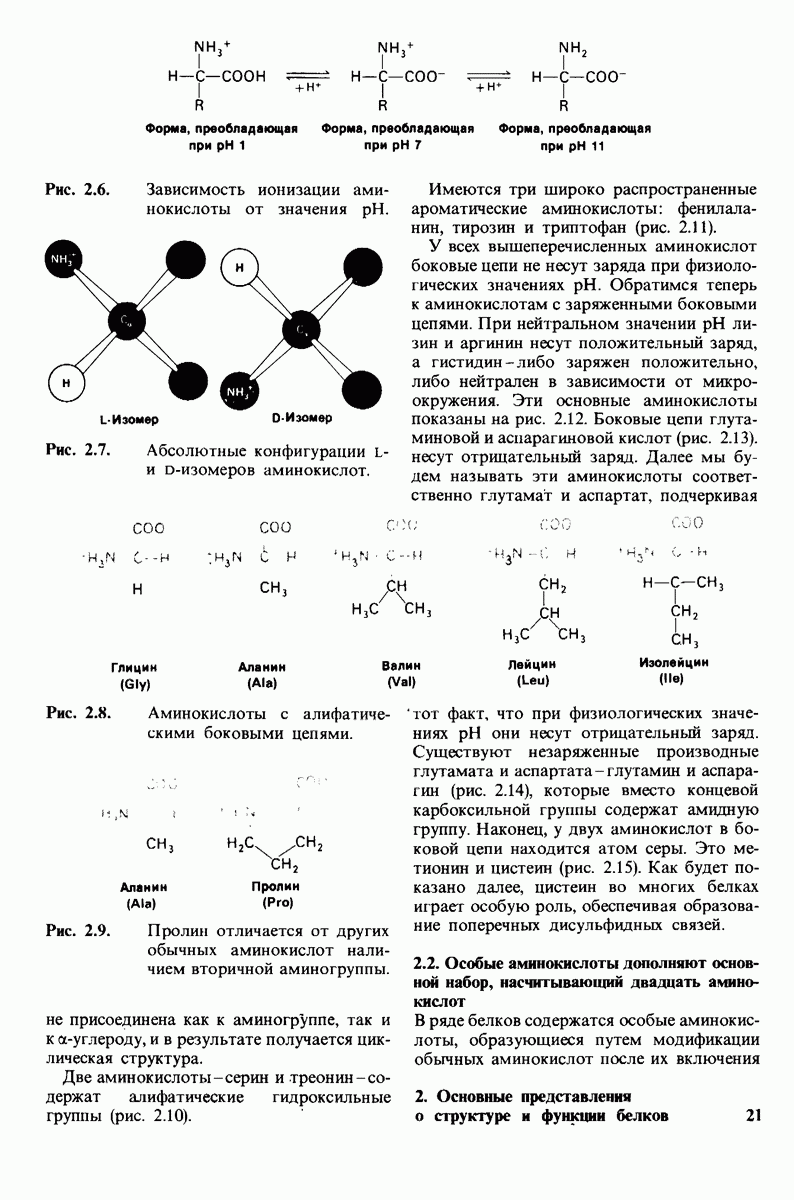
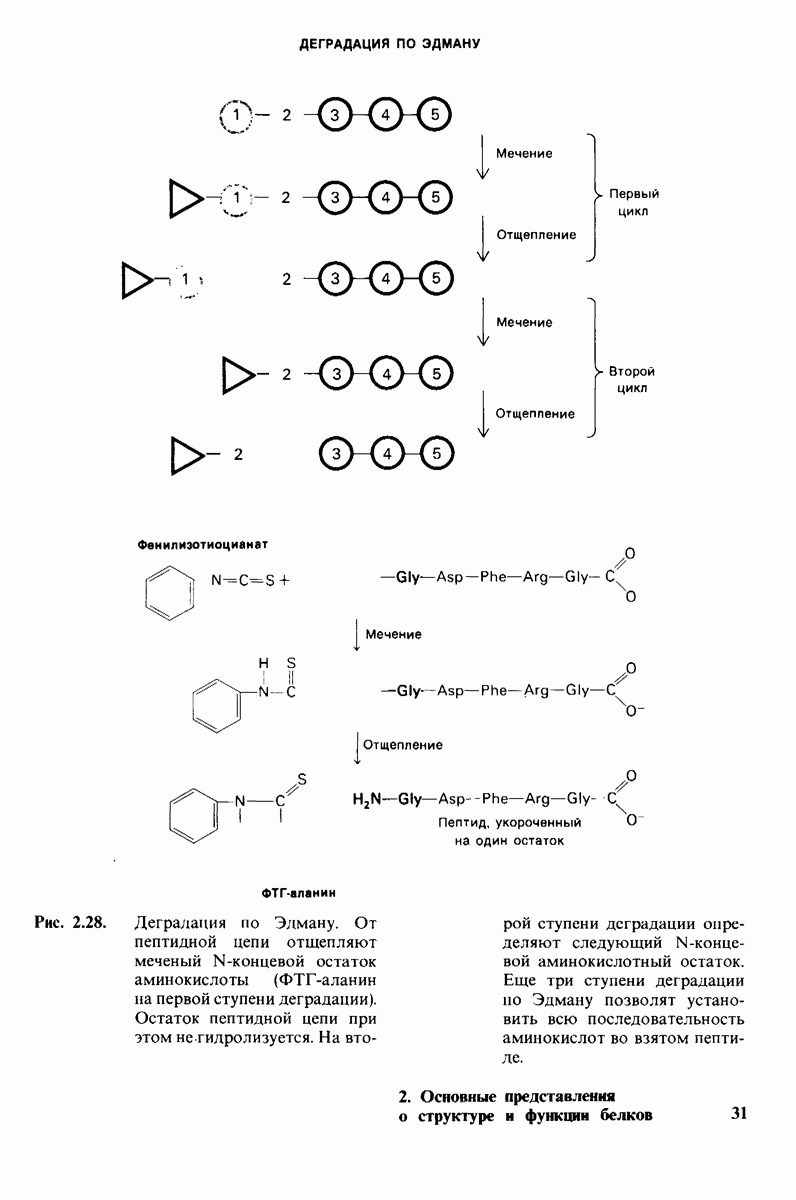
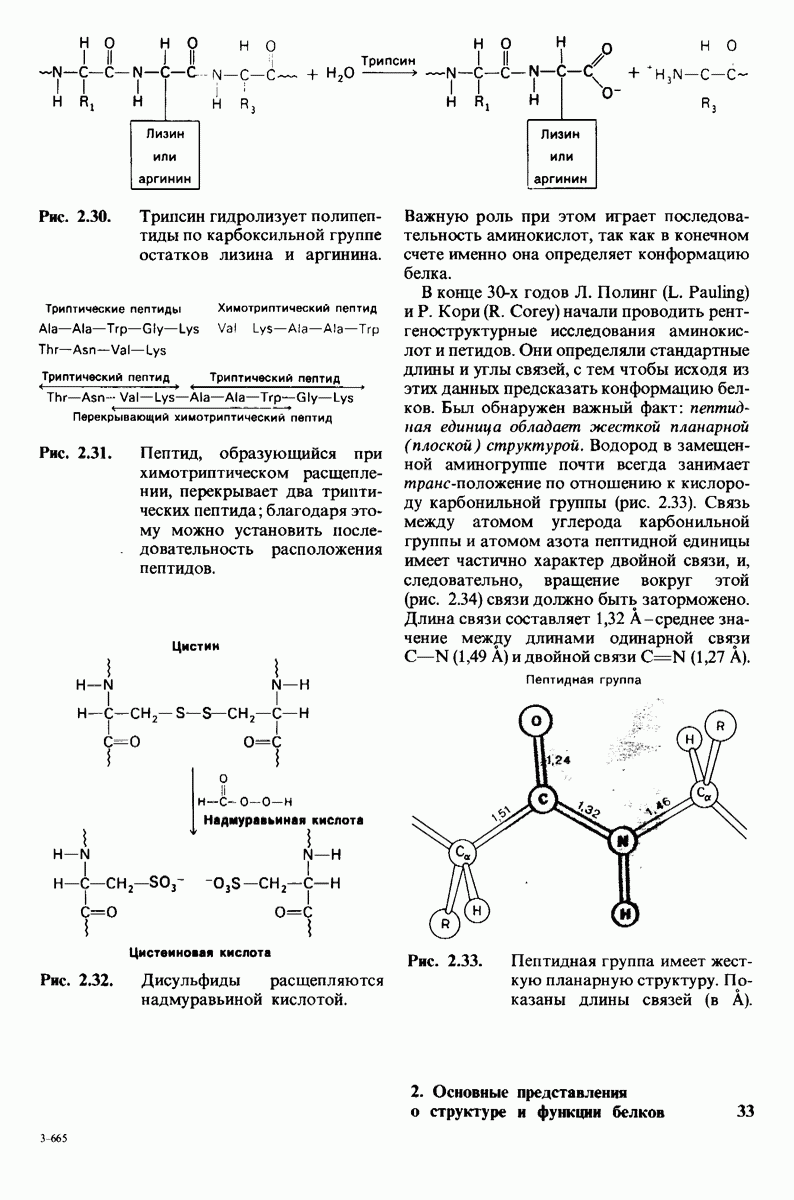
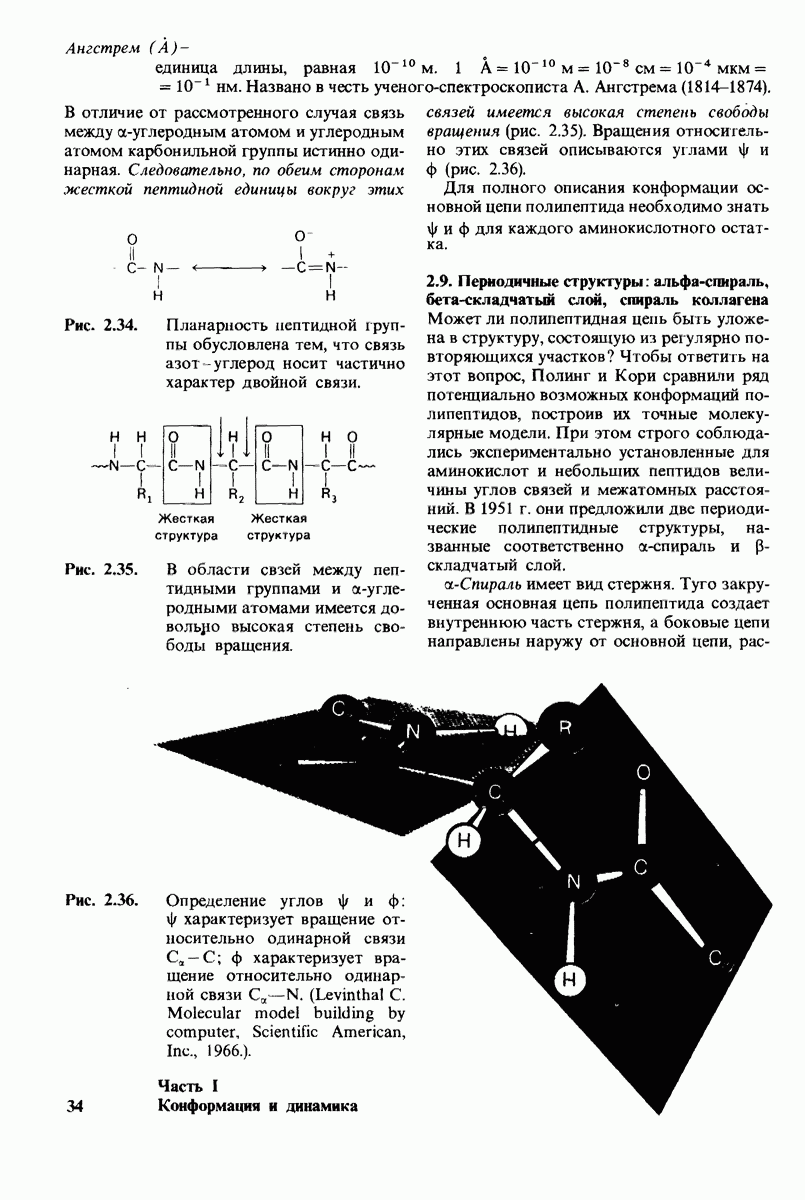
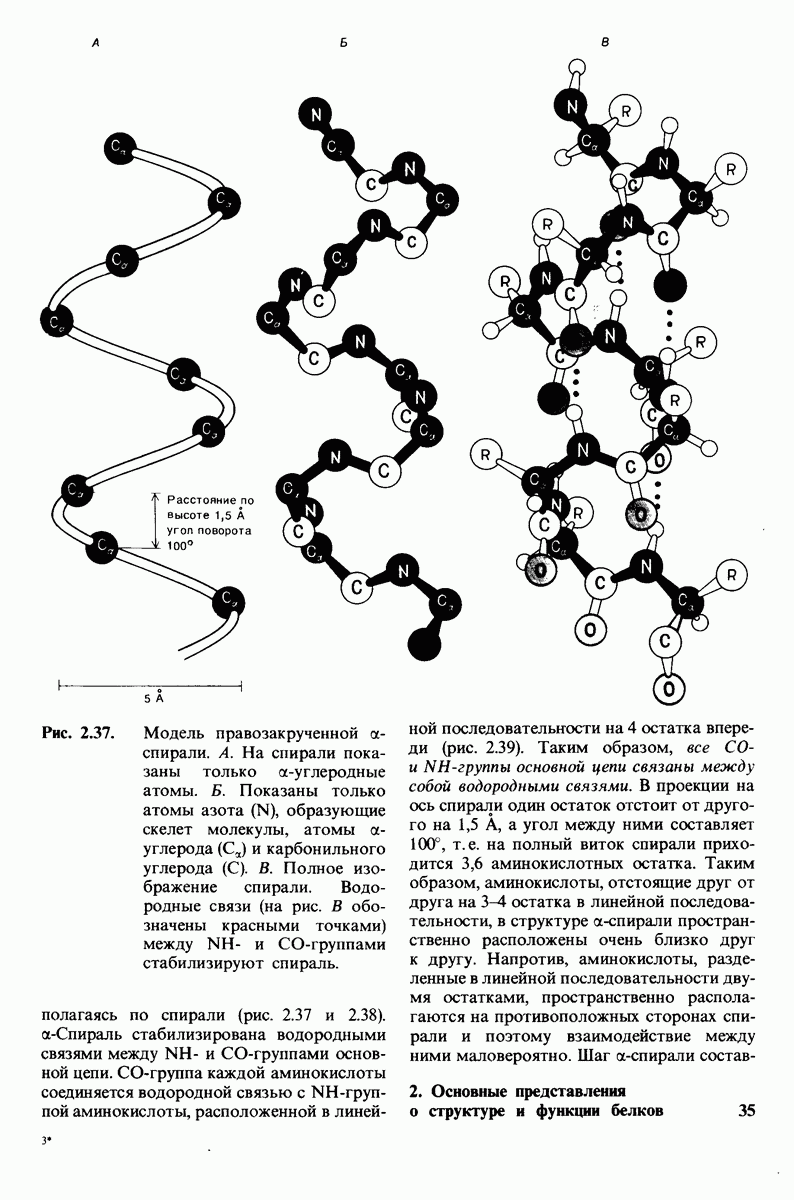
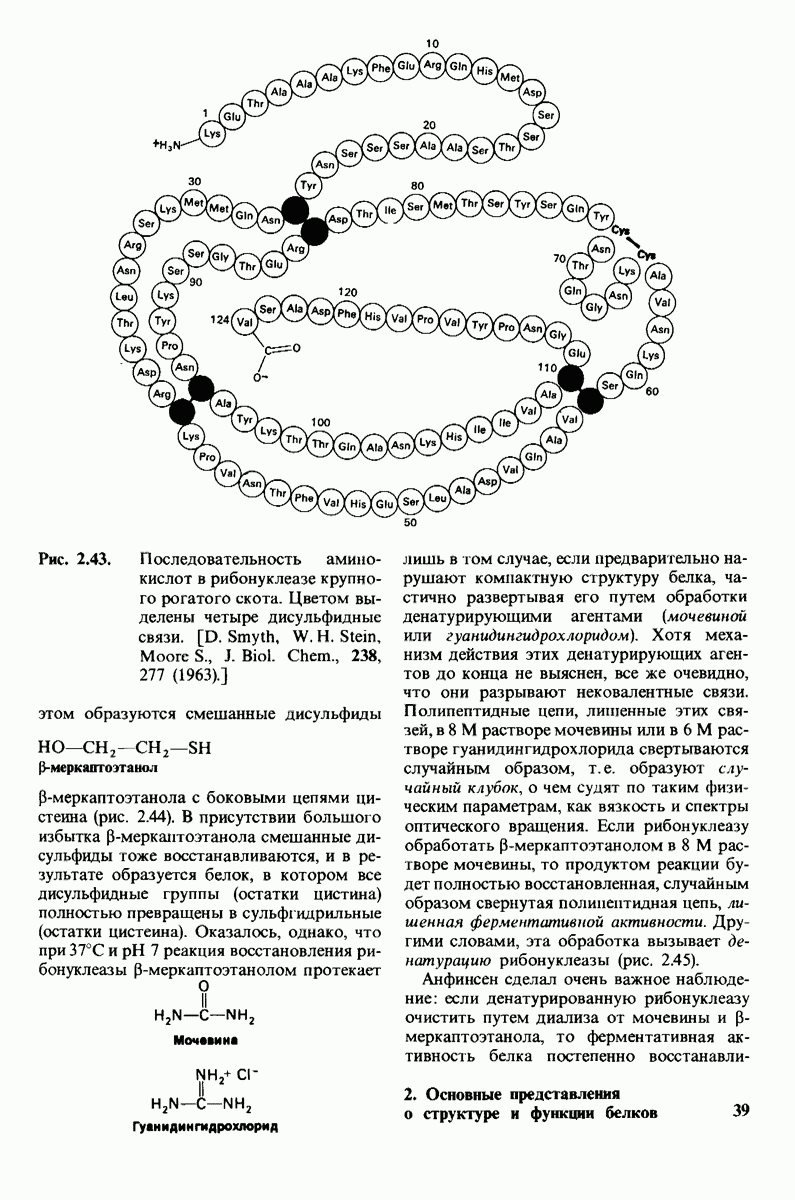
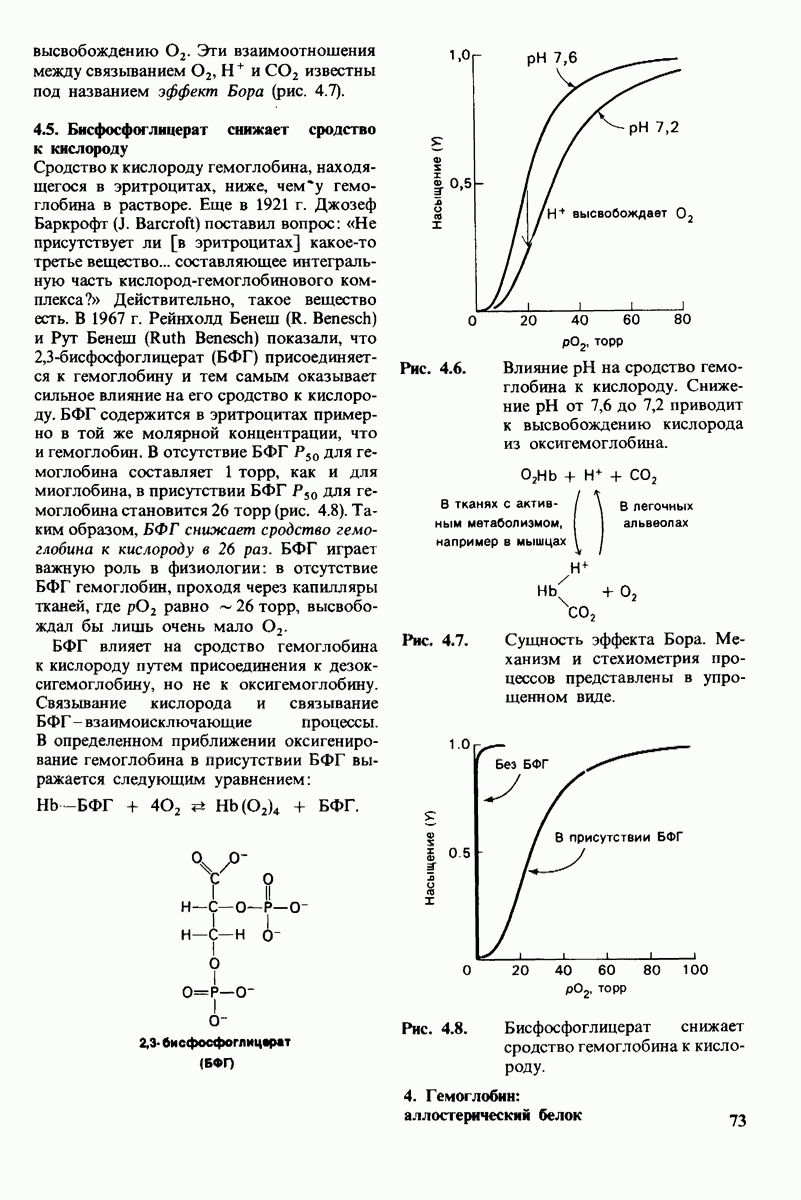
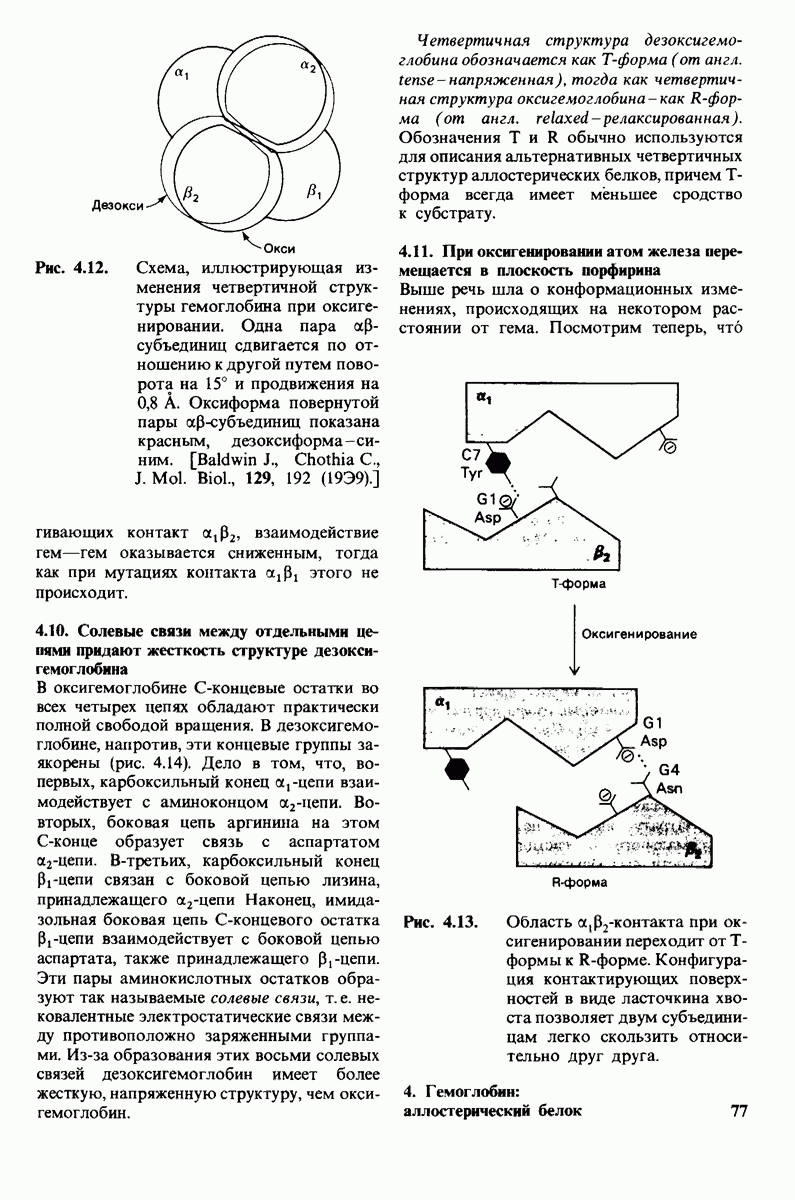
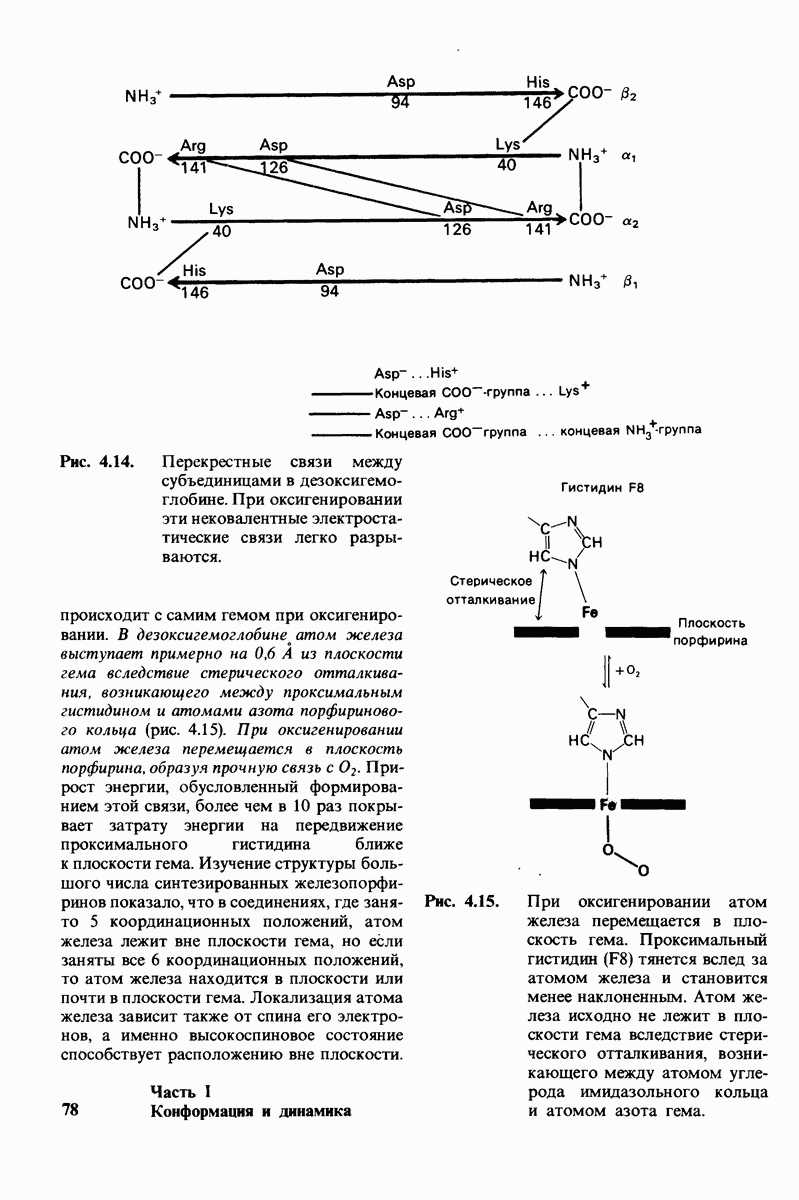
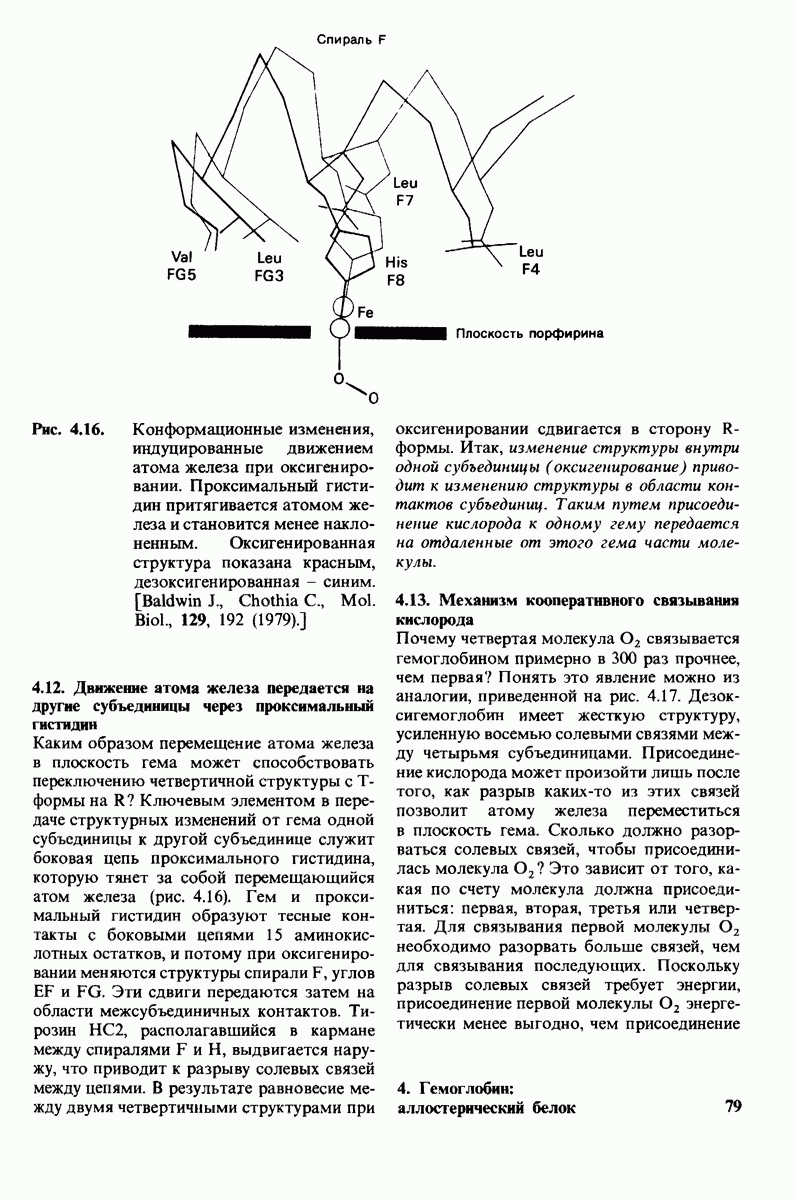
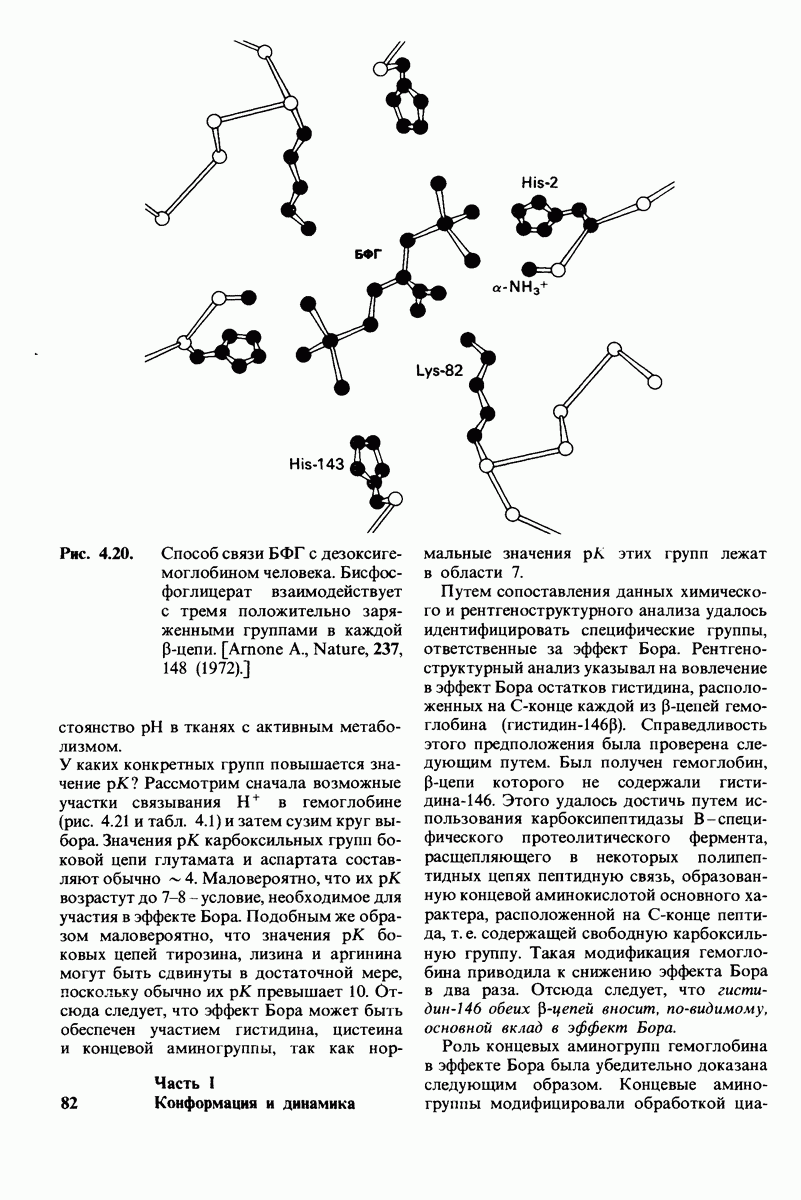
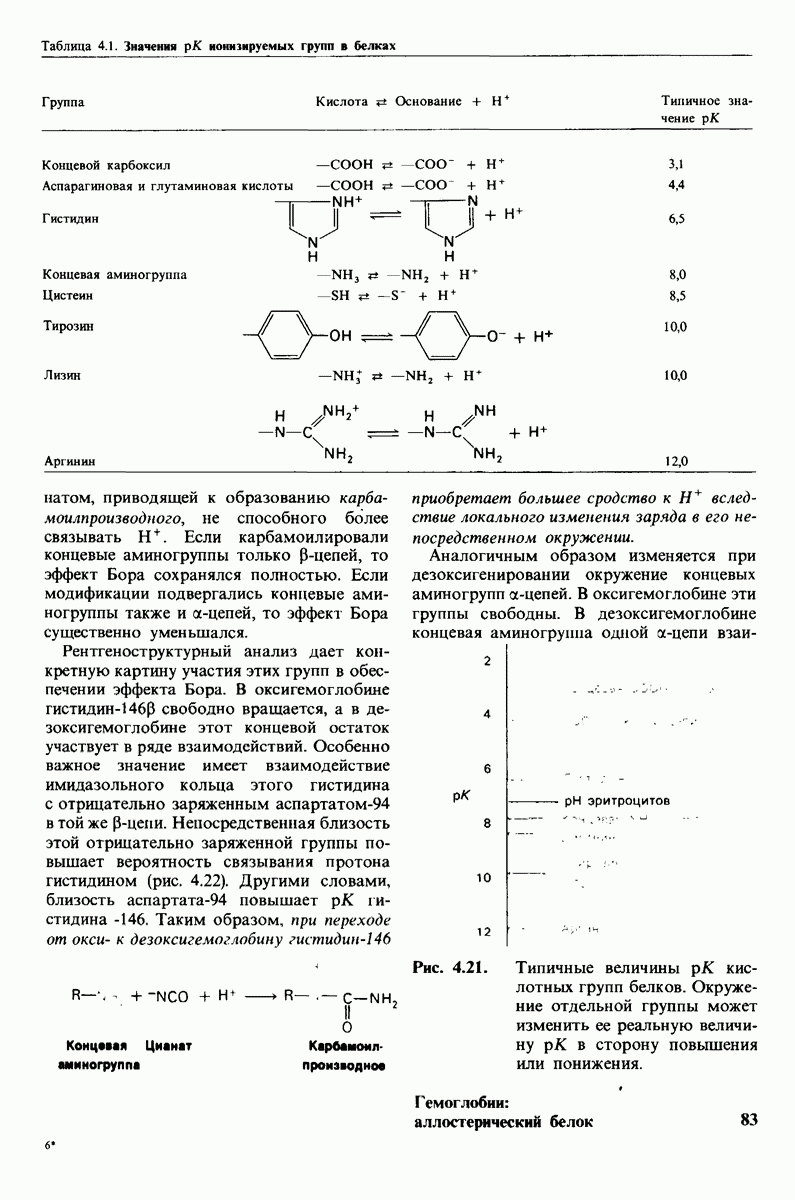
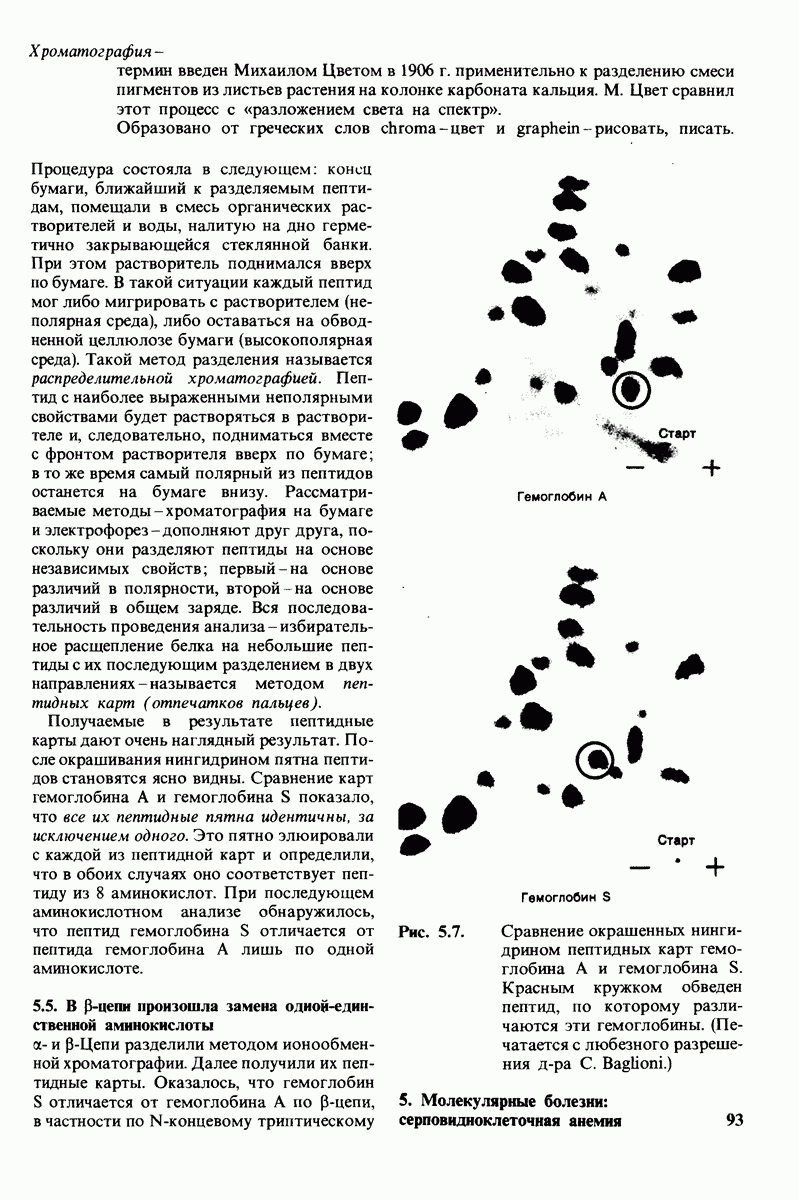
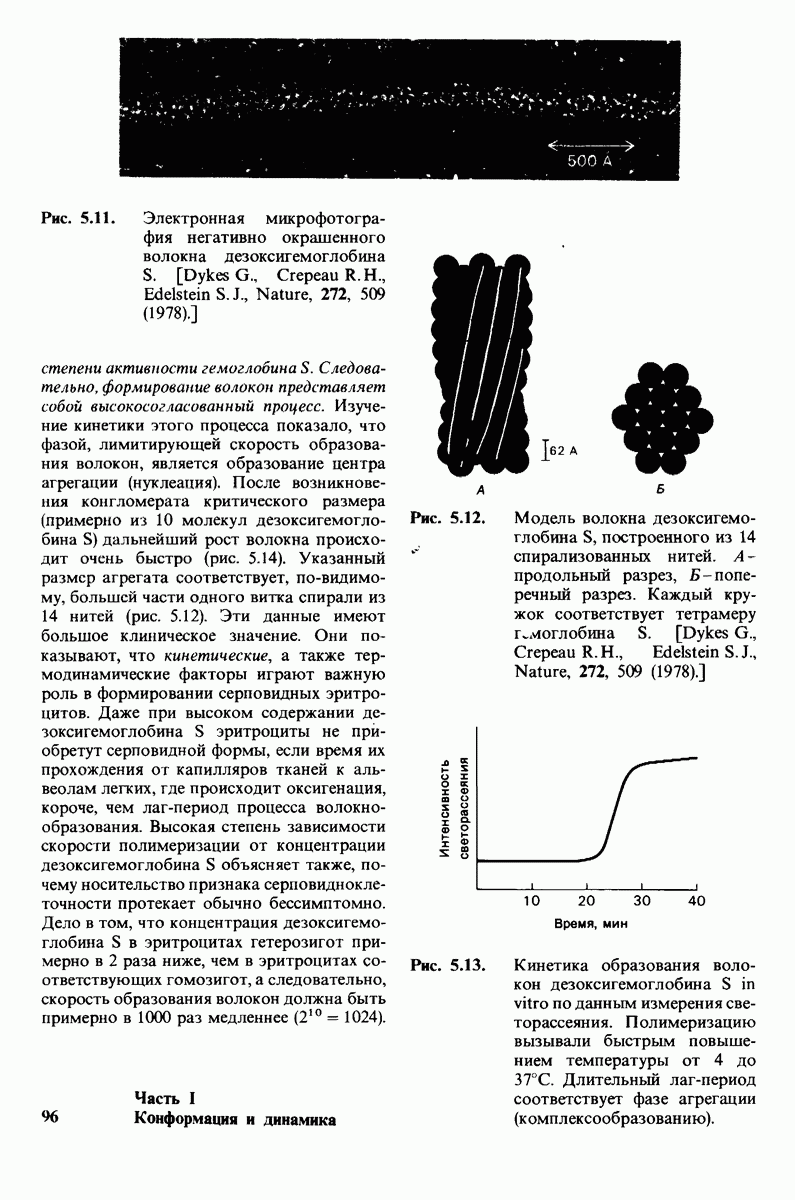
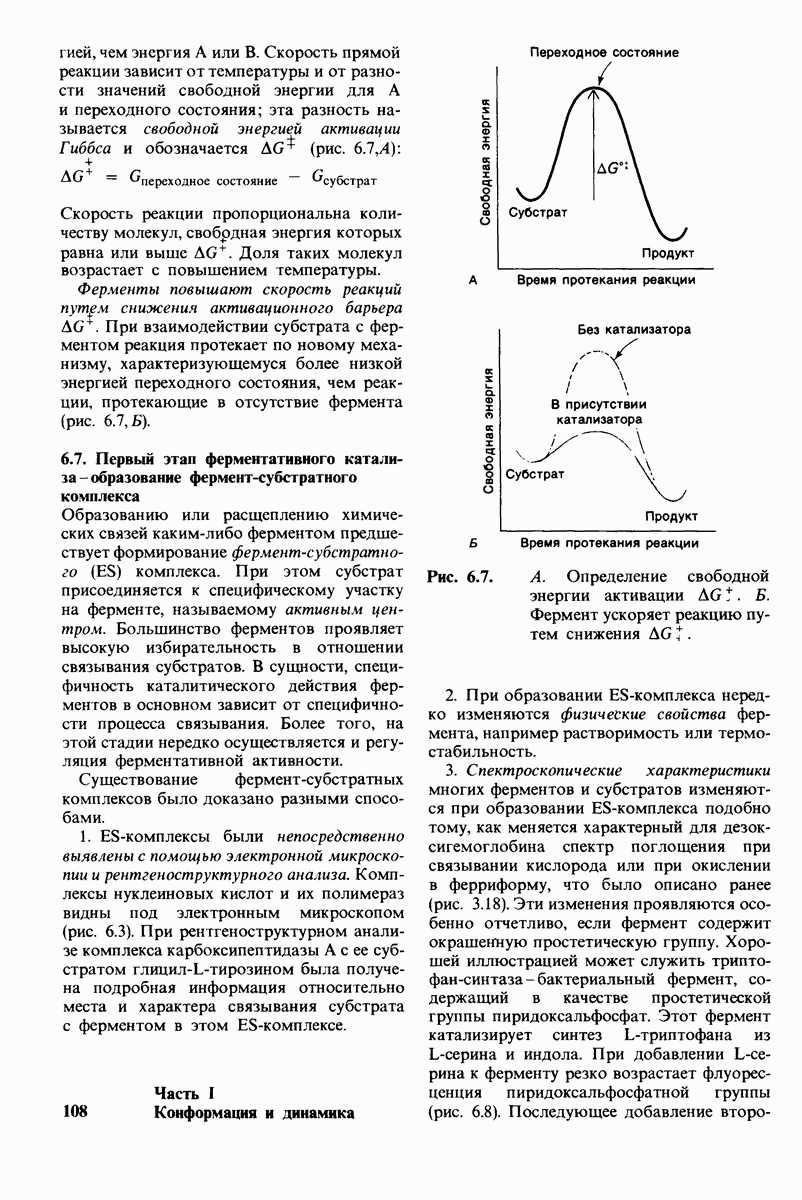
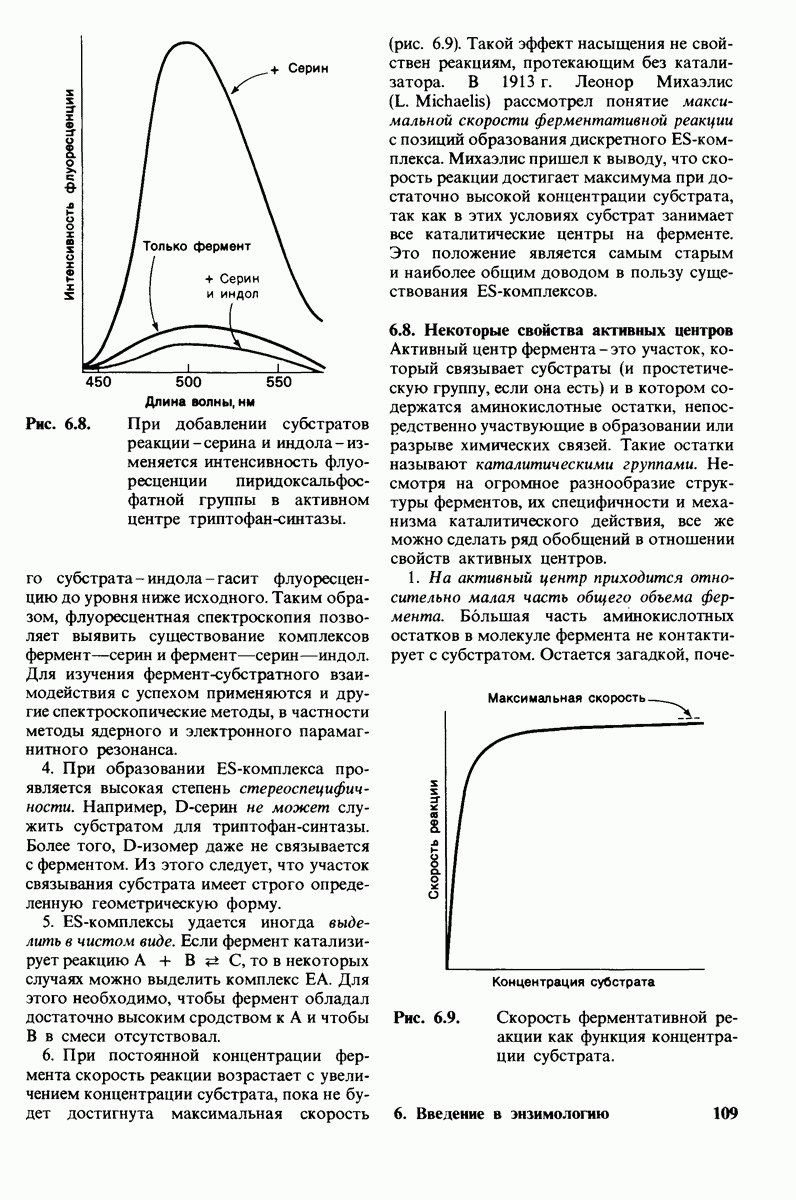
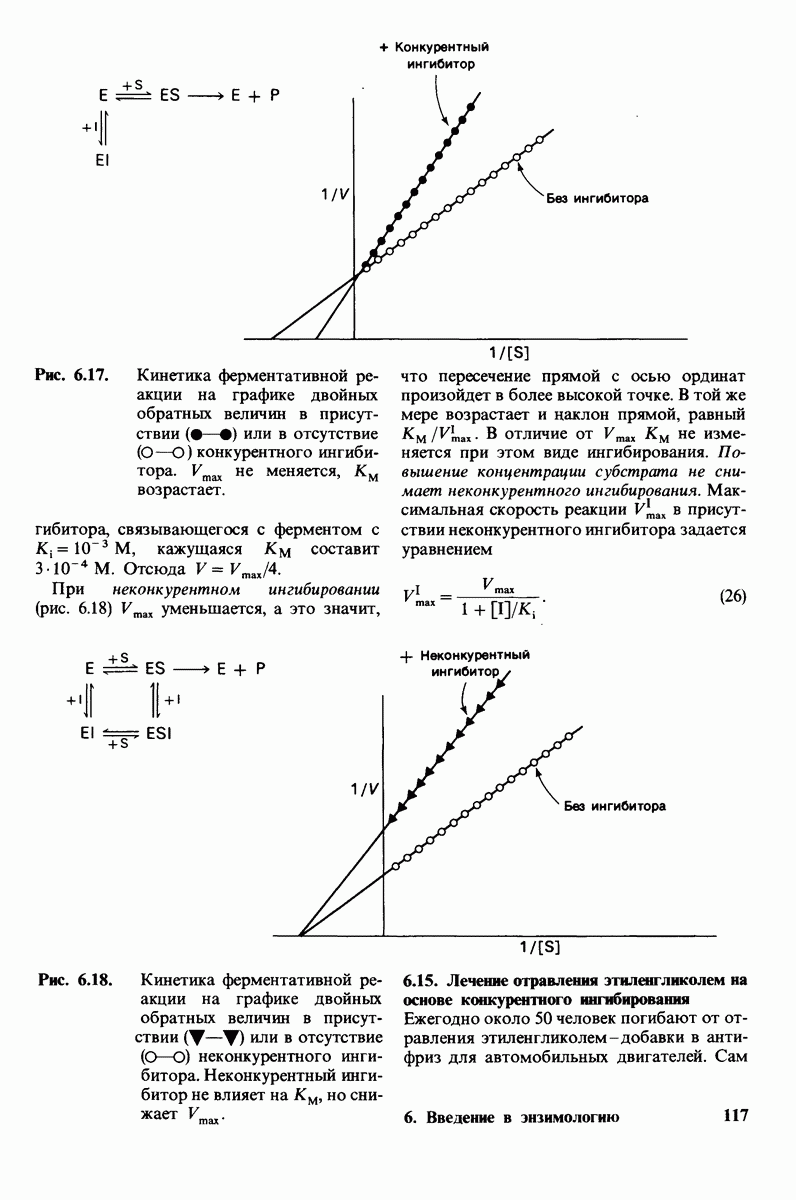
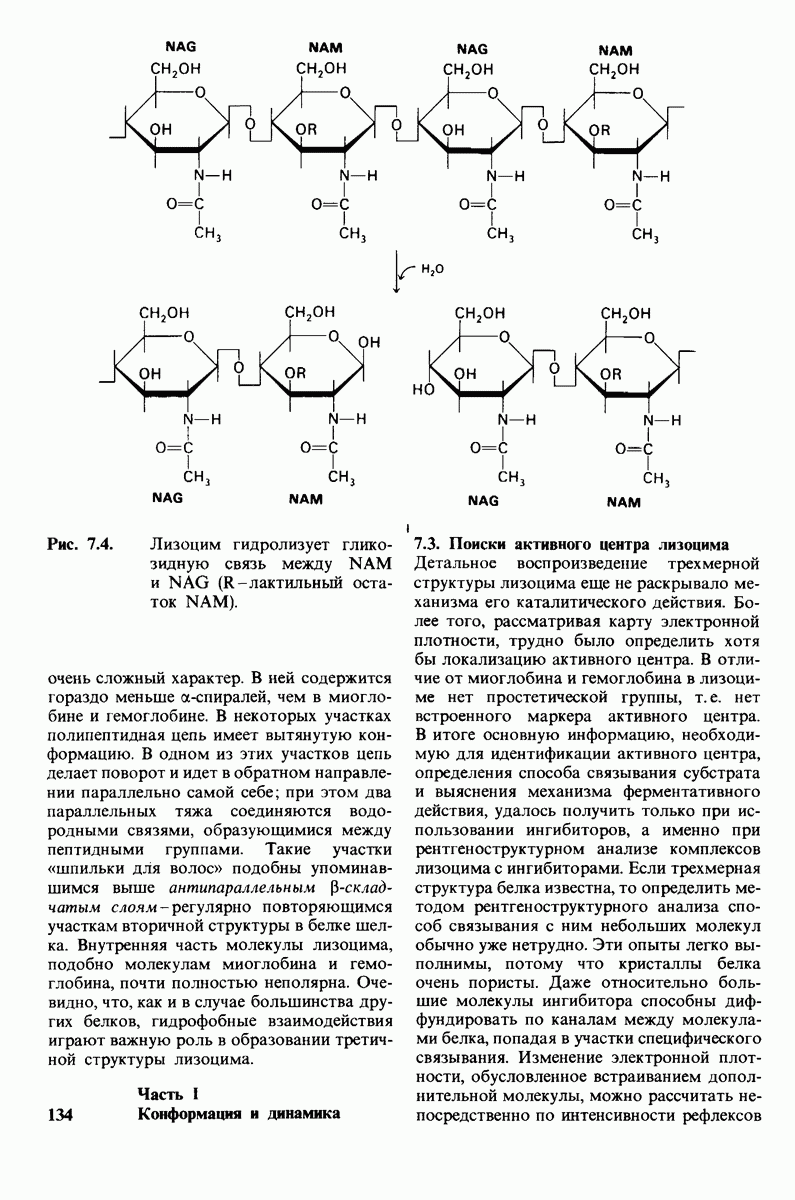
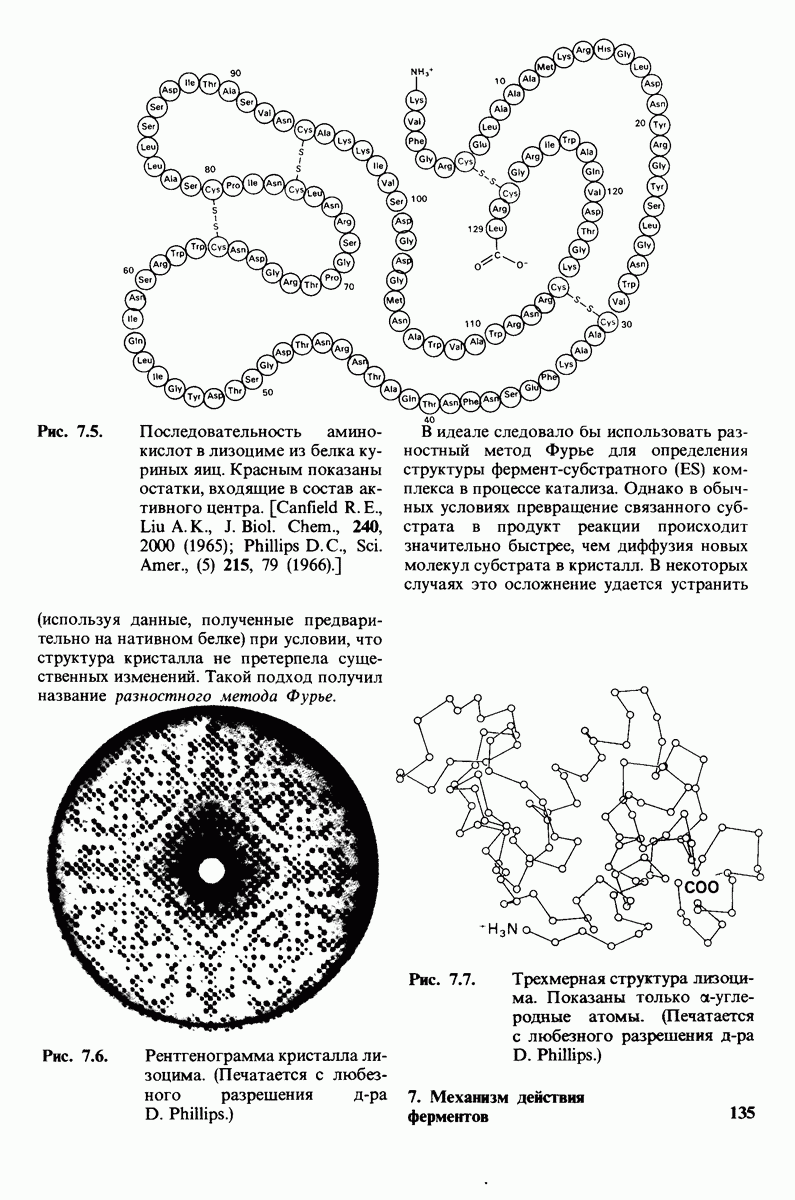
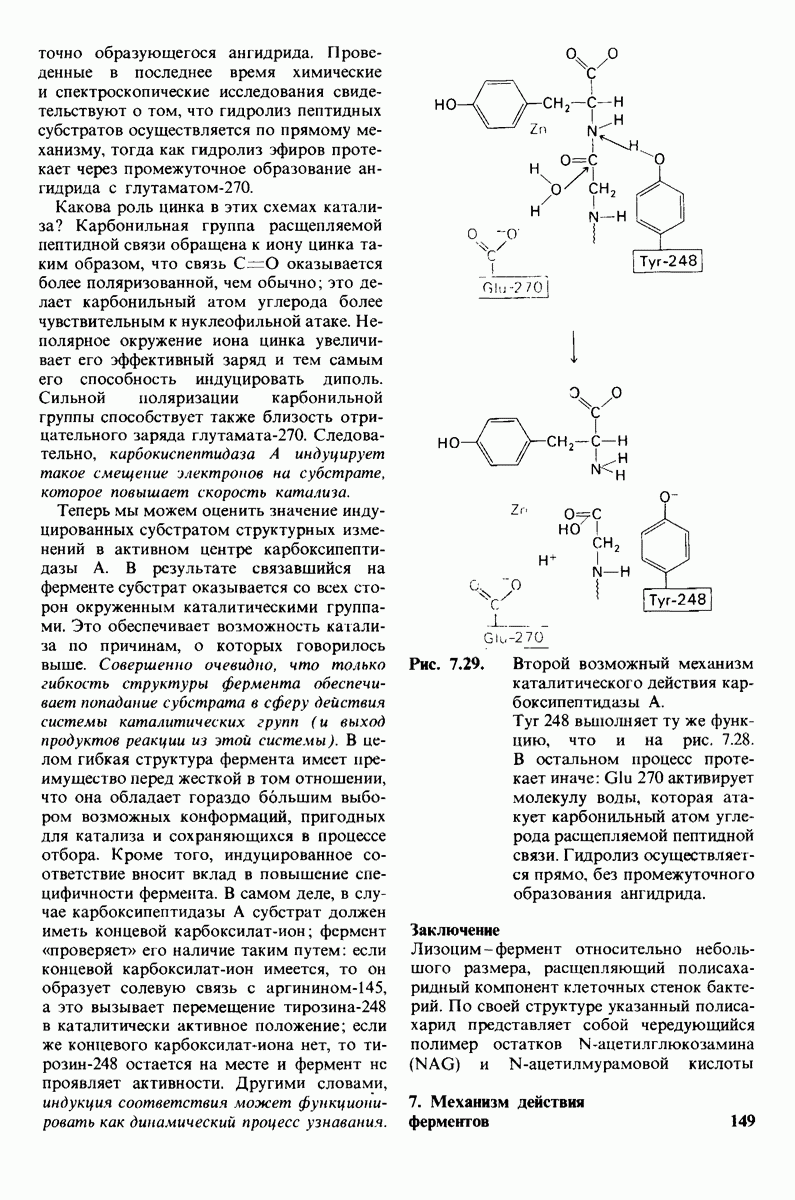
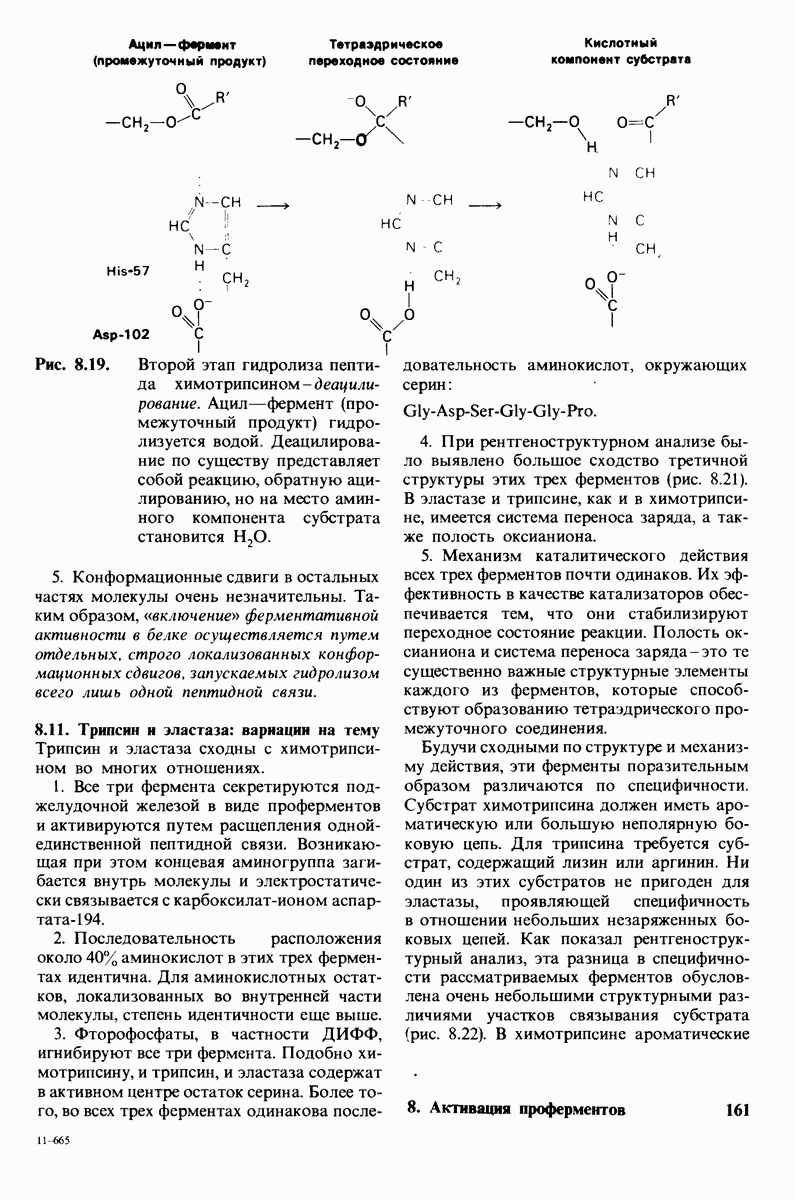
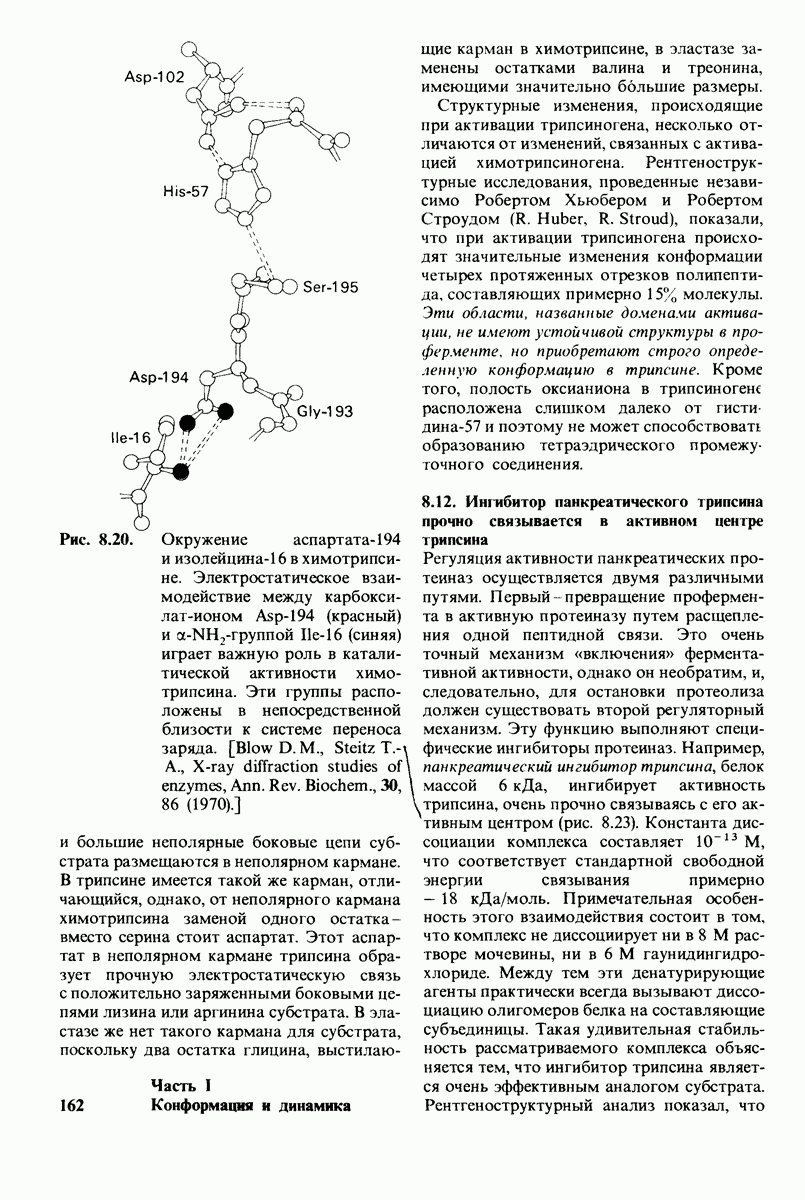
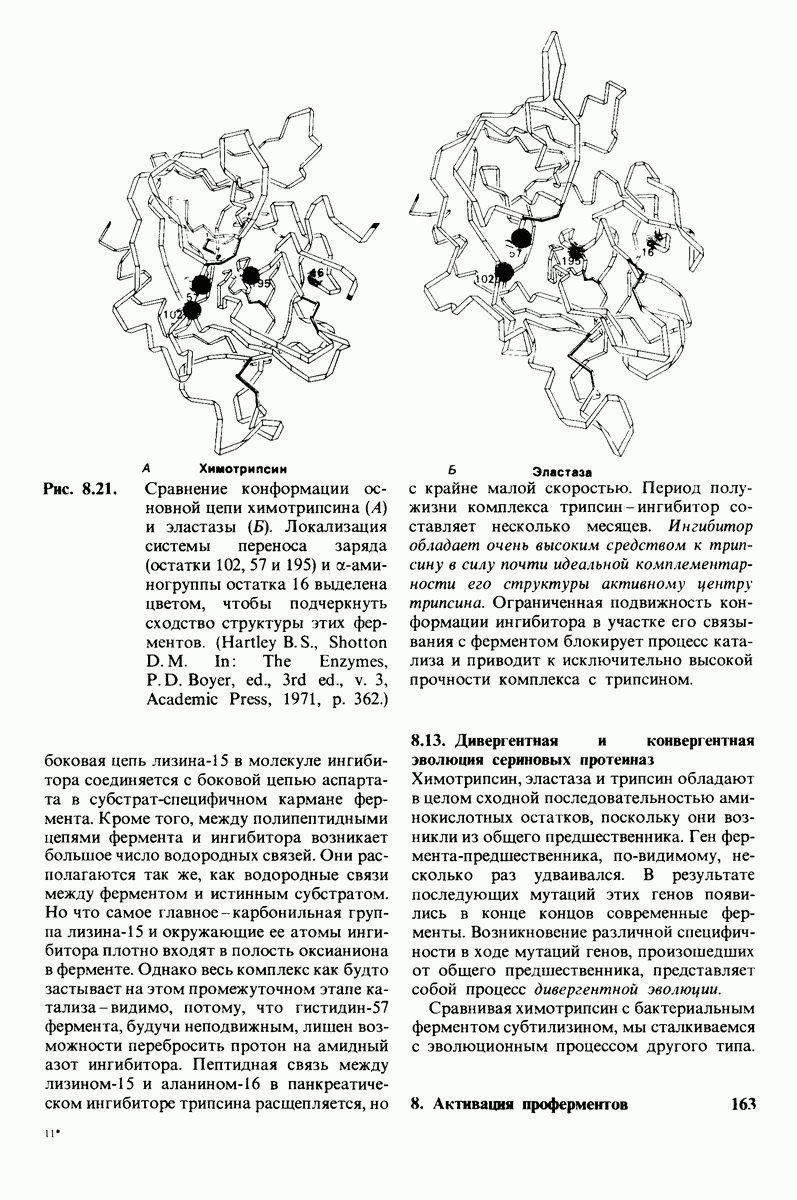
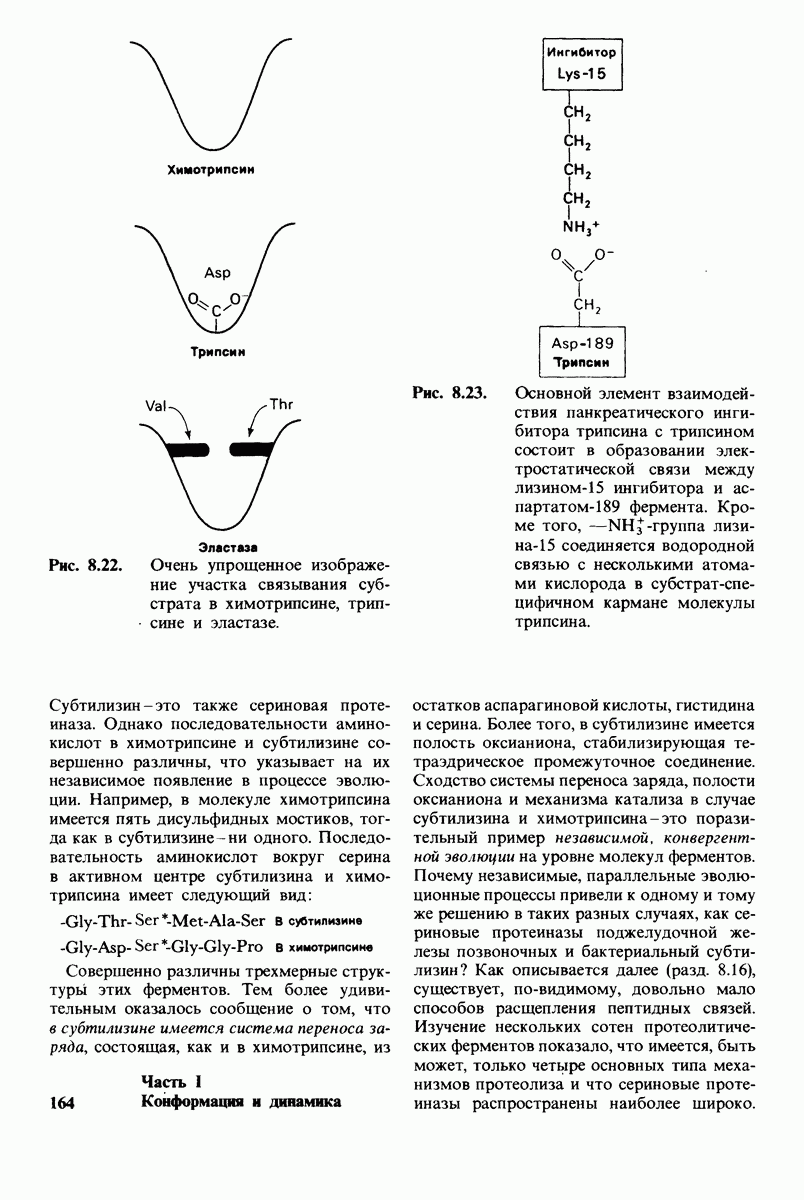
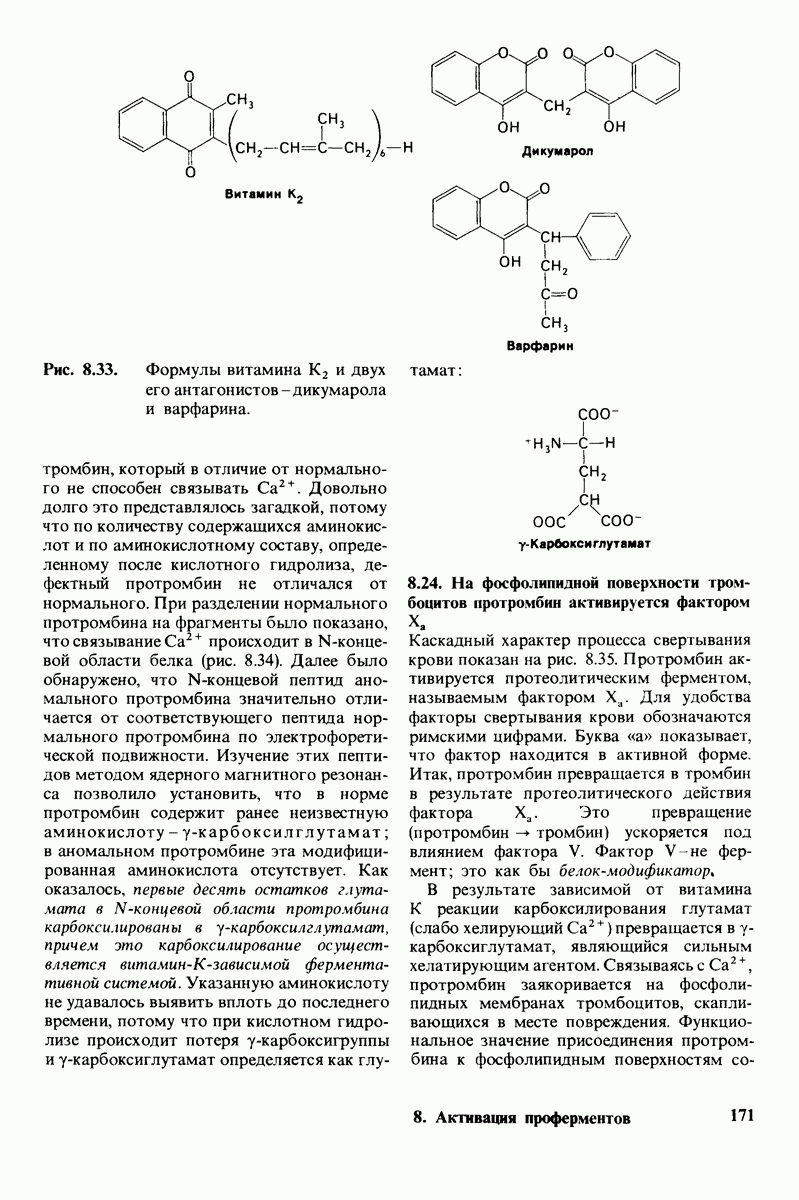
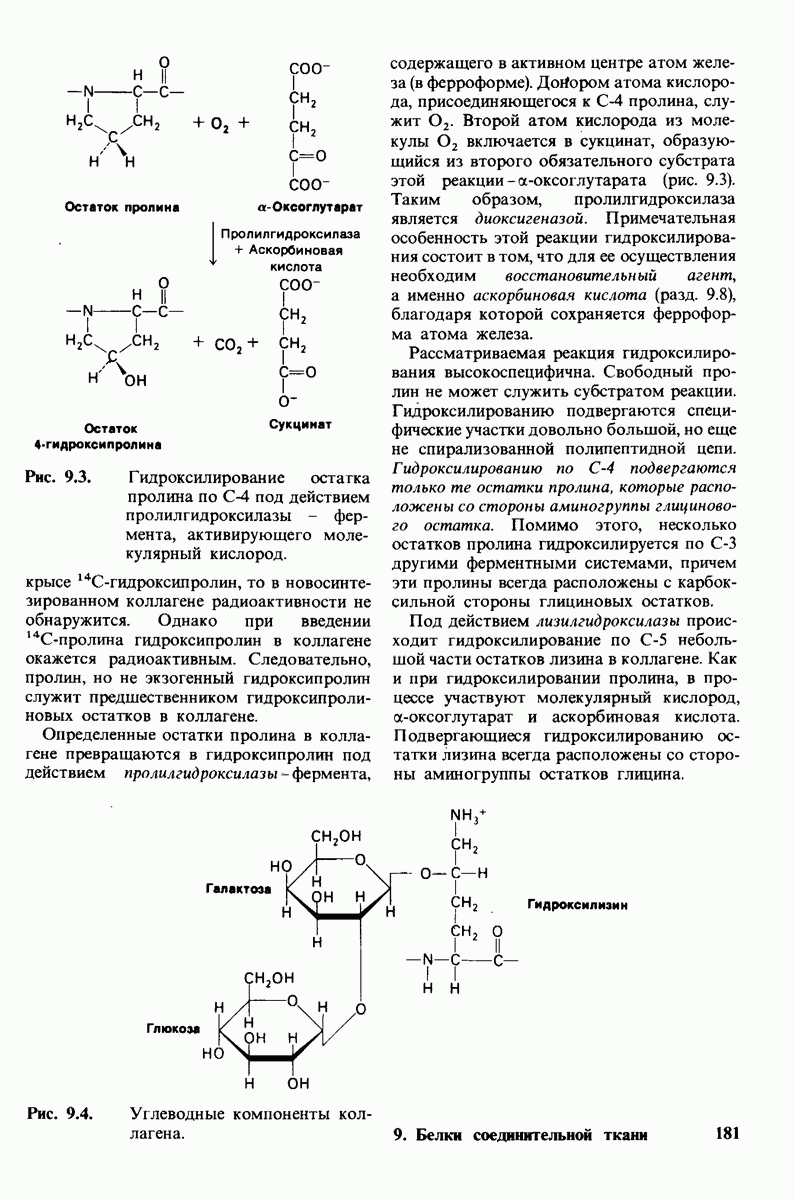
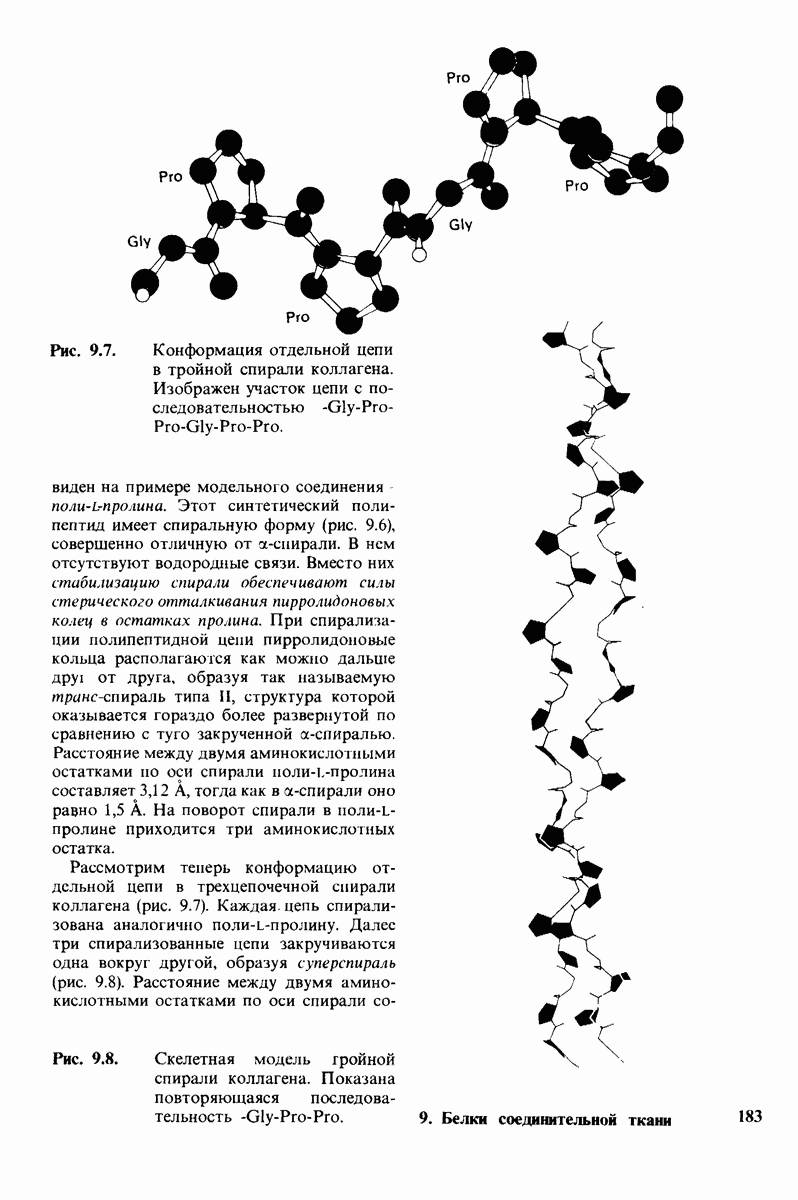
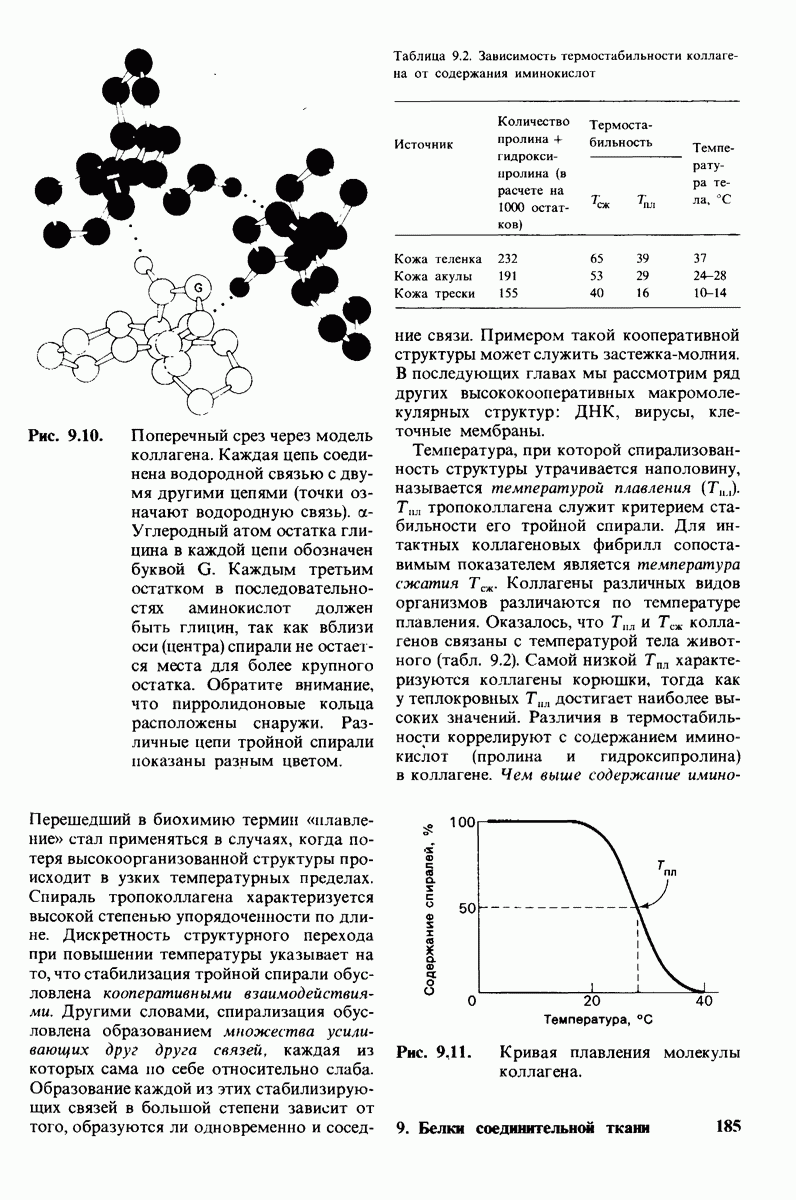
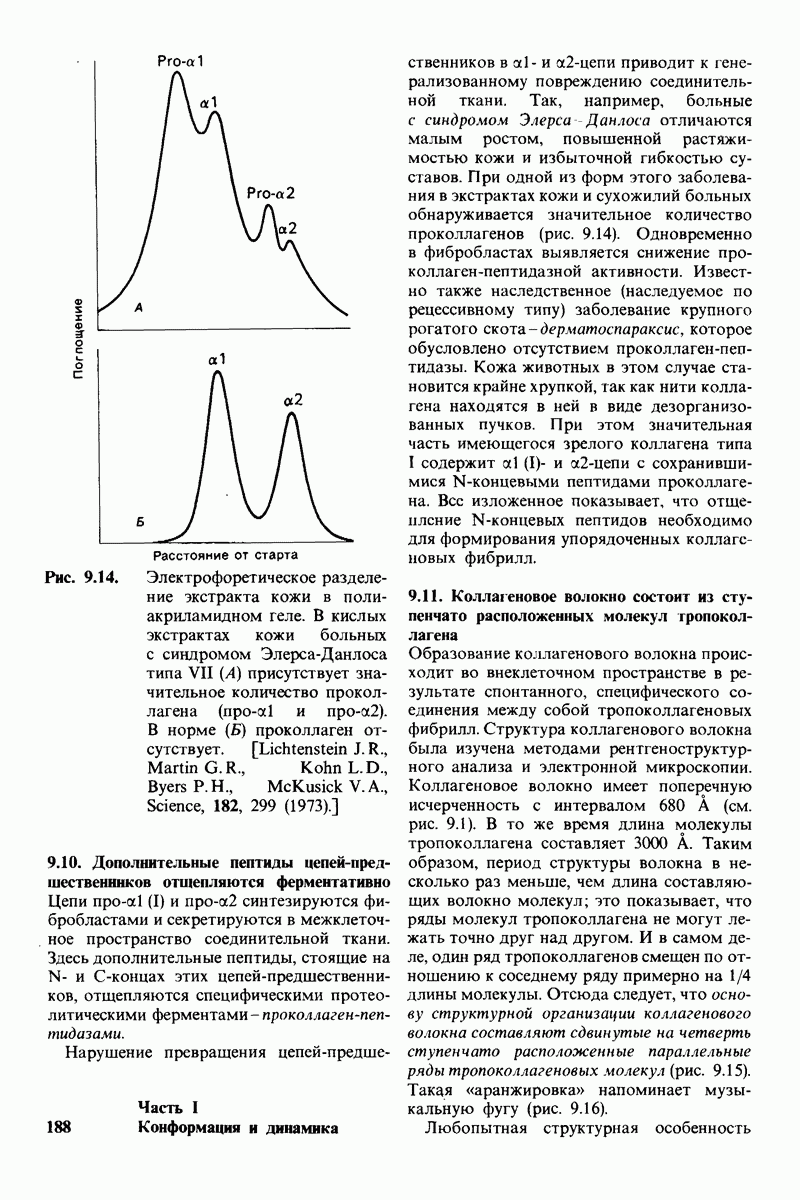
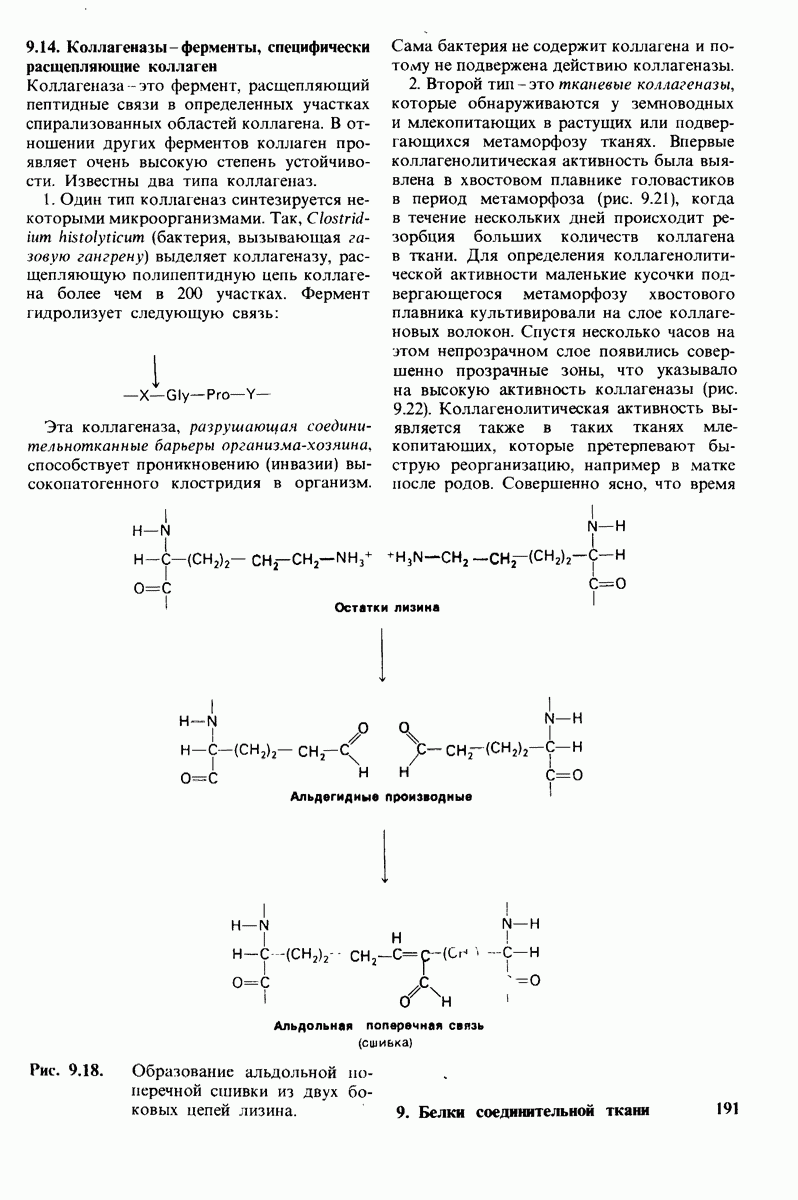
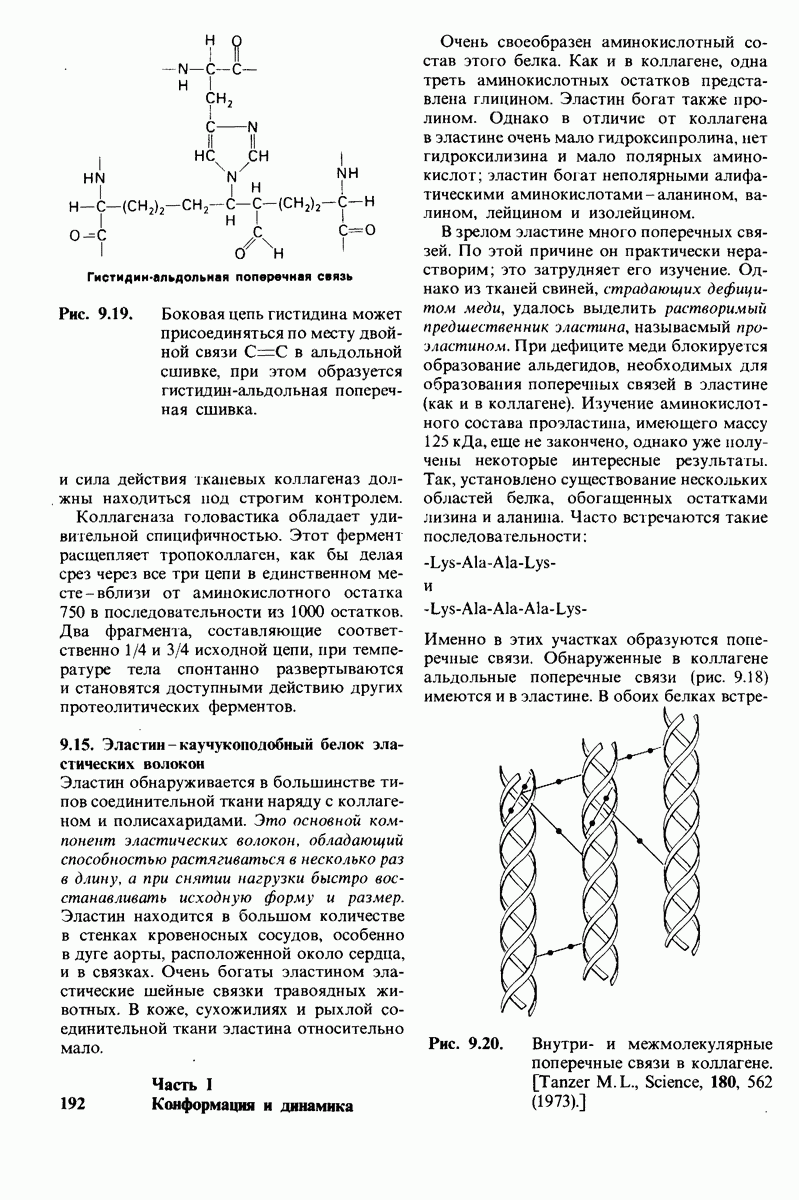
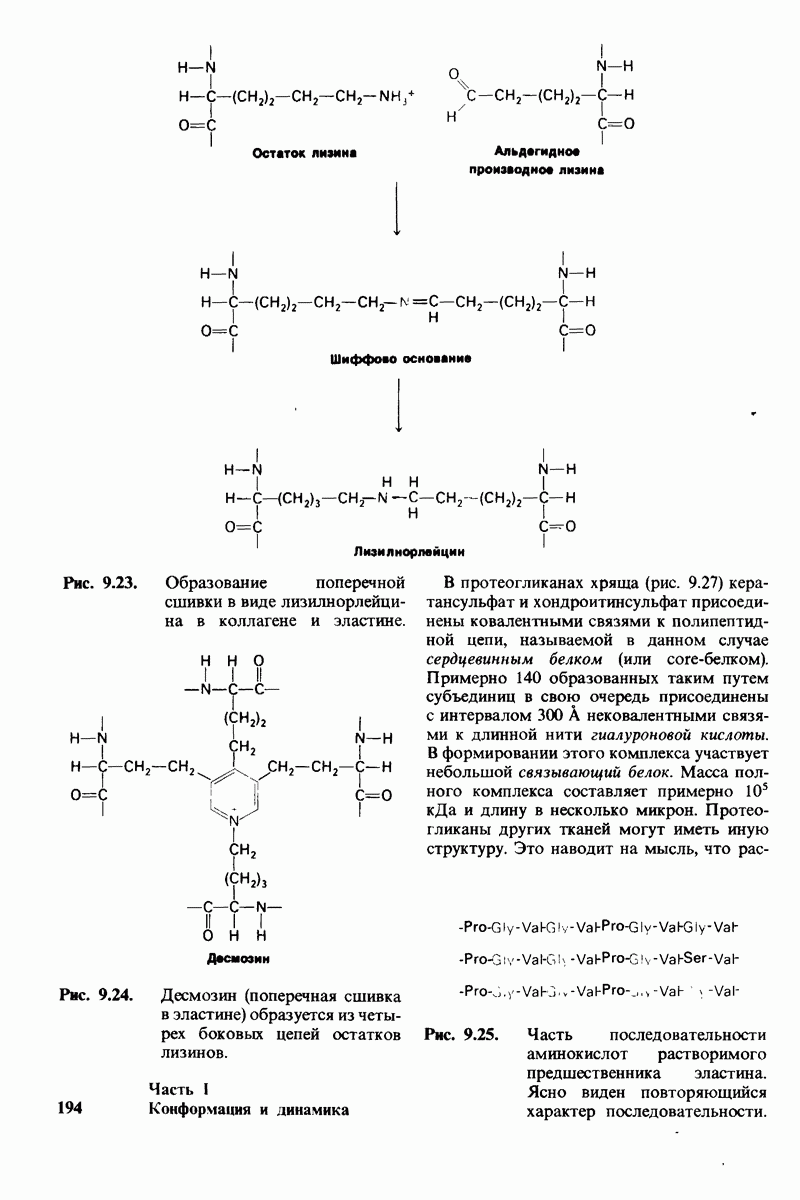
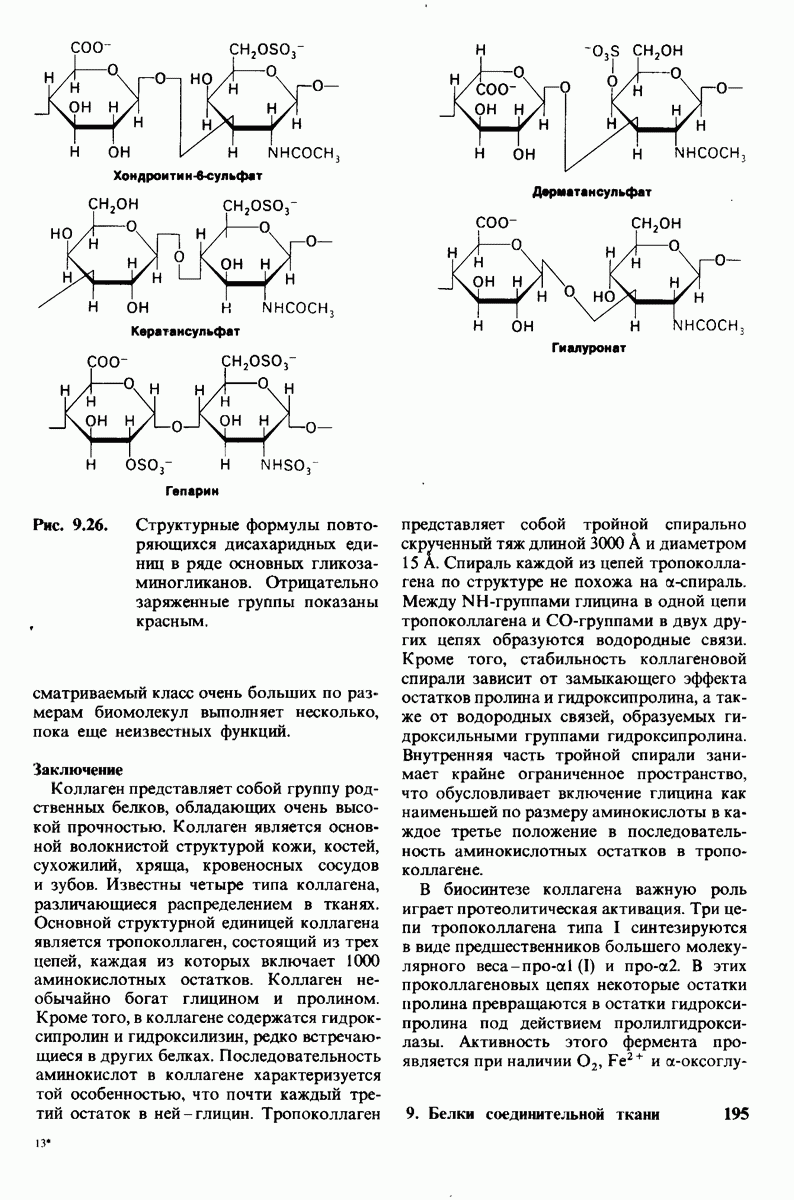
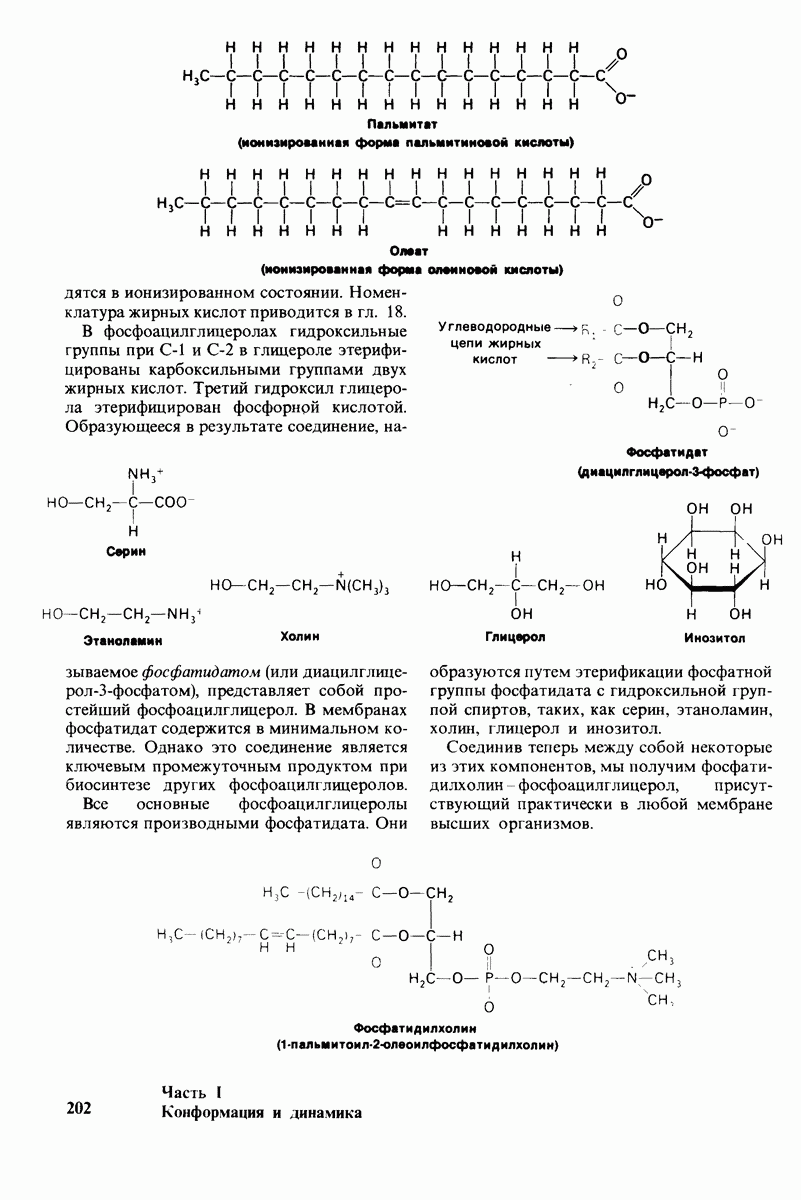
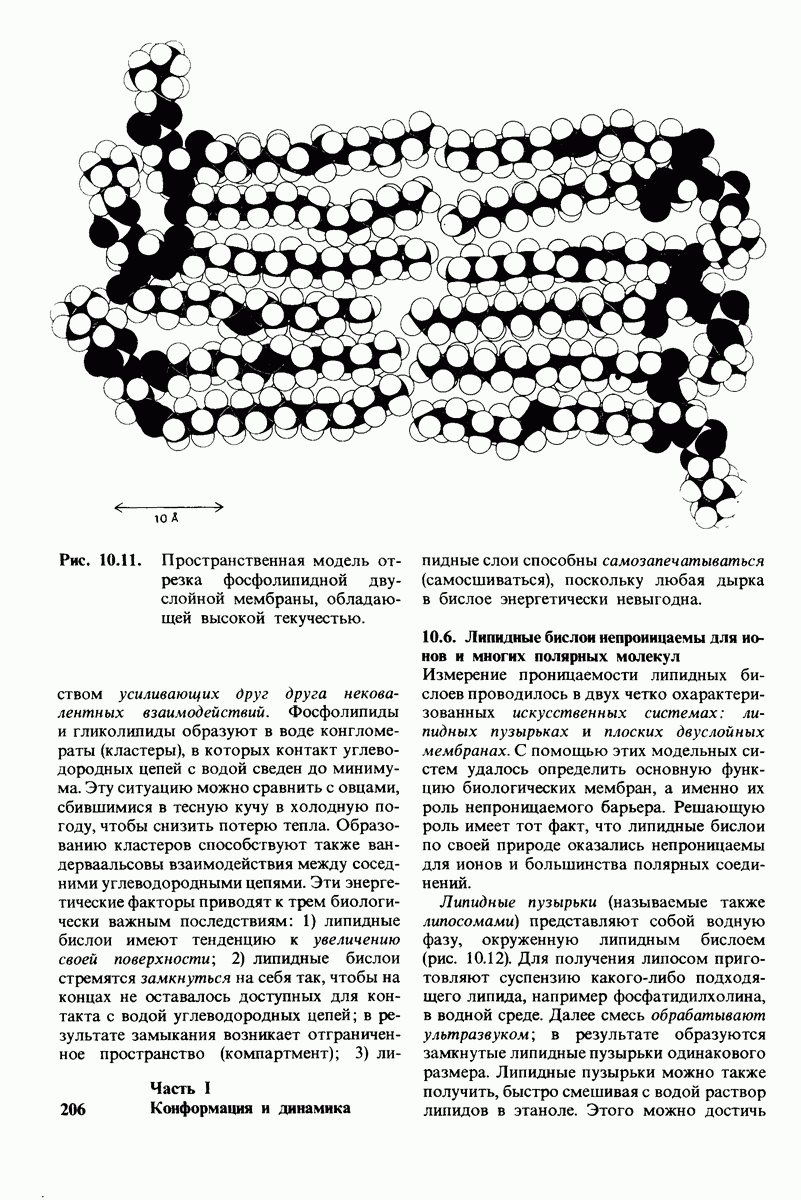
8.8. В химотрипснне имеется глубокий карман для связывания ароматической боковой цепи
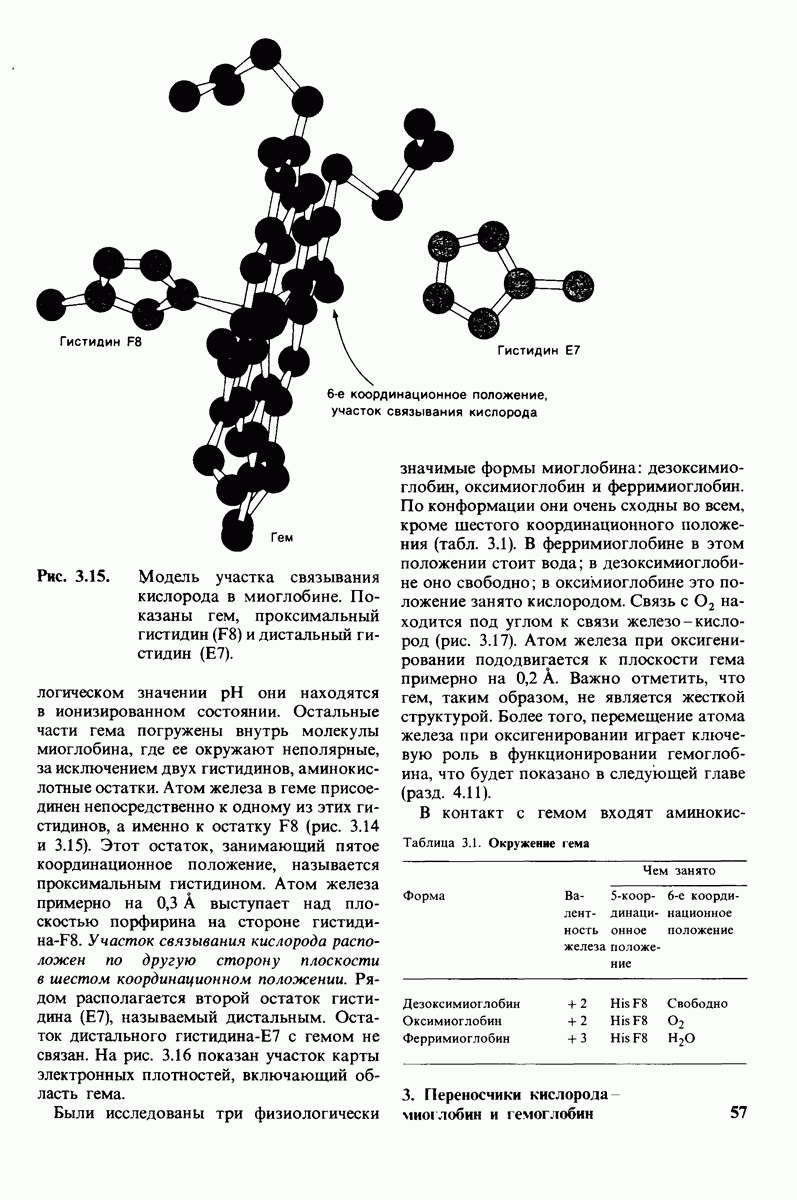
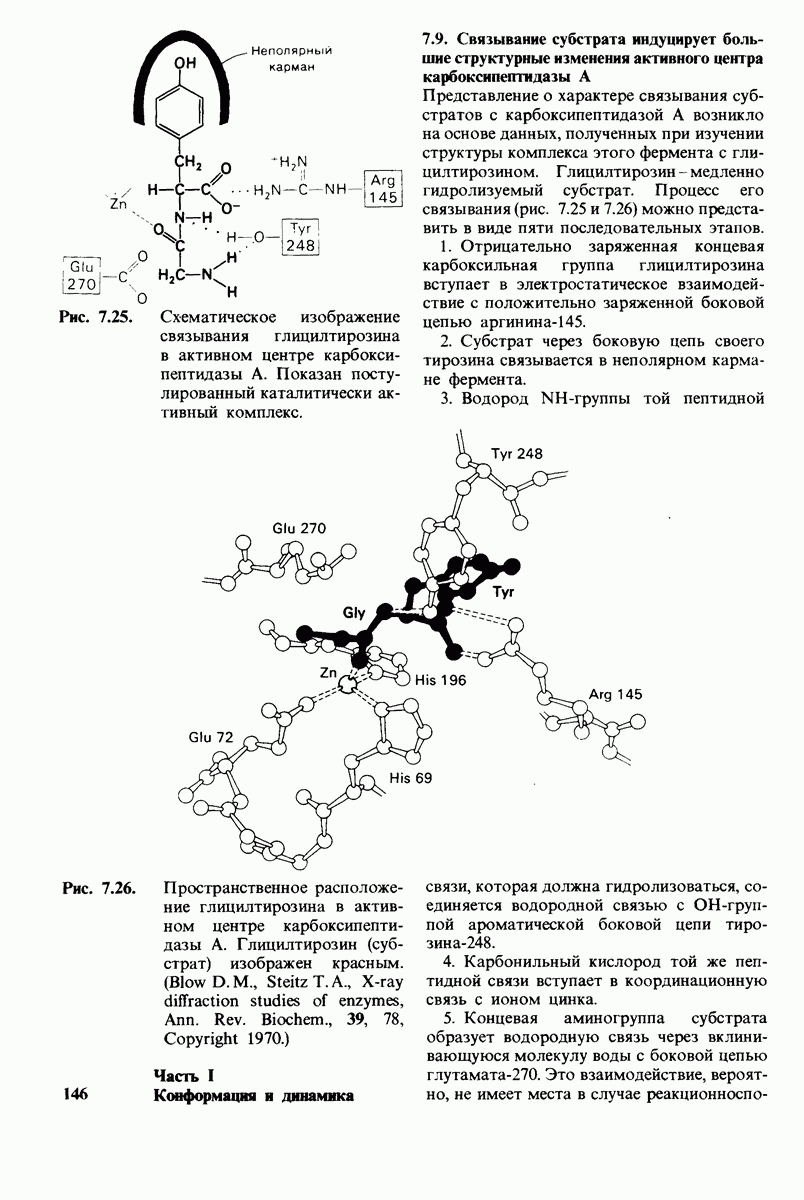
Локализация участков специфического связывания и вероятная ориентация гидролизуемой пептидной связи эффективного субстрата были выявлены в результате рентгеноструктурного анализа комплексов химотрипсина с аналогами субстрата. Установлено, что формил- -триптофан связывается с химотрипсином через индольную
-триптофан связывается с химотрипсином через индольную

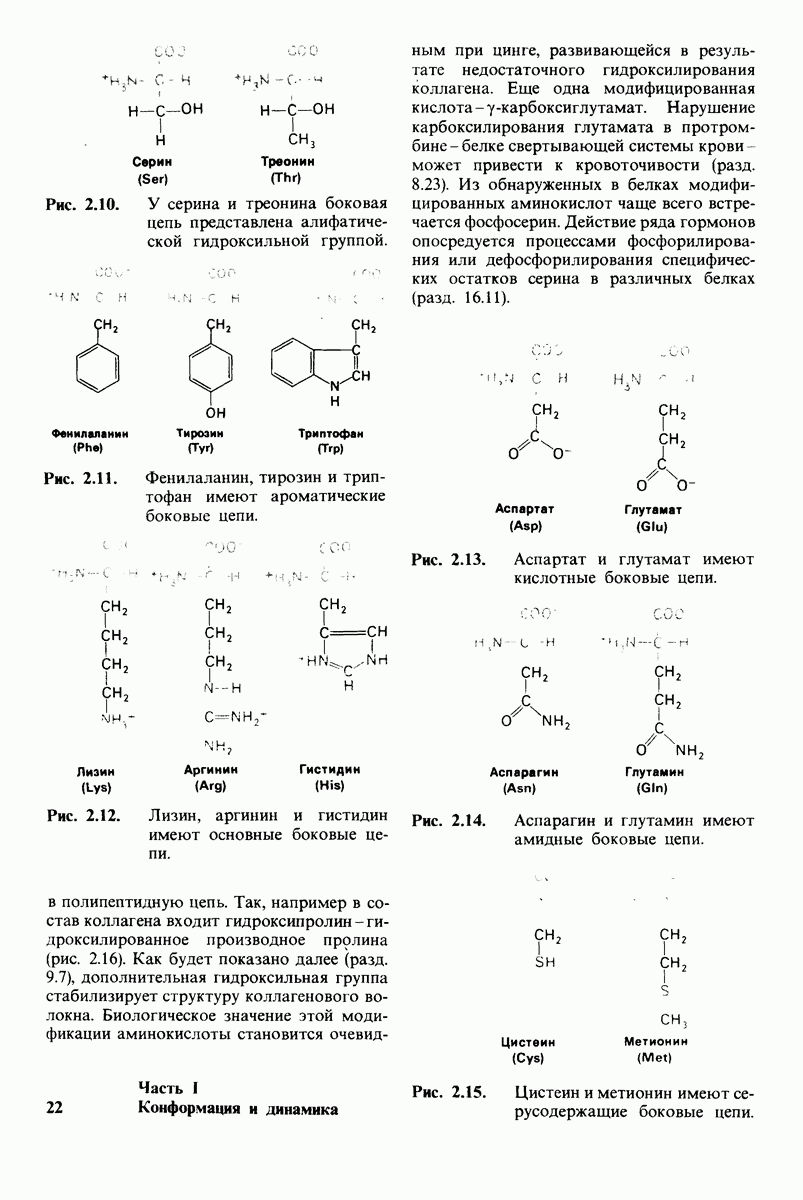
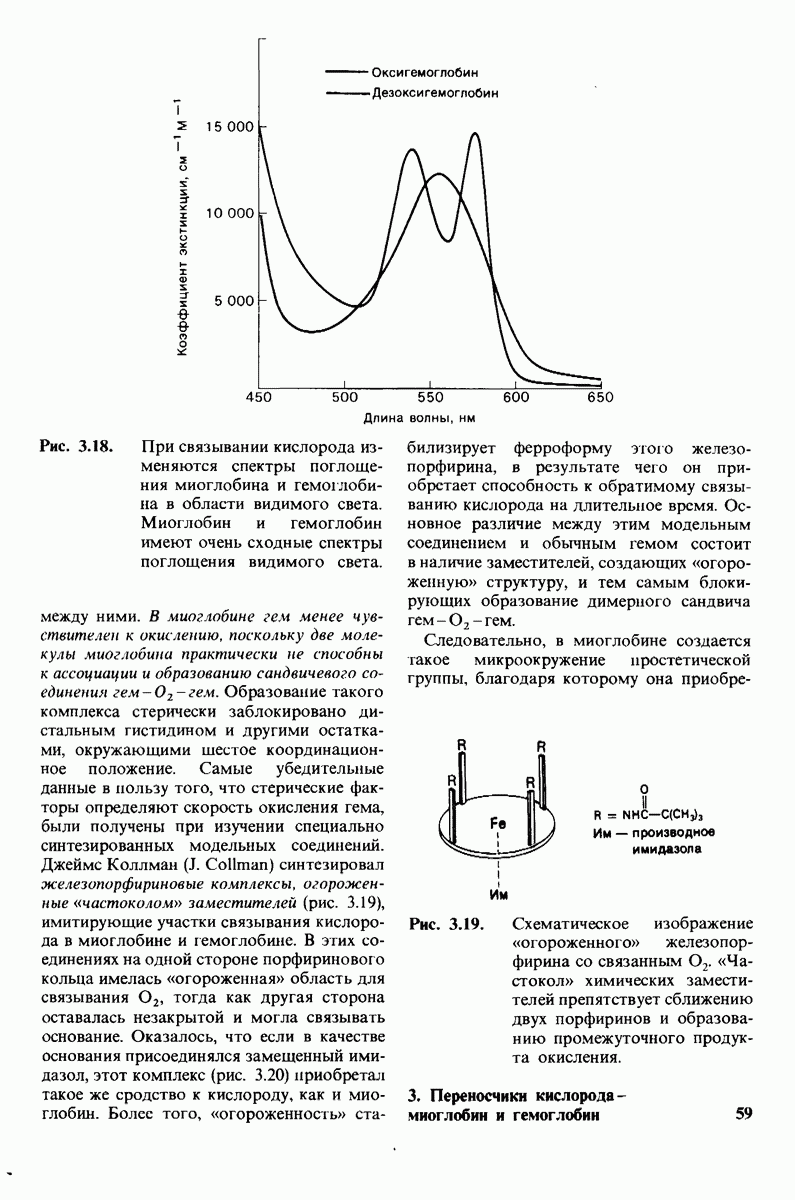
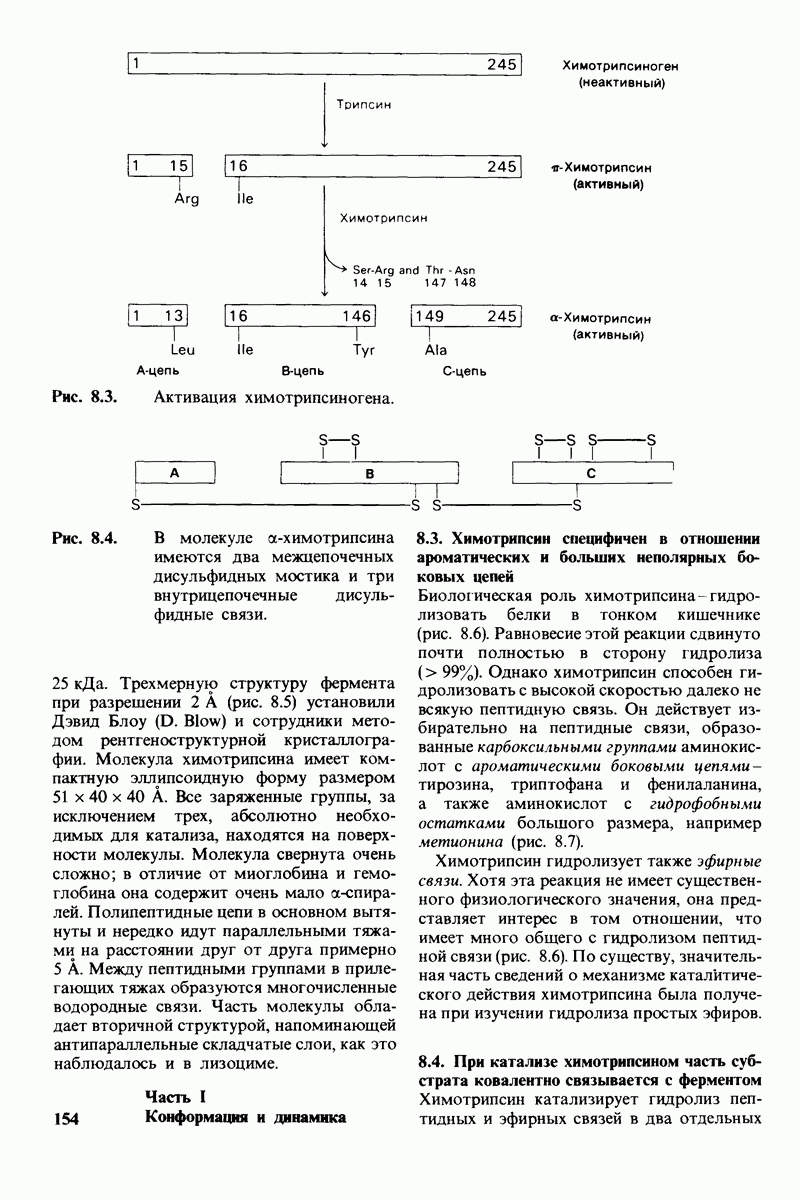
Рис. 8.13. Алкилирование гистидина-57 в химотрипсине воздействием ТФХК.

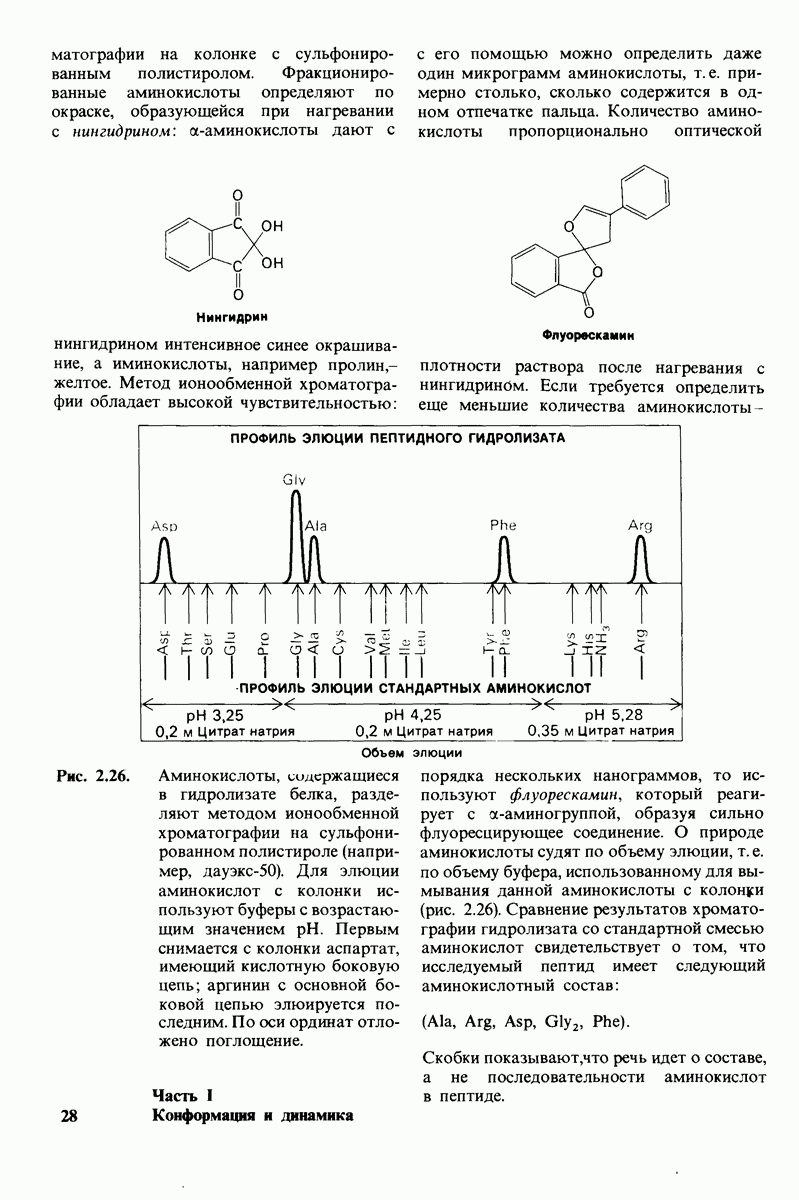
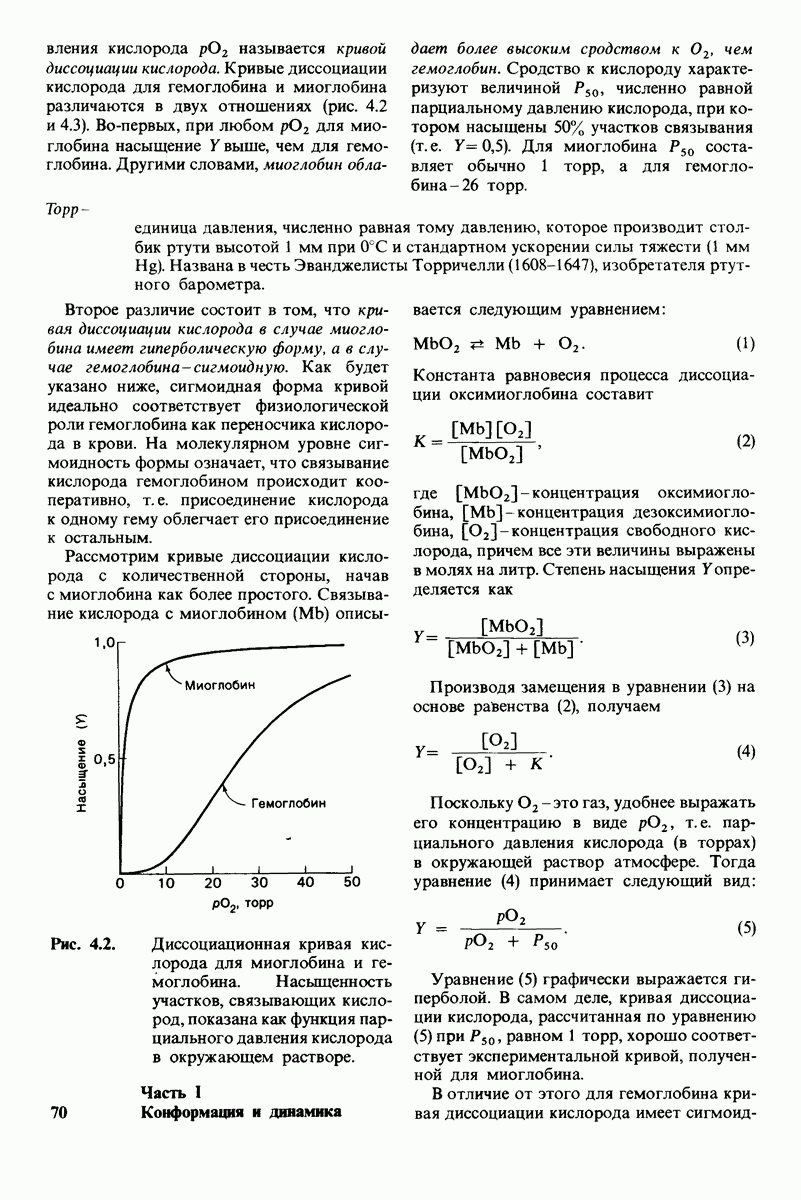
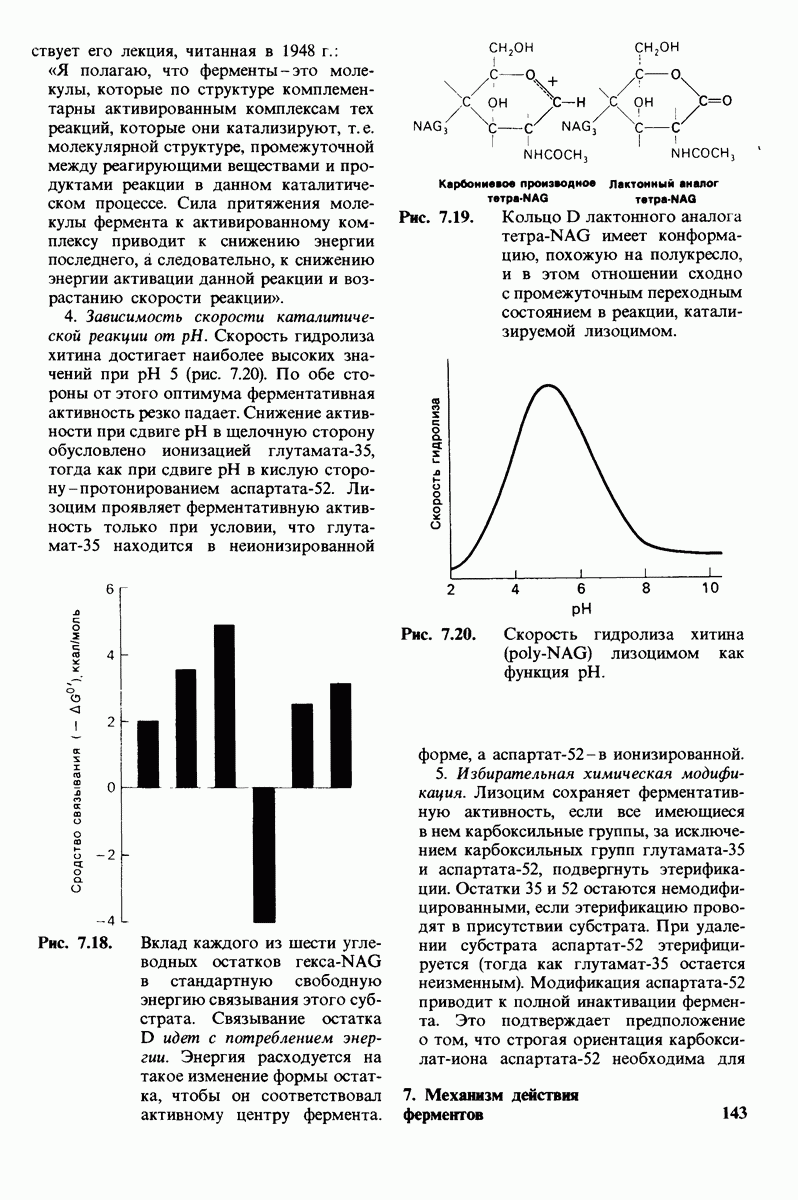
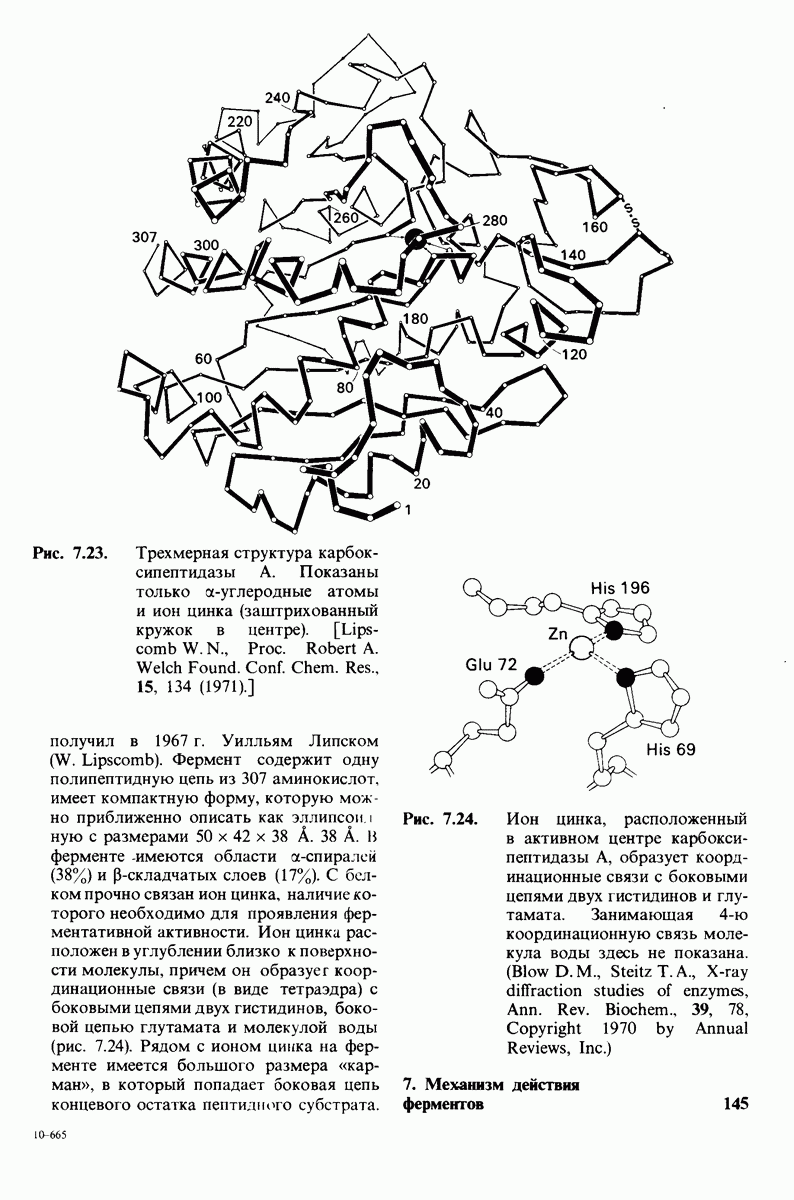
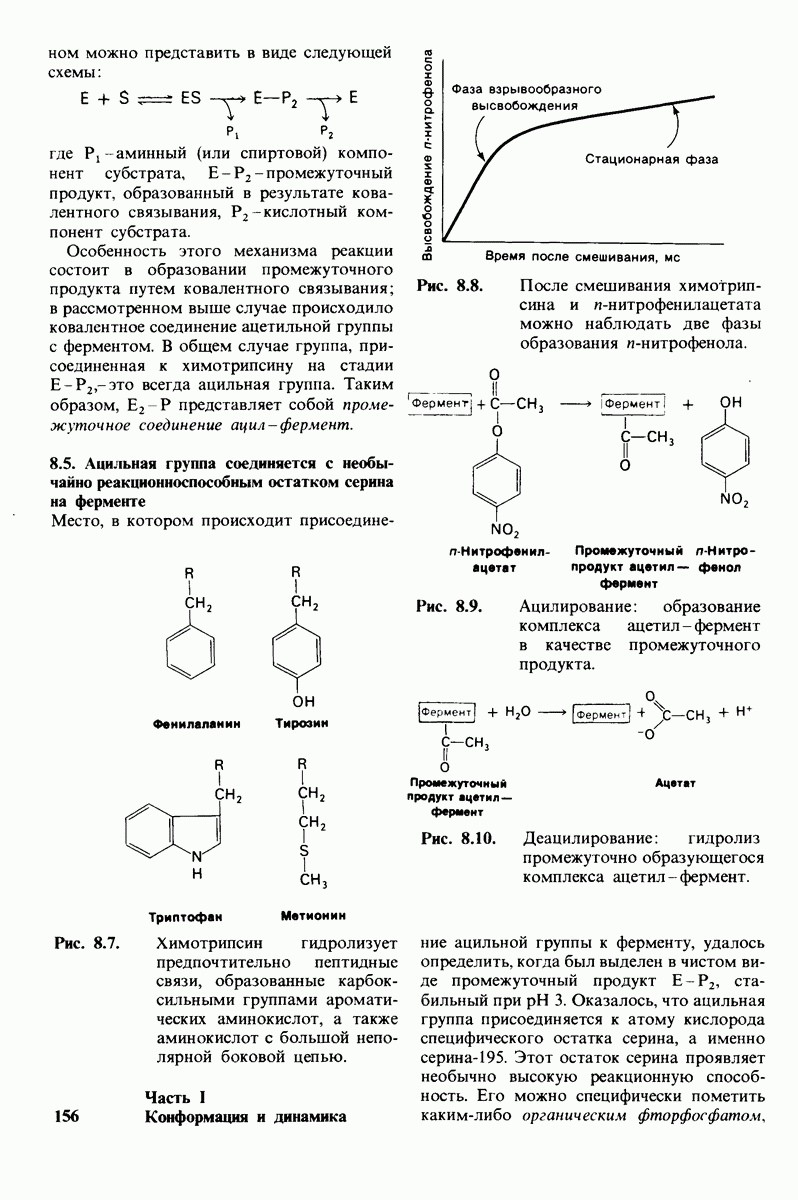
Рис. 8.14. Конформация системы переноса заряда в химотрипсине. [Blow D.M., Steitz Т. А., Х-гау diffraction studies of enzymes, Ann. Rev. Biochem., 39, 86 (1976).]

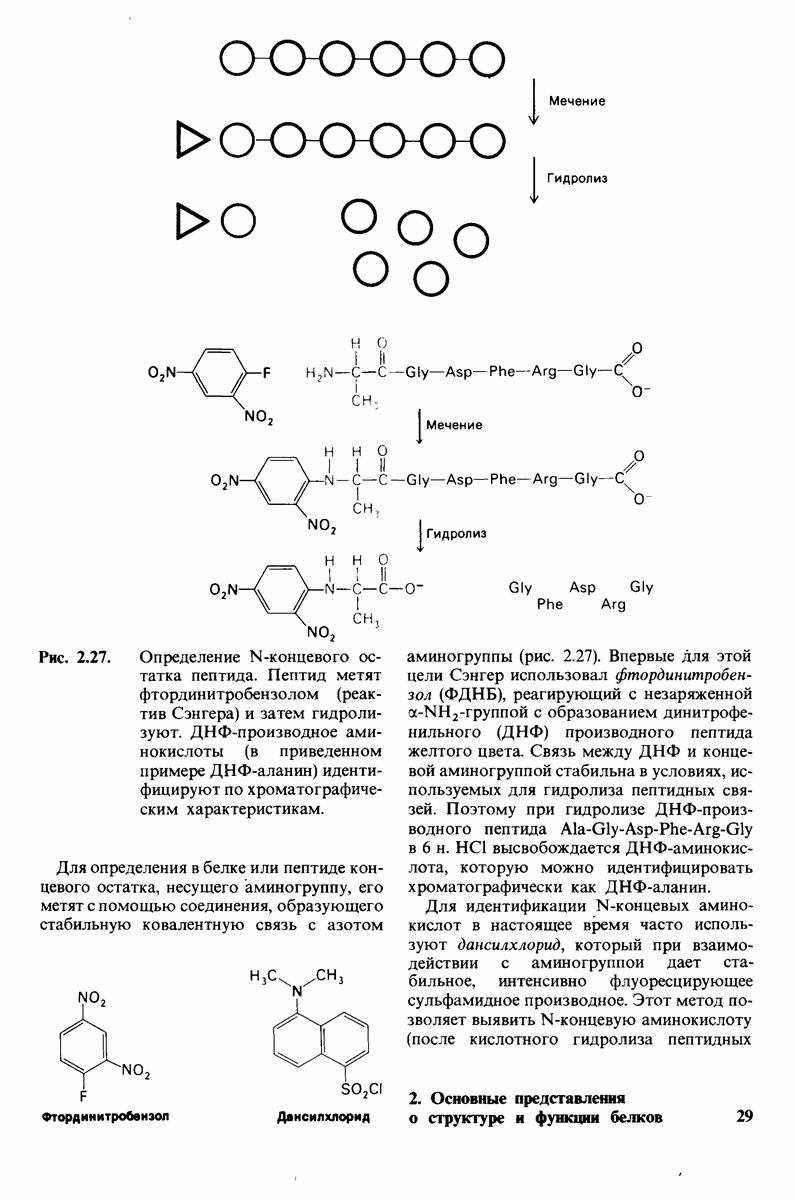
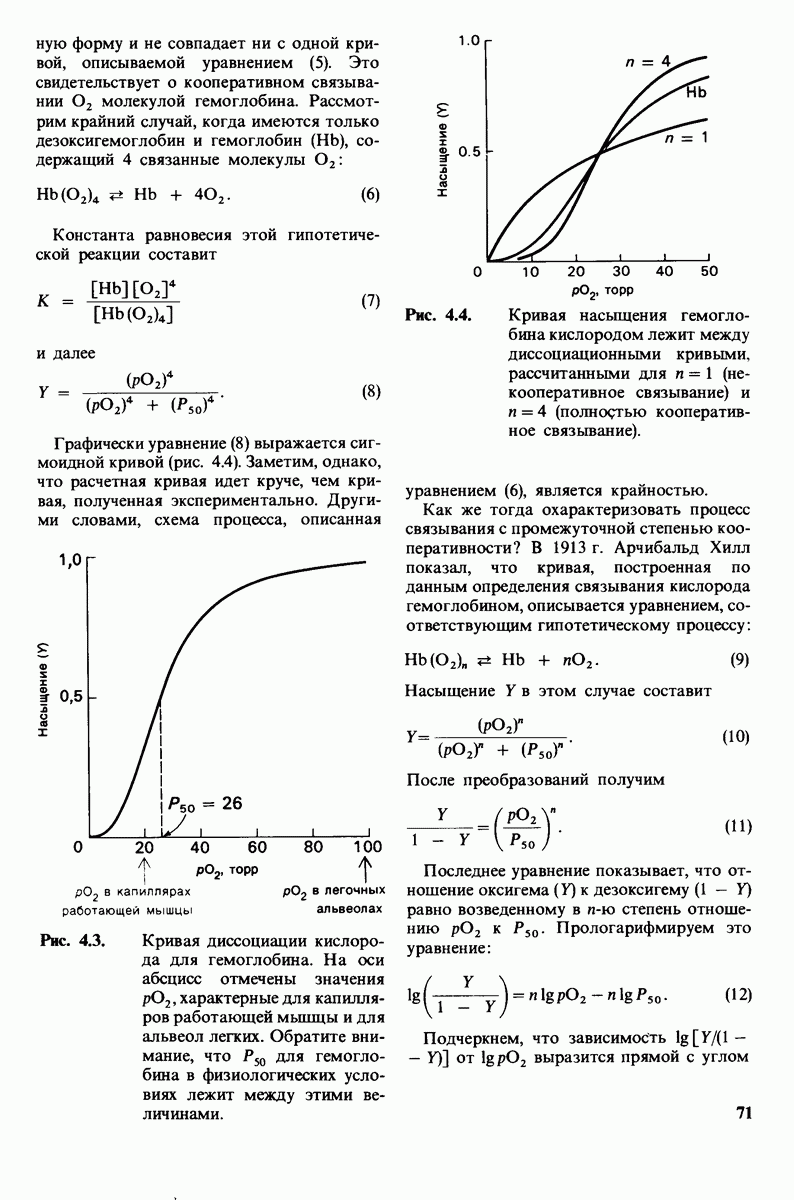
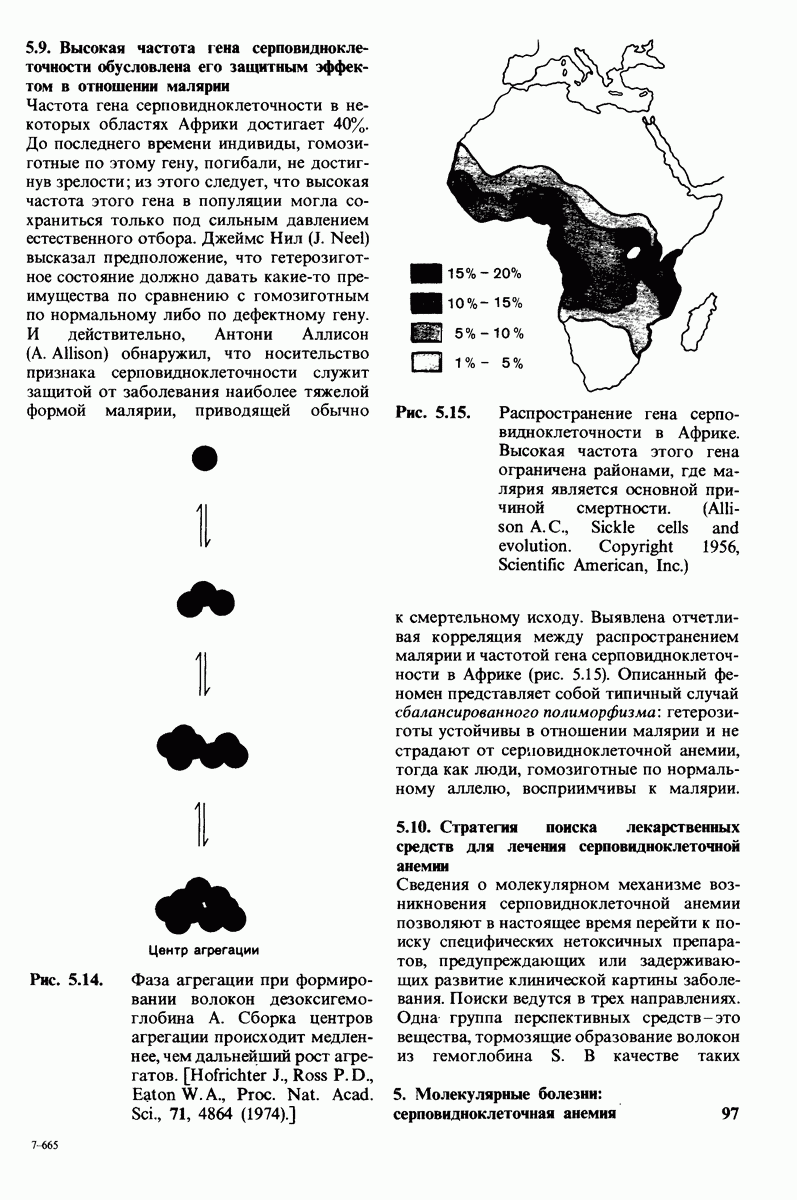
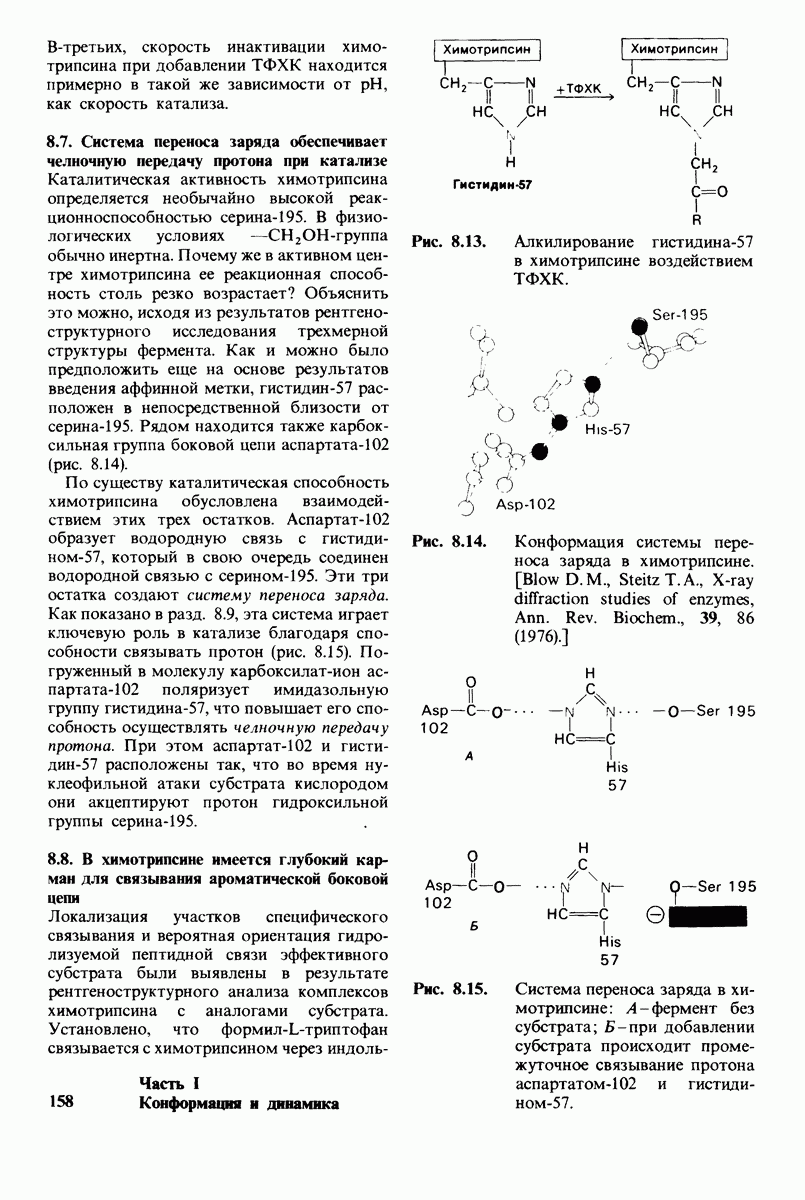
Рис. 8.15. Система переноса заряда в химотрипсине  А - фермент без субстрата; Б - при добавлении субстрата происходит промежуточное связывание протона аспартатом-102 и гистидином-57.
А - фермент без субстрата; Б - при добавлении субстрата происходит промежуточное связывание протона аспартатом-102 и гистидином-57.
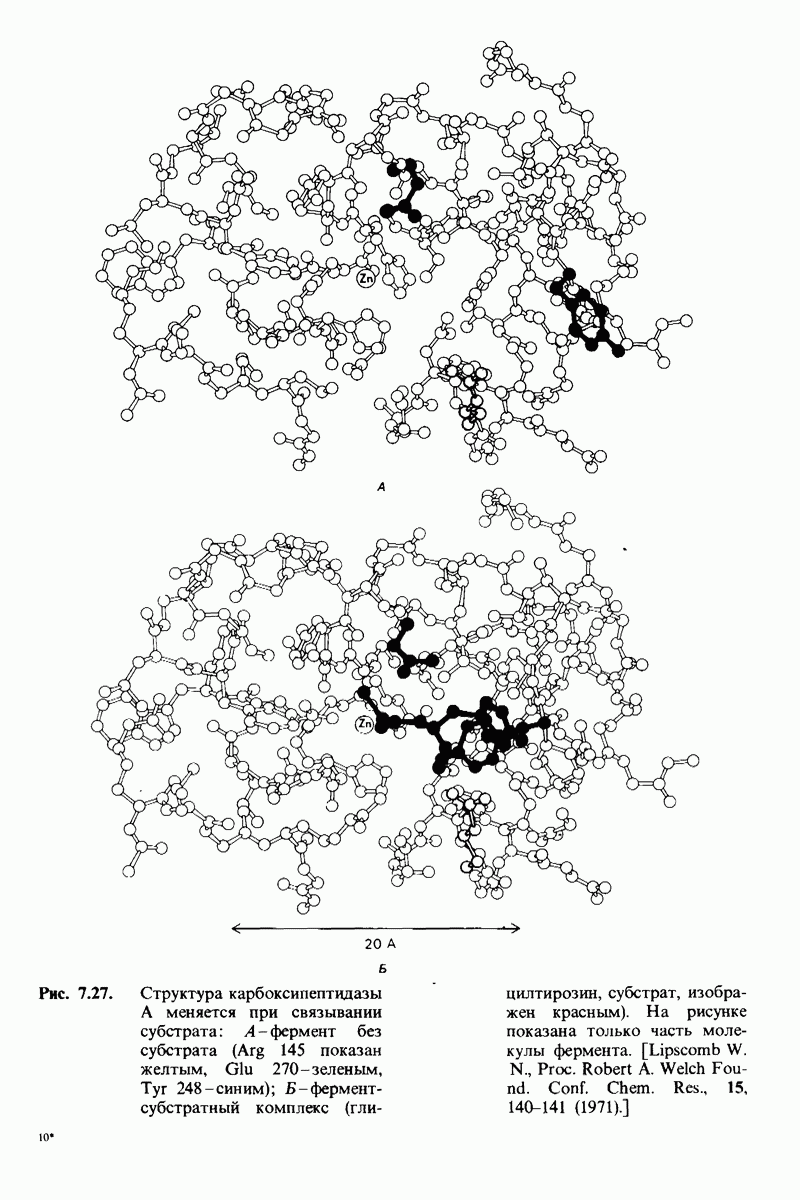
боковую цепь, которая хорошо соответствует по размеру карману на ферменте вблизи серина-195 (рис. 8.16). Именно наличием этого глубокого кармана объясняется специфичность химотрипсина в отношении аминокислот с ароматической или иной гидрофобной боковой цепью большого размера. При рентгеноструктурном анализе комплексов химотрипсина с аналогами полипептидных субстратов обнаружено большое число водородных связей между основными цепями фермента и субстрата, причем эти водородные связи располагаются так же, как в антипараллельных  -складчатых слоях.
-складчатых слоях.