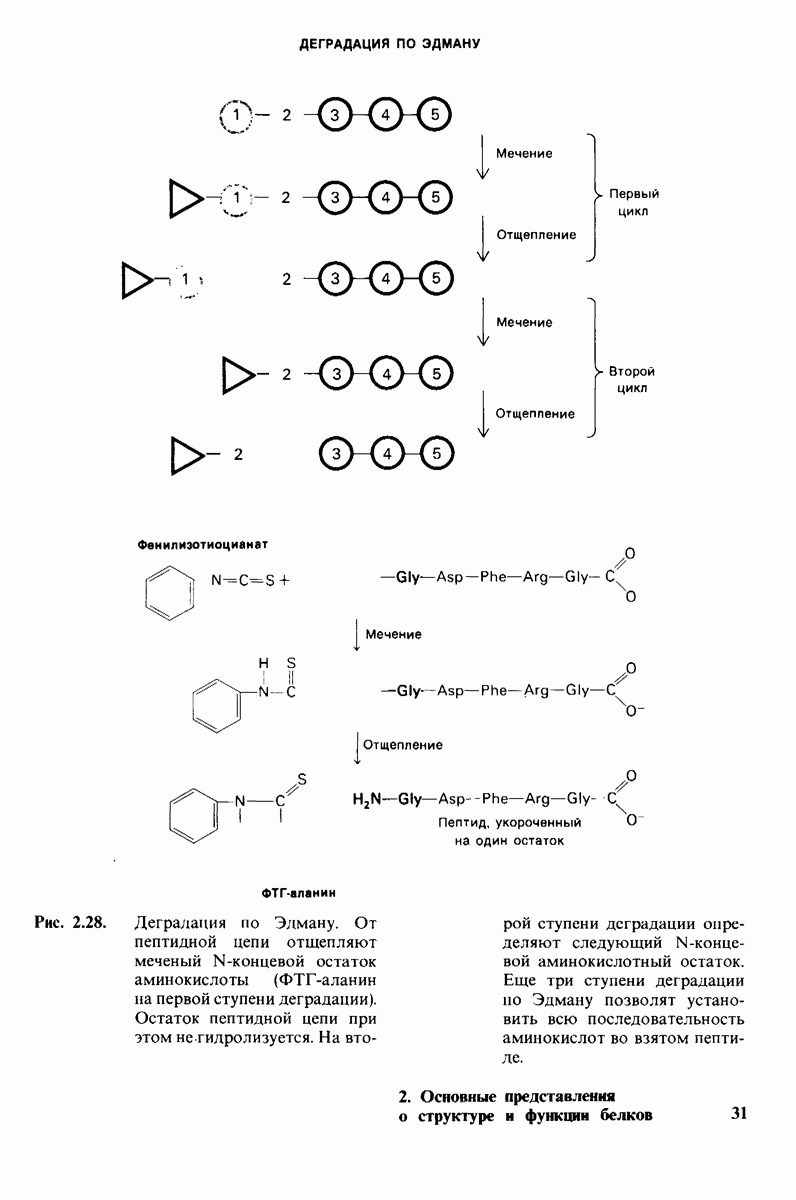
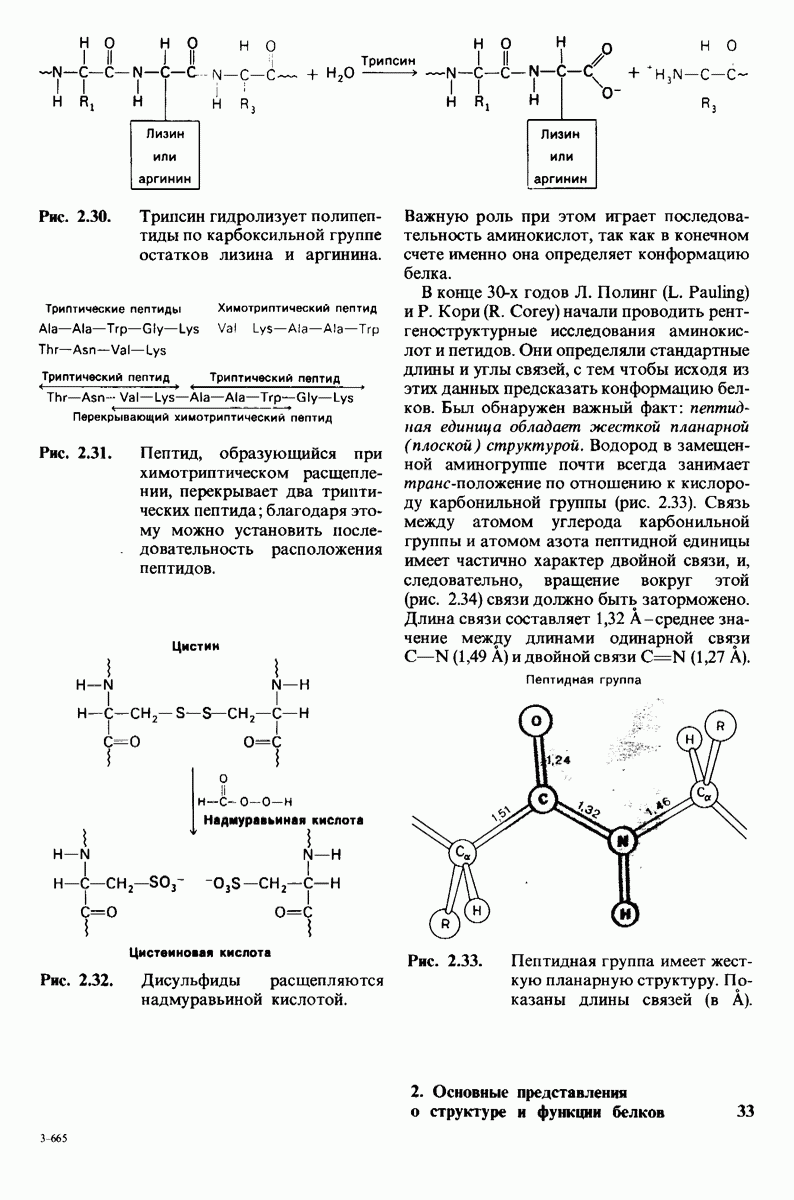
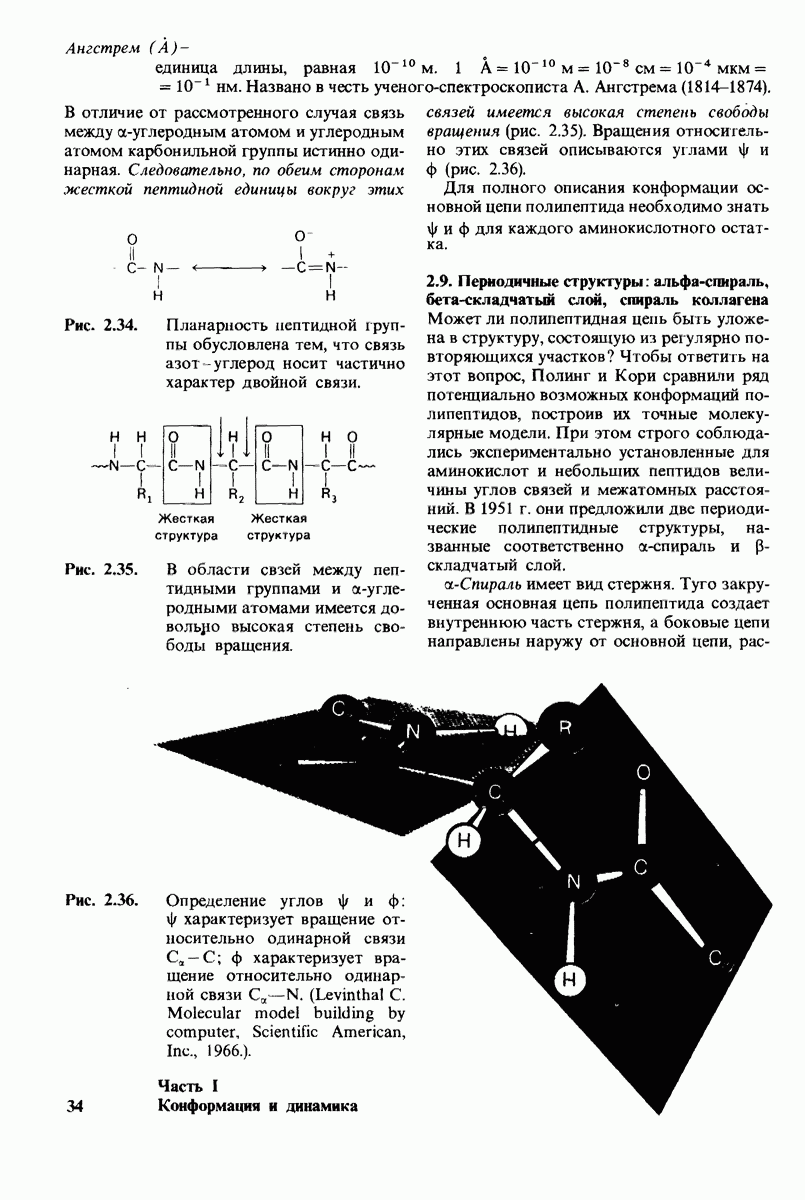
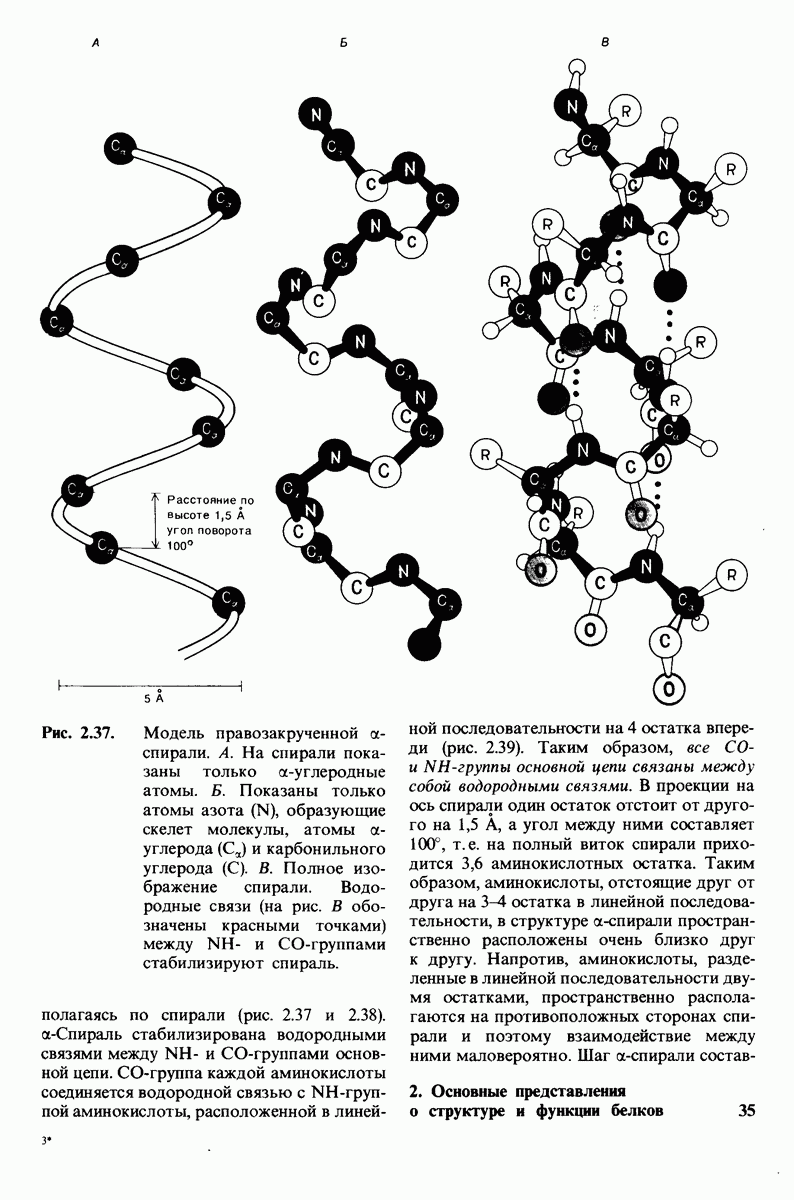
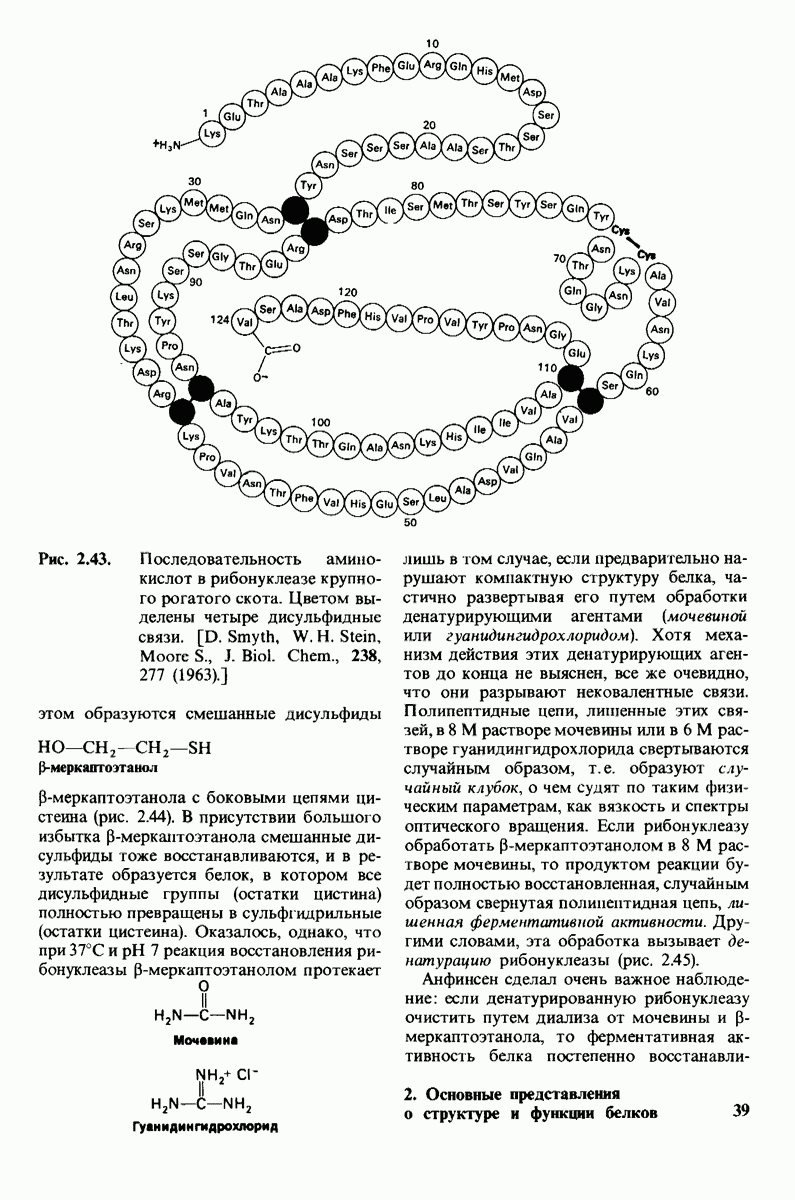
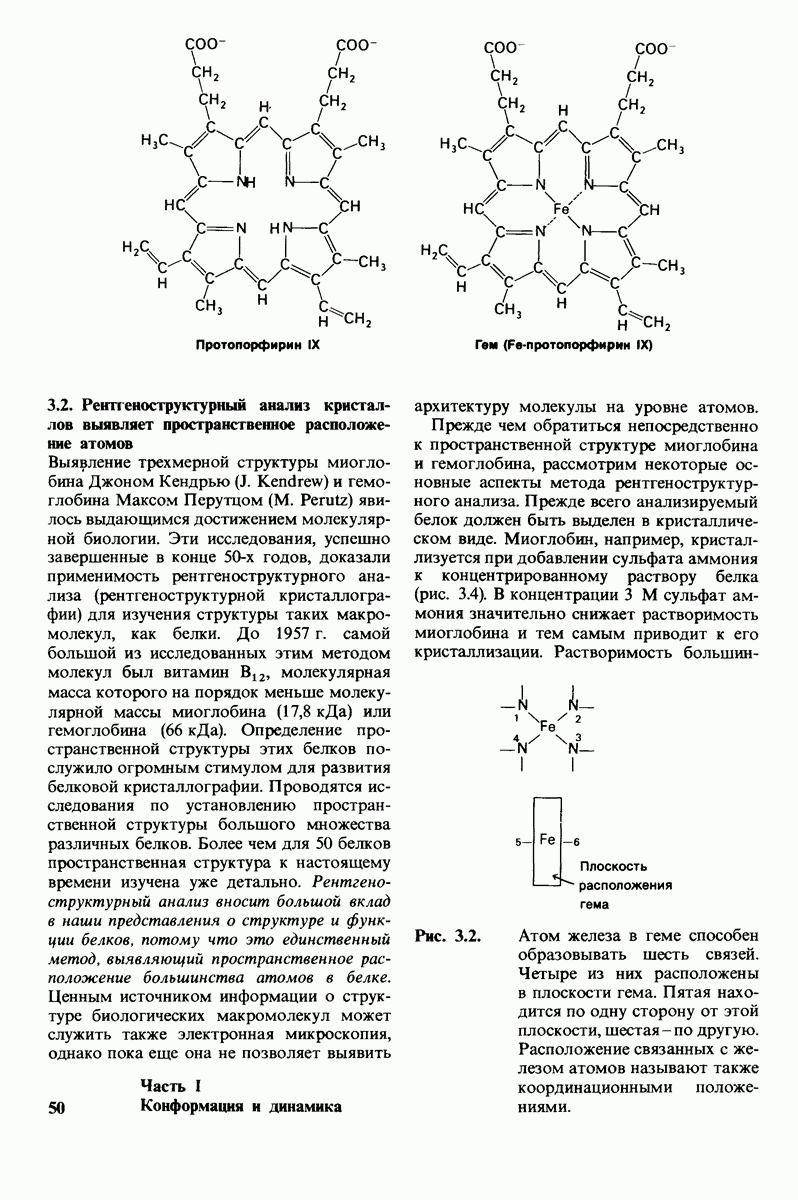
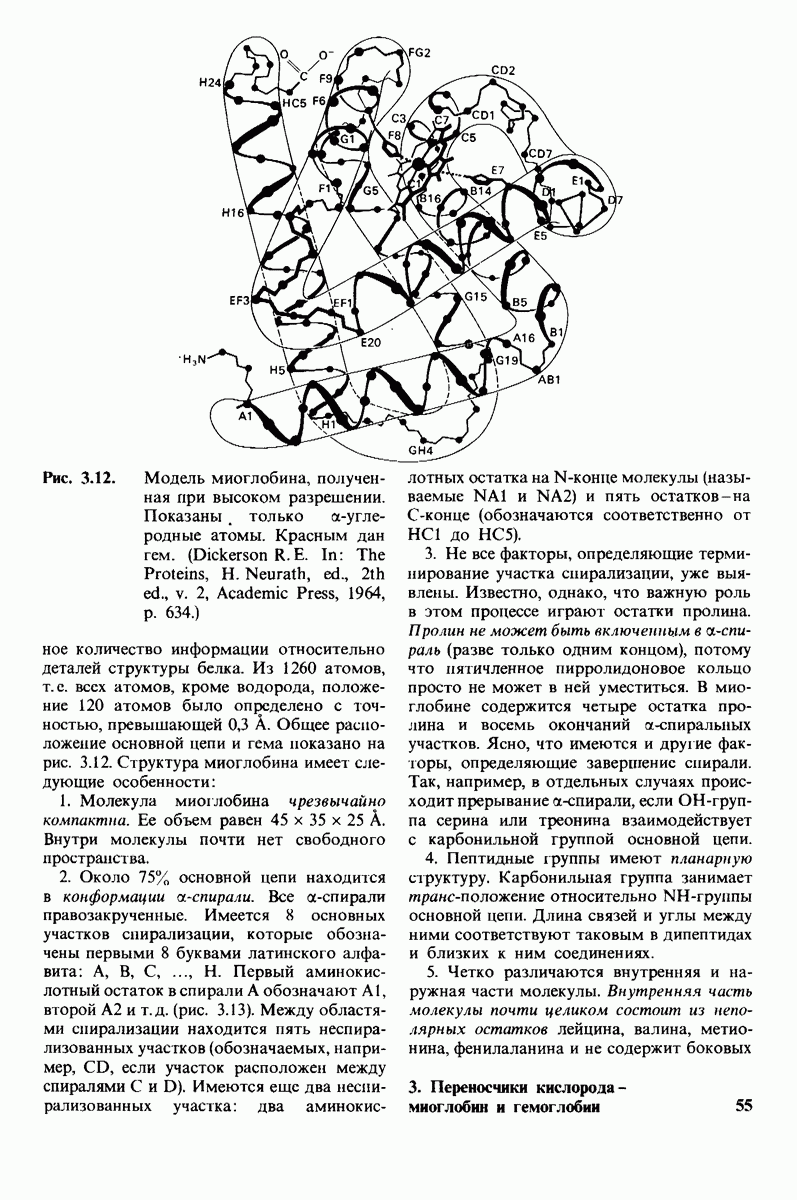
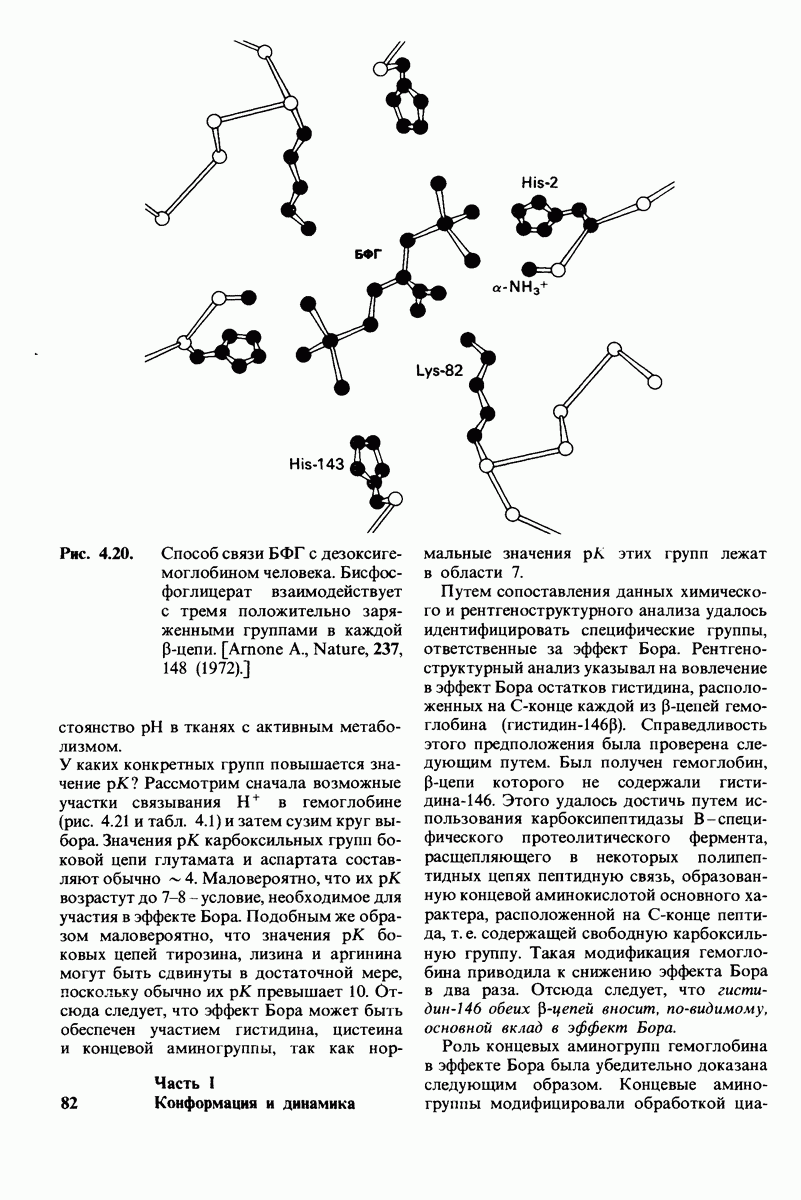
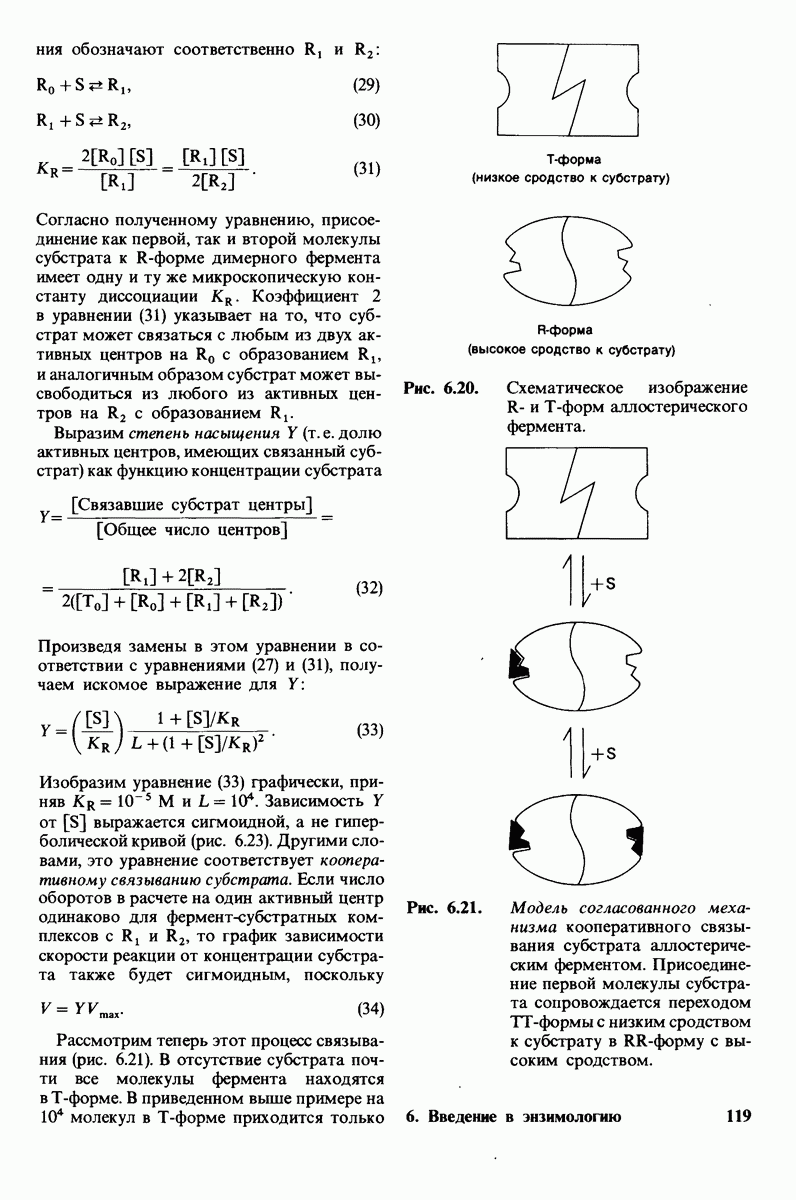
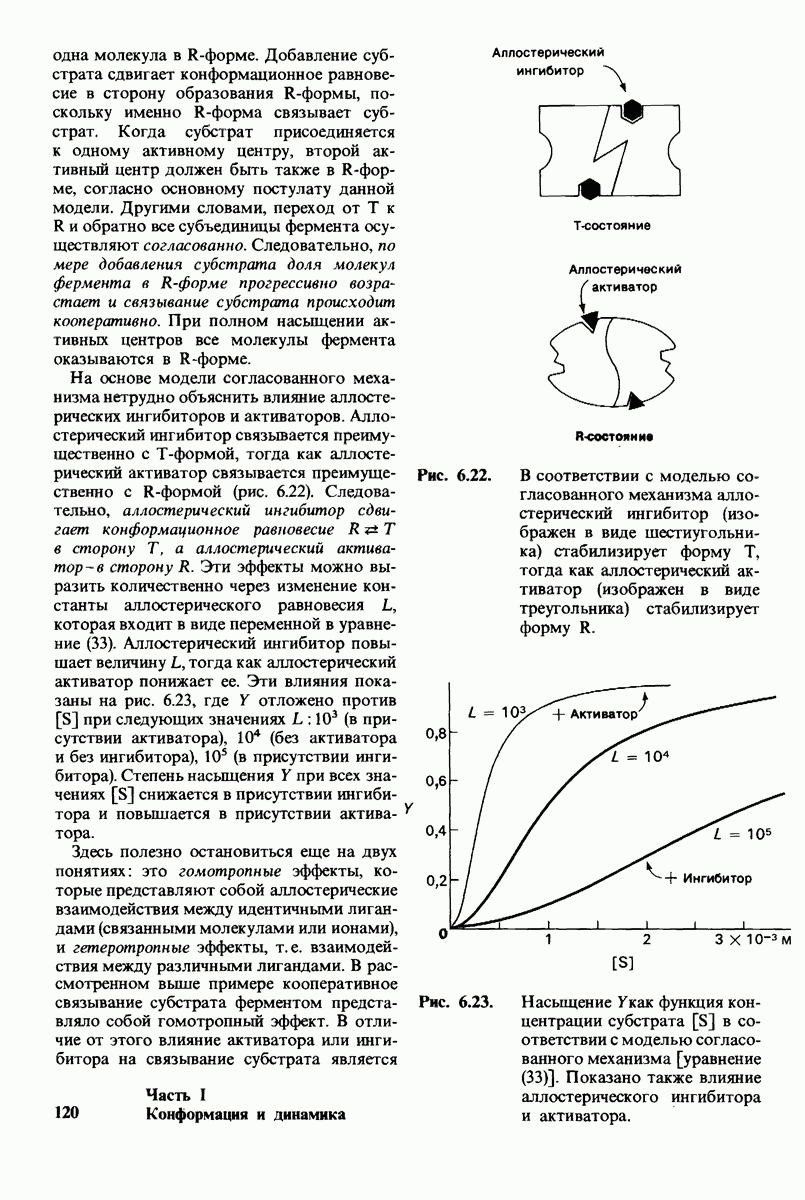
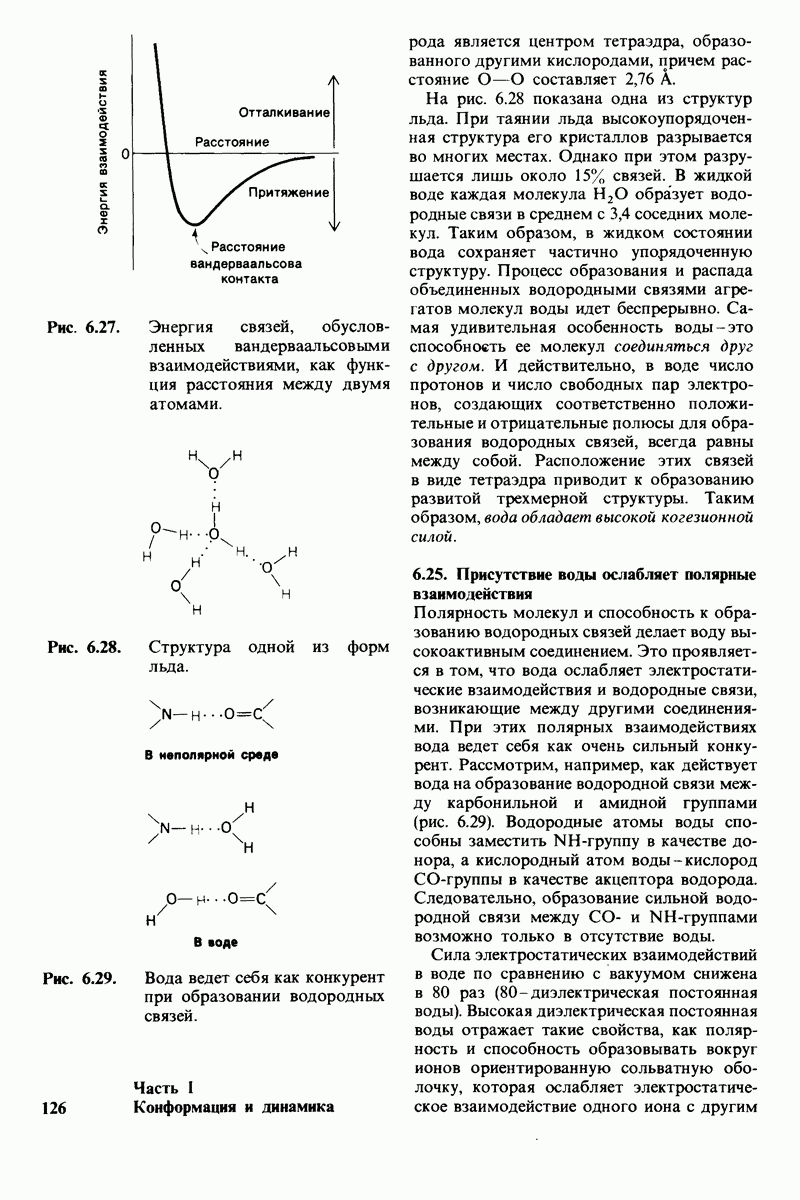
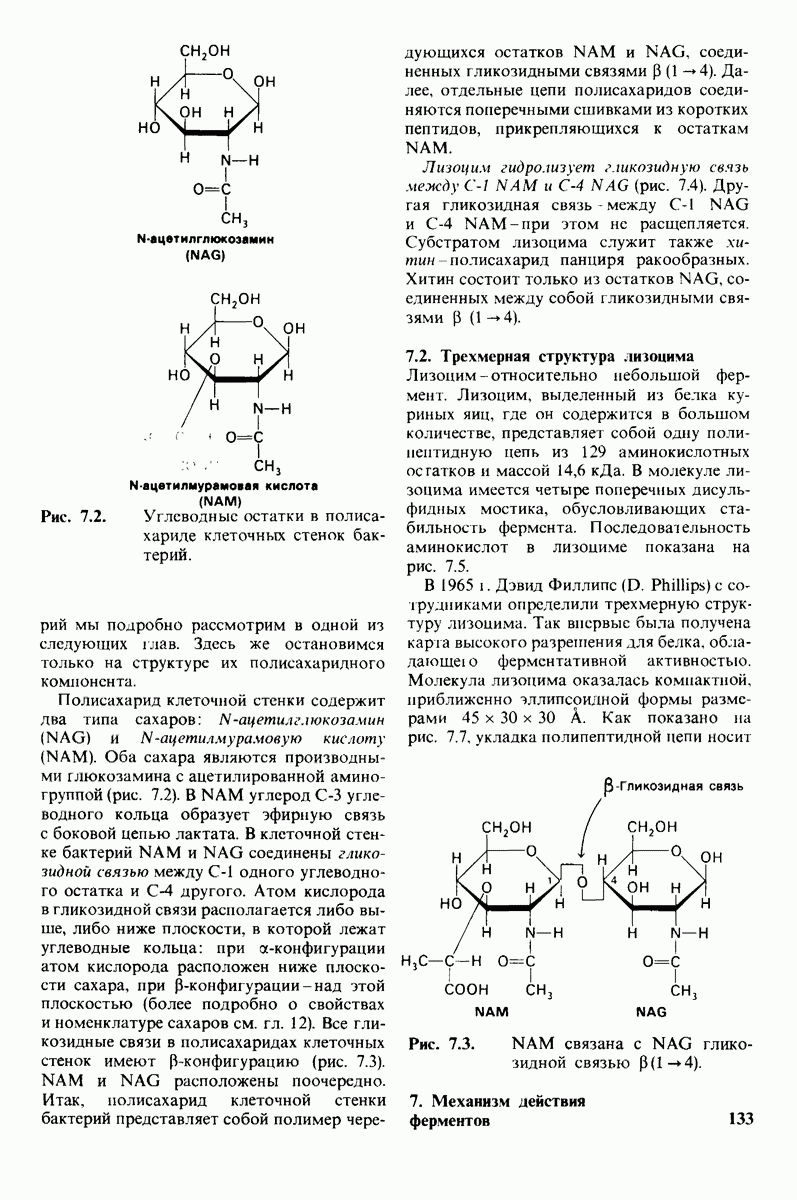
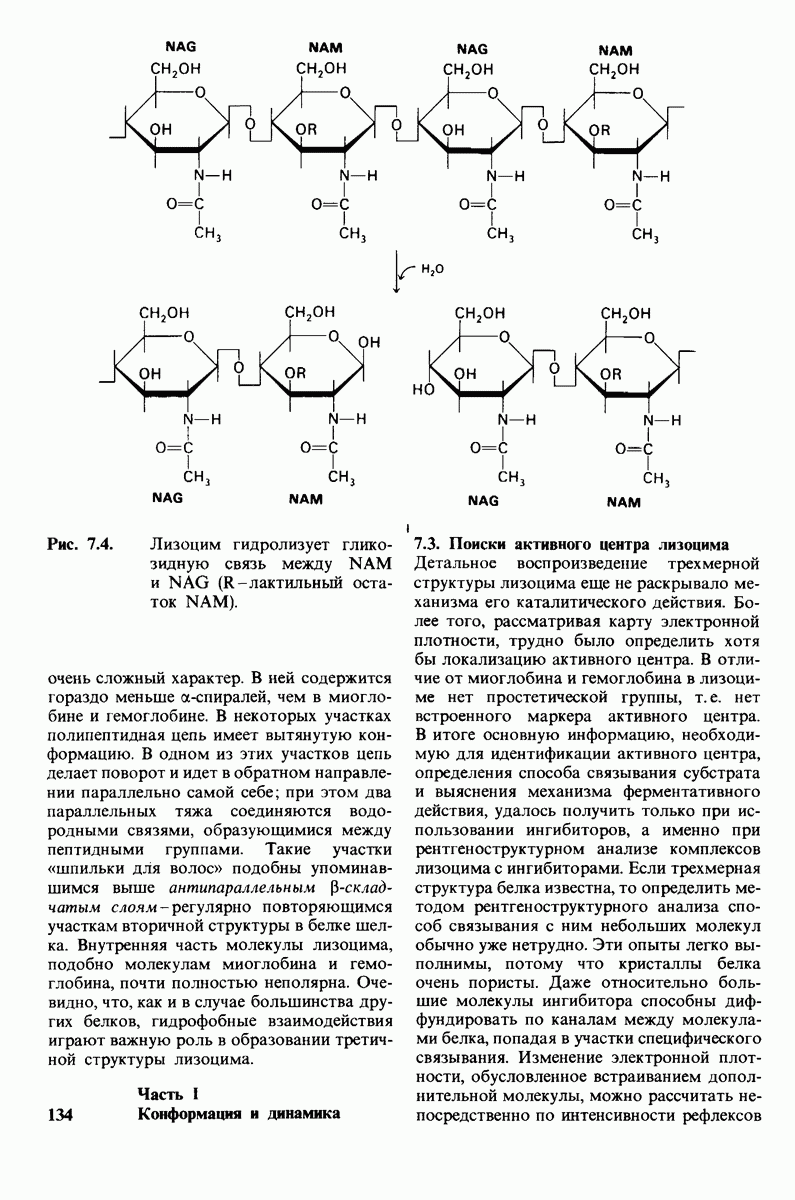
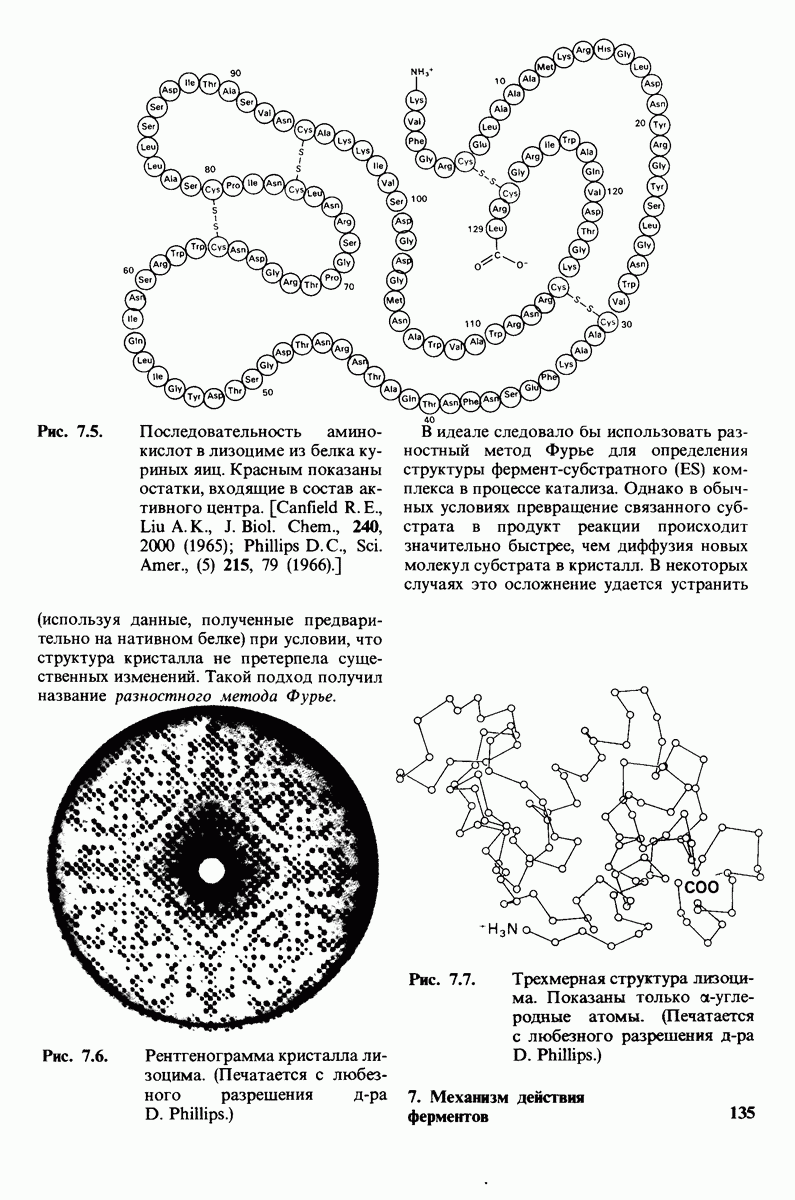
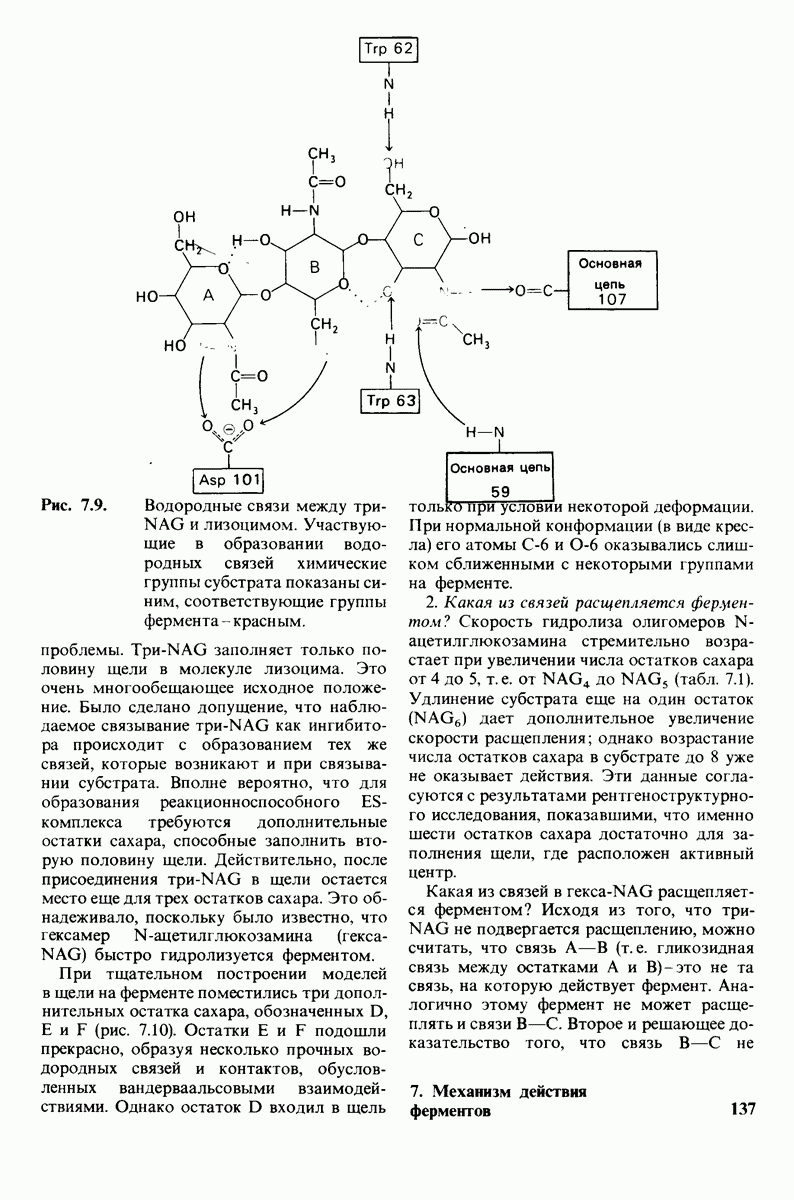
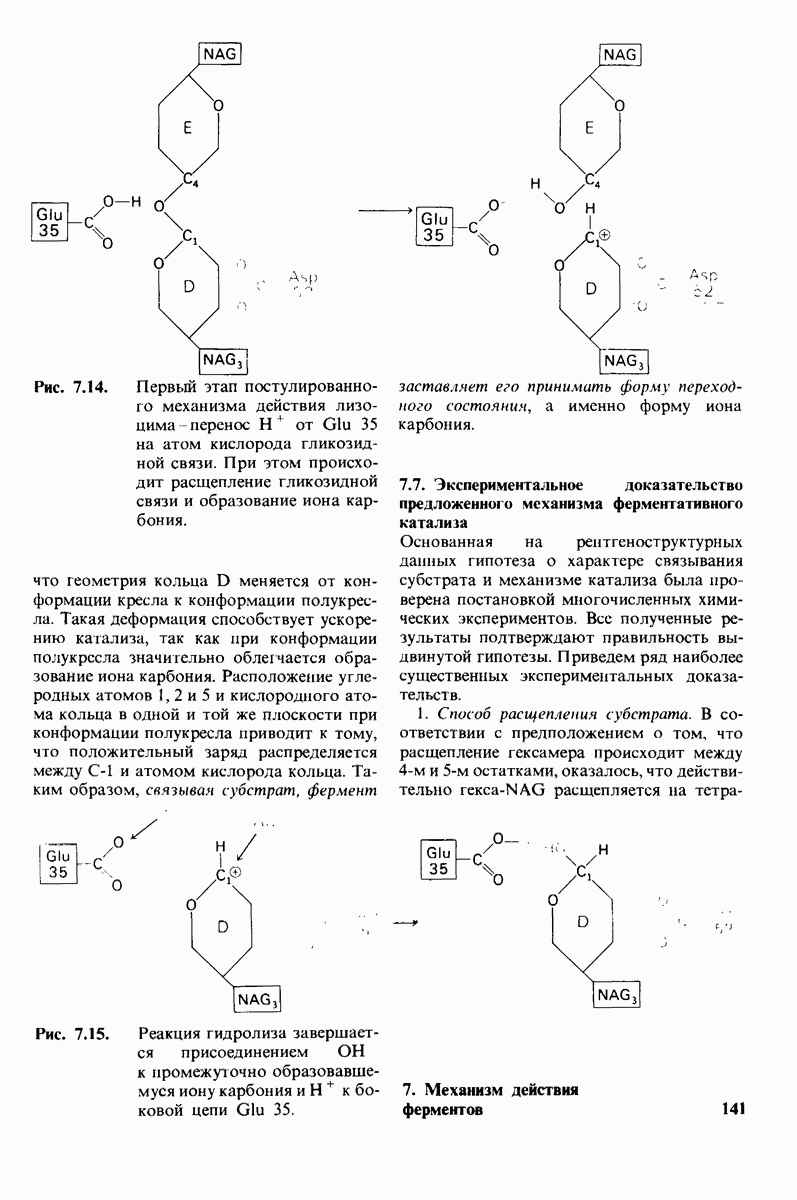
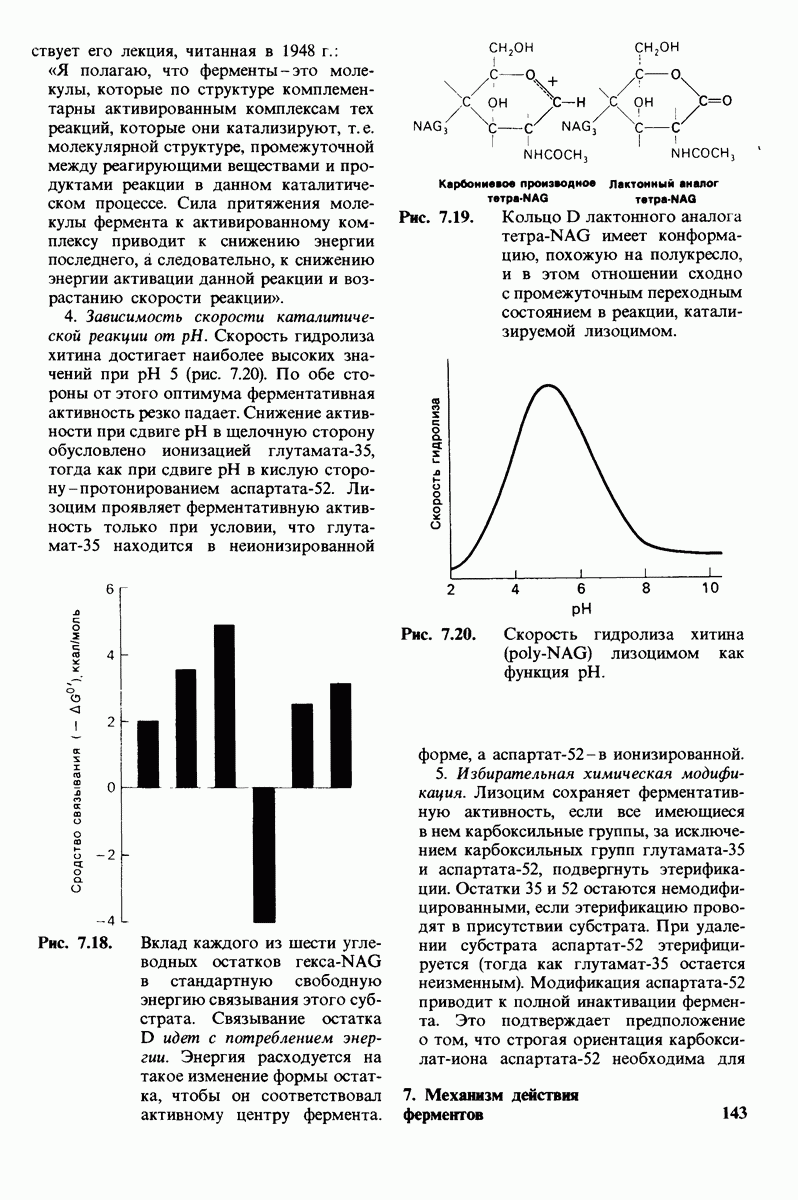
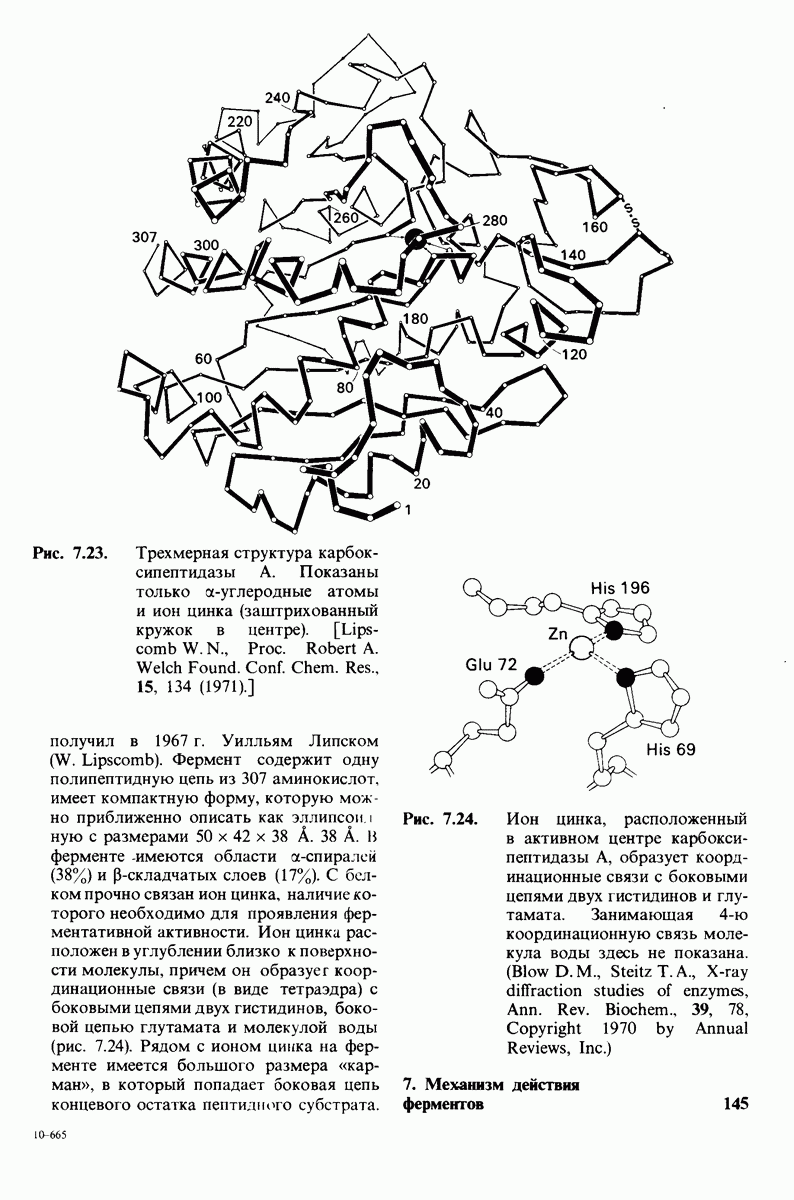
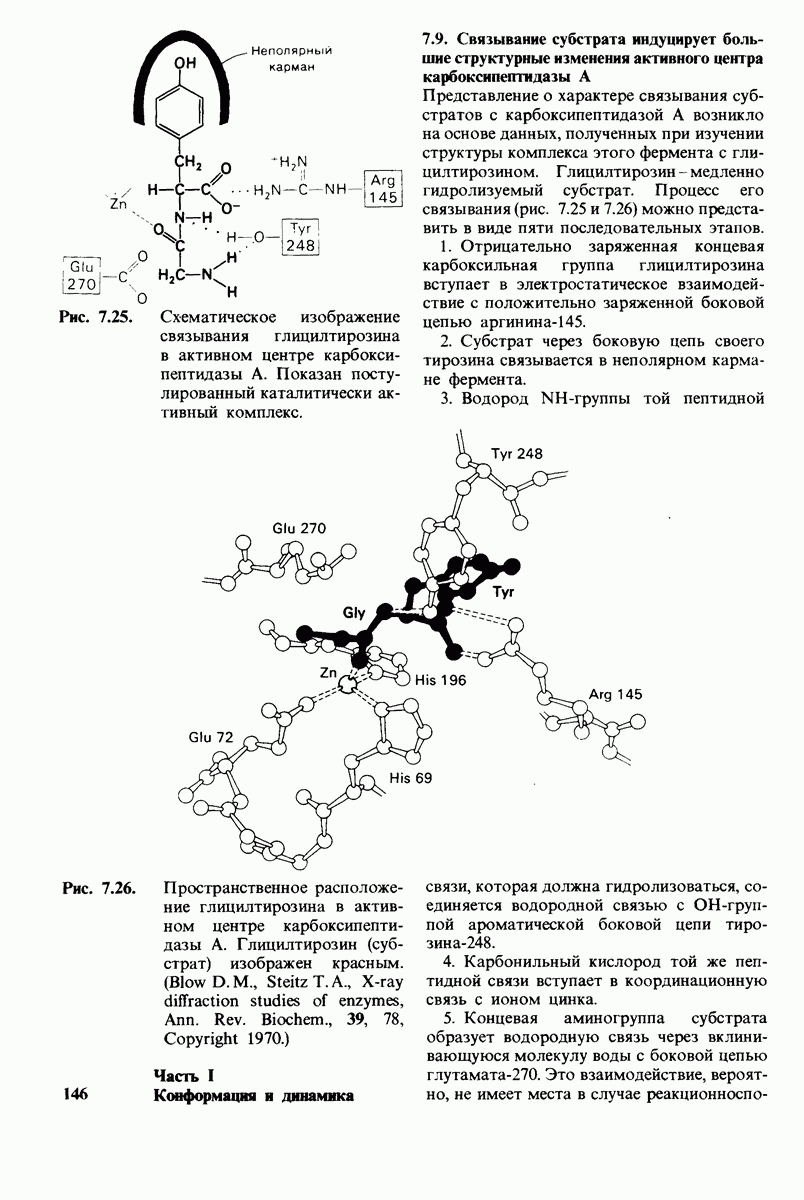
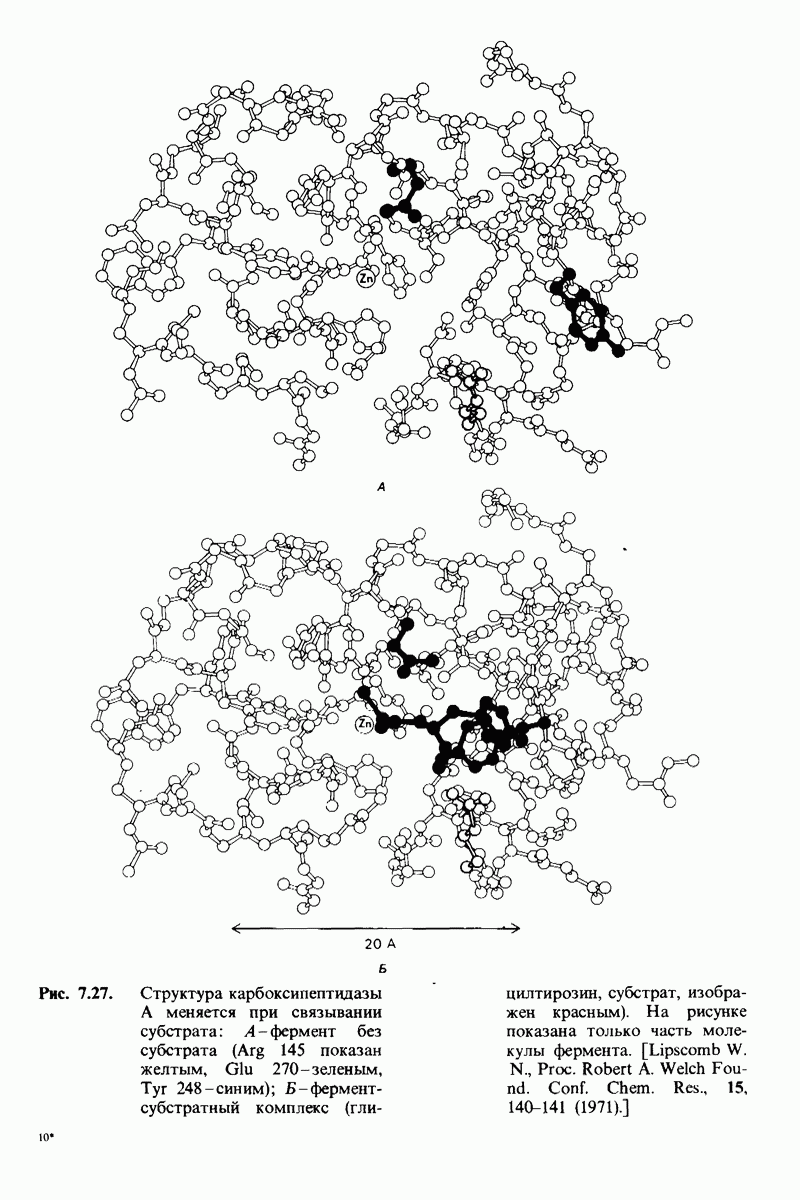
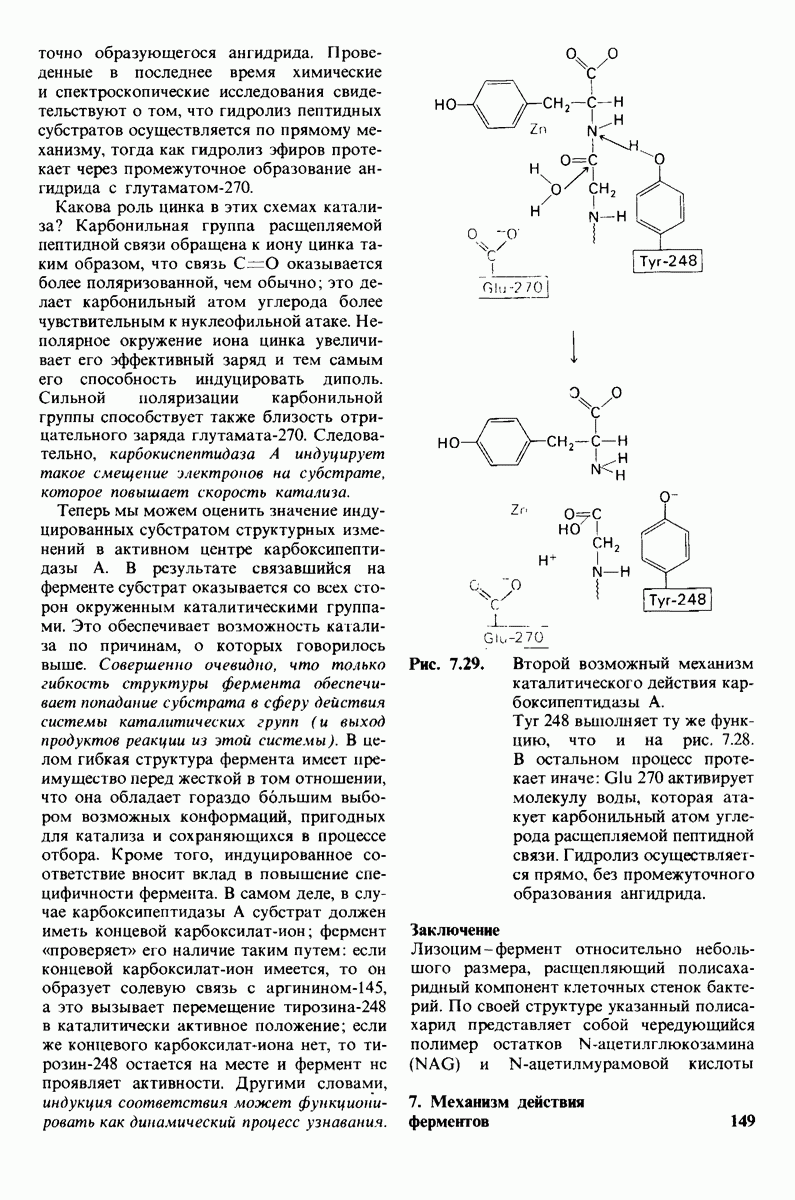
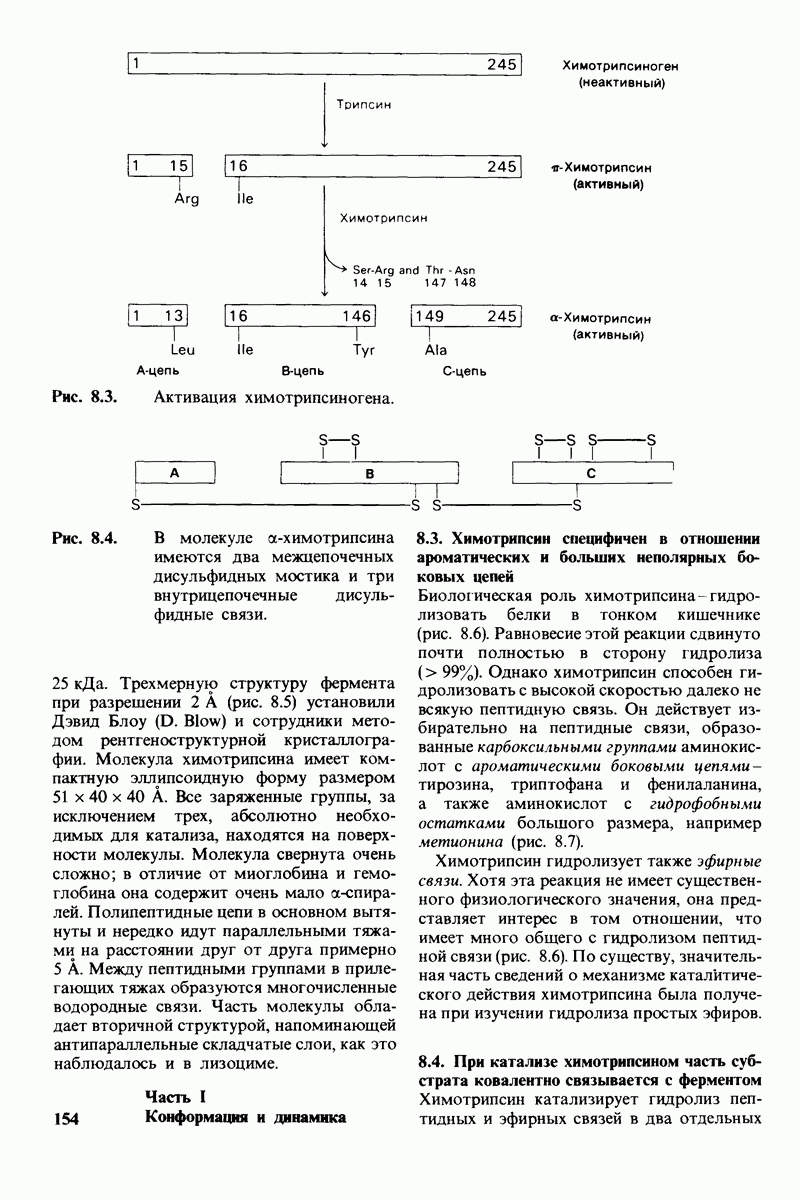
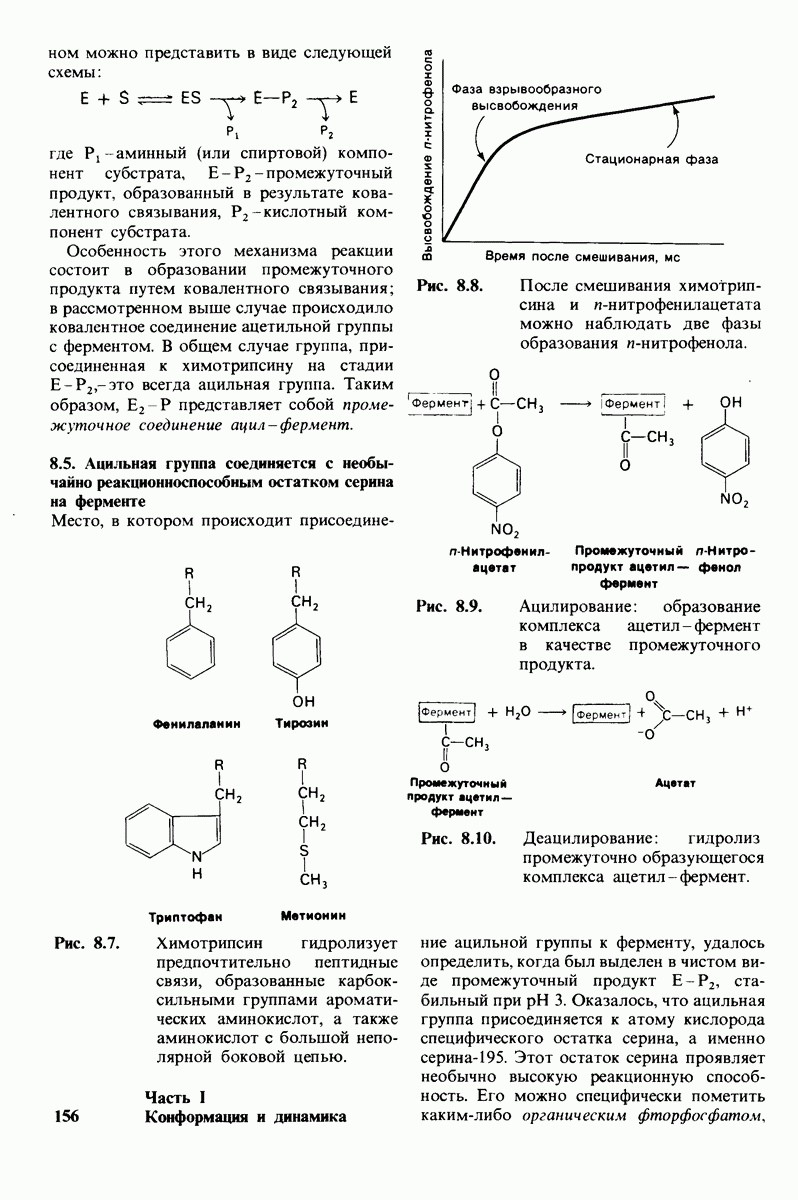
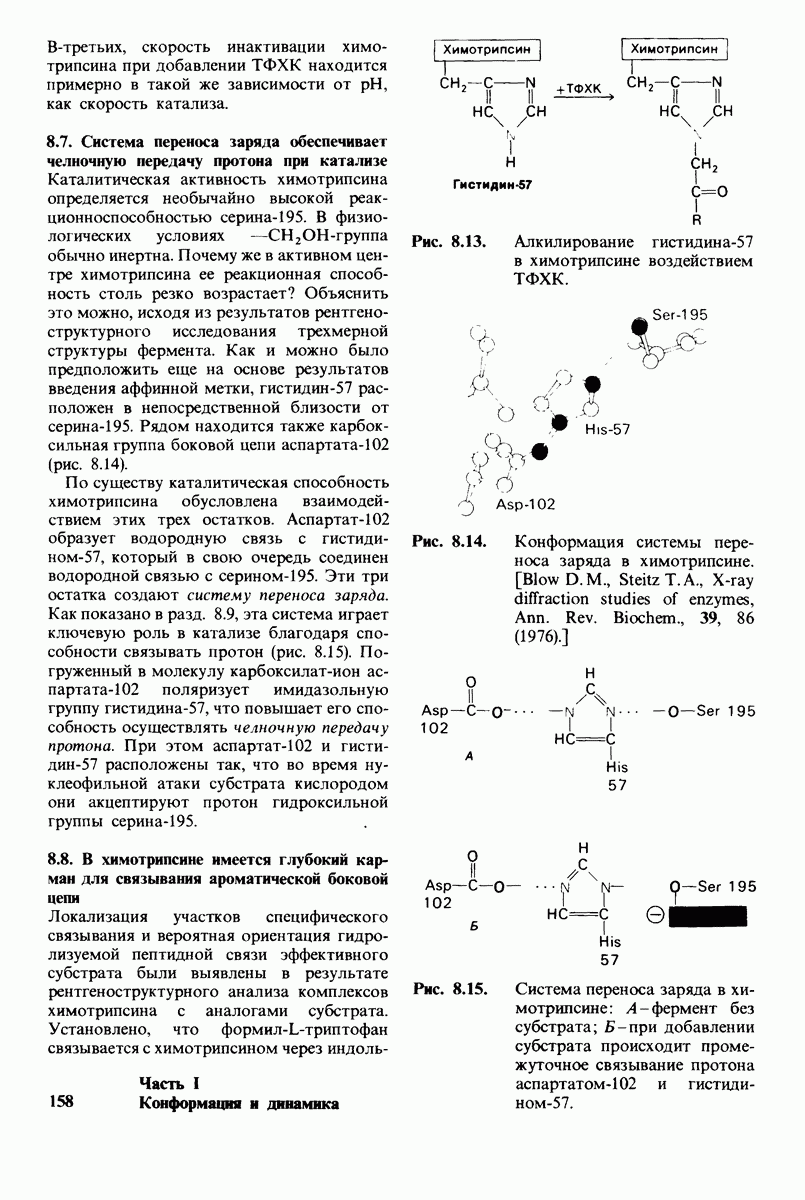
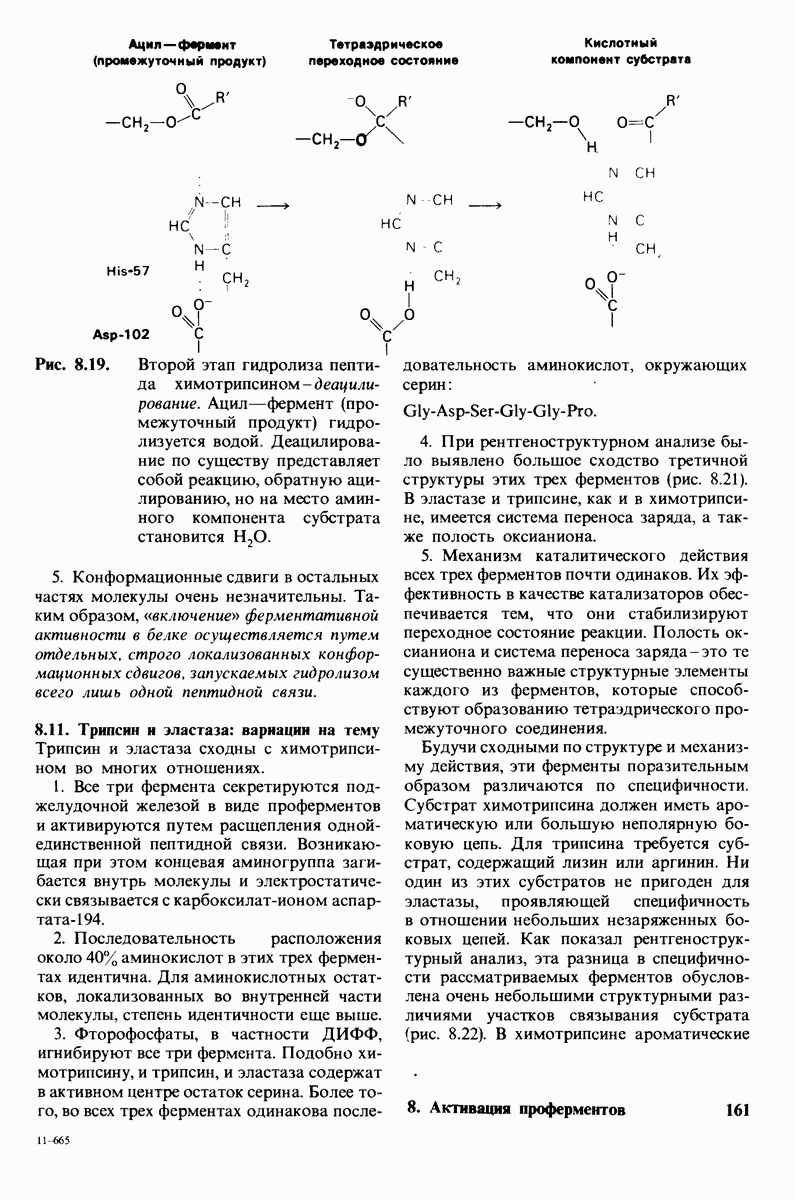
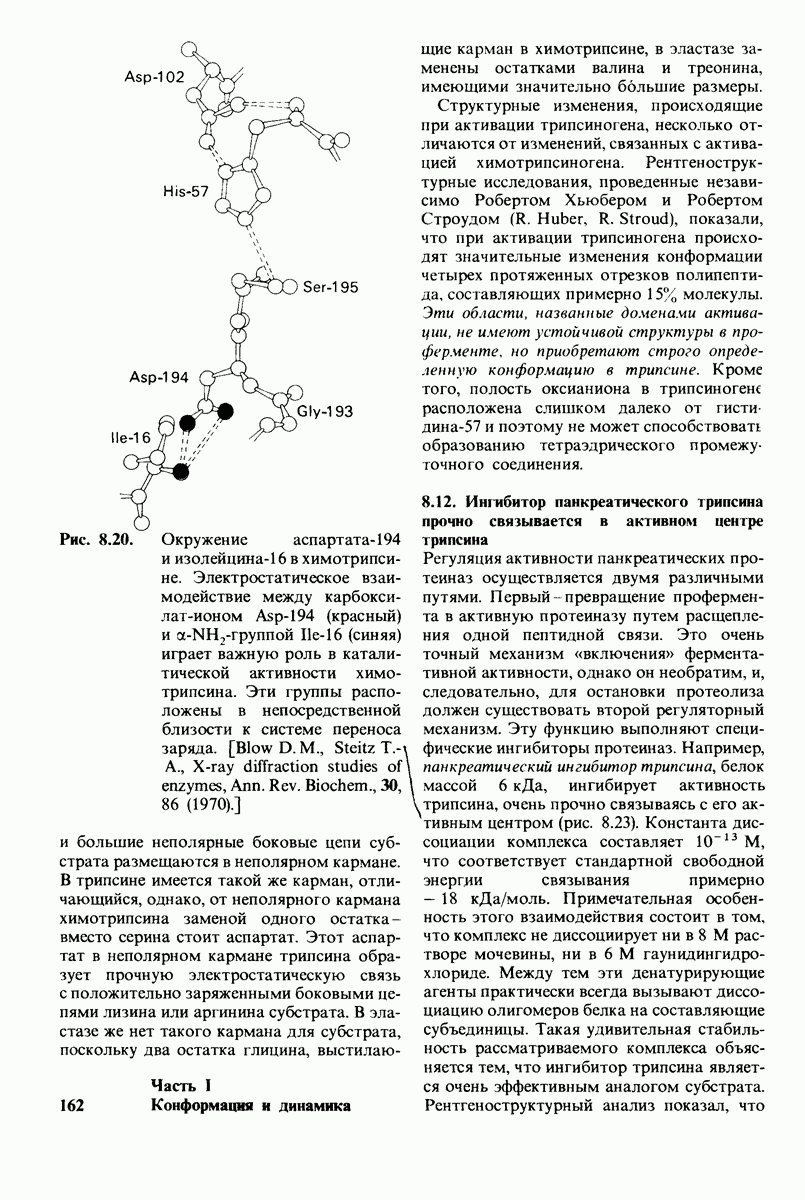
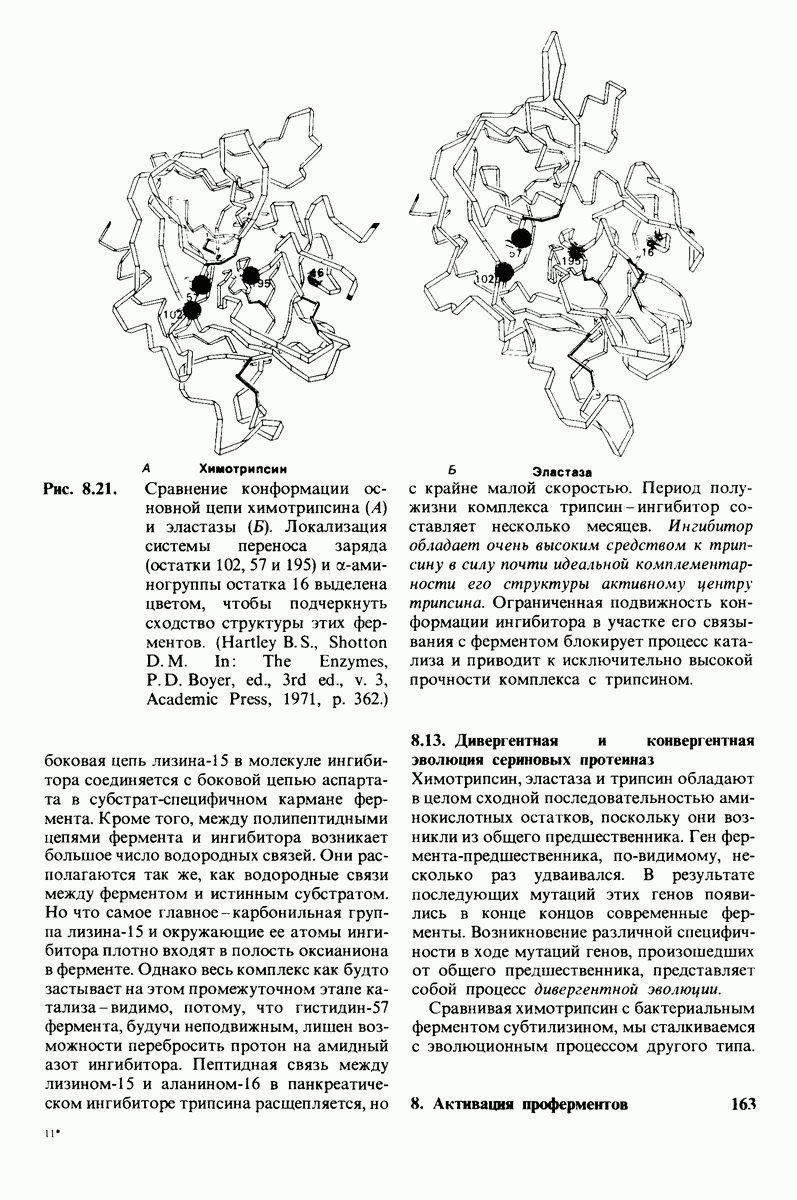
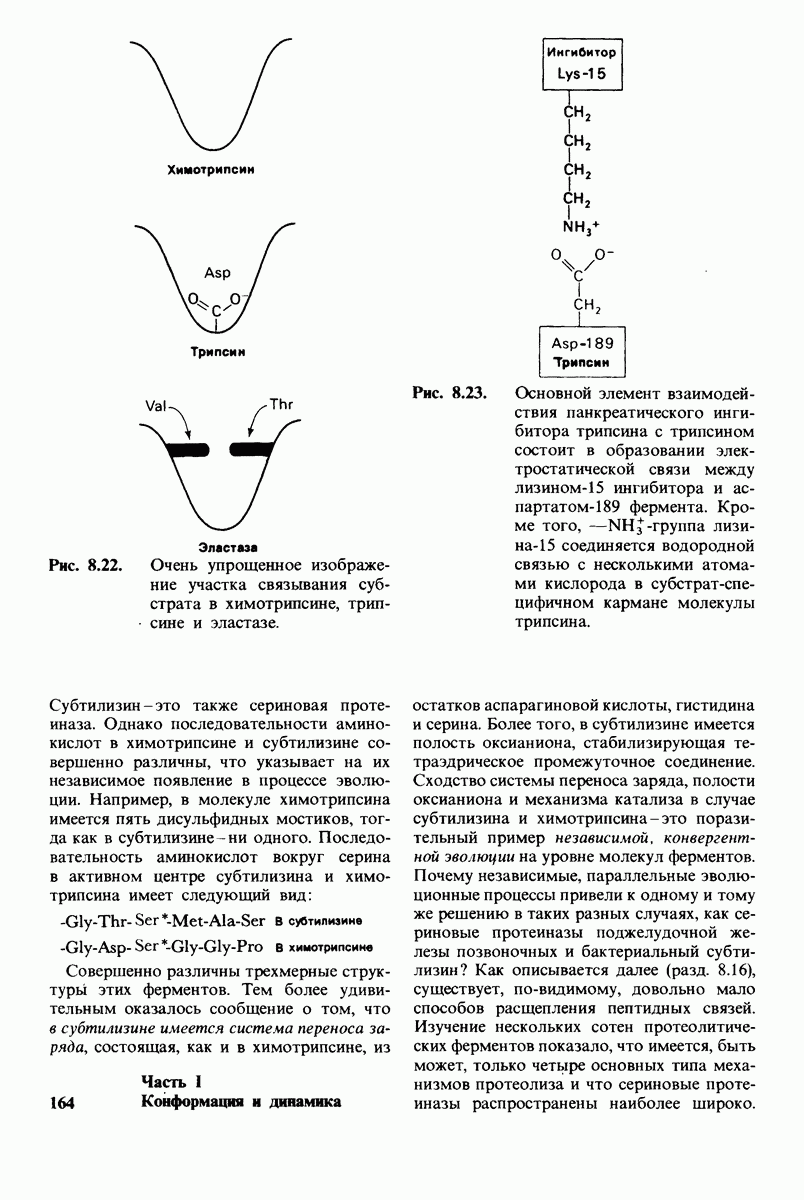
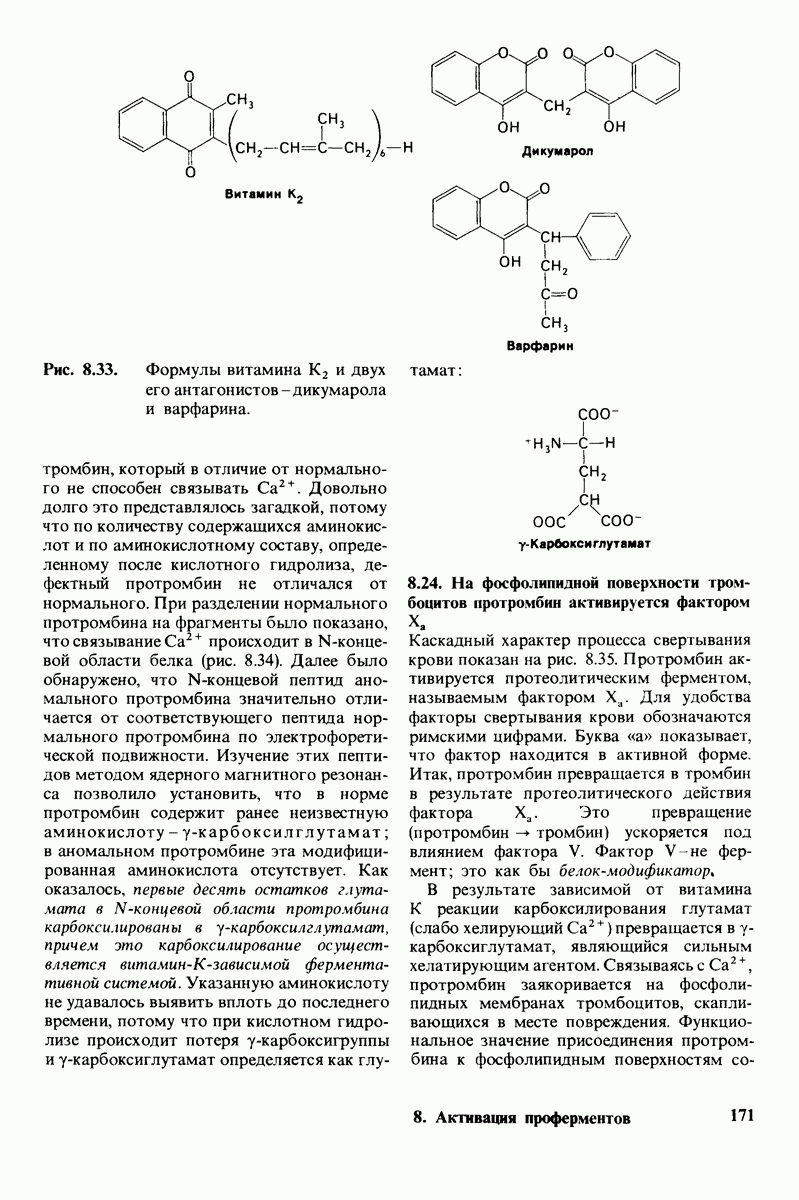
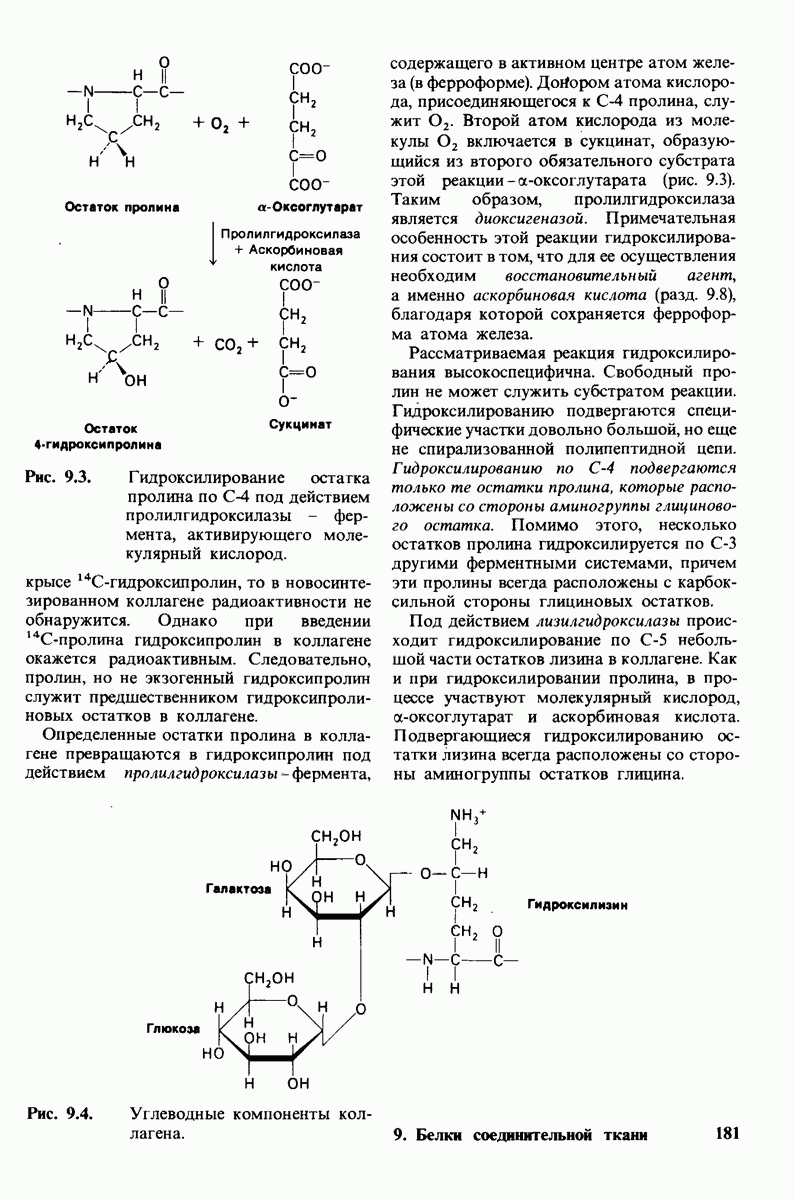
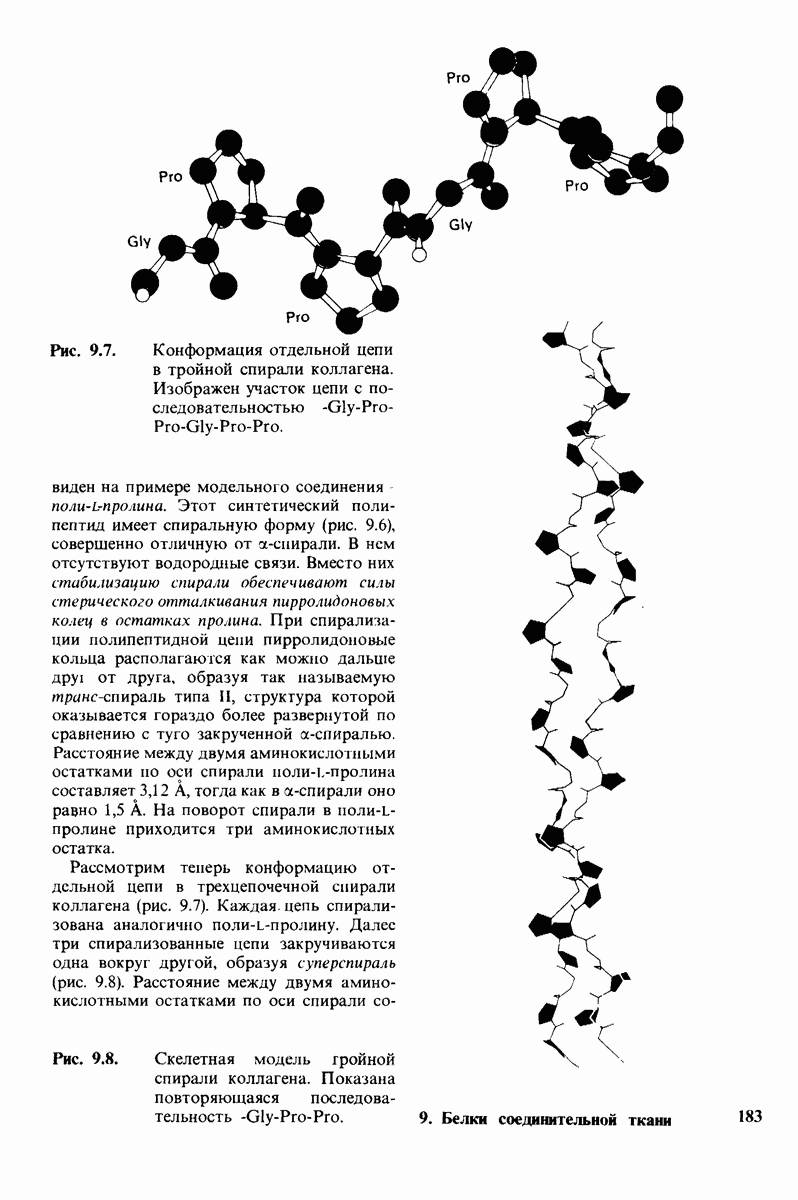
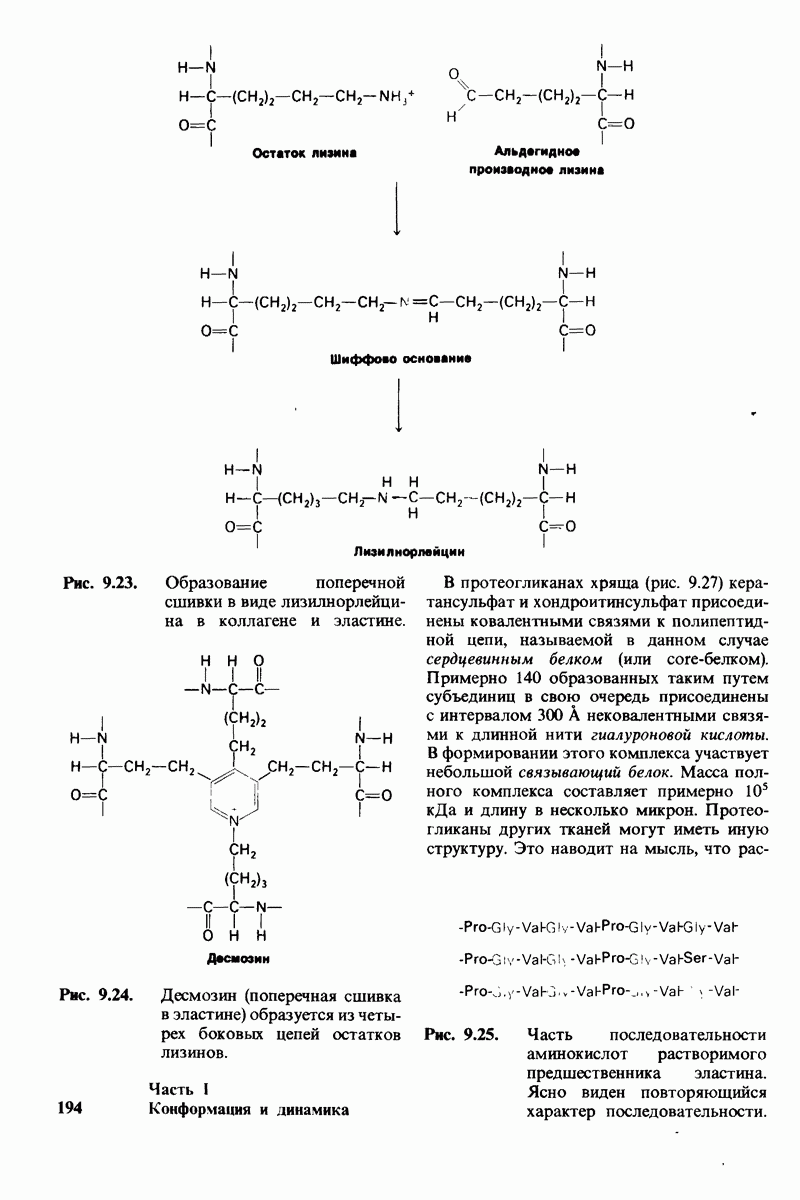
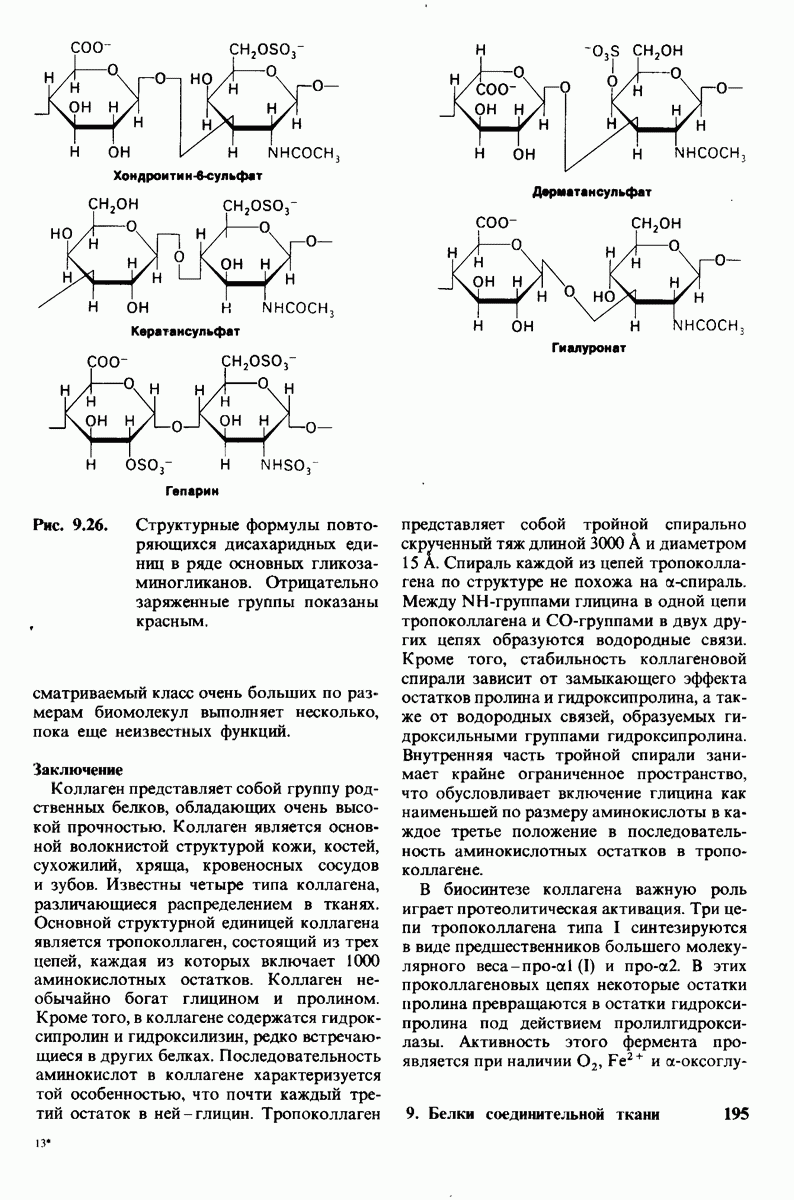
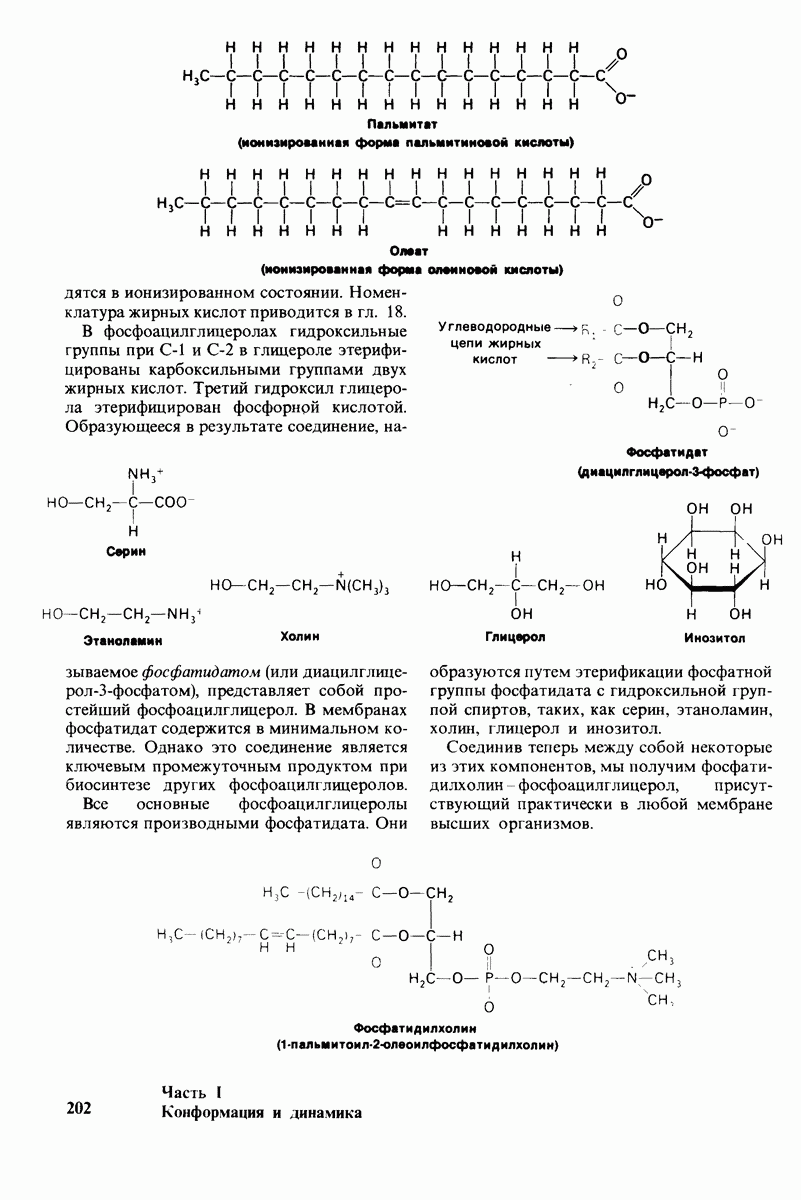
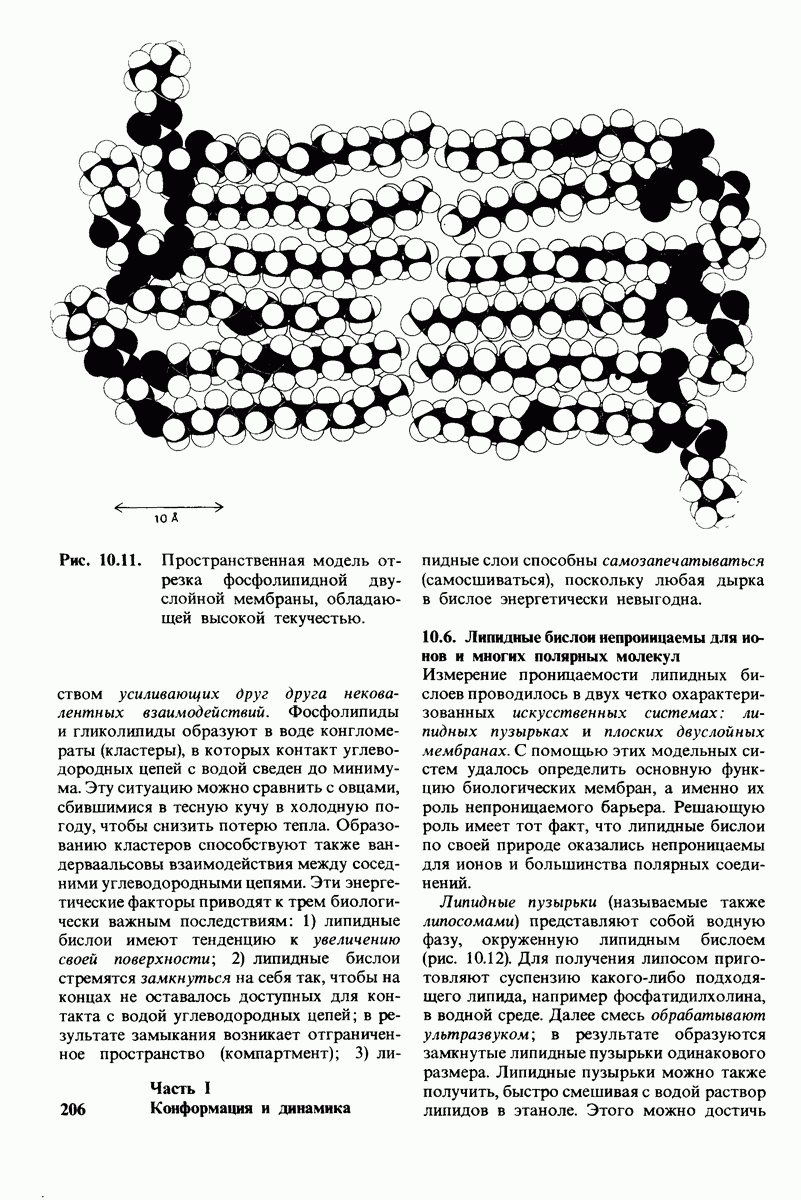
2.1. Белки построены из аминокислот
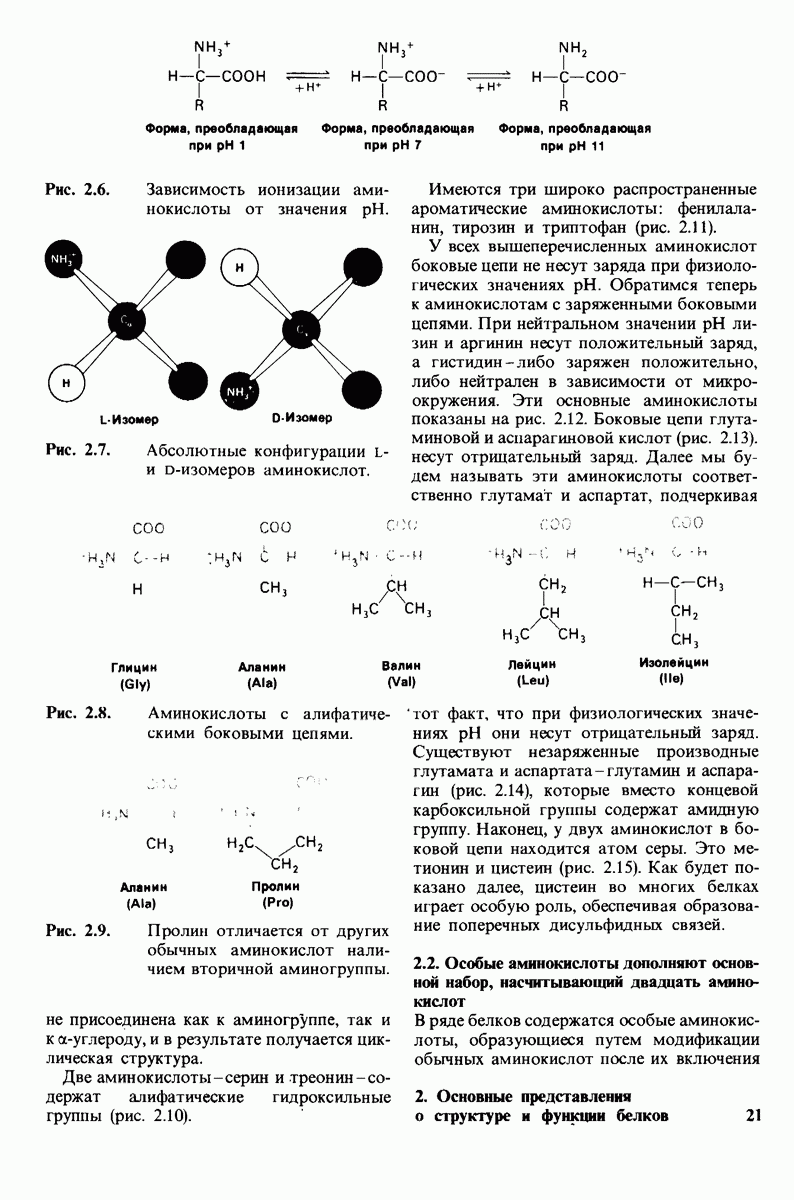
Основной структурной единицей белков являются аминокислоты. Каждая аминокислота состоит из аминогруппы, карбоксильной группы, атома водорода и определенной R-группы, присоединенной к атому углерода, называемому a-углеродом (рис. 2.5). R-группы называют боковой цепью по причинам, которые станут очевидными в ходе дальнейшего изложения. При нейтральном значении рН аминокислоты в растворах находятся не в виде неионизированных молекул, а преимущественно в виде биполярных ионов (цвиттерионов). При этом аминогруппа оказывается протонированной  а карбоксильная группа-диссоциированной
а карбоксильная группа-диссоциированной  Ионизация аминокислоты зависит от значения рН (рис. 2.6). В кислых растворах (например, при рН 1) карбоксильная группа находится в неионизированной форме
Ионизация аминокислоты зависит от значения рН (рис. 2.6). В кислых растворах (например, при рН 1) карбоксильная группа находится в неионизированной форме  -СООН), а аминогруппа ионизирована
-СООН), а аминогруппа ионизирована  В щелочных растворах (например, при рН 11), наоборот, ионизирована карбоксильная группа
В щелочных растворах (например, при рН 11), наоборот, ионизирована карбоксильная группа  а аминогруппа не
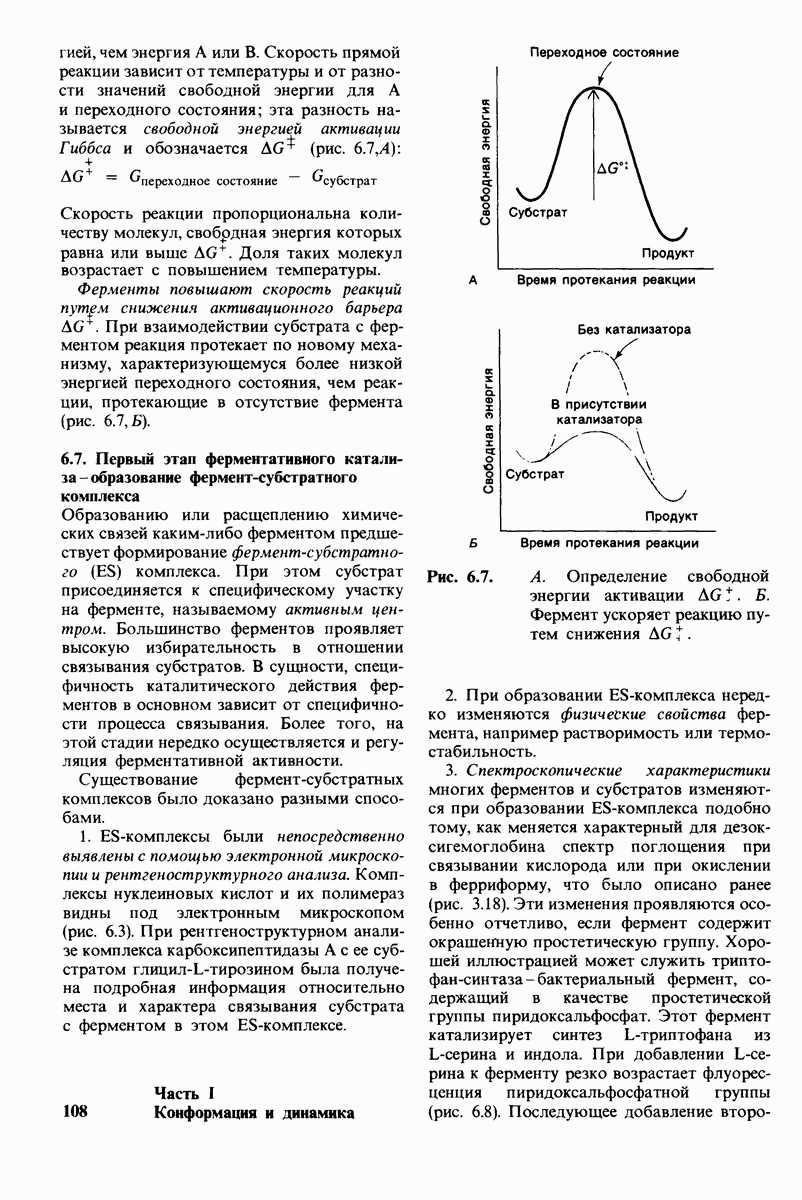
а аминогруппа не

Рис. 2.3.
Электронная микрофотография коллагенового волокна. (Печатается с любезного разрешения д-ра J. Cross и д-ра R. Bruns.)

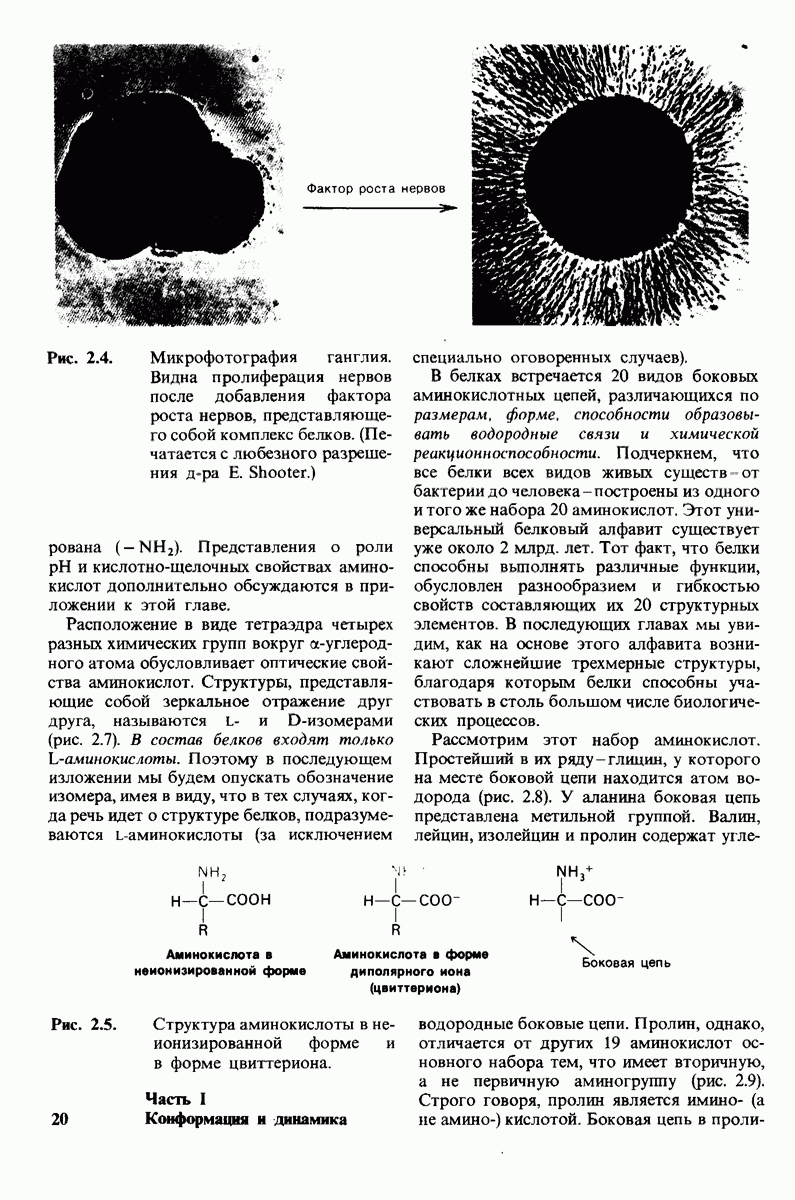
Рис. 2.4. Микрофотография ганглия, Видна пролиферация нервов после добавления фактора роста нервов, представляющего собой комплекс белков. (Печатается с любезного разрешения д-ра Е. Shooter.)
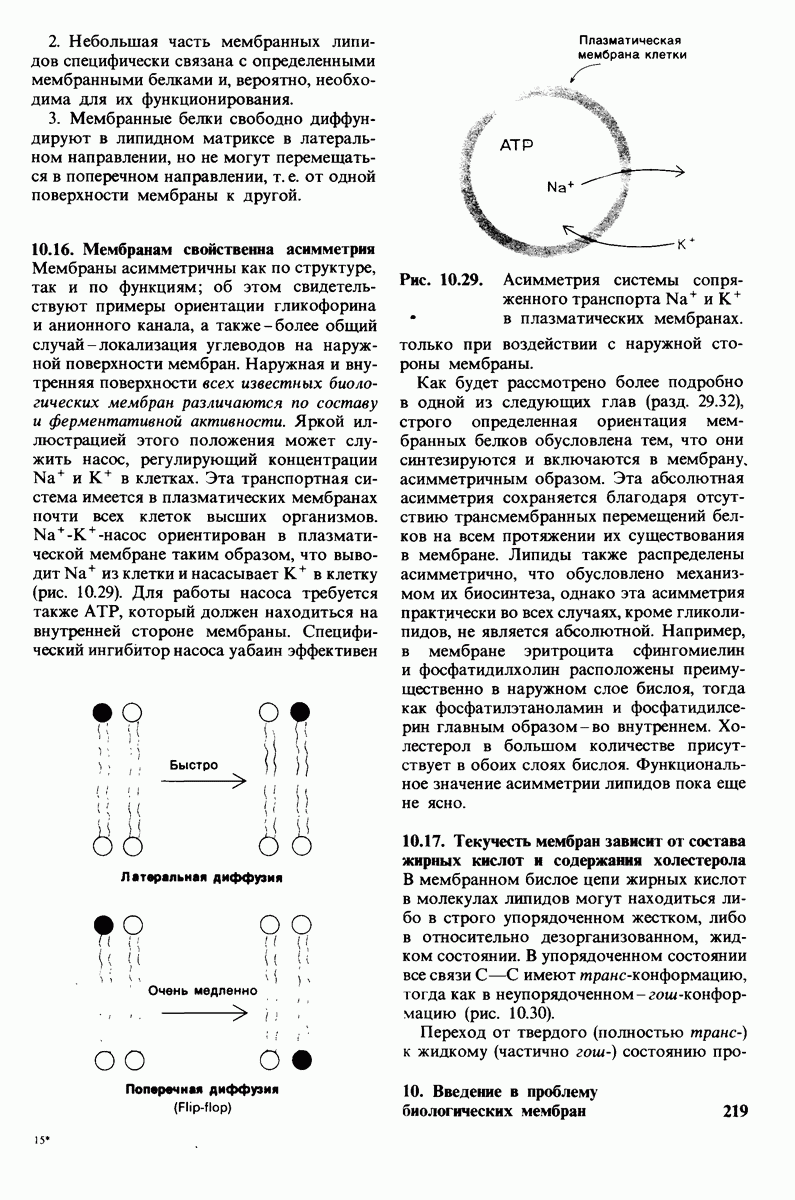
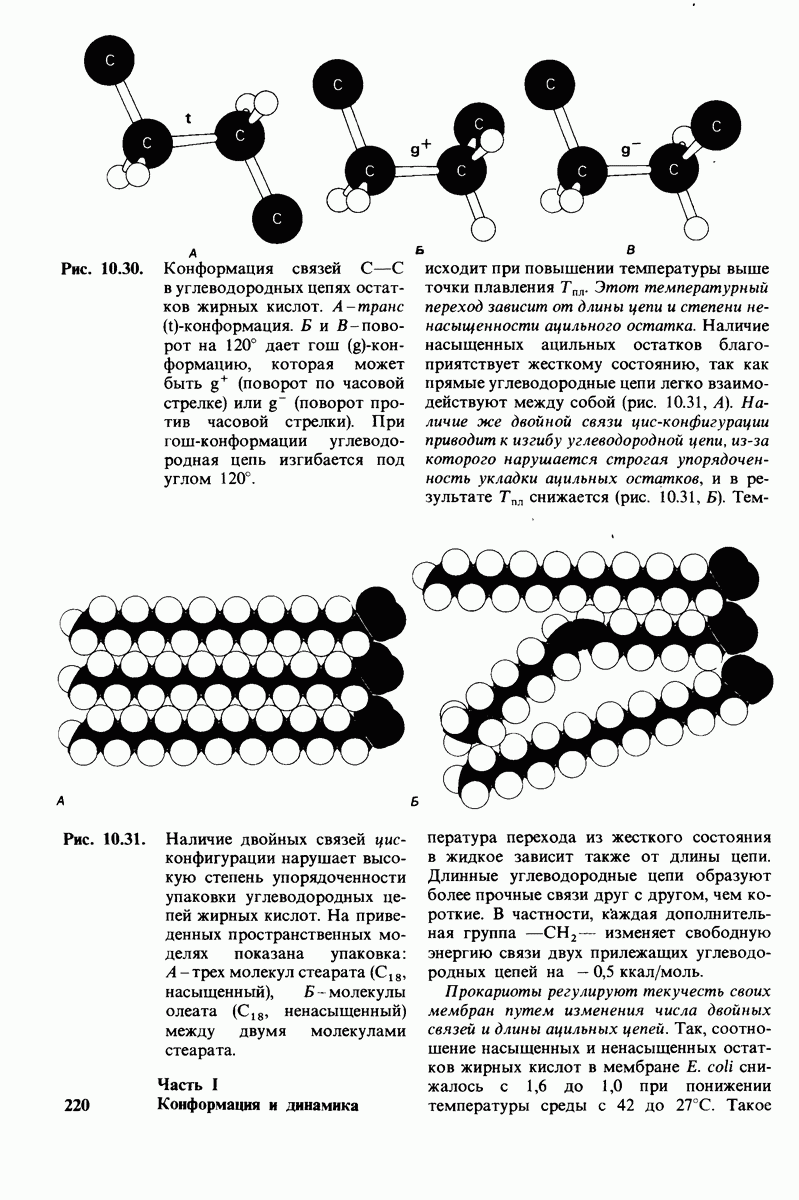
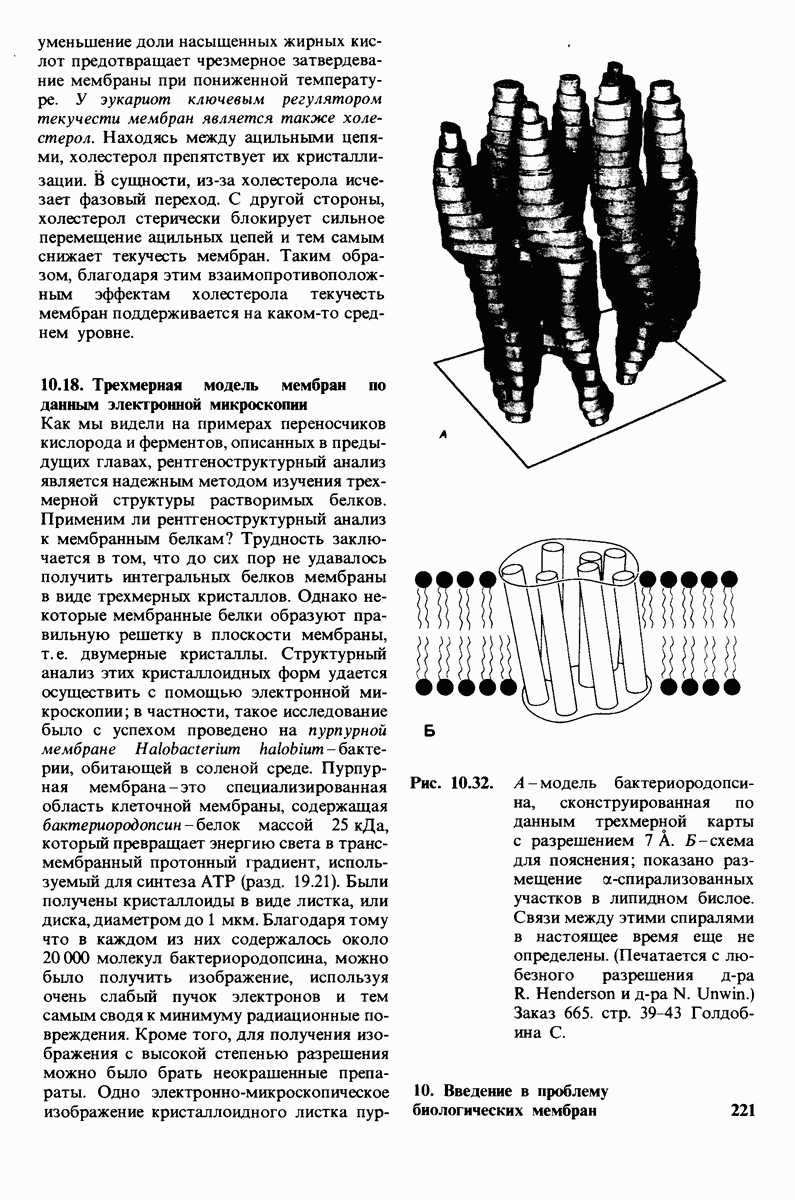
ионизирована  Представления о роли рН и кислотно-щелочных свойствах аминокислот дополнительно обсуждаются в приложении к этой главе,
Представления о роли рН и кислотно-щелочных свойствах аминокислот дополнительно обсуждаются в приложении к этой главе,
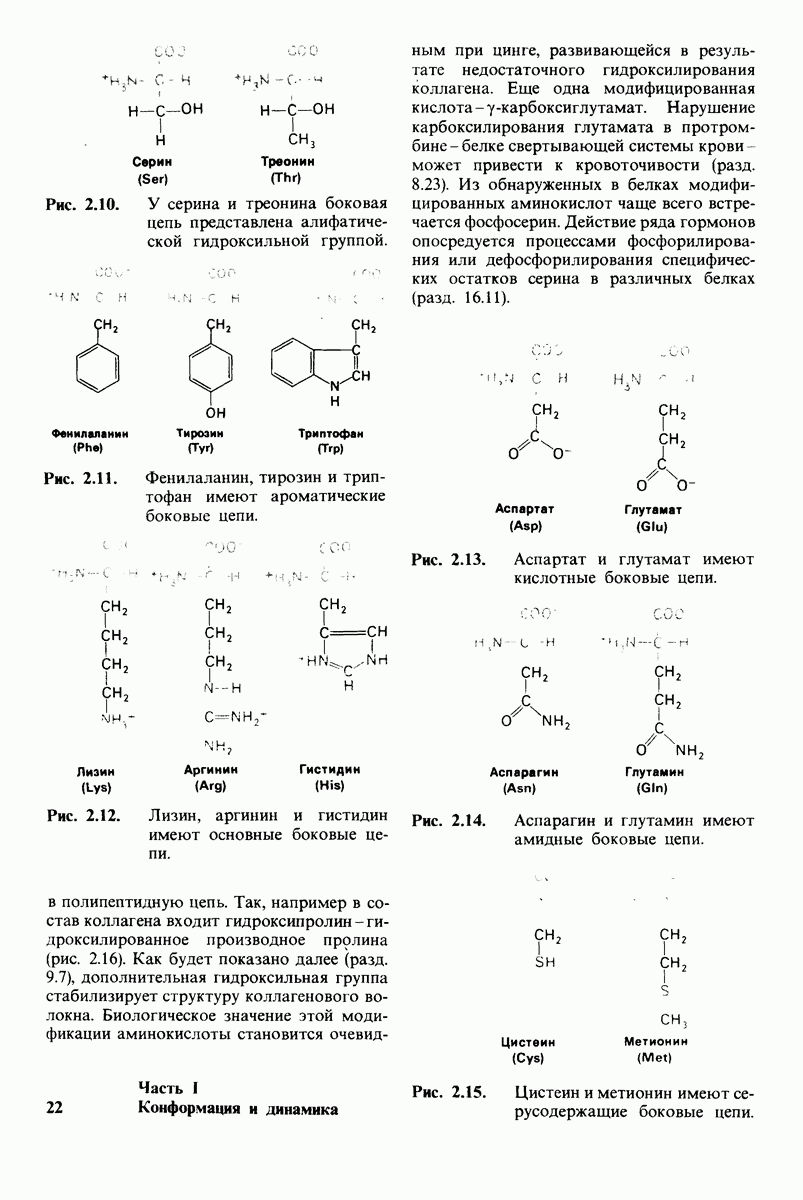
Расположение в виде тетраэдра четырех разных химических групп вокруг  -углеродного атома обусловливает оптические свойства аминокислот. Структуры, представляющие собой зеркальное отражение друг друга, называются L- и D-изомерами (рис. 2.7). В состав белков входят только L-аминокислоты. Поэтому в последующем изложении мы будем опускать обозначение изомера, имея в виду, что в тех случаях, когда речь идет о структуре белков, подразумеваются L-аминокислоты (за исключением специально оговоренных случаев),
-углеродного атома обусловливает оптические свойства аминокислот. Структуры, представляющие собой зеркальное отражение друг друга, называются L- и D-изомерами (рис. 2.7). В состав белков входят только L-аминокислоты. Поэтому в последующем изложении мы будем опускать обозначение изомера, имея в виду, что в тех случаях, когда речь идет о структуре белков, подразумеваются L-аминокислоты (за исключением специально оговоренных случаев),

Рис. 2.5. Структура аминокислоты в неионизированной форме и в форме цвиттериона.
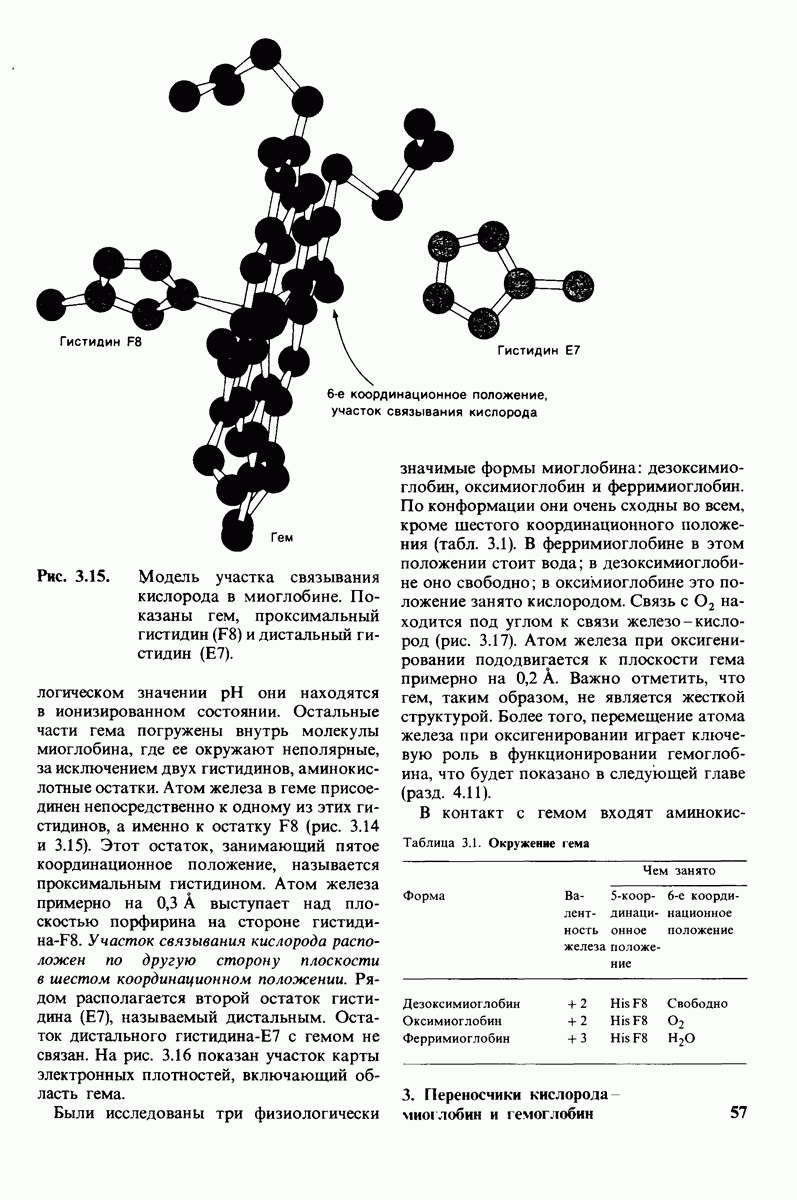
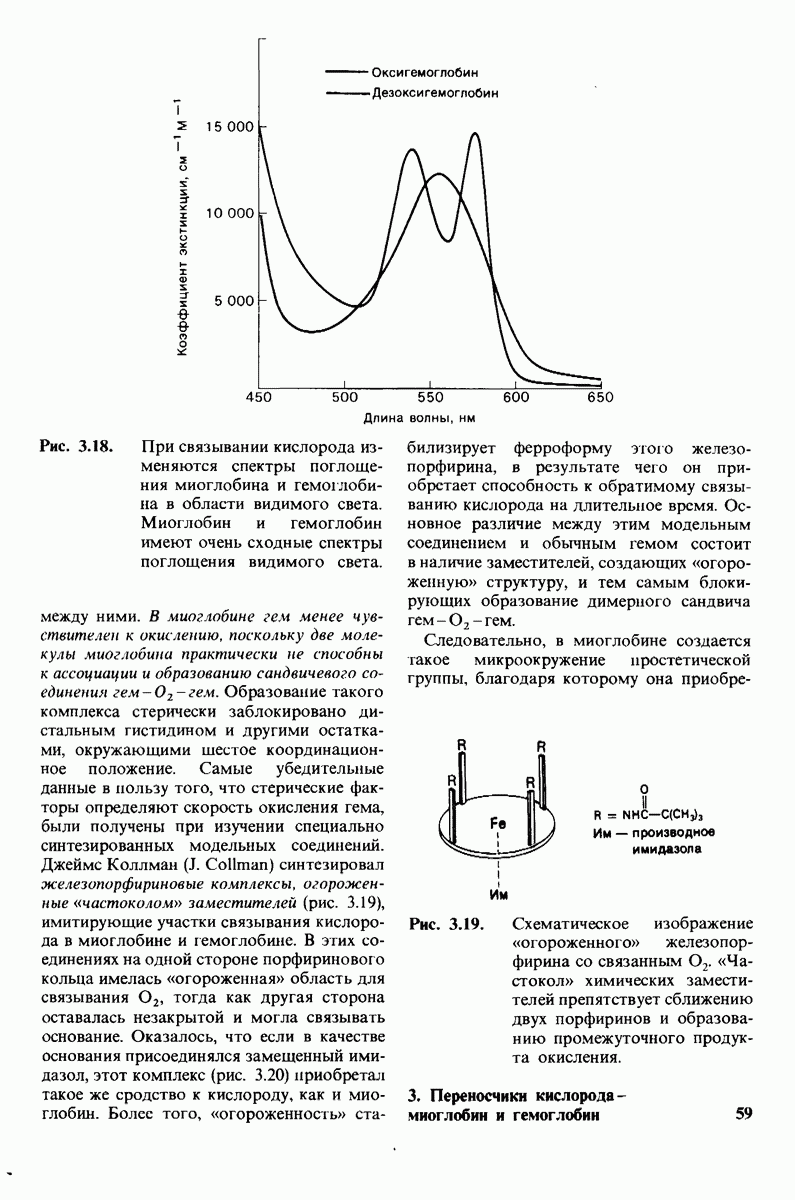
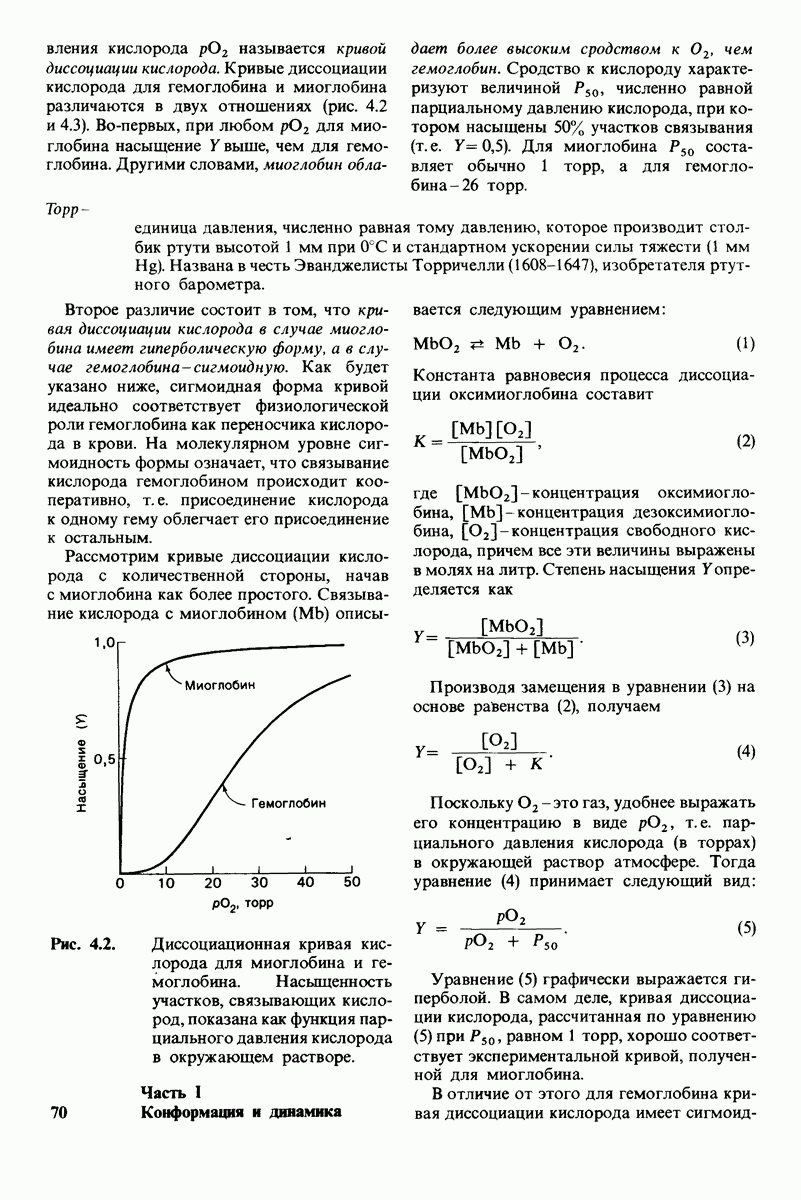
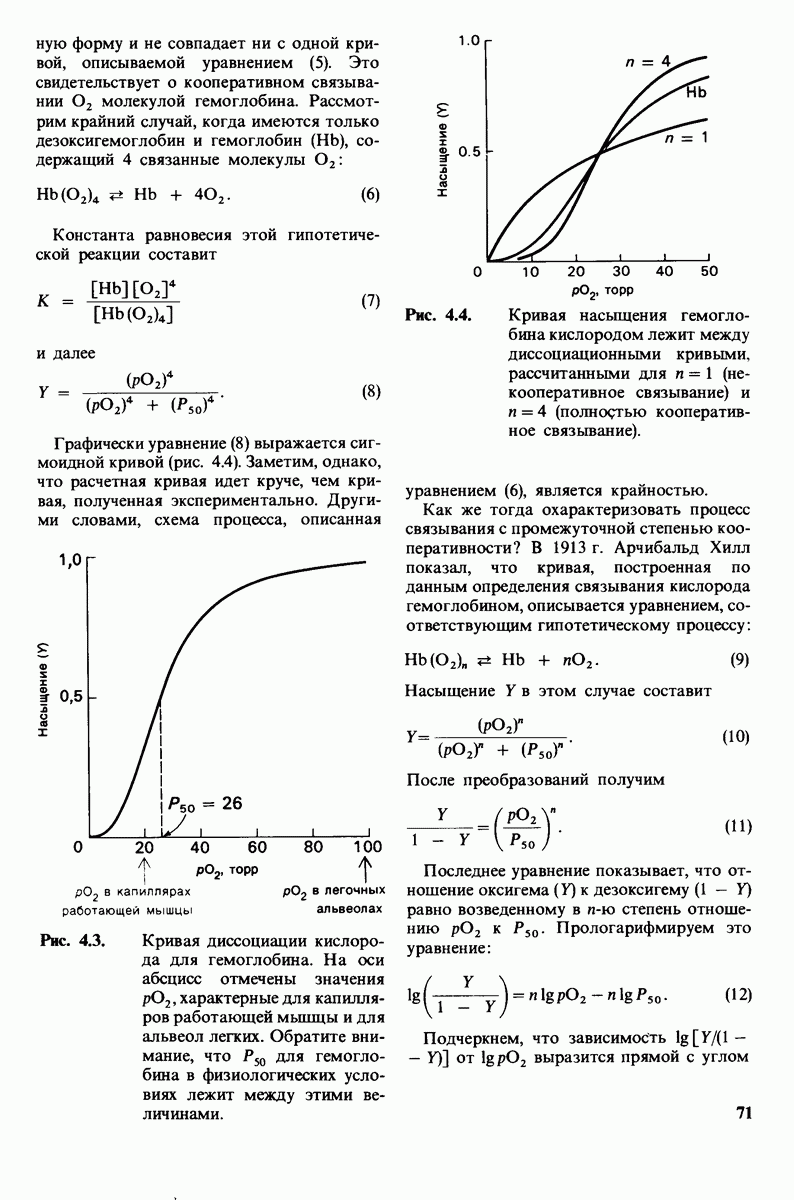
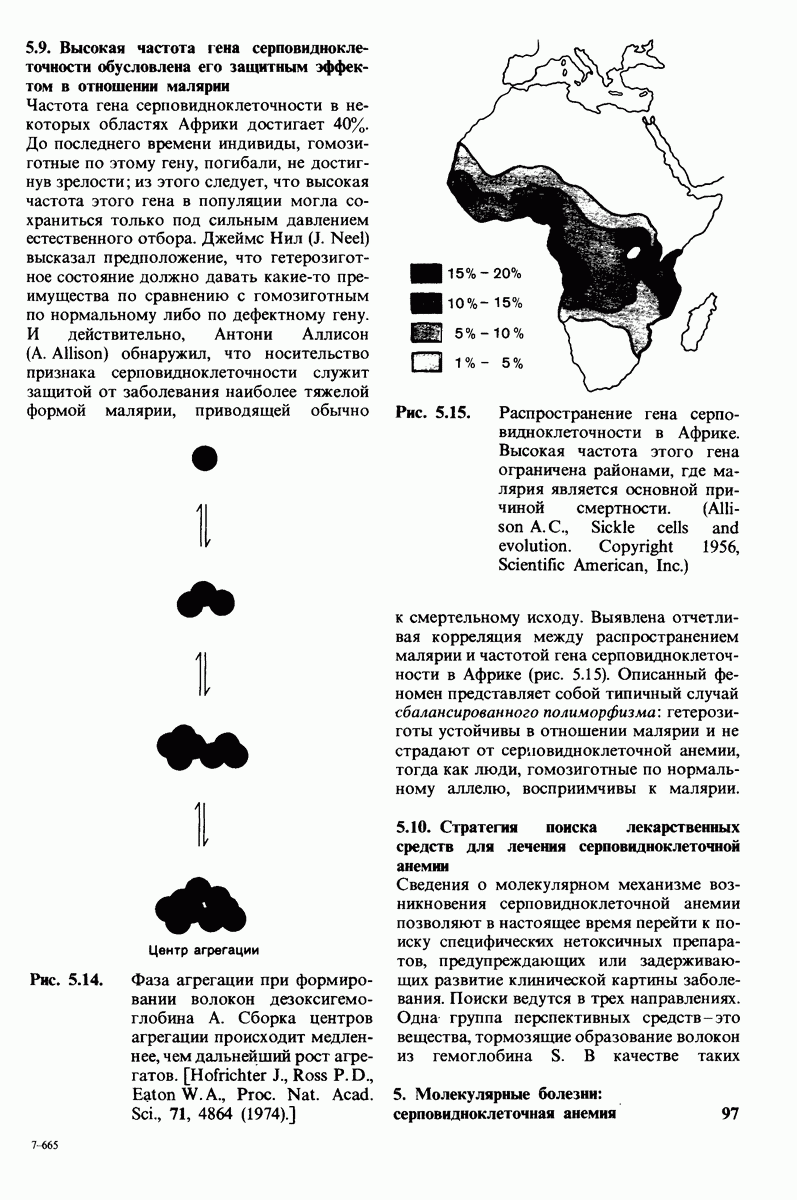
В белках встречается 20 видов боковых аминокислотных цепей, различающихся по размерам, форме  способности образовывать водородные связи и химической реакционноспособности. Подчеркнем, что все белки всех видов живых существ
способности образовывать водородные связи и химической реакционноспособности. Подчеркнем, что все белки всех видов живых существ  бактерии до человека - построены из одного и того же набора 20 аминокислот. Этот универсальный белковый алфавит существует уже около 2 млрд. лет. Тот факт, что белки способны выполнять различные функции, обусловлен разнообразием и гибкостью свойств составляющих их 20 структурных элементов. В последующих главах мы увидим, как на основе этого алфавита возникают сложнейшие трехмерные структуры, благодаря которым белки способны участвовать в столь большом числе биологических процессов.
бактерии до человека - построены из одного и того же набора 20 аминокислот. Этот универсальный белковый алфавит существует уже около 2 млрд. лет. Тот факт, что белки способны выполнять различные функции, обусловлен разнообразием и гибкостью свойств составляющих их 20 структурных элементов. В последующих главах мы увидим, как на основе этого алфавита возникают сложнейшие трехмерные структуры, благодаря которым белки способны участвовать в столь большом числе биологических процессов.
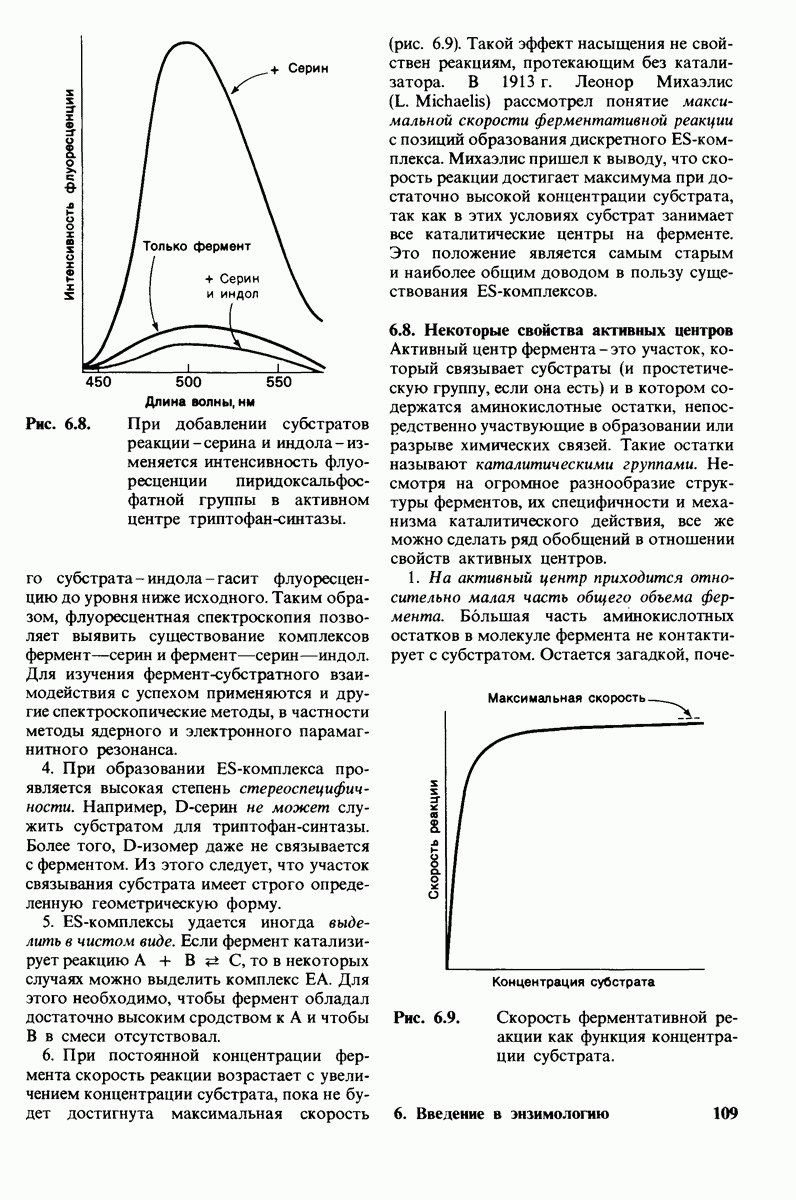
Рассмотрим этот набор аминокислот. Простейший в их ряду - глицин, у которого на месте боковой цепи находится атом водорода (рис. 2.8). У аланина боковая цепь представлена метальной группой. Валин, лейцин, изолейцин и пролин содержат углеводородные боковые цепи. Пролин, однако, отличается от других 19 аминокислот основного набора тем, что имеет вторичную, а не первичную аминогруппу (рис. 2.9). Строго говоря, пролин является имино- (а не амино-) кислотой. Боковая цепь в

Рис. 2.6. Зависимость ионизации аминокислоты от значения рН.

Рис. 2.7. Абсолютные конфигурации L- и D-изомеров аминокислот.

Рис. 2.8. Аминокислоты с алифатическими боковыми цепями.

Рис. 2.9. Пролин отличается от других обычных аминокислот наличием вторичной аминогруппы.
не присоединена как к аминогруппе, так и к  -углероду, и в результате получается циклическая структура.
-углероду, и в результате получается циклическая структура.
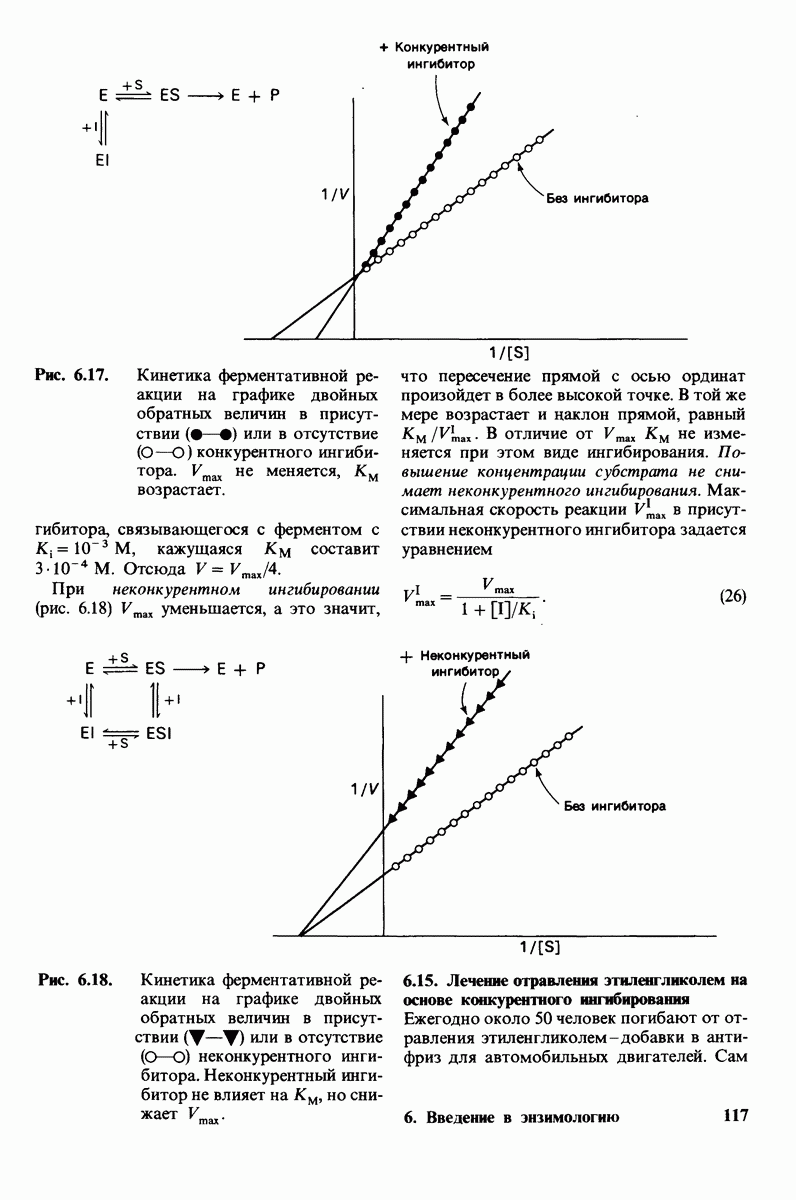
Две аминокислоты-серин и треонин-содержат алифатические гидроксильные группы (рис. 2.10).
Имеются три широко распространенные ароматические аминокислоты: фенилаланин, тирозин и триптофан (рис. 2.11).
У всех вышеперечисленных аминокислот боковые цепи не несут заряда при физиологических значениях рН. Обратимся теперь к аминокислотам с заряженными боковыми цепями. При нейтральном значении рН лизин и аргинин несут положительный заряд, а гистидин - либо заряжен положительно, либо нейтрален в зависимости от микроокружения. Эти основные аминокислоты показаны на рис. 2.12. Боковые цепи глутаминовой и аспарагиновой кислот (рис. 2.13). несут отрицательный заряд. Далее мы будем называть эти аминокислоты соответственно глутамат и аспартат, подчеркивая тот факт, что при физиологических значениях рН они несут отрицательный заряд. Существуют незаряженные производные глутамата и аспартата - глутамин и аспарагин (рис. 2.14), которые вместо концевой карбоксильной группы содержат амидную группу. Наконец, у двух аминокислот в боковой цепи находится атом серы. Это метионин и цистеин (рис. 2.15). Как будет показано далее, цистеин во многих белках играет особую роль, обеспечивая образование поперечных дисульфидных связей.