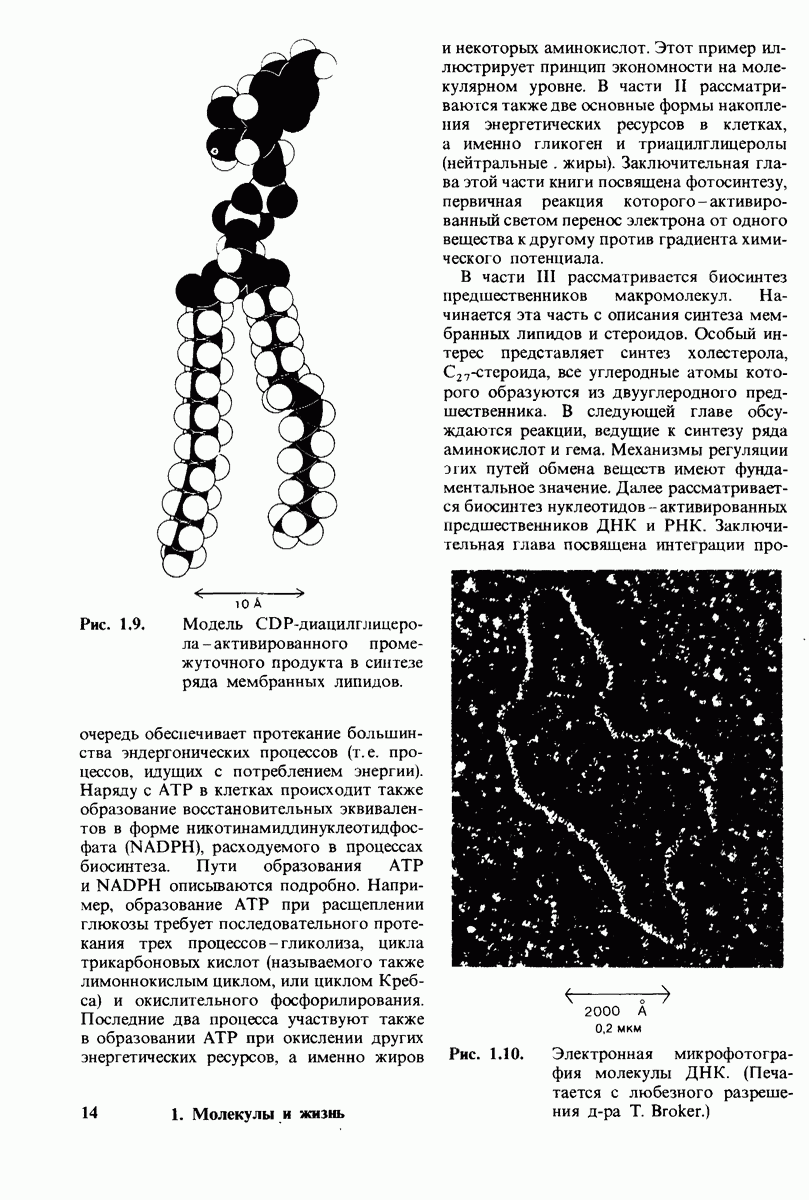
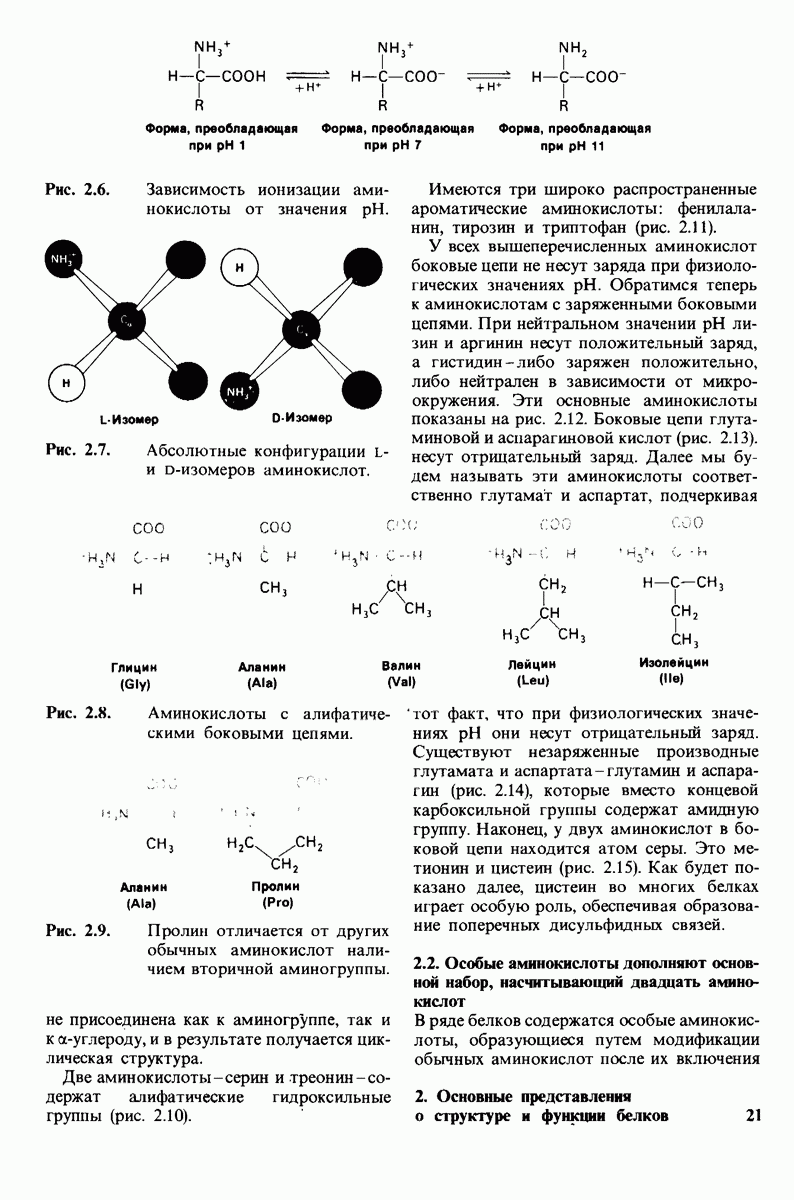
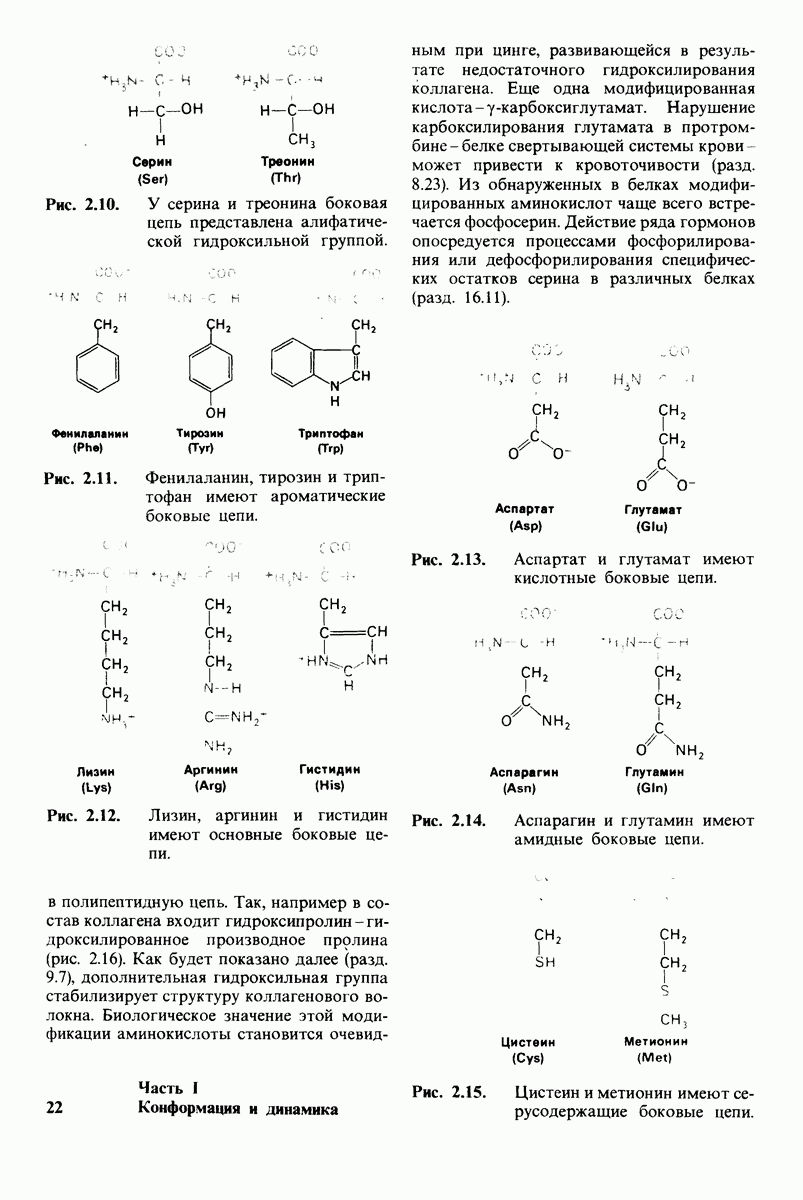
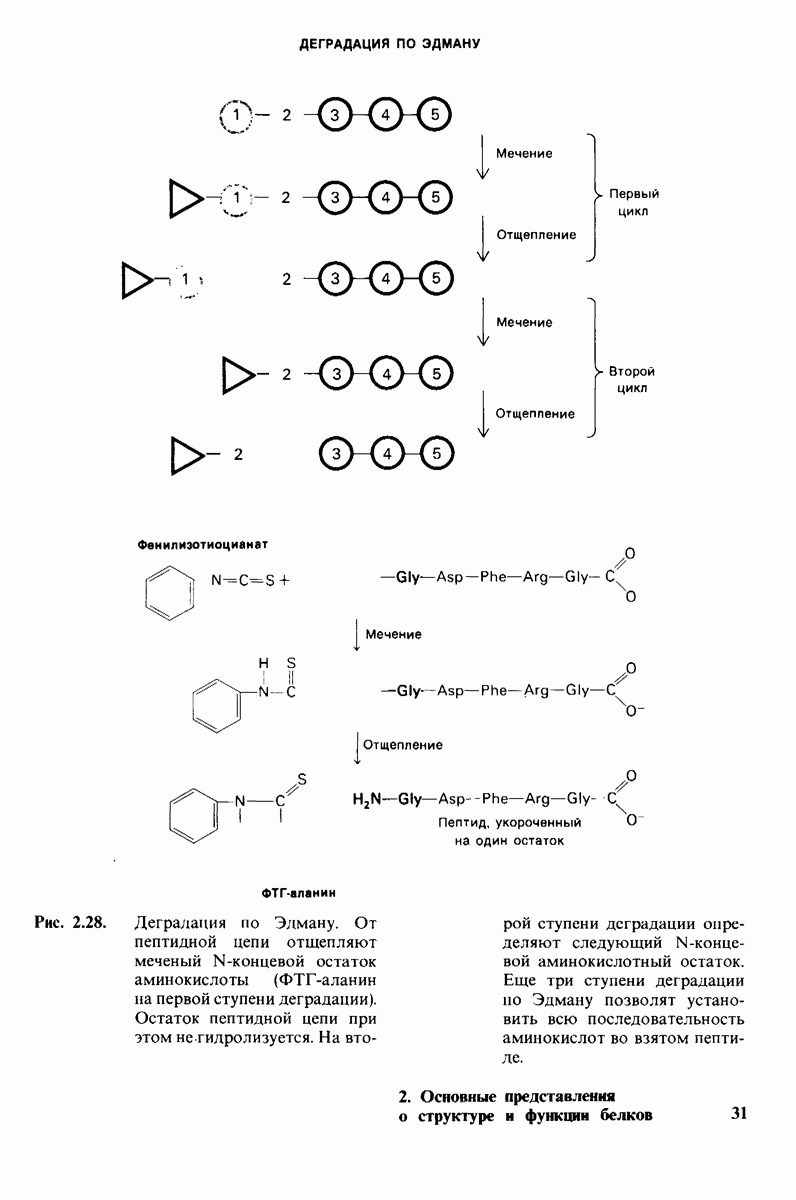
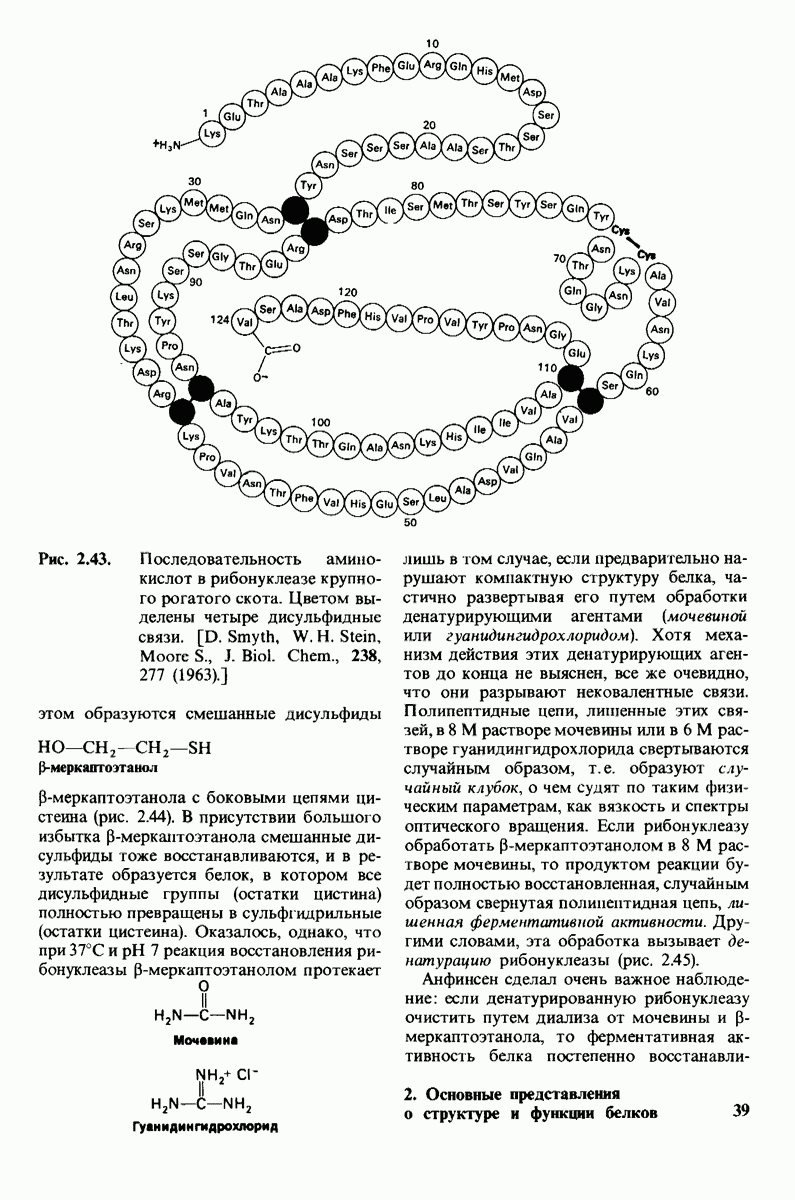
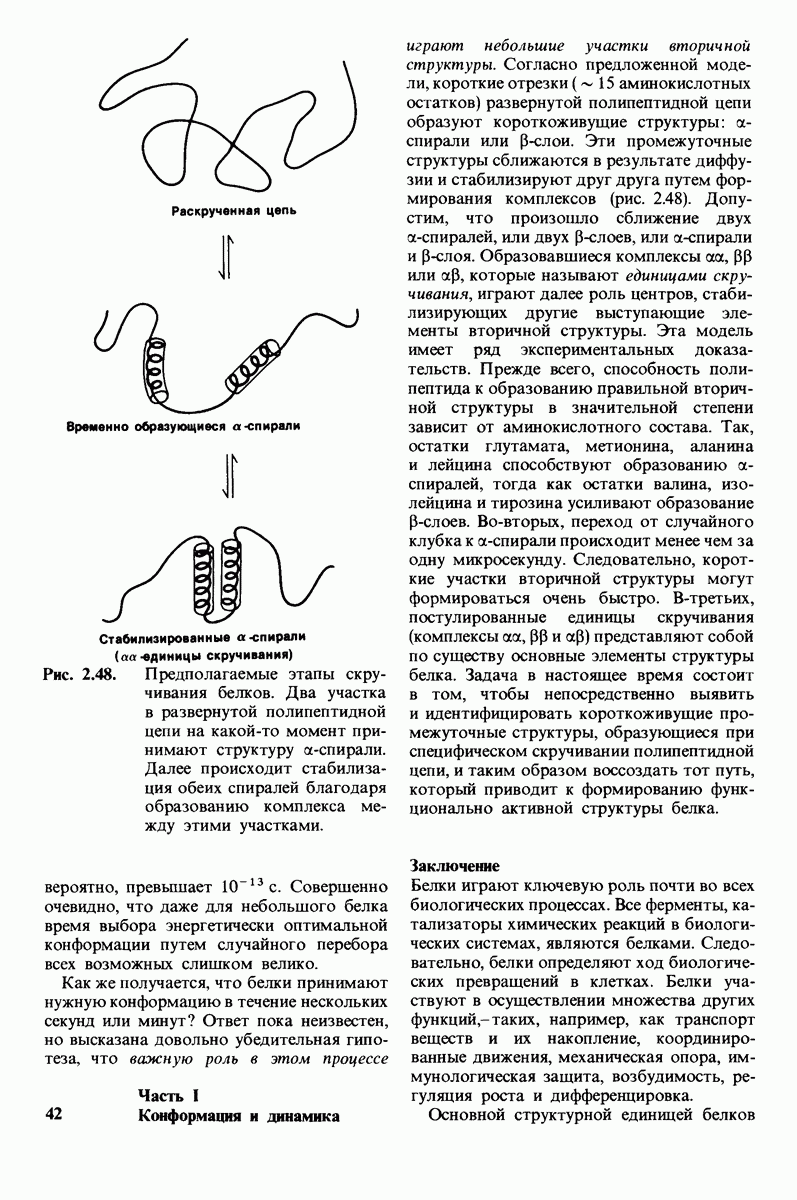
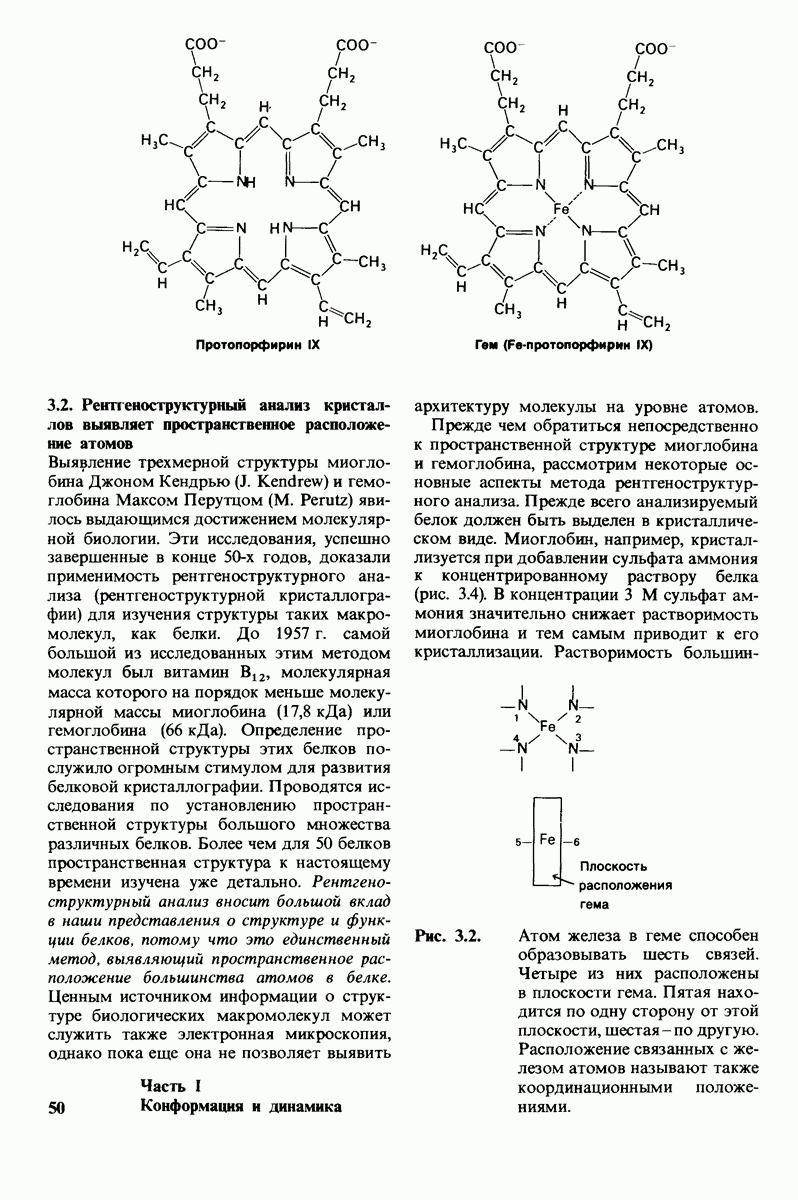
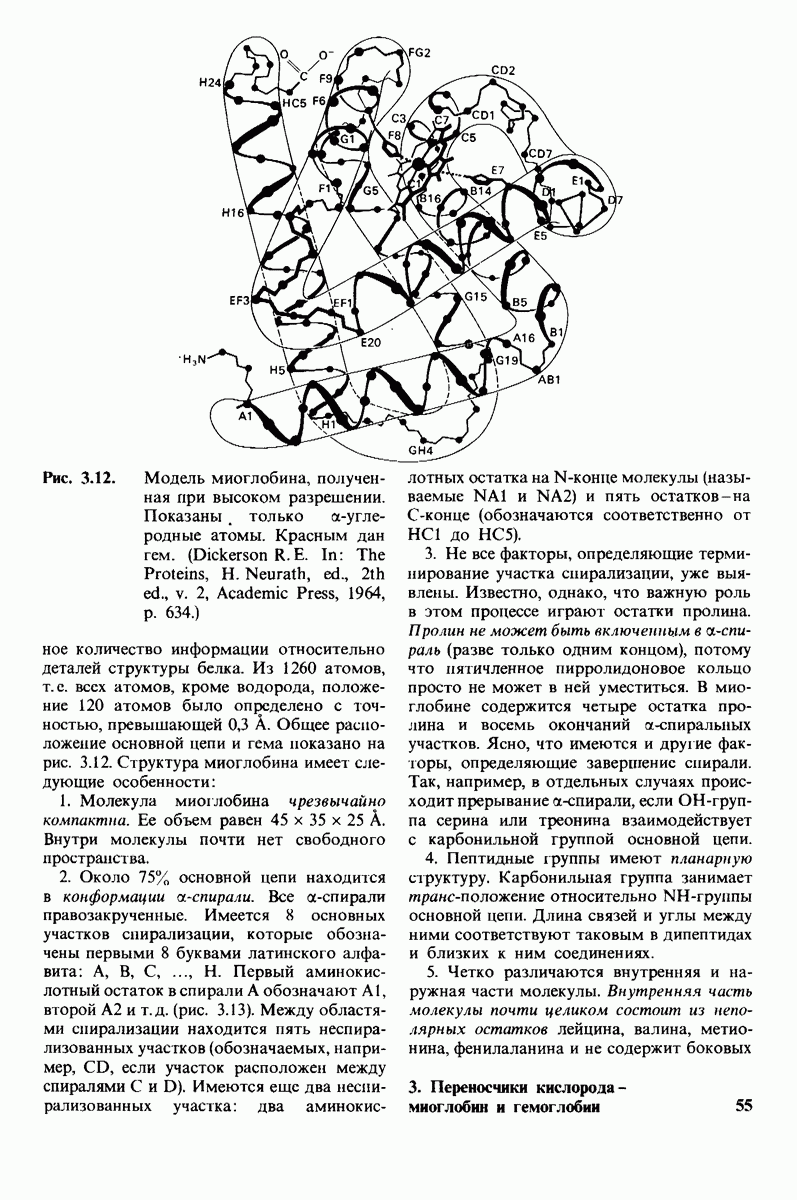
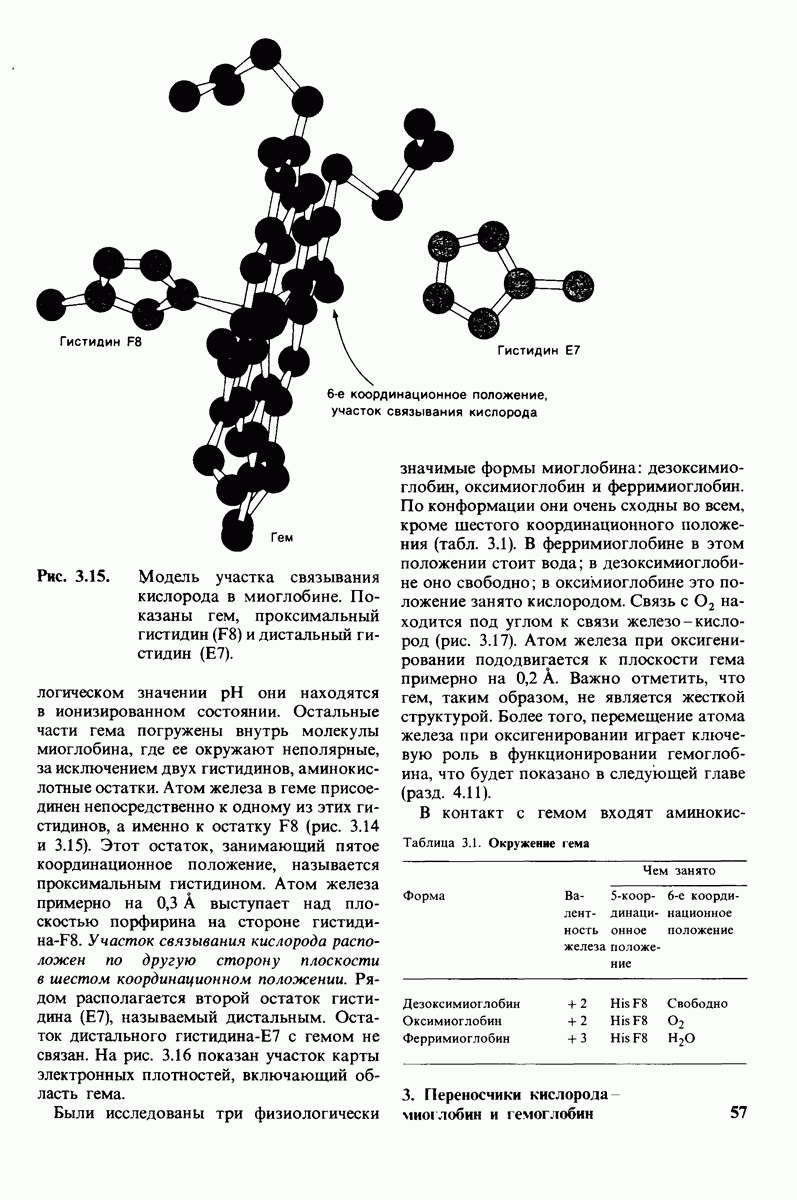
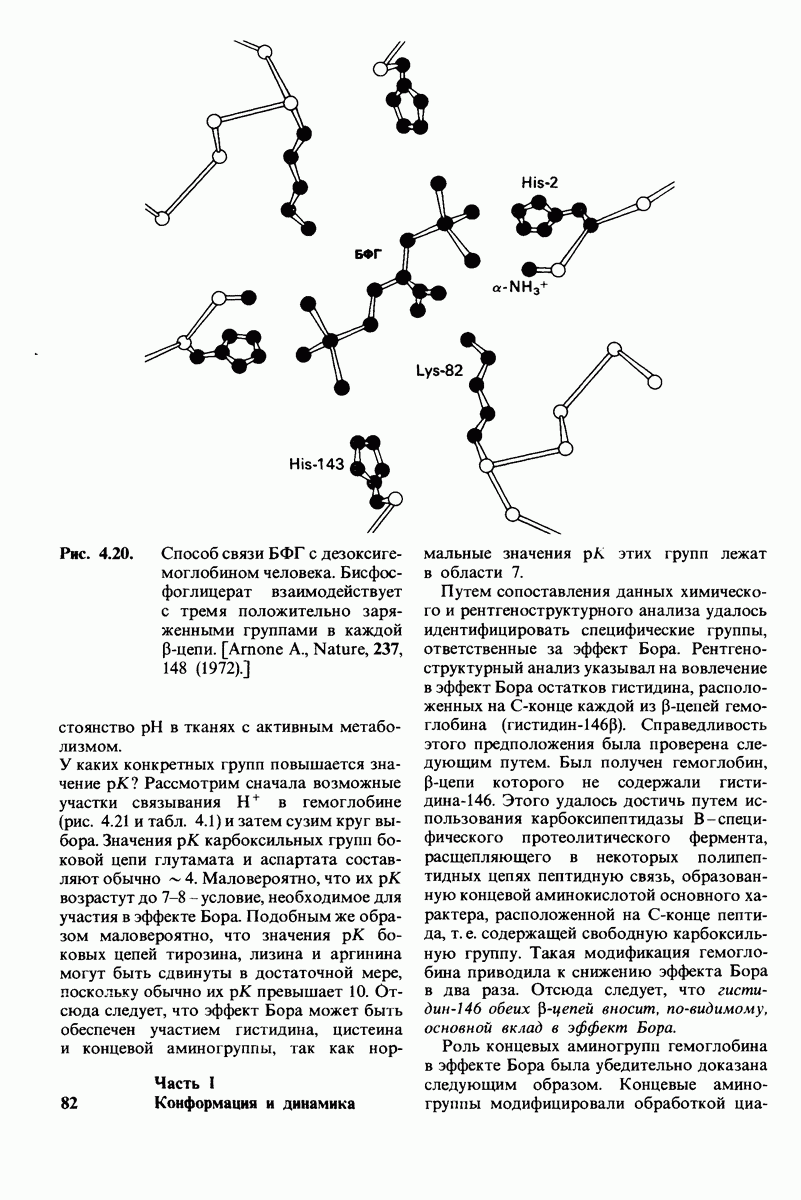
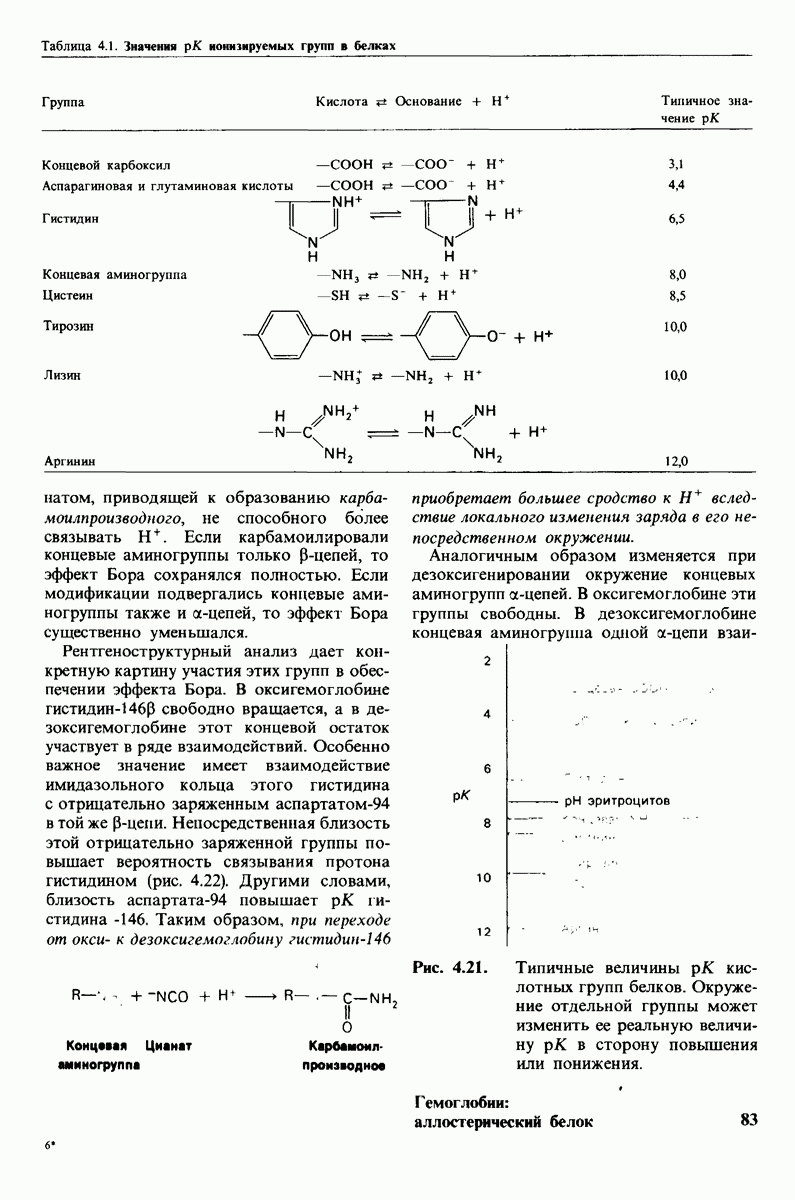
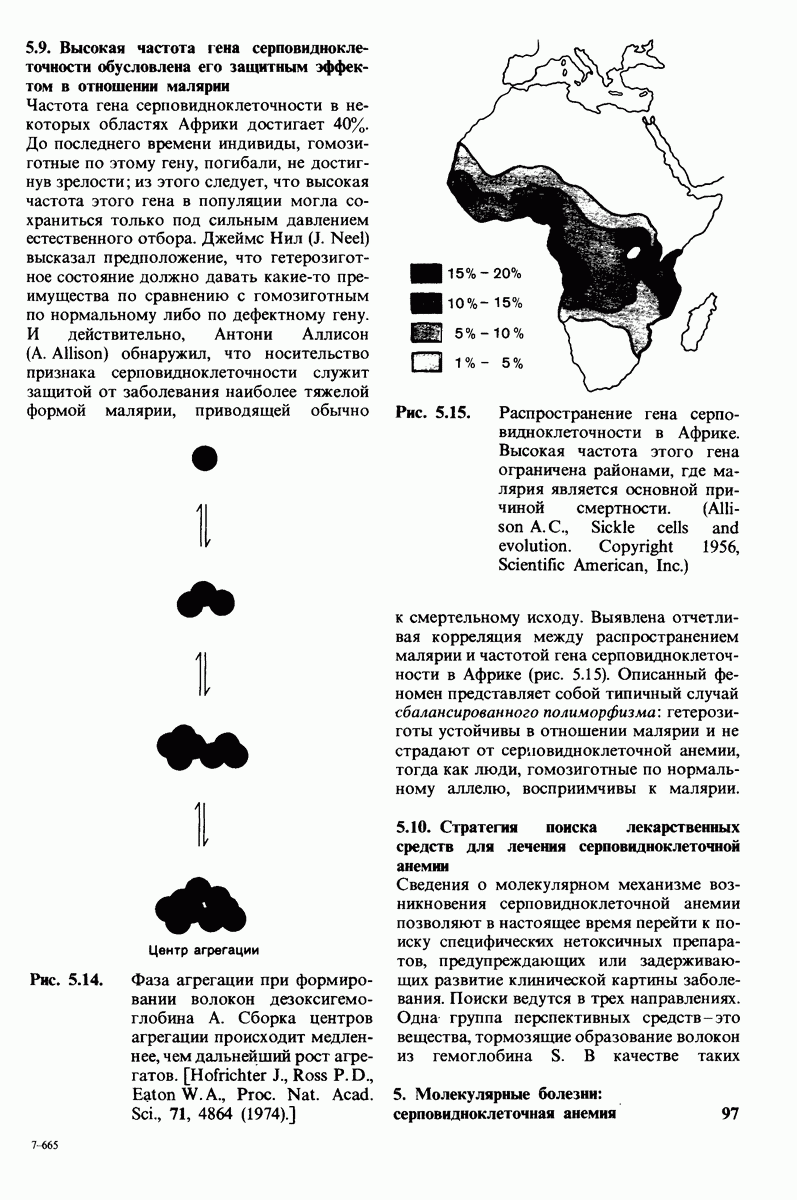
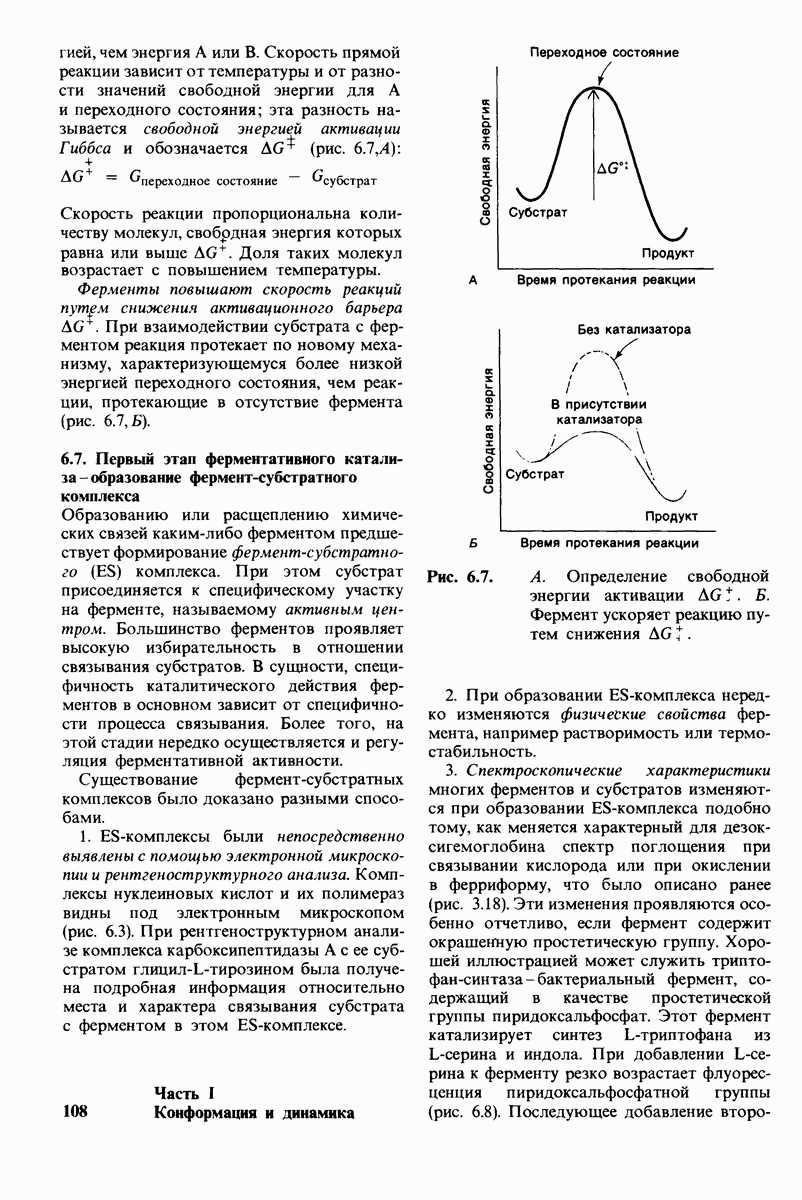
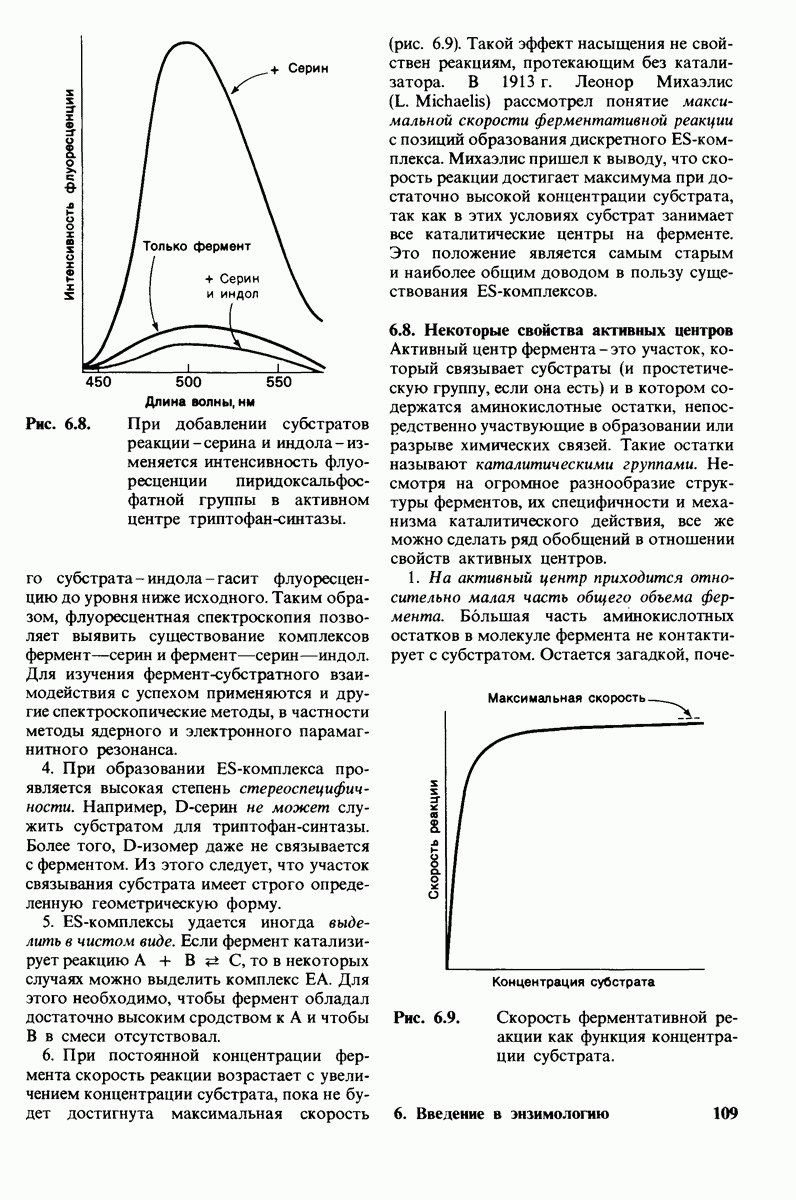
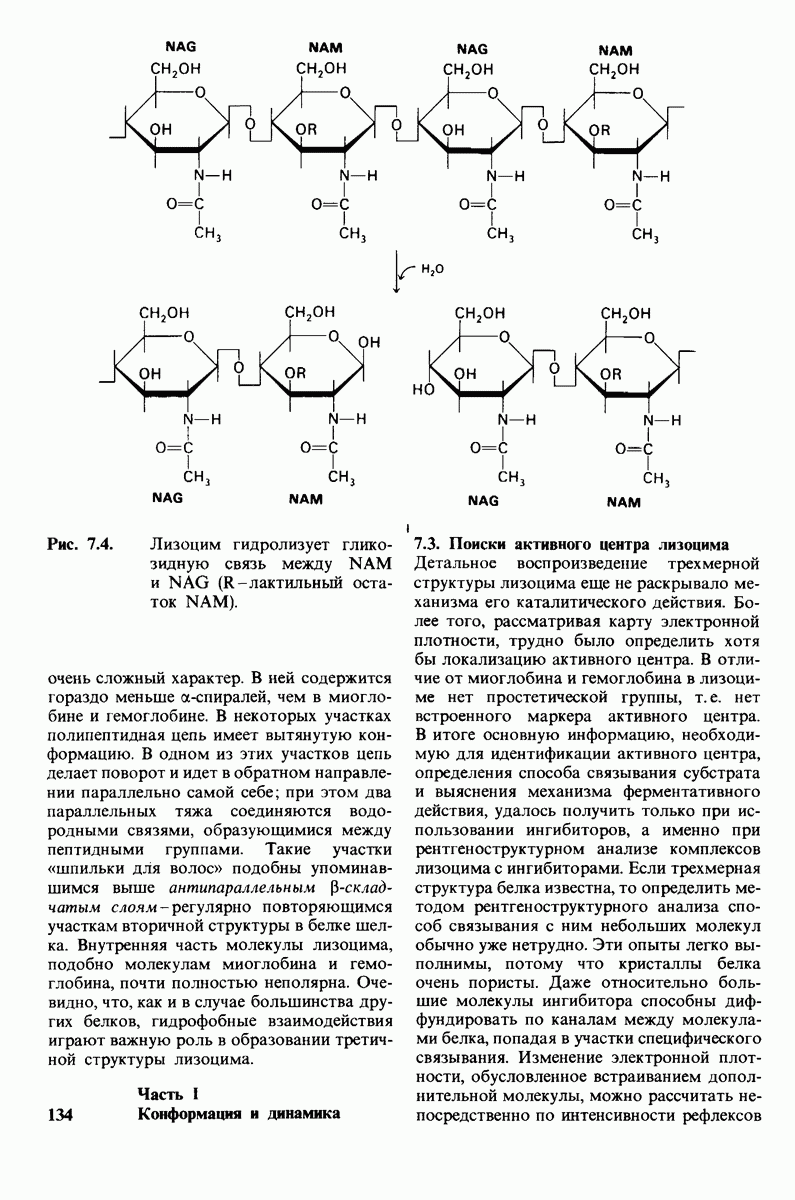
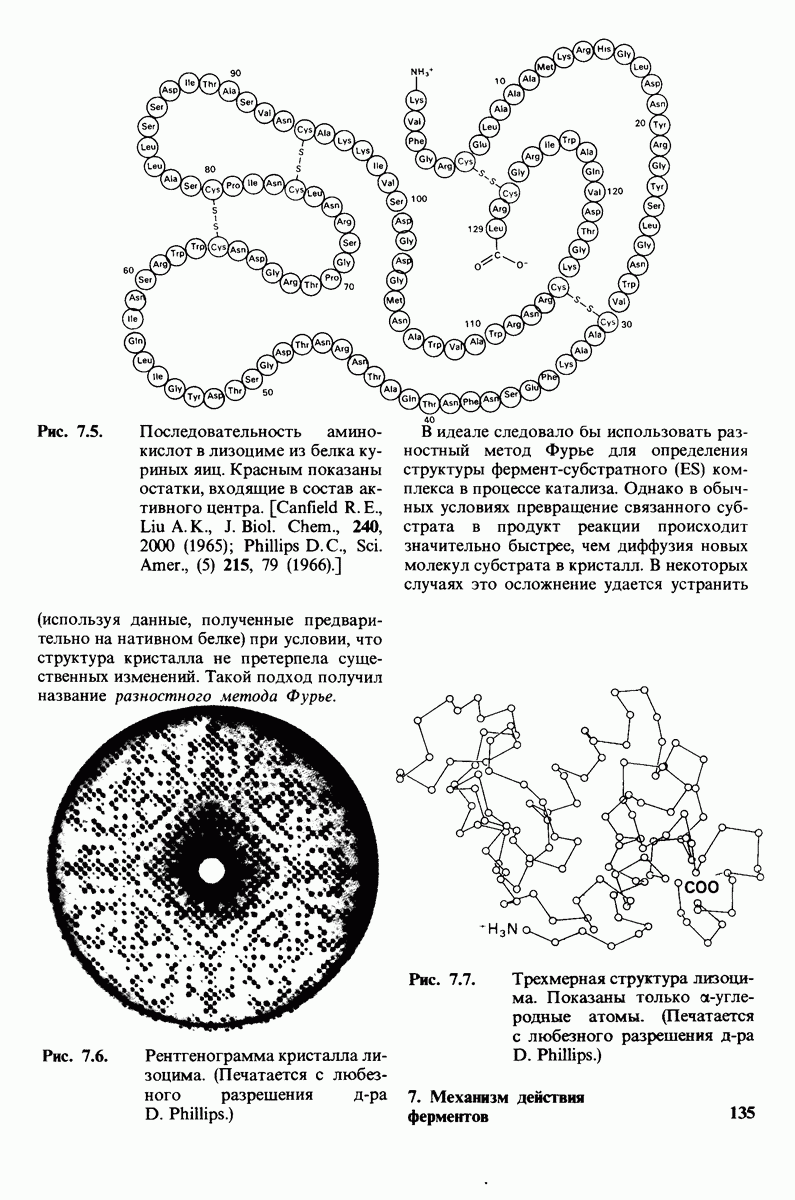
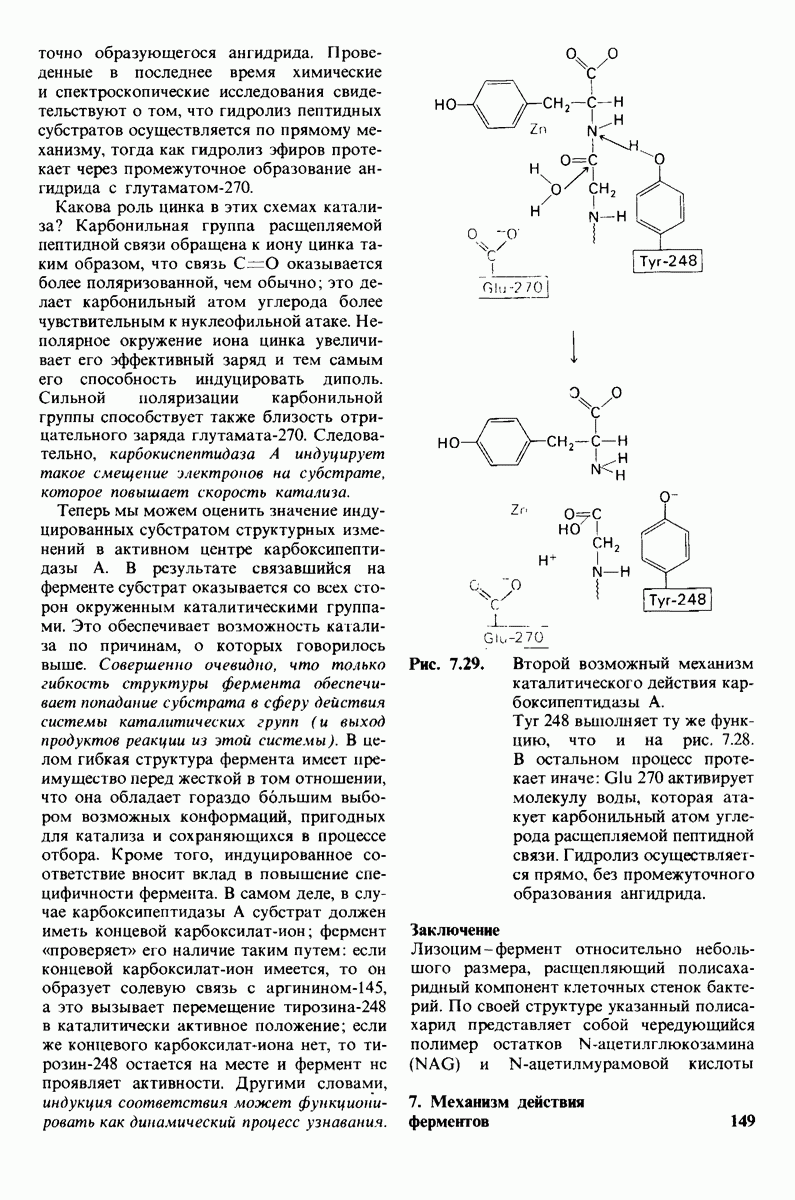
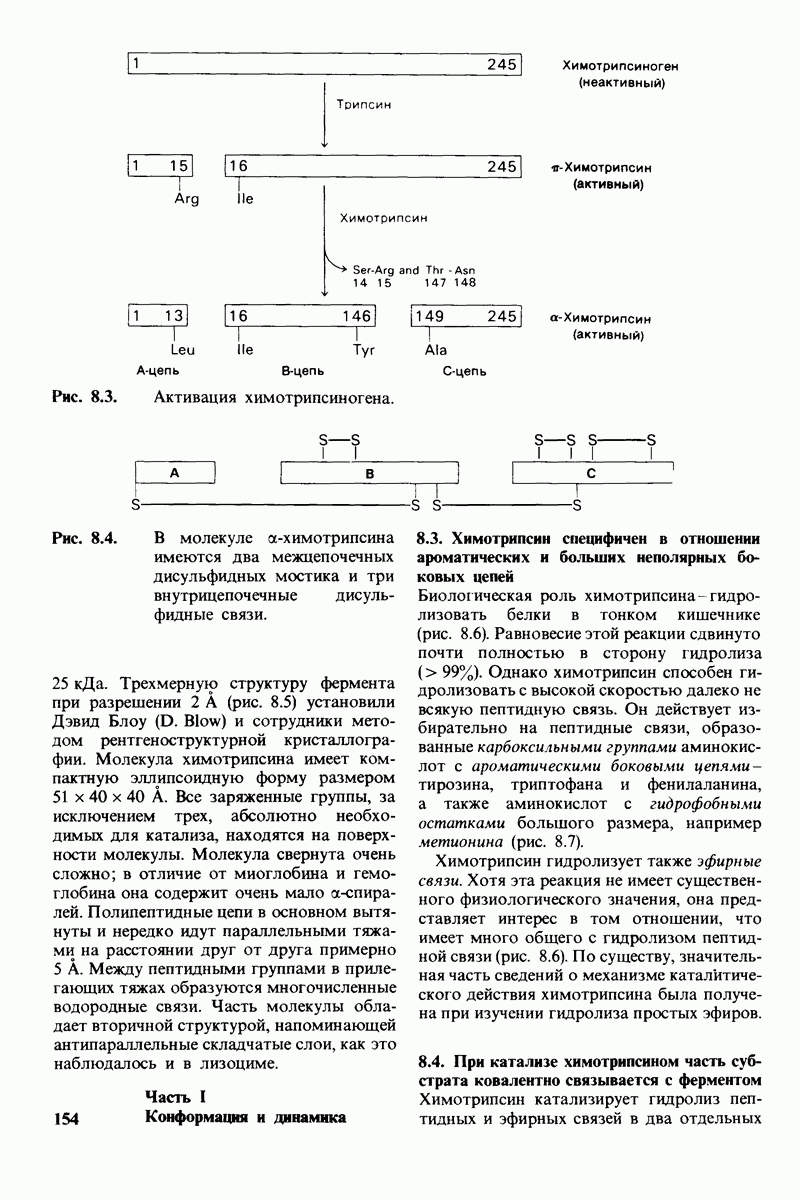
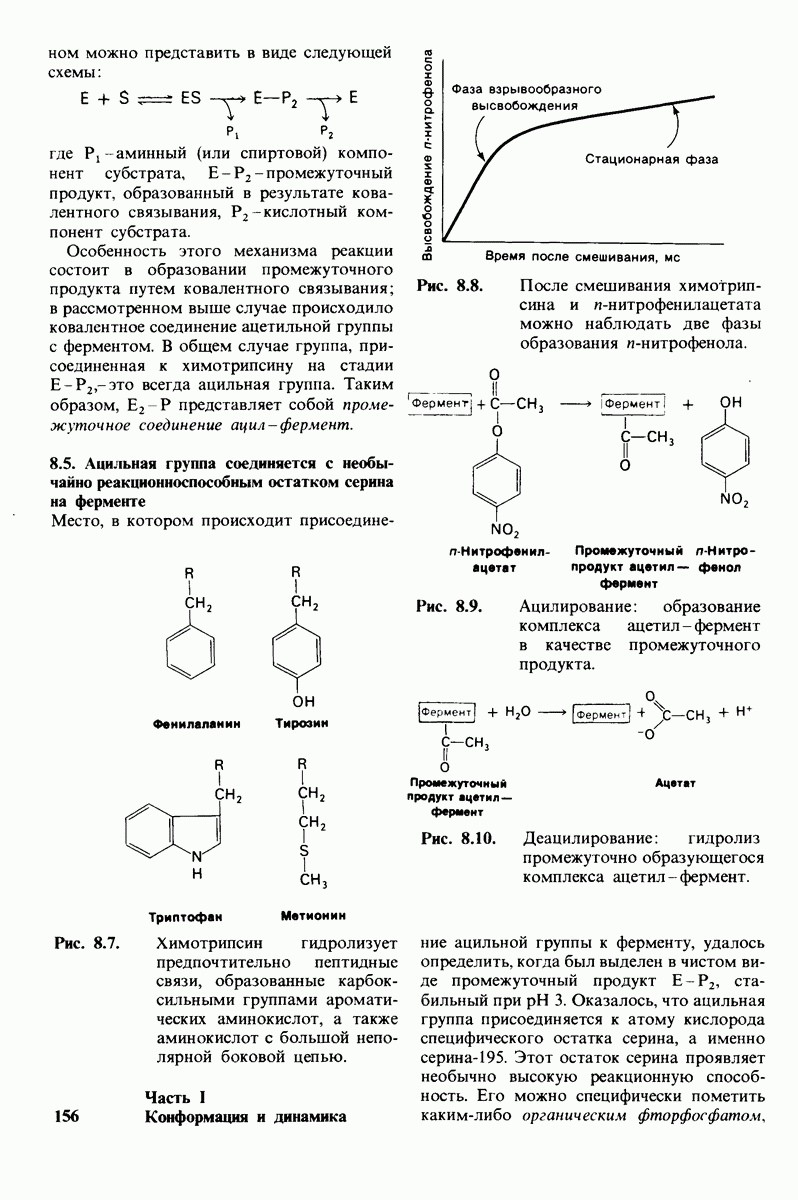
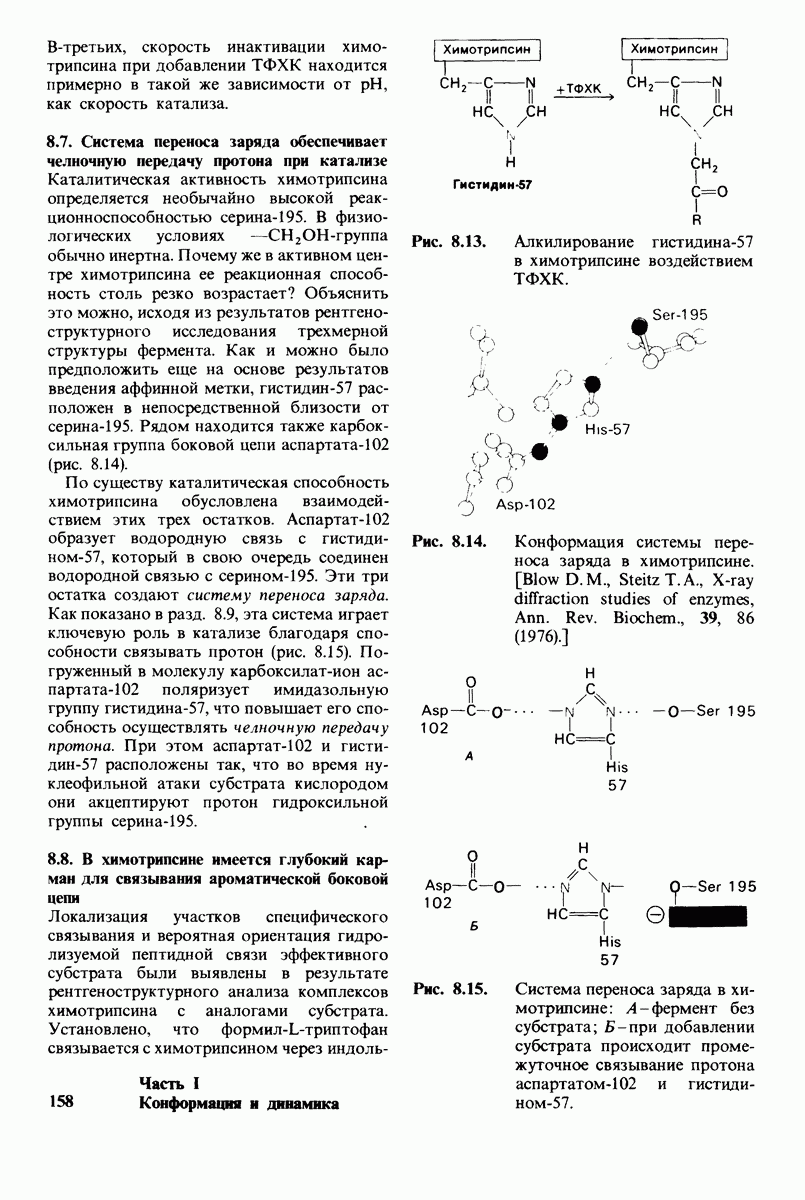
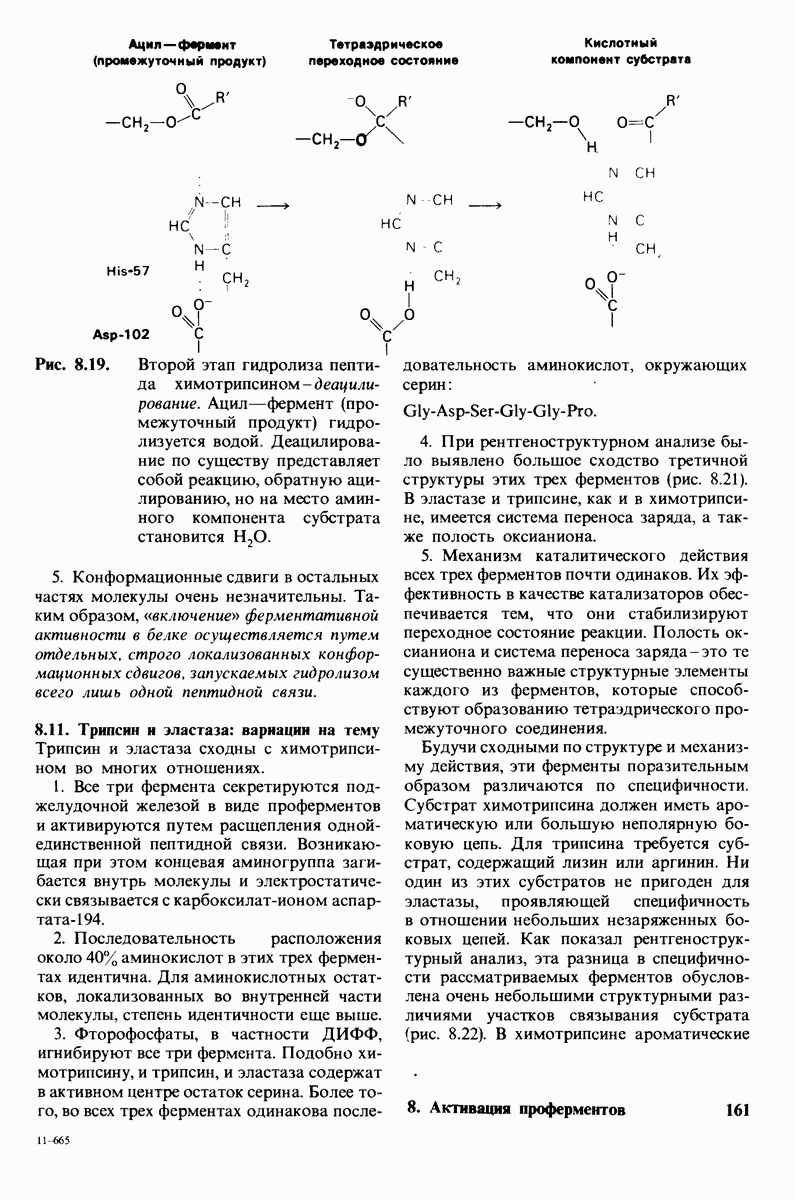
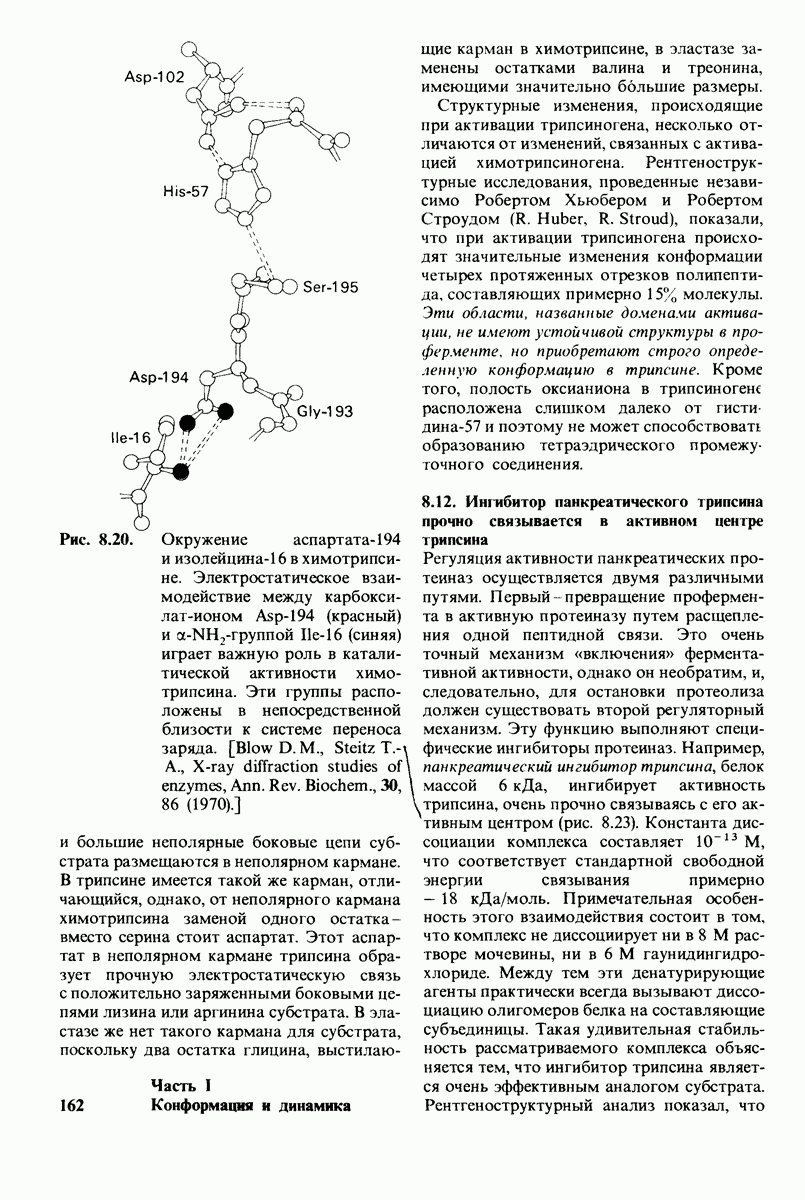
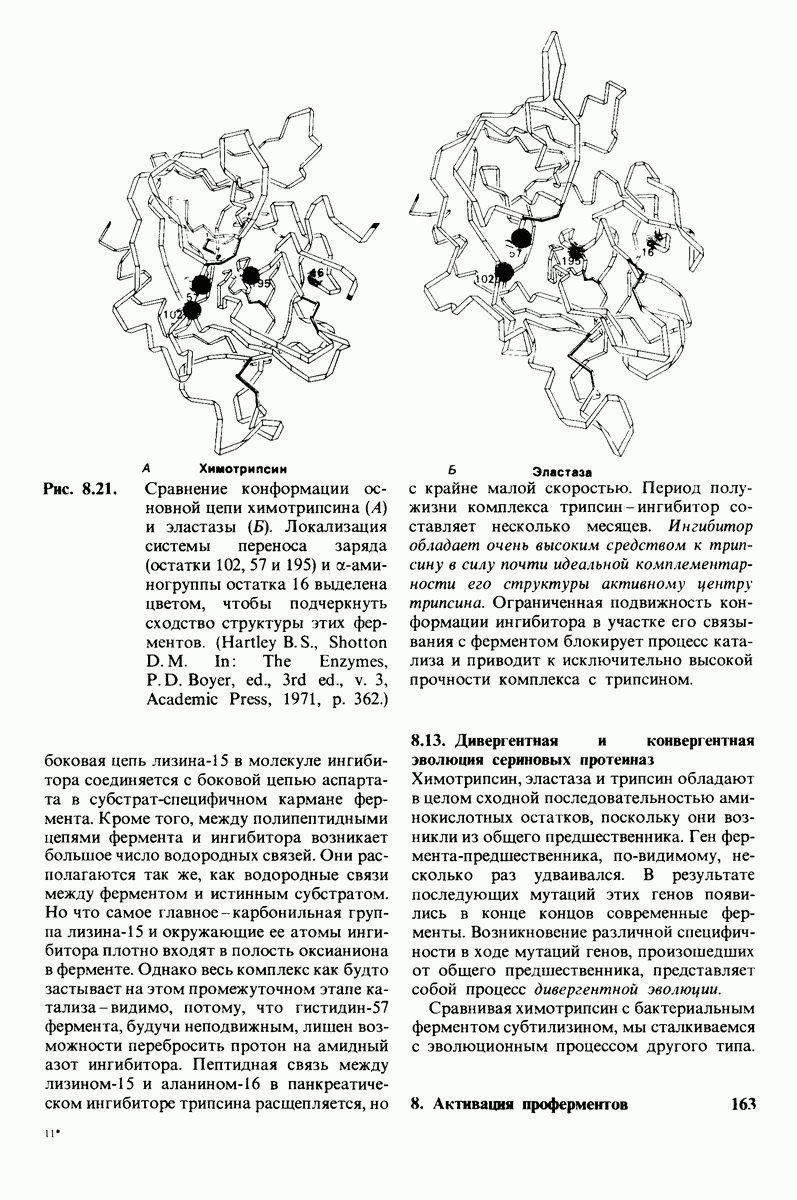
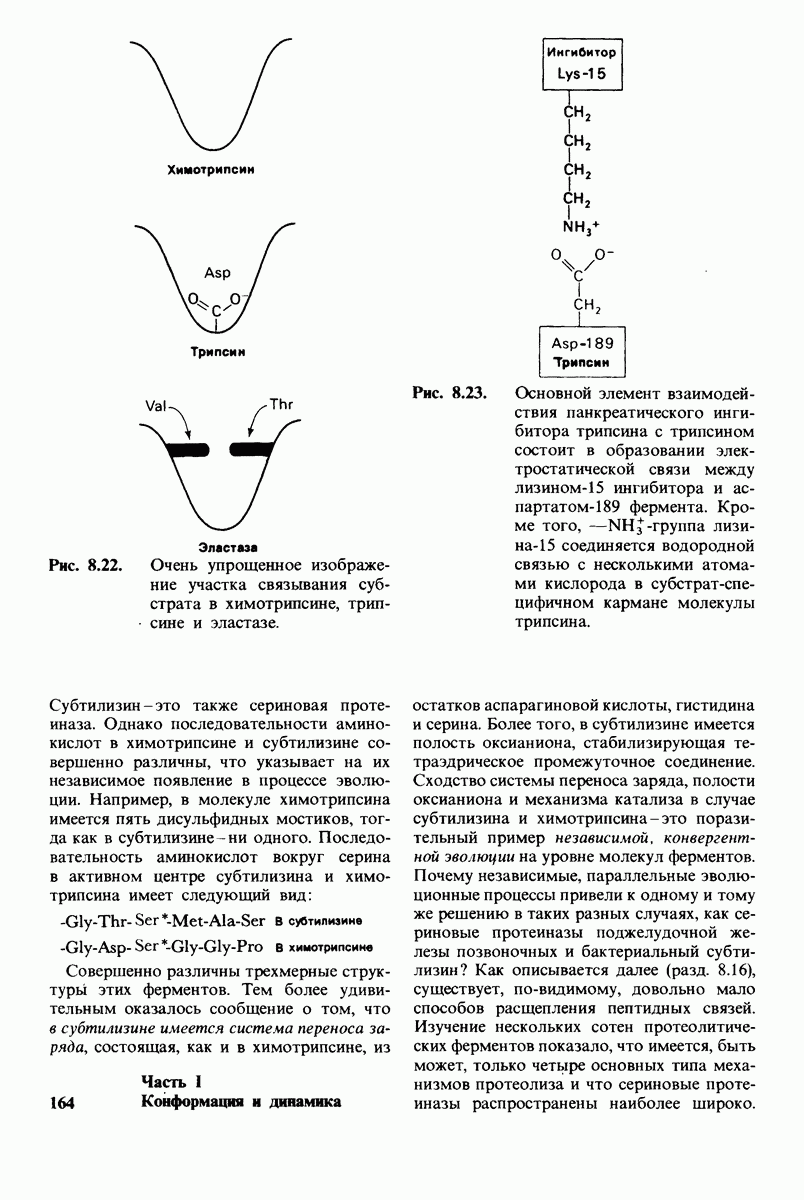
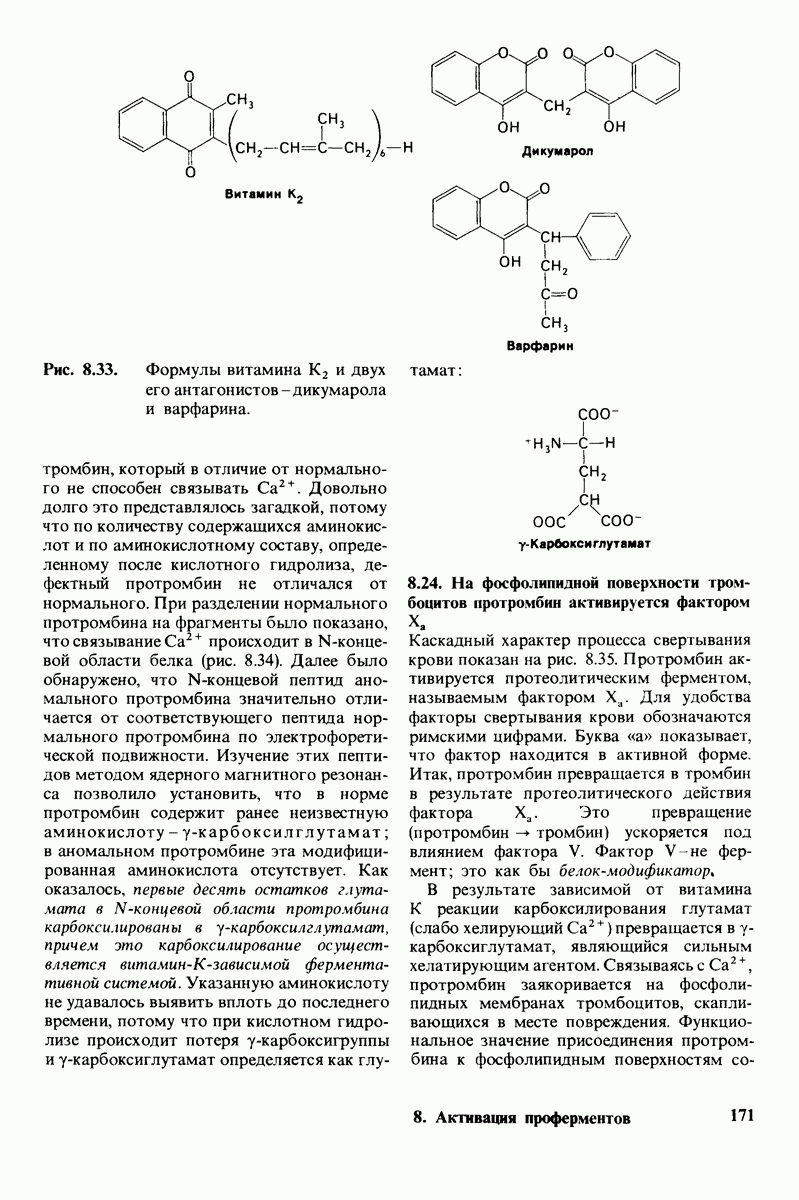
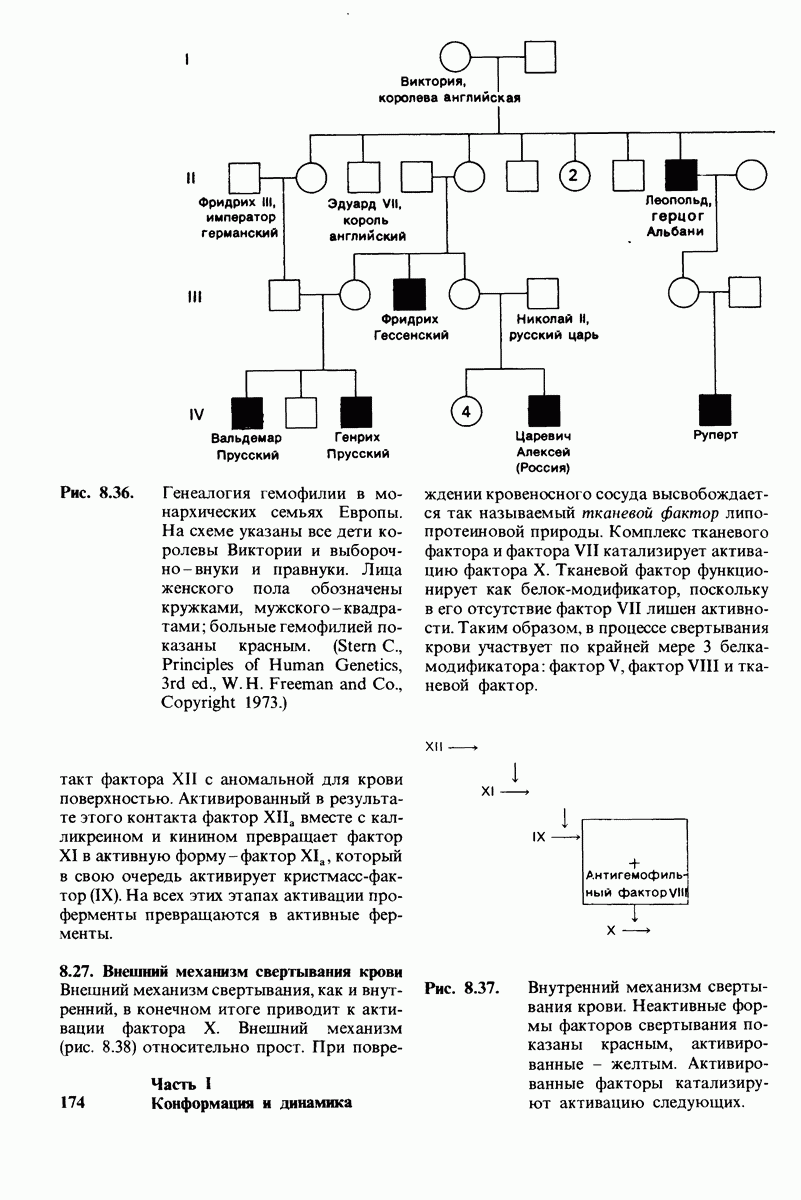
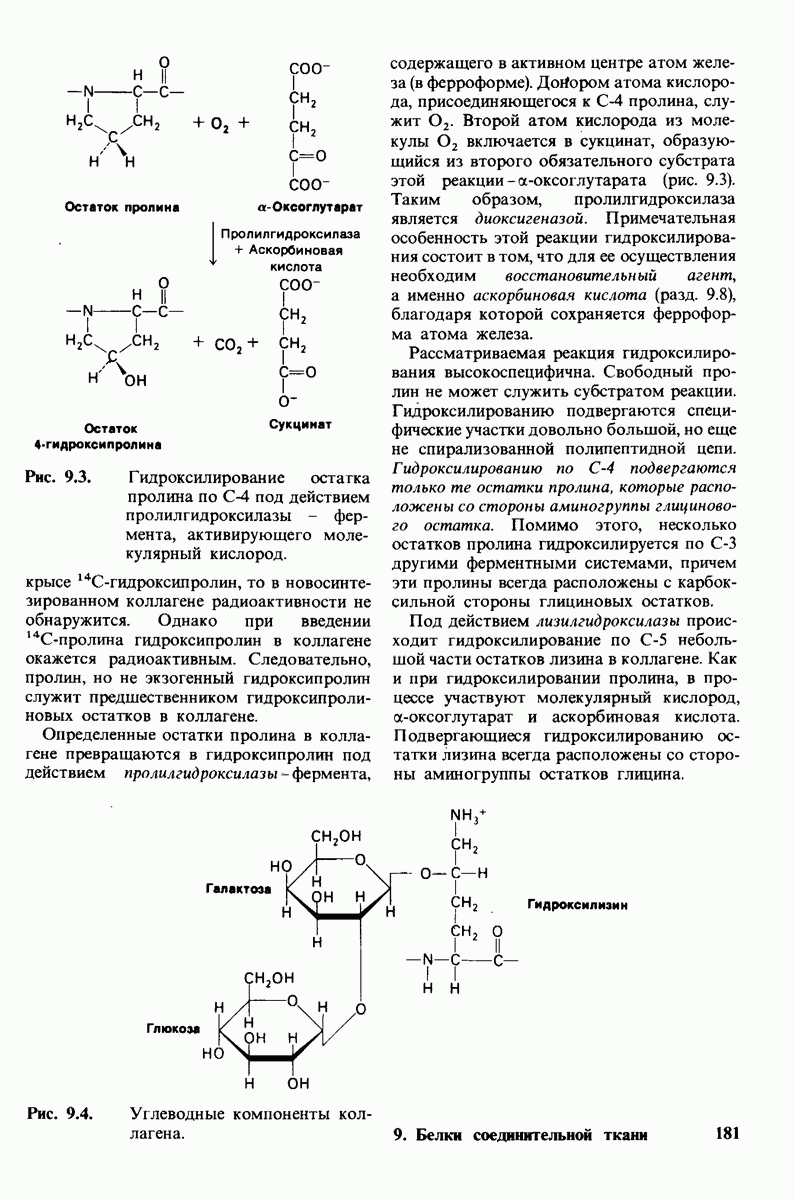
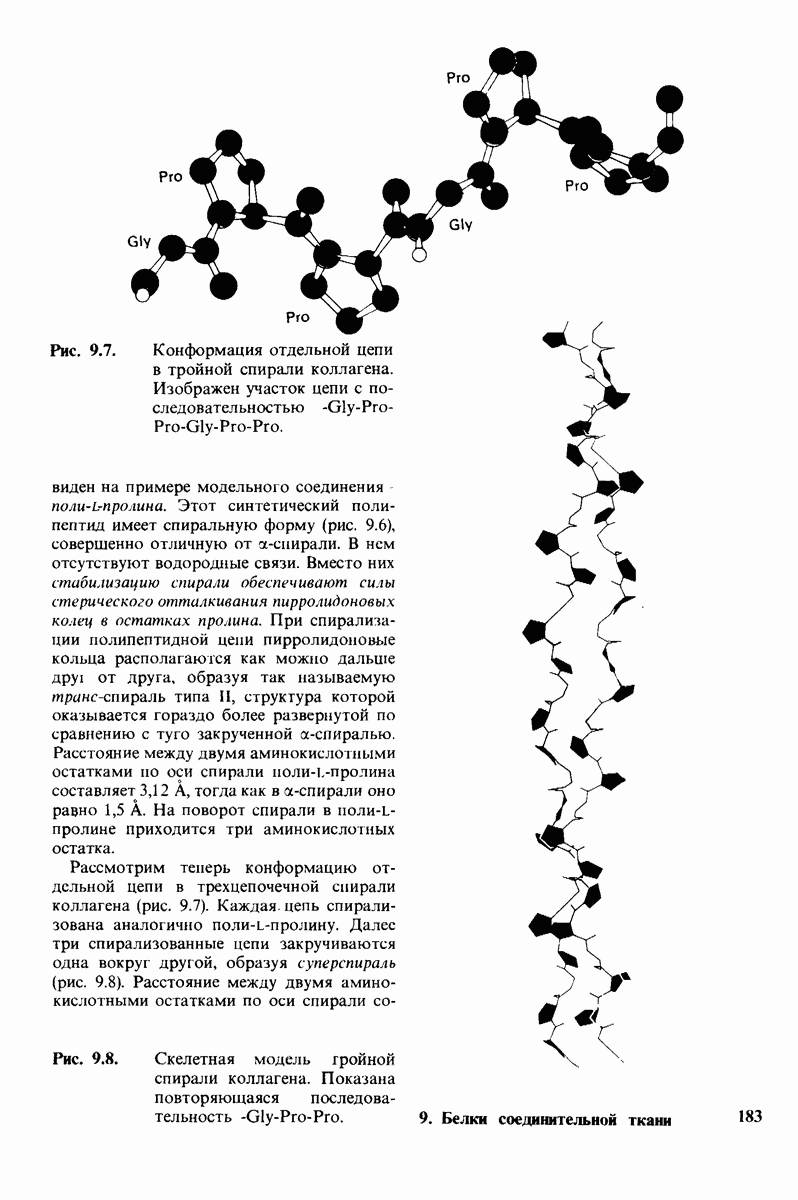
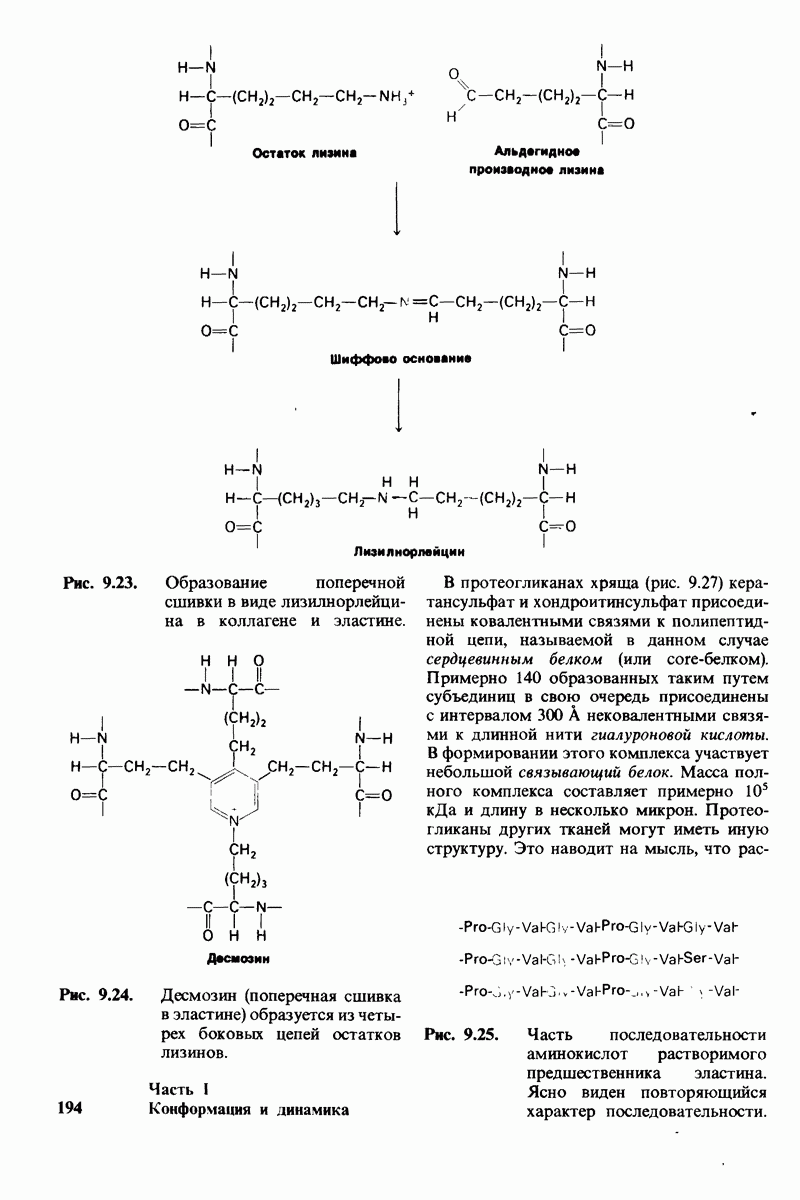
10.18. Трехмерная модель мембран по данным электронной микроскопии
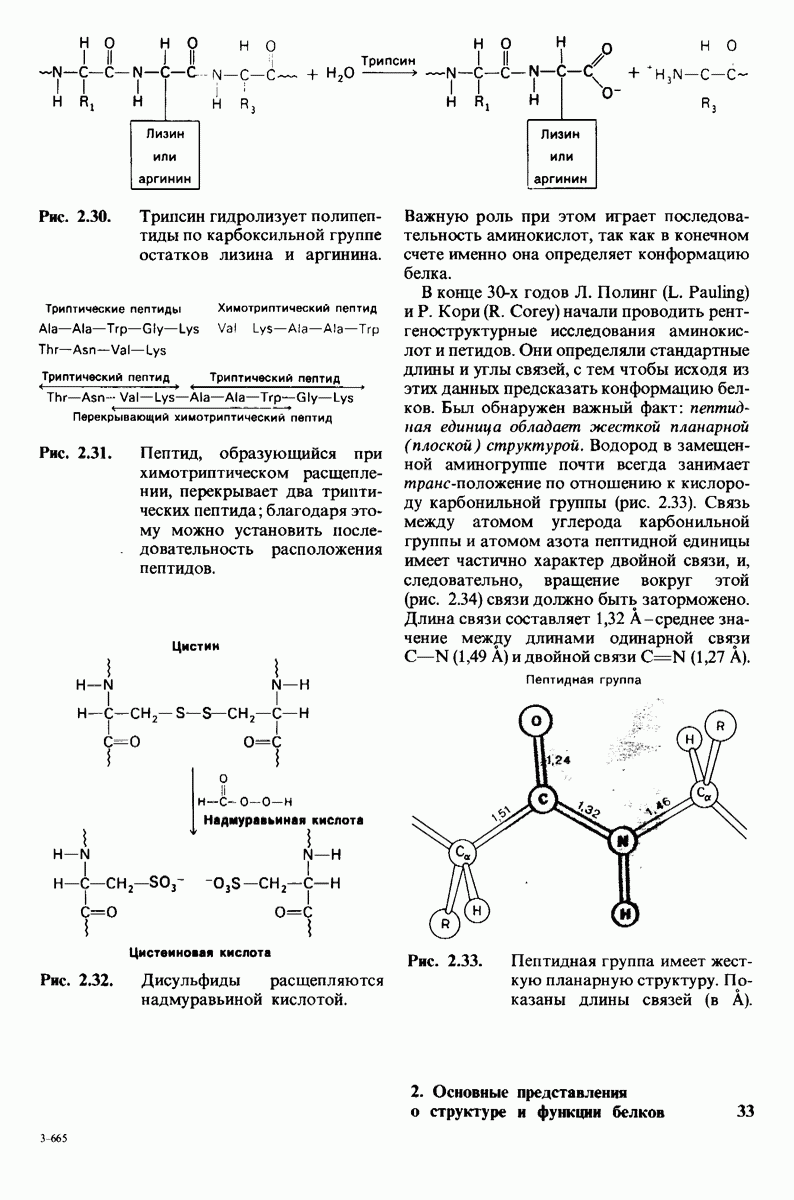
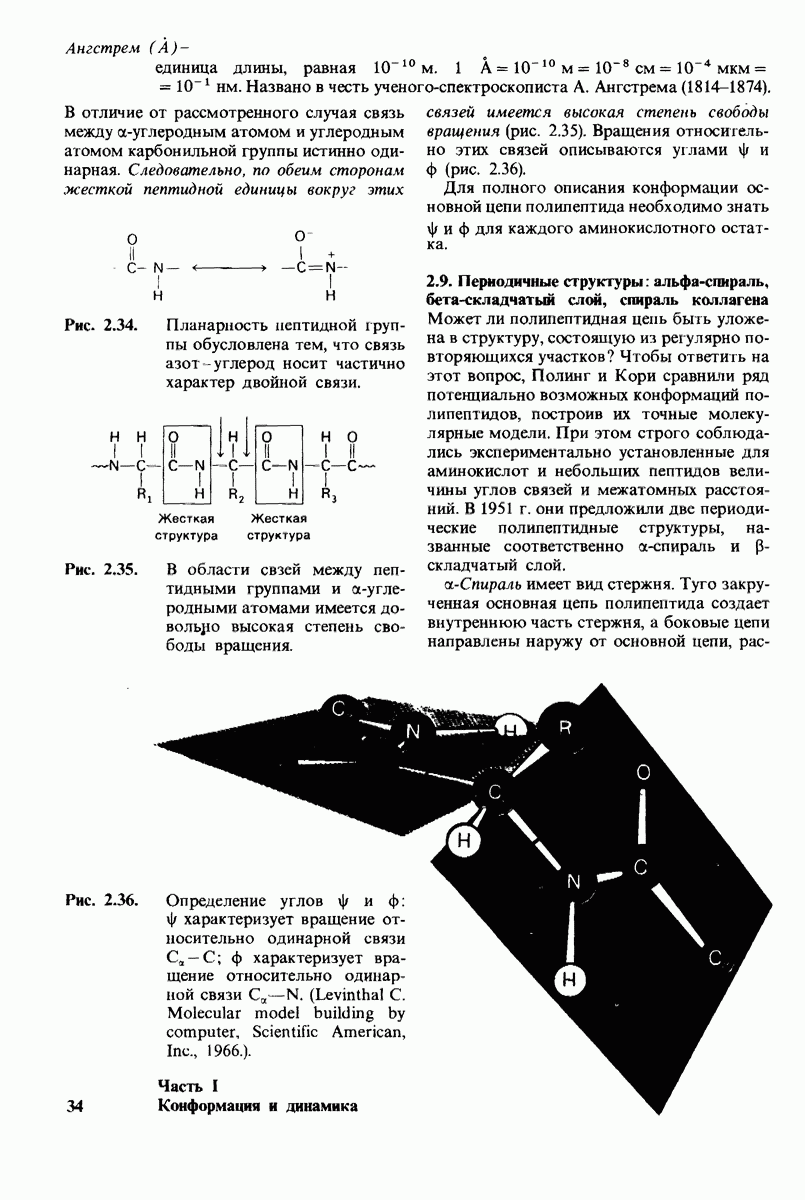
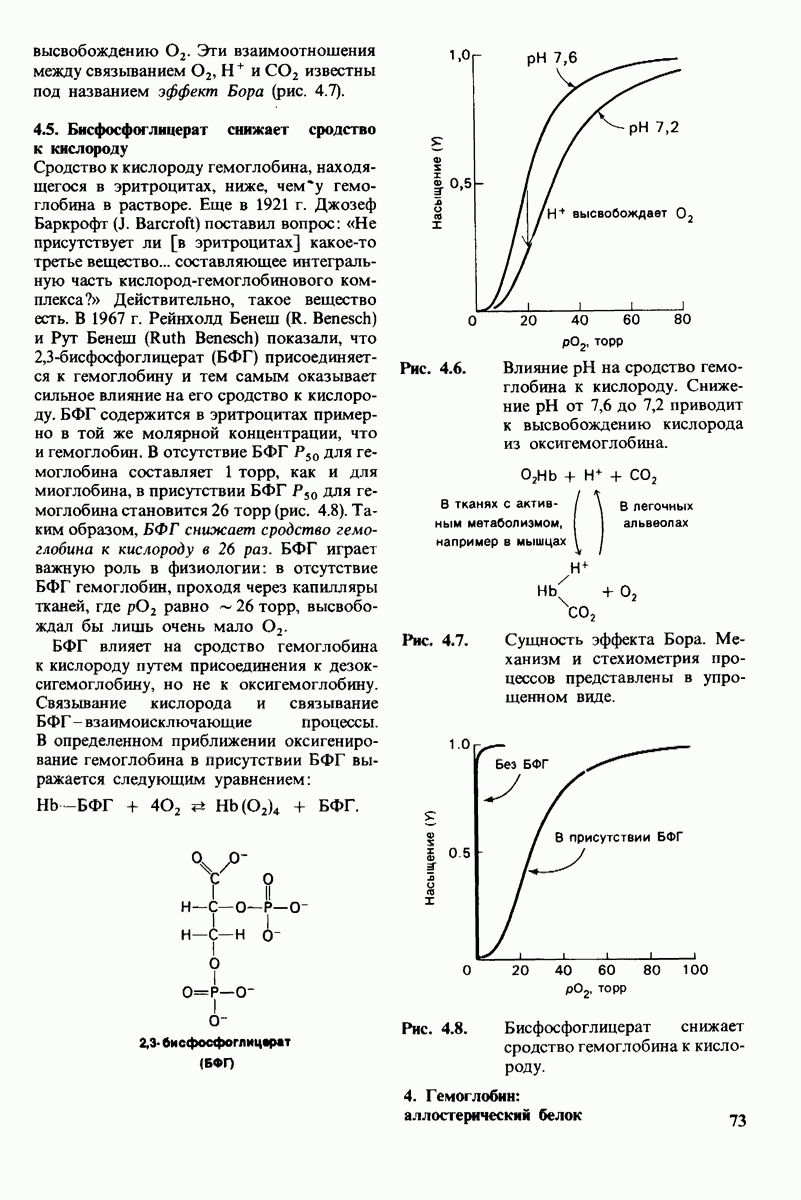
Как мы видели на примерах переносчиков кислорода и ферментов, описанных в предыдущих главах, рентгеноструктурный анализ является надежным методом изучения трехмерной структуры растворимых белков. Применим ли рентгеноструктурный анализ к мембранным белкам? Трудность заключается в том, что до сих пор не удавалось получить интегральных белков мембраны в виде трехмерных кристаллов. Однако некоторые мембранные белки образуют правильную решетку в плоскости мембраны, т. е. двумерные кристаллы. Структурный анализ этих кристаллоидных форм удается осуществить с помощью электронной микроскопии  в частности, такое исследование было с успехом проведено на пурпурной мембране Halobacterium halobium - бактерии, обитающей в соленой среде. Пурпурная мембрана - это специализированная область клеточной мембраны, содержащая бактериородопсин - белок массой
в частности, такое исследование было с успехом проведено на пурпурной мембране Halobacterium halobium - бактерии, обитающей в соленой среде. Пурпурная мембрана - это специализированная область клеточной мембраны, содержащая бактериородопсин - белок массой  который превращает энергию света в трансмембранный протонный градиент, используемый для синтеза АТР (разд. 19.21). Были получены кристаллоиды в виде листка, или диска, диаметром до
который превращает энергию света в трансмембранный протонный градиент, используемый для синтеза АТР (разд. 19.21). Были получены кристаллоиды в виде листка, или диска, диаметром до  Благодаря тому что в каждом из них содержалось около
Благодаря тому что в каждом из них содержалось около  молекул бактериородопсина, можно было получить изображение, используя очень слабый пучок электронов и тем самым сводя к минимуму радиационные повреждения. Кроме того, для получения изображения с высокой степенью разрешения можно было брать неокрашенные препараты. Одно электронно-микроскопическое изображение кристаллоидного листка
молекул бактериородопсина, можно было получить изображение, используя очень слабый пучок электронов и тем самым сводя к минимуму радиационные повреждения. Кроме того, для получения изображения с высокой степенью разрешения можно было брать неокрашенные препараты. Одно электронно-микроскопическое изображение кристаллоидного листка 

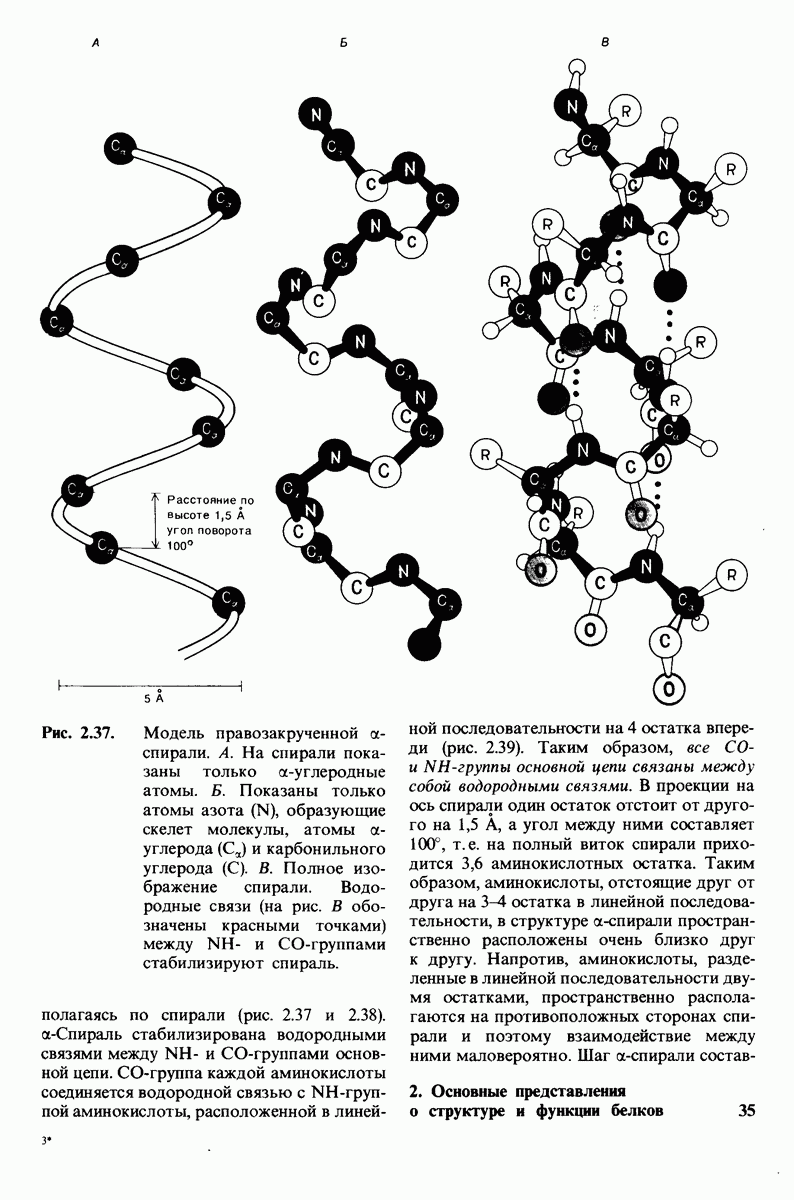
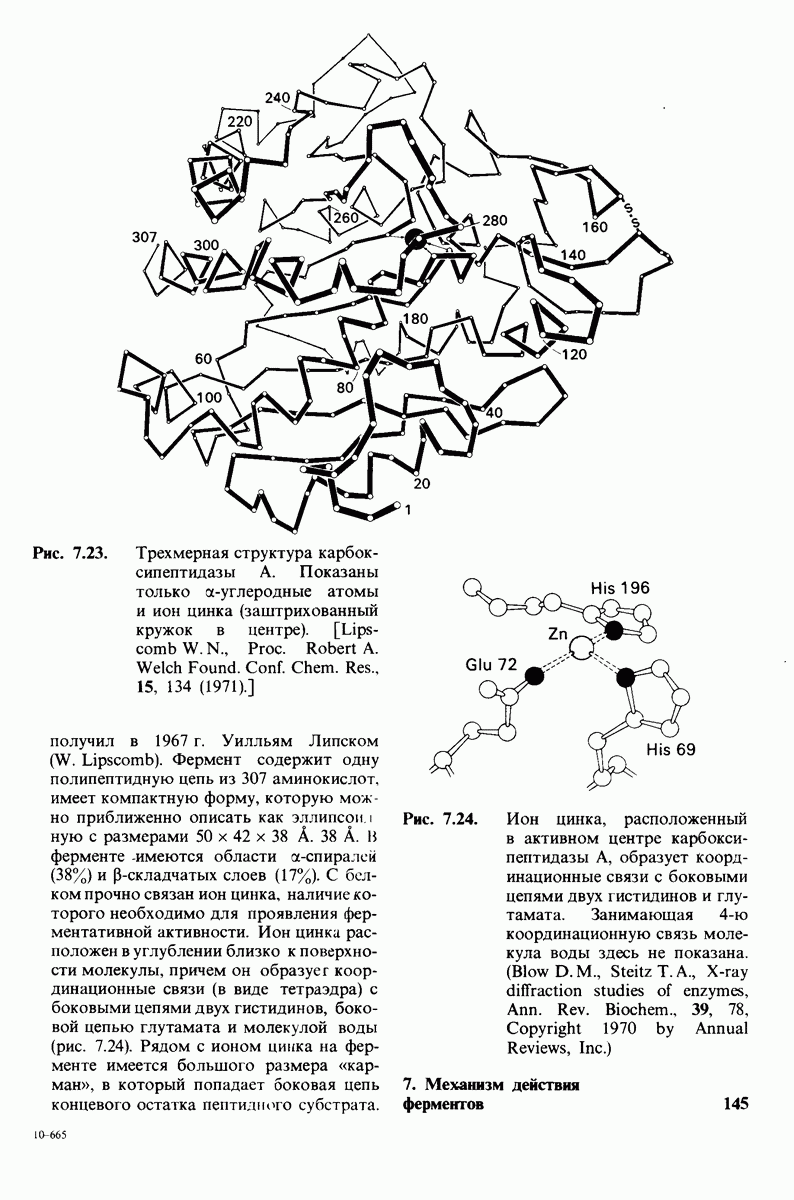
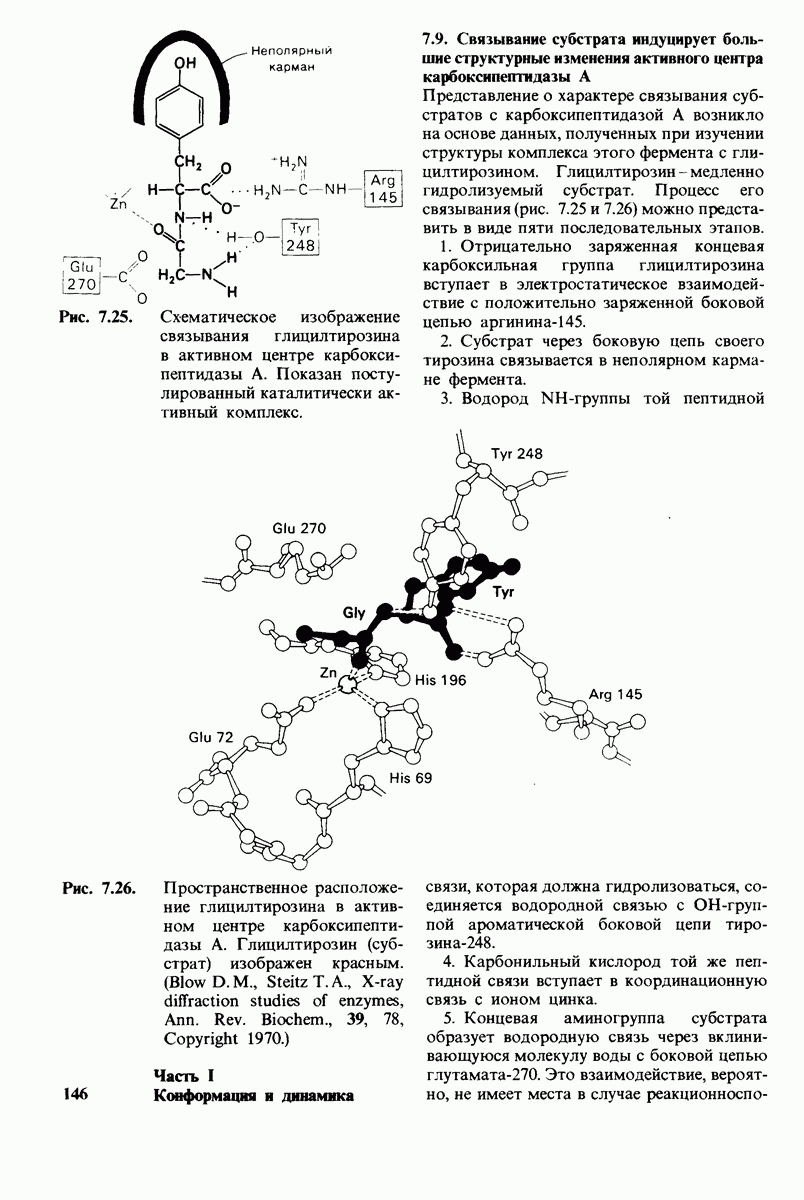
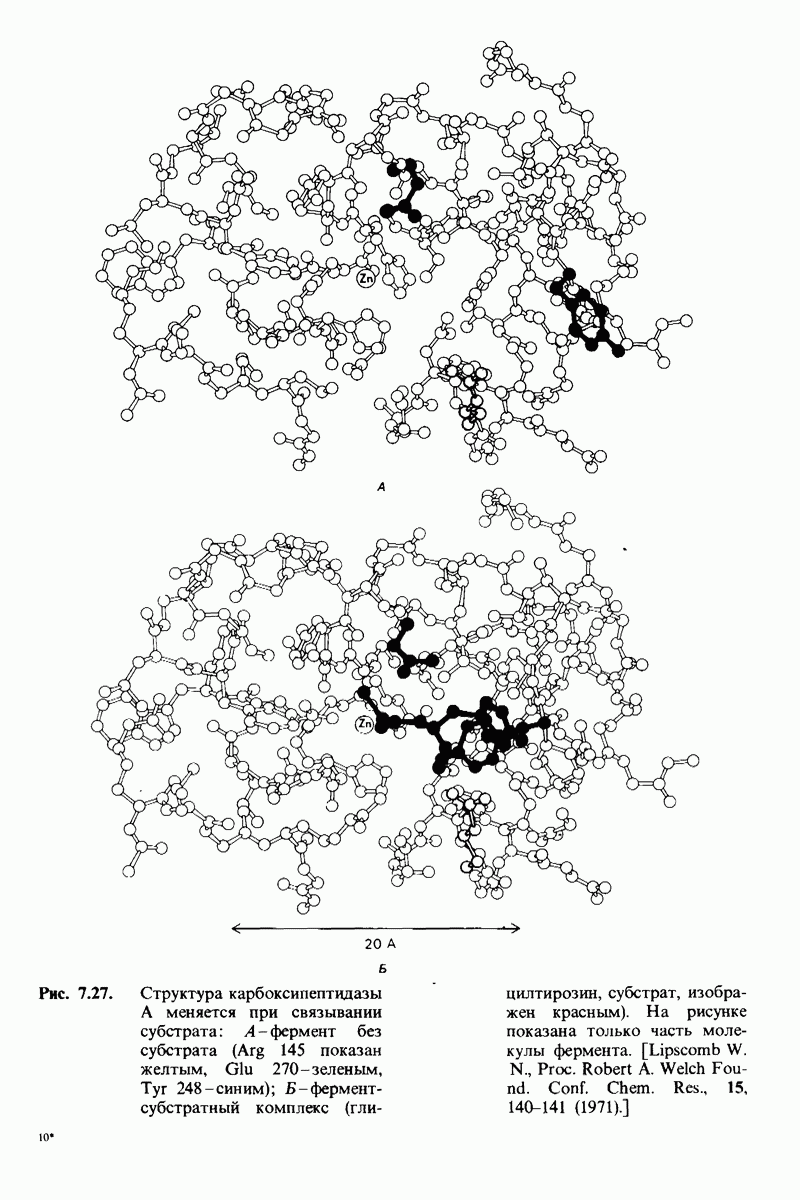
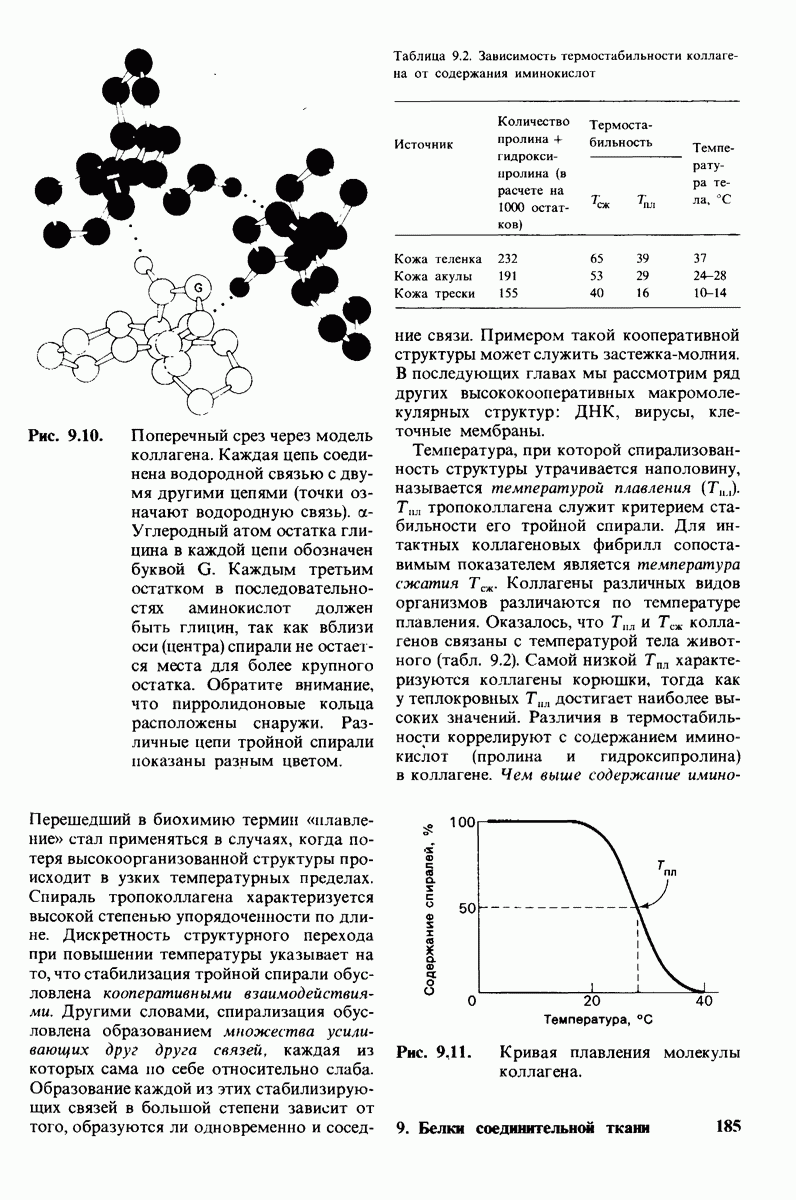
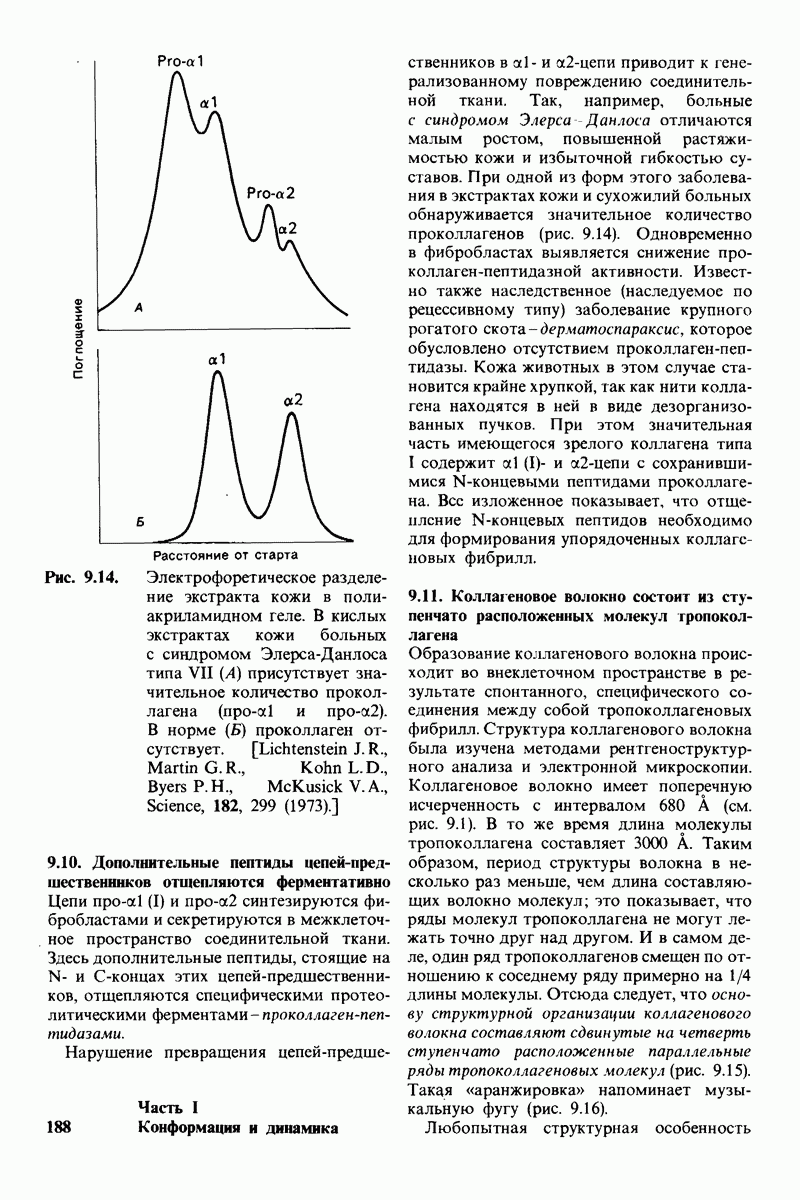
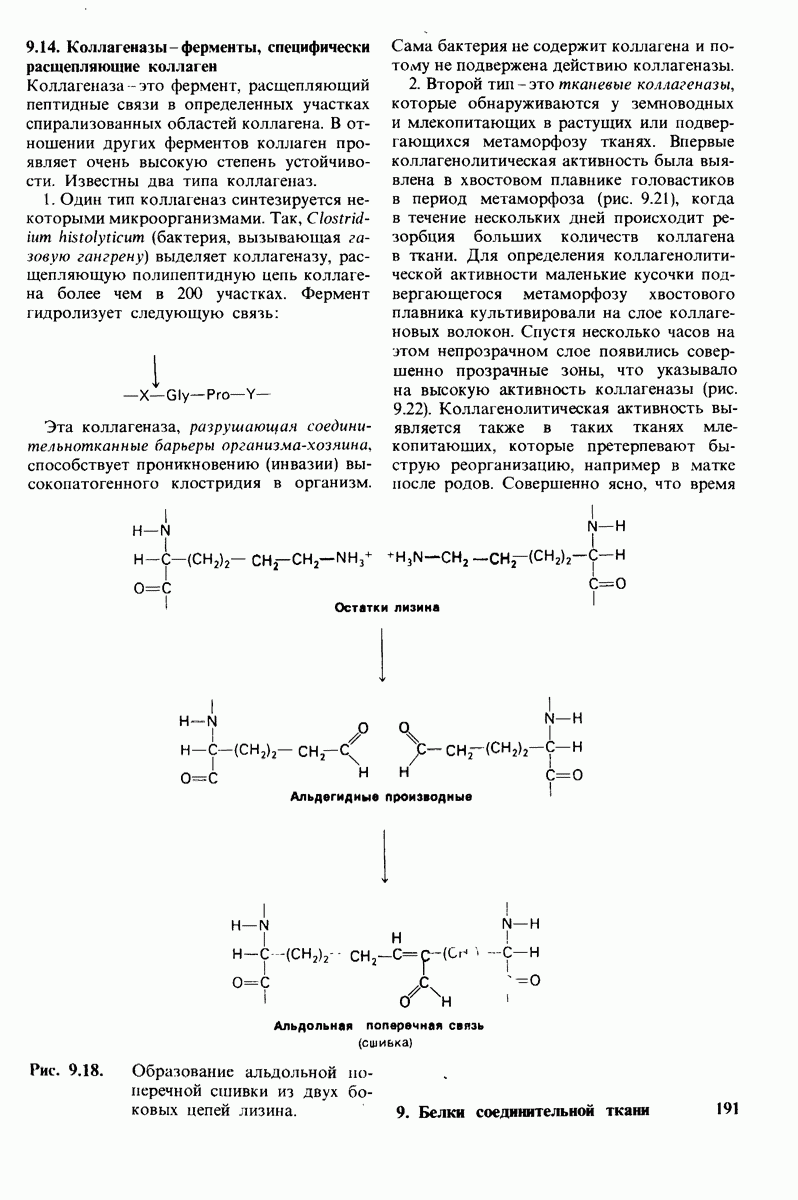
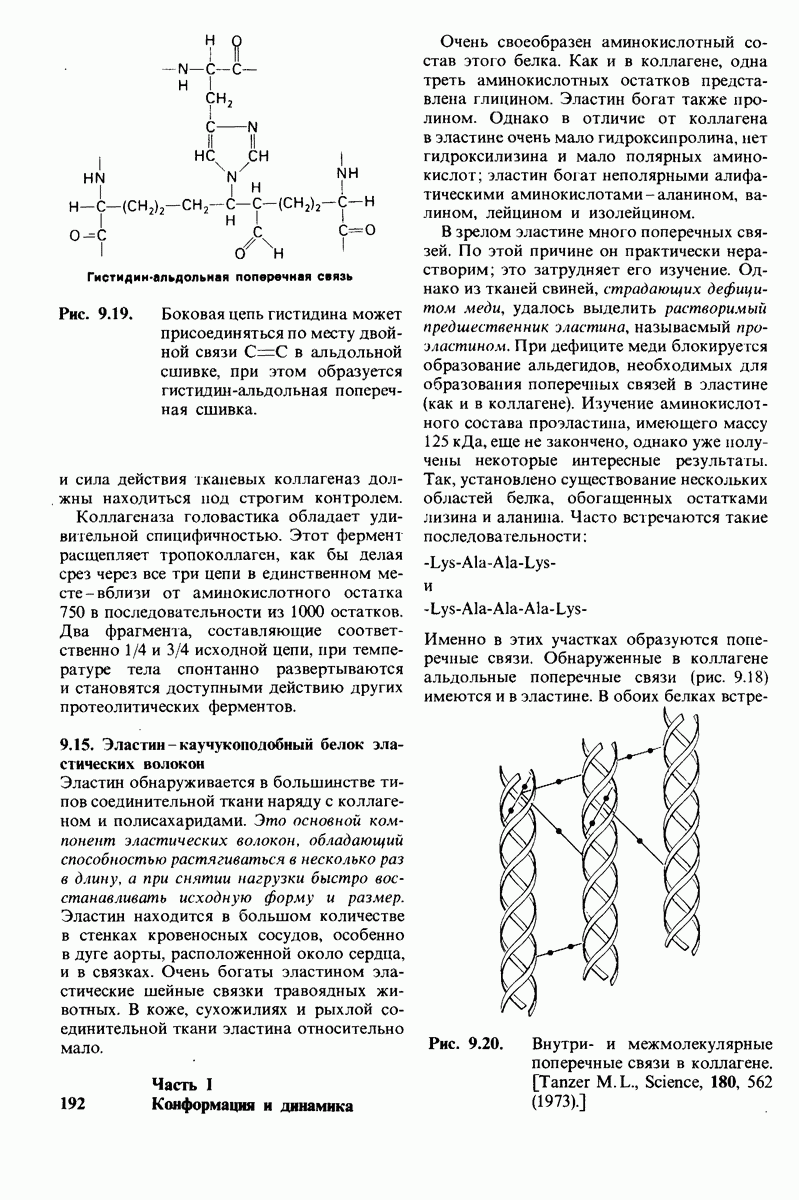
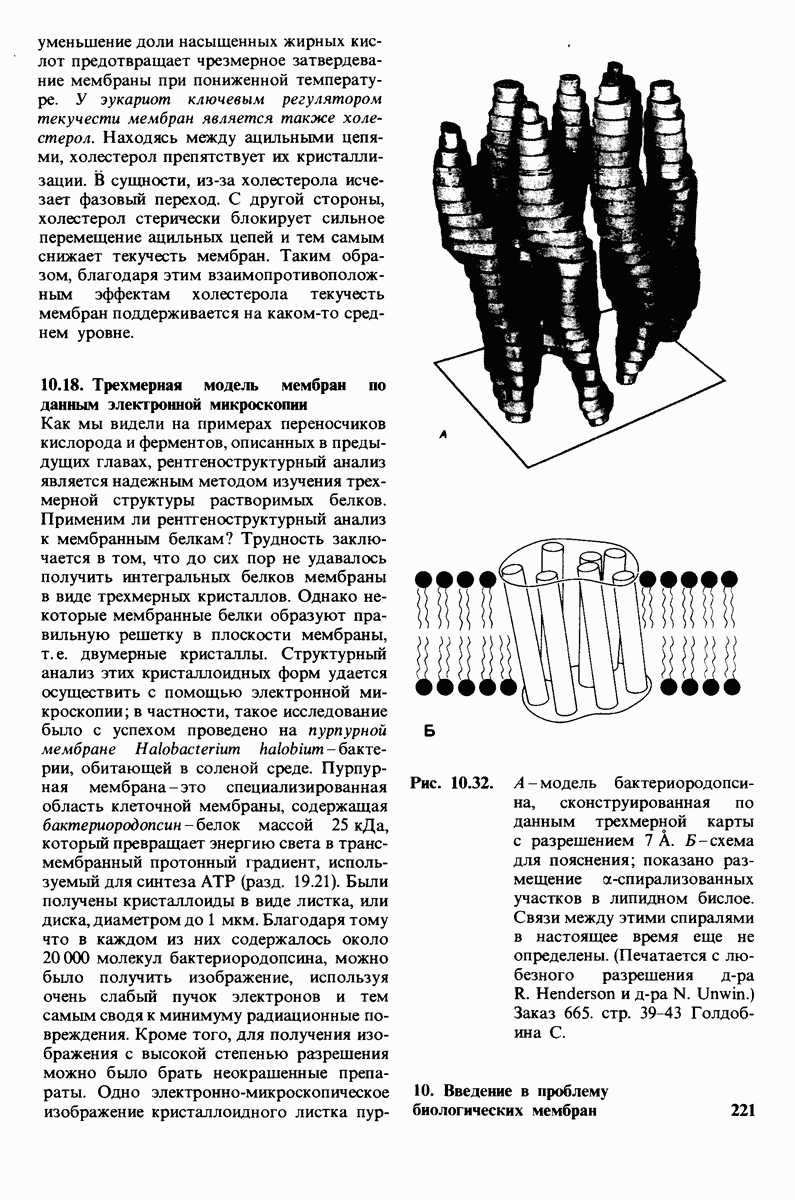
Рис. 10.32. А - модель бактериородопсина, сконструированная по данным трехмерной карты с разрешением 7 А. Б - схема для пояснения; показано размещение  -спирализованных участков в липидном бислое. Связи между этими спиралями в настоящее время еще не определены. (Печатается с любезного разрешения д-ра R. Henderson и д-ра N. Unwin.) Заказ 665. стр. 39-43 Голдобина С.
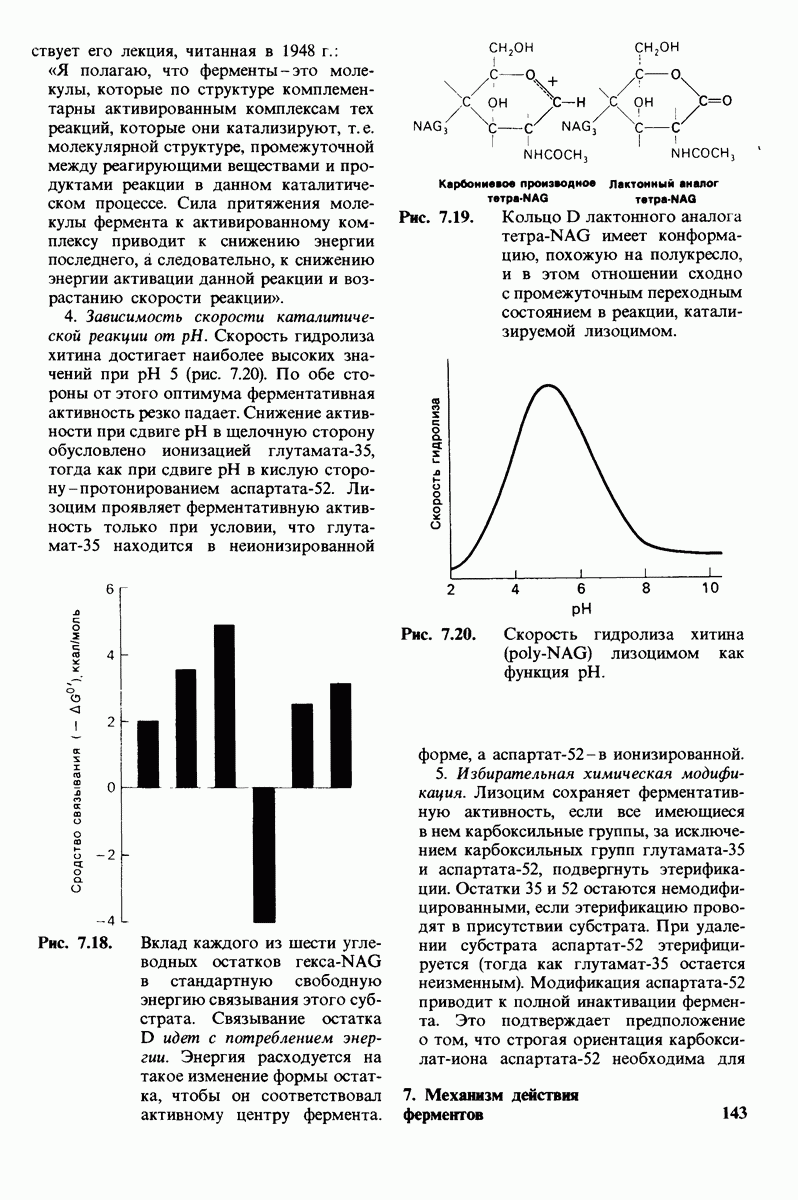
-спирализованных участков в липидном бислое. Связи между этими спиралями в настоящее время еще не определены. (Печатается с любезного разрешения д-ра R. Henderson и д-ра N. Unwin.) Заказ 665. стр. 39-43 Голдобина С.
пурной мембраны дает вид структуры, спроецированной на плоскость. Следующий этап - получение изображений последовательных срезов и обработка информации, заключенной примерно в 20 снимках, методом рядов Фурье.
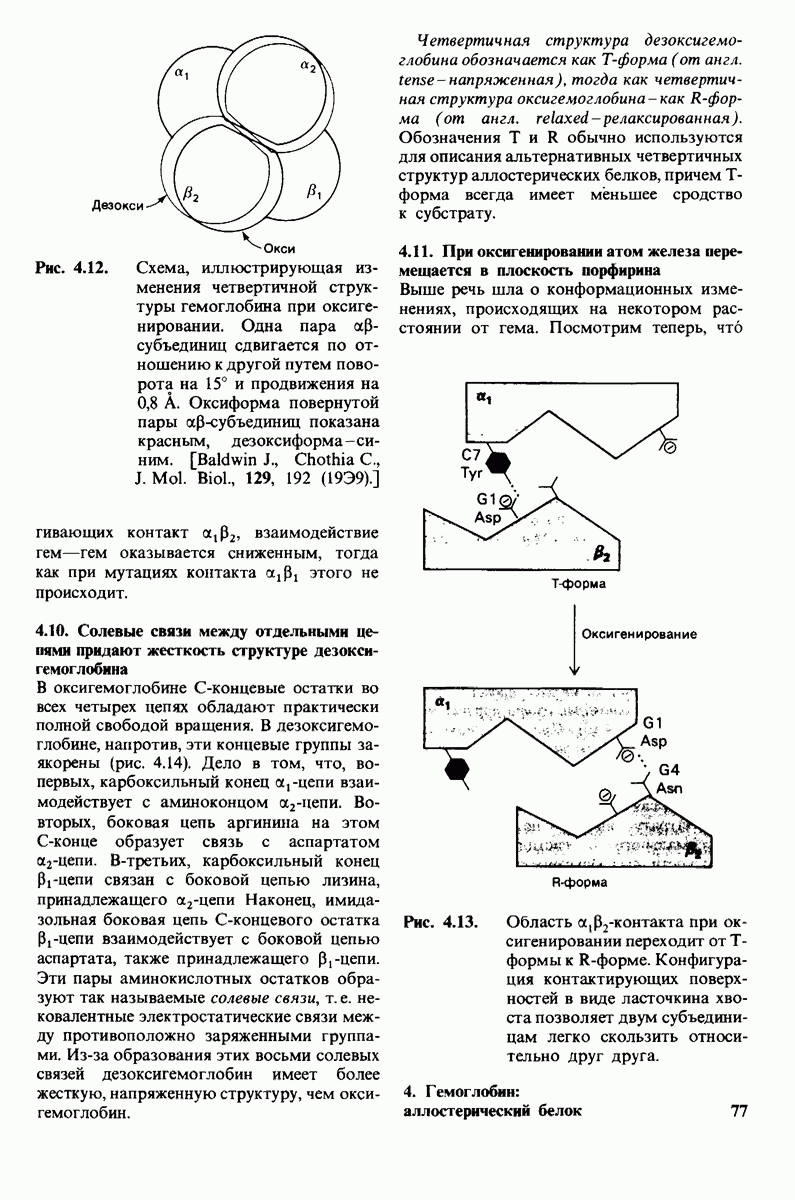
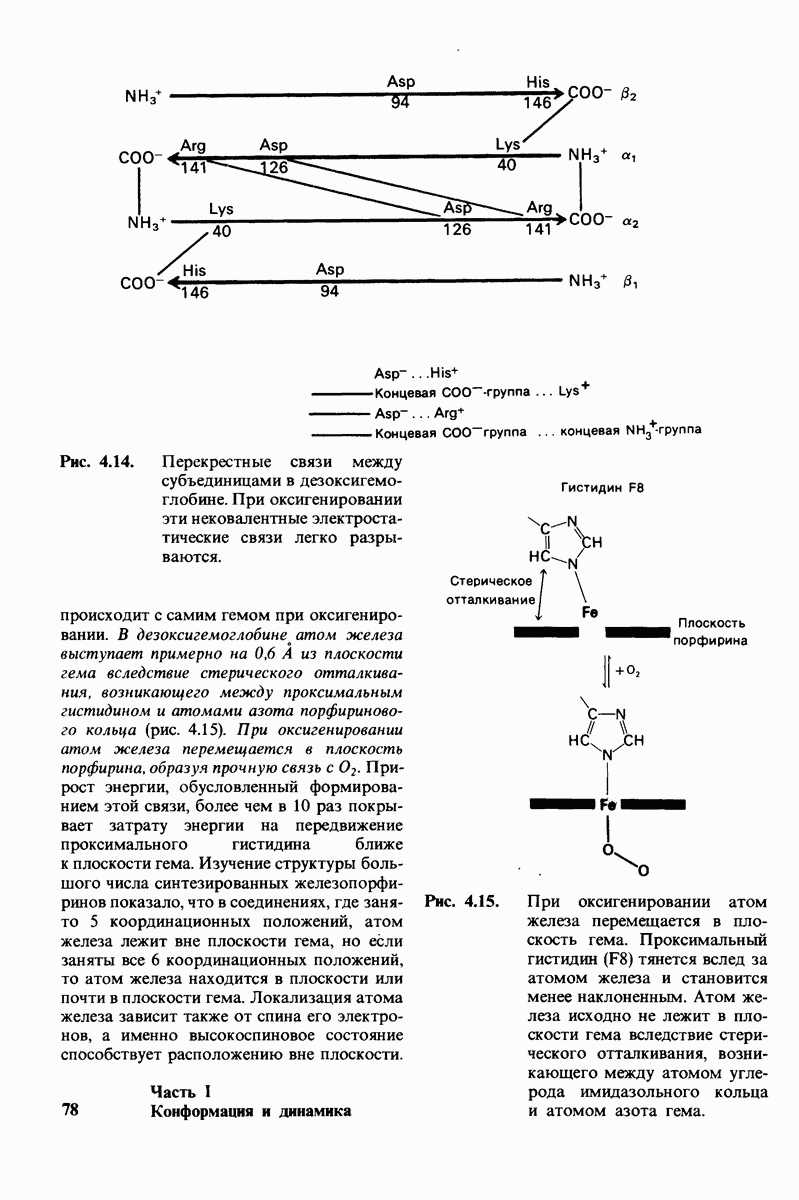
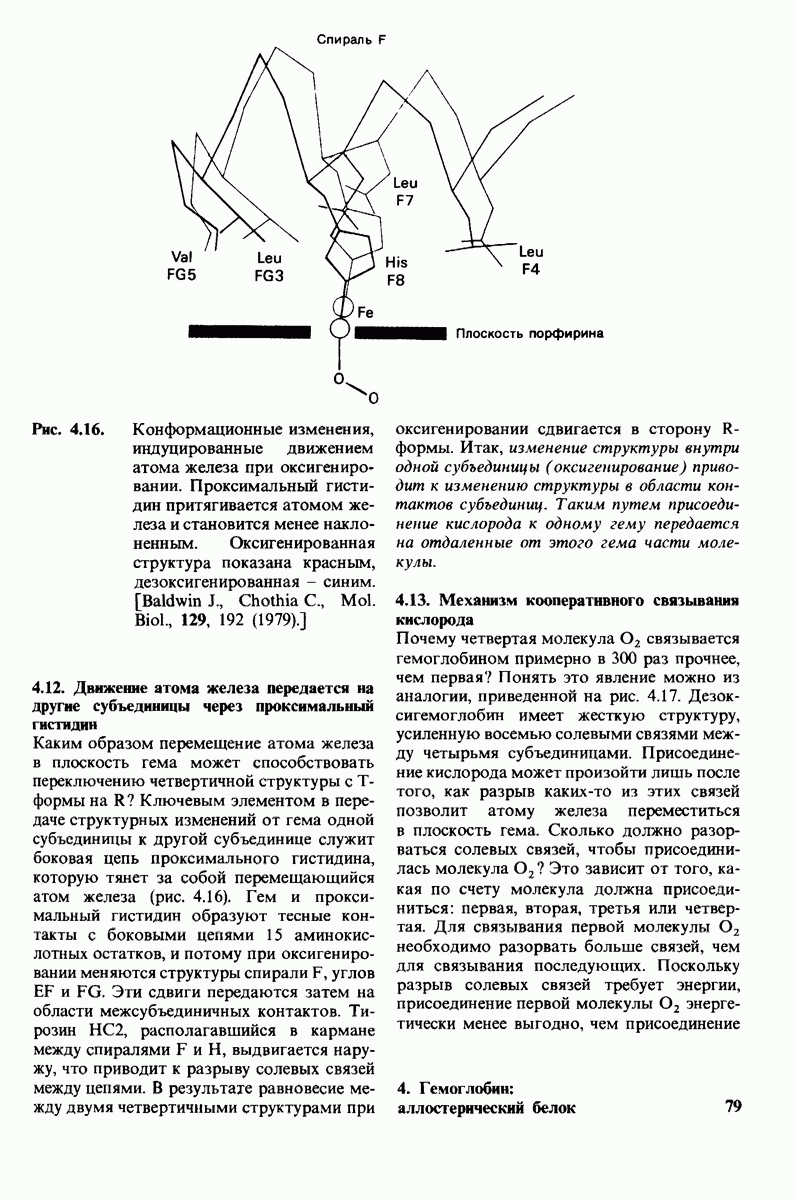
Используя описанный подход, Ричард Хендерсон и Нигел Ануин (R. Henderson, N. Unwin) сконструировали трехмерную модель пурпурной мембраны с разрешением 7 А (рис. 10.32). Белки этой мембраны содержат семь плотно упакованных  -спиралей, которые идут почти перпендикулярно плоскости мембраны, занимая по ширине около 45 А. Пространство между молекулами белка заполнено липидным бислоем. Вполне вероятно, что принцип структурной организации пурпурной мембраны использован и в организации других интегральных белков. Предполагается, в частности, что мембранные насосы и каналы содержат
-спиралей, которые идут почти перпендикулярно плоскости мембраны, занимая по ширине около 45 А. Пространство между молекулами белка заполнено липидным бислоем. Вполне вероятно, что принцип структурной организации пурпурной мембраны использован и в организации других интегральных белков. Предполагается, в частности, что мембранные насосы и каналы содержат  -спирализованные участки, пронизывающие бислой.
-спирализованные участки, пронизывающие бислой.
Заключение
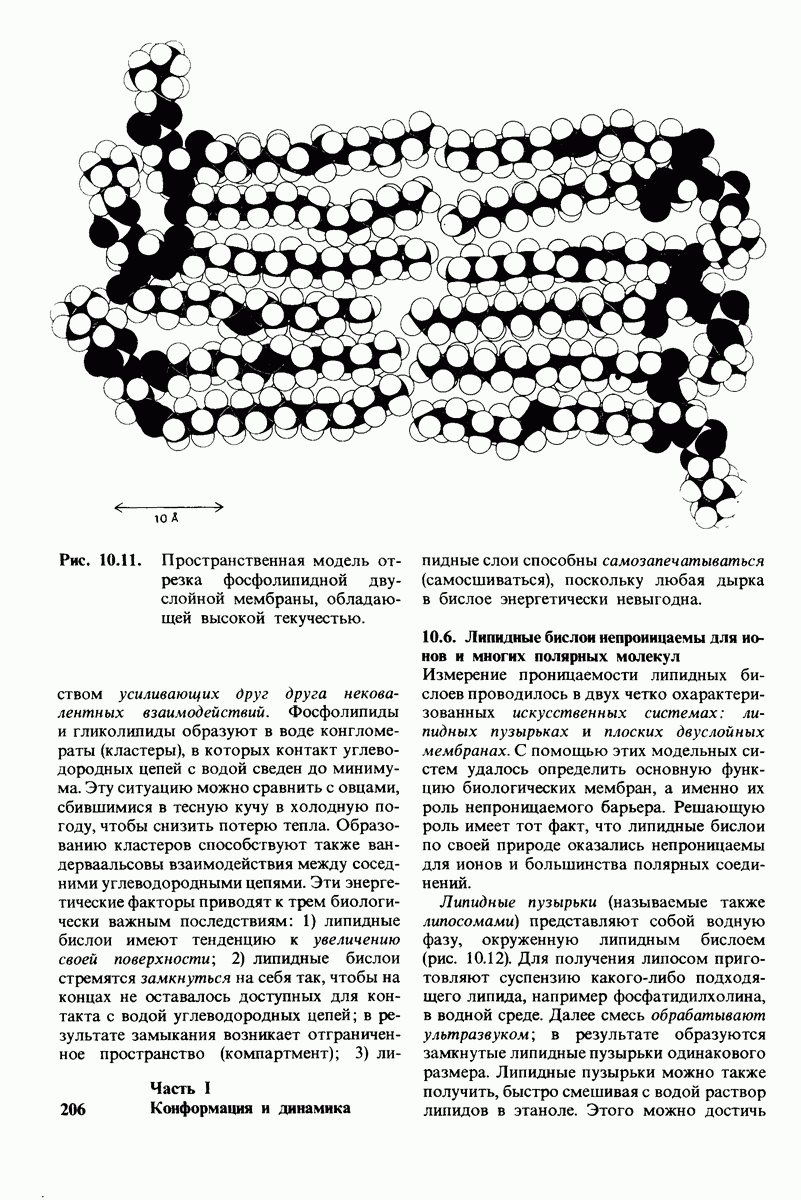
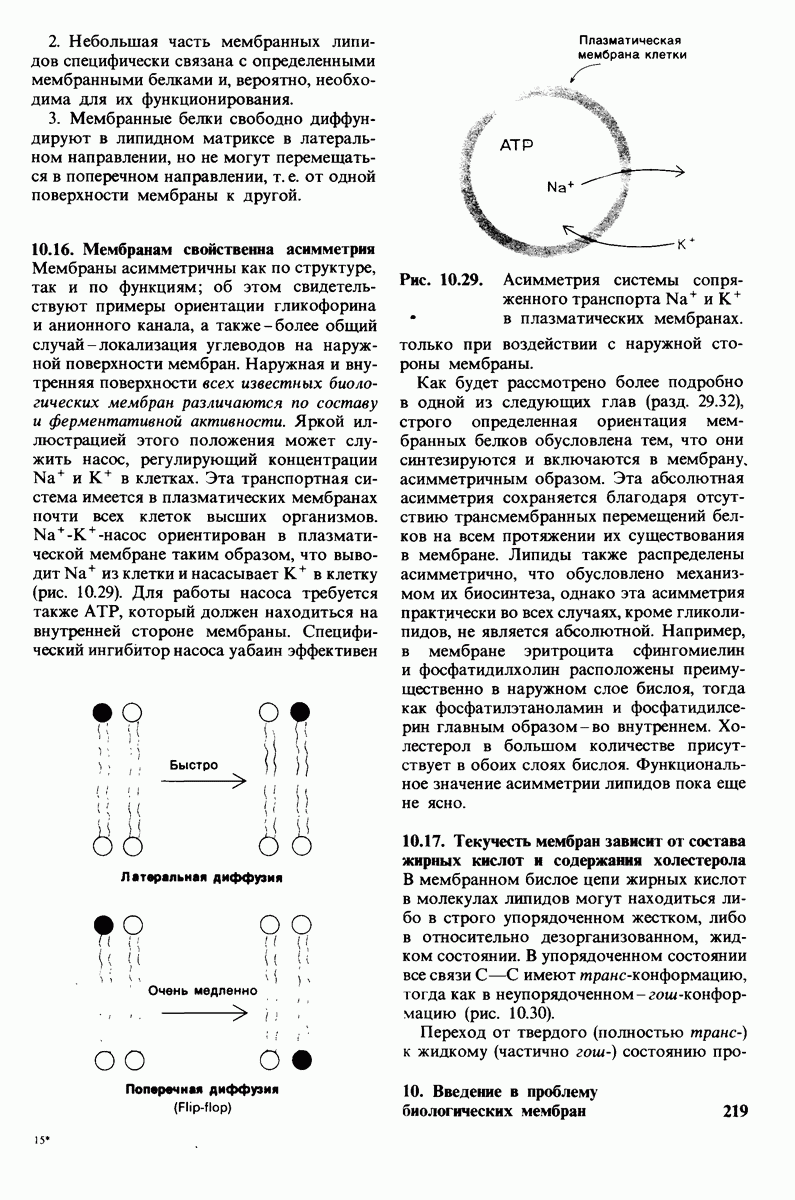
Биологические мембраны представляют собой плоские структуры шириной порядка 75 А, которые состоят из молекул белков и липидов, удерживаемых вместе нековалентными связями. Мембраны служат барьерами проницаемости с высокой степенью избирательности. Они отграничивают замкнутые пространства (компартменты) в виде целых клеток или субклеточных органелл. Встроенные в мембраны насосы и каналы регулируют молекулярный и ионный состав этих компартментов. Мембраны регулируют также обмен информацией между клетками. В частности, на некоторых мембранах находятся рецепторы гормонов, например рецепторы инсулина. Кроме того, мембраны непосредственно участвуют в таких процессах превращения энергии, как фотосинтез и окислительное фосфорилирование.
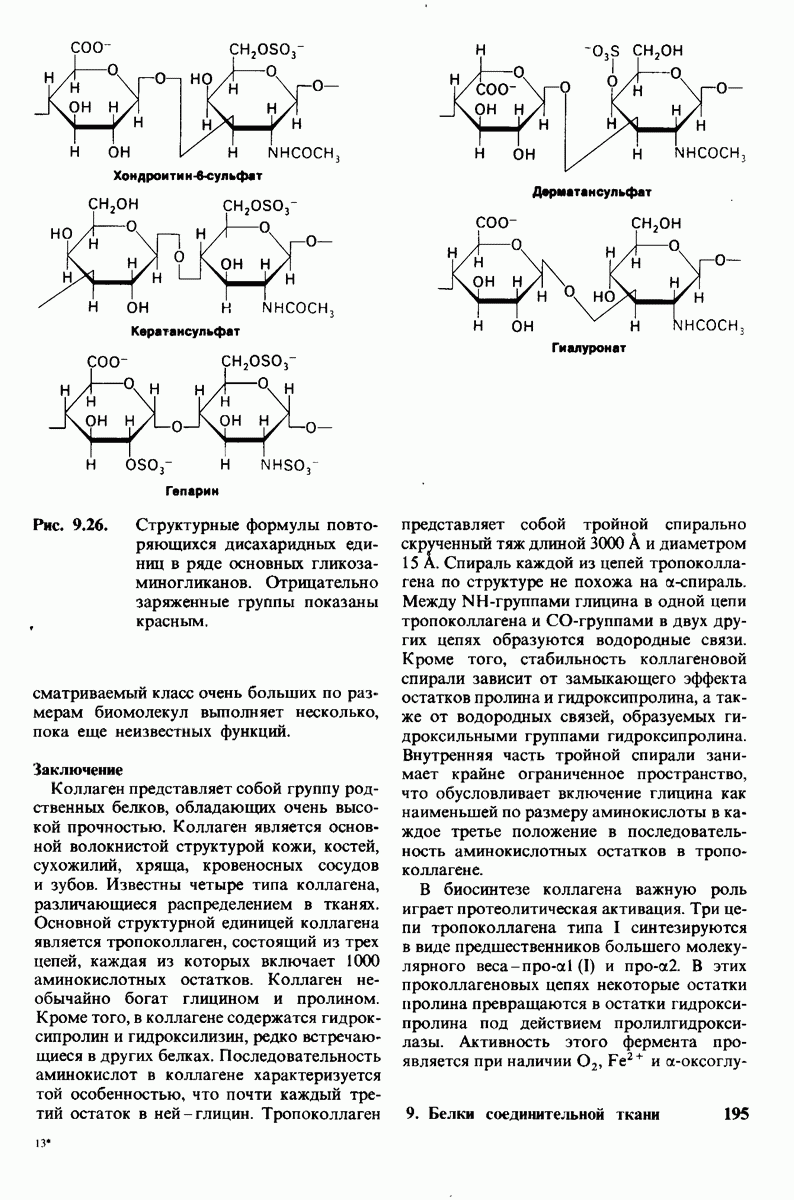
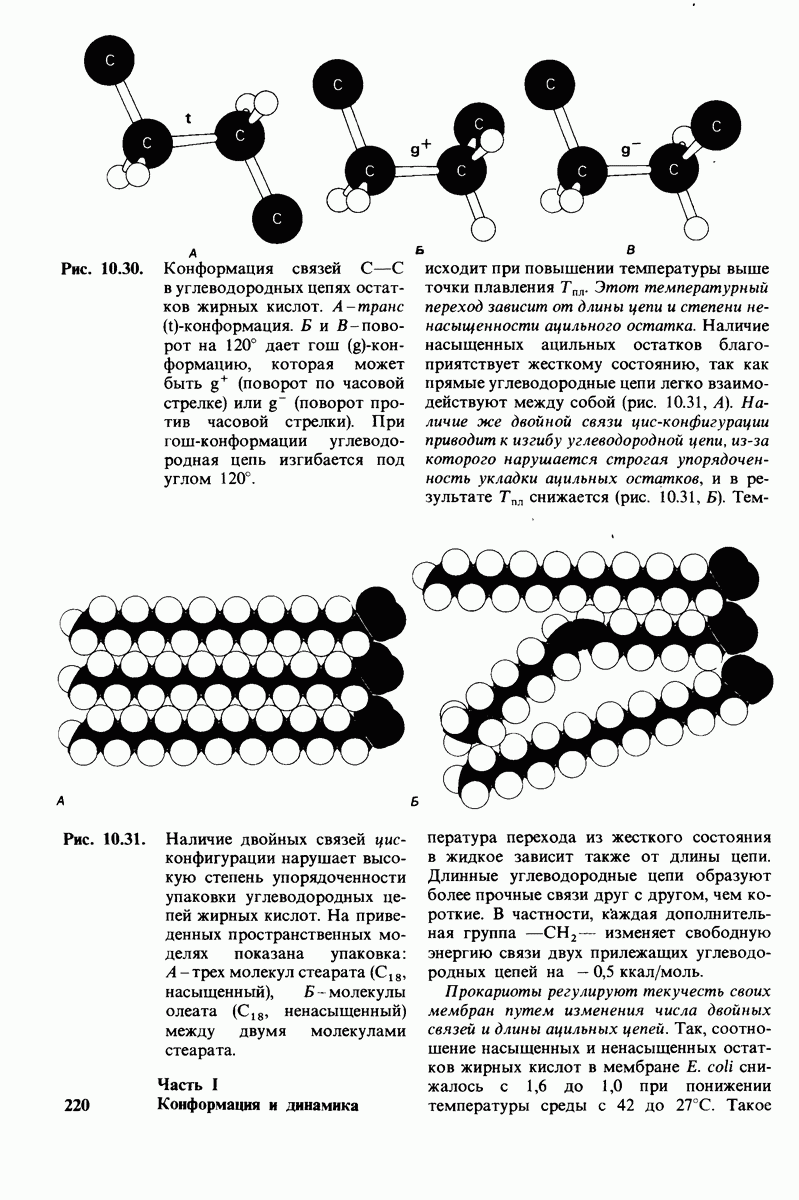
Основные классы мембранных липидов - это фосфолипиды, гликолипиды и холестерол. К классу фосфолипидов относятся фосфоацилглицеролы, построенные из глицеролового скелета, двух цепей жирных кислот и фосфорилированного спирта. Цепи жирных кислот содержат обычно от 14 до 24 атомов углерода и могут быть как насыщенными, так и ненасыщенными. Основные фосфоацилглицеролы - фосфатидилхолин, фосфатидилсерин и фосфатидилэтаноламин. К фосфолипидам другого типа принадлежит сфингсмиелин, основу скелета которого составляет не глицерол, а сфингозин. Глико липиды представлены углеводсодержащими липидами - производными сфингозина. Все мембранные липиды - амфипатические соединения. В водных растворах они спонтанно образуют обширный бимолекулярный слой, состоящий из гидрофильной и гидрофобной фаз. Липидные бислои характеризуются крайне низкой проницаемостью для ионов и большинства полярных соединений; в то же время бислой жидкий, что позволяет ему служит растворителем для мембранных белков.
Отдельные функции мембран, такие, как транспорт, коммуникация, преобразование энергии, выполняются специфическими белками. Некоторые мембранные белки погружены в глубь углеводородной части липидного бислоя. Трансмембранные белки, в частности белок полосы 3 из мембраны эритроцитов, могут служить ионными каналами. Мембраны ассимметричны структурно и функционально; это проявляется как в направленности действия систем транспорта ионов, так и в локализации углеводных остатков только на наружной стороне плазматических мембран клеток млекопитающих. Мембраны - динамичные структуры, и в отсутствие специальных ограничений составляющие их белки и липиды быстро диффундируют в плоскости мембраны (латеральная диффузия). Однако переход белков и липидов с одной стороны мембраны на другую (поперечная диффузия, «flip-flop» - nepecкок) происходит крайне медленно. Степень текучести мембран частично зависит от длины цепей и степени ненасыщенности составляющих их жирных кислот.