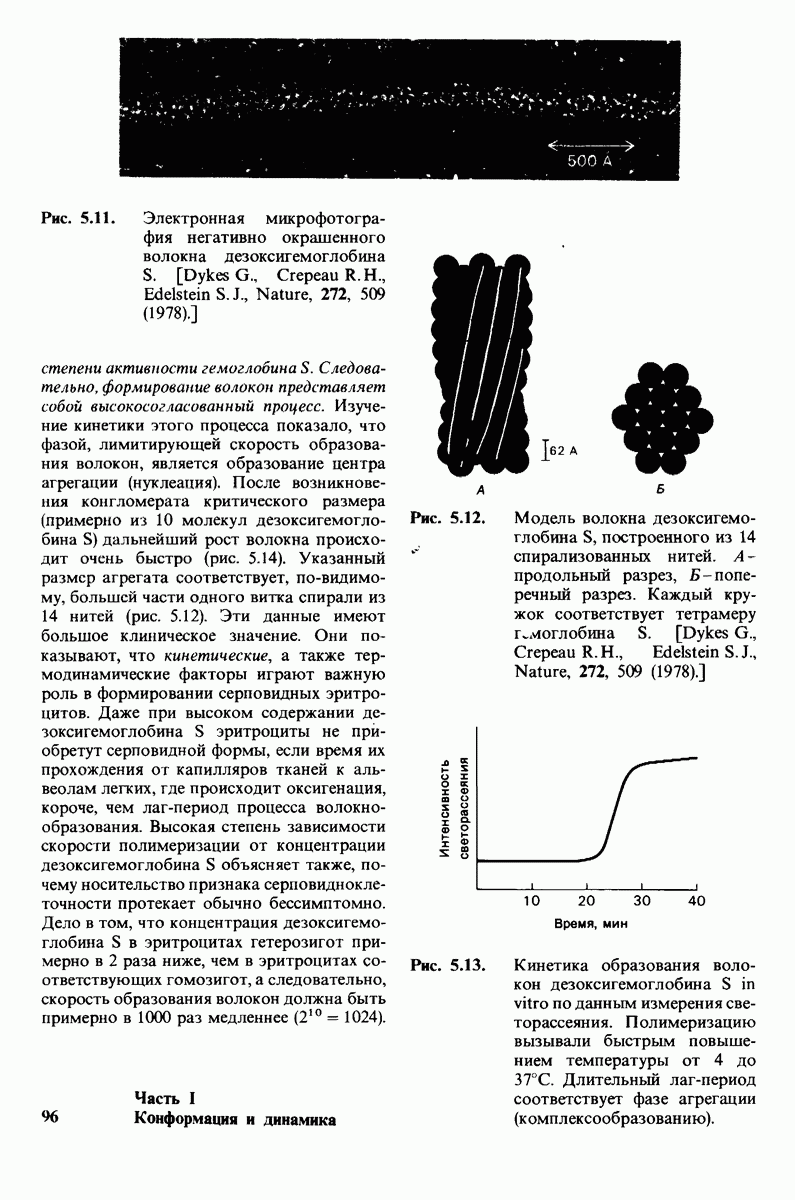
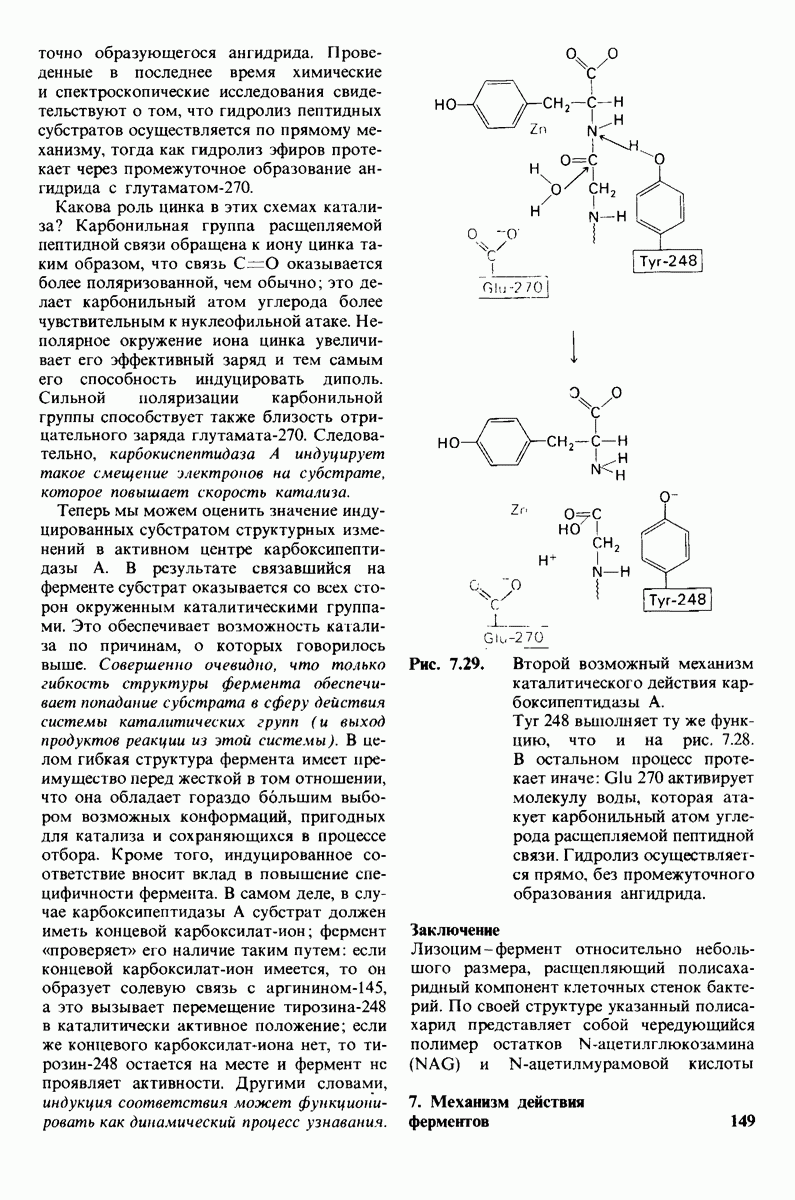
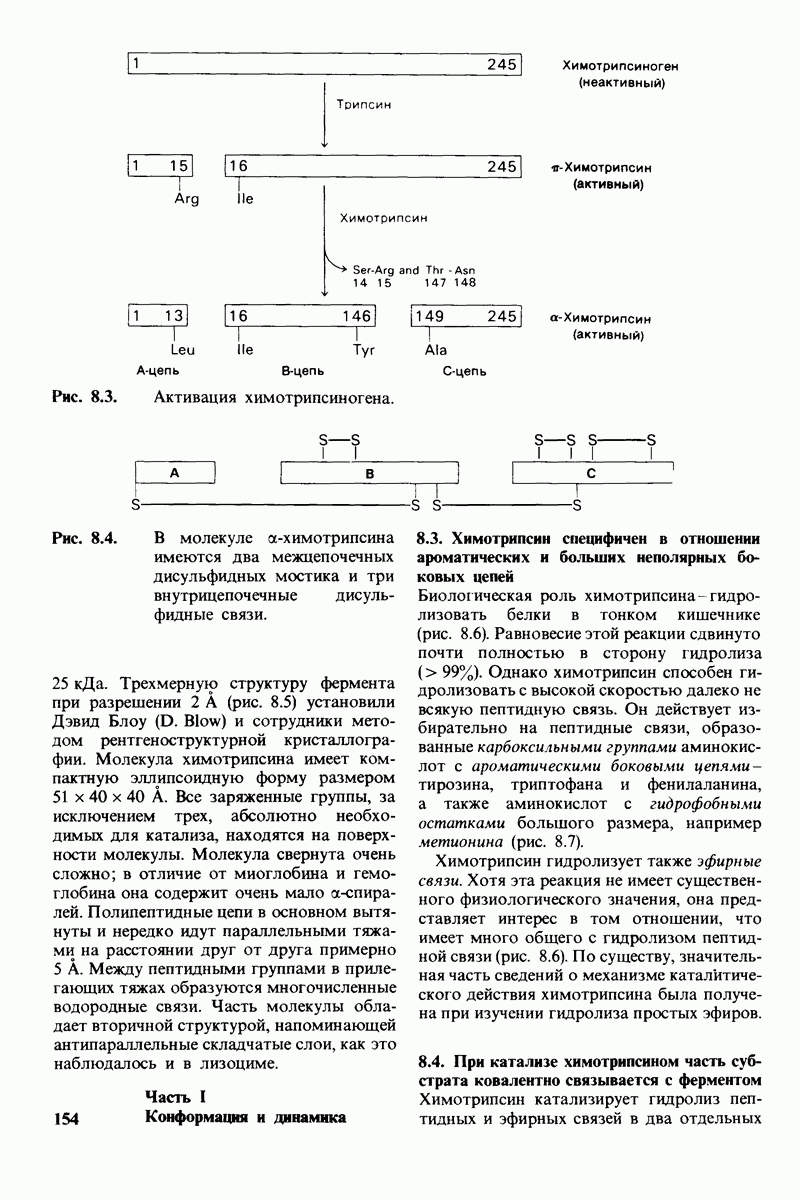
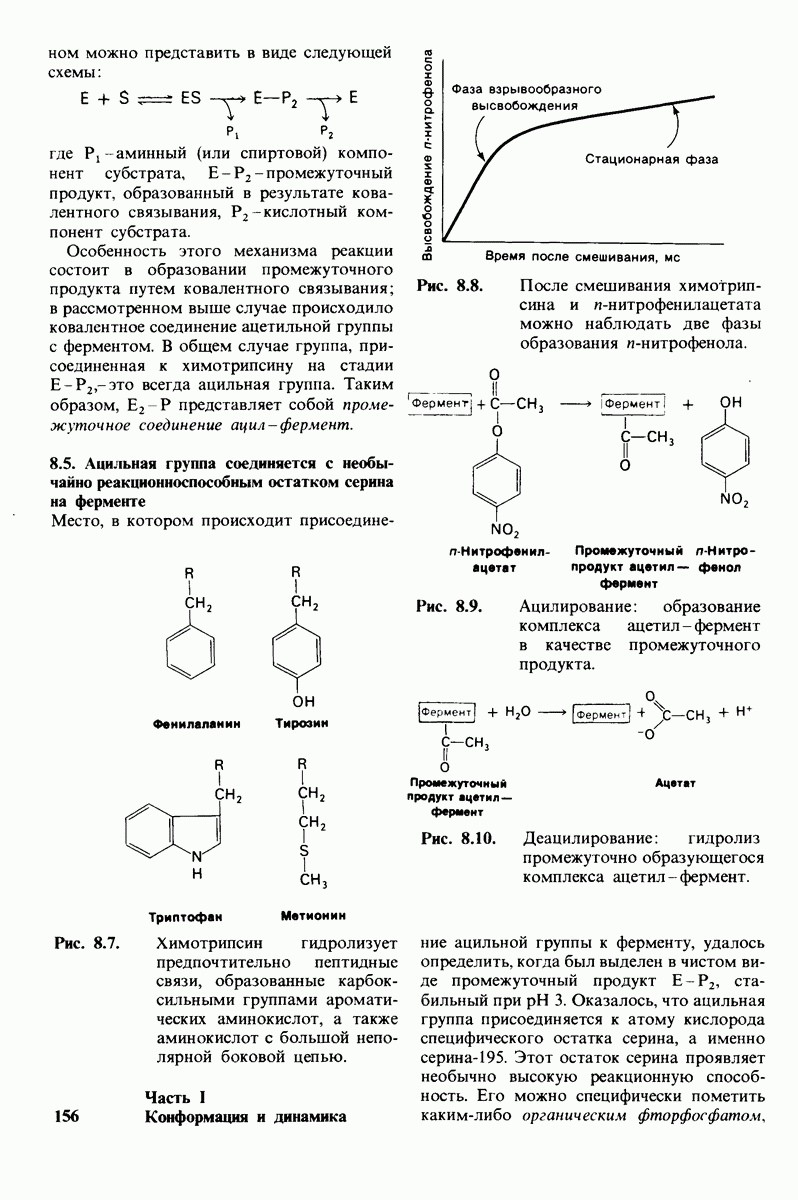
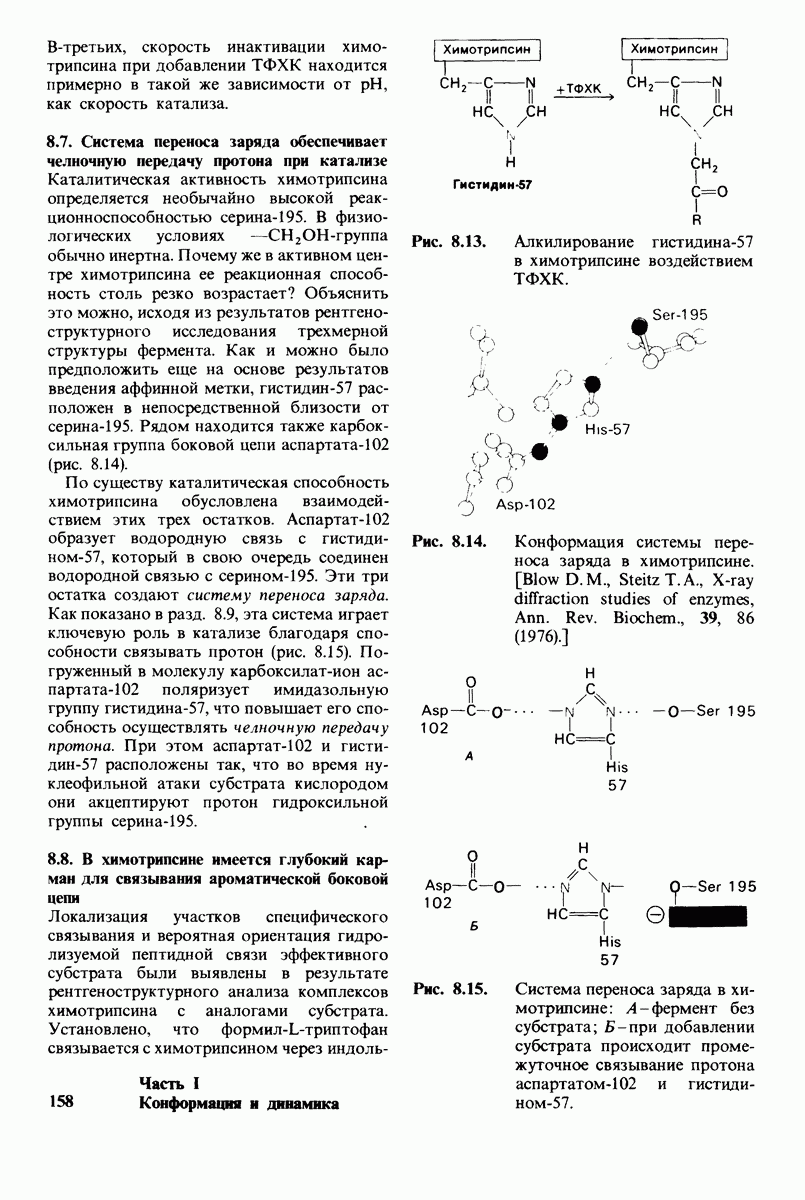
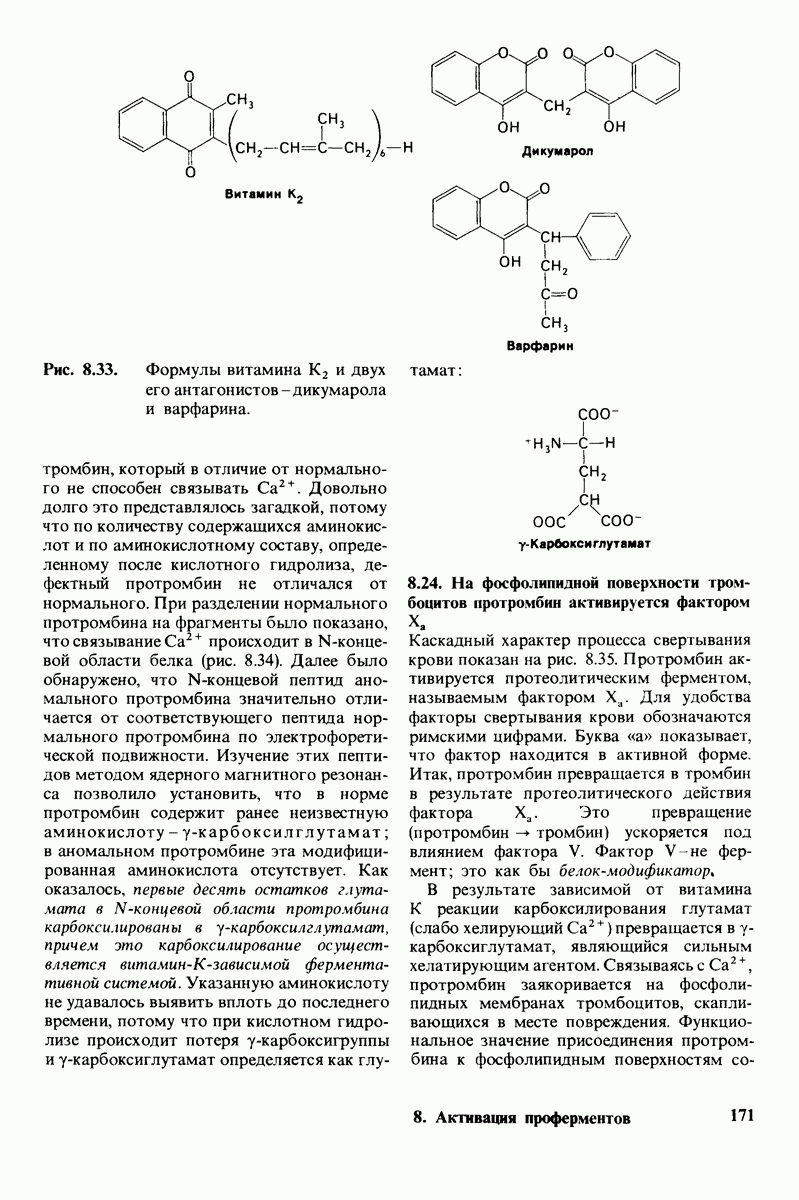
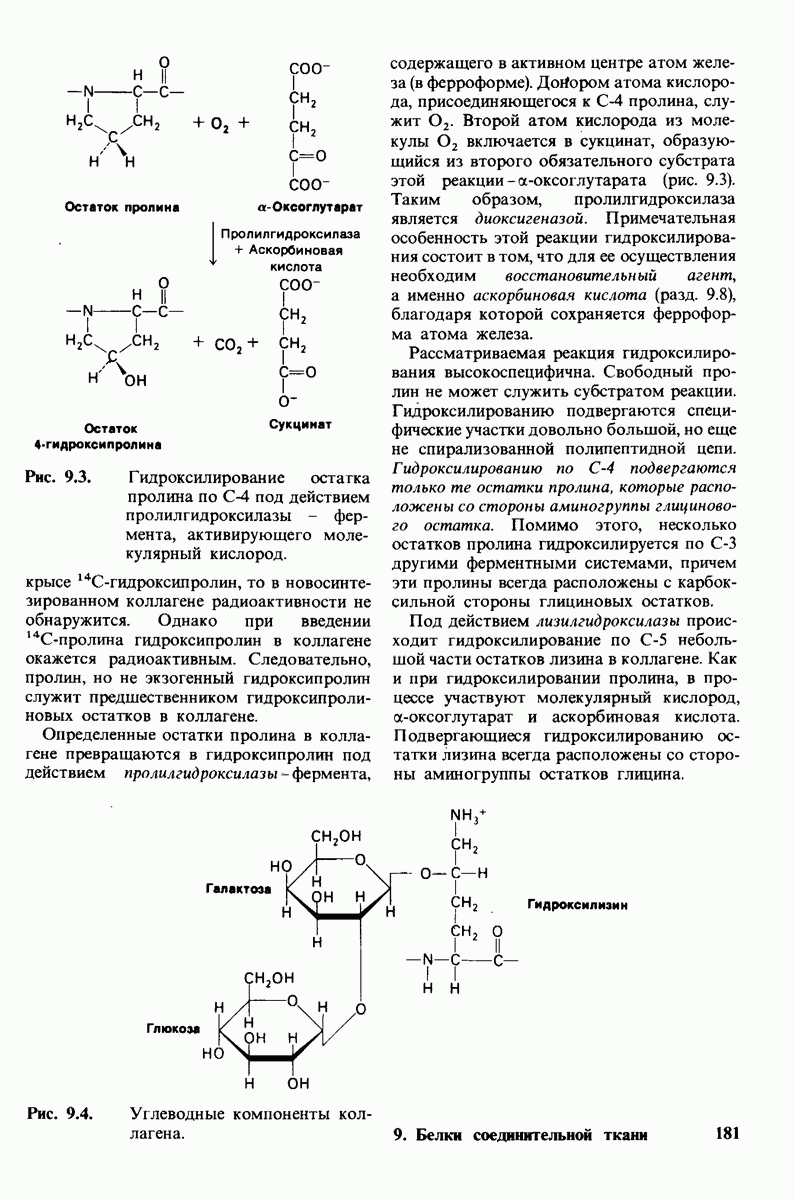
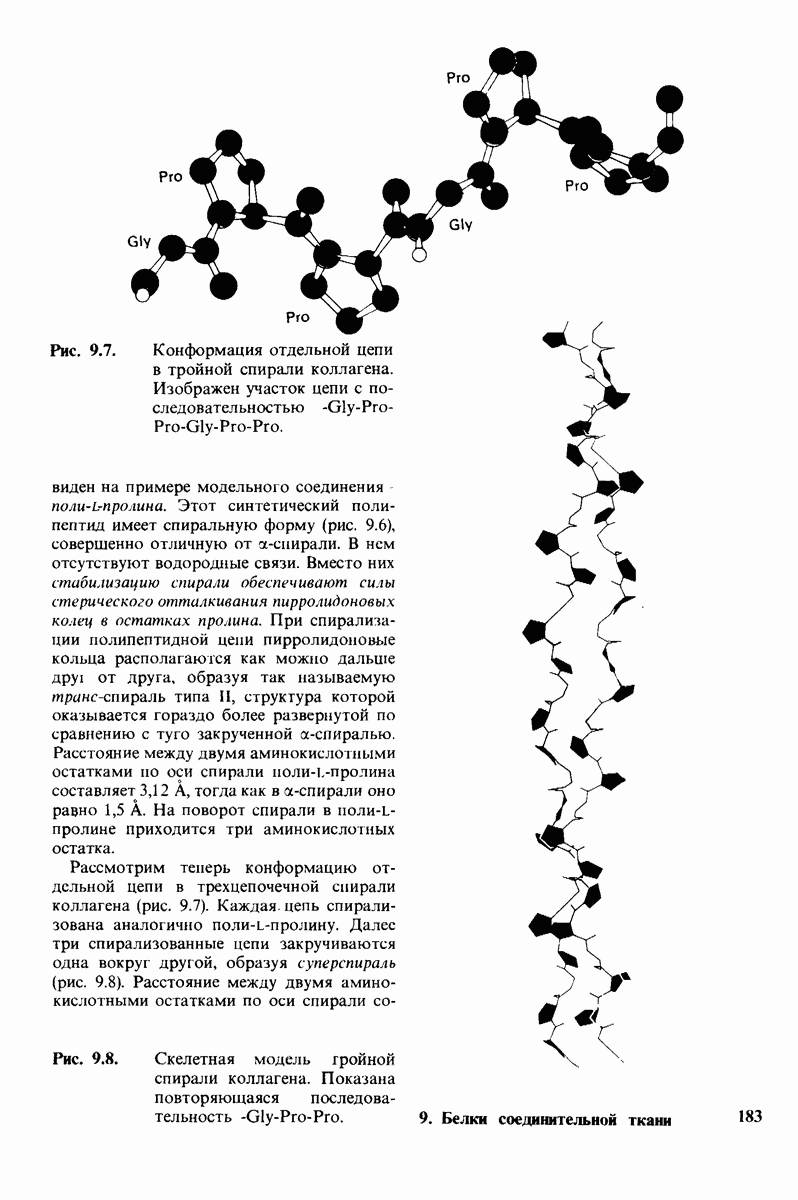
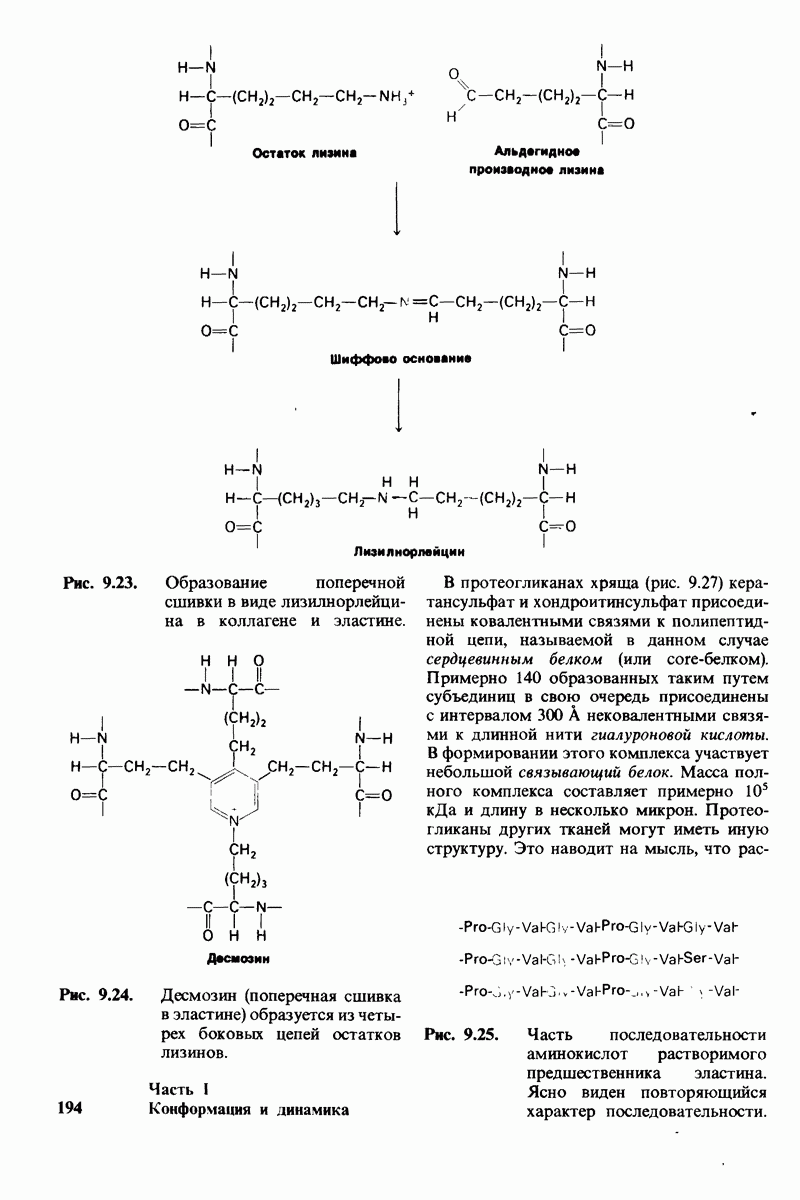
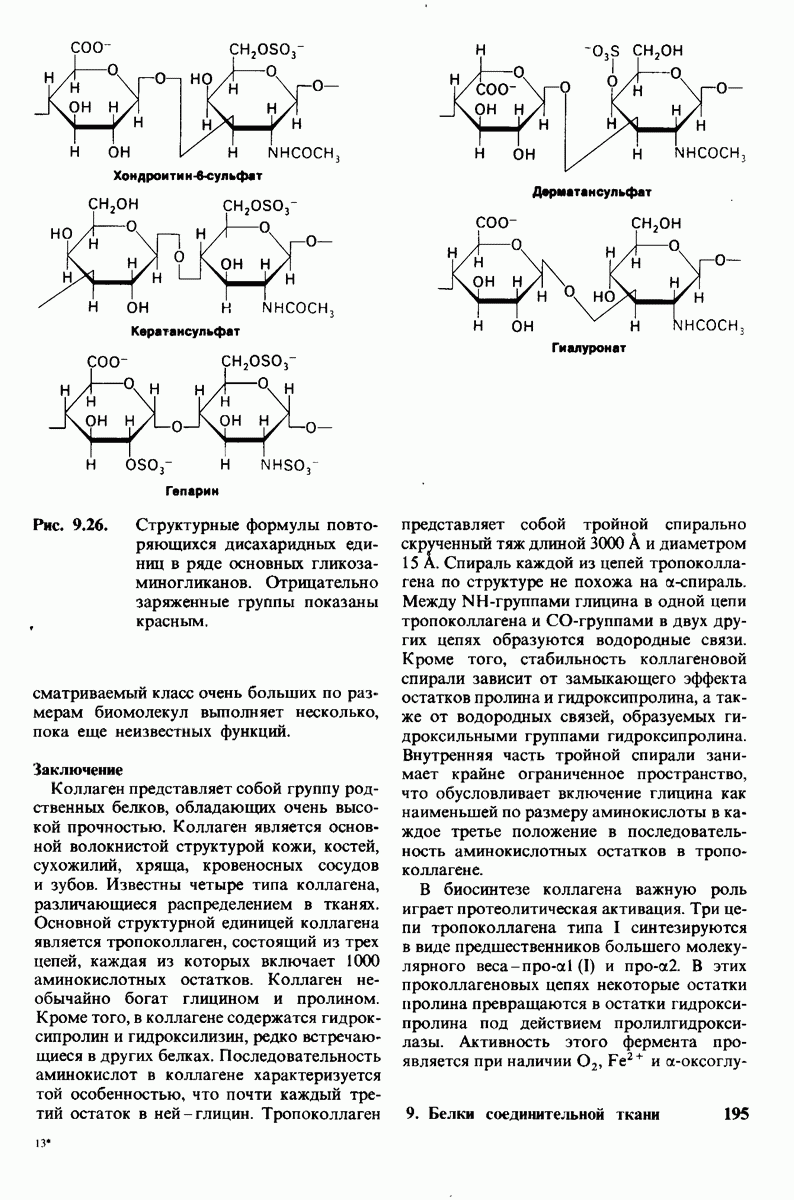
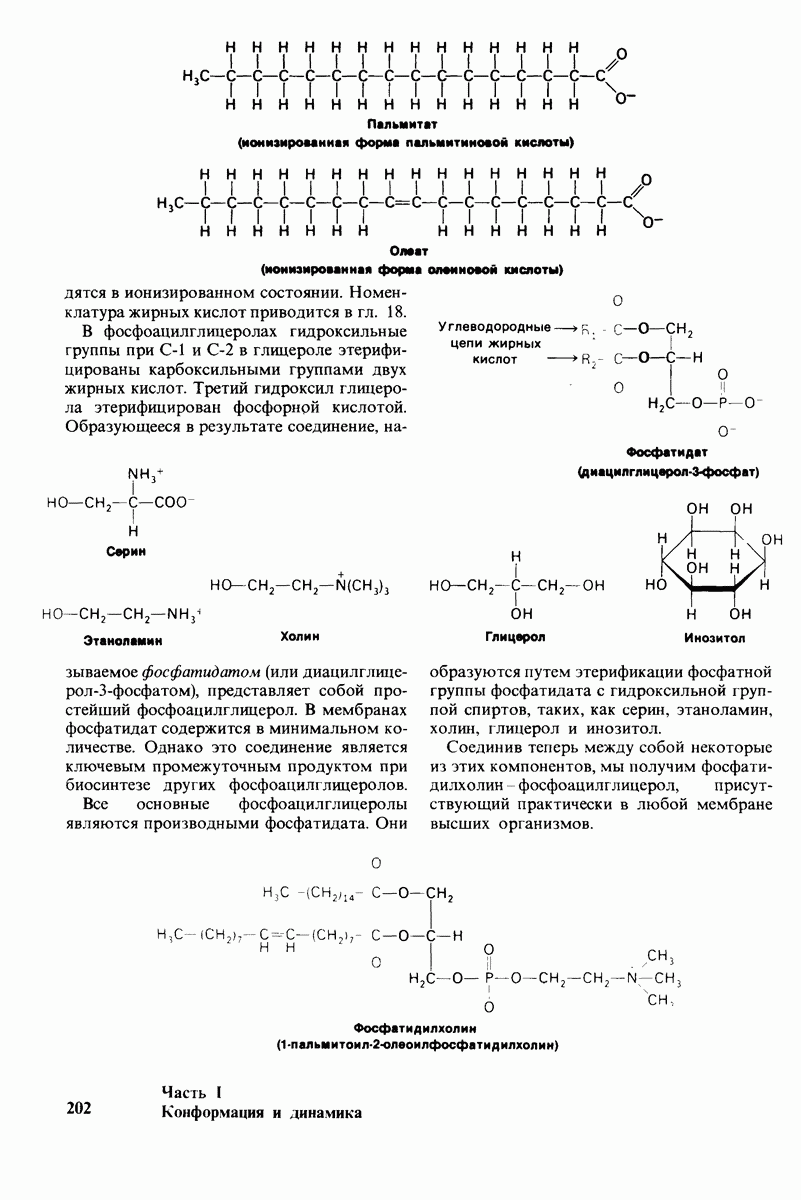
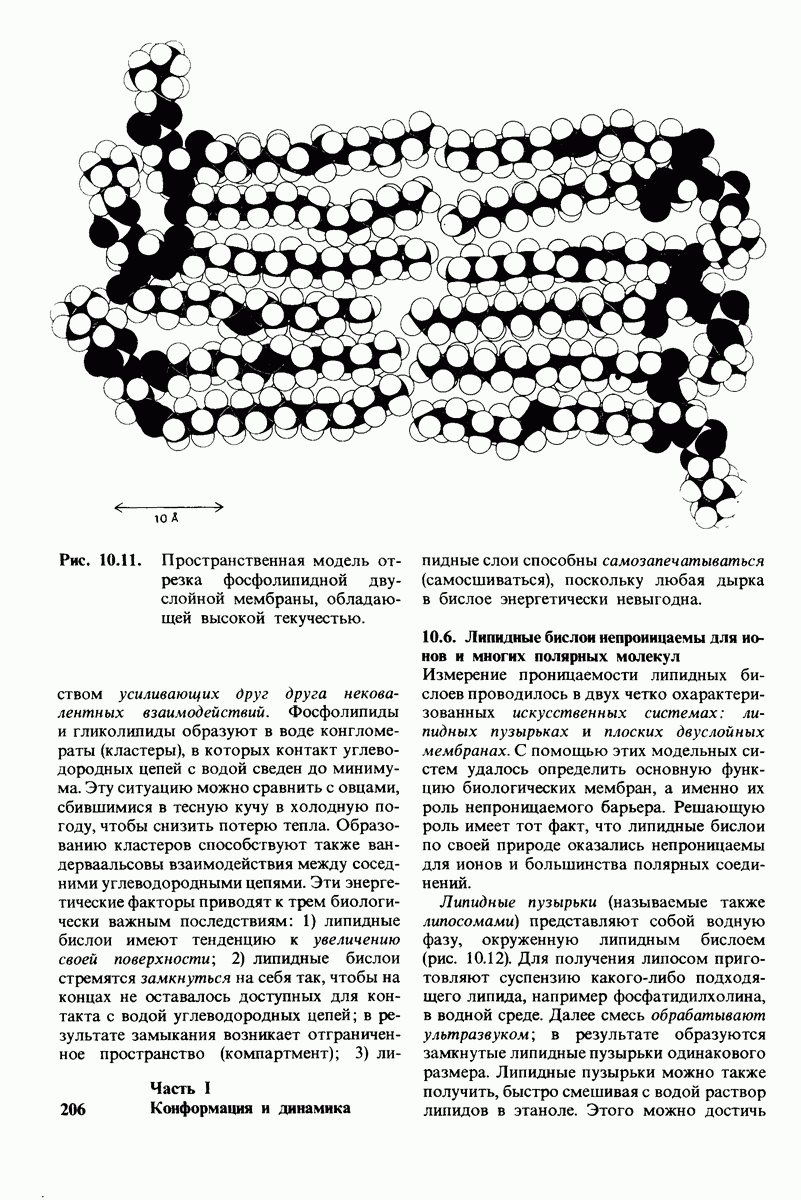
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше
Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике
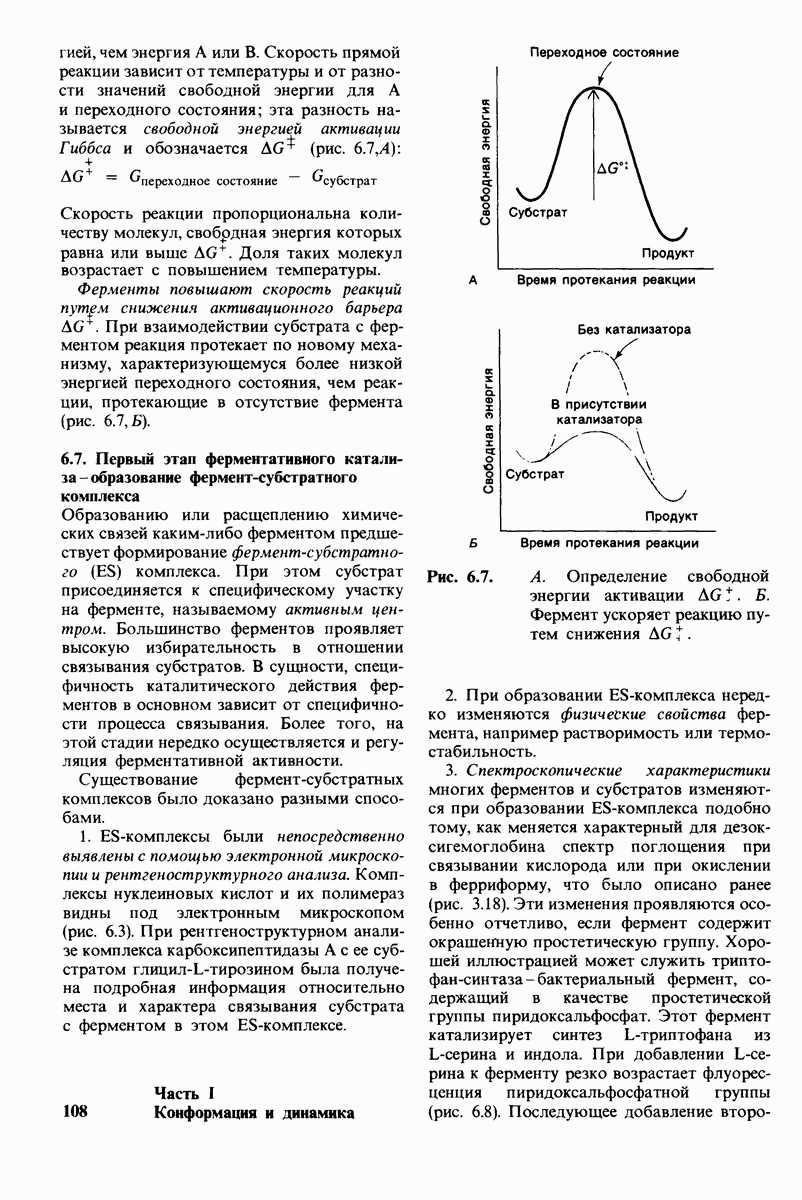
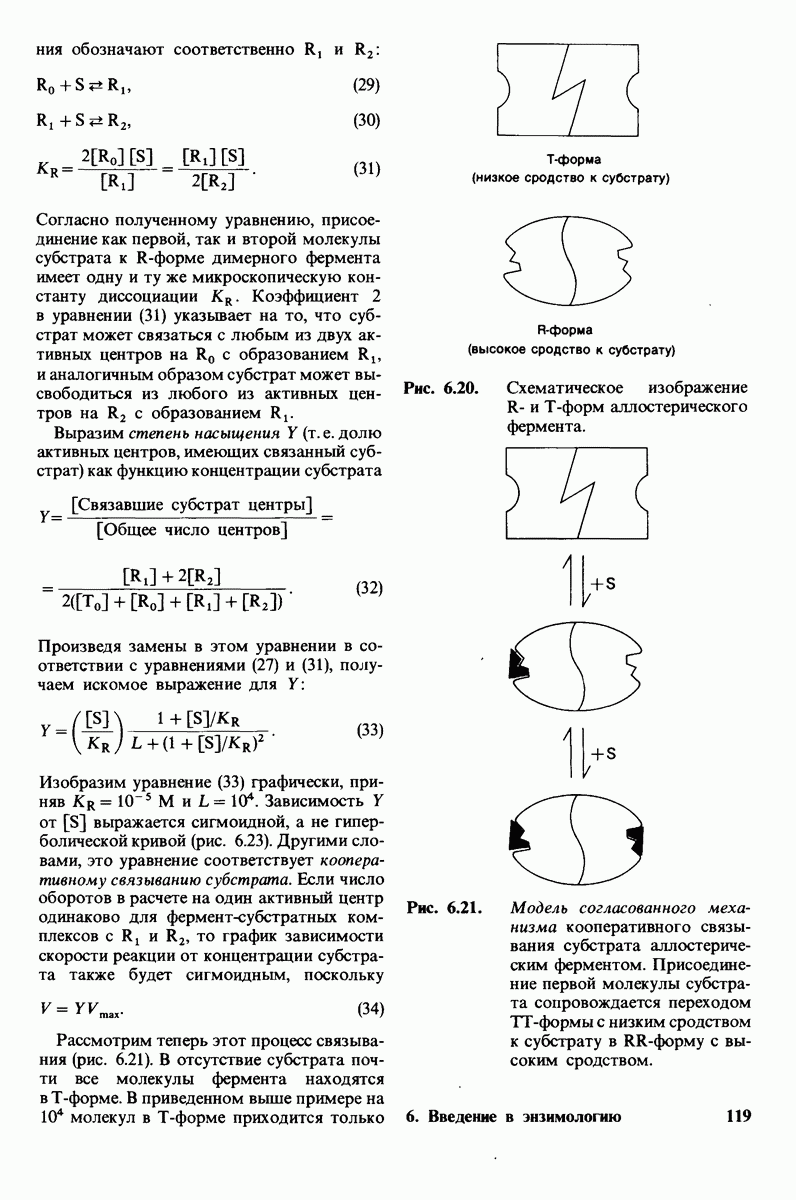
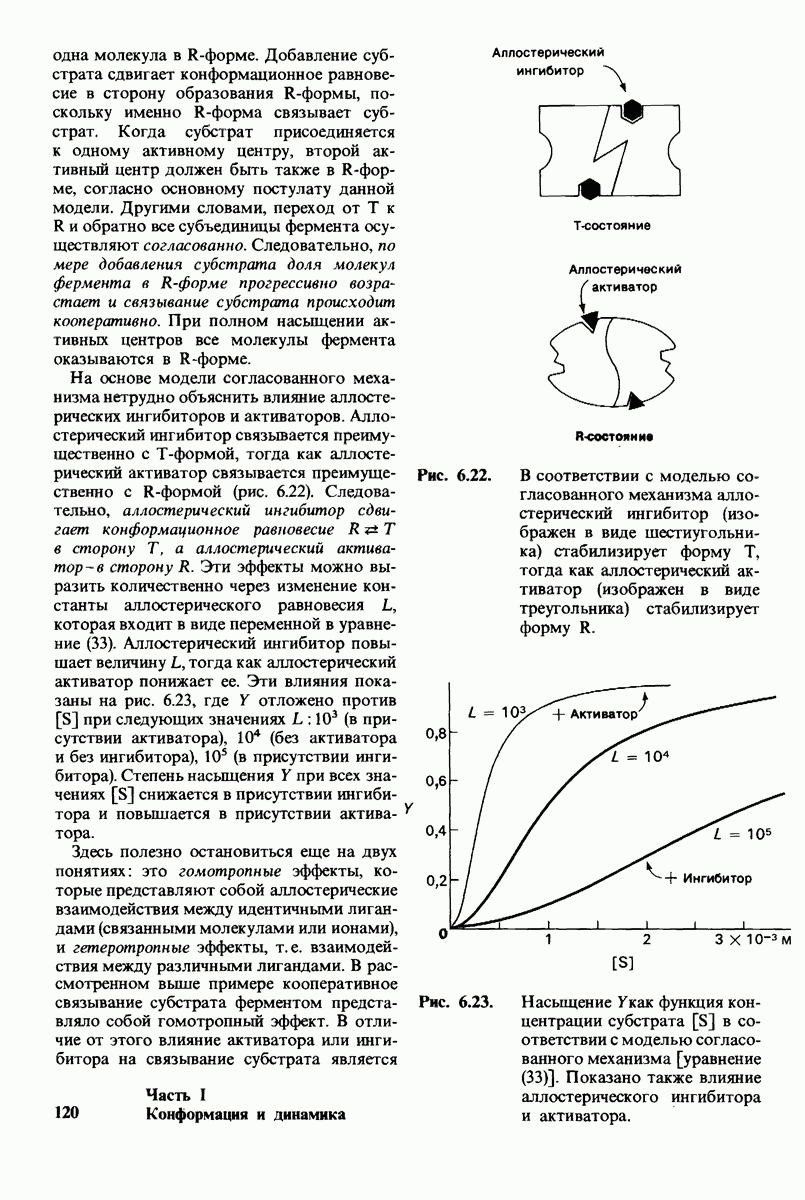
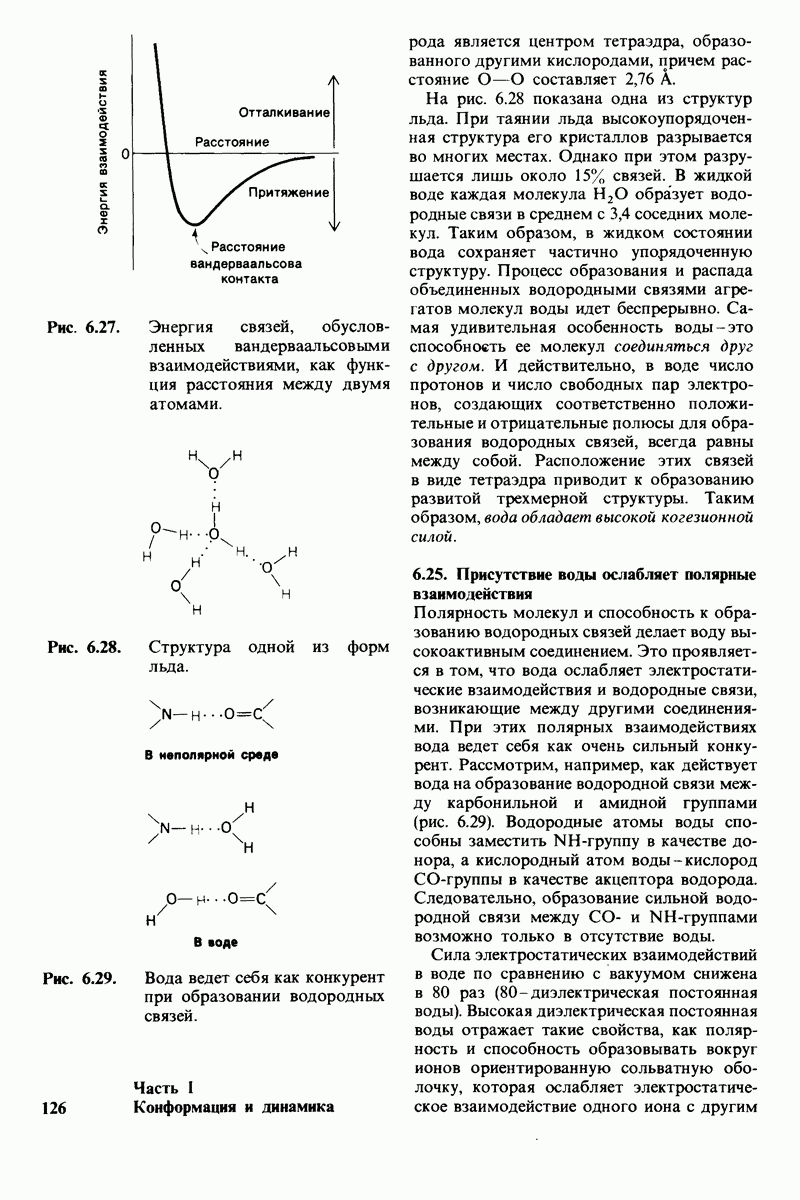
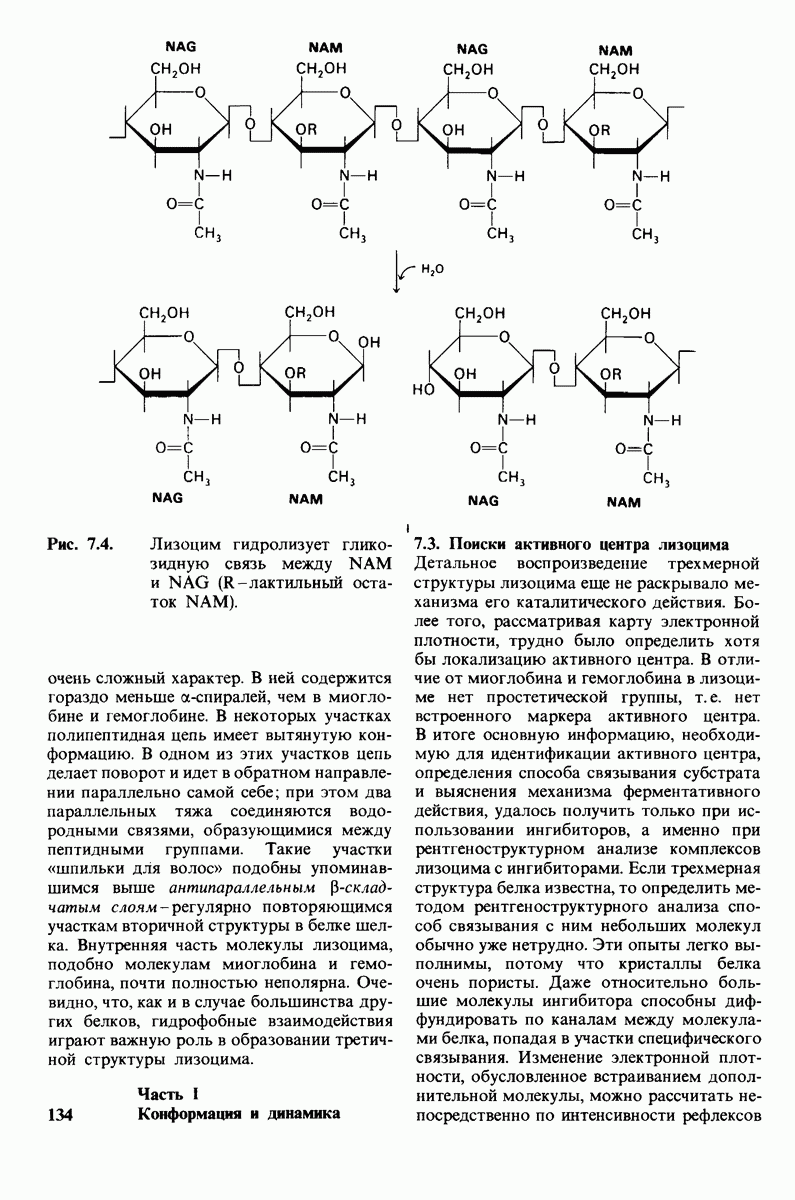
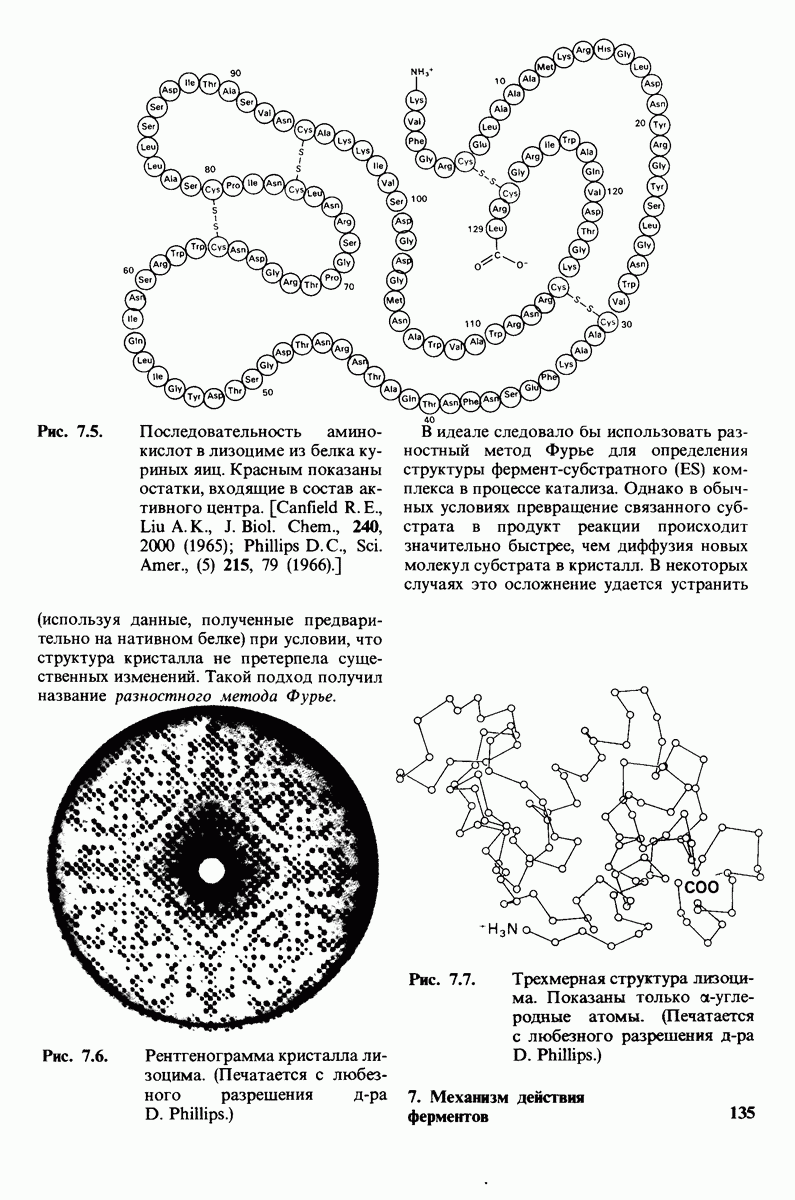
7.5. От структуры фермента — к механизму ферментативного действия
1. Как происходит связывание субстрата? Мы уже говорили о том, что метод рентгеноструктурного анализа не дает возможности непосредственно определить, как происходит связывание с ферментом эффективного субстрата. Однако данные, полученные при рентгеноструктурном анализе комплекса фермент — конкурентный ингибитор, могут сыграть ключевую роль в решении этой

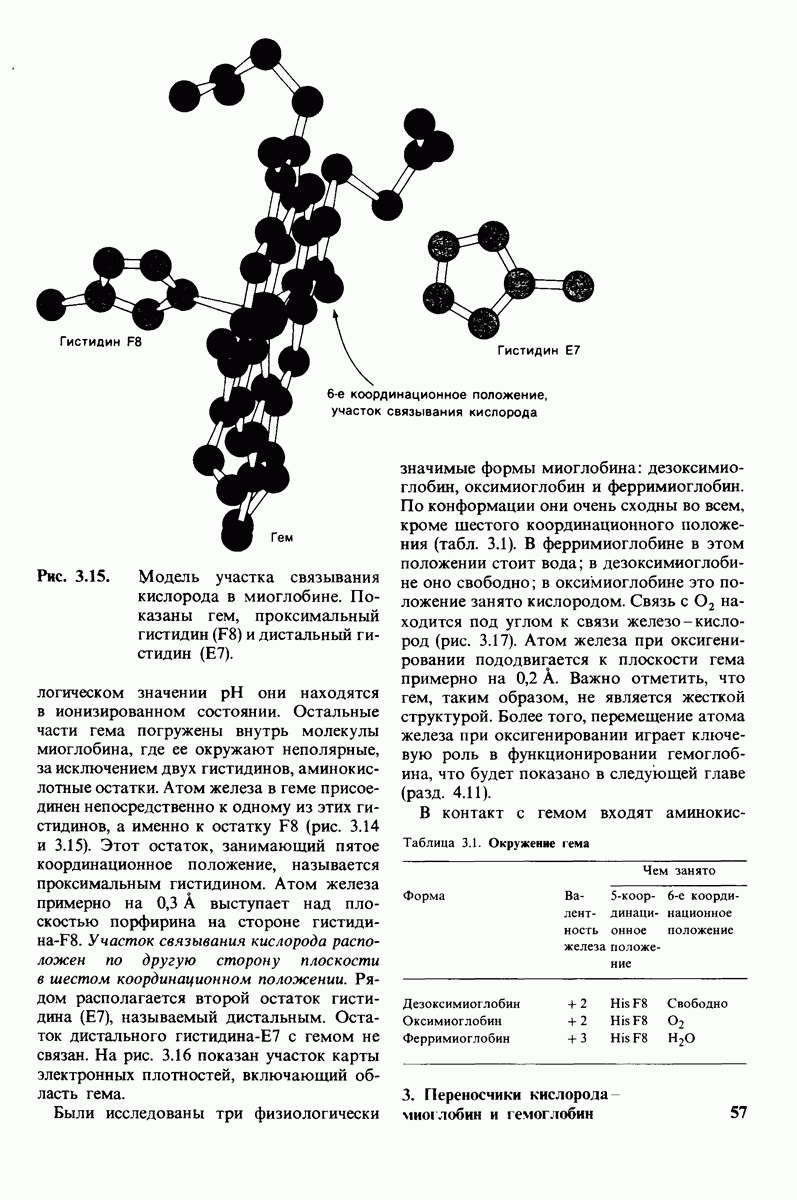
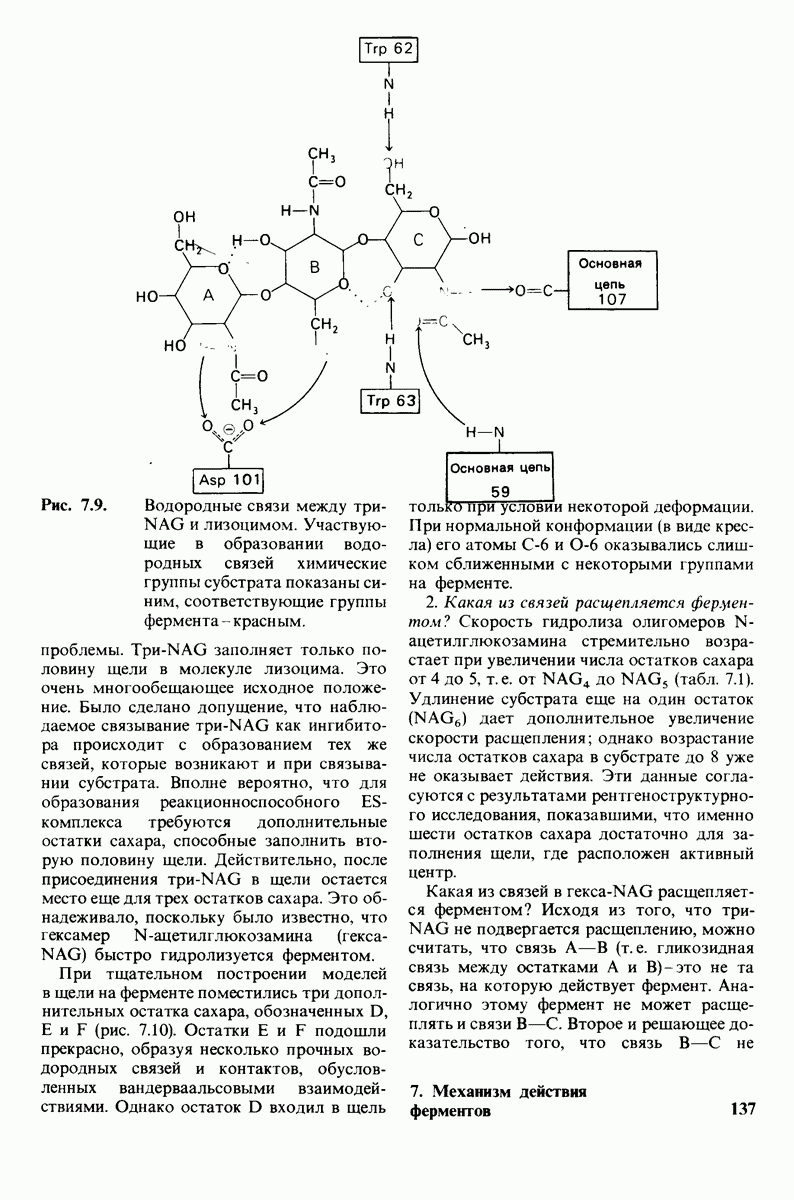
Рис. 7.9.
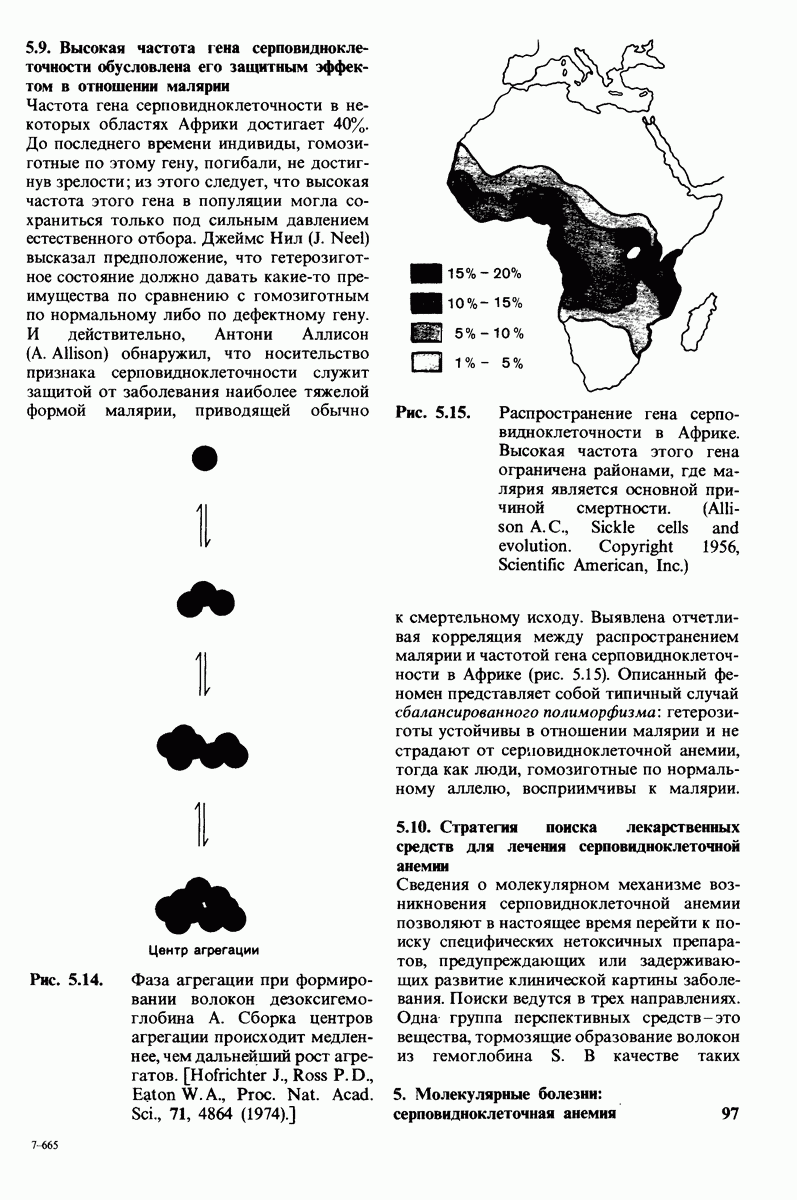
Водородные связи между три-NAG и лизоцимом. Участвующие в образовании водородных связей химические группы субстрата показаны синим, соответствующие группы фермента - красным.
проблемы. Три-NAG заполняет только половину щели в молекуле лизоцима. Это очень многообещающее исходное положение. Было сделано допущение, что наблюдаемое связывание три-NAG как ингибитора происходит с образованием тех же связей, которые возникают и при связывании субстрата. Вполне вероятно, что для образования реакционноспособного  -комплекса требуются дополнительные остатки сахара, способные заполнить вторую половину щели. Действительно, после присоединения три-NAG в щели остается место еще для трех остатков сахара. Это обнадеживало, поскольку было известно, что гексамер N-ацетилглюкозамина
-комплекса требуются дополнительные остатки сахара, способные заполнить вторую половину щели. Действительно, после присоединения три-NAG в щели остается место еще для трех остатков сахара. Это обнадеживало, поскольку было известно, что гексамер N-ацетилглюкозамина  быстро гидролизуется ферментом.
быстро гидролизуется ферментом.
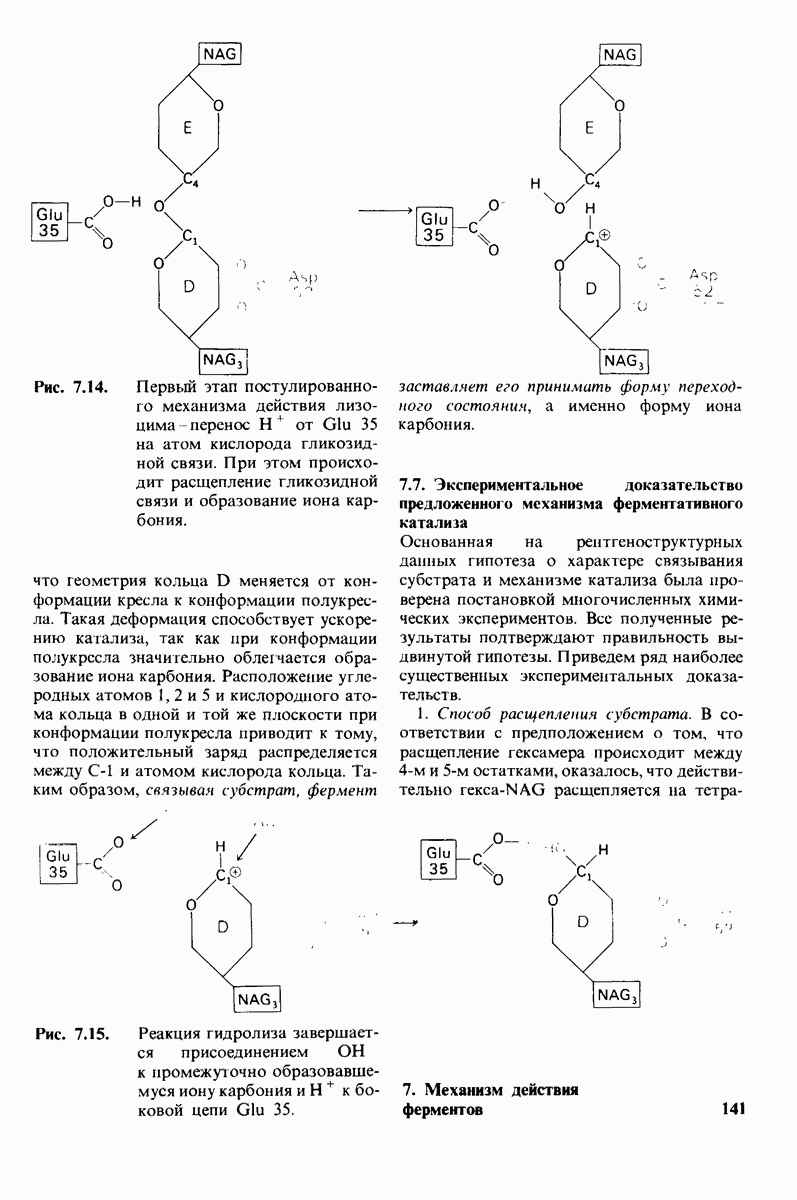
При тщательном построении моделей в щели на ферменте поместились три дополнительных остатка сахара, обозначенных  (рис. 7.10). Остатки
(рис. 7.10). Остатки  подошли прекрасно, образуя несколько прочных водородных связей и контактов, обусловленных вандерваальсовыми взаимодействиями. Однако остаток D входил в щель только при условии некоторой деформации. При нормальной конформации (в виде кресла) его атомы
подошли прекрасно, образуя несколько прочных водородных связей и контактов, обусловленных вандерваальсовыми взаимодействиями. Однако остаток D входил в щель только при условии некоторой деформации. При нормальной конформации (в виде кресла) его атомы  оказывались слишком сближенными с некоторыми группами на ферменте.
оказывались слишком сближенными с некоторыми группами на ферменте.
2. Какая из связей расщепляется ферментом? Скорость гидролиза олигомеров N-ацетилглюкозамина стремительно возрастает при увеличении числа остатков сахара от 4 до 5, т.е. от  до
до  (табл. 7.1). Удлинение субстрата еще на один остаток
(табл. 7.1). Удлинение субстрата еще на один остаток  дает дополнительное увеличение скорости расщепления; однако возрастание числа остатков сахара в субстрате до 8 уже не оказывает действия. Эти данные согласуются с результатами рентгеноструктурного исследования, показавшими, что именно шести остатков сахара достаточно для заполнения щели, где расположен активный центр.
дает дополнительное увеличение скорости расщепления; однако возрастание числа остатков сахара в субстрате до 8 уже не оказывает действия. Эти данные согласуются с результатами рентгеноструктурного исследования, показавшими, что именно шести остатков сахара достаточно для заполнения щели, где расположен активный центр.
Какая из связей в  расщепляется ферментом? Исходя из того, что три-NAG не подвергается расщеплению, можно считать, что связь
расщепляется ферментом? Исходя из того, что три-NAG не подвергается расщеплению, можно считать, что связь  (т.е. гликозидная связь между остатками
(т.е. гликозидная связь между остатками  - это не та связь, на которую действует фермент. Аналогично этому фермент не может расщеплять и связи
- это не та связь, на которую действует фермент. Аналогично этому фермент не может расщеплять и связи  Второе и решающее доказательство того, что связь
Второе и решающее доказательство того, что связь  не
не

Рис. 7.10. Способ связывания гекса-NAG (показан желтым) с лизоцимом. Расположение углеводных остатков  (слева) соответствует локализации три-NAG в комплексе с лизоцимом; расположение остатков
(слева) соответствует локализации три-NAG в комплексе с лизоцимом; расположение остатков  (справа) предсказали путем модельного построения. Зеленым показаны два аминокислотных остатка, непосредственно участвующих в катализе.
(справа) предсказали путем модельного построения. Зеленым показаны два аминокислотных остатка, непосредственно участвующих в катализе.
расщепляется ферментом, состоит в том, что NAM не может встать в положение остатка С. Если  отлично умещается в участке С, то NAM здесь не умещается из-за лактильной боковой цепи. Между тем в полисахариде клеточной стенки бактерий лизоцим расщепляет связь
отлично умещается в участке С, то NAM здесь не умещается из-за лактильной боковой цепи. Между тем в полисахариде клеточной стенки бактерий лизоцим расщепляет связь  Следовательно, и связь
Следовательно, и связь  не может подвергаться расщеплению, если действительно полисахарид клеточных стенок бактерий связывается с лизоцимом таким же образом, как и гекса-NAG. Несоответствие NAM положению С исключает еще одно место гидролиза, а именно связь
не может подвергаться расщеплению, если действительно полисахарид клеточных стенок бактерий связывается с лизоцимом таким же образом, как и гекса-NAG. Несоответствие NAM положению С исключает еще одно место гидролиза, а именно связь  Вспомним, что полисахарид клеточной стенки - это чередующийся полимер NAM и
Вспомним, что полисахарид клеточной стенки - это чередующийся полимер NAM и  следовательно, если NAM не может занимать положение С, то этот остаток не может стоять и в положении
следовательно, если NAM не может занимать положение С, то этот остаток не может стоять и в положении 
Из приведенных соображений вытекает, что при расщеплении ферментом гексамерного субстрата связи  и
и  не могут разрываться. Следовательно, единственным возможным местом расщепления субстрата является связь
не могут разрываться. Следовательно, единственным возможным местом расщепления субстрата является связь  (рис. 7.11).
(рис. 7.11).
3. Какая группа на ферменте непосредственно осуществляет катализ? Заключение о том, что гидролиз субстрата происходит по связи  позволило перейти
позволило перейти
Таблица 7.1. (см. скан) Эффективность олнгомеров N-ацетнлглю-козамина в качестве субстратов

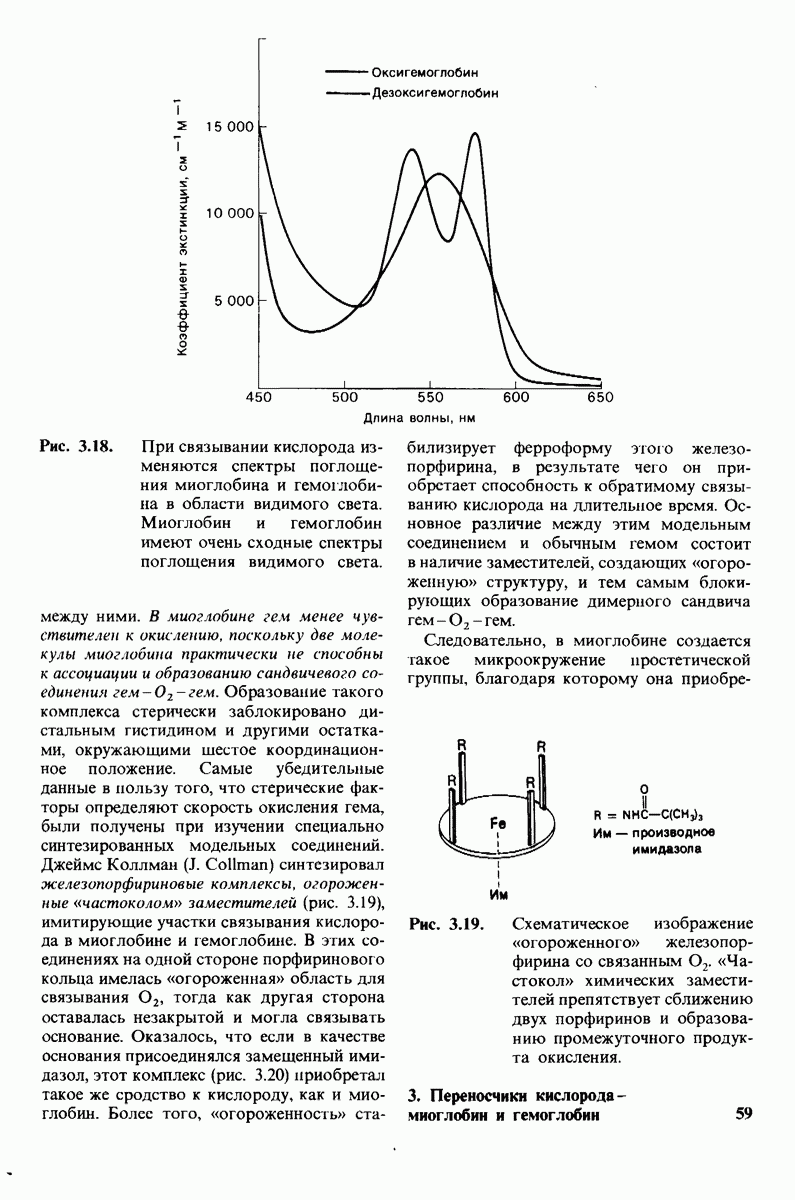
Рис. 7.11
Ход рассуждений, доказывающих, что место приложения действия фермента - гликозидная связь между углеводными остатками 
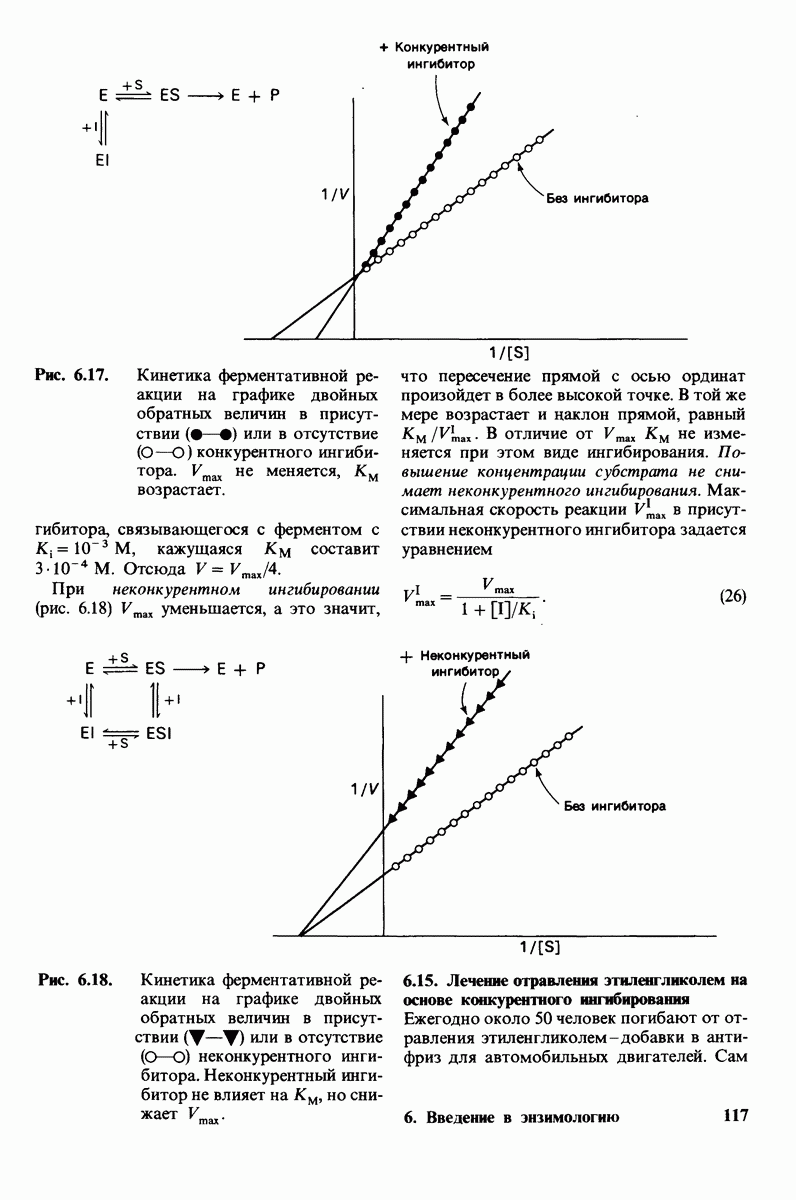
далее к выявлению тех групп на ферменте, которые непосредственно осуществляют реакцию гидролиза. Для этого, однако, нужно еще более точно локализовать место расщепления субстрата, а именно выяснить, по какую сторону от гликозидного атома кислорода происходит разрыв связи. Ответ на этот вопрос был получен в опытах по ферментативному гидролизу в среде, содержащей воду, меченную стабильным тяжелым атомом кислорода 180 (рис. 7.12). В выделенных по окончании гидролиза сахарах 180 оказался присоединенным к  остатка
остатка  тогда как гидроксильная группа при
тогда как гидроксильная группа при  в остатке
в остатке  содержала обычный изотоп кислорода. Отсюда следует, что при гидролизе разрыв связи происходит между
содержала обычный изотоп кислорода. Отсюда следует, что при гидролизе разрыв связи происходит между  остатка D и кислородом гликозидной связи, примыкающим к остатку
остатка D и кислородом гликозидной связи, примыкающим к остатку  Эта работа может служить примером использования изотопов в изучении механизма ферментативного катализа. Без изотопов было бы крайне трудно, а может быть, и невозможно в данном случае установить точное место приложения действия фермента.
Эта работа может служить примером использования изотопов в изучении механизма ферментативного катализа. Без изотопов было бы крайне трудно, а может быть, и невозможно в данном случае установить точное место приложения действия фермента.
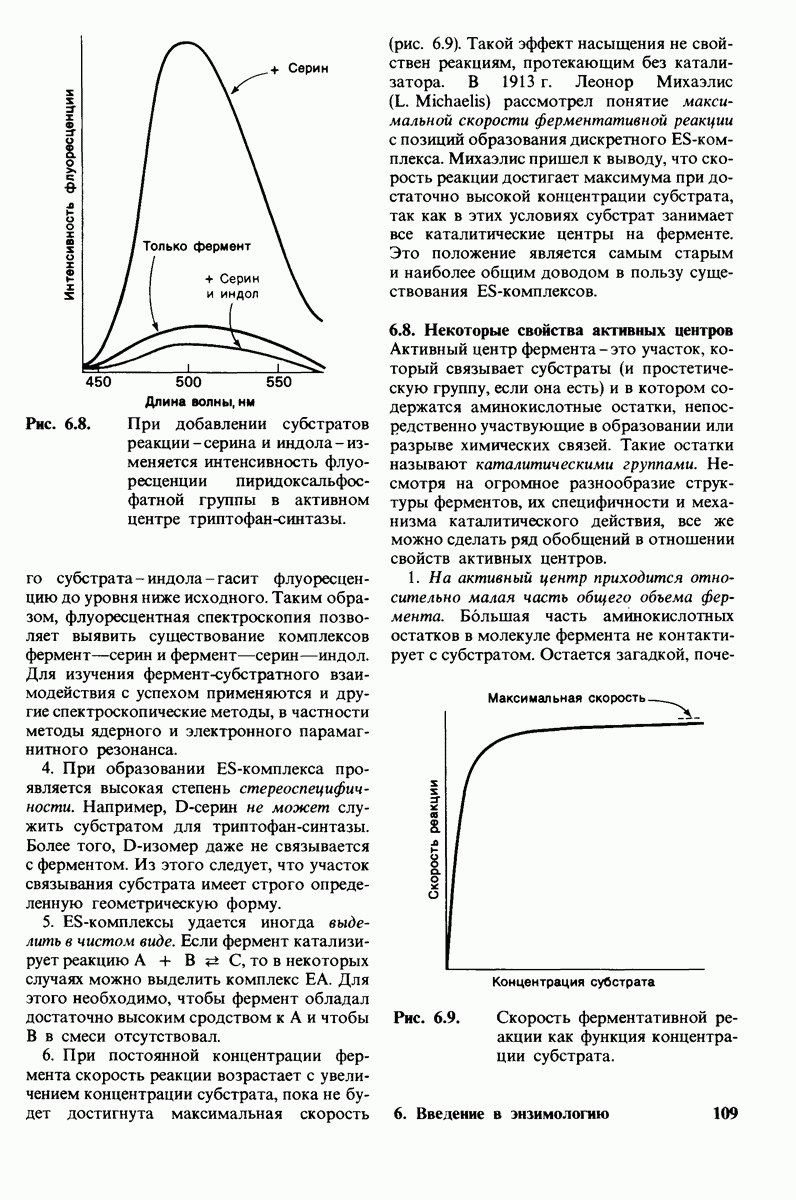
Затем перешли к поиску возможных каталитических групп, которые должны располагаться вблизи расщепляемой гликозидной связи. Как указывалось в предыдущей главе, под каталитическими группами подразумевают те группы фермента, которые непосредственно участвуют в образовании или разрыве ковалентных связей. Наиболее подходящие кандидаты на эту роль - это группы, способные к образованию водородных связей в качестве доноров или акцепторов водорода. Отрыв или присоединение иона водорода - это критический этап большинства ферментативных реакций. В лизоциме единственные остатки, способные быть каталитическими и расположенные вблизи расщепляемой гликозидной связи, - это аспартат-52 и глутамат-55. Остаток аспарагиновой кислоты лежит по одну сторону от гликозидной связи, а остаток глутаминовой - по другую. Окружение этих двух кислотных боковых цепей совершенно различно. Аспартат-52 находится в полярном окружении, где он служит акцептором водорода в сложной сети водородных связей. Глутамат-35, напротив, расположен в неполярной области. Отсюда следует, что при

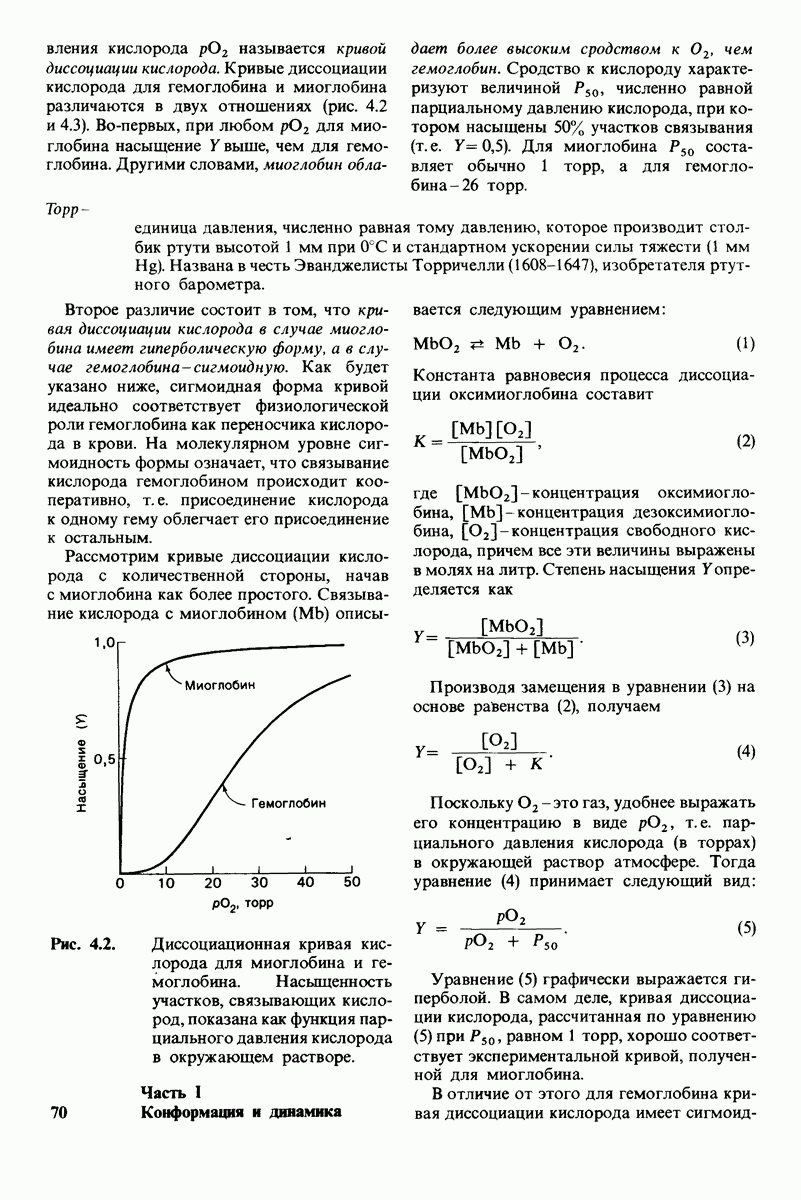
Рис. 7.12. Проведение гидролиза в меченной 180 воде показало, что лизоцим расщепляет связь между  но не связь между
но не связь между  и О (изображен только скелет остатков
и О (изображен только скелет остатков 

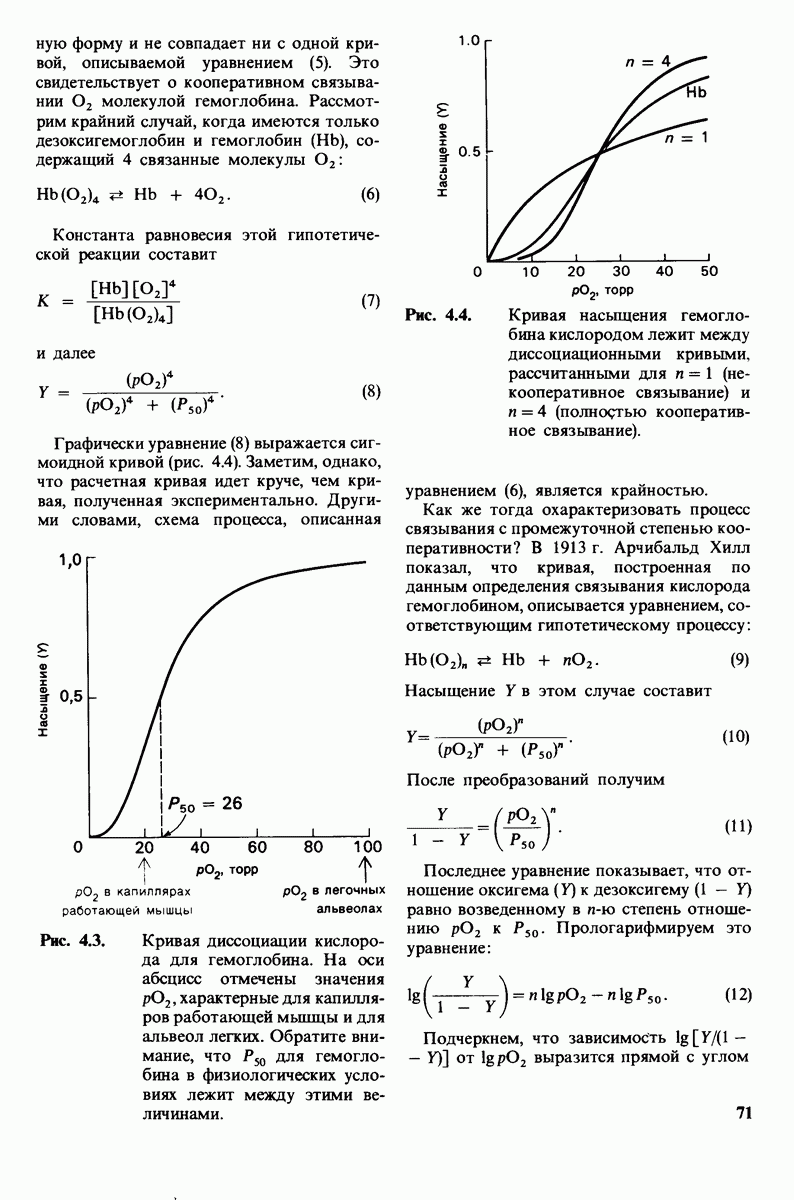
Рис. 7.13.
Структура части активного цен  лизоцима. Желтым показаны кольца
лизоцима. Желтым показаны кольца  субстрата
субстрата  Вблизи субстрата расположены боковые цепи аспартата-52 (красное) и глутамата-35 (зеленое). [Lipscomb W.N., Ргос. Robert A, Wclch Found. Conf. Chem. Res, 15, 150 (1971.)]
Вблизи субстрата расположены боковые цепи аспартата-52 (красное) и глутамата-35 (зеленое). [Lipscomb W.N., Ргос. Robert A, Wclch Found. Conf. Chem. Res, 15, 150 (1971.)]
 оптимальном рН для гидролиза лизо-цимом хитина, аспарагиновая кислота в положении 52 должна находиться в ионизированной СОО-форме, тогда как глутаминовая кислота в положении 35-в неионизированной СООН-форме. Расстояние между гликозидной связью и ближайшим к ней атомом кислорода как одной, так и другой кислотной группы составляет примерно 3 А (рис. 7.13).
оптимальном рН для гидролиза лизо-цимом хитина, аспарагиновая кислота в положении 52 должна находиться в ионизированной СОО-форме, тогда как глутаминовая кислота в положении 35-в неионизированной СООН-форме. Расстояние между гликозидной связью и ближайшим к ней атомом кислорода как одной, так и другой кислотной группы составляет примерно 3 А (рис. 7.13).