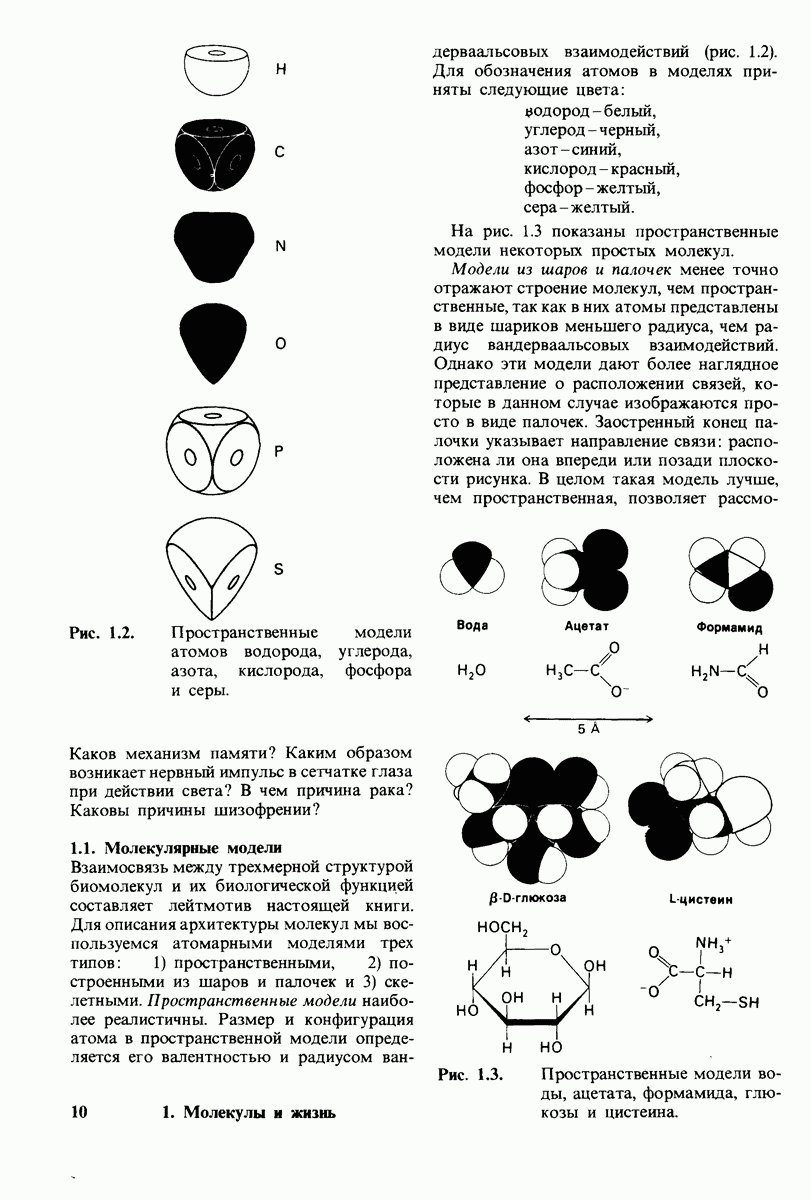
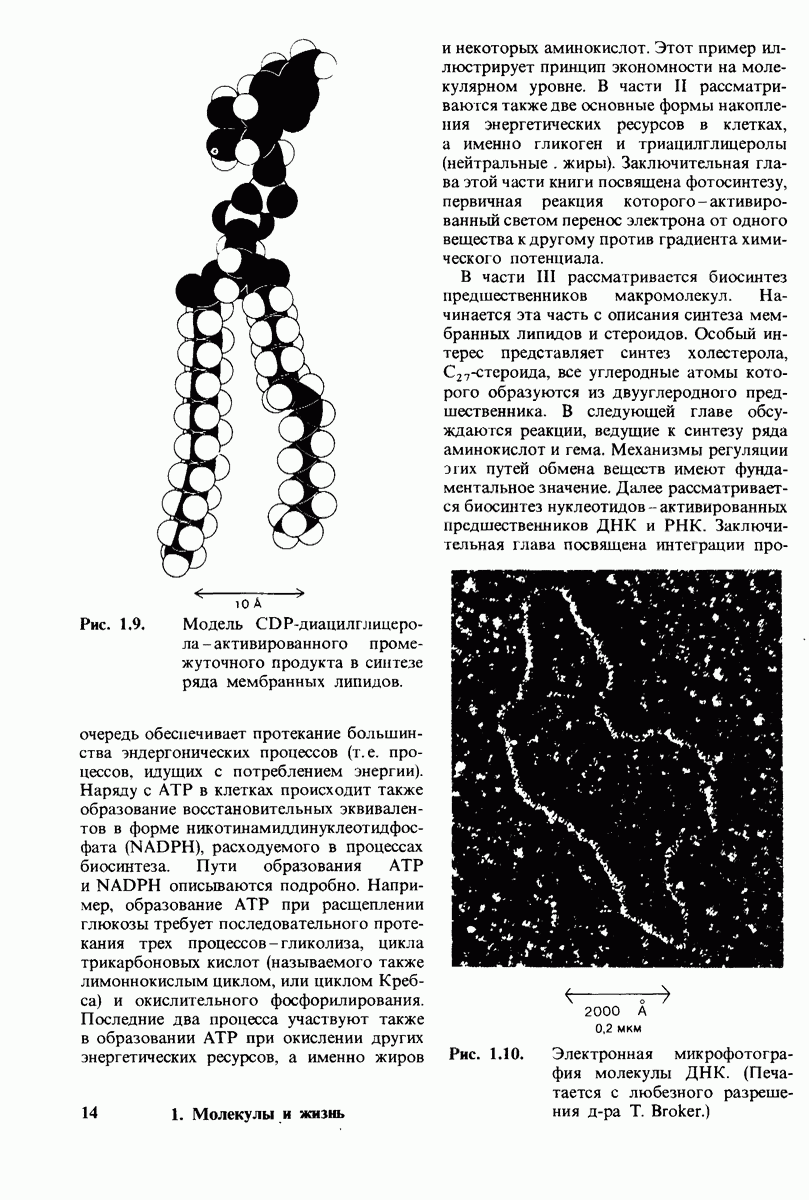
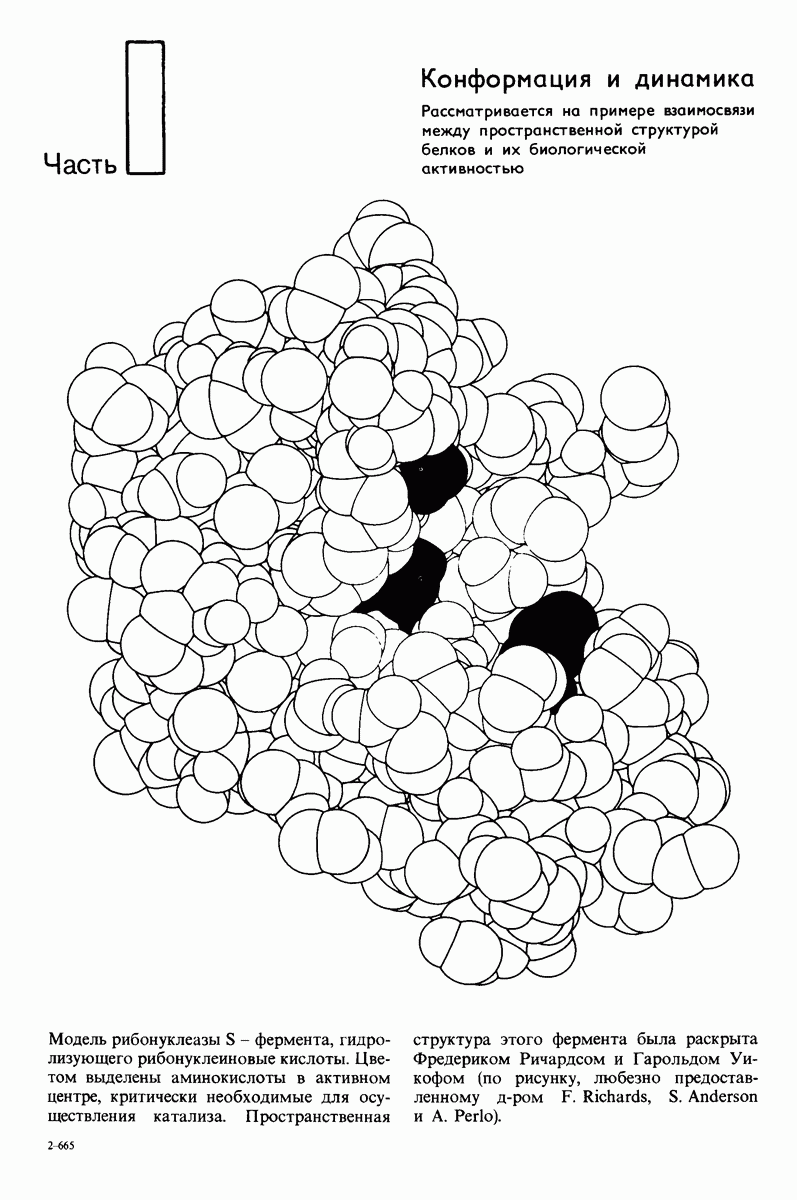
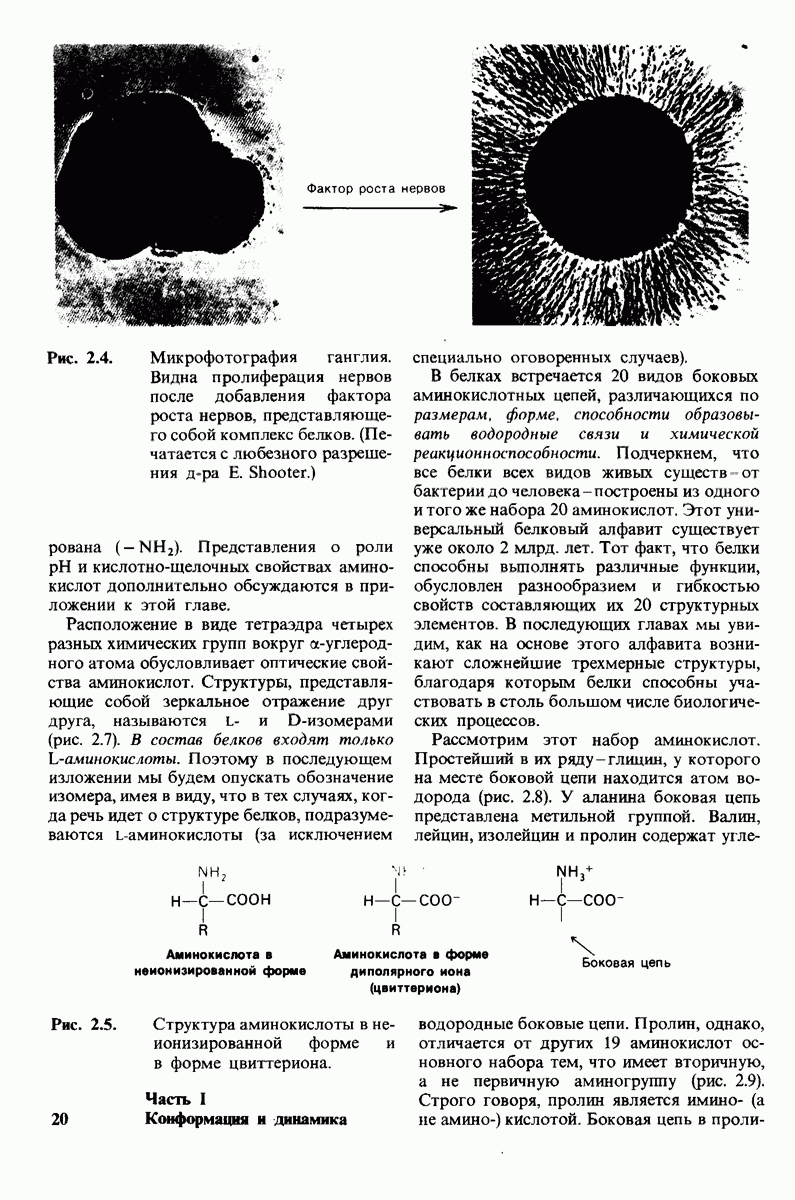
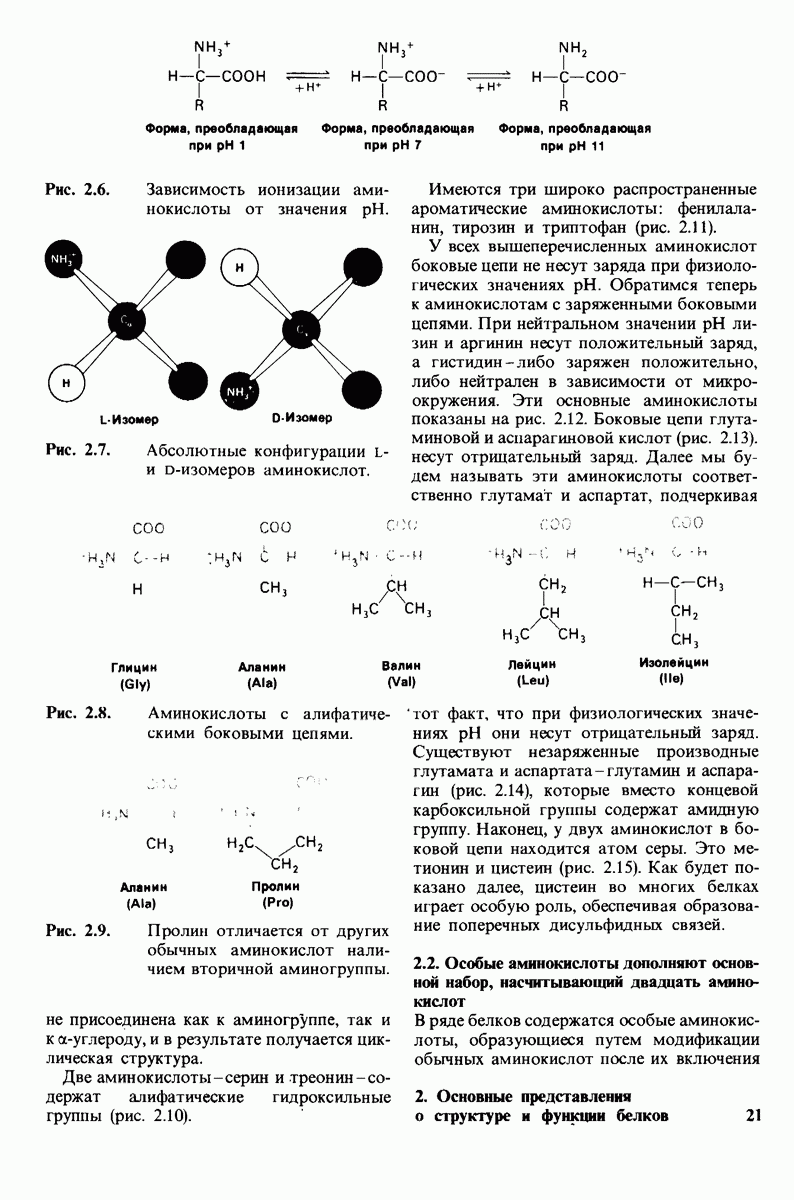
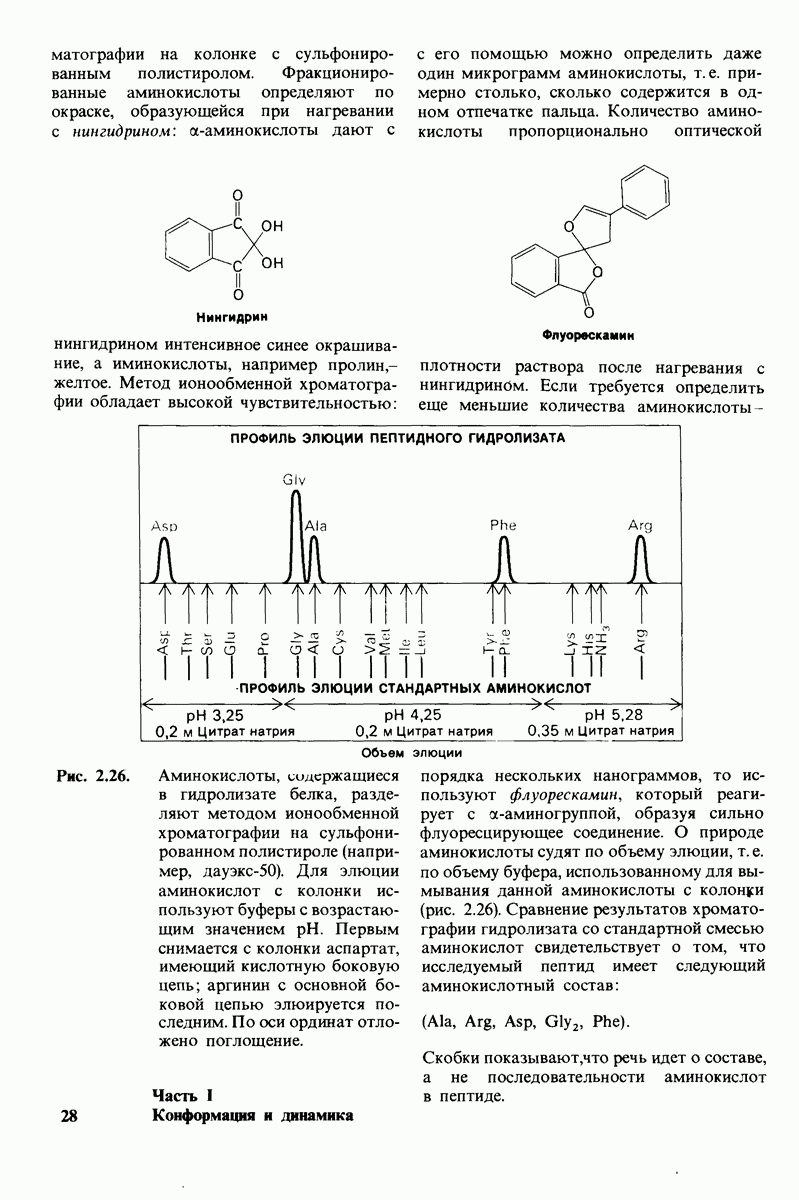
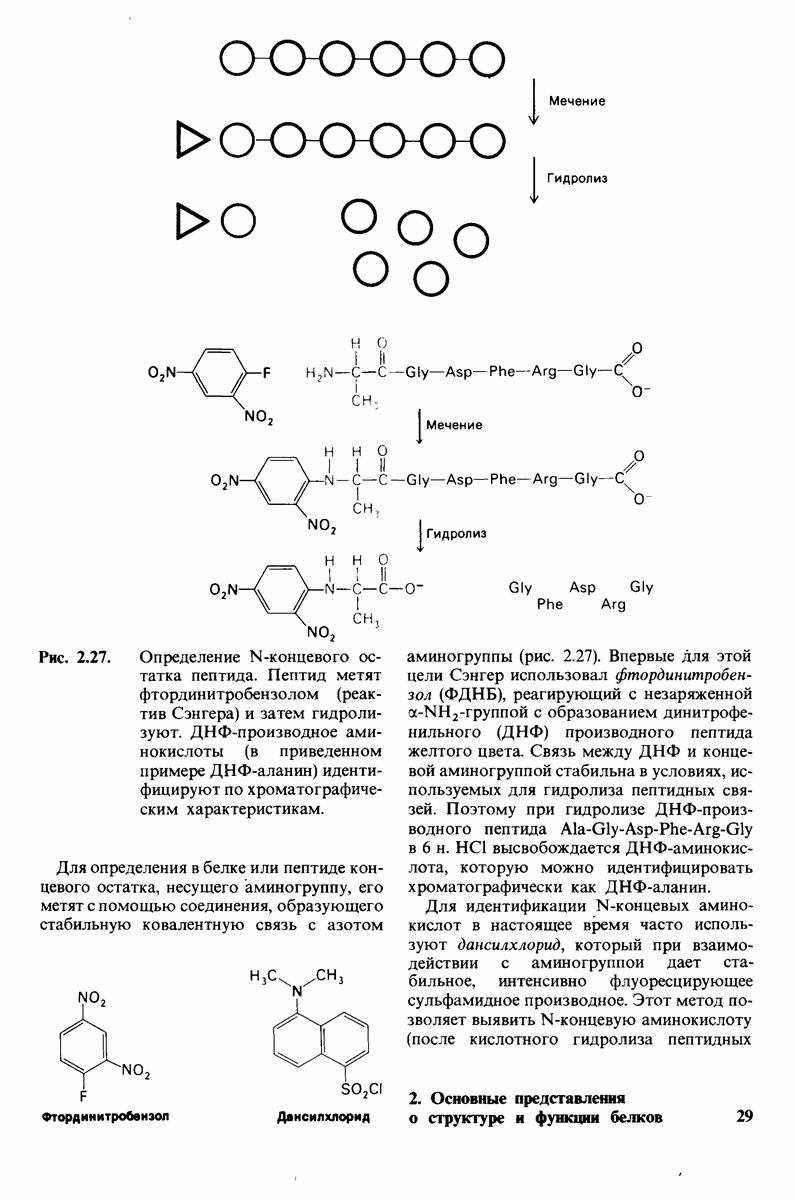
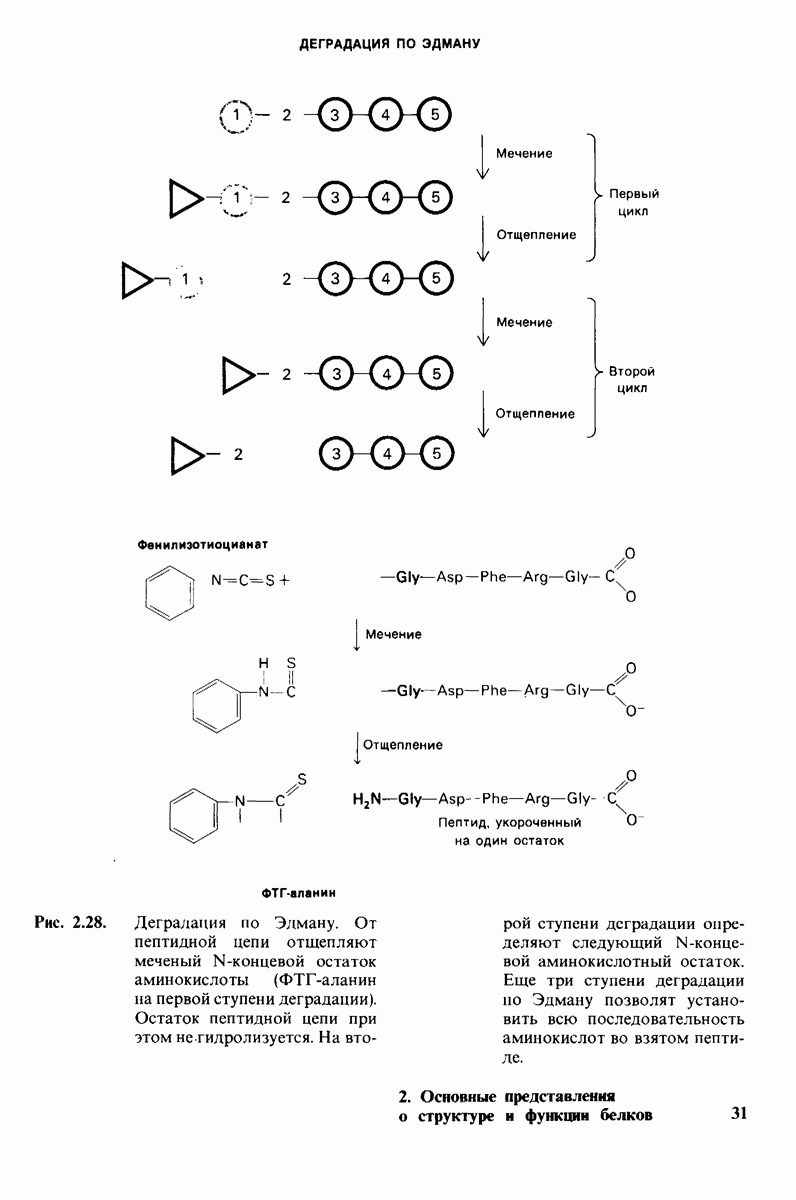
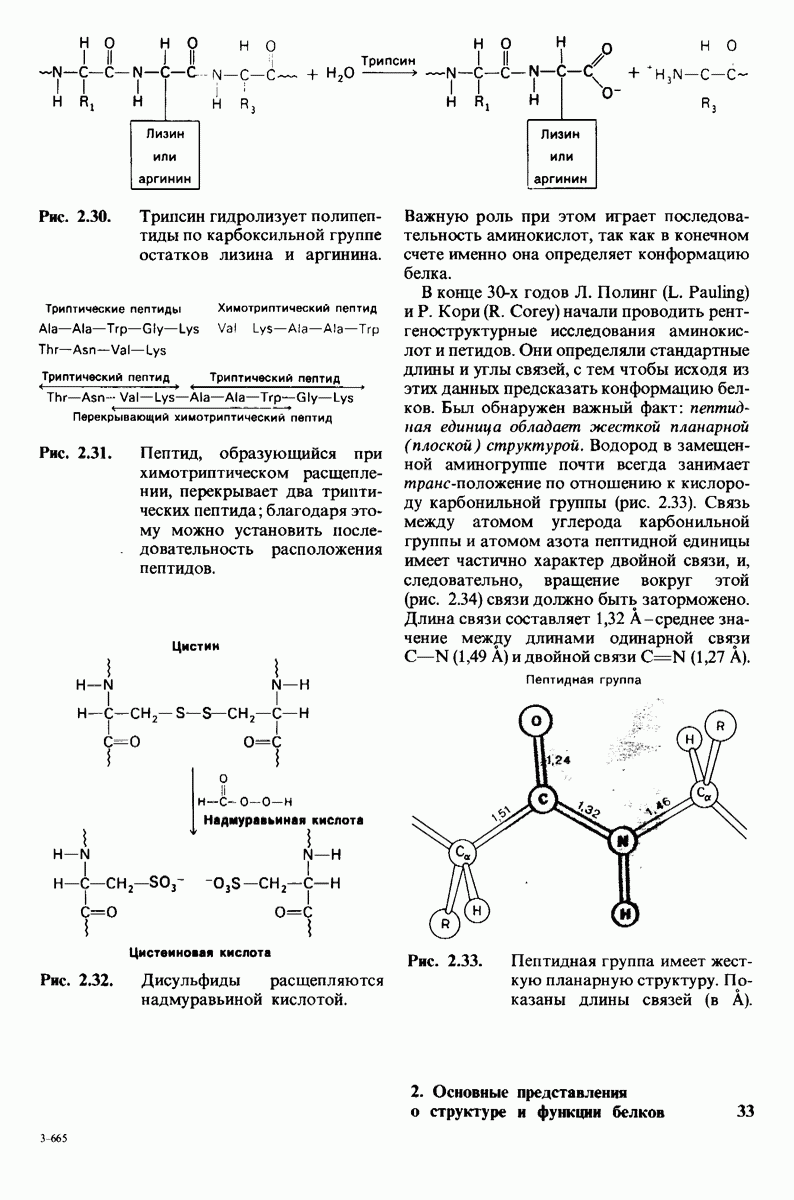
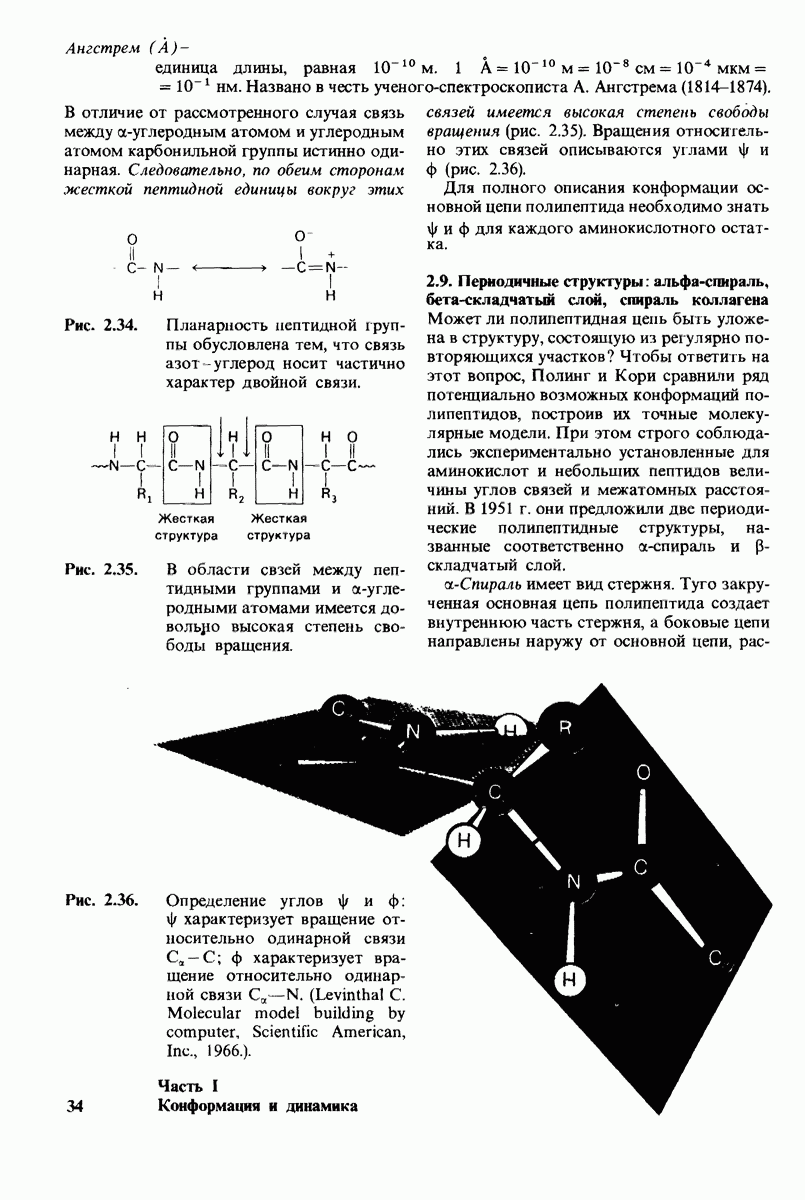
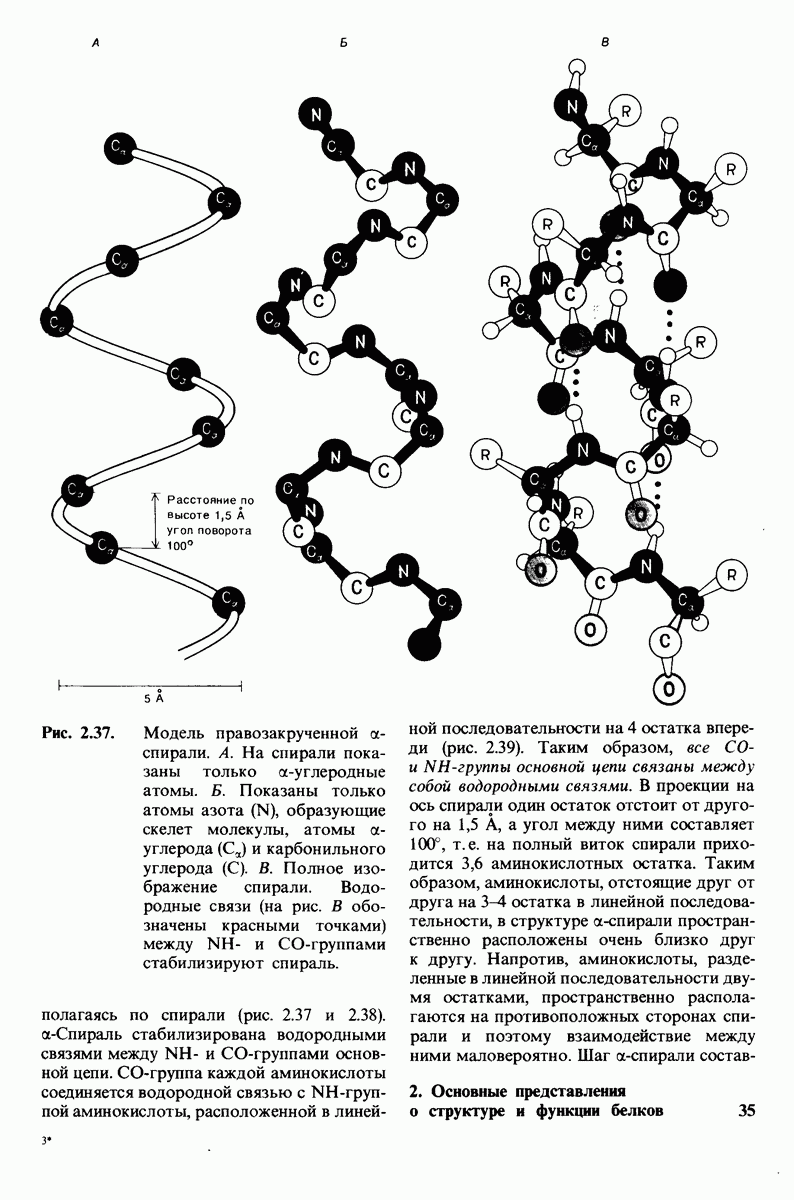
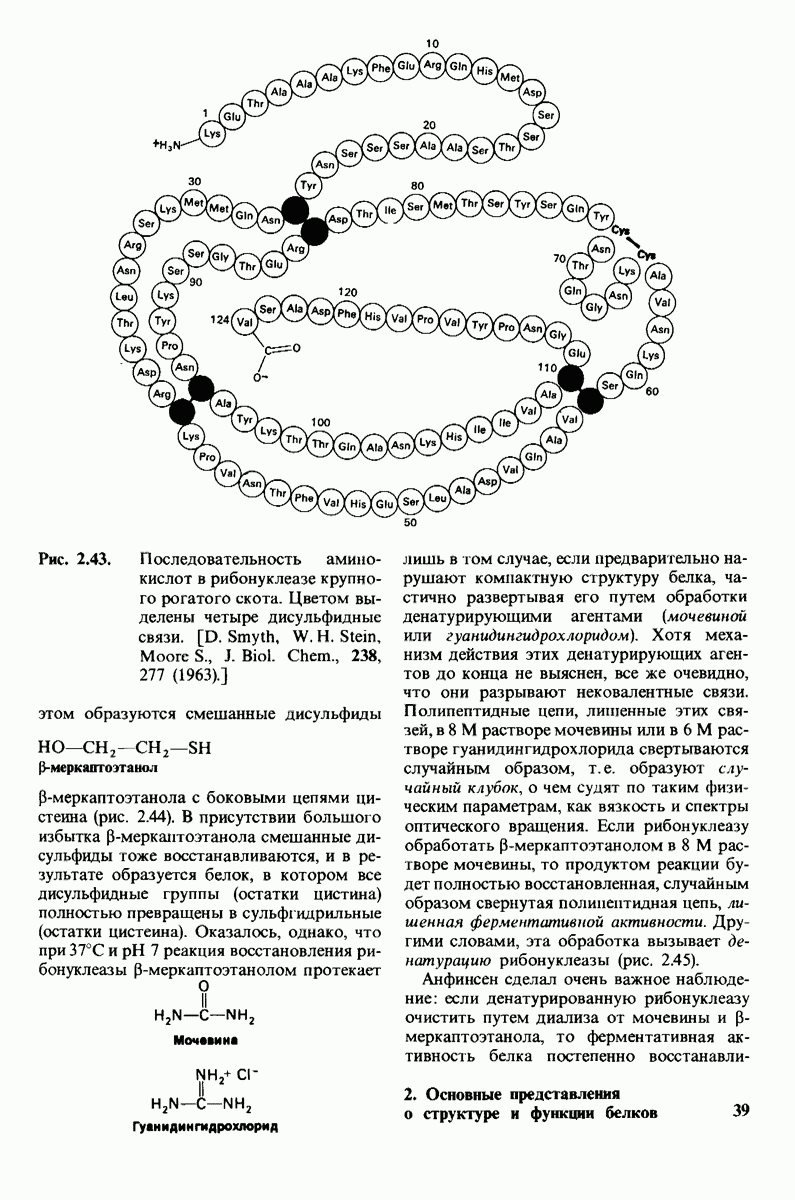
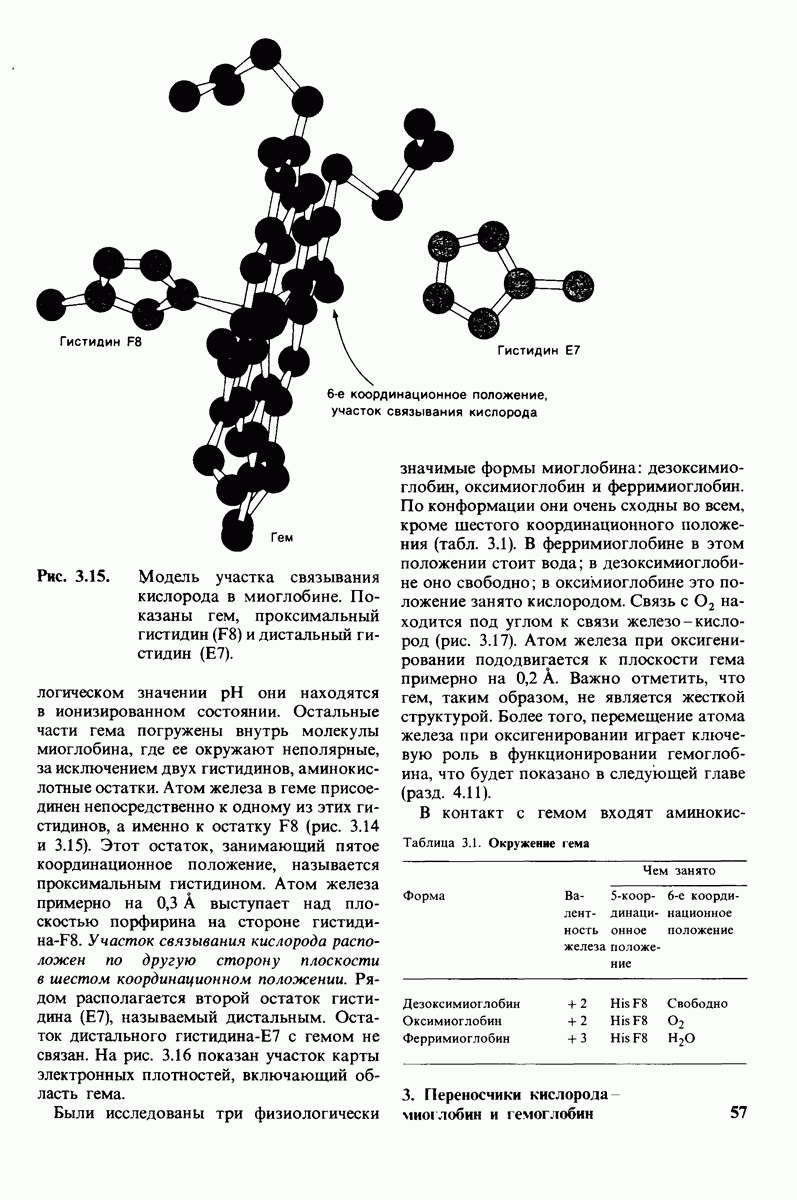
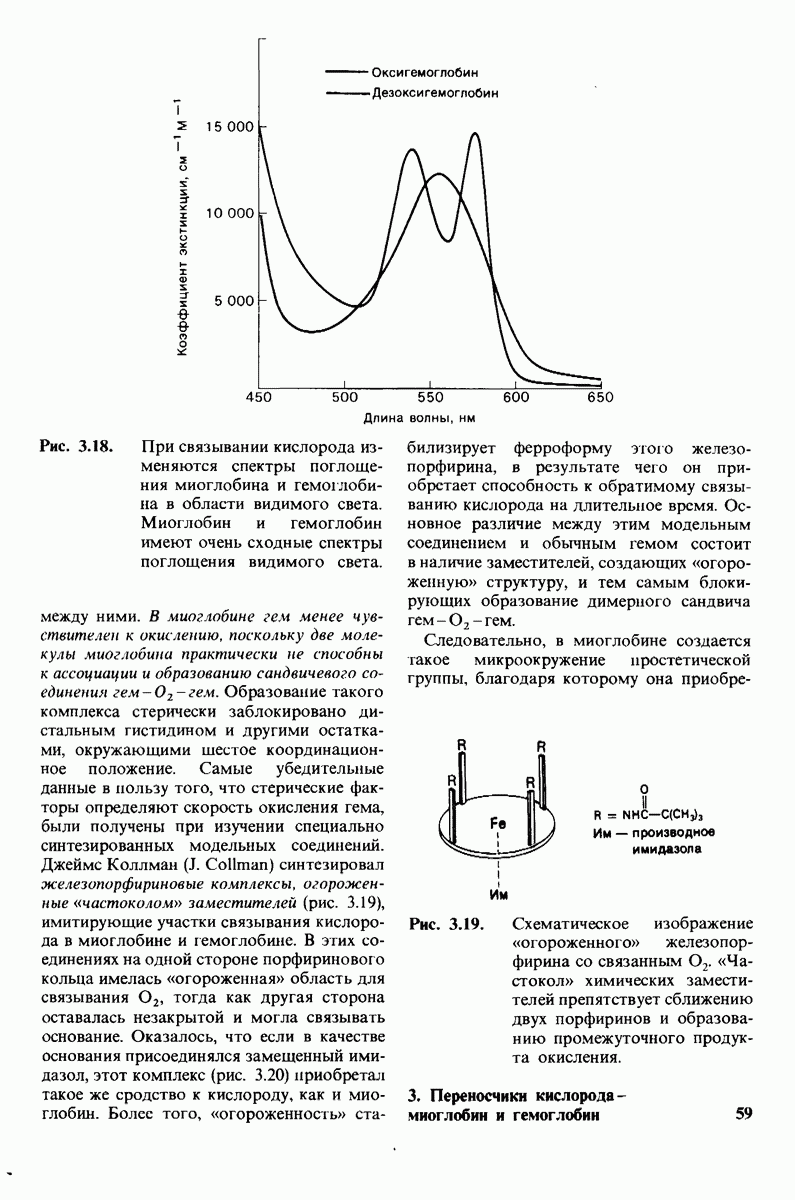
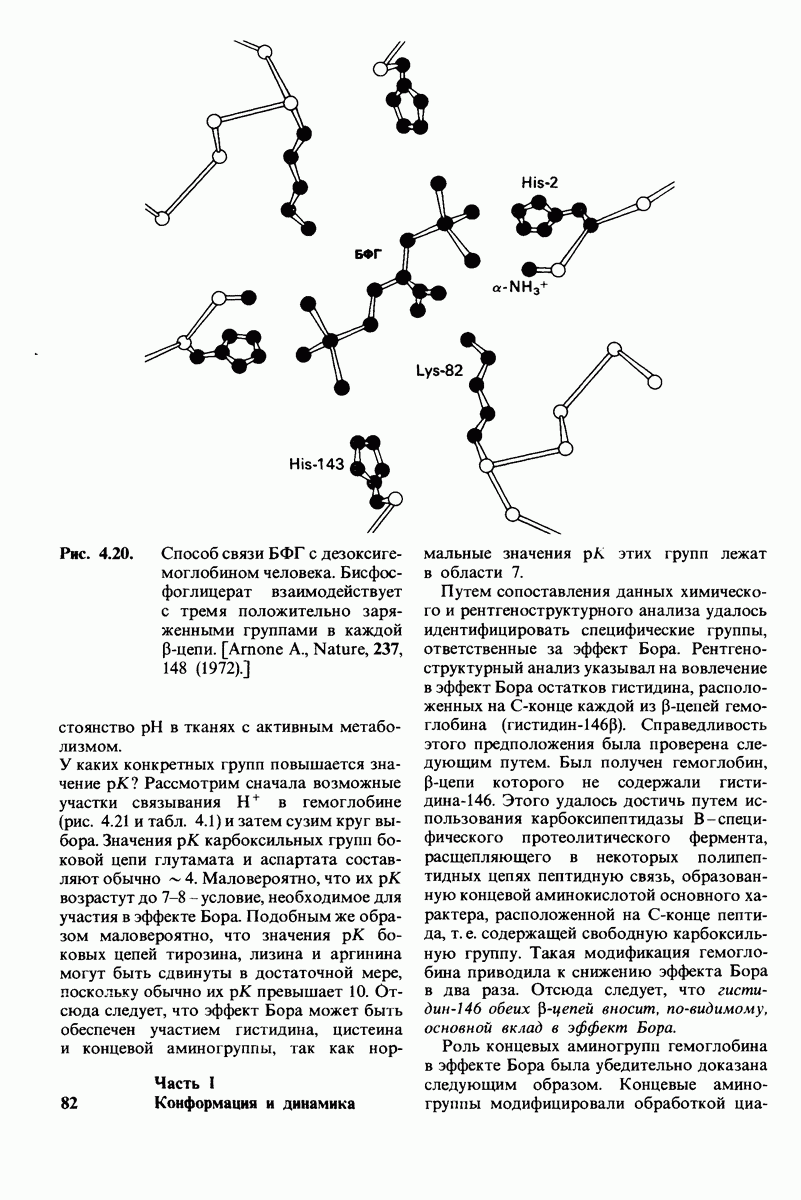
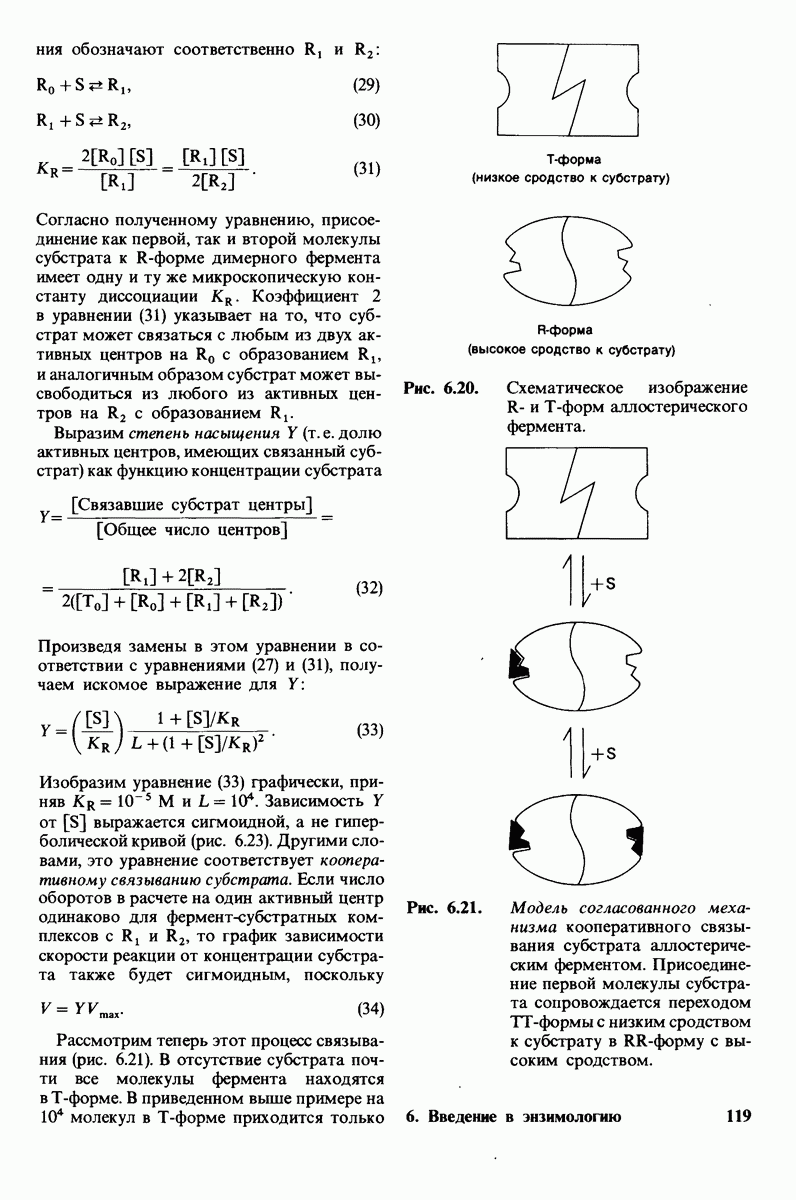
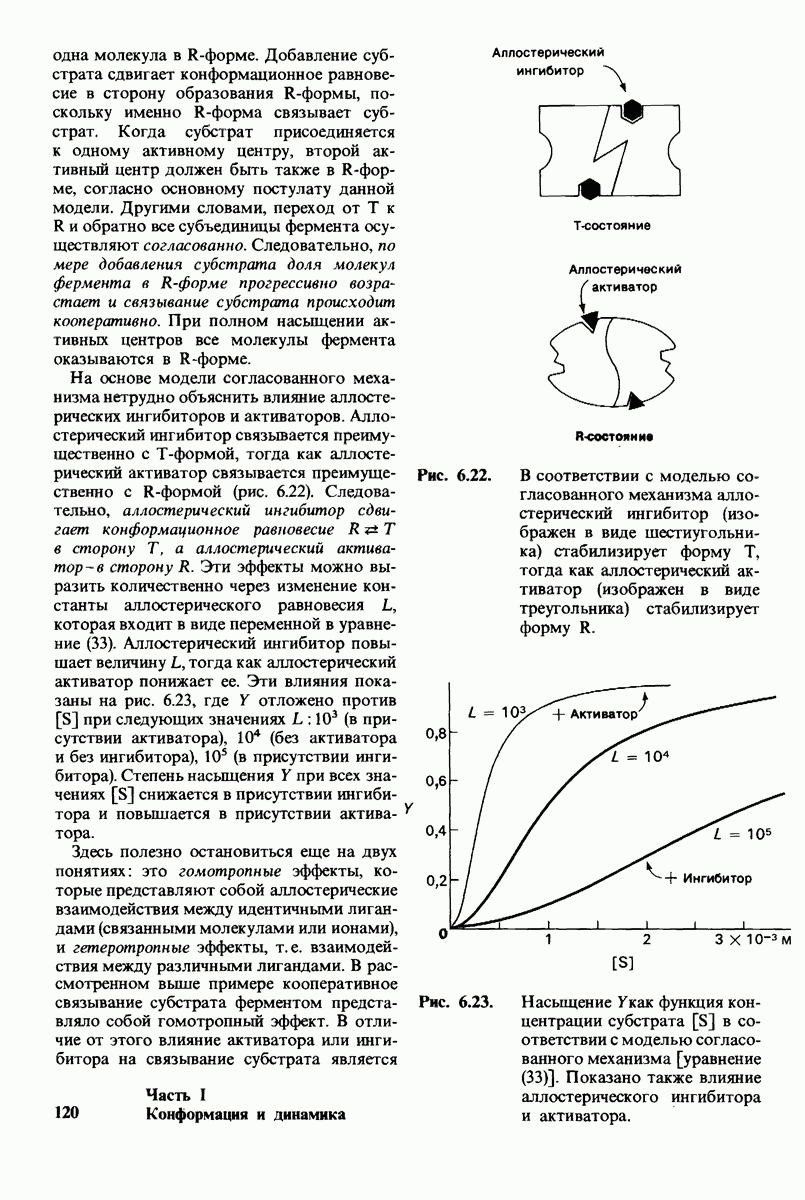
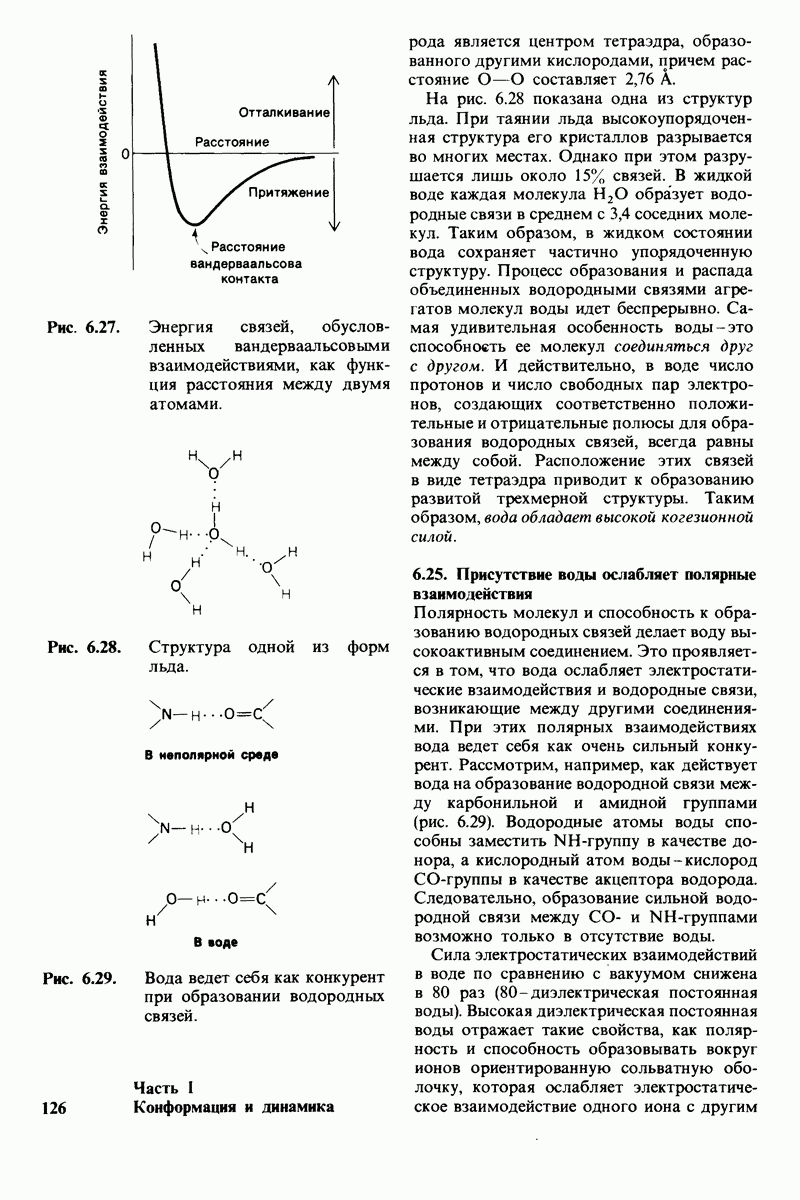
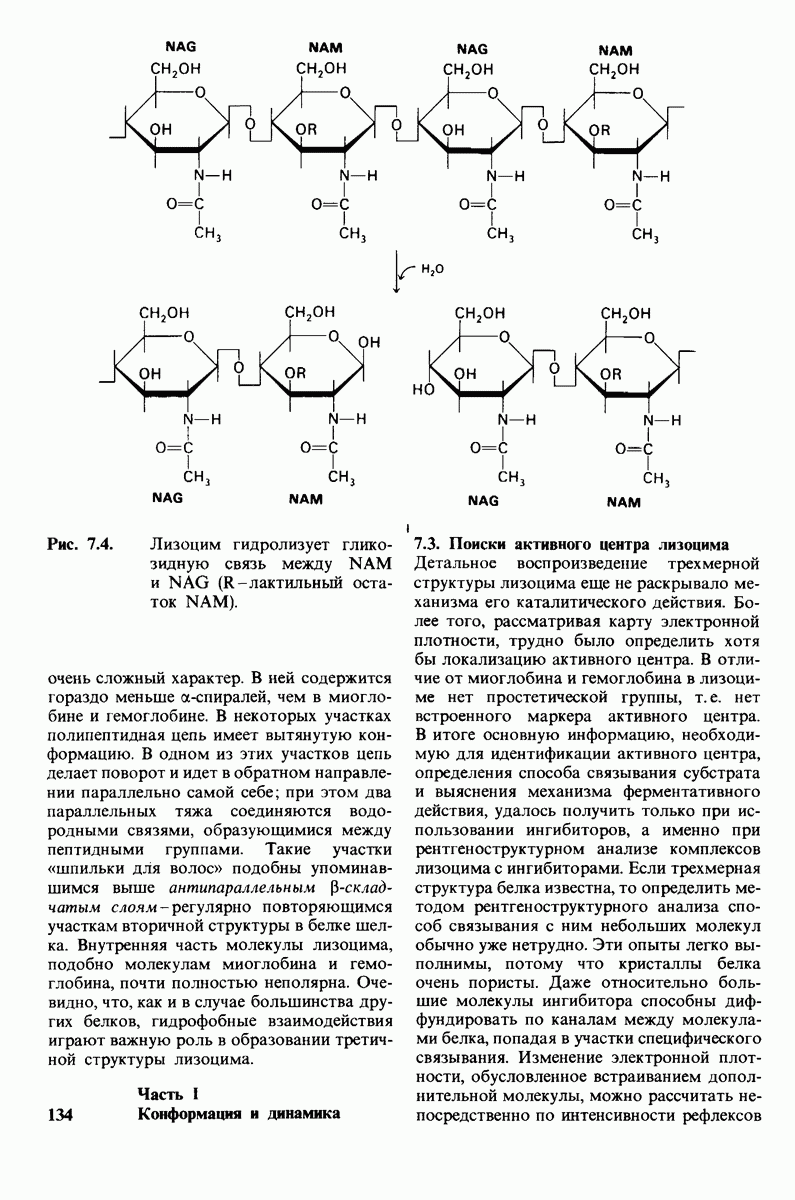
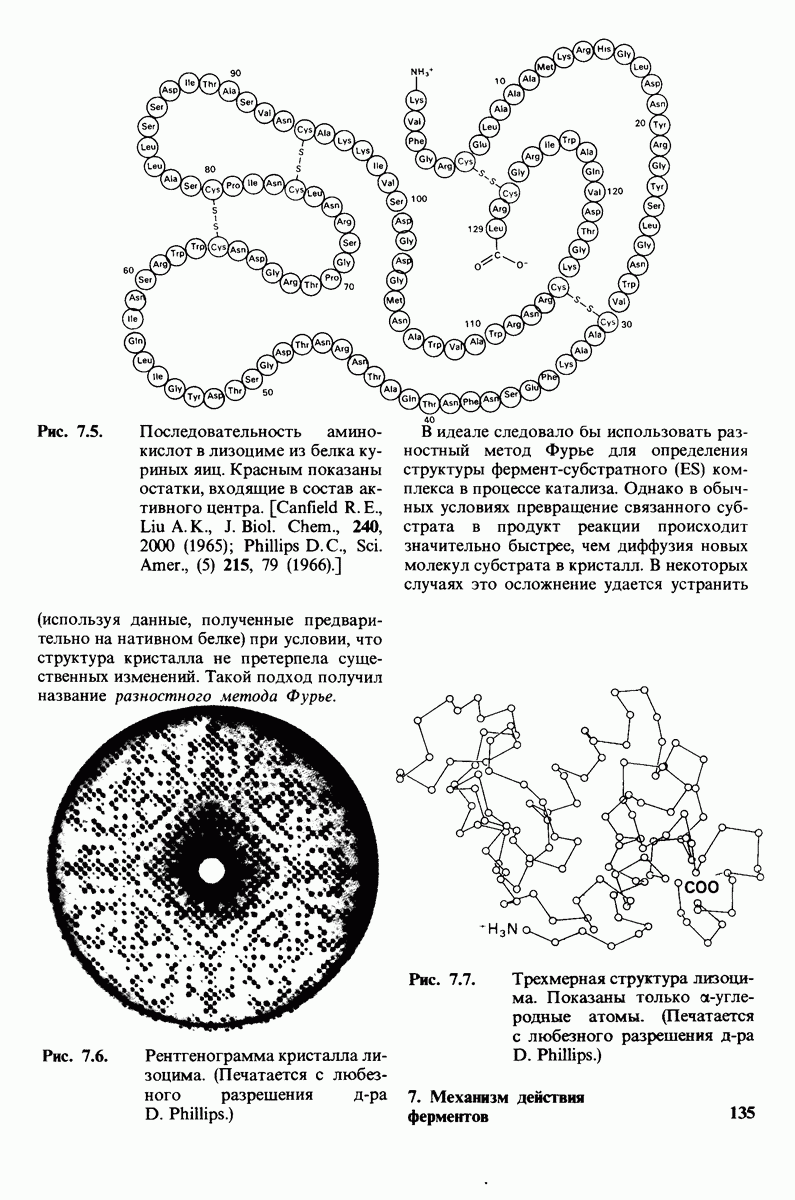
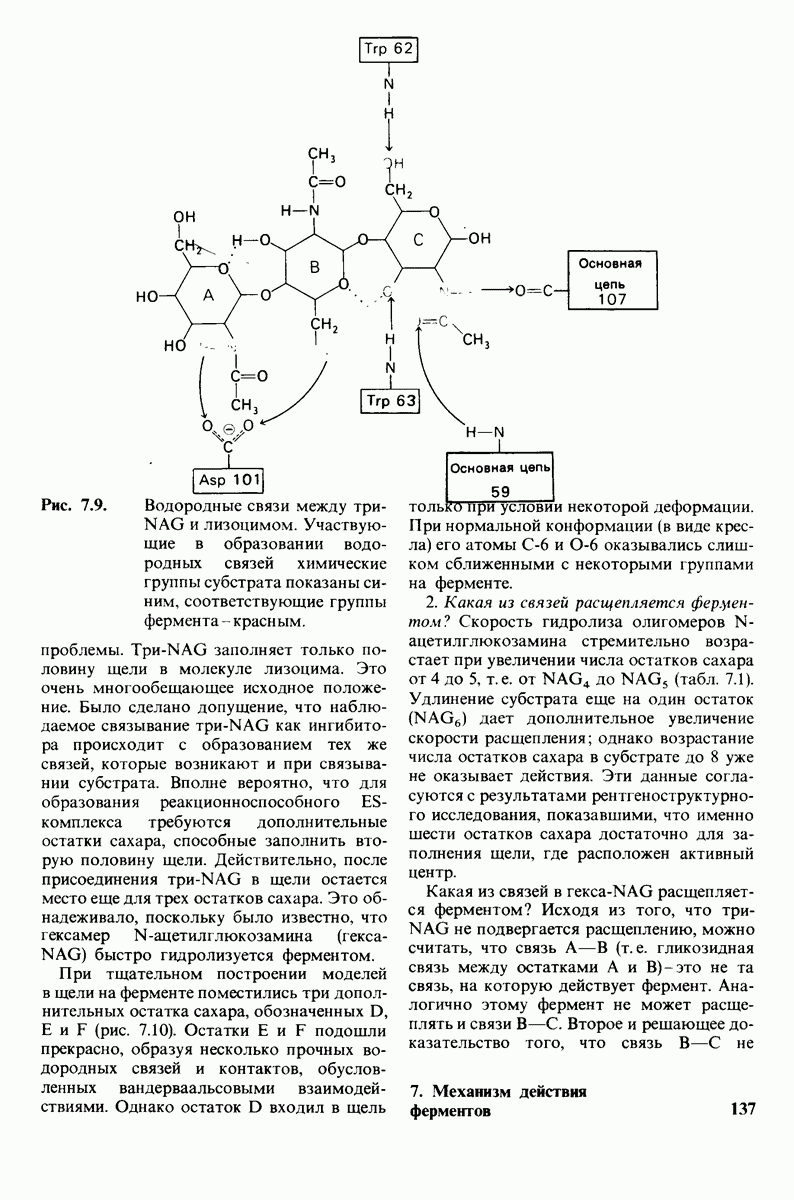
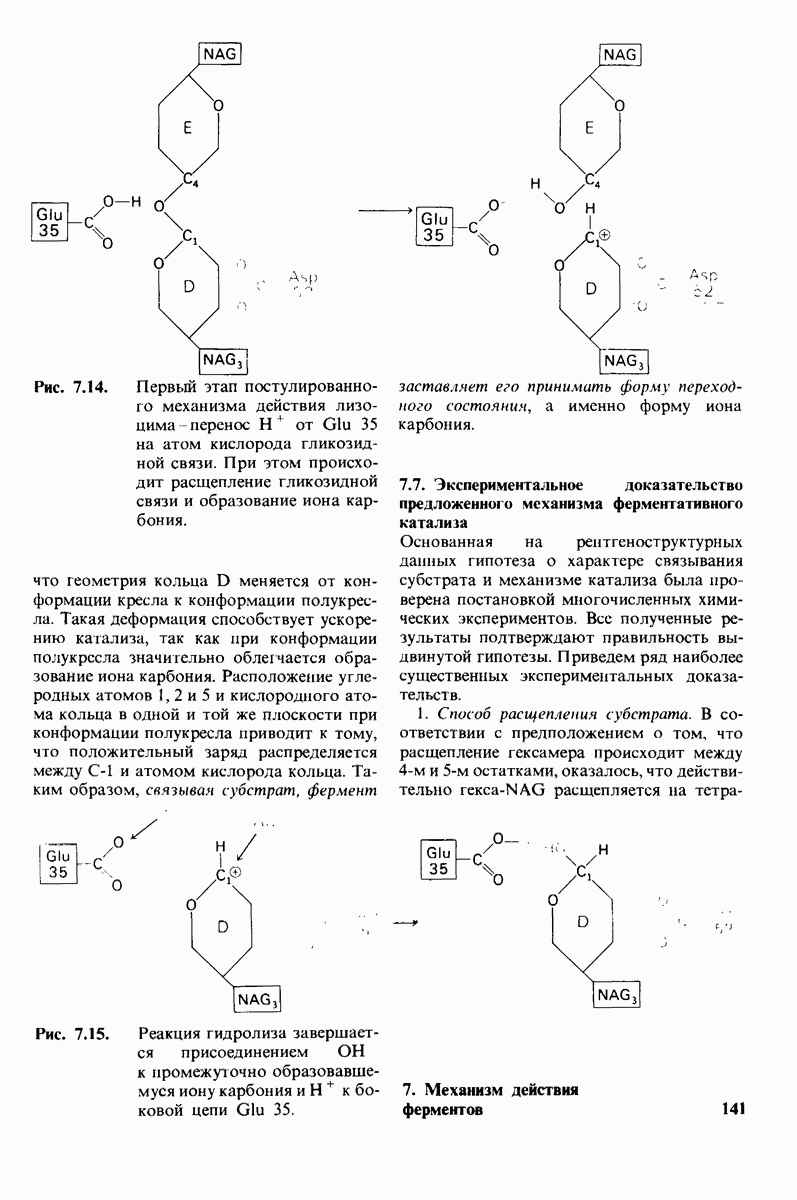
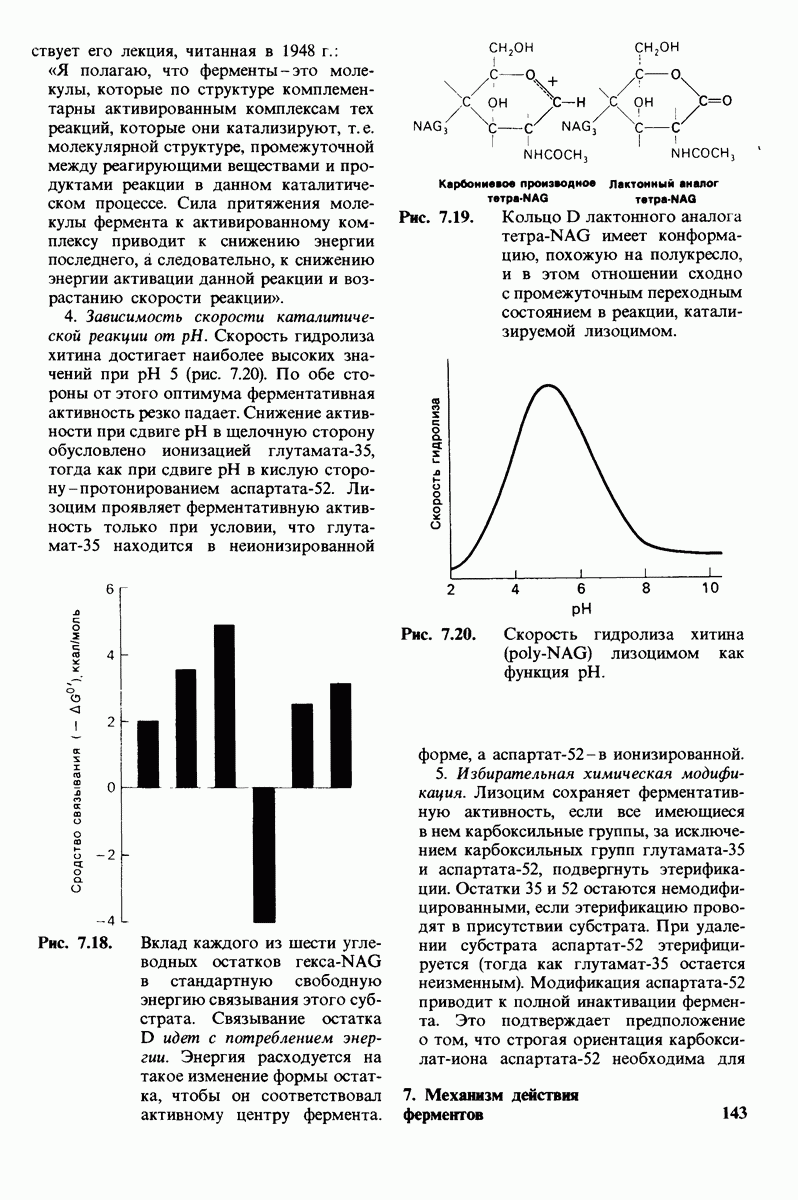
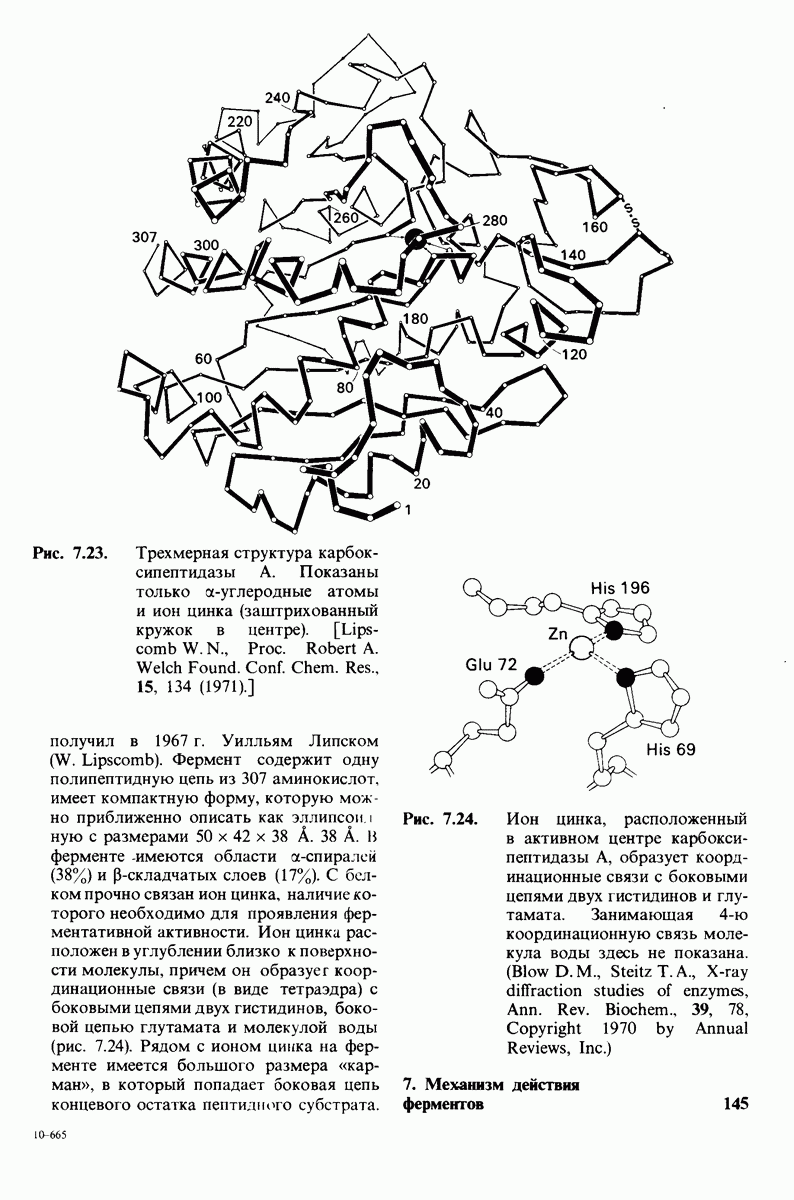
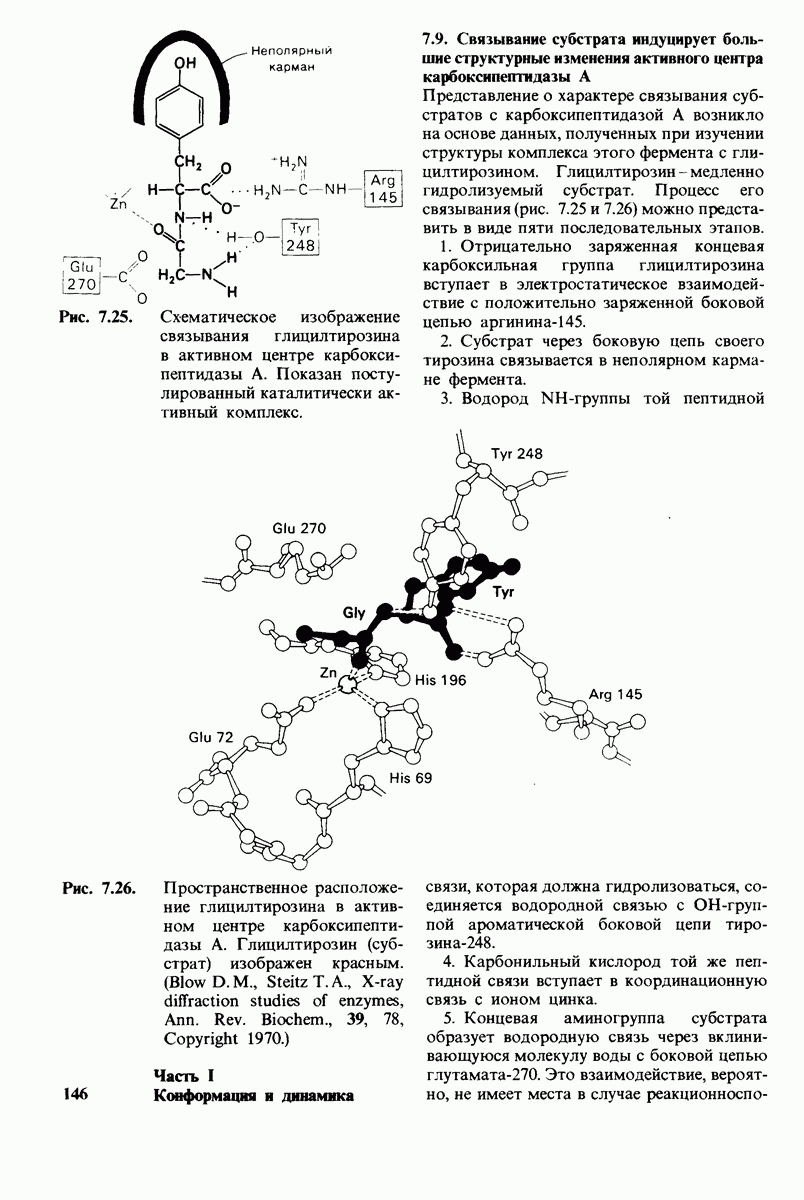
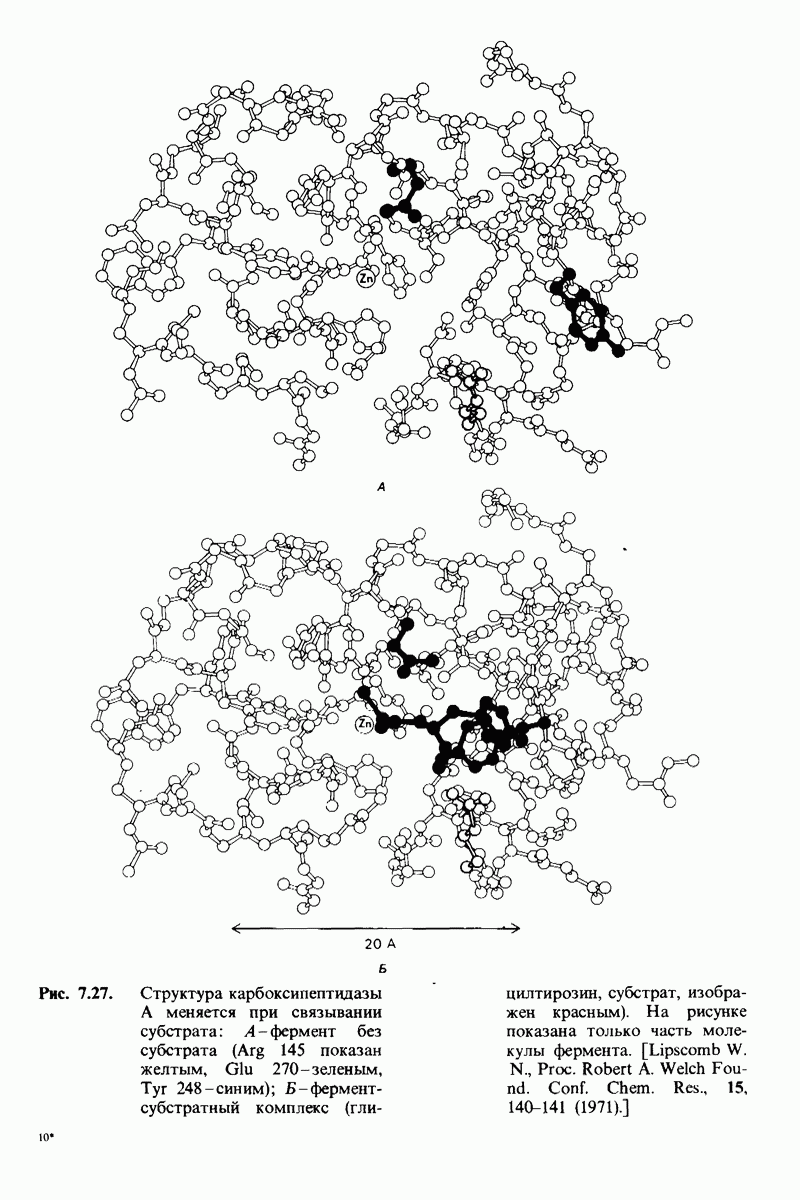
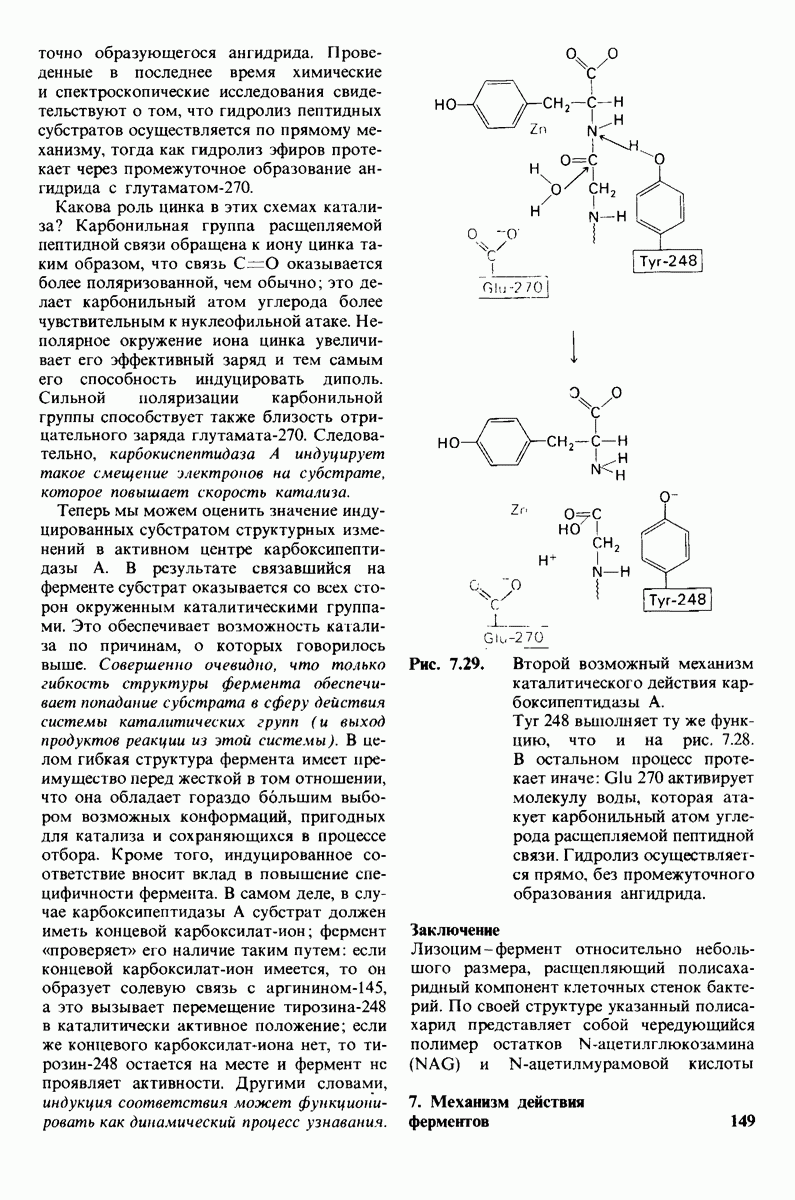
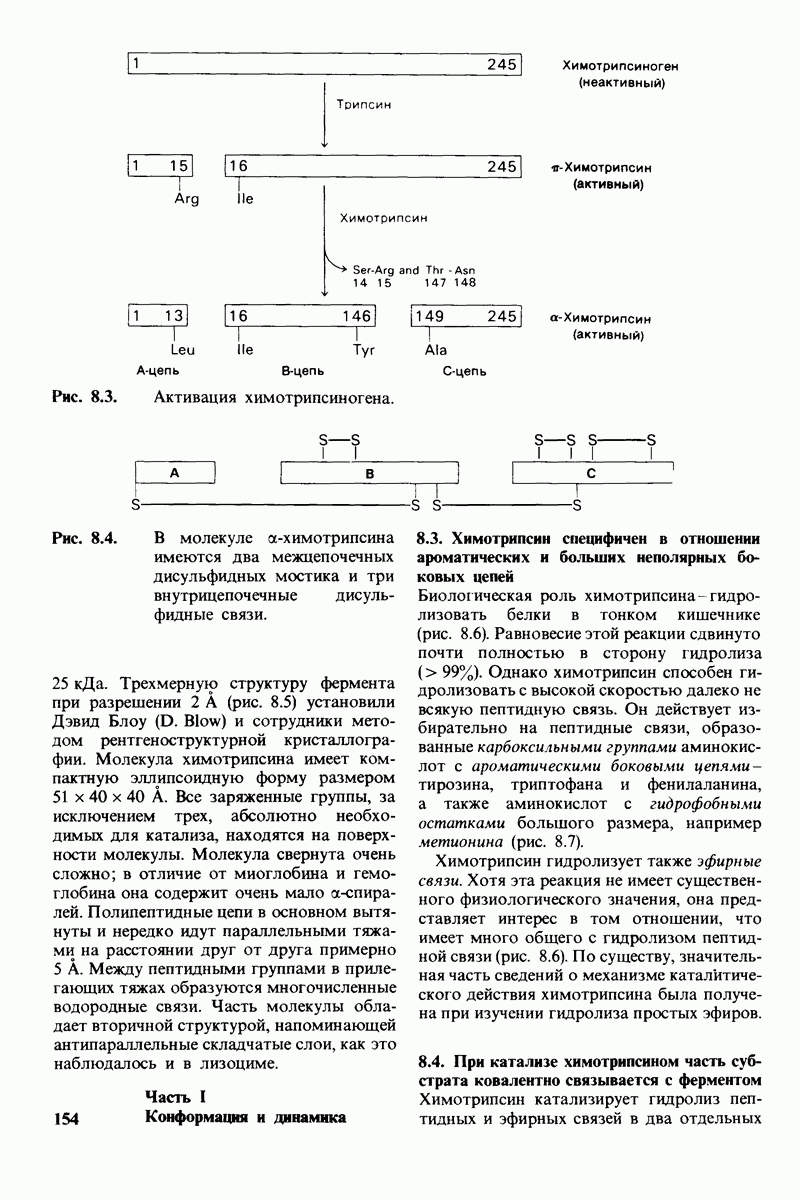
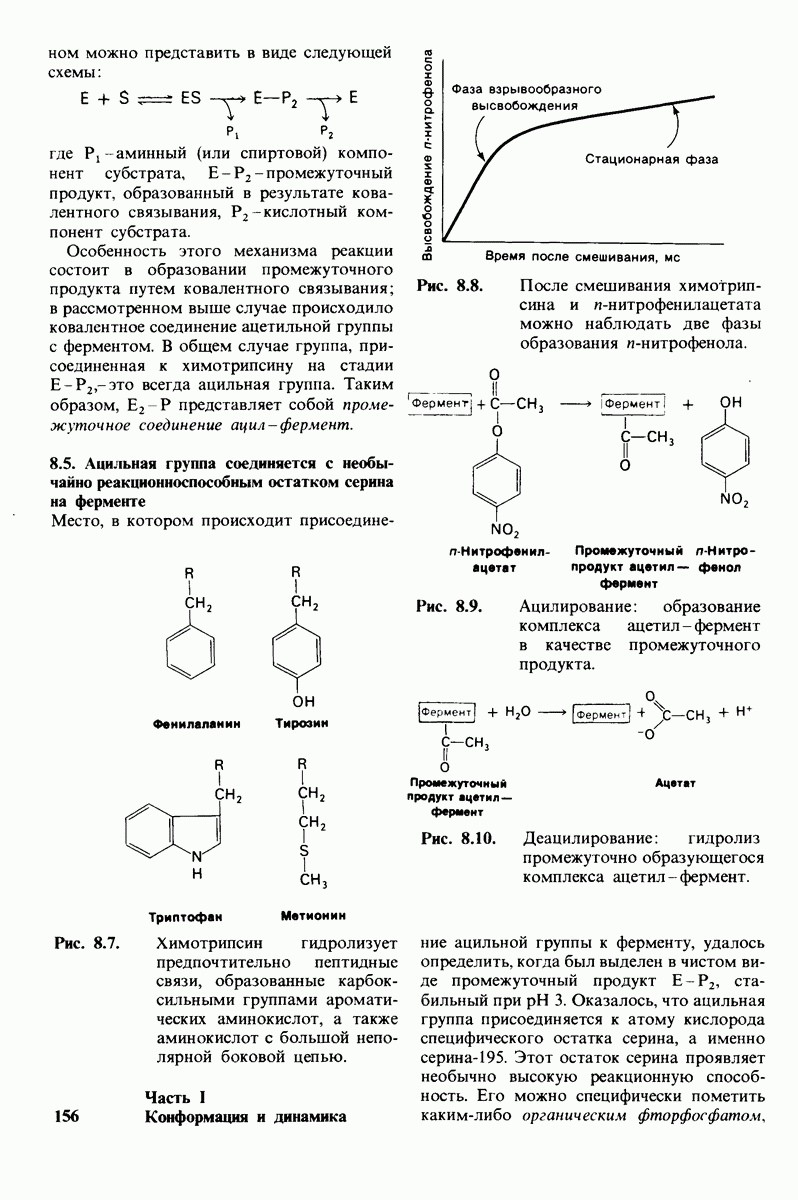
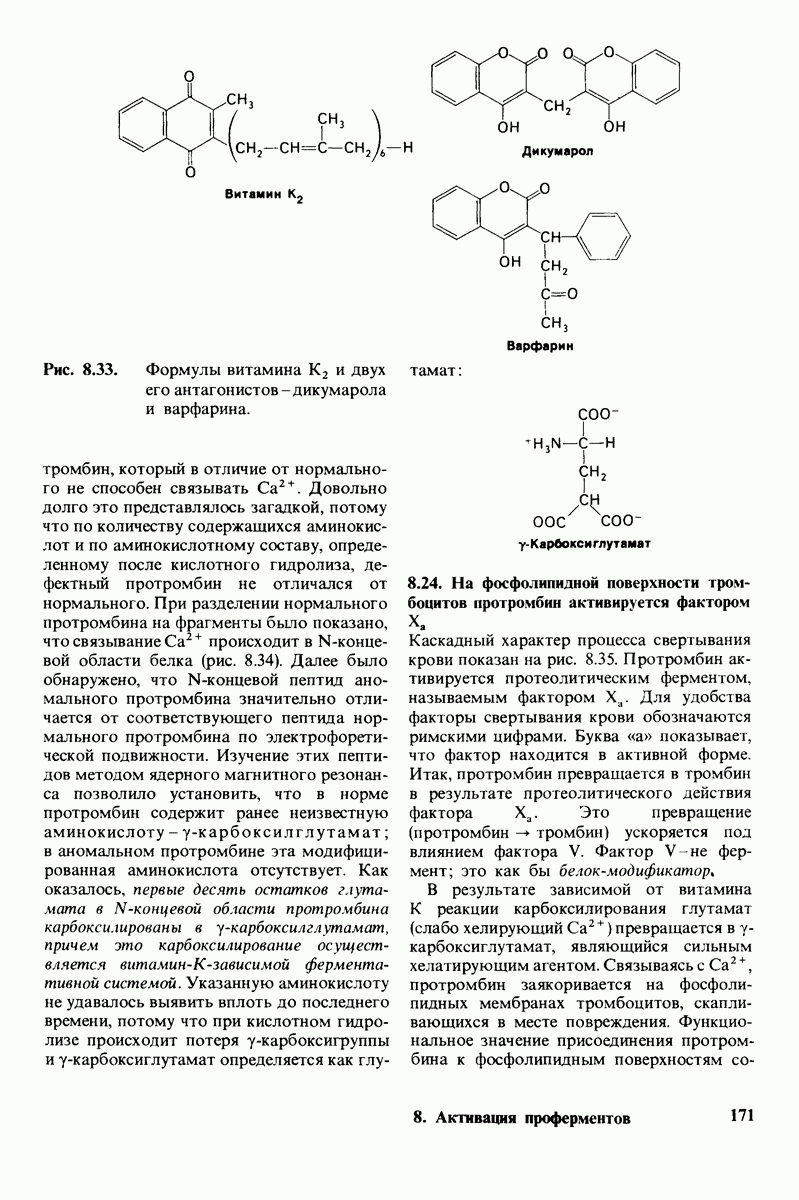
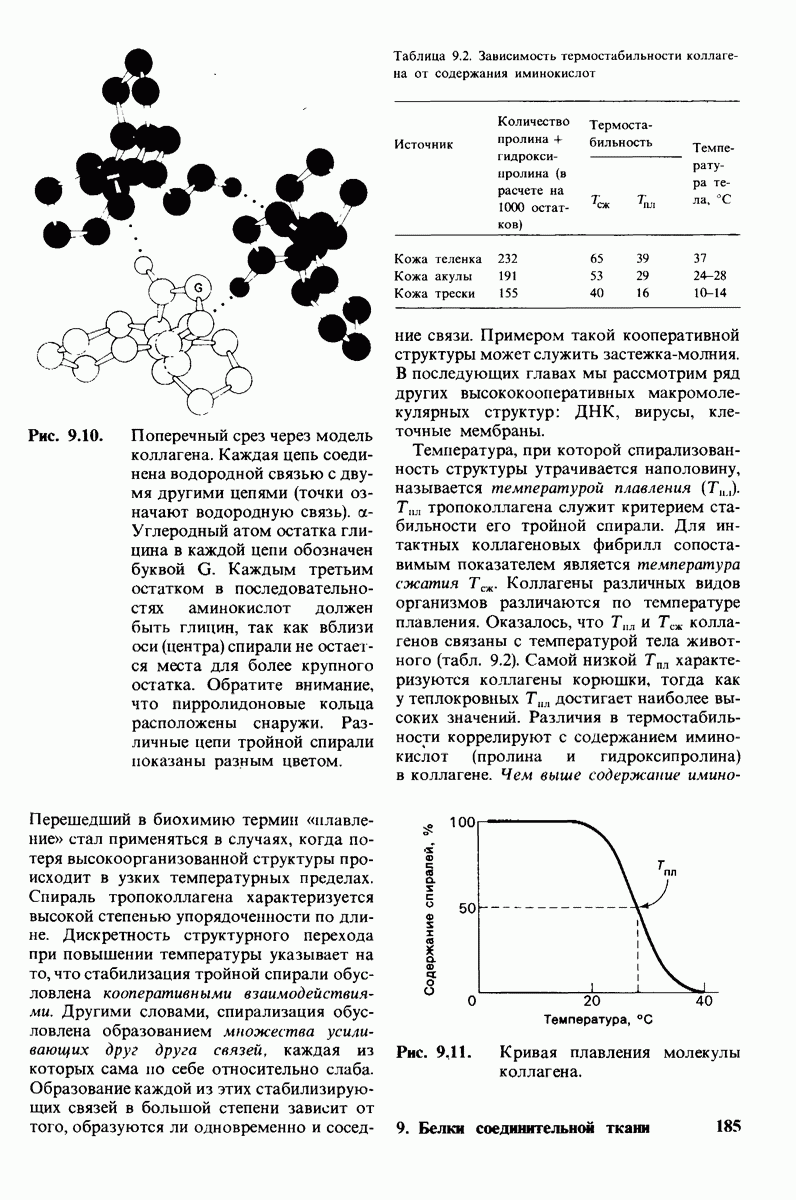
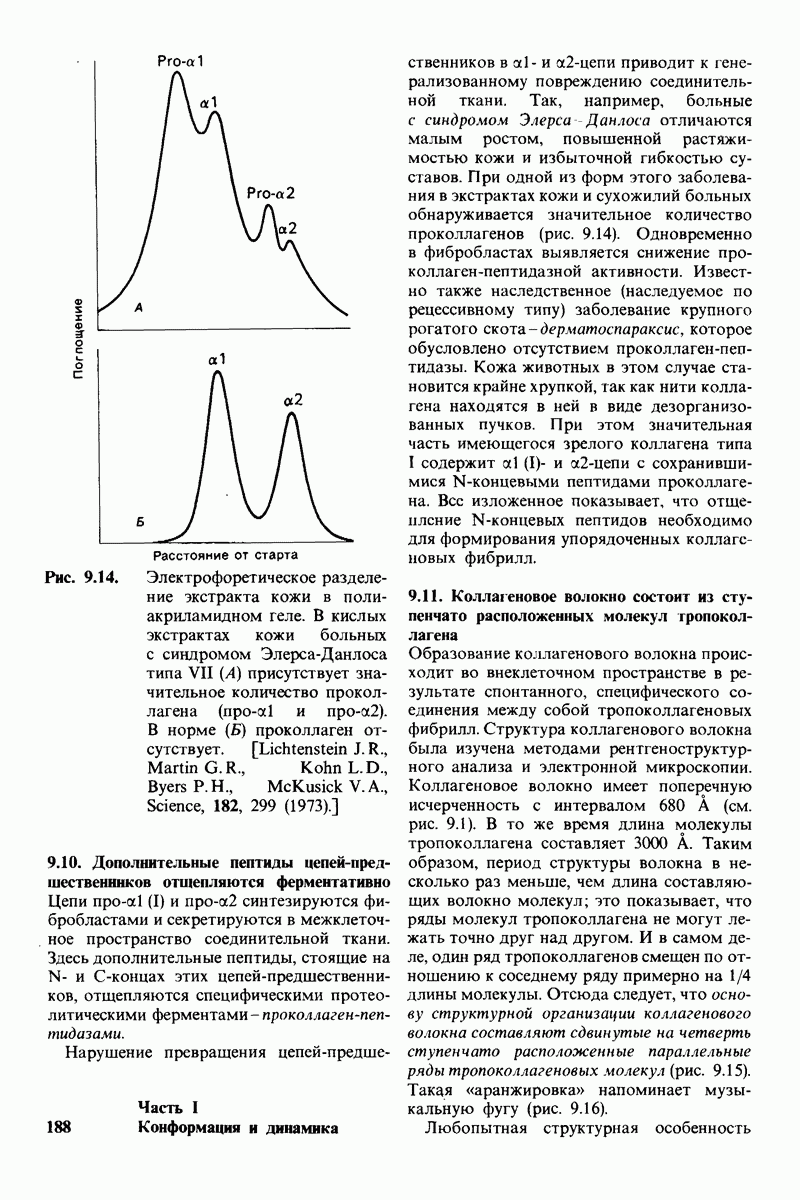
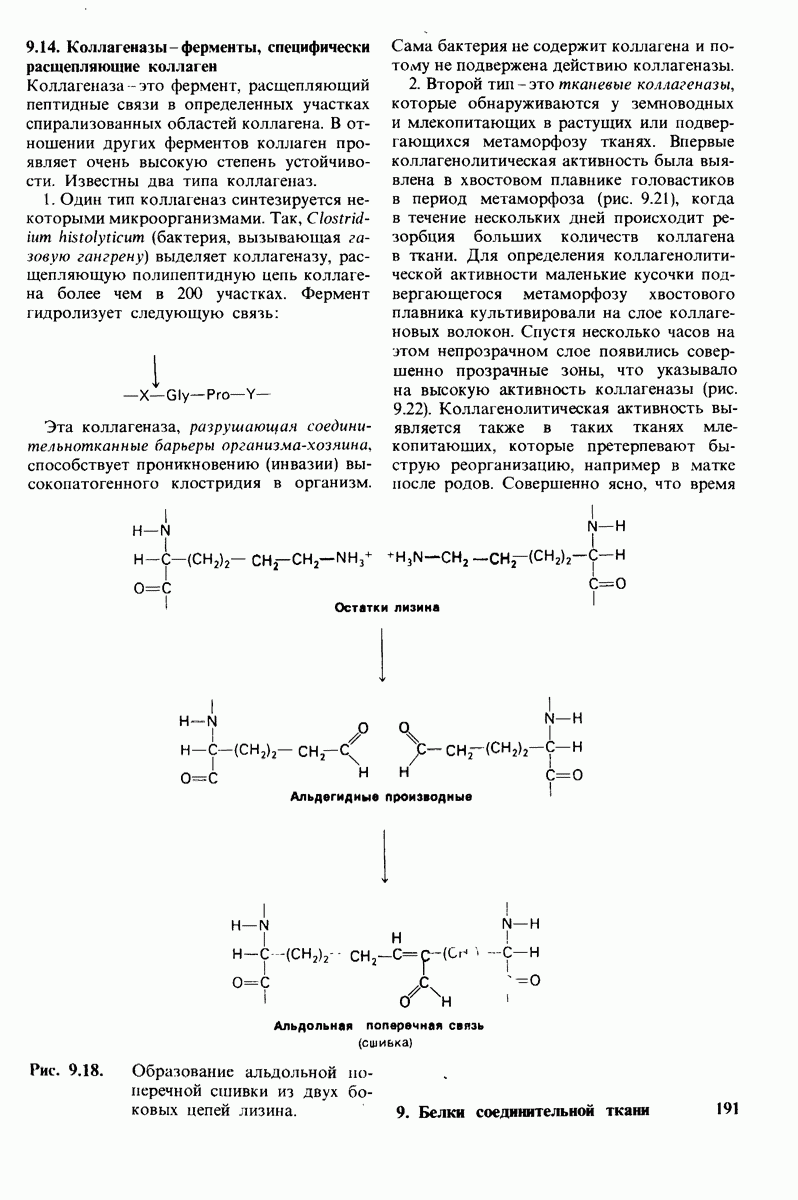
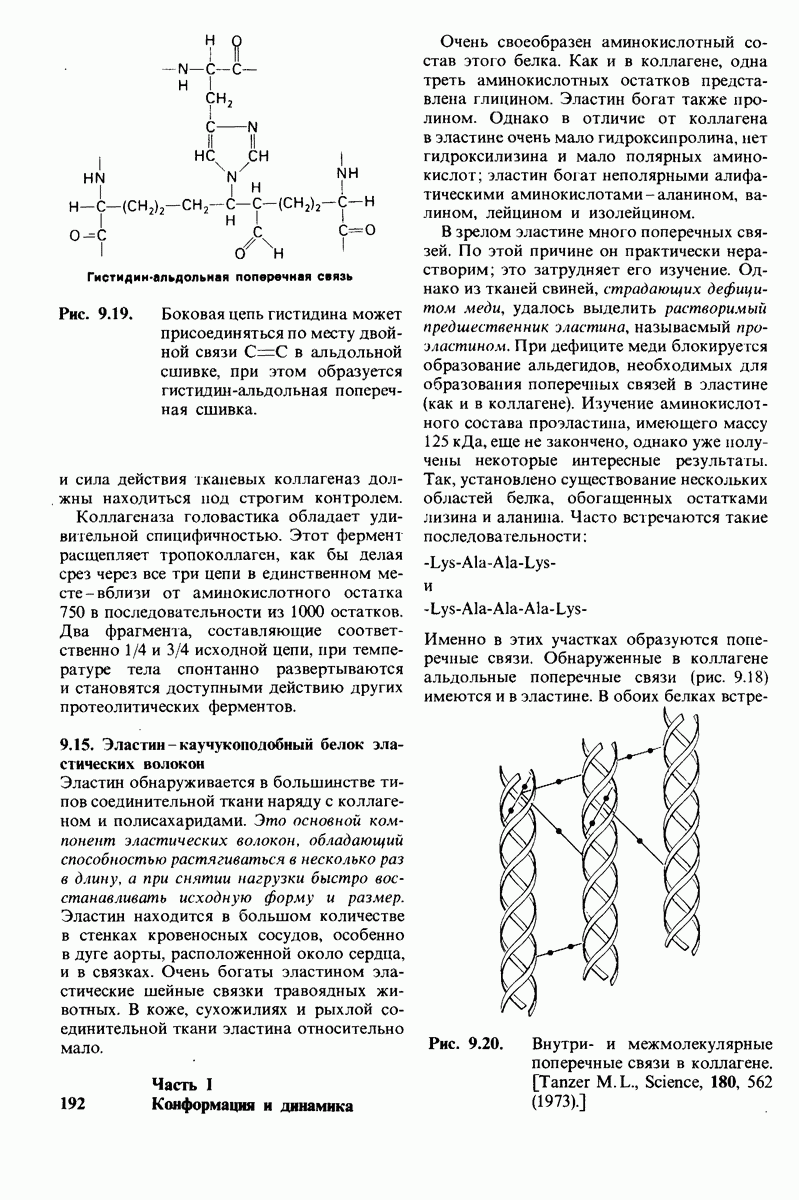
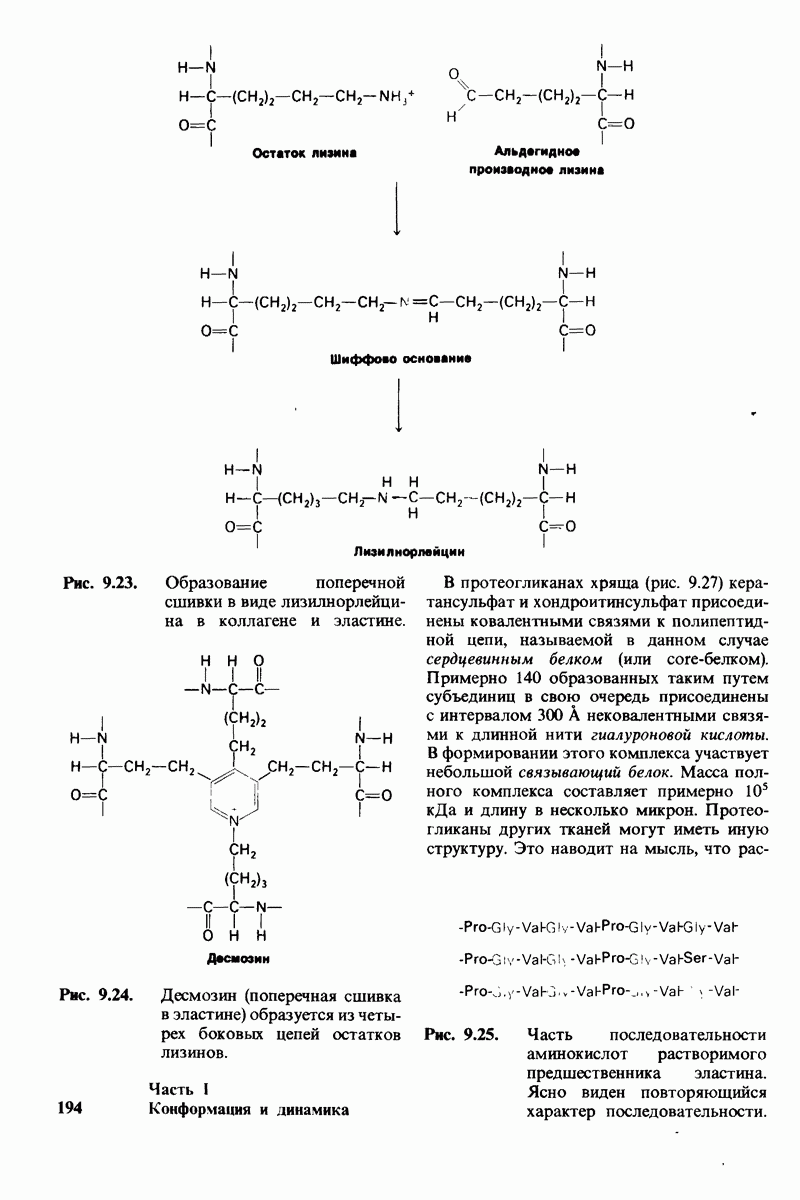
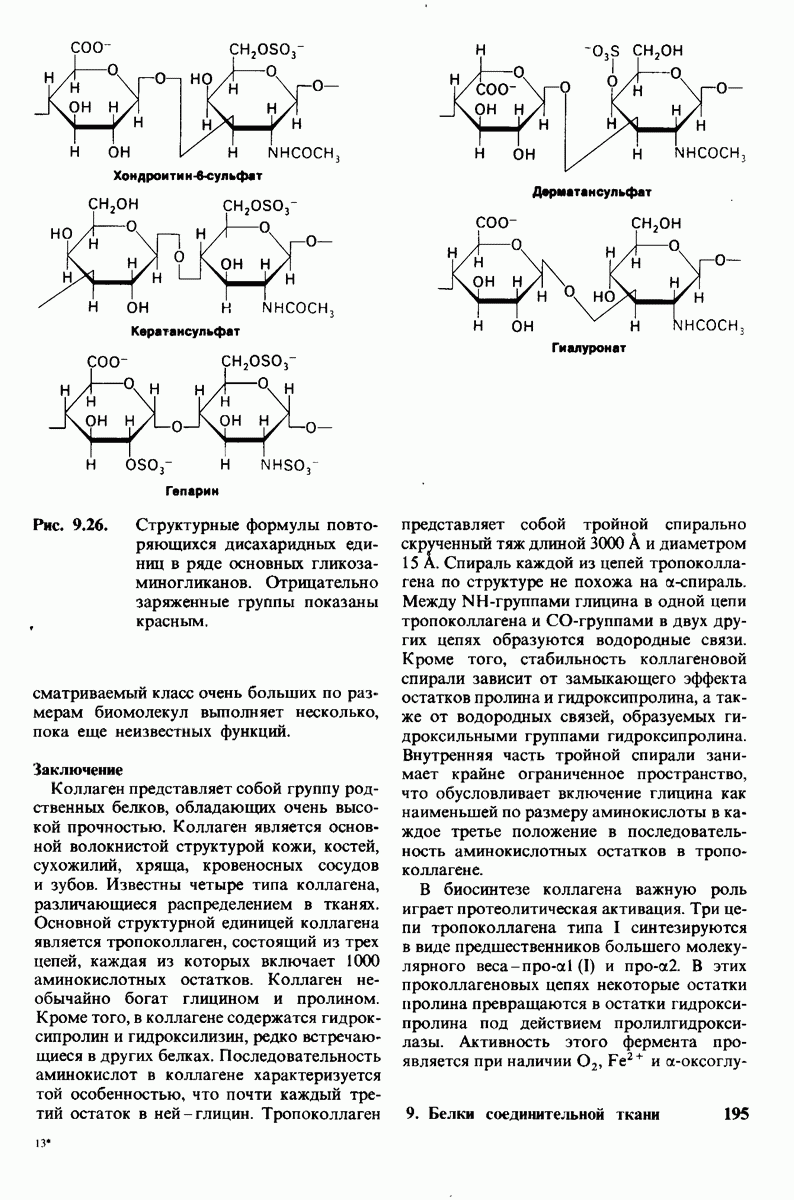
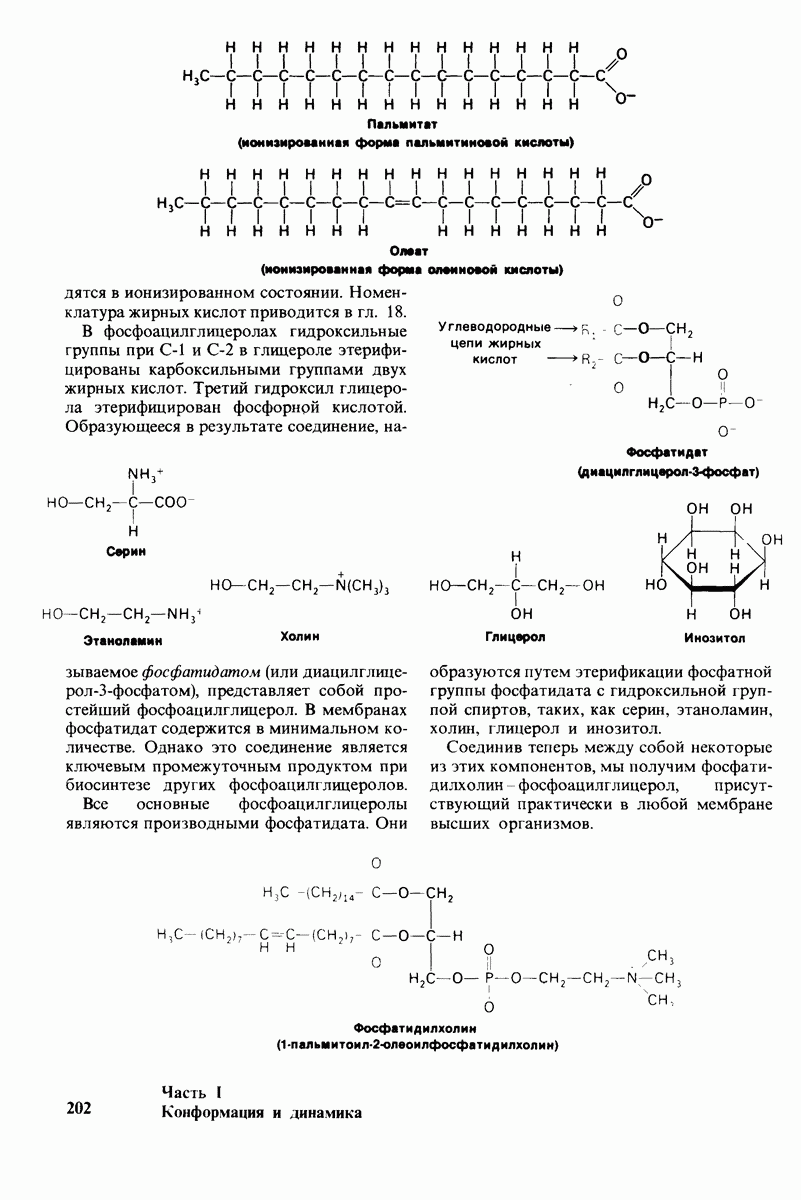
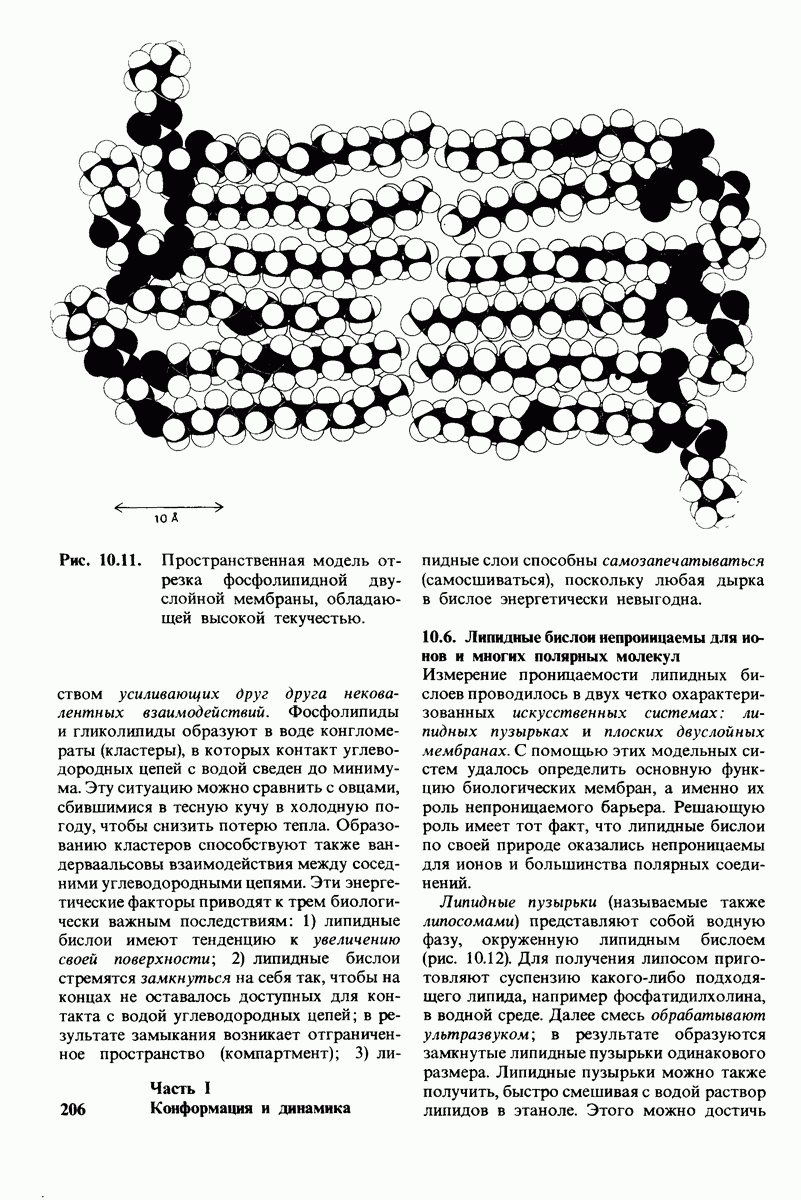
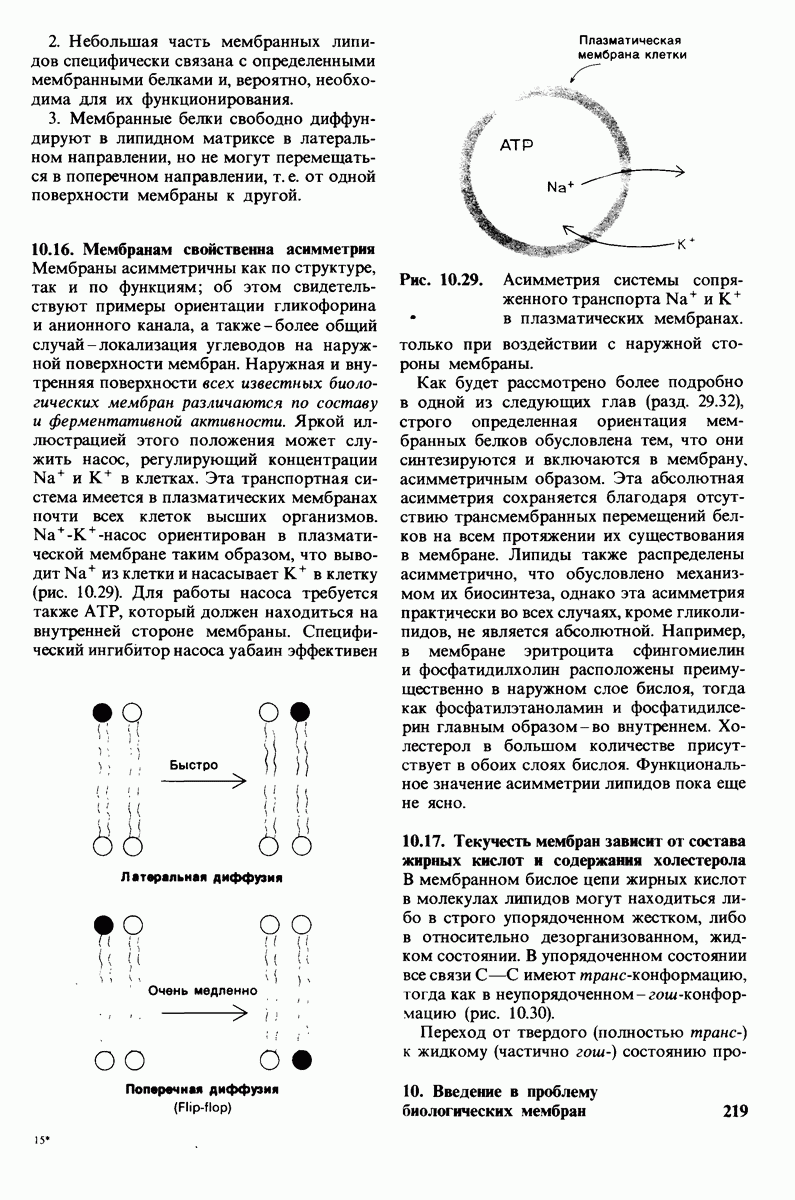
8.12. Ингибитор панкреатического трипсина прочно связывается в активном центре трипсина
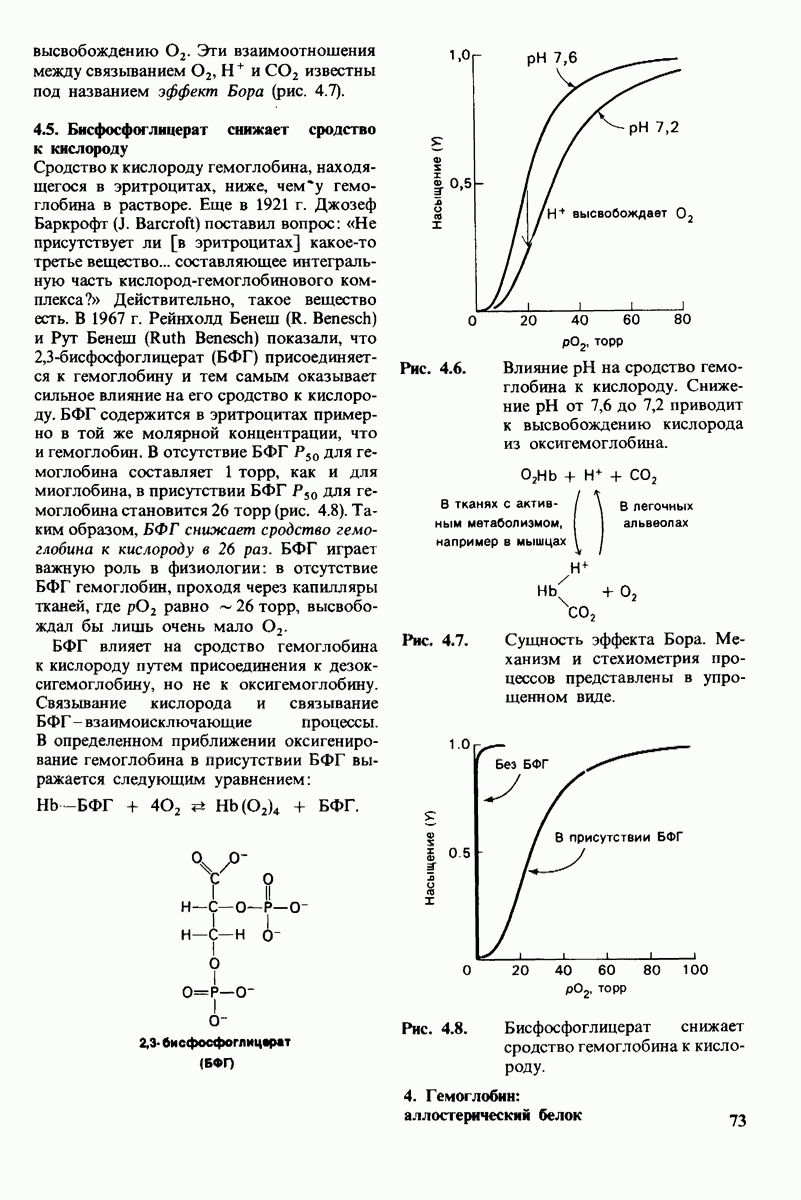
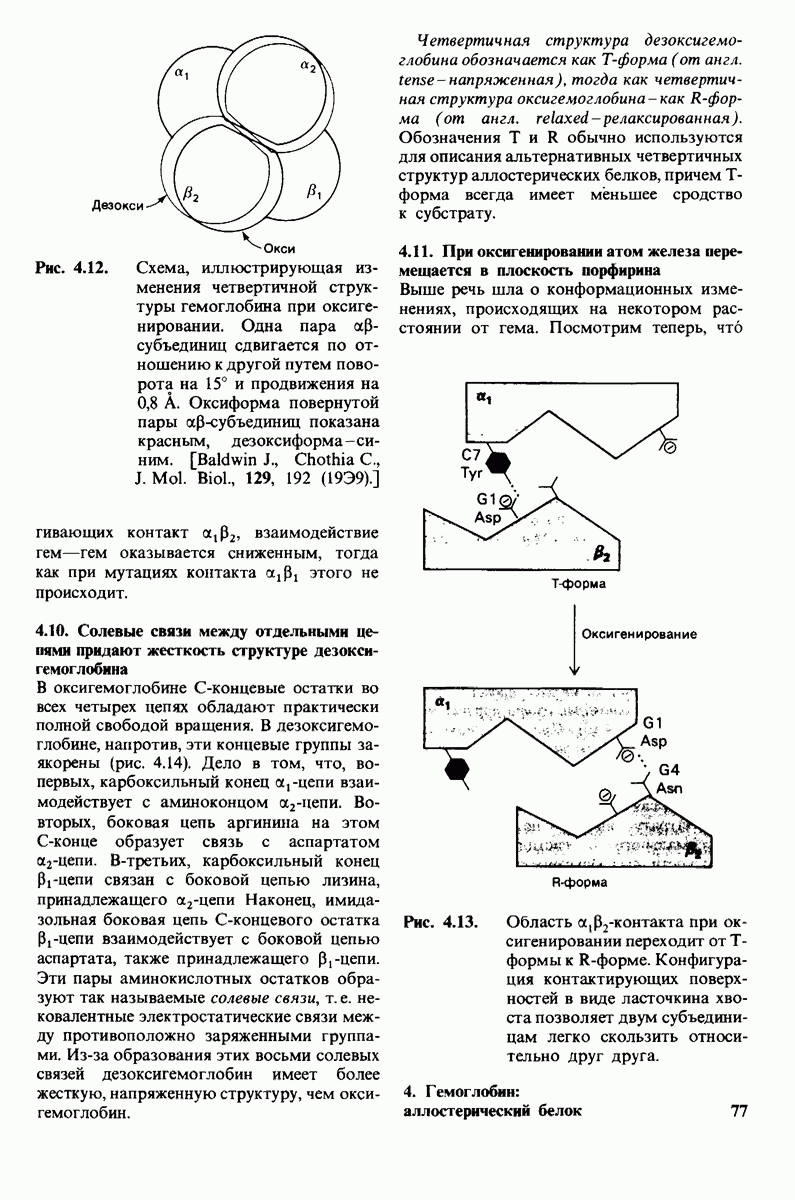
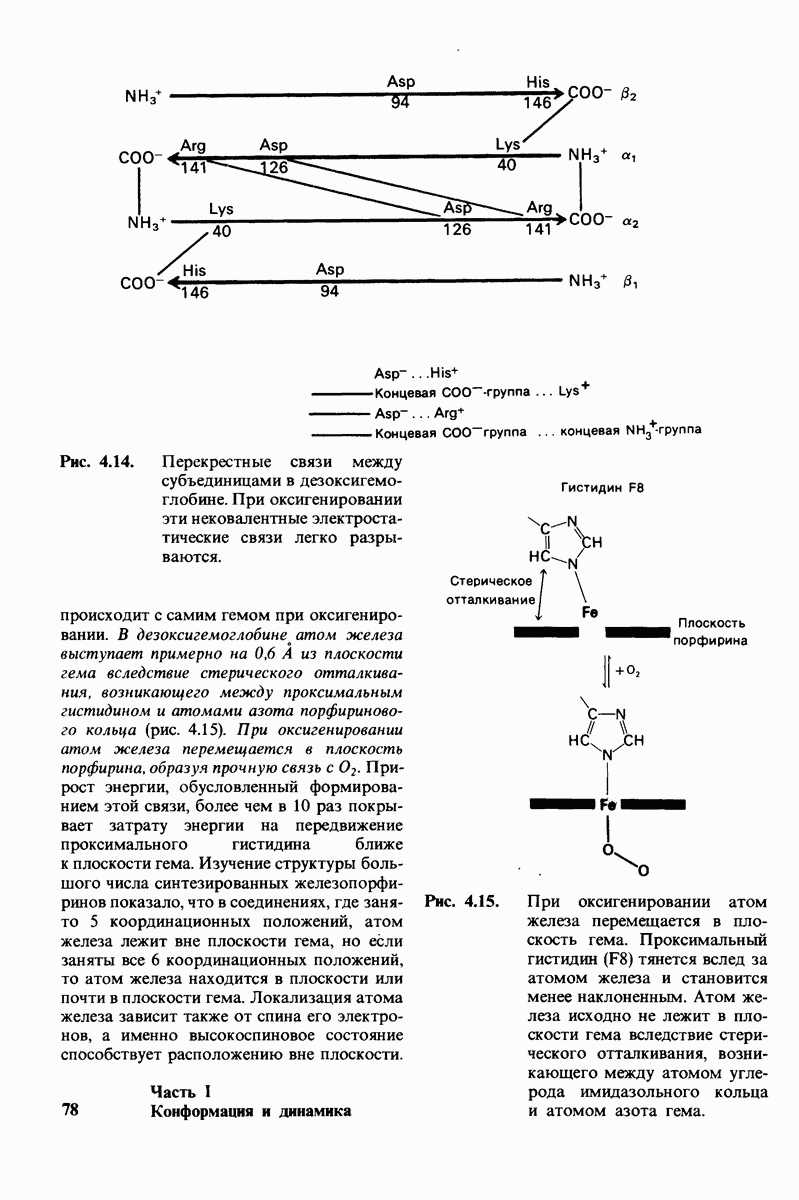
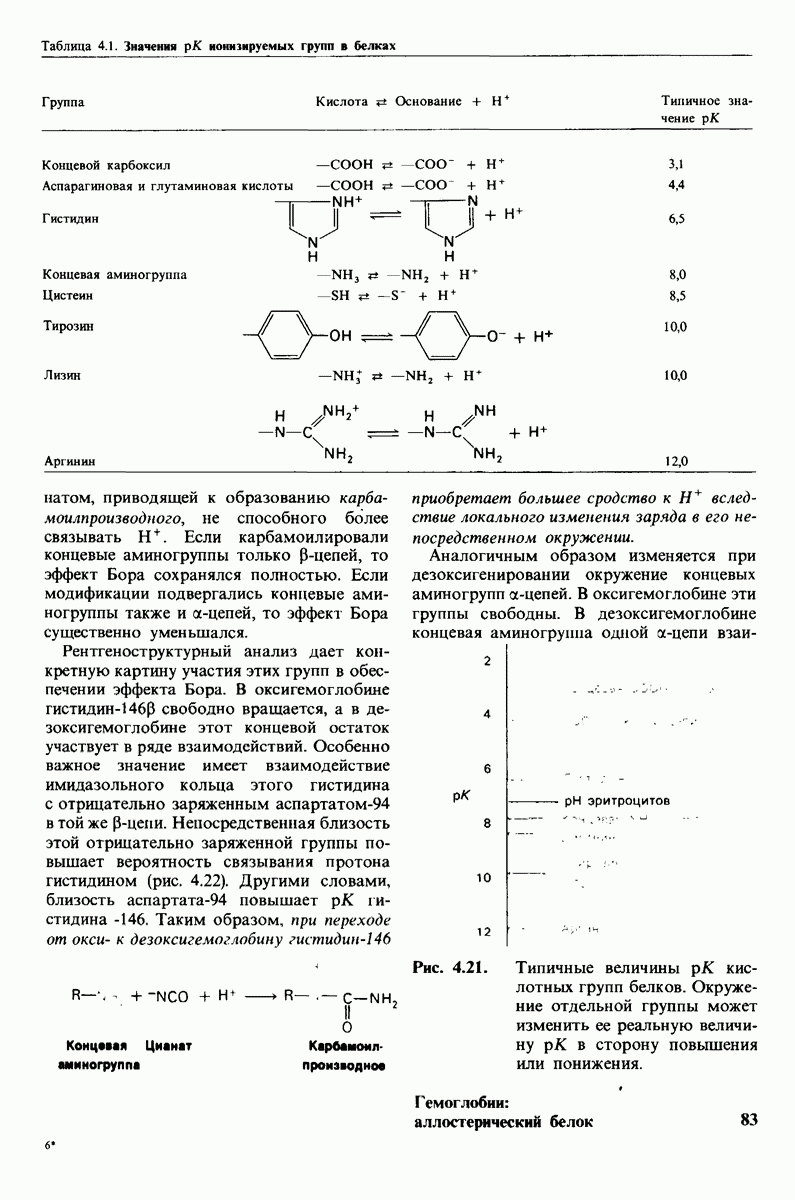
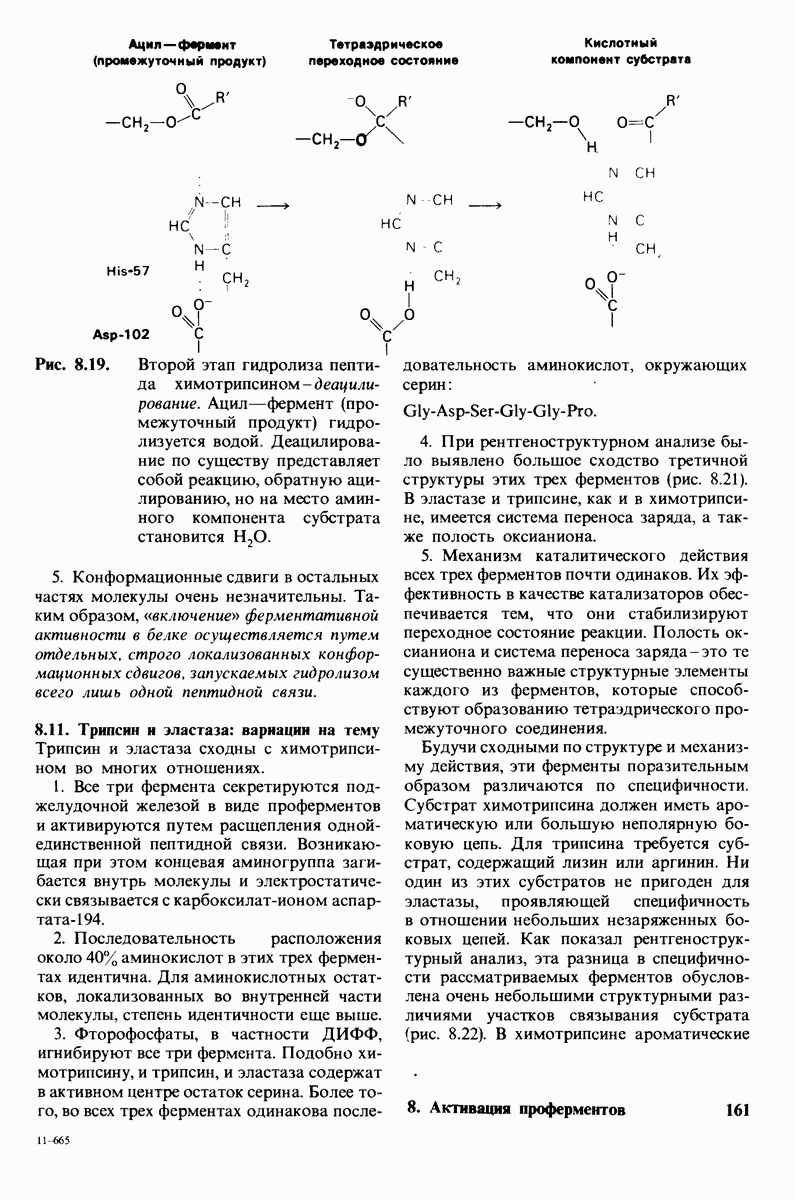
Регуляция активности панкреатических протеиназ осуществляется двумя различными путями. Первый - превращение профермента в активную протеиназу путем расщепления одной пептидной связи. Это очень точный механизм «включения» ферментативной активности, однако он необратим, и, следовательно, для остановки протеолиза должен существовать второй регуляторный механизм. Эту функцию выполняют специфические ингибиторы протеиназ. Например, панкреатический ингибитор трипсина, белок массой  ингибирует активность трипсина, очень прочно связываясь с его активным центром (рис. 8.23). Константа диссоциации комплекса составляет
ингибирует активность трипсина, очень прочно связываясь с его активным центром (рис. 8.23). Константа диссоциации комплекса составляет  что соответствует стандартной свободной энергии связывания примерно —
что соответствует стандартной свободной энергии связывания примерно —  Примечательная особенность этого взаимодействия состоит в том, что комплекс не диссоциирует ни в
Примечательная особенность этого взаимодействия состоит в том, что комплекс не диссоциирует ни в  растворе мочевины, ни в
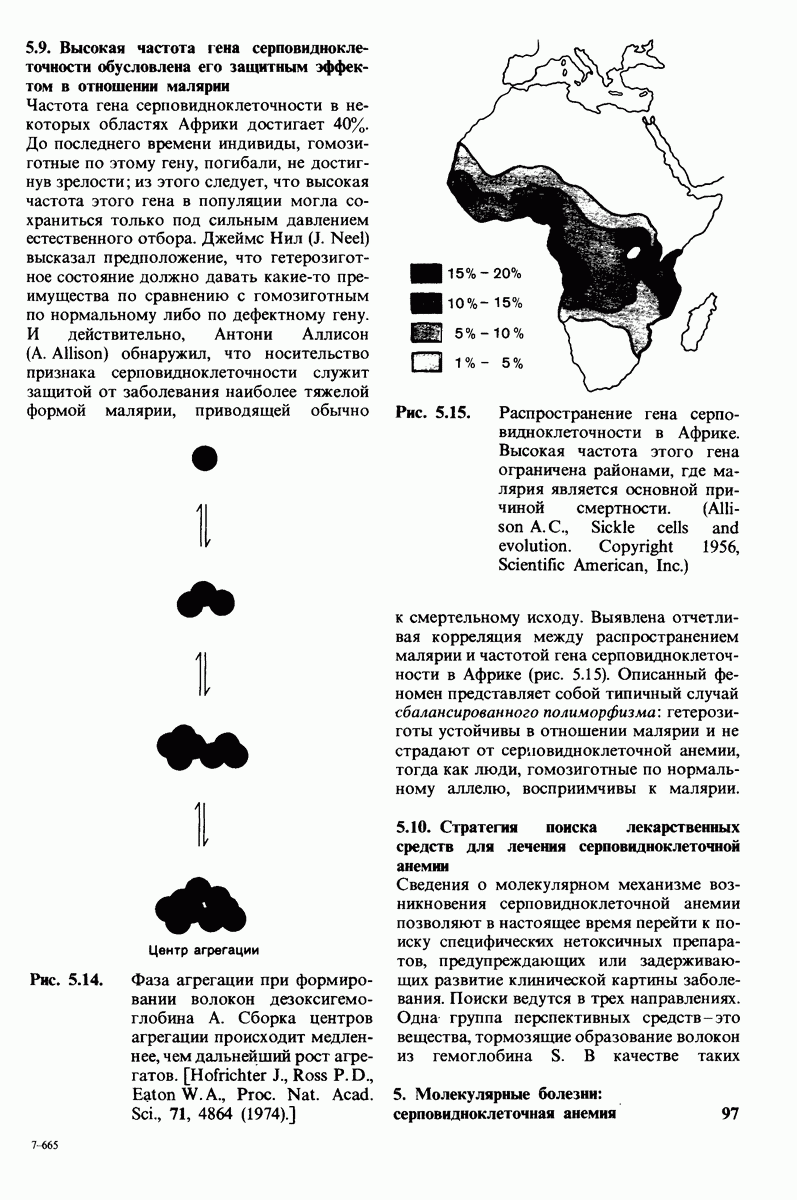
растворе мочевины, ни в  гаунидингидрохлориде. Между тем эти денатурирующие агенты практически всегда вызывают диссоциацию олигомеров белка на составляющие субъединицы. Такая удивительная стабильность рассматриваемого комплекса объясняется тем, что ингибитор трипсина является очень эффективным аналогом субстрата. Рентгеноструктурный анализ показал, что
гаунидингидрохлориде. Между тем эти денатурирующие агенты практически всегда вызывают диссоциацию олигомеров белка на составляющие субъединицы. Такая удивительная стабильность рассматриваемого комплекса объясняется тем, что ингибитор трипсина является очень эффективным аналогом субстрата. Рентгеноструктурный анализ показал, что

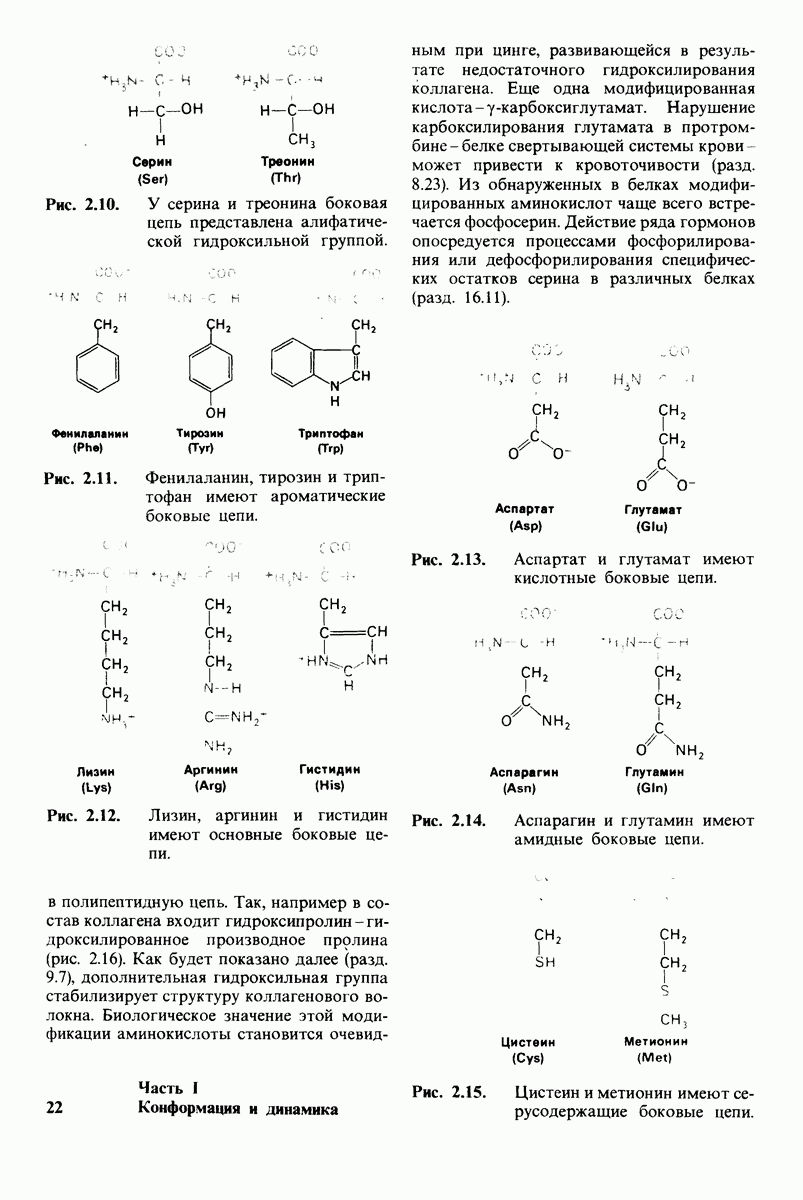
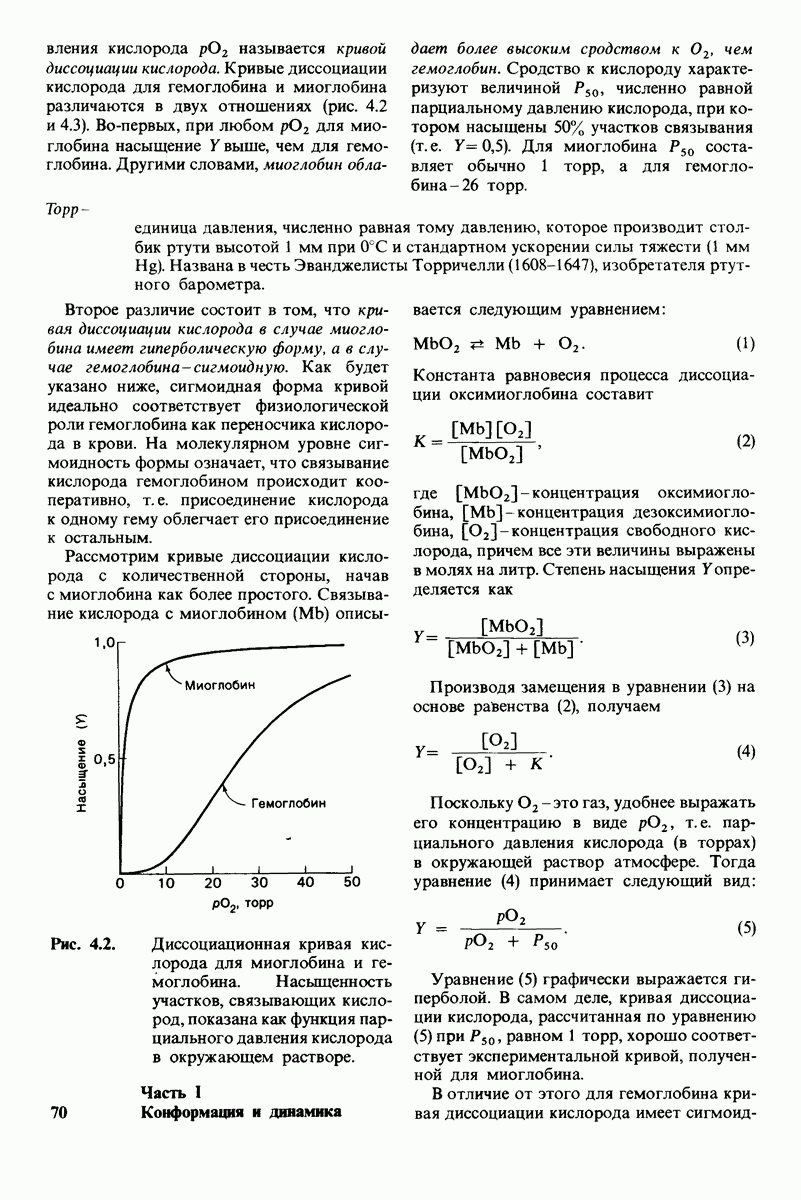
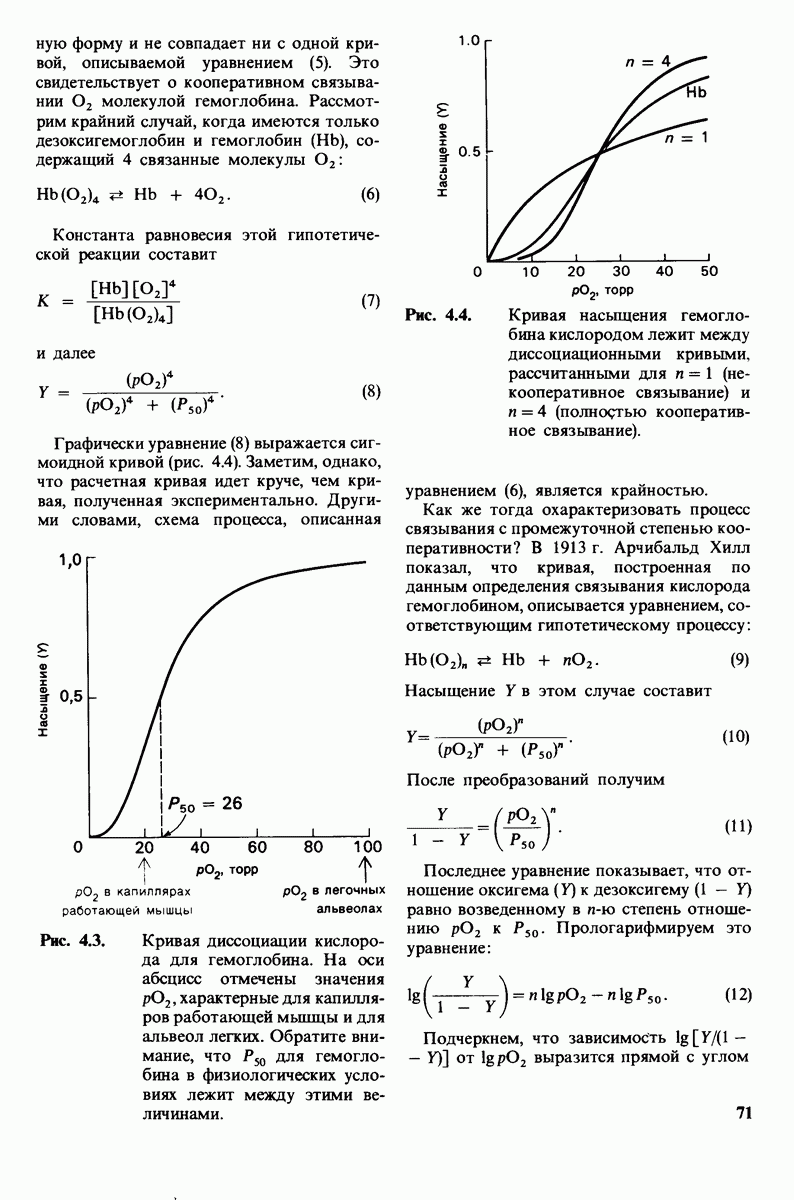
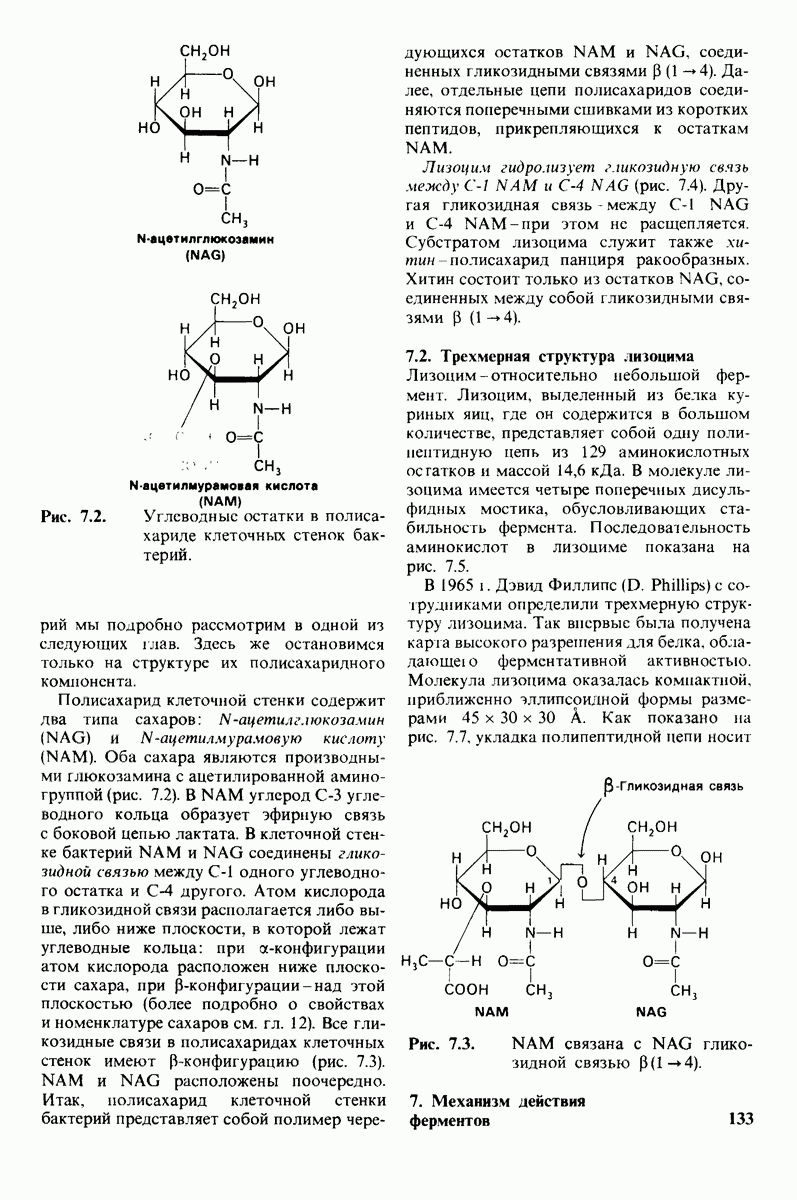
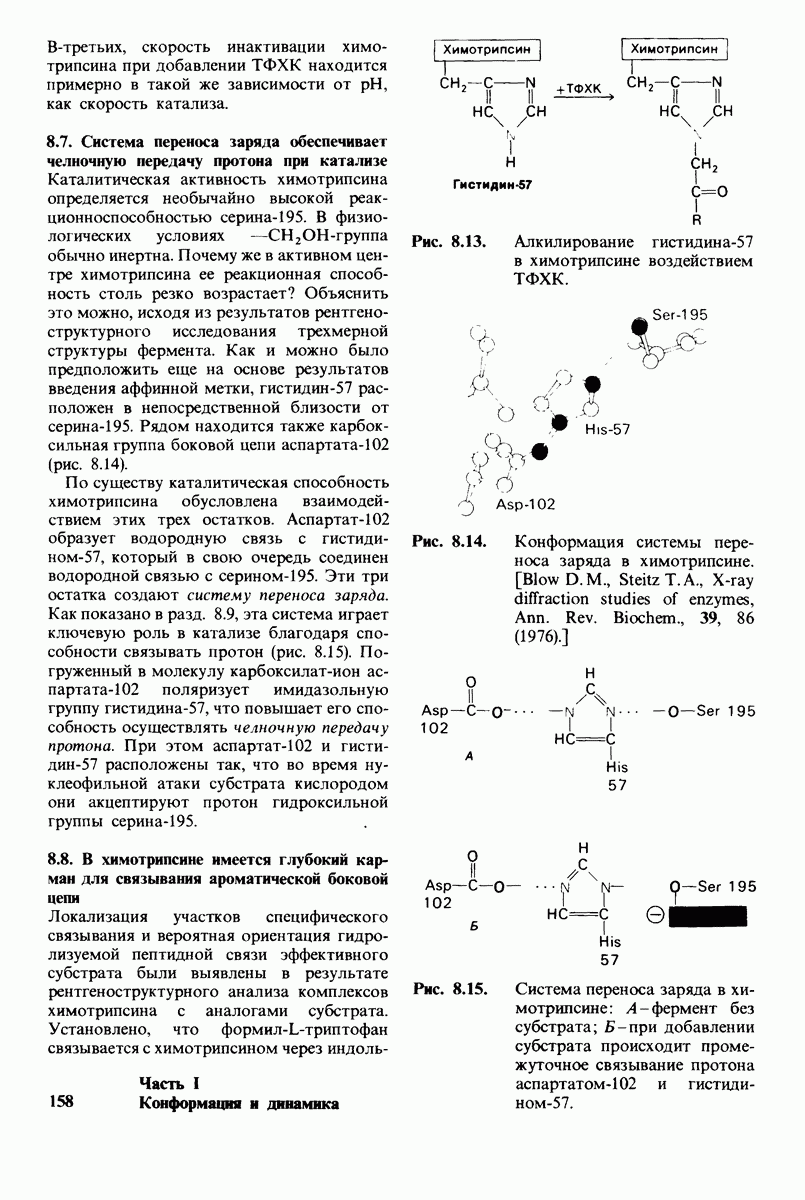
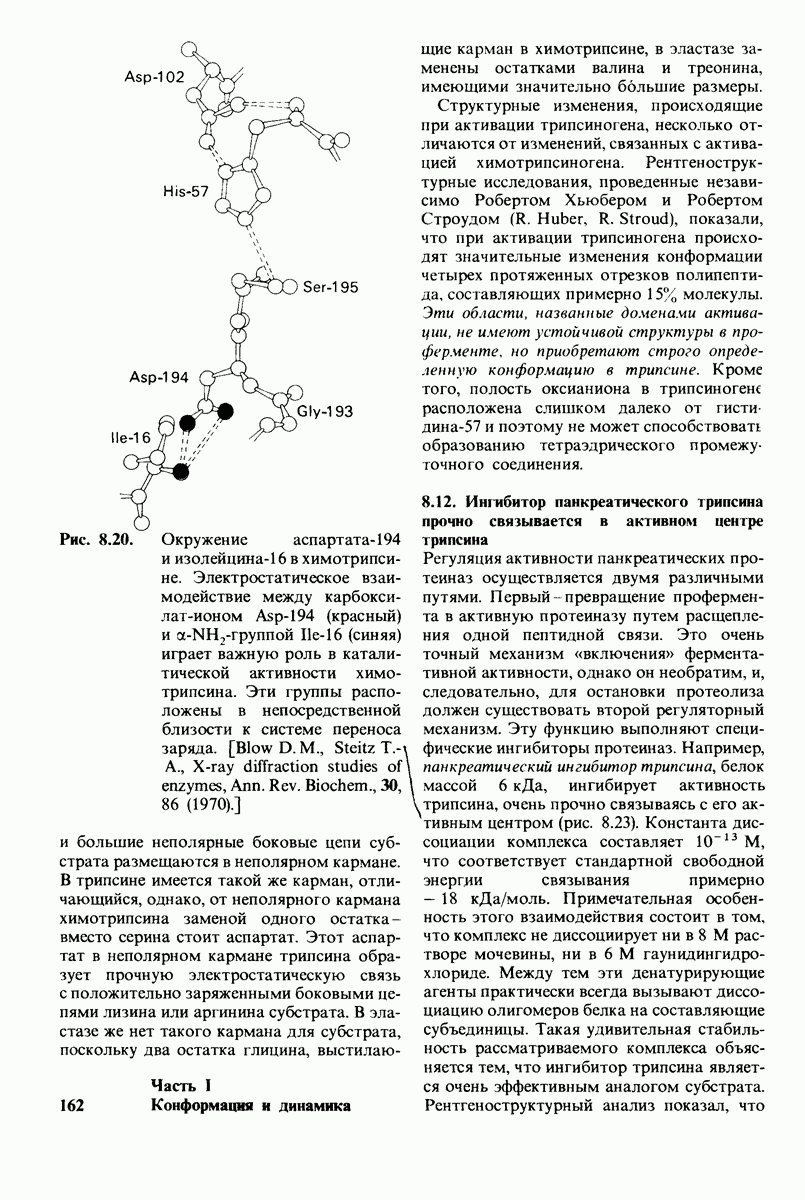
Рис. 8.21.
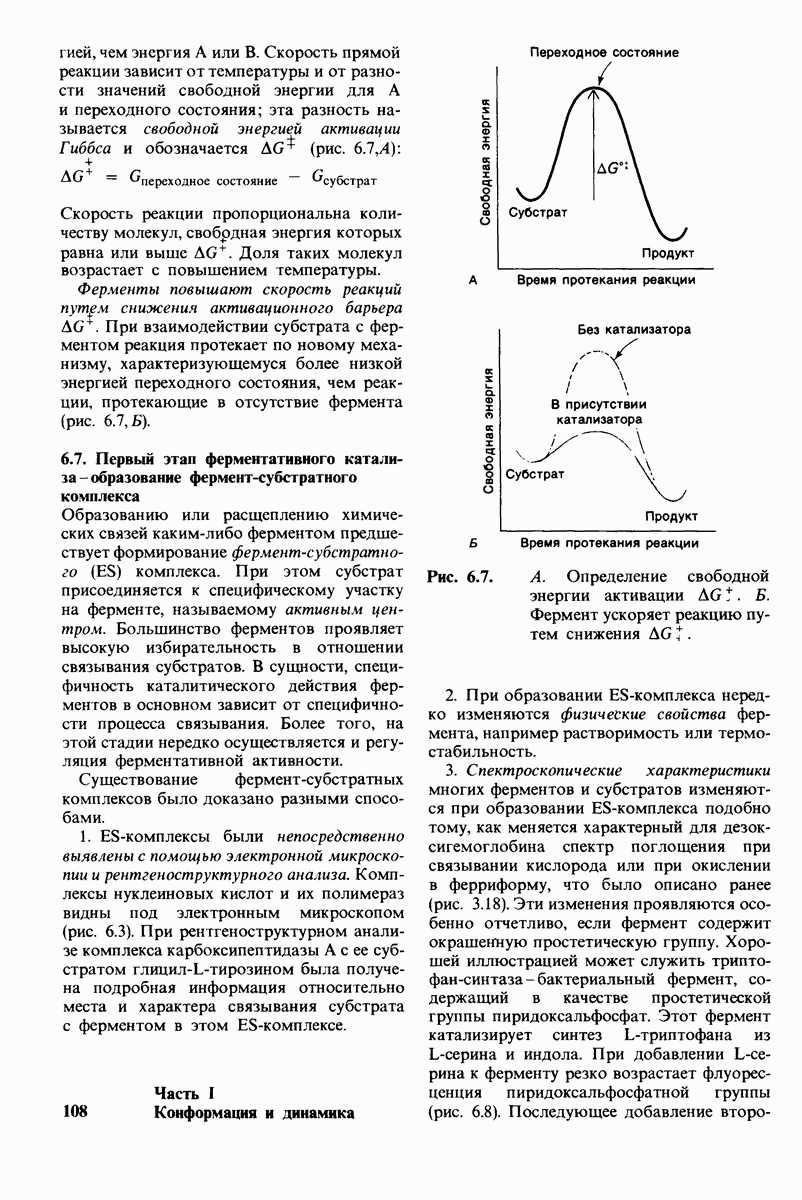
Сравнение конформации основной цепи химотрипсина  и эластазы
и эластазы  Локализация системы переноса заряда (остатки 102, 57 и 195) и
Локализация системы переноса заряда (остатки 102, 57 и 195) и  -аминогруппы остатка 16 выделена цветом, чтобы подчеркнуть сходство структуры этих ферментов. (Hartley В. S., Shotton D. М. In: The Enzymes, P.O. Boyer, ed., 3rd ed., v. 3, Academic Press, 1971, p. 362.)
-аминогруппы остатка 16 выделена цветом, чтобы подчеркнуть сходство структуры этих ферментов. (Hartley В. S., Shotton D. М. In: The Enzymes, P.O. Boyer, ed., 3rd ed., v. 3, Academic Press, 1971, p. 362.)
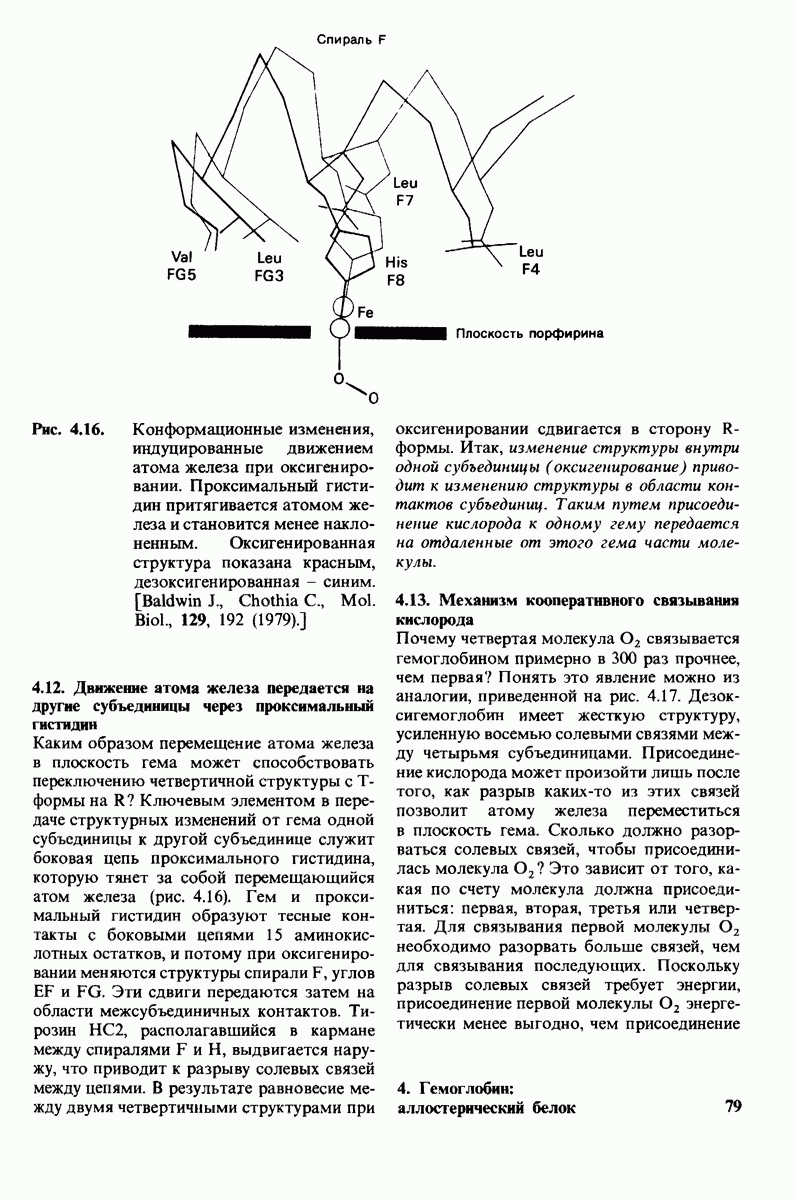
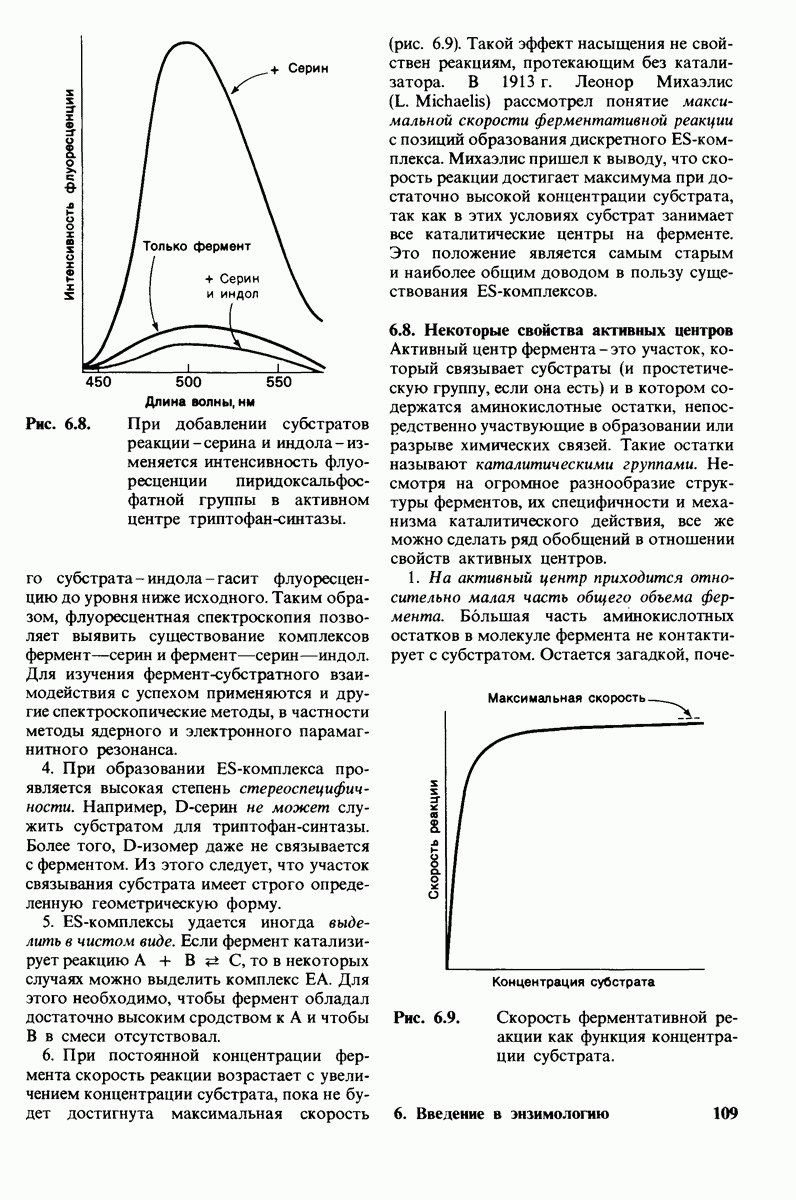
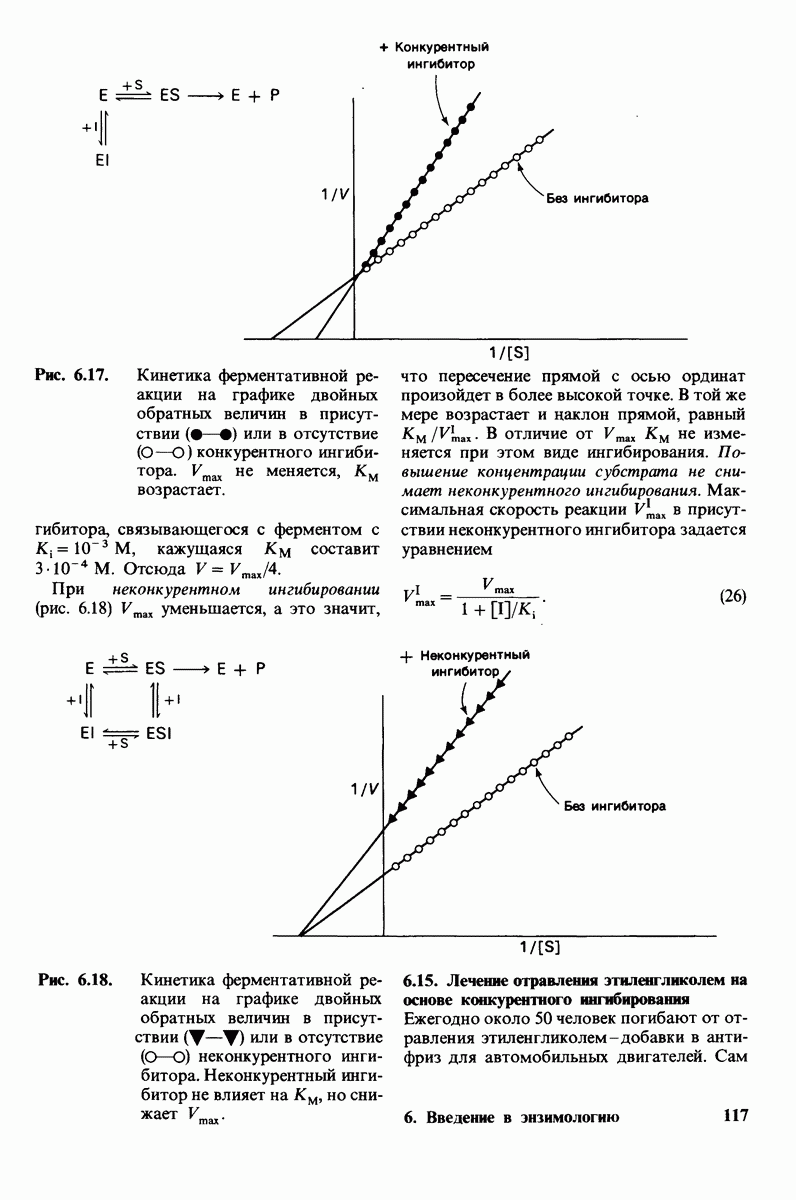
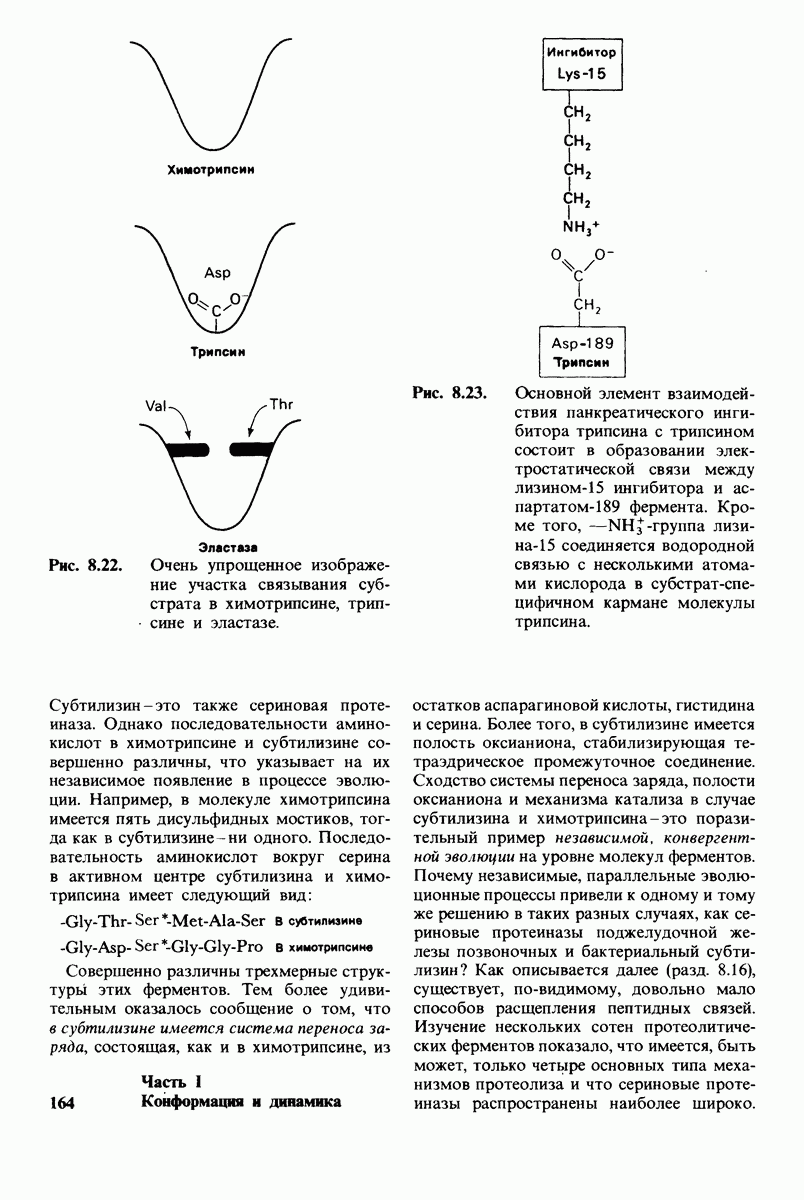
боковая цепь лизина-15 в молекуле ингибитора соединяется с боковой цепью аспартата в субстрат-специфичном кармане фермента. Кроме того, между полипептидными цепями фермента и ингибитора возникает большое число водородных связей. Они располагаются так же, как водородные связи между ферментом и истинным субстратом. Но что самое главное - карбонильная группа лизина-15 и окружающие ее атомы ингибитора плотно входят в полость оксианиона в ферменте. Однако весь комплекс как будто застывает на этом промежуточном этапе катализа - видимо, потому, что гистидин-57 фермента, будучи неподвижным, лишен возможности перебросить протон на амидный азот ингибитора. Пептидная связь между лизином-15 и аланином-16 в панкреатическом ингибиторе трипсина расщепляется, но с крайне малой скоростью. Период полужизни комплекса трипсин-ингибитор составляет несколько месяцев. Ингибитор обладает очень высоким средством к трипсину в силу почти идеальной комплементарности его структуры активному центру трипсина. Ограниченная подвижность конформации ингибитора в участке его связывания с ферментом блокирует процесс катализа и приводит к исключительно высокой прочности комплекса с трипсином.