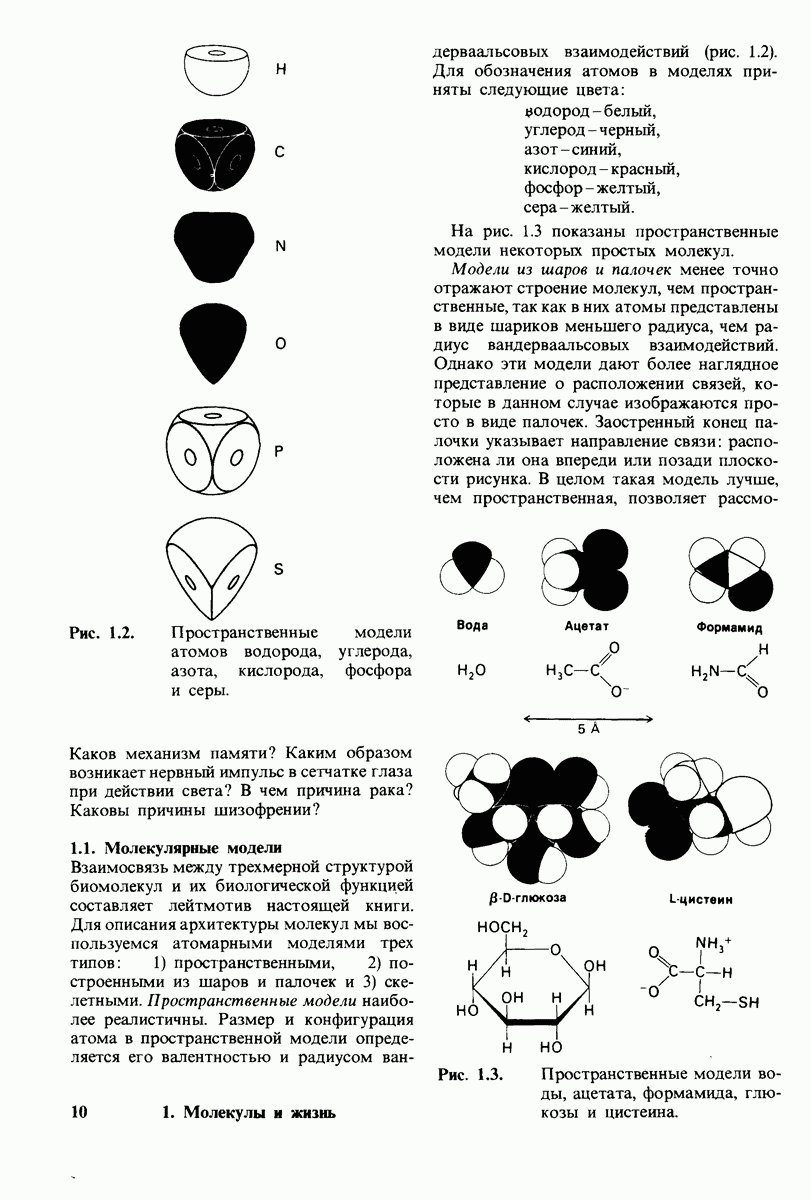
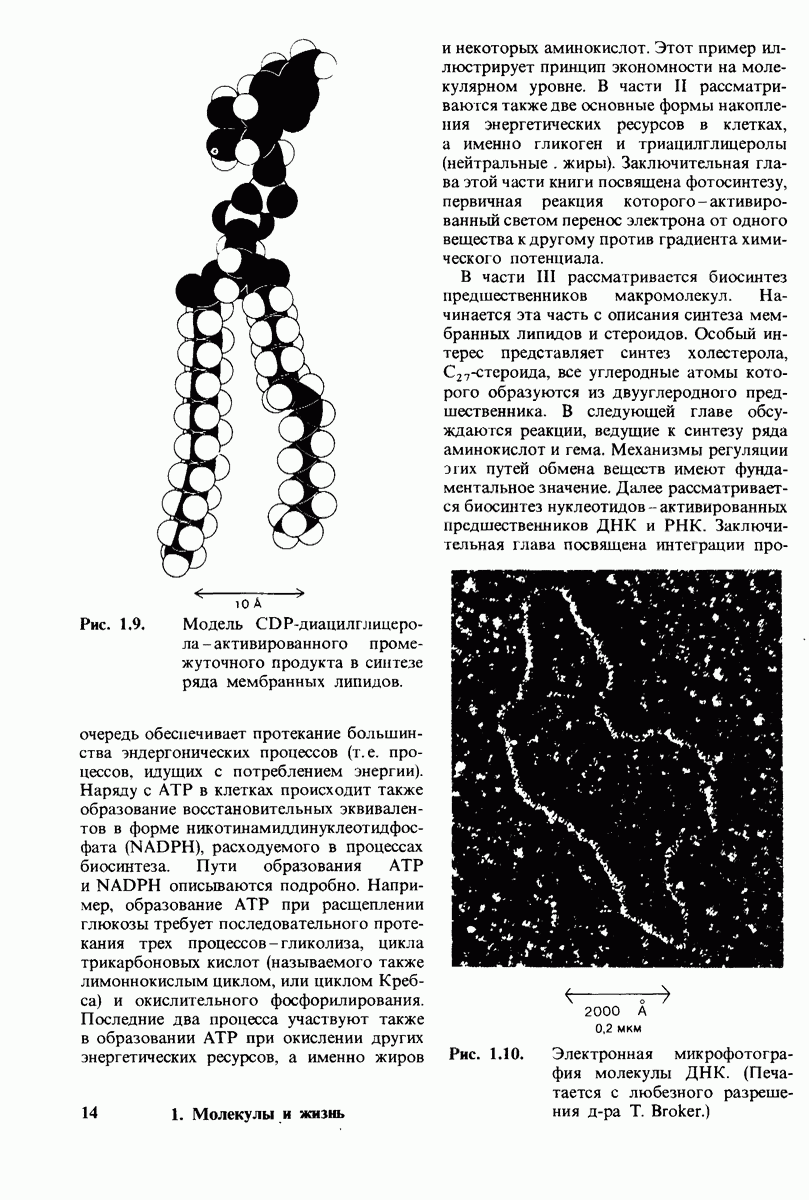
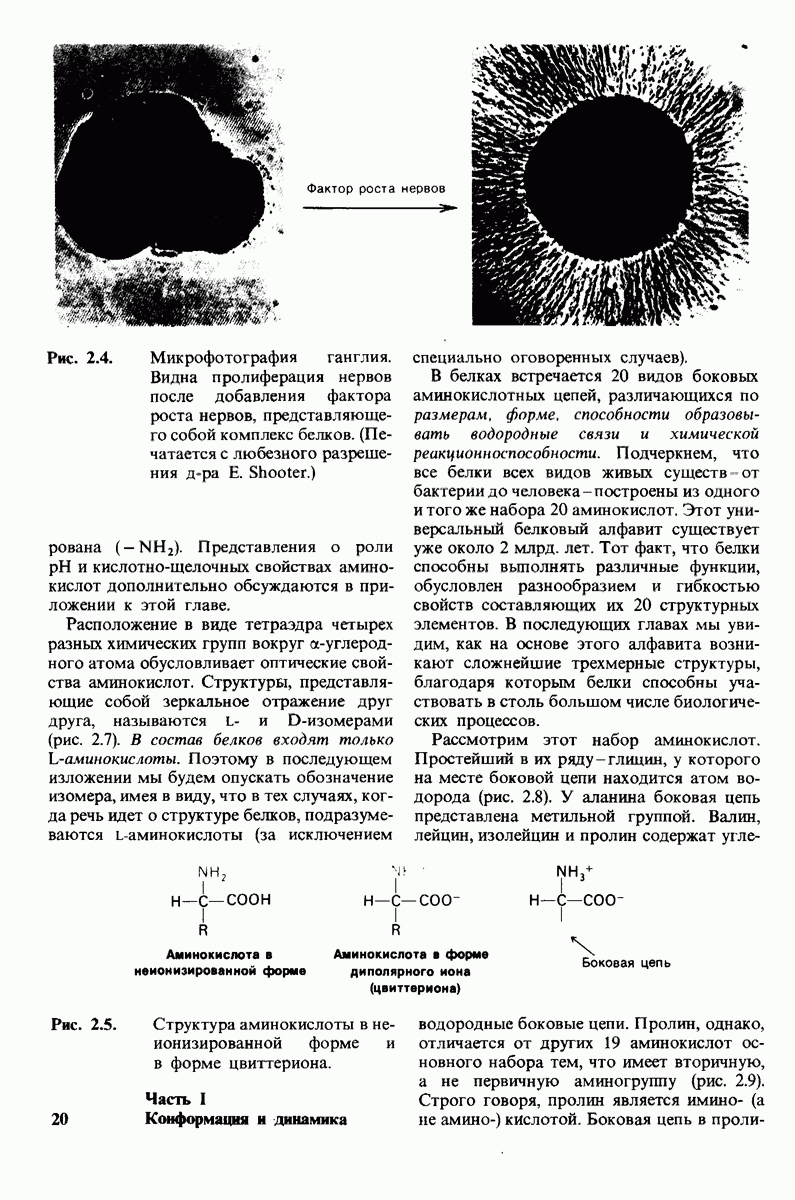
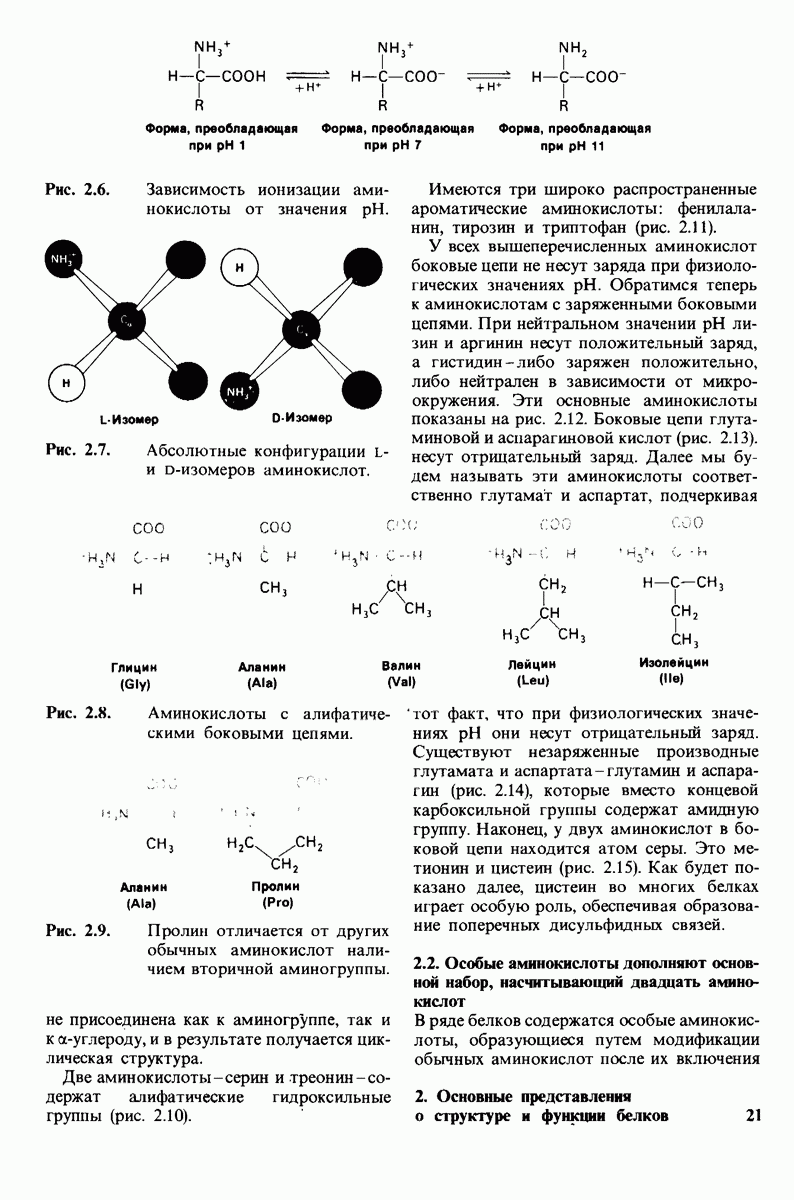
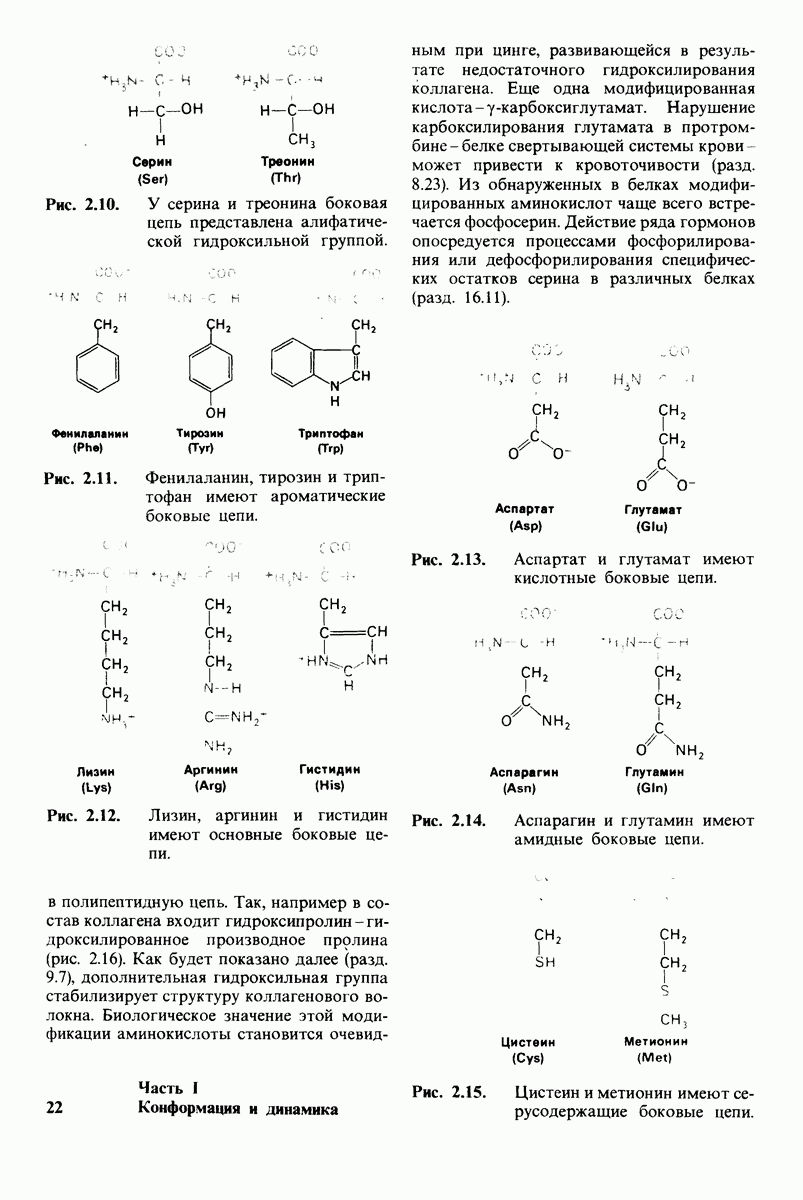
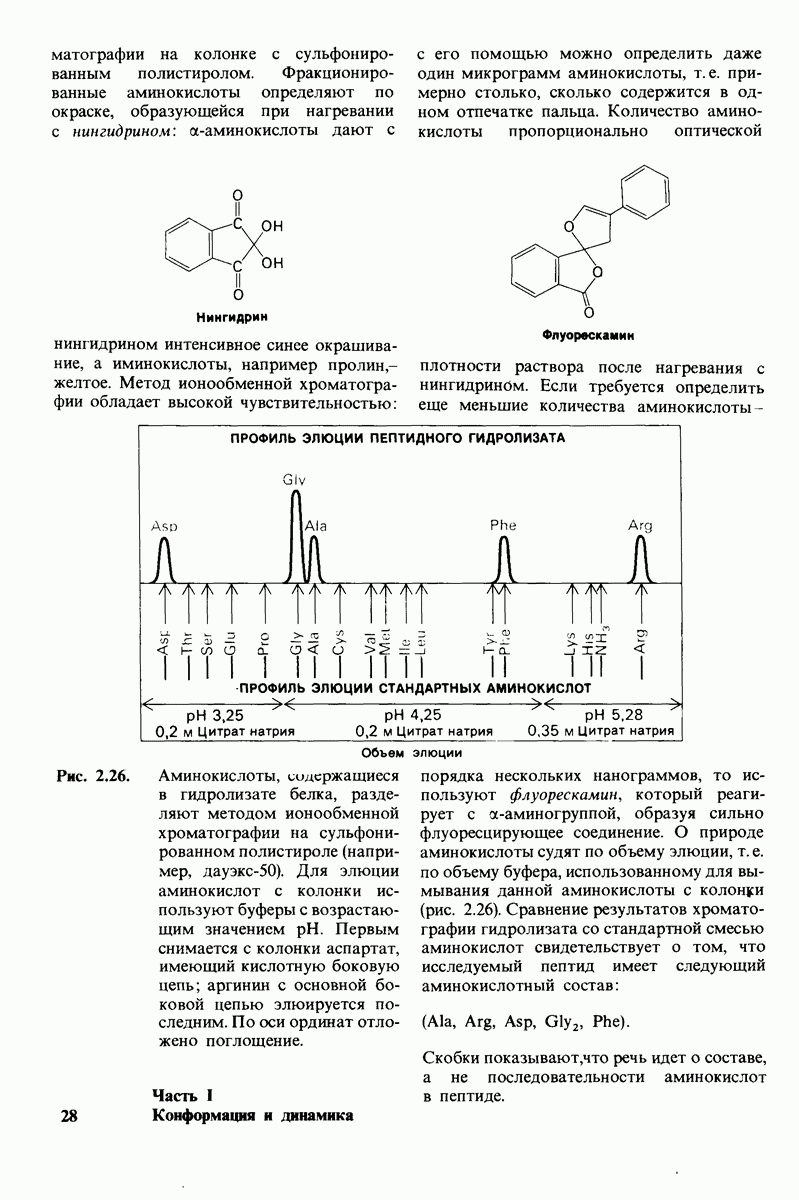
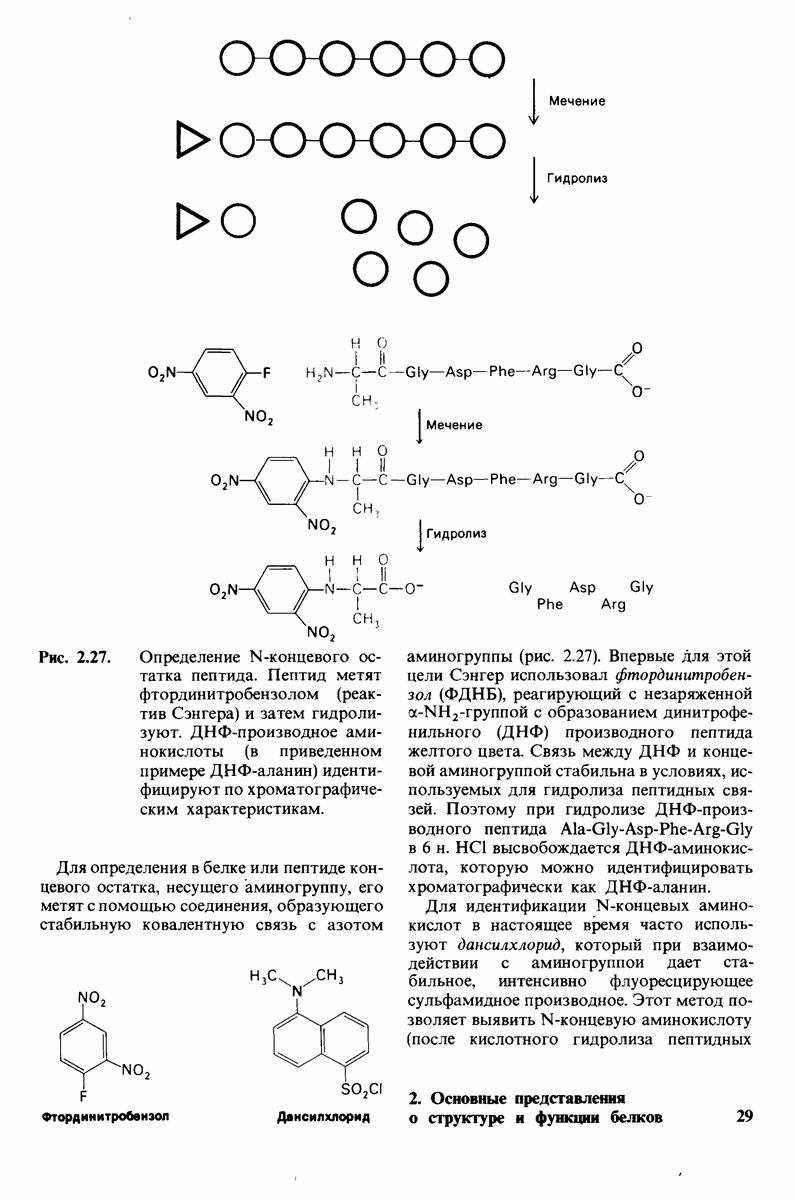
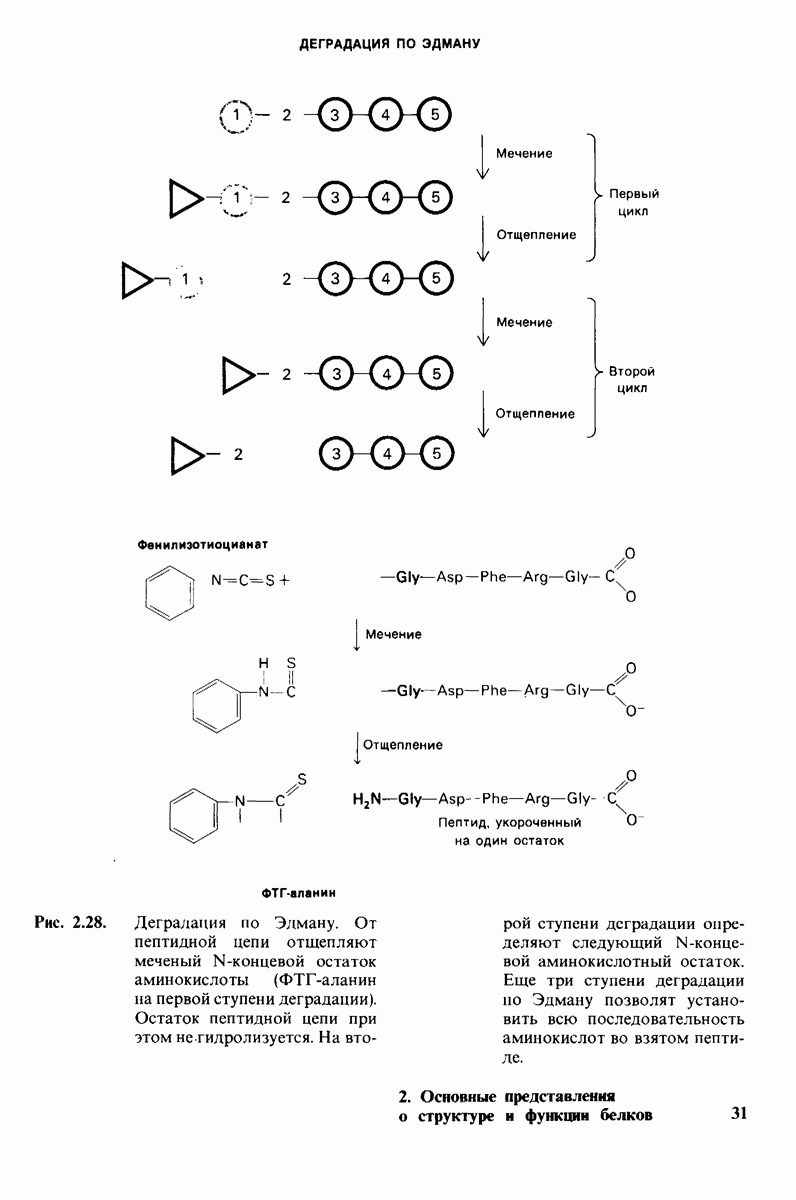
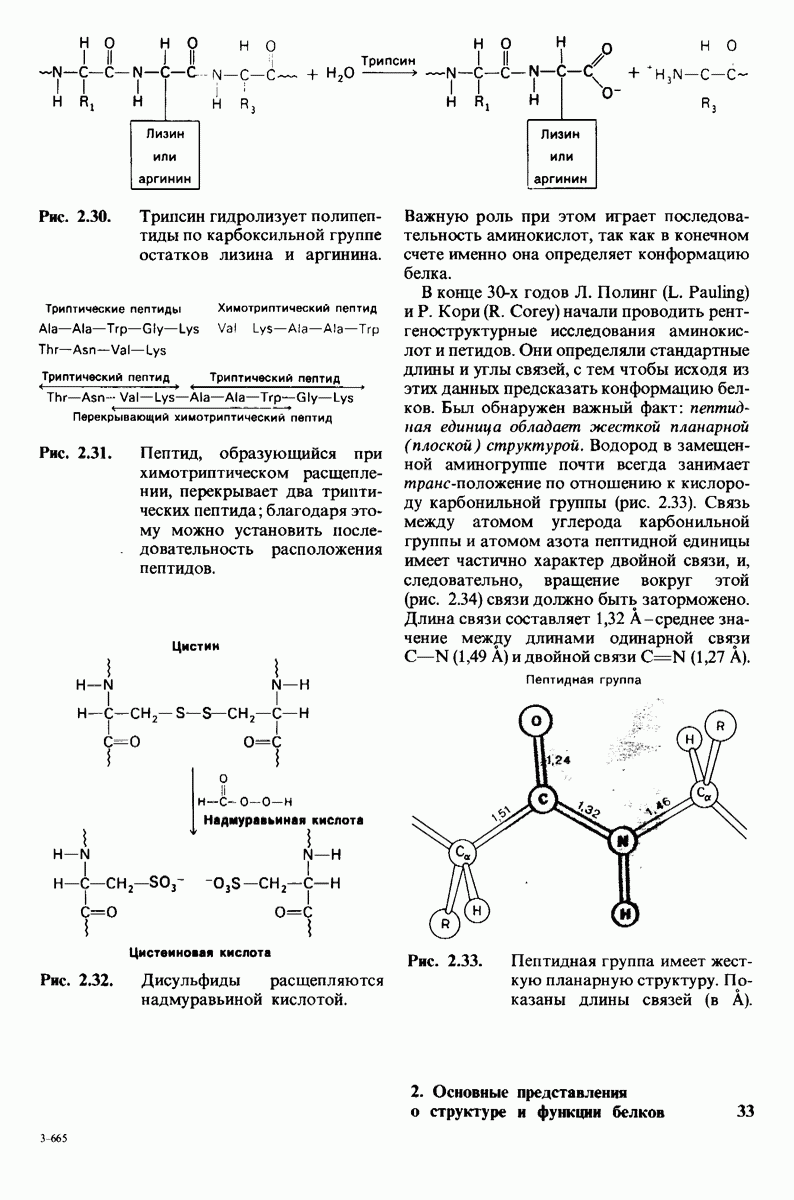
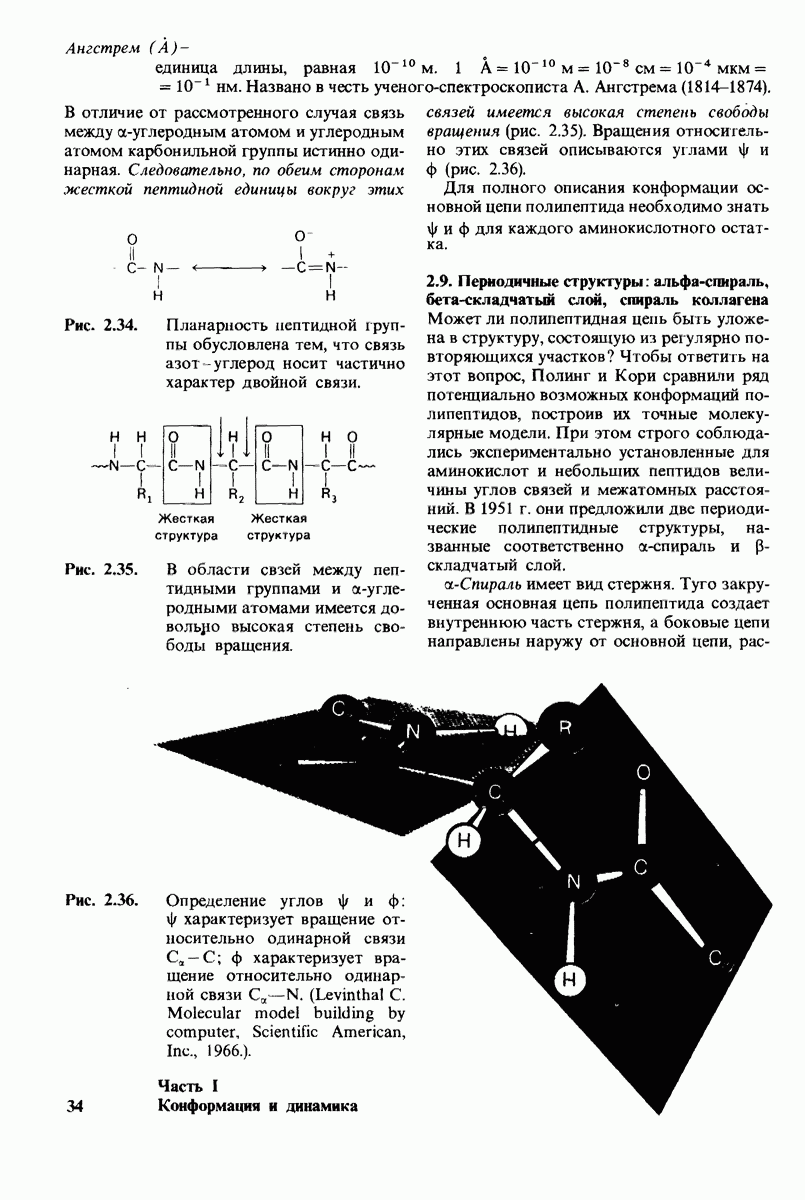
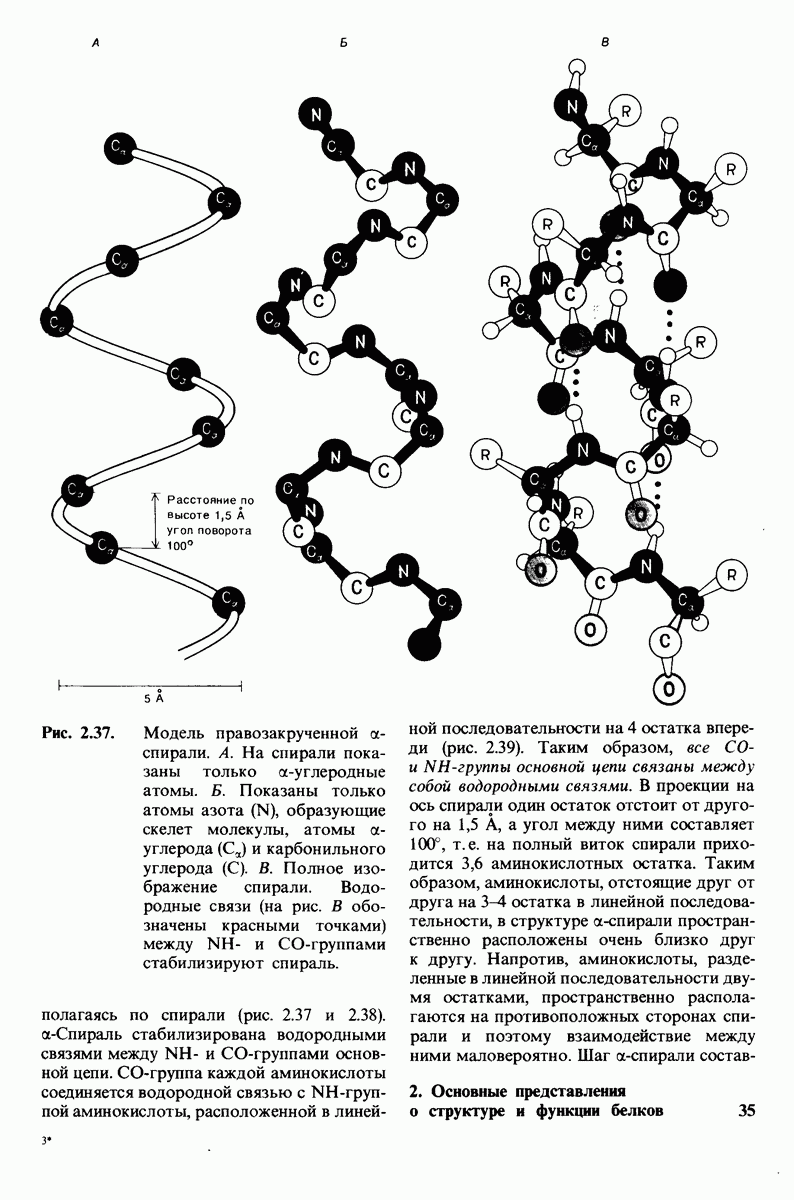
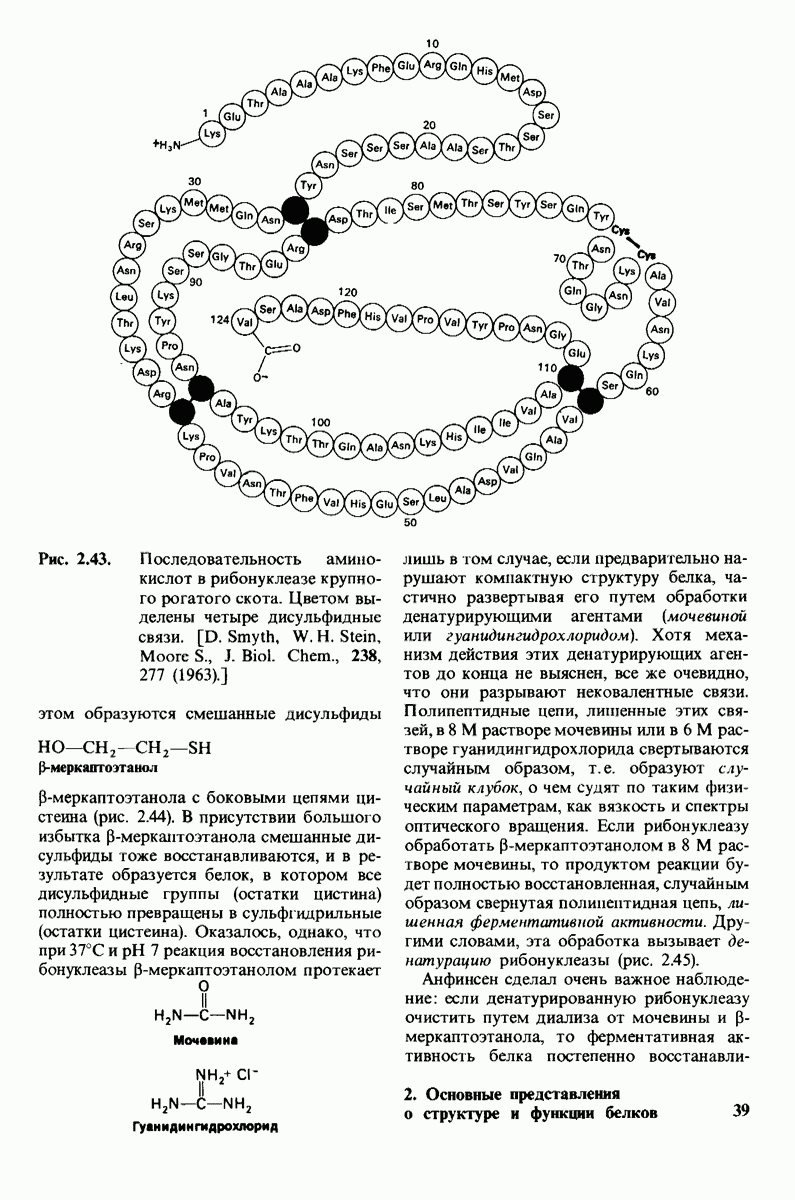
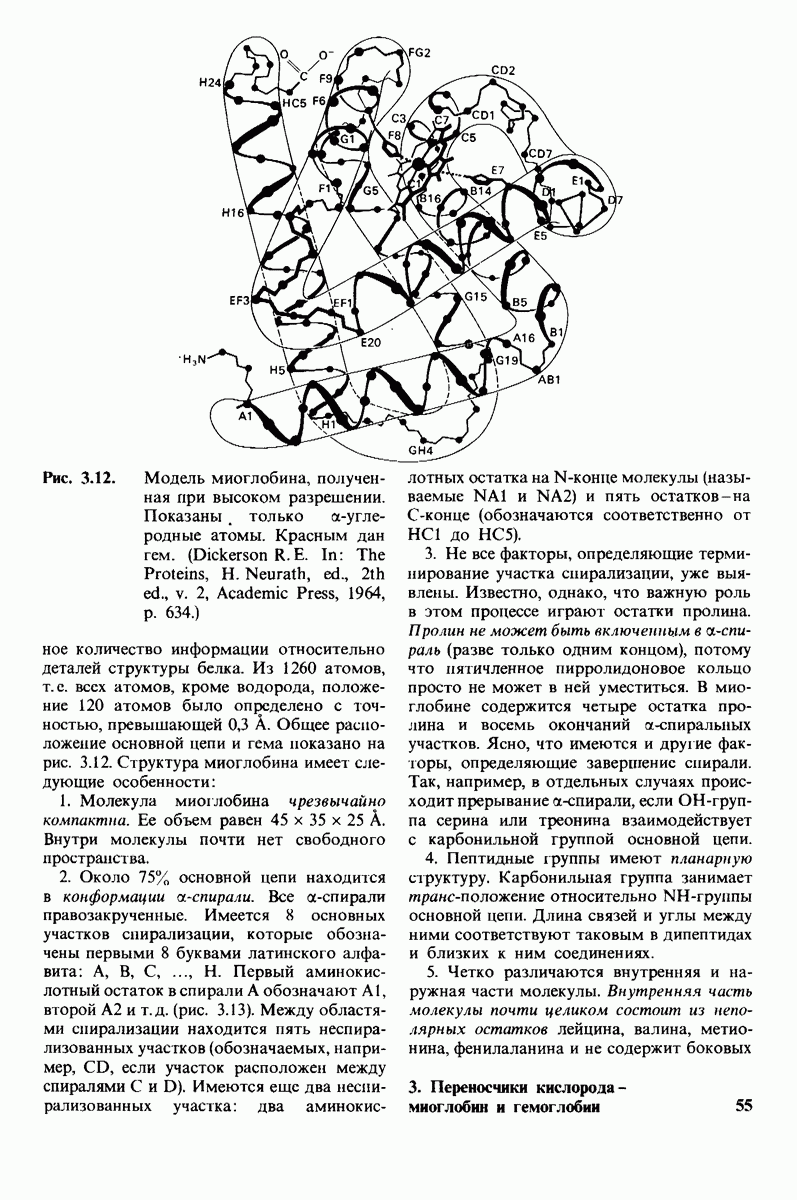
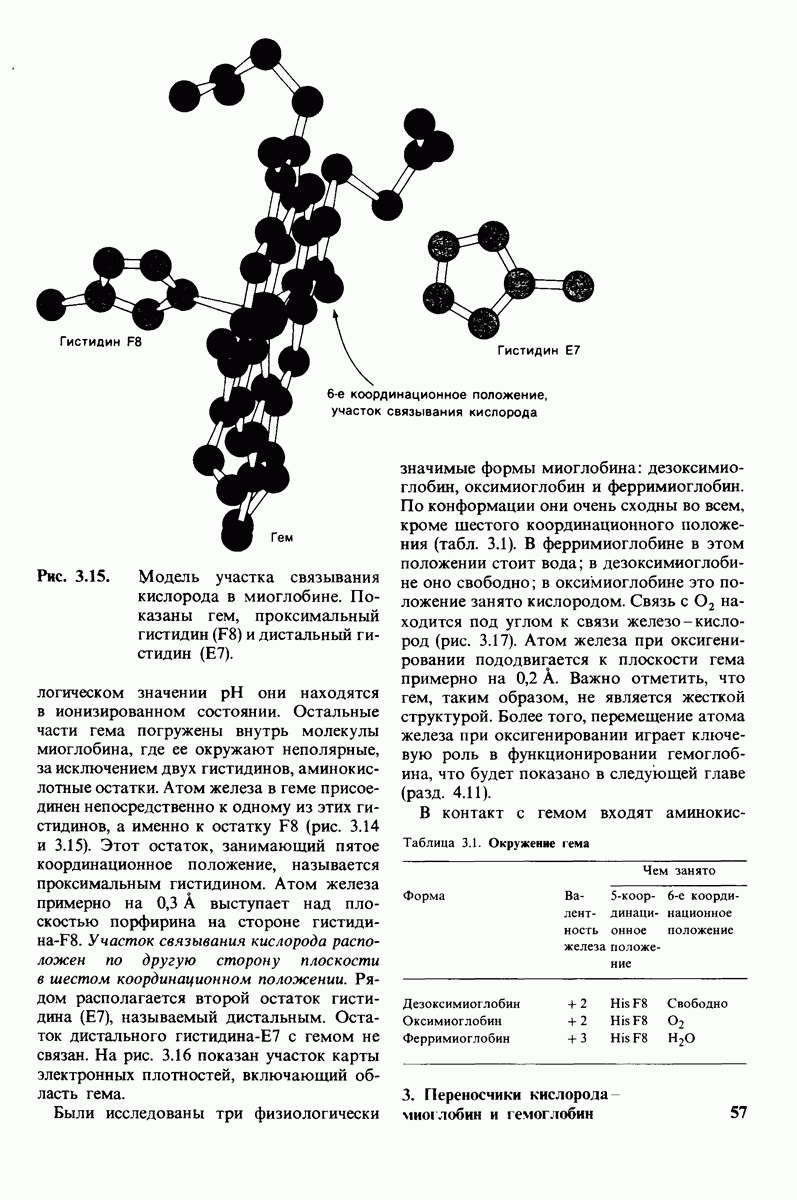
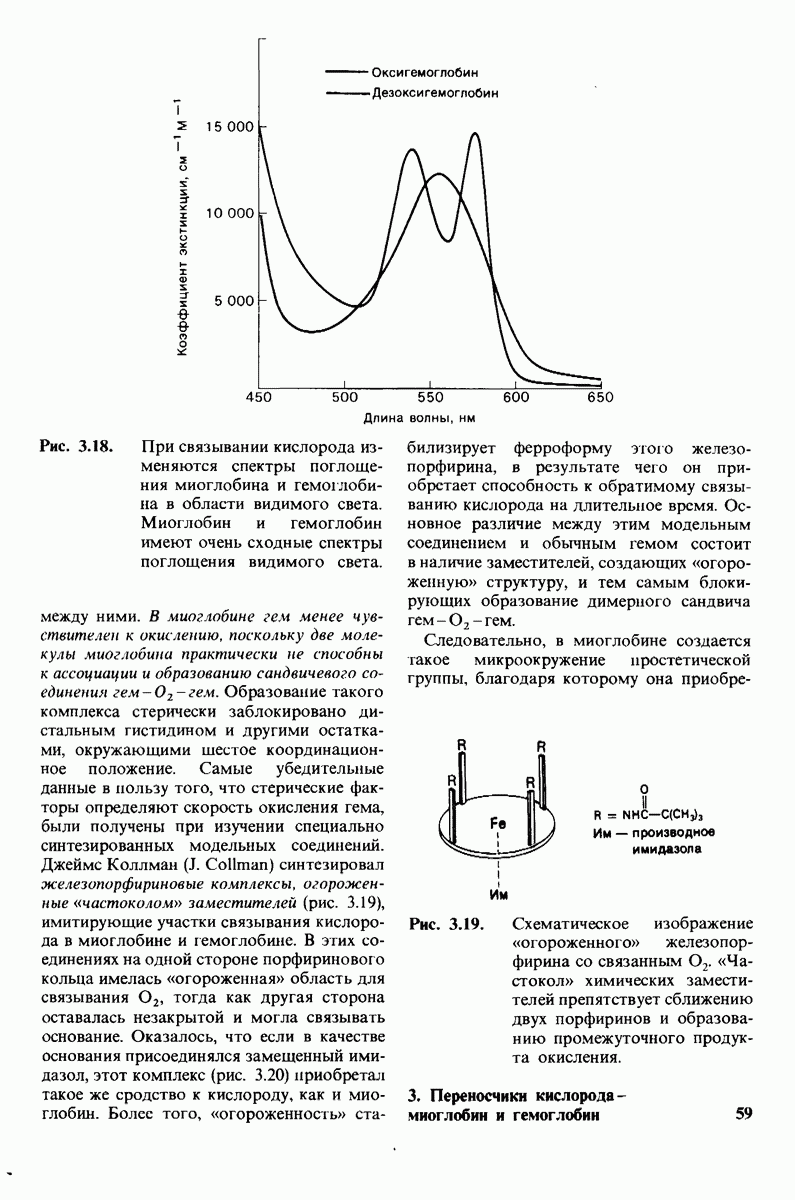
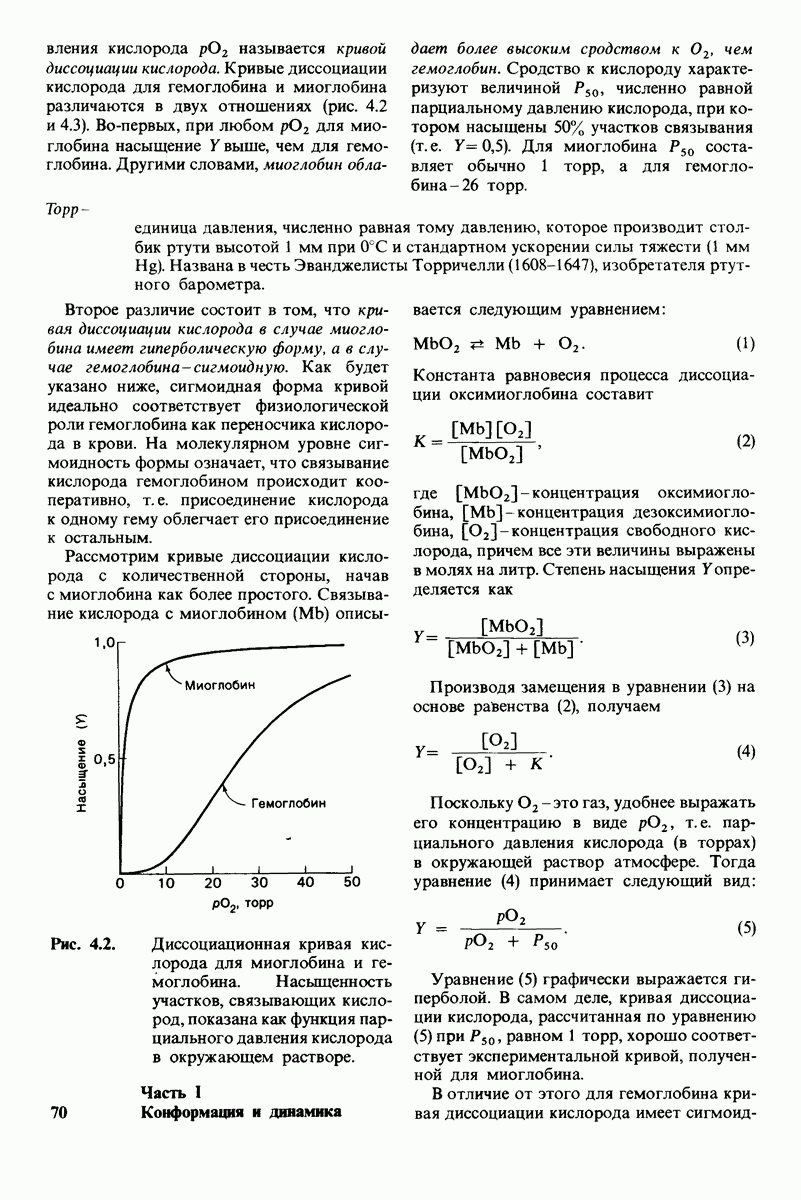
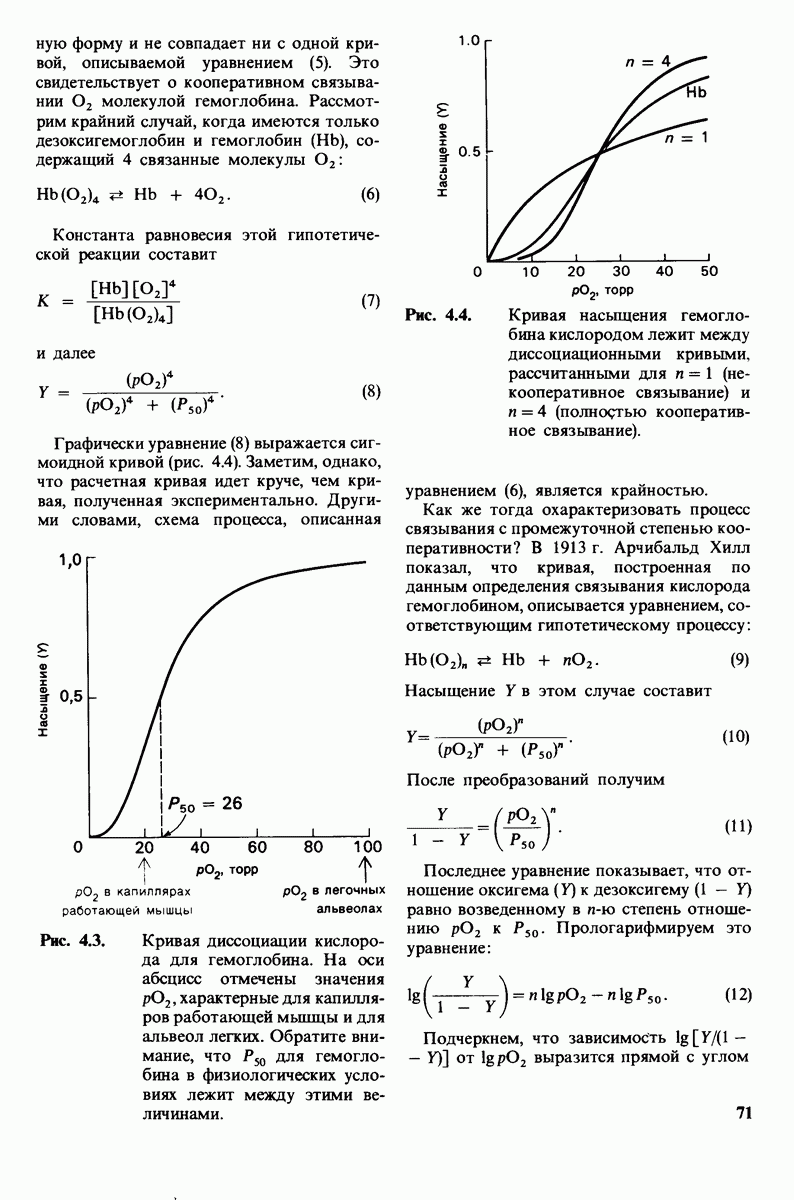
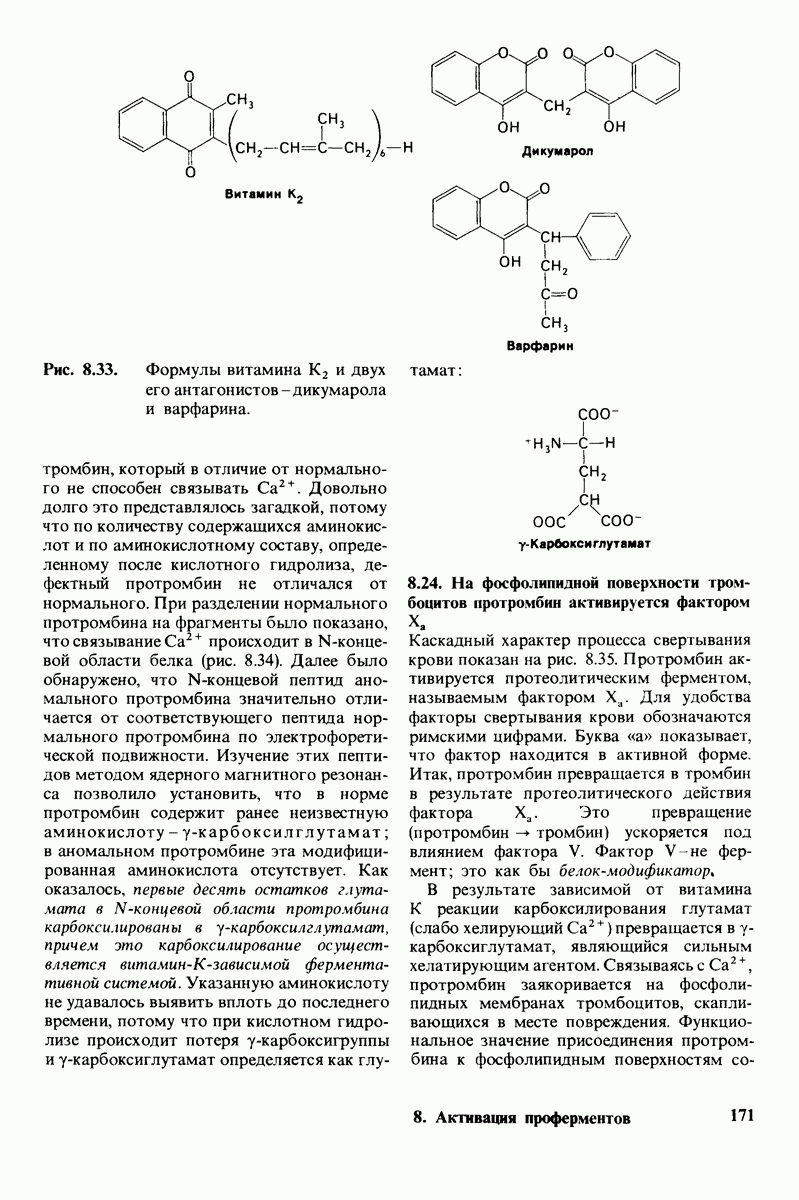
Распознанный текст, спецсимволы и формулы могут содержать ошибки, поэтому с корректным вариантом рекомендуем ознакомиться на отсканированных изображениях учебника выше
Также, советуем воспользоваться поиском по сайту, мы уверены, что вы сможете найти больше информации по нужной Вам тематике
9.16. Протеогликаны образуют основное вещество соединительной ткани
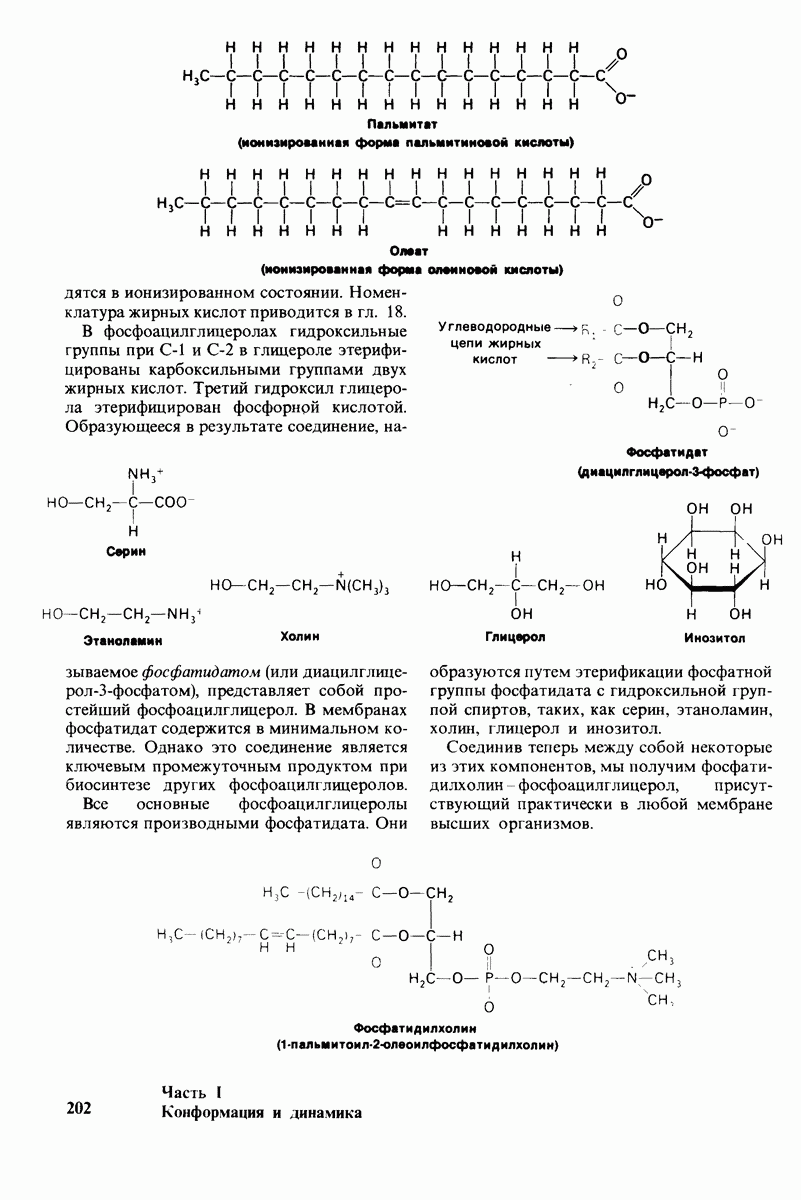
Соединительная ткань богата также протеогликанами, которые состоят из полисахаридных (около 95%) и белковых (около 5%) единиц. Эти полианионы, имеющие очень большие размеры, связывают воду и катионы, образуя основное вещество соединительной ткани. Протеогликаны определяют вязкоэластические свойства суставов и других анатомических структур, подвергающихся механической деформации.

Рис. 9.21. При метаморфозе головастика из хвостового плавника быстро исчезает коллаген. (Lapiere С, М., Gross J. In: Mechanisms of Hard Tissue Destruction, R.F. Sognnaes, ed., p. 665, Copyright 1963.)

Рис. 9.22. Кусочки хвостового плавника головастика помещены на реконструированный коллагеновый гель (наверху). После 24 ч инкубации при 37 °С вокруг каждого кусочка появляется прозрачная зона - результат активности коллагеназы (внизу). (Lapiere С. М., Gross J. In: Mechanisms of Hard Tissue Destruction, R.F. Sognnaes, ed., p. 681, Copyright 1963.)
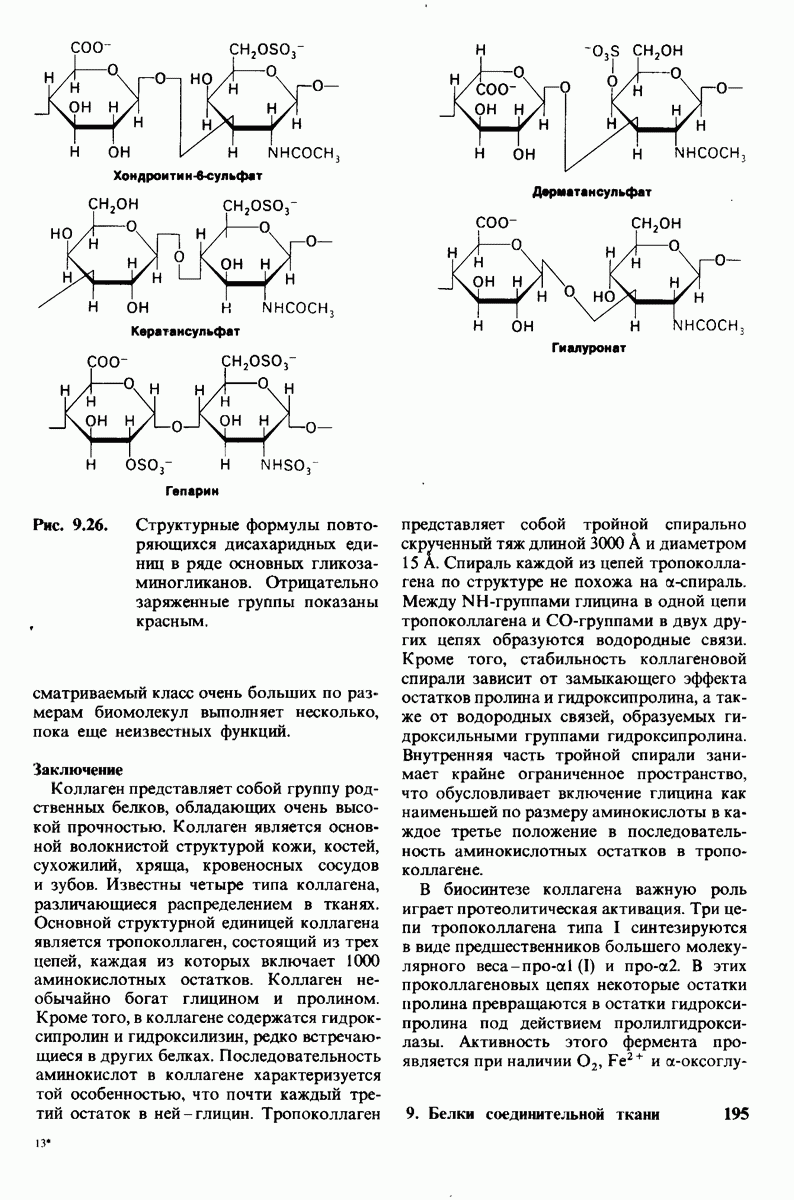
Полисахаридные цепи протеогликанов, называемые гликозаминогликанами, построены из повторяющихся дисахаридных единиц, содержащих производное аминосахара- глюкозамина или галактозамина. По крайней мере один из Сахаров в дисахариде имеет отрицательно заряженную карбоксильную или сульфатную группу. Важнейшие глико-заминогликаны - это гиалуронат, хондрои-тинсульфат, кератансульфат, гепарансульфат и гепарин (рис. 9.26). Гепарансульфат подобен гепарину, за исключением того, что в нем меньше N- и О-сульфатных групп и больше N-ацетильных групп.
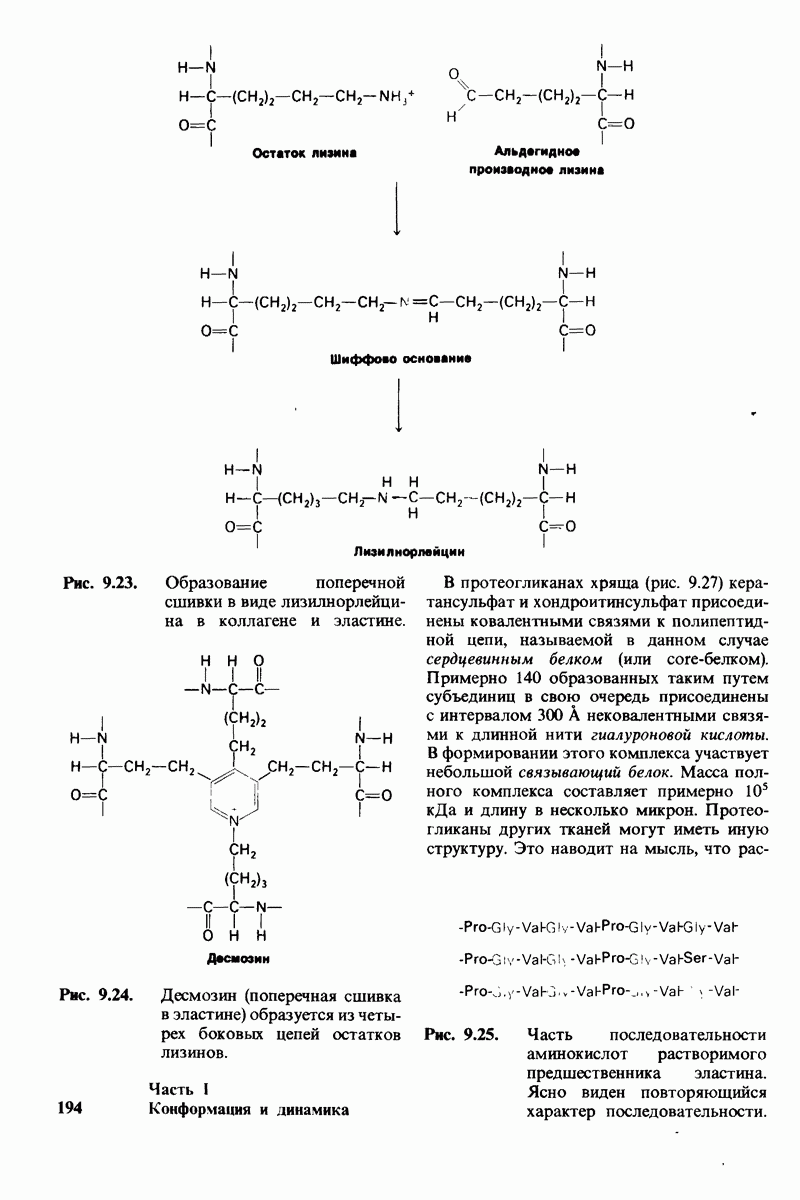
Рис. 9.23. (см. скан) Образование поперечной сшивки в виде лизилнорлейцина в коллагене и эластине.

Рис. 24.27. Десмозин (поперечная сшивка в эластине) образуется из четырех боковых цепей остатков лизинов.
В протеогликанах хряща (рис. 9.27) кератансульфат и хондроитинсульфат присоединены ковалентными связями к полипептидной цепи, называемой в данном случае сердцевинным белком (или соге-белком). Примерно 140 образованных таким путем субъединиц в свою очередь присоединены с интервалом 300 А нековалентными связями к длинной нити гиалуроновой кислоты. В формировании этого комплекса участвует небольшой связывающий белок. Масса полного комплекса составляет примерно 105 к Да и длину в несколько микрон. Протеогликаны других тканей могут иметь иную структуру. Это наводит на мысль, что

Рис. 9.25. Часть последовательности аминокислот растворимого предшественника эластина. Ясно виден повторяющийся характер последовательности.

Рис. 9.26, Структурные формулы повторяющихся дисахаридных единиц в раде основных гликозаминогликанов. Отрицательно заряженные группы показаны красным,
рассматриваемый класс очень больших по размерам биомолекул выполняет несколько, пока еще неизвестных функций.
Заключение
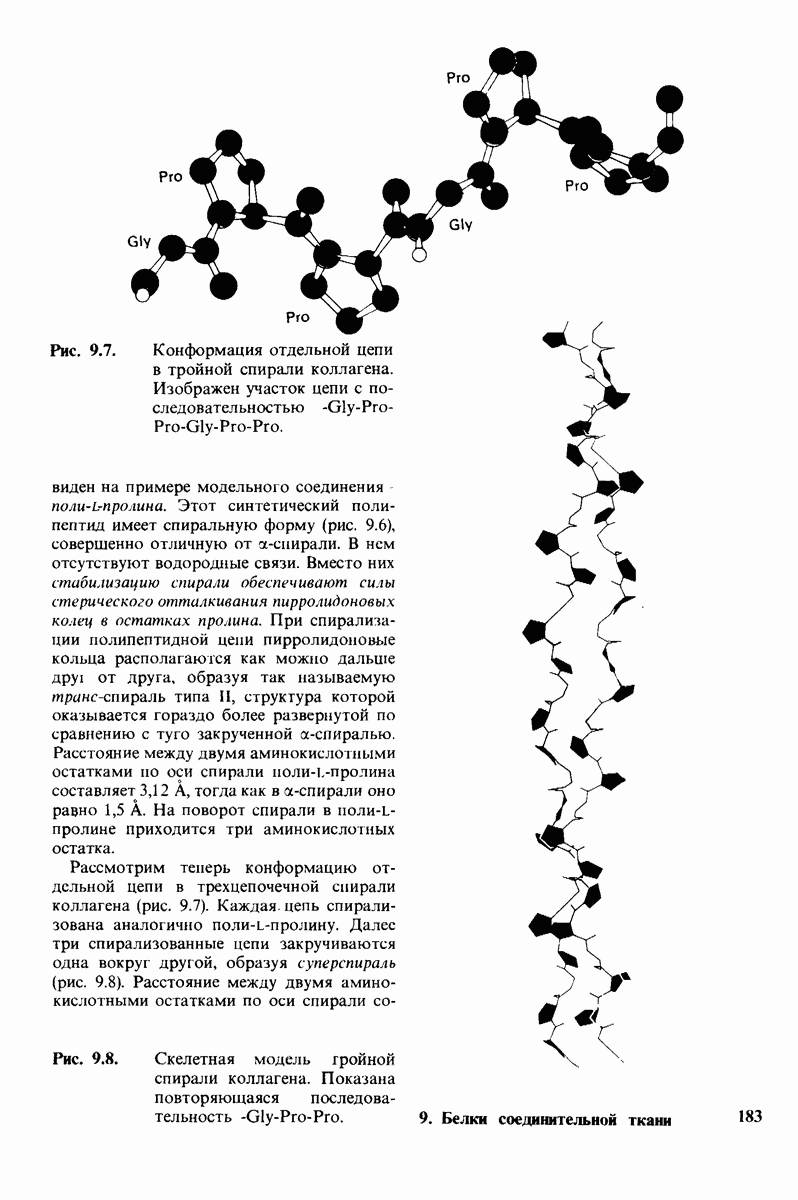
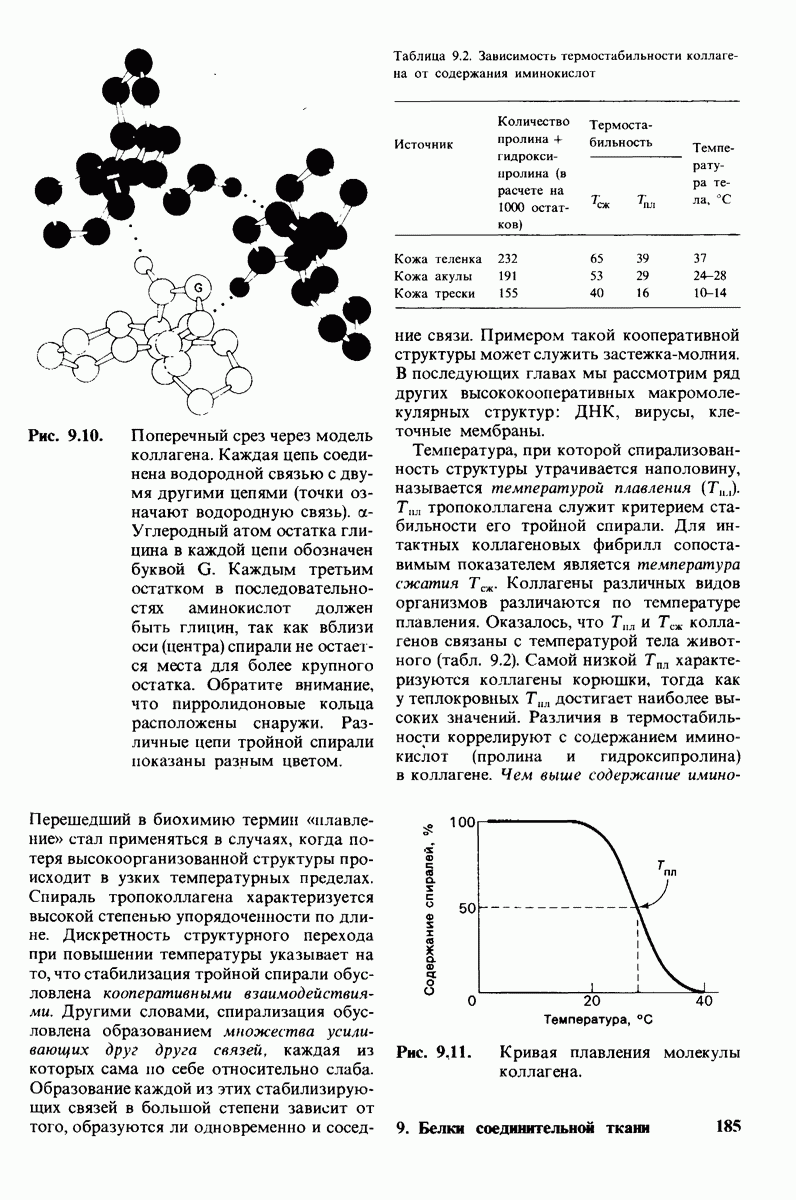
Коллаген представляет собой группу родственных белков, обладающих очень высокой прочностью, Коллаген является основной волокнистой структурой кожи, костей, сухожилий, хряща, кровеносных сосудов и зубов. Известны четыре типа коллагена, различающиеся распределением в тканях. Основной структурной единицей коллагена является тропоколлаген, состоящий из трех цепей, каждая из которых включает 1000 аминокислотных остатков. Коллаген необычайно богат глицином и пролином. Кроме того, в коллагене содержатся гидрок-сипролин и гидроксилизин, редко встречающиеся в других белках. Последовательность аминокислот в коллагене характеризуется той особенностью, что почти каждый третий остаток в ней - глицин. Тропоколлаген представляет собой тройной спирально скрученный тяж длиной 3000 А и диаметром 15 А. Спираль каждой из цепей тропоколлагена по структуре не похожа на  -спираль. Между NH-группами глицина в одной цепи тропоколлагена и СО-группами в двух других цепях образуются водородные связи. Кроме того, стабильность коллагеновой спирали зависит от замыкающего эффекта остатков пролина и гидроксипролина, а также от водородных связей, образуемых гидроксильными группами гидроксипролина. Внутренняя часть тройной спирали занимает крайне ограниченное пространство, что обусловливает включение глицина как наименьшей по размеру аминокислоты в каждое третье положение в последовательность аминокислотных остатков в тропо коллагене.
-спираль. Между NH-группами глицина в одной цепи тропоколлагена и СО-группами в двух других цепях образуются водородные связи. Кроме того, стабильность коллагеновой спирали зависит от замыкающего эффекта остатков пролина и гидроксипролина, а также от водородных связей, образуемых гидроксильными группами гидроксипролина. Внутренняя часть тройной спирали занимает крайне ограниченное пространство, что обусловливает включение глицина как наименьшей по размеру аминокислоты в каждое третье положение в последовательность аминокислотных остатков в тропо коллагене.
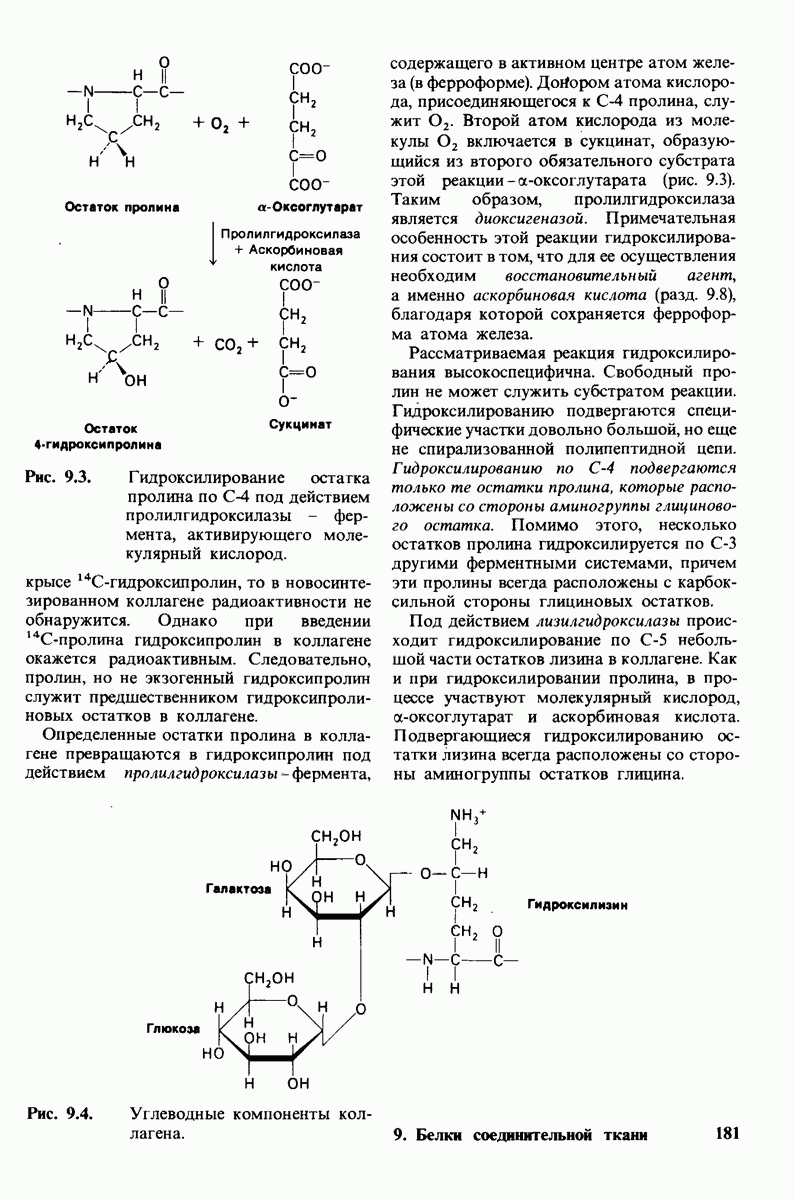
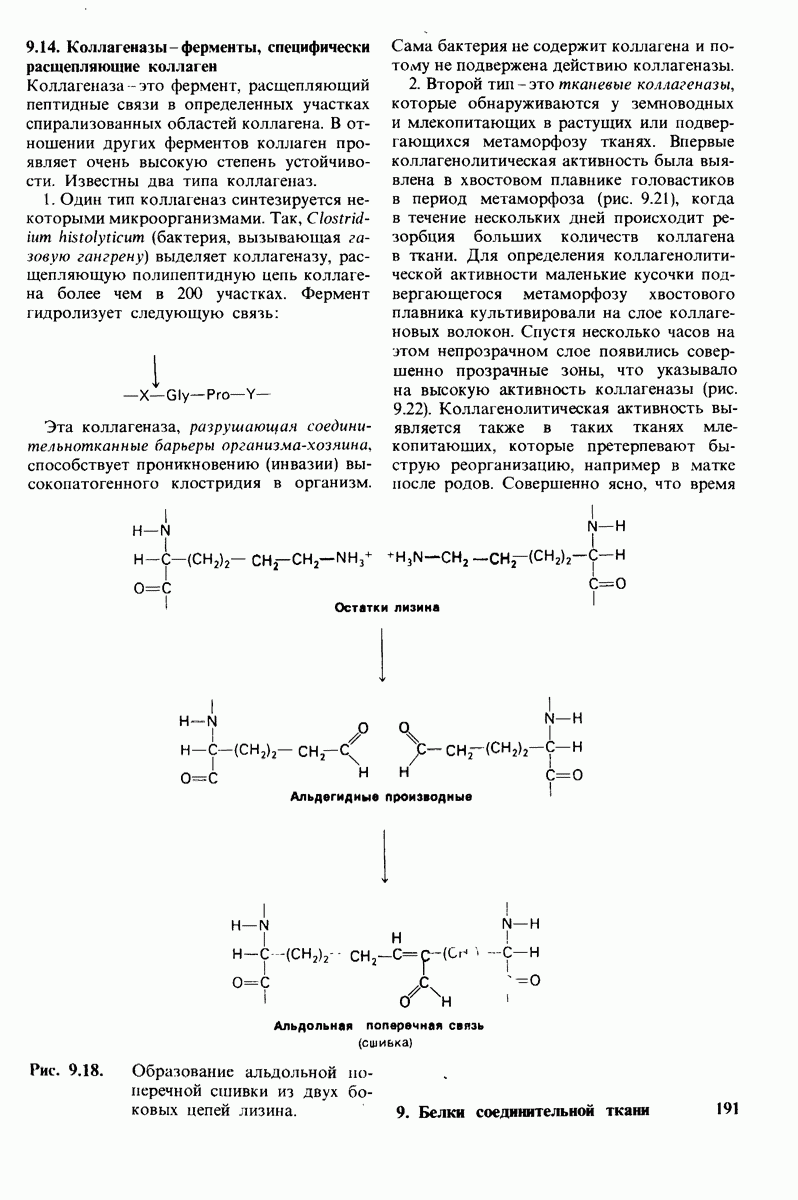
В биосинтезе коллагена важную роль играет протеолитическая активация. Три цепи тропоколлагена типа I синтезируются в виде предшественников большего молекулярного веса про- (I) и про-
(I) и про- . В этих проколлагеновых цепях некоторые остатки пролина превращаются в остатки гидроксипролина под действием пролилгидроксилазы. Активность этого фермента проявляется при наличии
. В этих проколлагеновых цепях некоторые остатки пролина превращаются в остатки гидроксипролина под действием пролилгидроксилазы. Активность этого фермента проявляется при наличии  и
и  -оксоглутарата:
-оксоглутарата:

Рис. 9.27. А - электронная микрофотография протеогликанового агрегата из хряща эпифиза эмбриона коровы. Мономеры протеогликанов располагаются с регулярными интервалами по двум сторонам удлиненной центральной нити гиалуроната. (Печатается с любезного разрешения д-ра J. Buck waiter и д-ра L. Rosenberg.) Б - схематическое изображение. (Rosenberg L. In: Dynamics of Connective Tissue Macromolecules, M. Burleigh, R. Poole, eds., North-Holland, 1975, p. 105.) Заказ 665. стр. 33-38. Голдобина С.
в качестве восстанавливающего агента необходима также аскорбиновая кислота. Синтезированный в отсутствие аскорбиновой кислоты коллаген непрочен, так как он недостаточно гидроксилирован; такая ситуация характерна для цинги. Под действием другого фермента происходит гидроксилирование лизина. Далее к остаткам гидроксилизина цепей-предшественников присоединяются сахара. Процессы гидроксилирования и гликозилирования протекают в фибробластах, после этого прокол-лагены секретируются во внеклеточную среду. Здесь амино- и карбоксиконцевые области проколлагеновых цепей отщепляются проколлаген-пептидазами, образуя тропоколлаген, который спонтанно ассоциируется в фибриллы. Основой структуры коллагеновых волокон являются параллельные ряды молекул тропоколлагена, расположенные так, что один ряд сдвинут по отношению к другому на 1/4 длины молекулы. Последний этап созревания коллагенового волокна-формирование поперечных связей, придающих дополнительную прочность. Примером могут служить альдольные поперечные связи, возникающие при конденсации альдегидов, образованных путем окисления боковых цепей ряда остатков лизина.
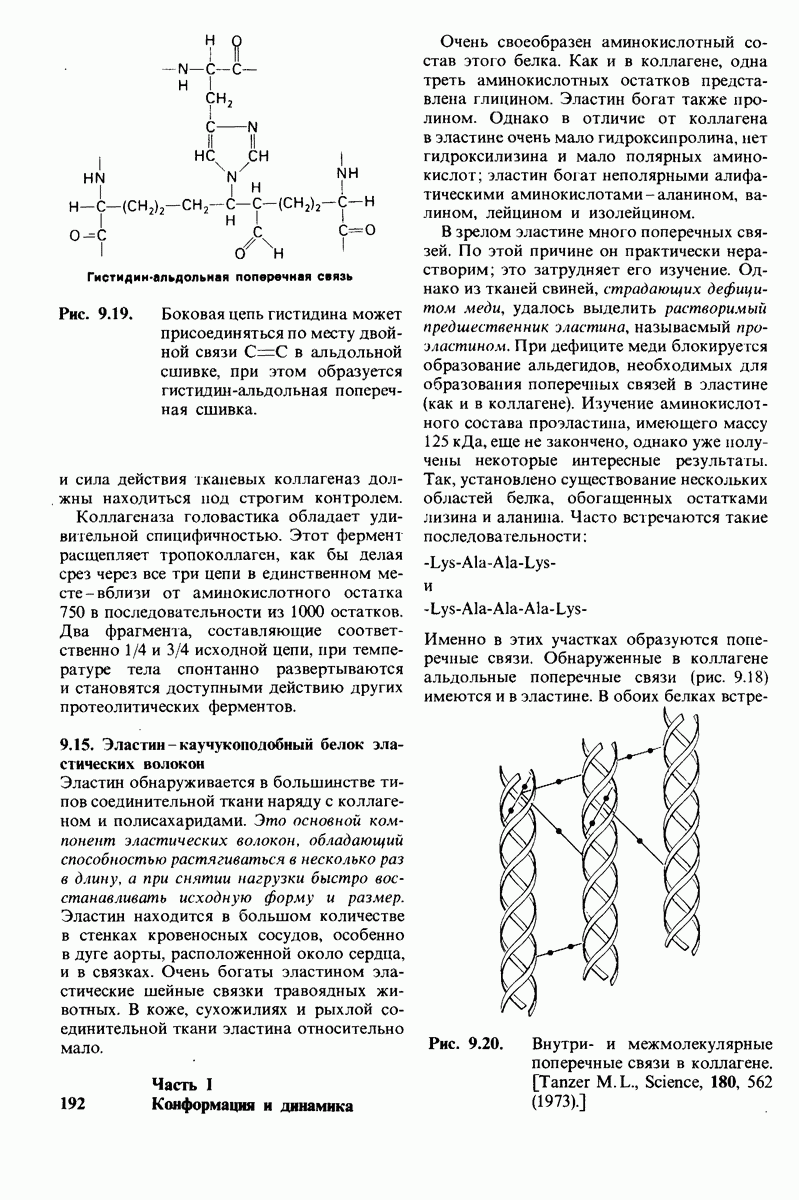
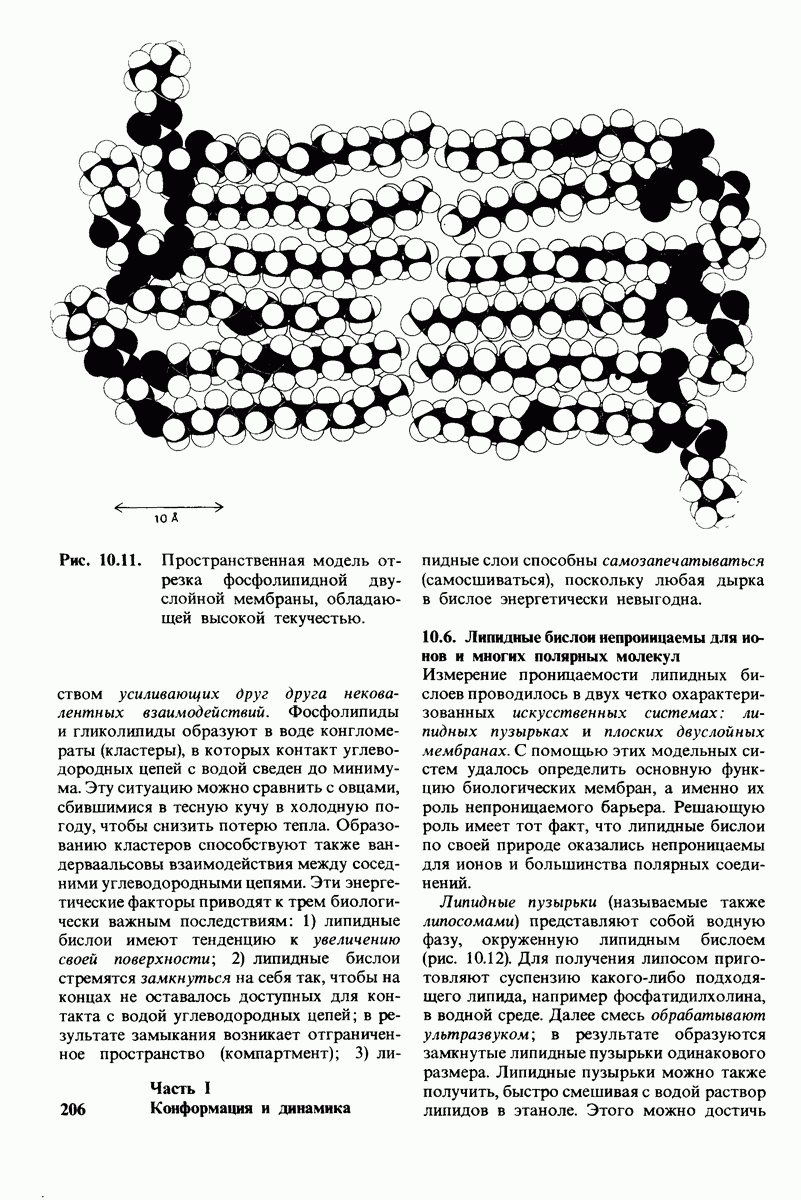
Эластин - это нерастворимый, похожий на резину белок эластических волокон соединительной ткани. Он способен к обратимому растяжению в длину в несколько раз. Такие виды соединительной ткани, как связки и дуга аорты, особенно богаты эластином. Как и в коллагене, в эластине содержится много пролина и глицина; в отличие от коллагена в эластине нет гидроксилизина и мало гидроксипролина. Аминокислотный состав эластина характеризуется выраженным преобладанием неполярных аминокислот. Последовательность аминокислот в эластине имеет определенную периодичность. Так, часто повторяется последовательность  Эластин синтезируется в виде растворимого предшественника, который далее переходит в
Эластин синтезируется в виде растворимого предшественника, который далее переходит в
нерастворимую форму в результате образования различного рода поперечных сшивок. Одна из таких сшивок представляет собой десмозин - производное четырех остатков лизина. Как и в коллагене, формирование поперечных сшивок в эластине происходит с образованием альдегидов в качестве промежуточных соединений.
Второй основной макромолекулярный компонент соединительной ткани - протеогликаны, состоит из полисахаридного и белкового компонентов. Полисахаридные цепи, называемые гликозаминогликанами, построены из повторяющихся дисахаридных единиц, несущих большой отрицательный заряд. Протеогликаны составляют основное вещество соединительной ткани и определяют ее вязкоэластические свойства.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
(см. скан)
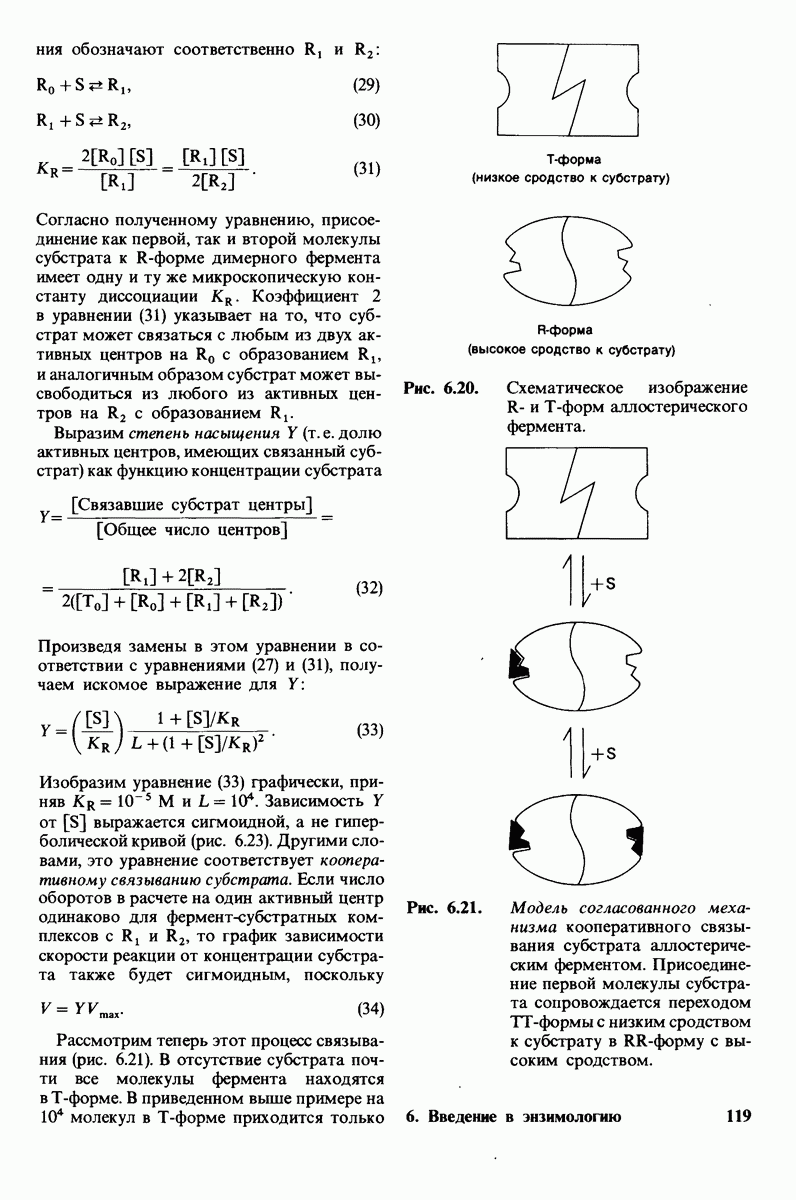
Вопросы и задачи
(см. скан)