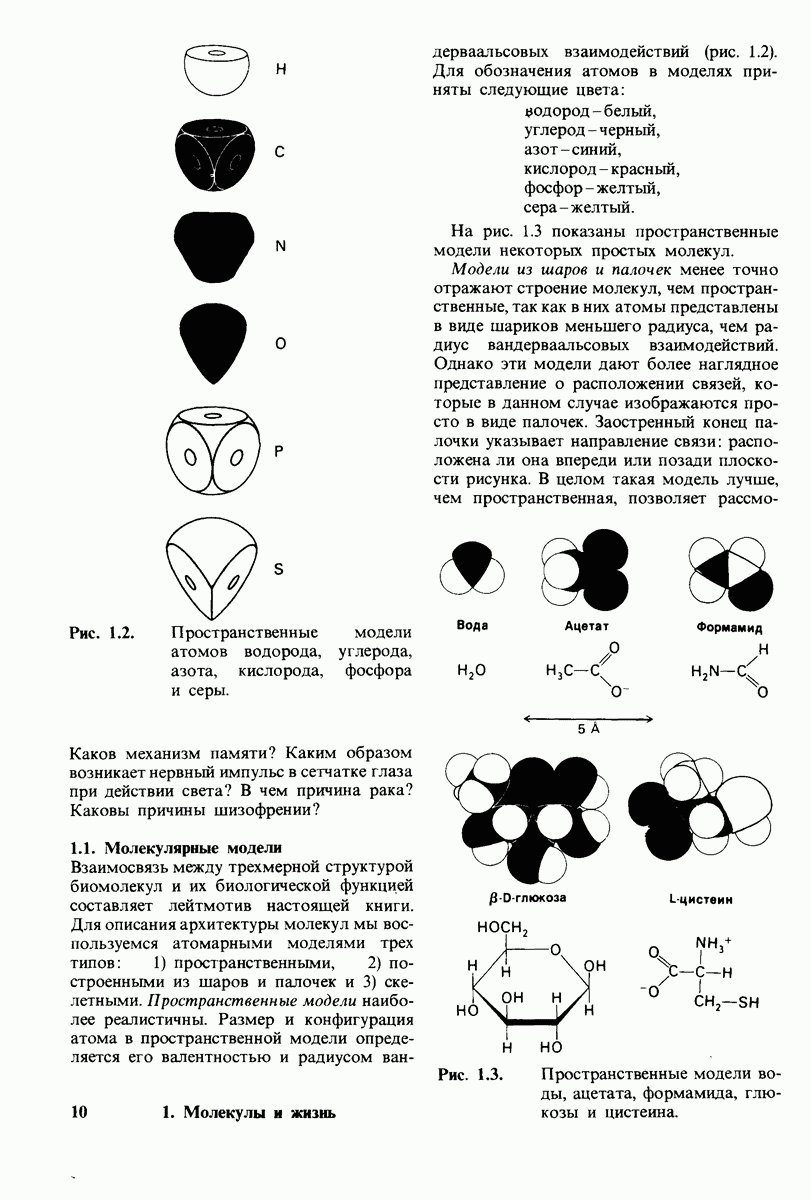
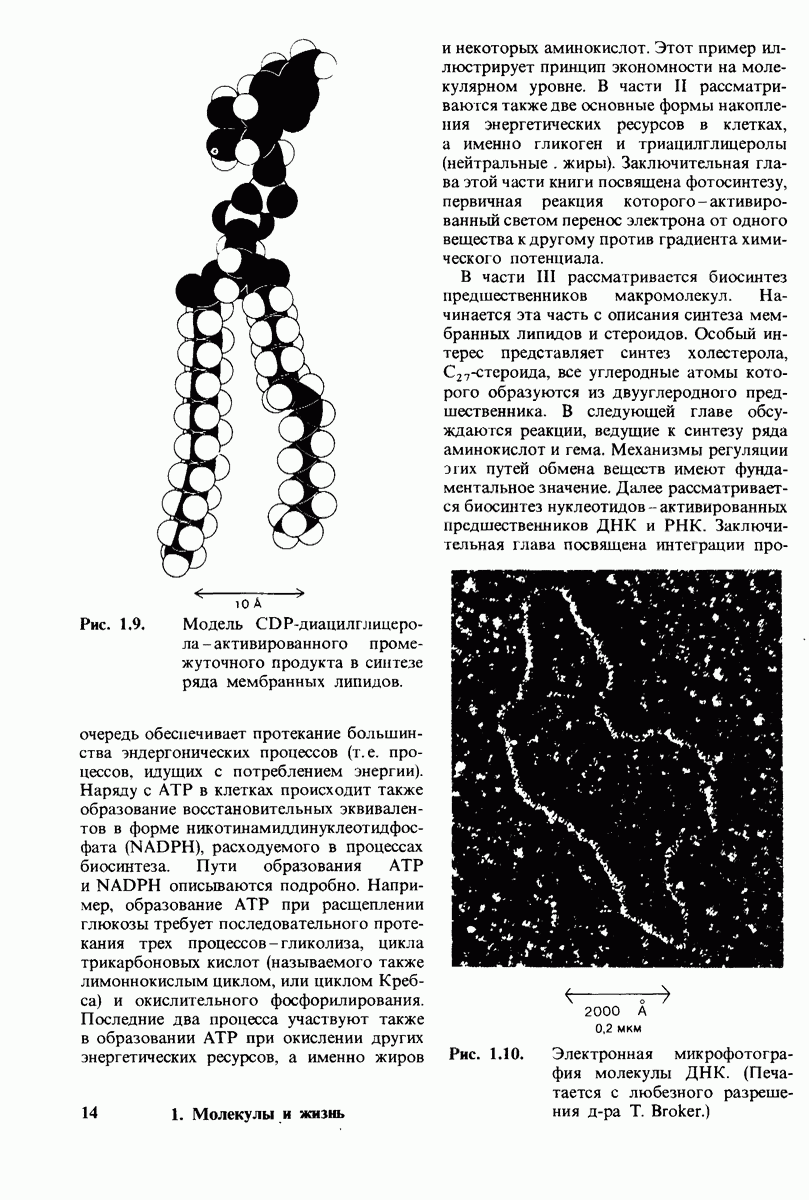
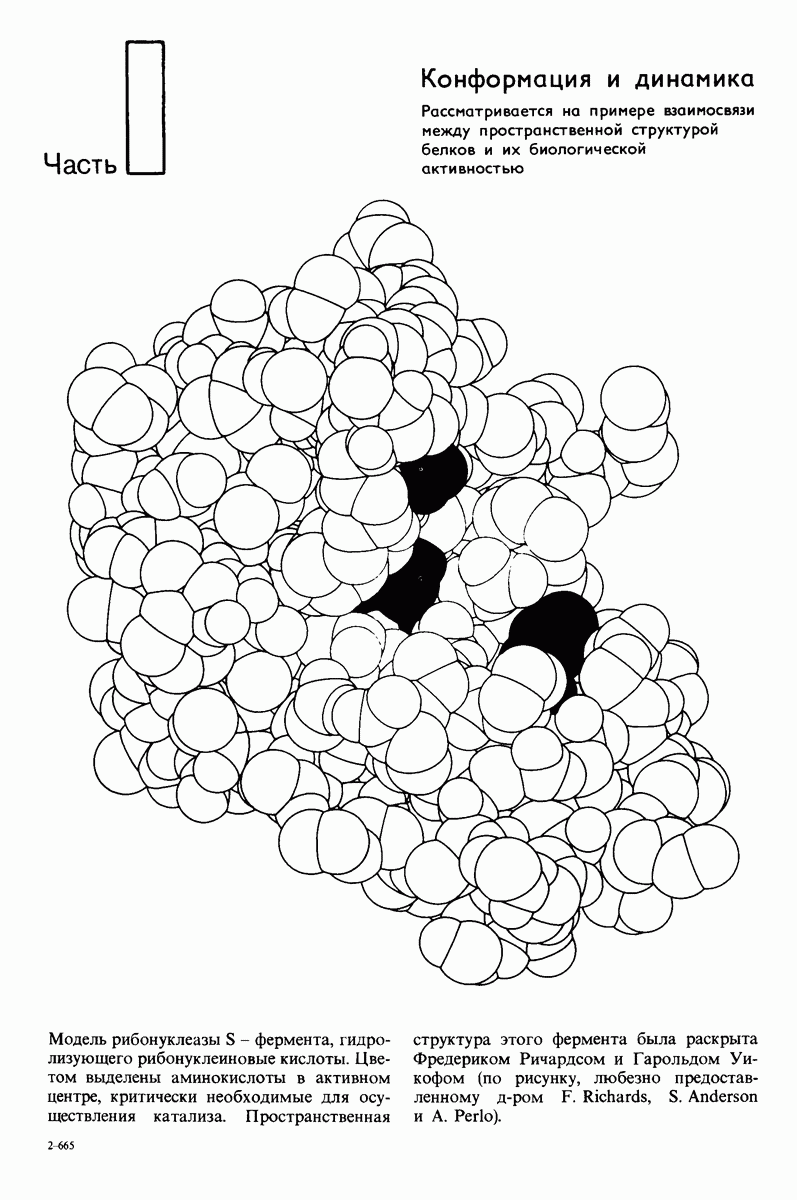
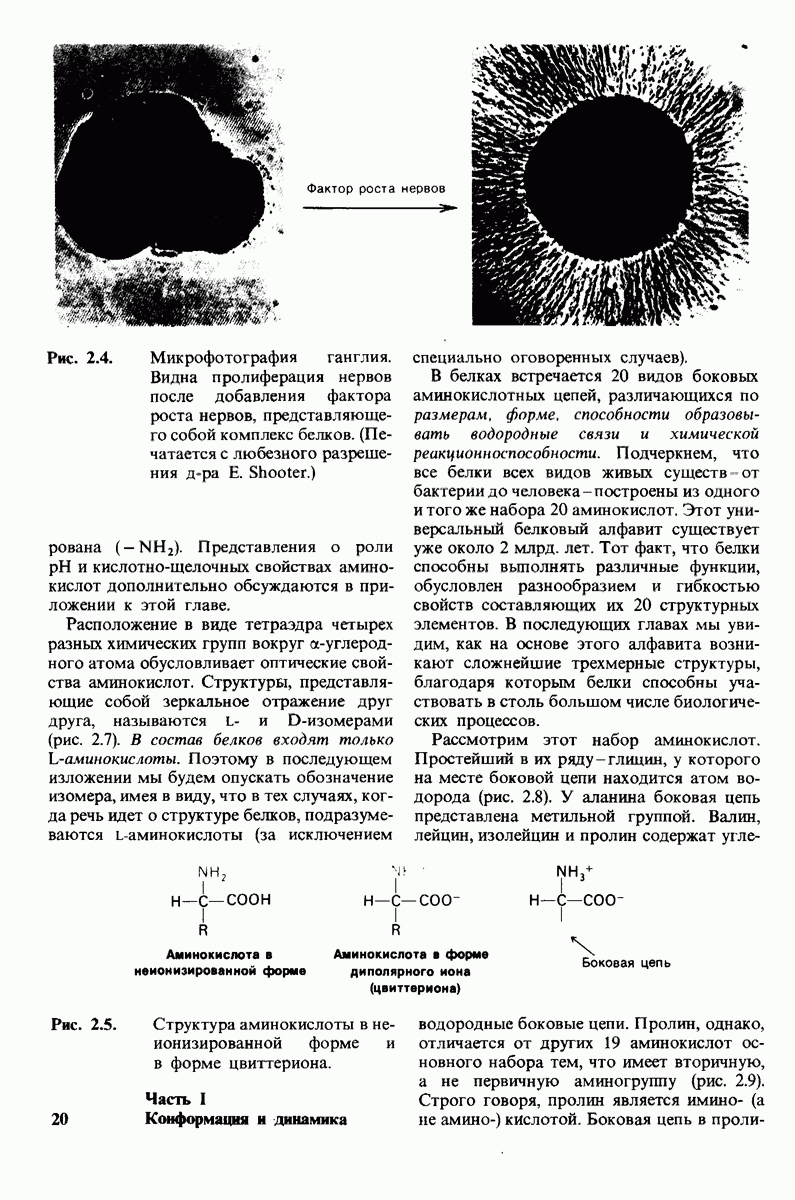
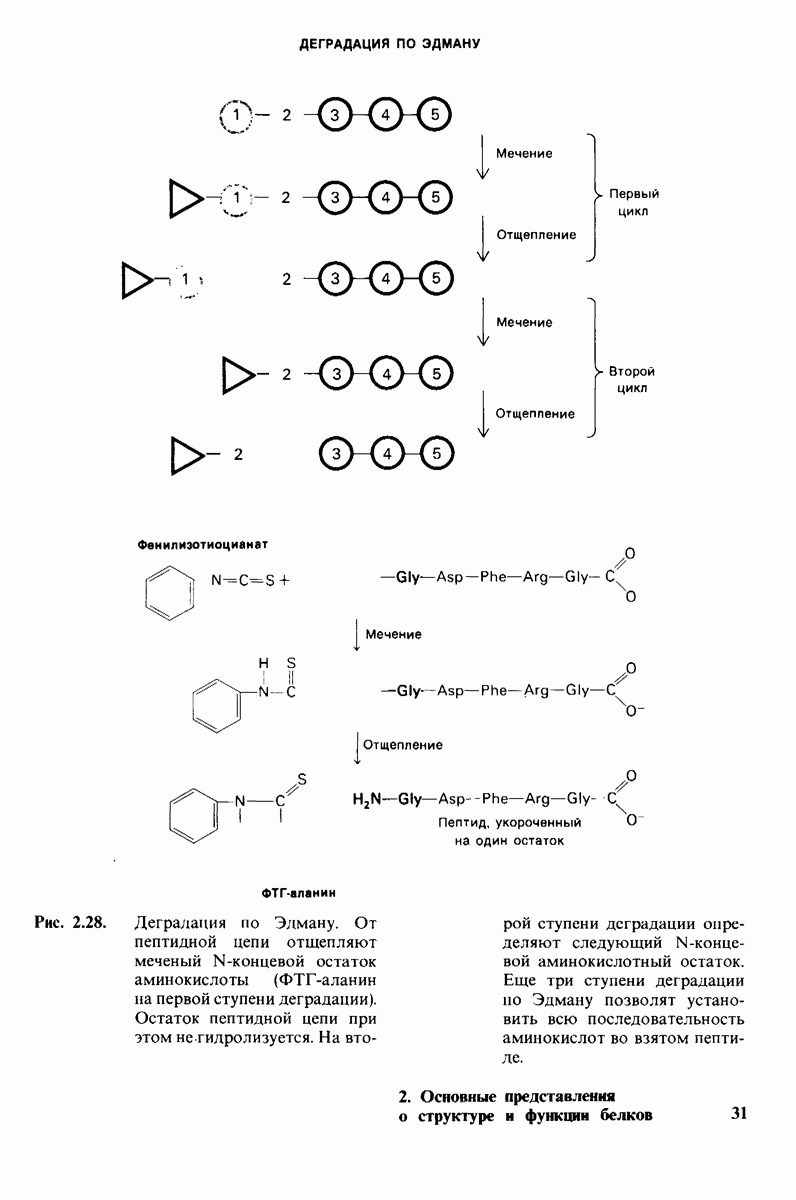
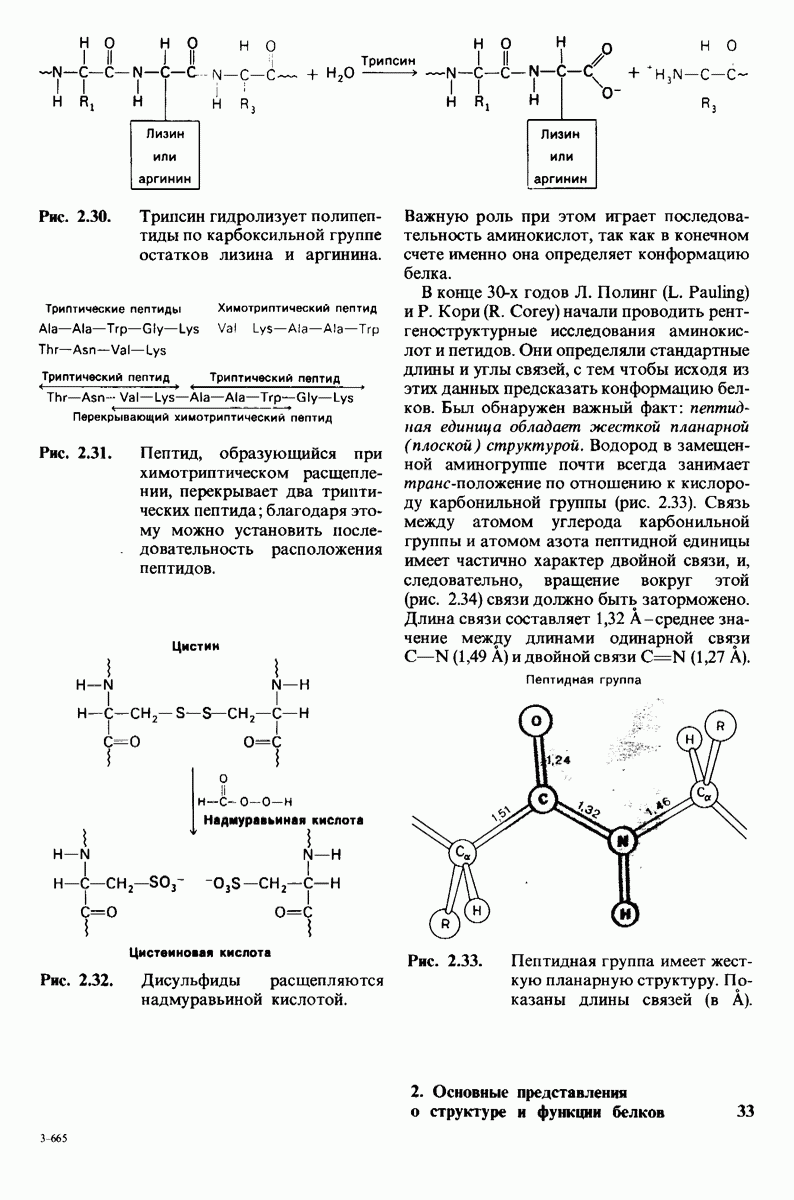
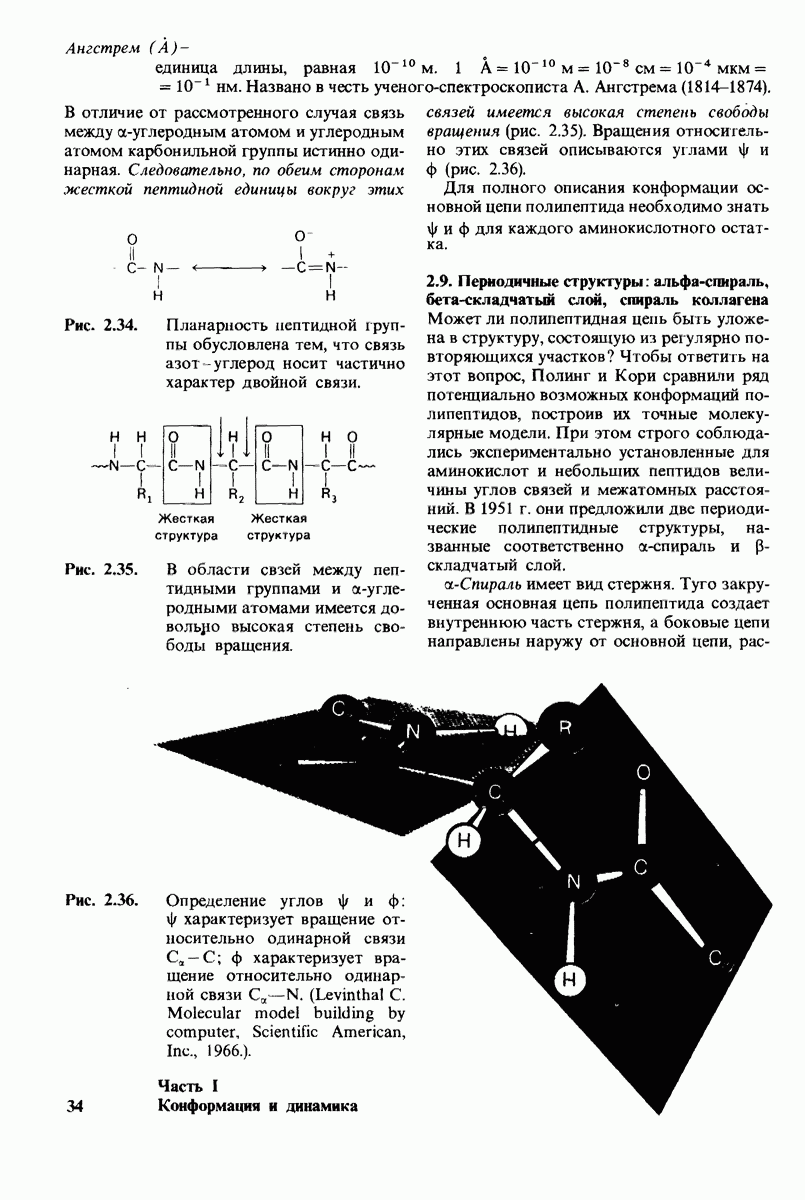
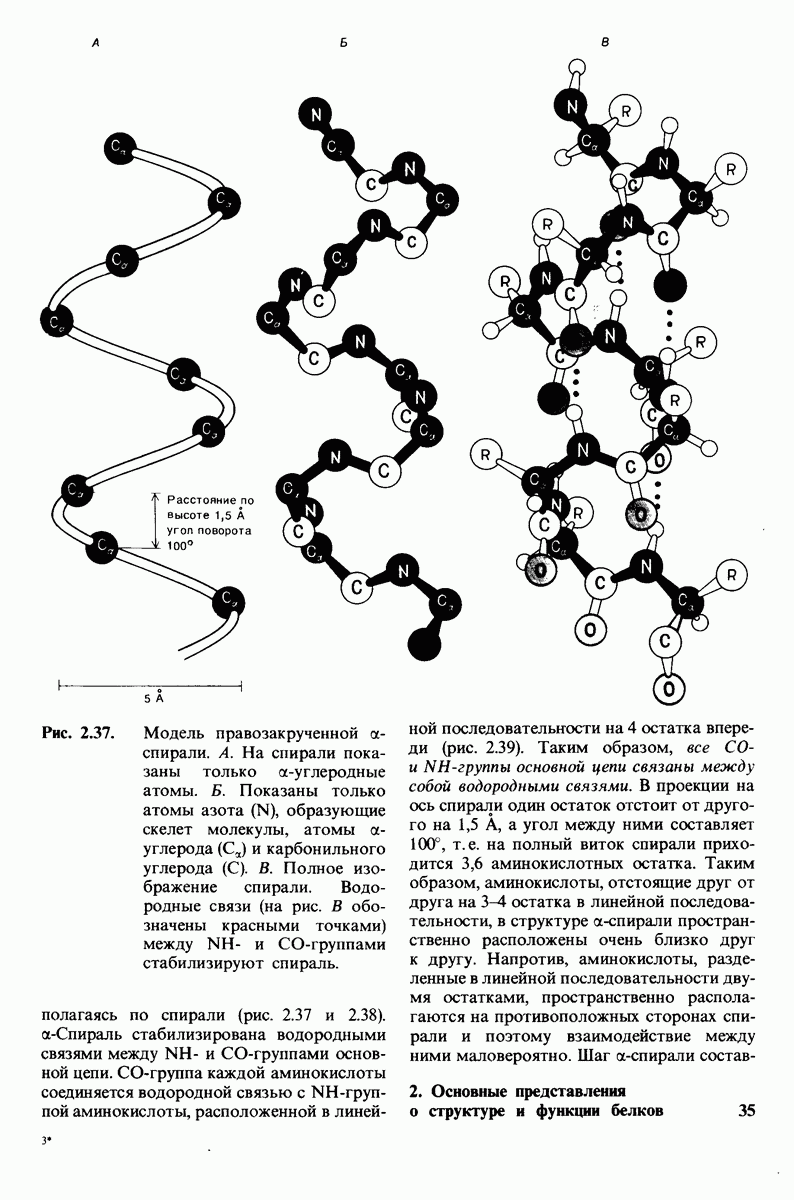
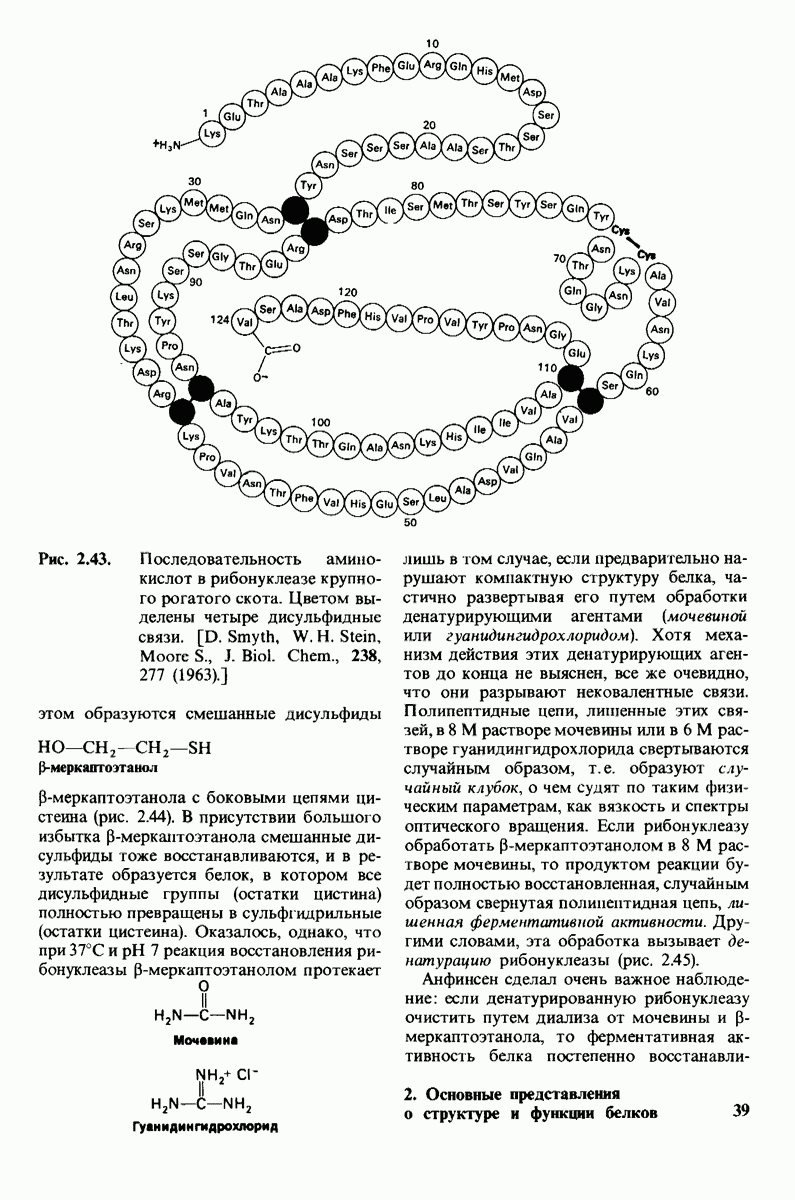
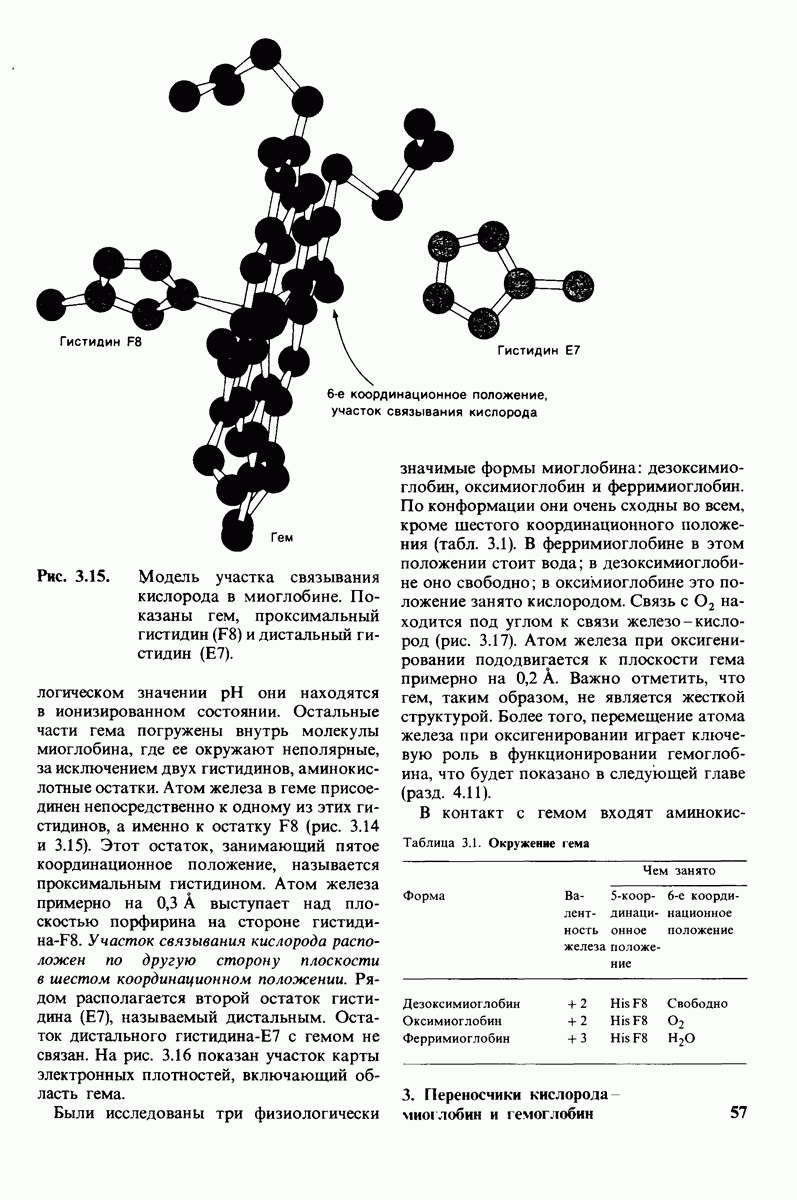
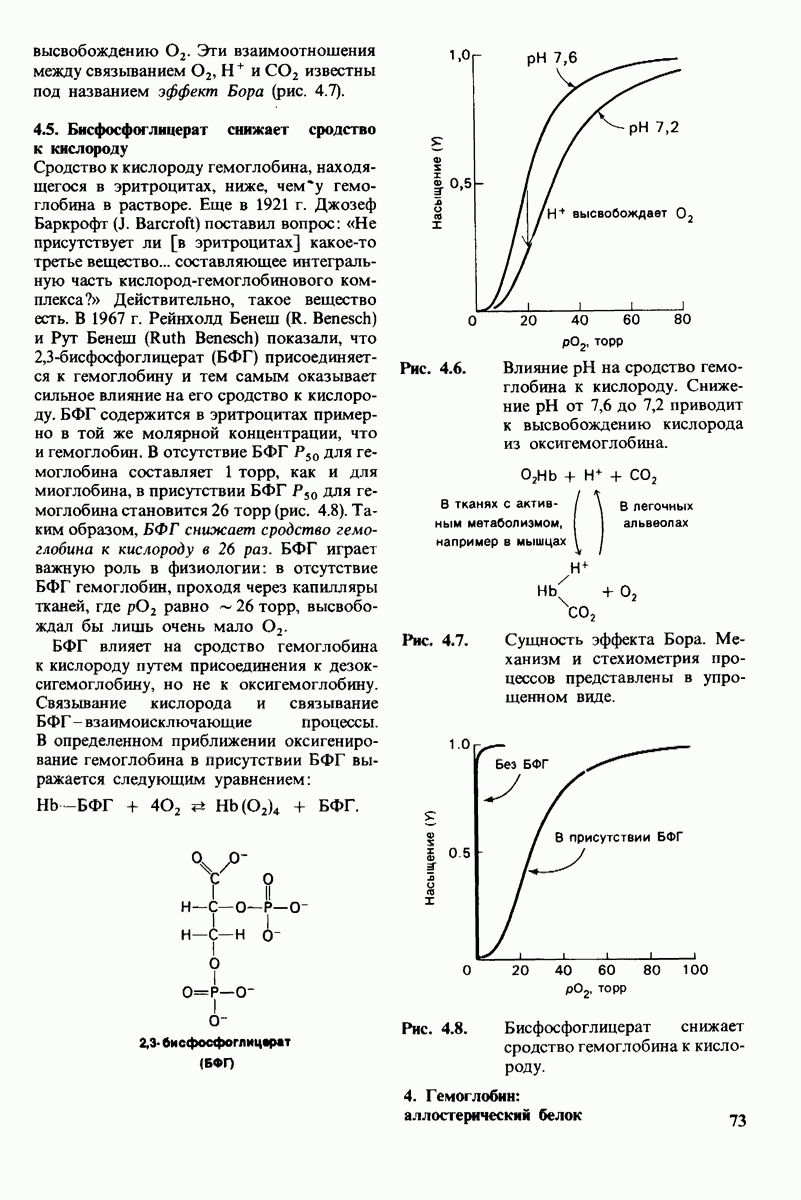
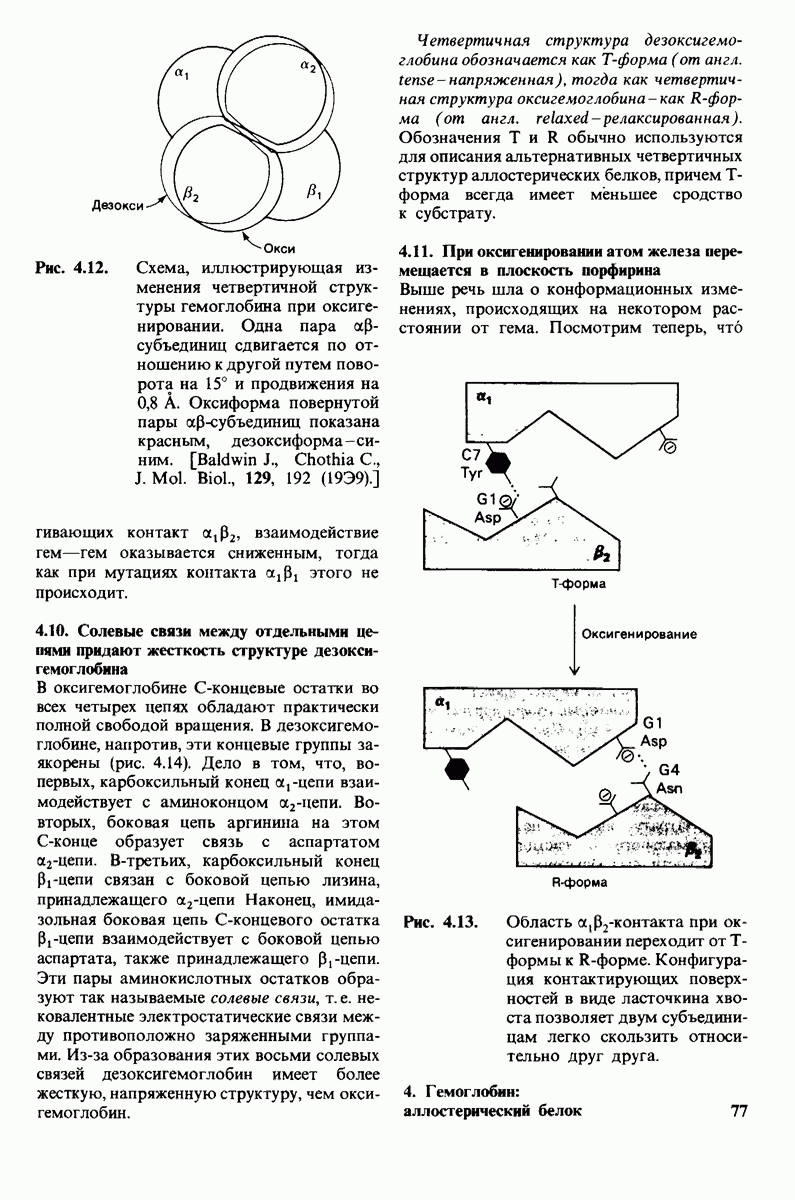
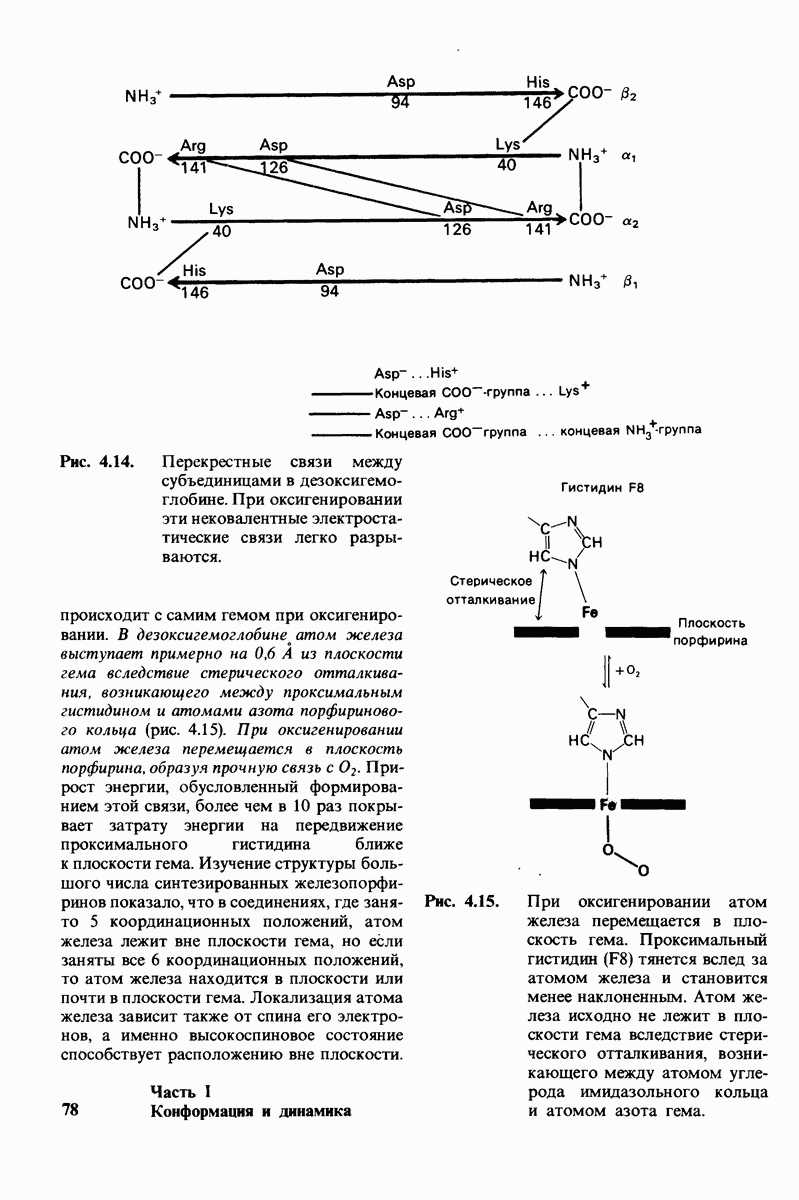
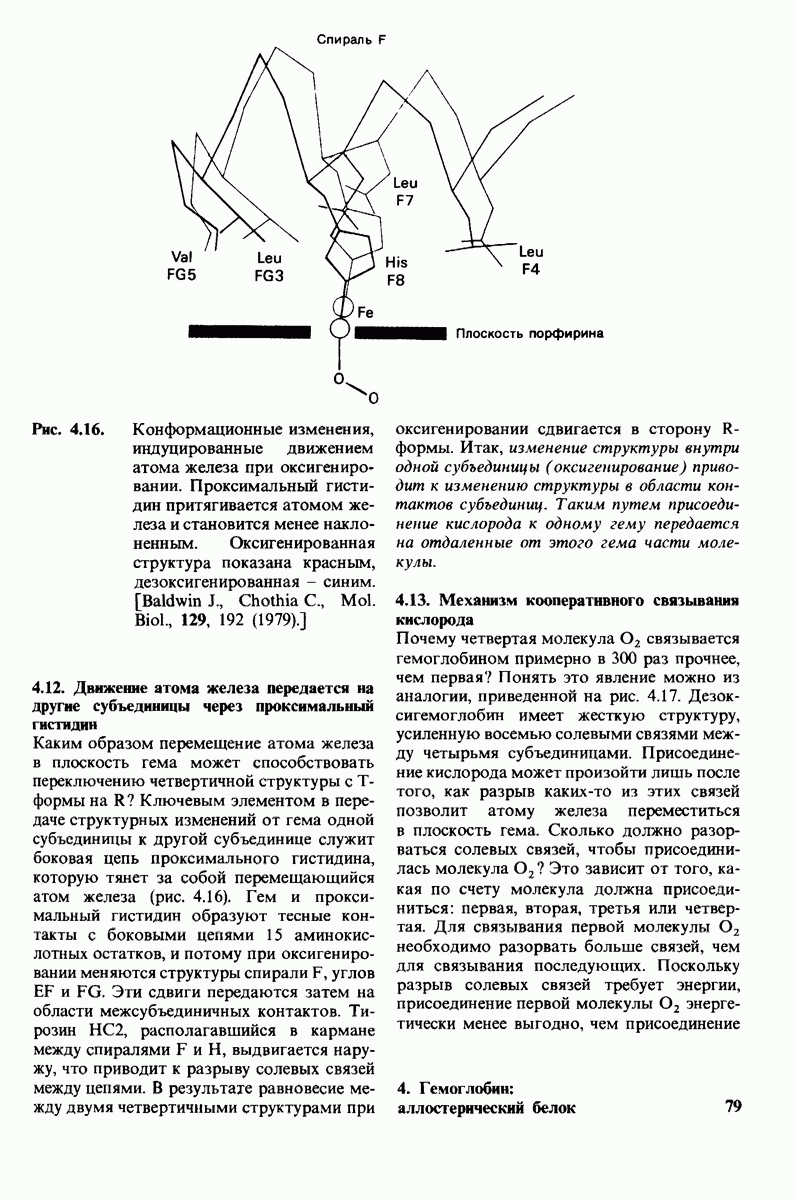
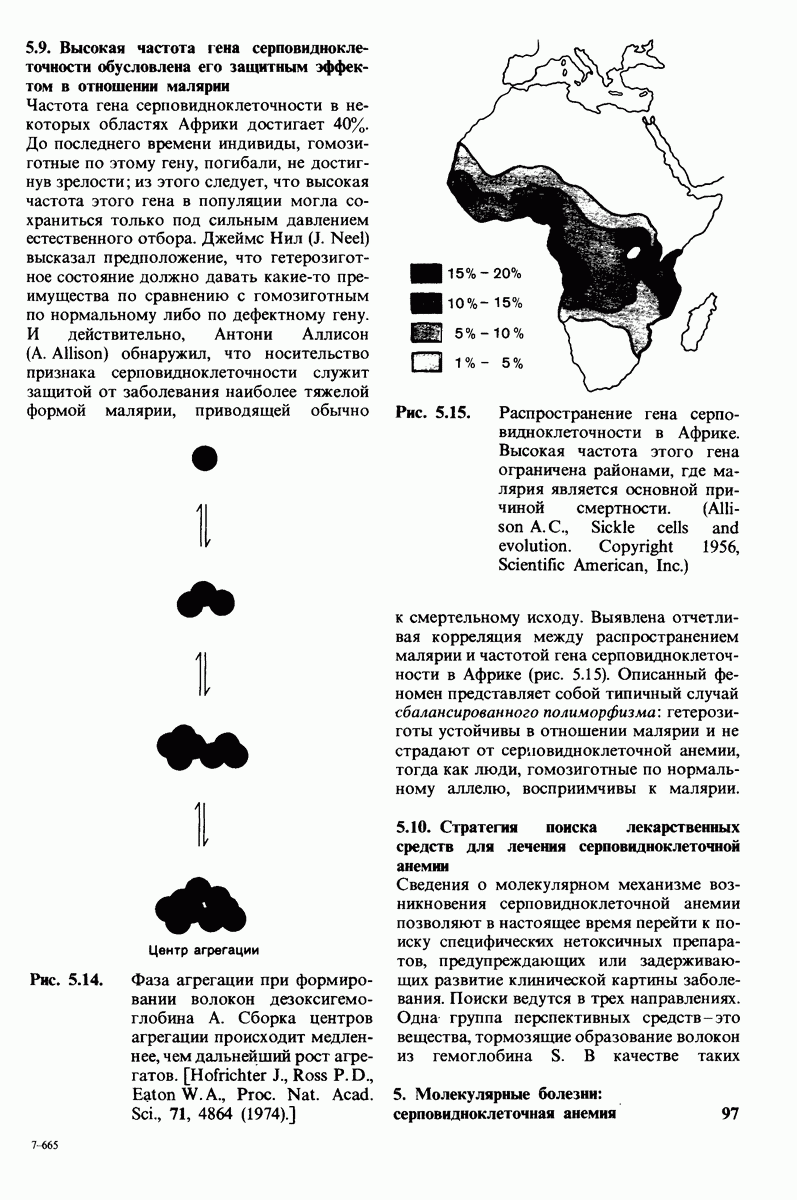
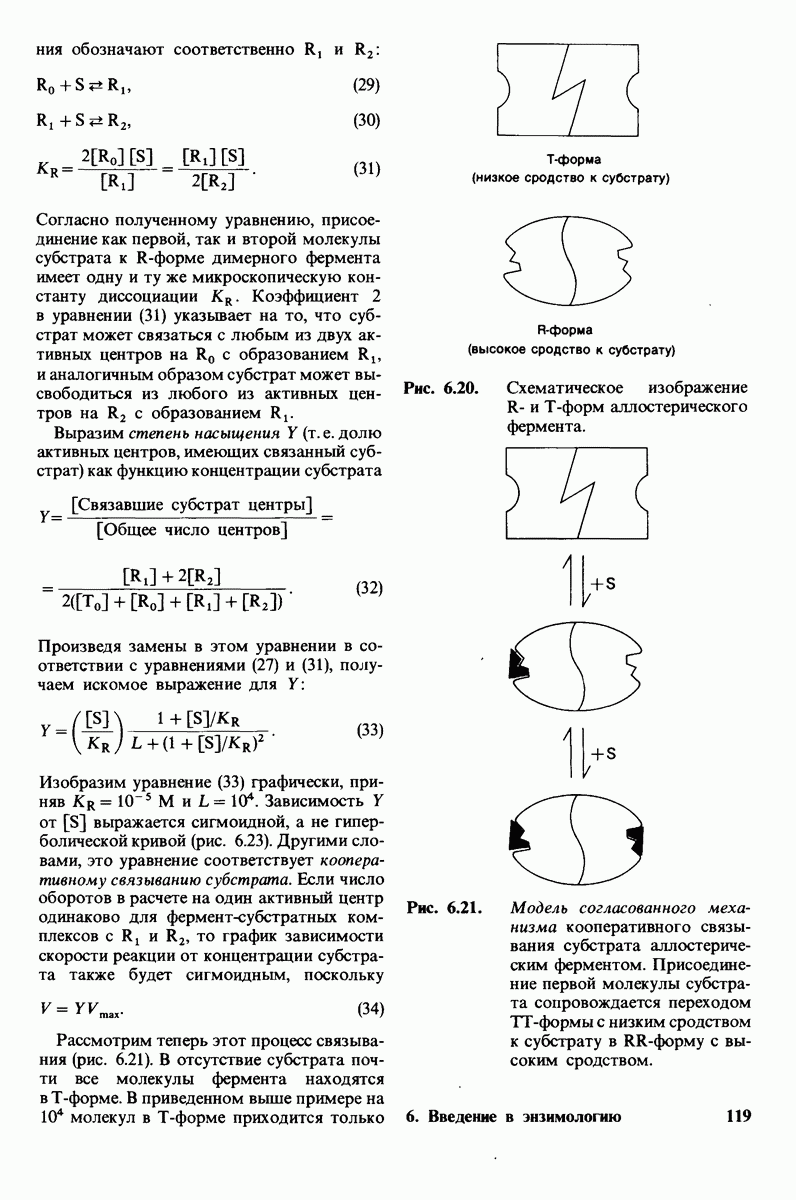
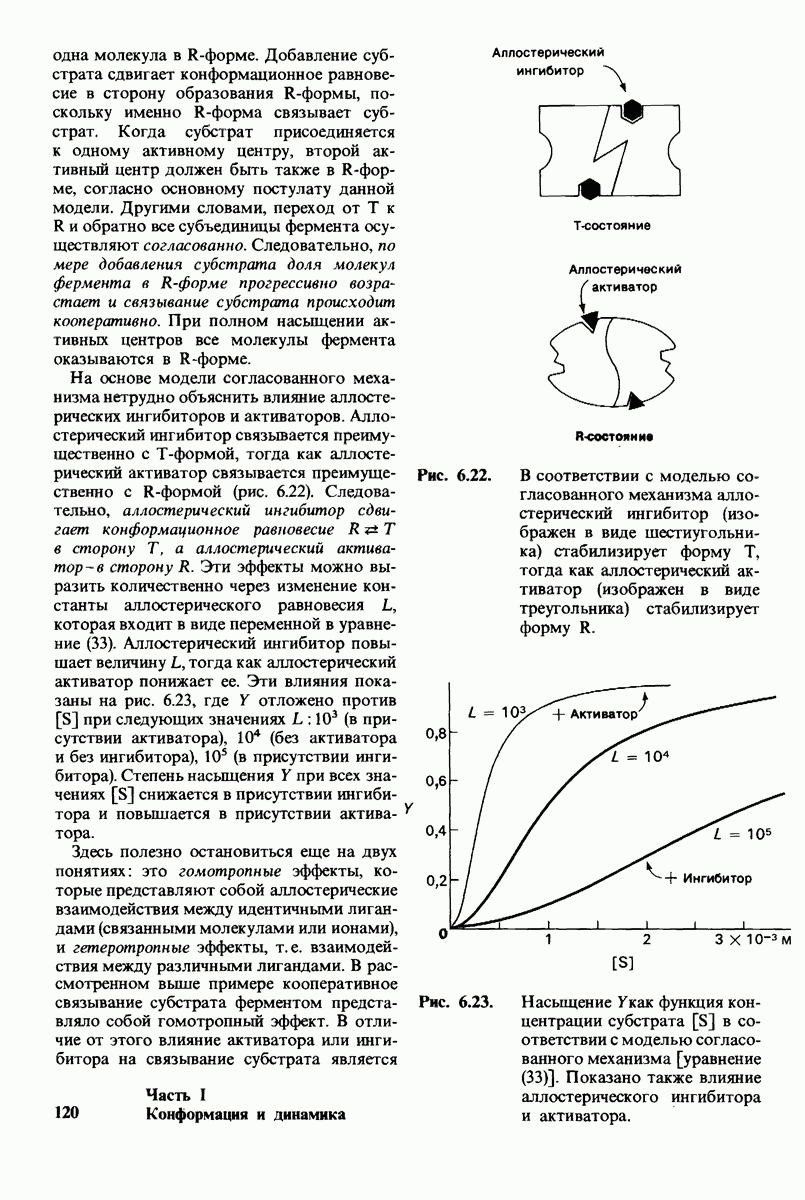
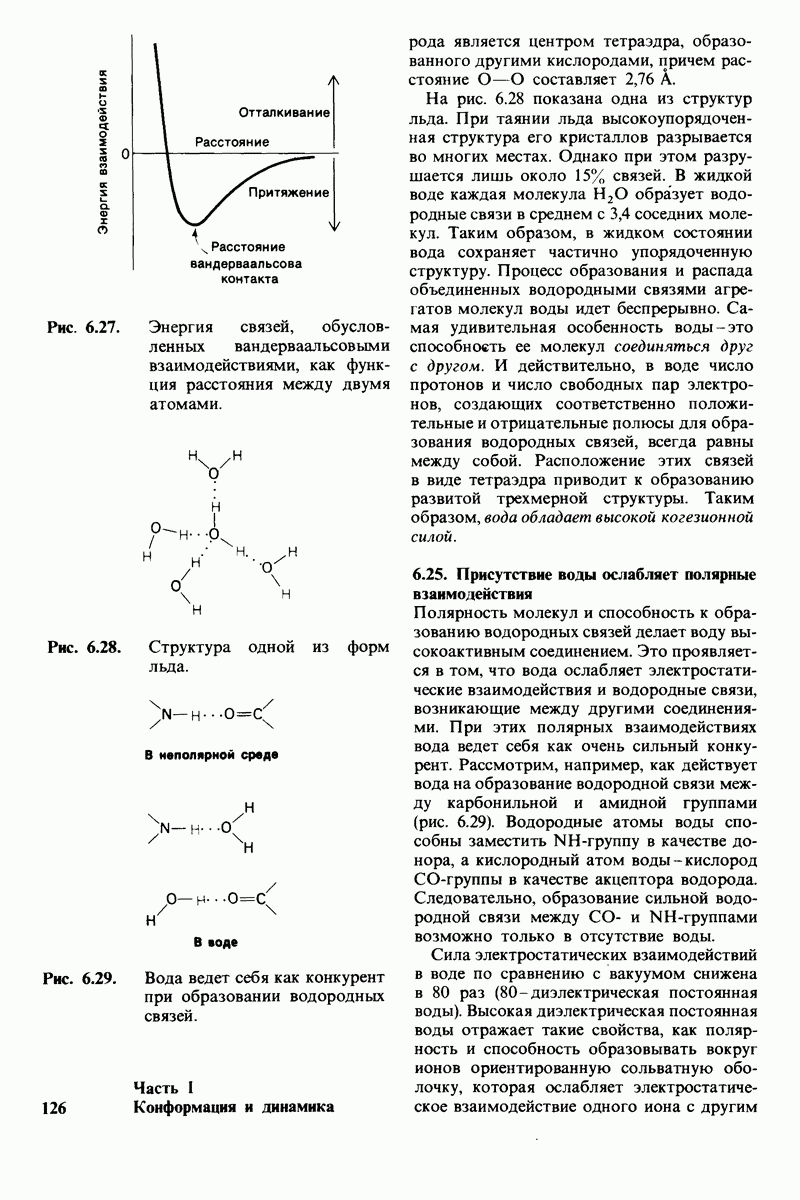
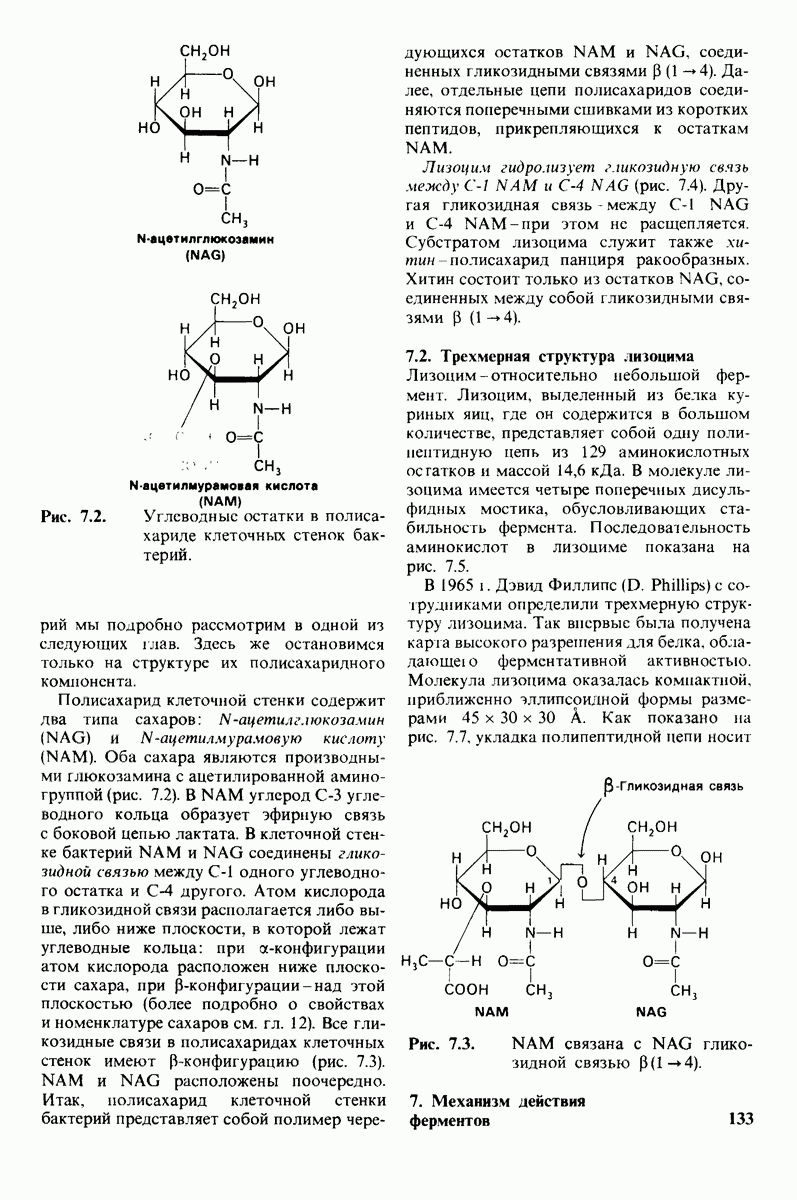
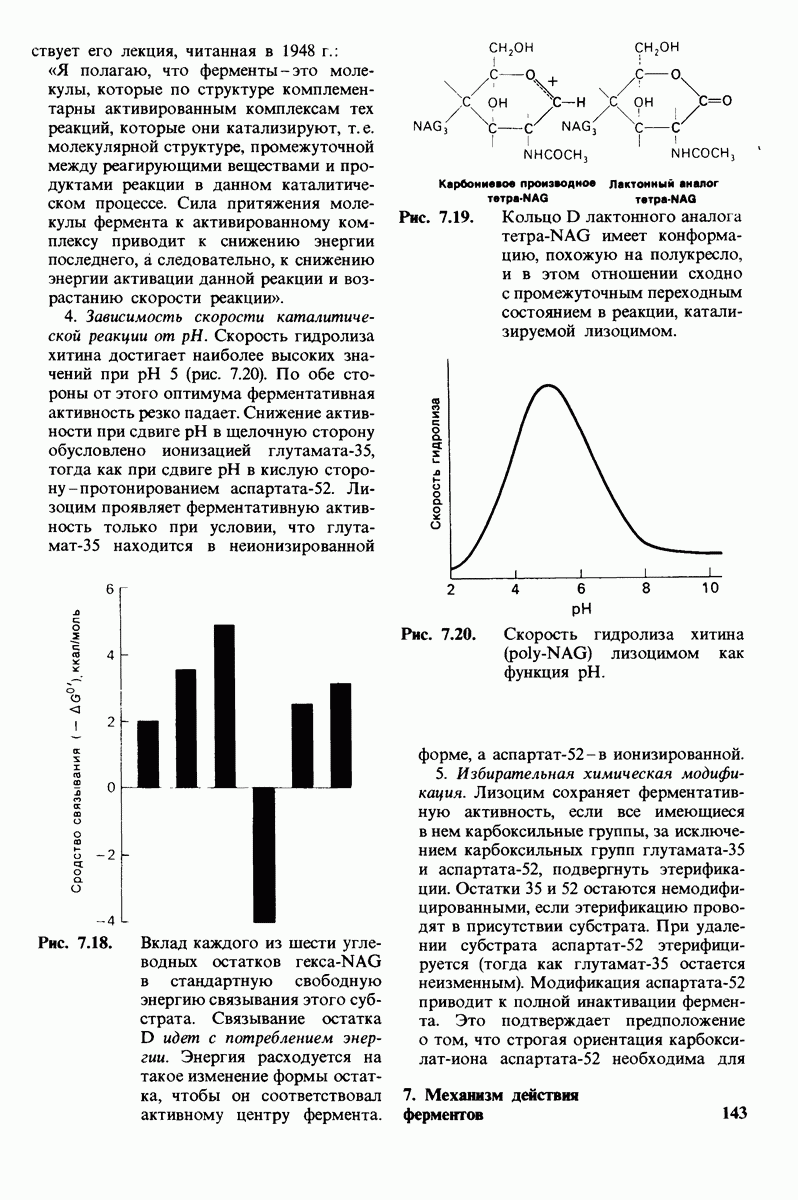
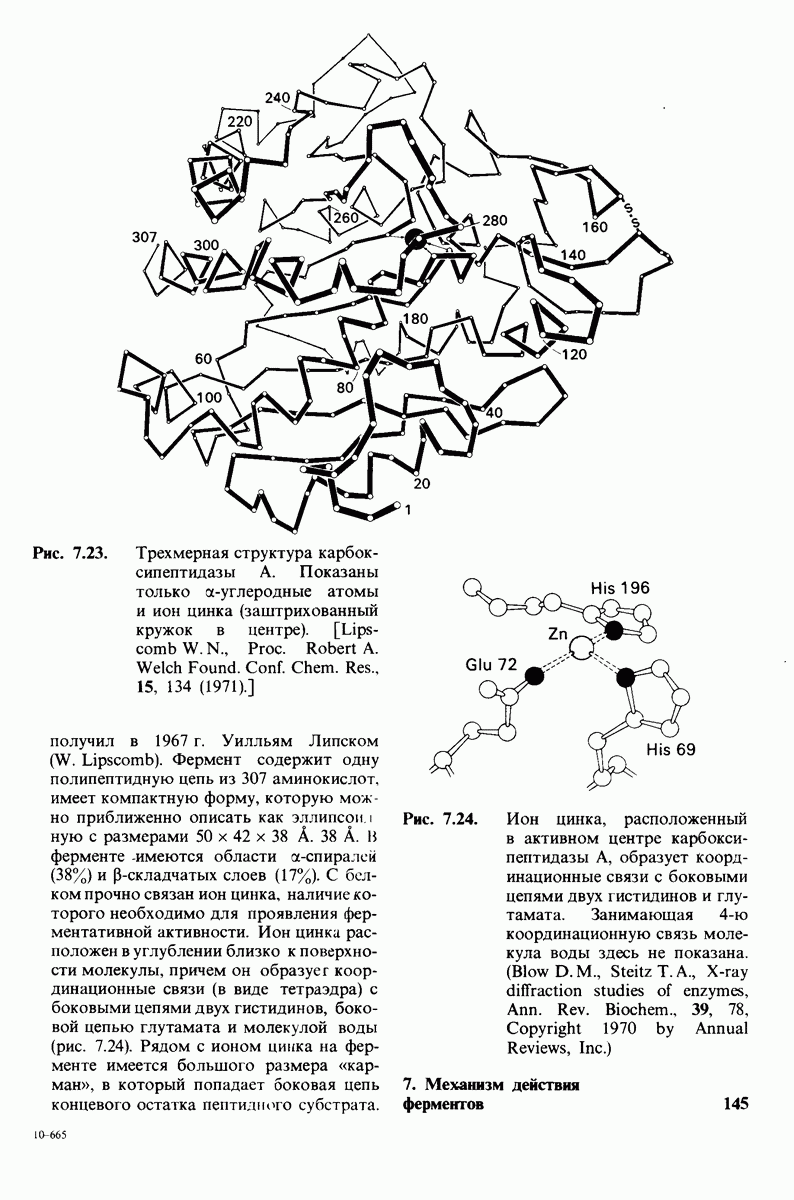
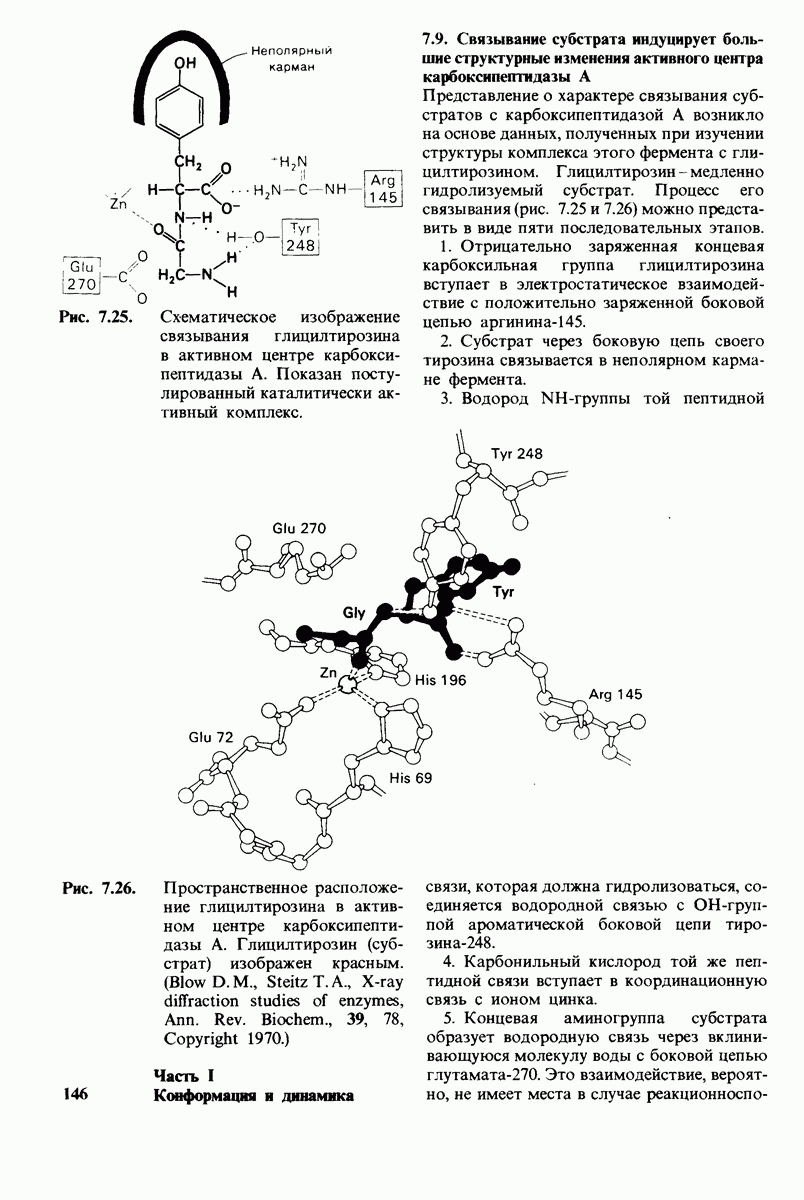
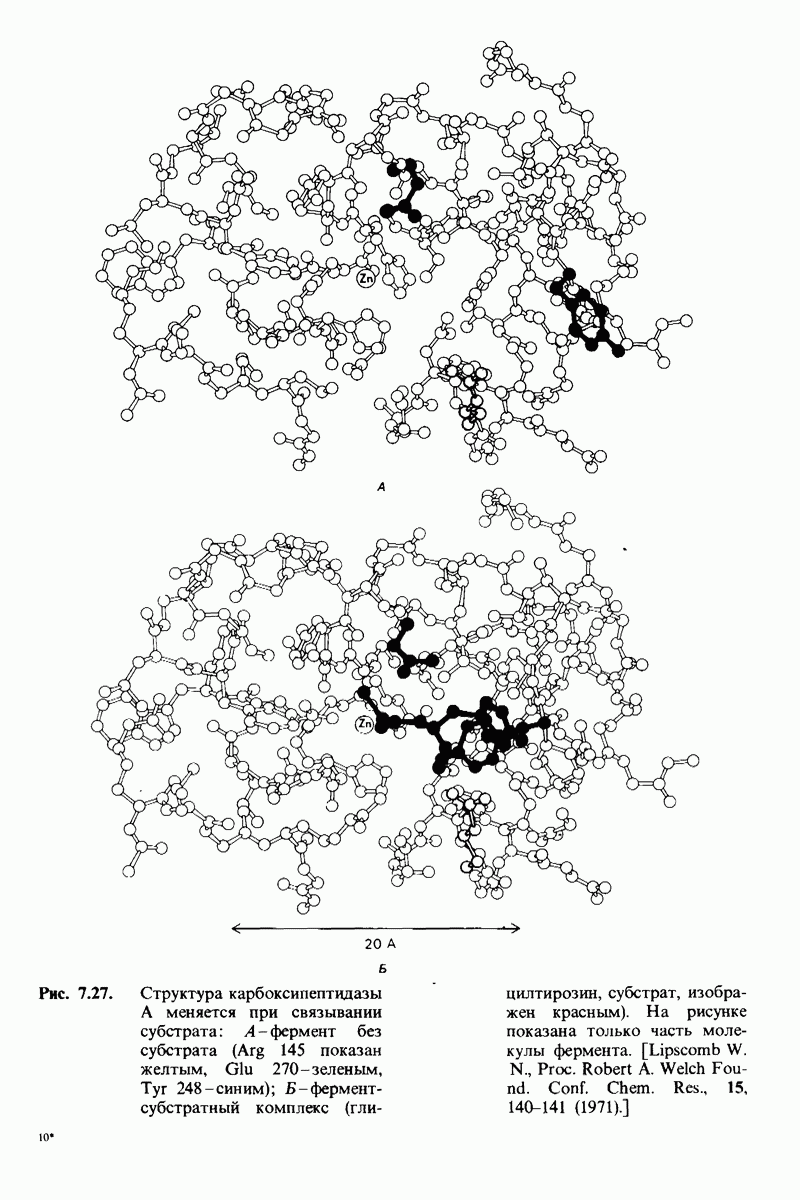
8.28. Контроль свертывания крови: проблема, требующая внимания
Вполне очевидно, что свертывание крови должно регулироваться очень точно. Грань между геморрагией и тенденцией к тромбообразованию едва уловима. Образование кровяного сгустка должно происходить очень быстро, но при этом лишь в пределах поврежденного участка сосуда. Почему же этот процесс включает столько этапов? Почему существуют внутренний и внешний механизмы и как они связаны между собой? Какие механизмы ограничивают процесс тромбообразования участком повреждения? Эти вопросы интересны не только сами по себе, но и в связи с практическими задачами предупреждения и лечения сердечнососудистых заболеваний.
Существенную роль в регуляции тромбообразования играет лабильность активированных факторов свертывания. Их продолжительность жизни очень короткая, поскольку они быстро разводятся в кровотоке, удаляются из кровеносного русла печенью, расщепляются протеиназами, инактивируются специфическими ингибиторами. Наиболее важную роль среди специфических ингибиторов играет антитромбин III, белок плазмы, инактивирующий тромбин путем образования с ним стабильного комплекса. По-видимому, этот комплекс сходен со стабильным комплексом ацилфермент, образованным трипсином и его ингибитором из поджелудочной железы (разд. 8.12). Антитромбин III ингибирует также факторы свертывания  .
.
Таблица 8.2 (см. скан) Факторы свертывания крови

Рис. 8.38. Внешний механизм свертывания крови.
Ингибиторное действие антитромбина III усиливается в присутствии гепарина - отрицательно заряженного полисахарида (разд. 9.16), синтезируемого тучными клетками, расположенными вблизи стенок кровеносных сосудов. Гепарин действует как антикоагулянт, увеличивая скорость необратимого комплексообразования между тромбином и антитромбином III.
Заключение
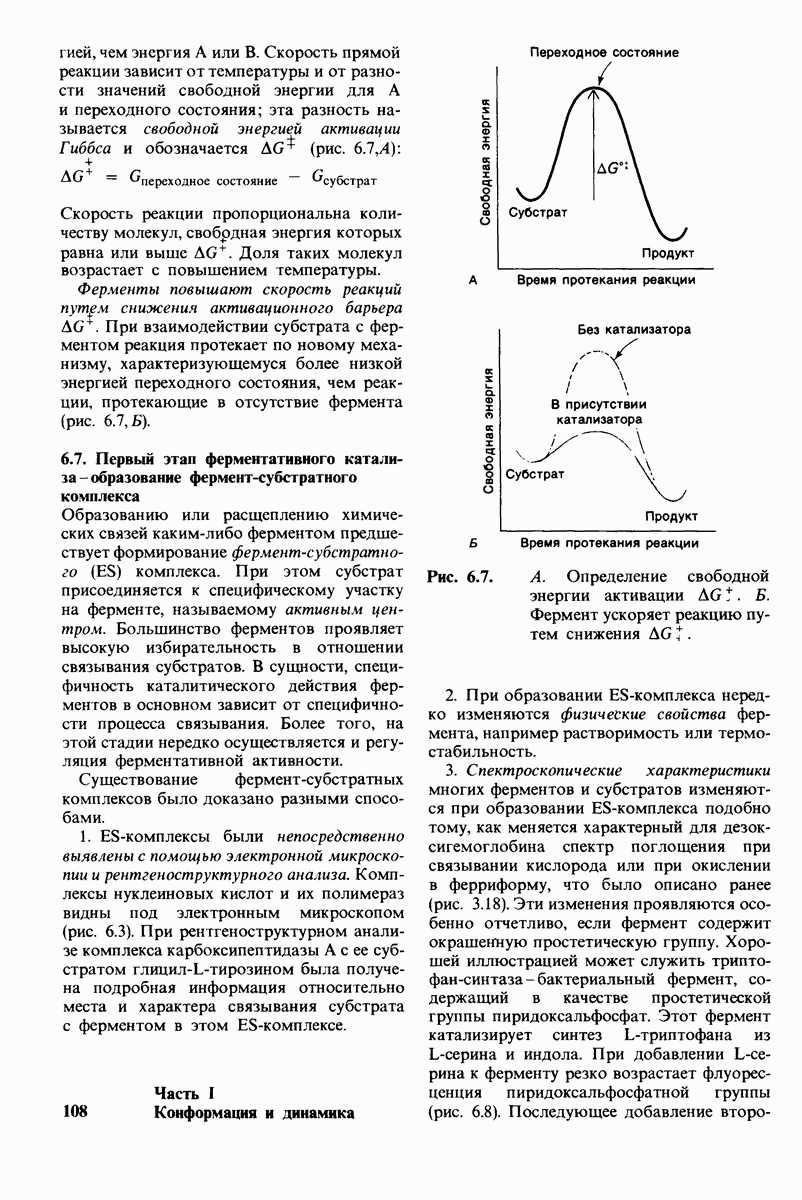
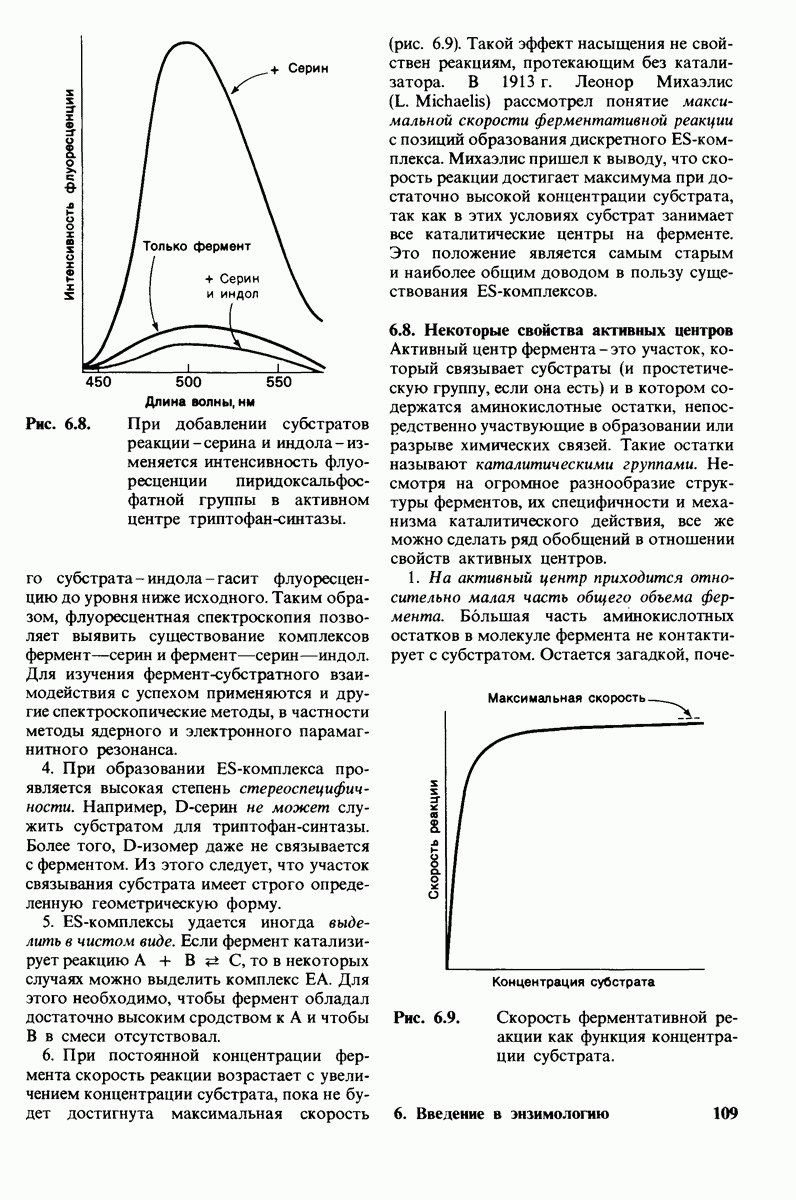
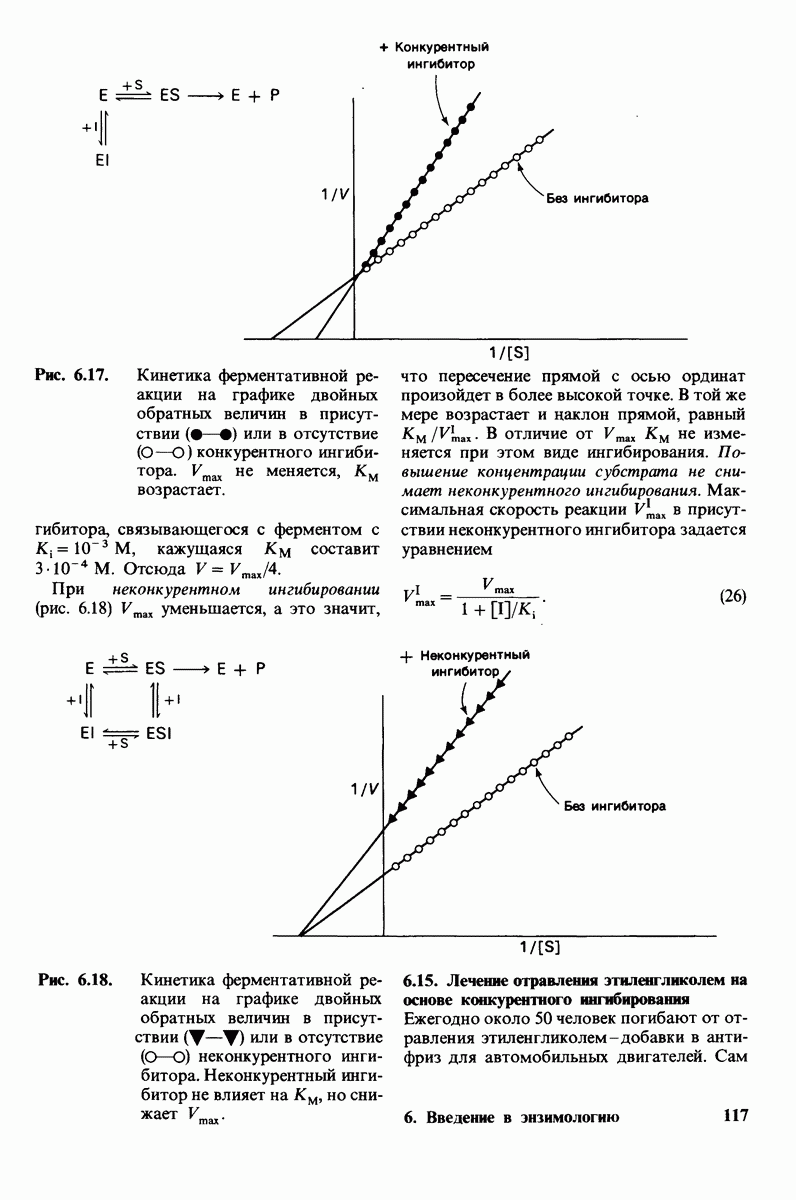
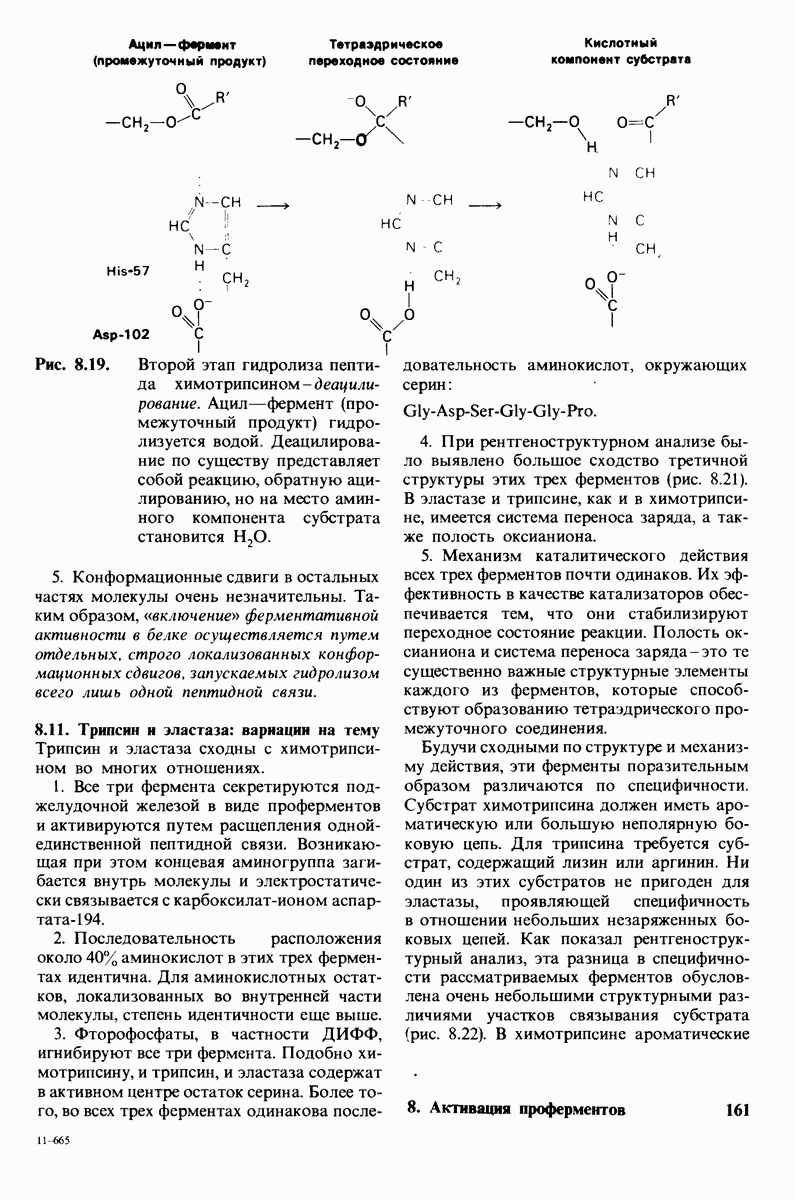
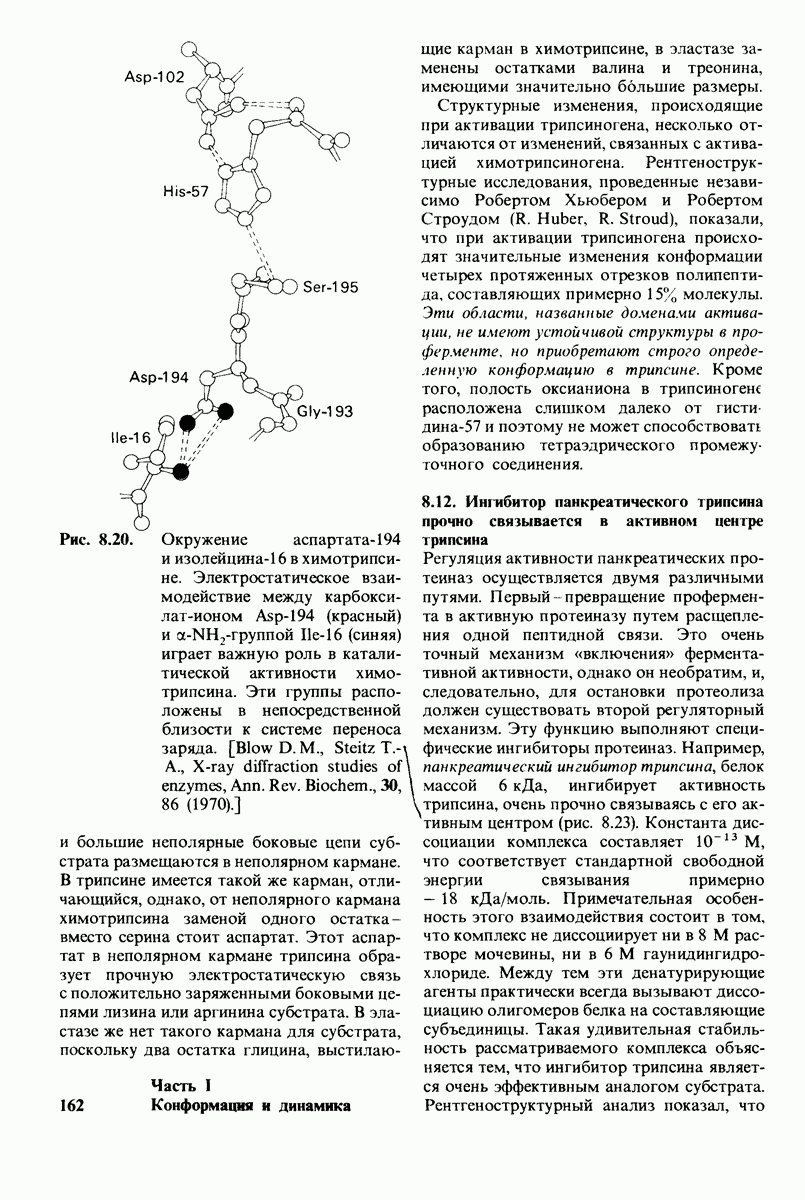
Активация белков в результате протеолитического расщепления одной или нескольких пептидных связей - широко распространенный механизм регуляции в биологических системах. Если белок в активированной форме является ферментом, то его неактивный предшественник называется проферментом (или зимогеном). Наиболее изученный процесс активации профермента - это превращение химотрипсиногена в химотрипсин, происходящее следующим путем. Под действием трипсина пептидная связь между остатками 15 и 16 в молекуле химотрипсиногена разрывается и в результате образуется химотрипсин. Возникшая при этом новая концевая аминогруппа изолейцина-16 загибается внутрь молекулы белка, где электростатически связывается с аспартатом-194. Это взаимодействие служит пусковым механизмом для ряда локальных изменений конформации, конечный результат которых - формирование кармана для связывания ароматических (либо больших неполярных) боковых цепей субстрата. Кроме того, формируется полость оксианиона, стабилизирующая переходное состояние.
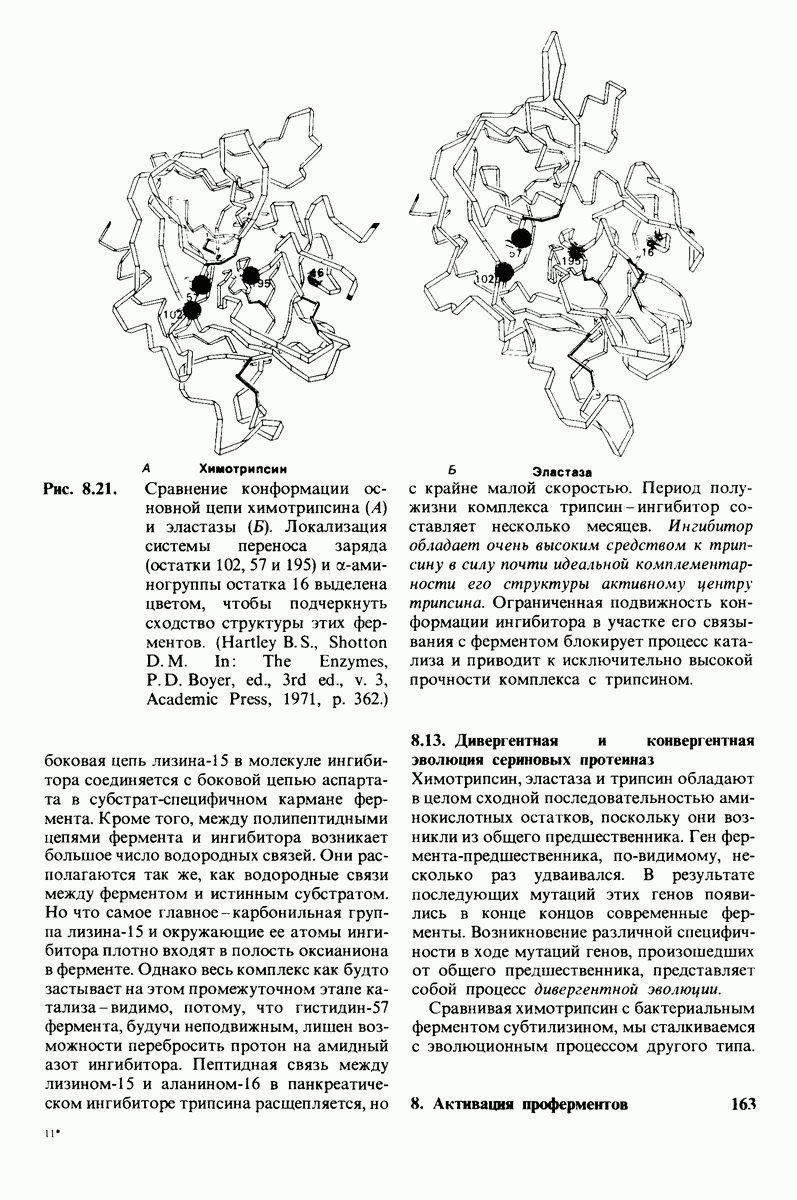
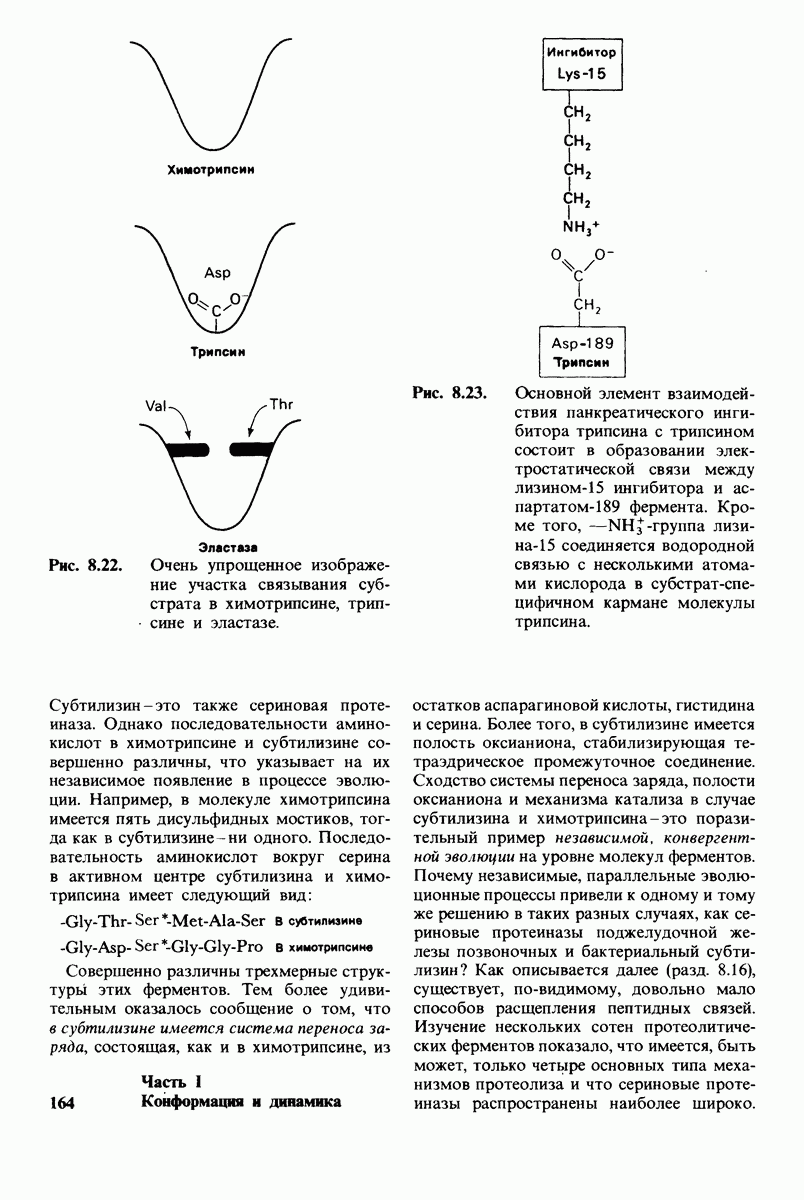
Критическая роль в механизме каталитического действия химотрипсина принадлежит остатку серина (серин-195) с высокой реакционной способностью. На первом этапе гидролиза пептидного субстрата происходит этерификация карбоксильного компонента субстрата с гидроксильной группой серина-195, приводящая к образованию ковалентно связанного промежуточного продукта ацилфермент. Этот промежуточный продукт гидролизуется на втором этапе катализа. Ключевую роль как в ацилировании, так и в деацилировании фермента играет также гистидин-57. На деле серин-195 становится сильным нуклеофильным агентом в результате формирования ряда водородных связей; он соединяется водородной связью с гистидином-57, который в свою очередь соединяется водородной связью с аспартатом-102, расположенным в глубине молекулы. В итоге эти три остатка составляют систему переноса заряда, ускоряющую катализ примерно в 103 раз. Такая же система переноса заряда имеется в трипсине, эластазе и тромбине. Все эти ферменты подобны химотрипсину по аминокислотной последовательности, конформации и механизму каталитического действия, но отличаются от него по специфичности. Такие вариации на одну тему, видимо, результат происхождения от общего предшественника и последующей дивергентной эволюции. Три других основных типа протеолитических ферментов - это цинксодержащие, тиоловые и карбоксипротеиназы.
Активация проферментов играет также ведущую роль в регуляции свертывания крови. Поразительная особенность процесса свертывания состоит в том, что он организован как каскад превращений проферментов, в котором активированная форма одного фактора свертывания катализирует активацию следующего. Свертывание крови происходит в результате взаимодействия двух последовательностей реакций, получивших название внешнего и внутреннего механизмов. Оба механизма необходимы для нормального свертывания крови. Они сливаются в общий механизм, приводящий к формированию фибринового сгустка. Фибрин образуется из фибриногена, высокорастворимого белка плазмы, путем гидролиза четырех пептидных связей между остатками аргинина и глицина. Катализирует эту реакцию тромбин - фермент, сходный с трипсином. В результате гидролиза от фибриногена отщепляются два  -пептида и два
-пептида и два  -пептида, на долю которых приходится около 3% молекулы фибриногена. Образующийся в результате мономер фибрина спонтанно полимеризуется в длинные
-пептида, на долю которых приходится около 3% молекулы фибриногена. Образующийся в результате мономер фибрина спонтанно полимеризуется в длинные
нерастворимые нити фибрина. Сгусток фибрина стабилизируется далее ковалентными поперечными связями, образующимися в результате реакции трансамидирования между боковыми цепями специфических остатков глутамина и лизина. Для карбоксилирования остатков глутамата в протромбине и некоторых других факторах свертывания необходим витамин К. Связывание ионов  -карбоксиглутаматом способствует прикреплению протромбина к мембранам тромбоцитов, что ведет к ускорению активации протромбина факторами
-карбоксиглутаматом способствует прикреплению протромбина к мембранам тромбоцитов, что ведет к ускорению активации протромбина факторами 
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
(см. скан)
Вопросы и задачи
(см. скан)