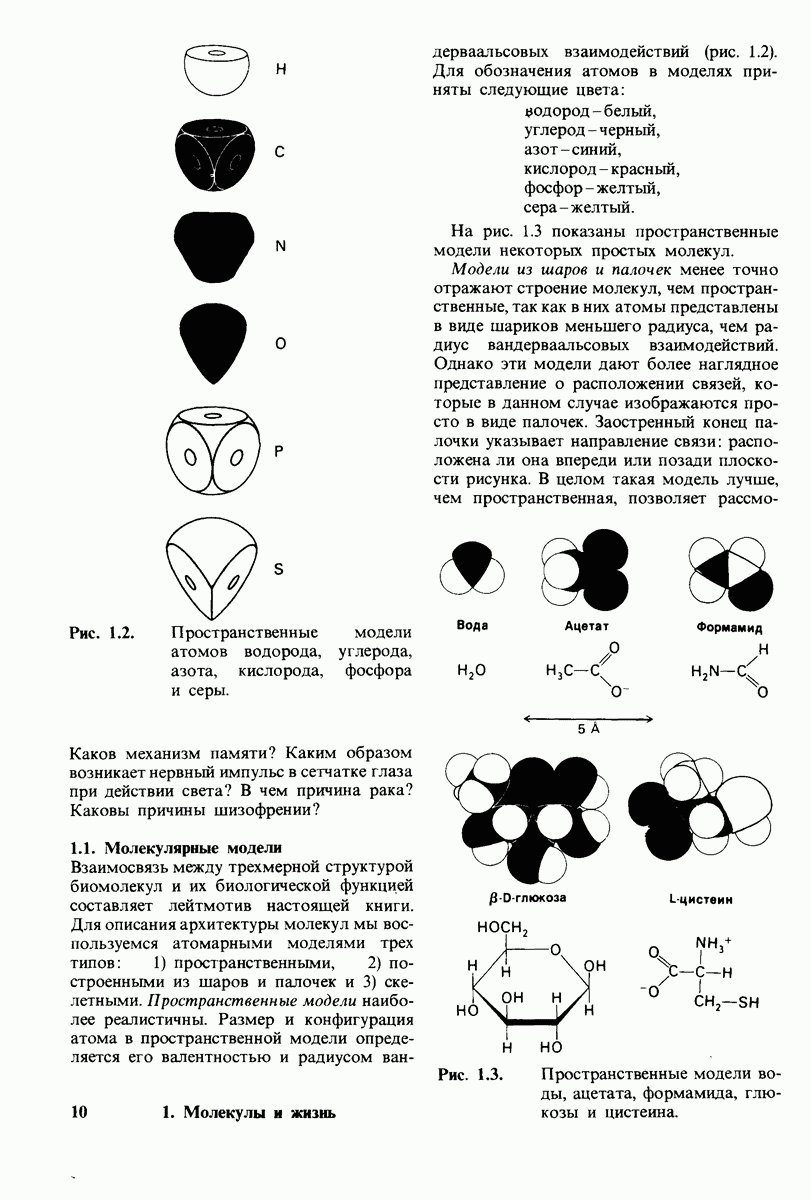
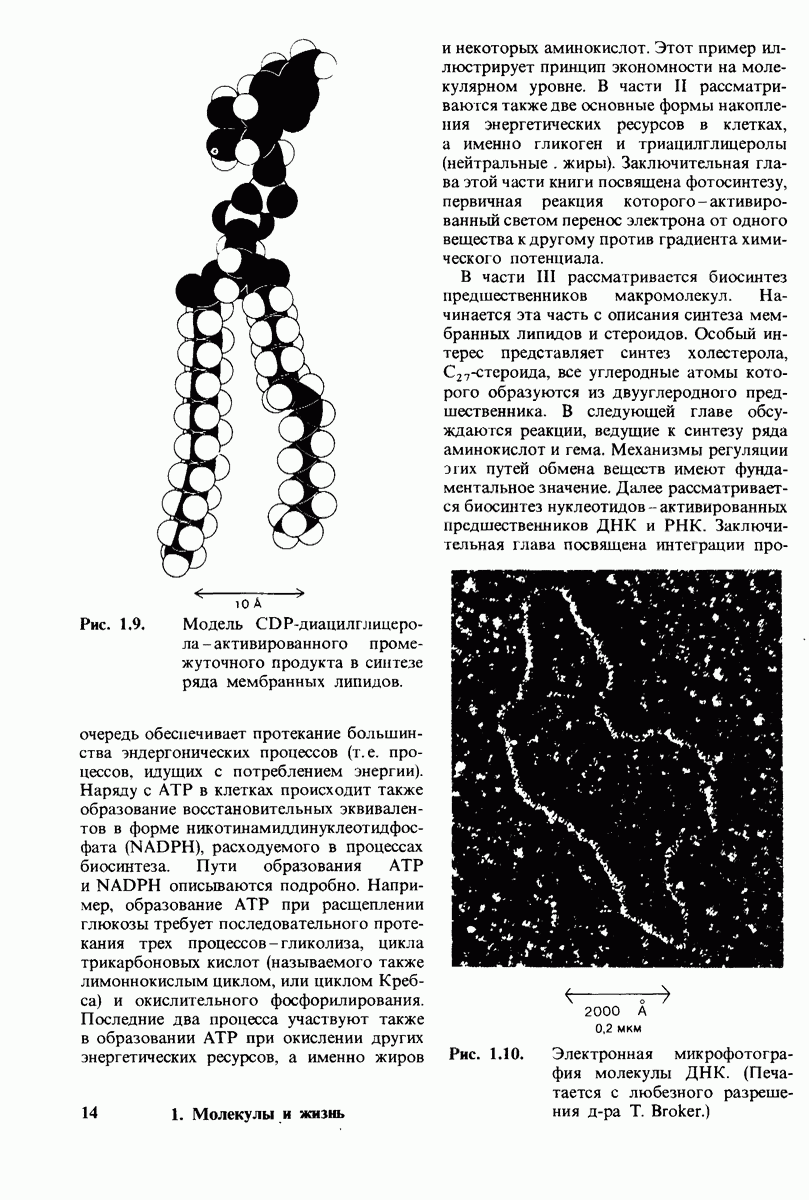
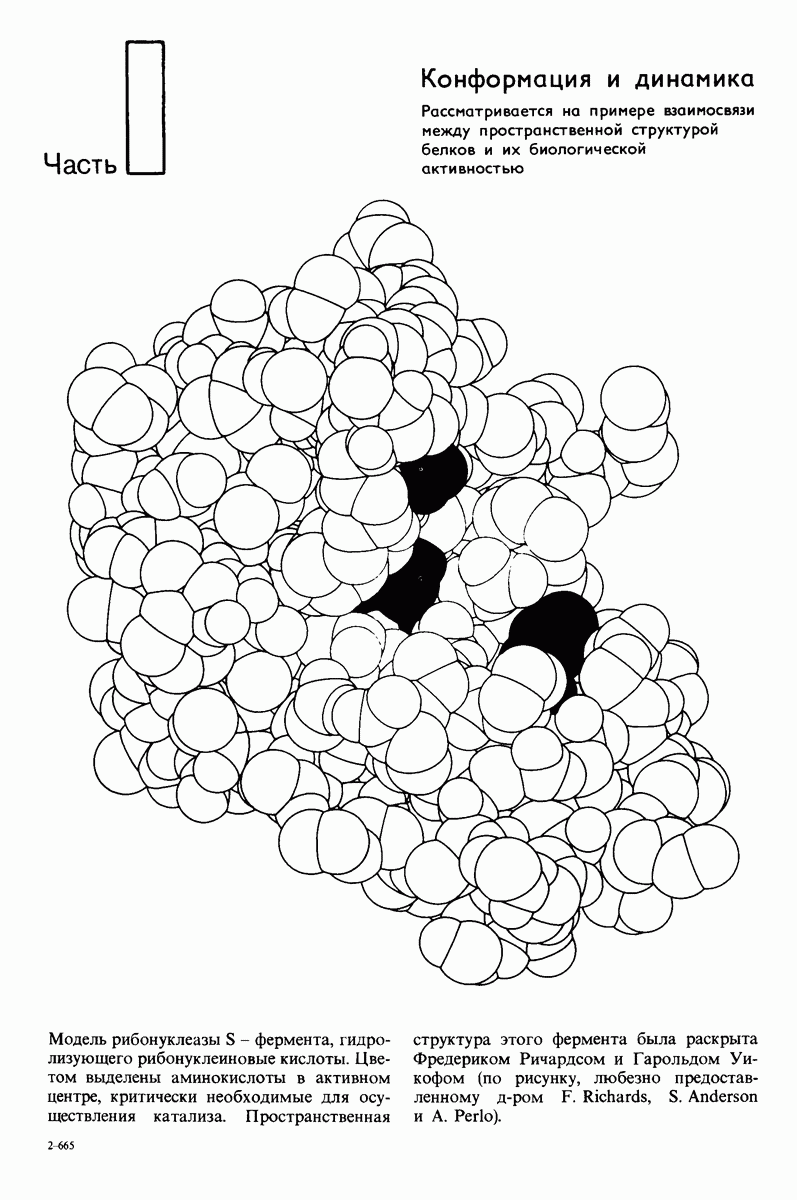
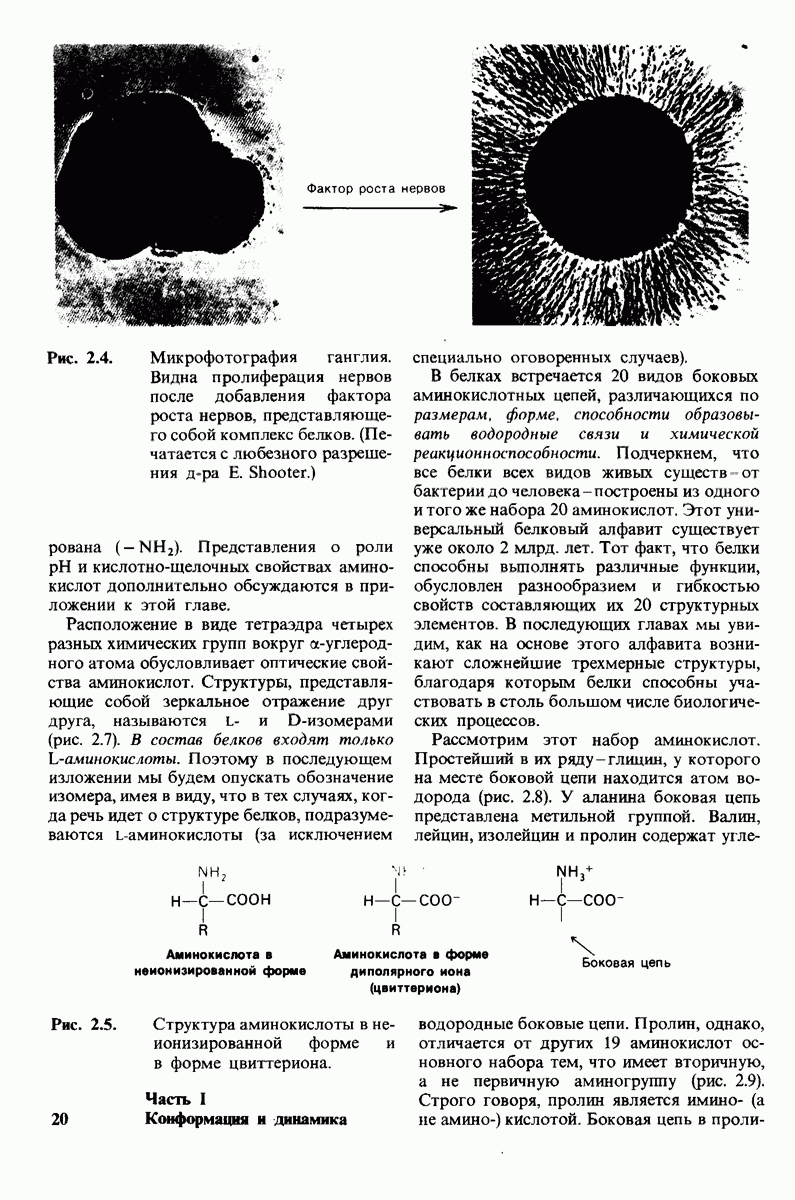
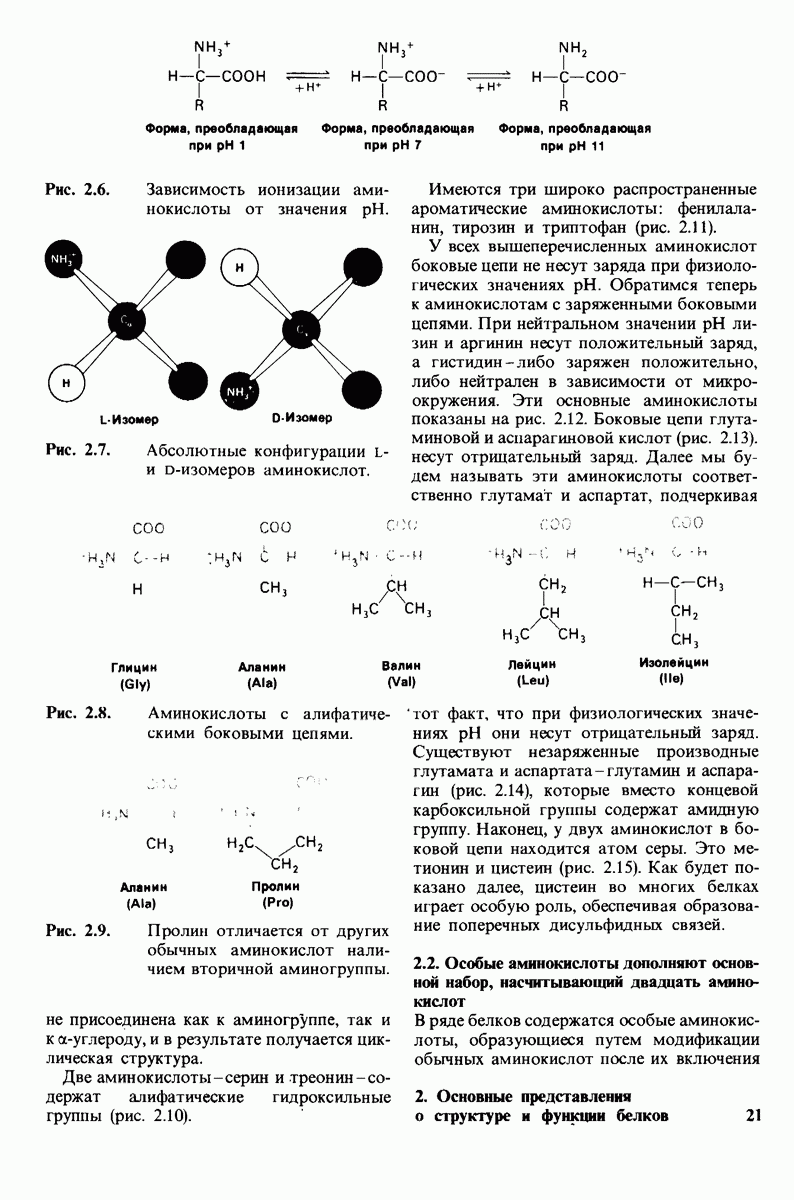
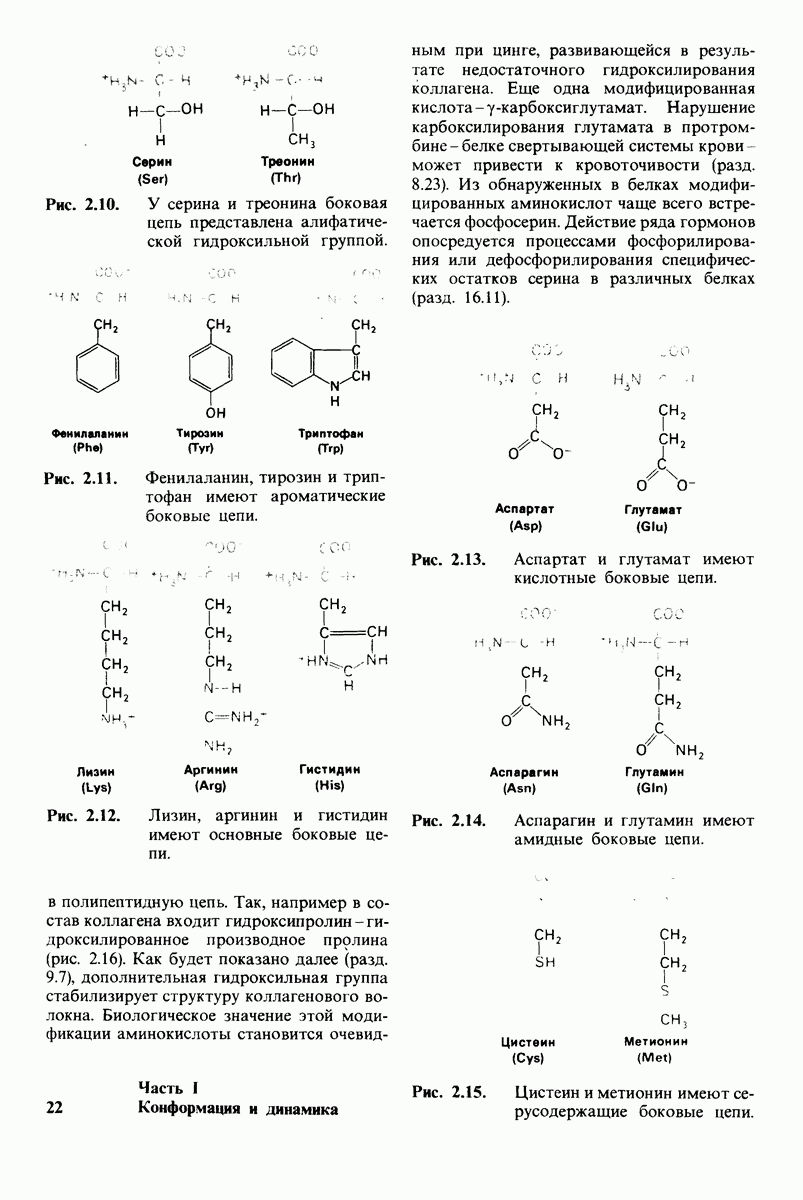
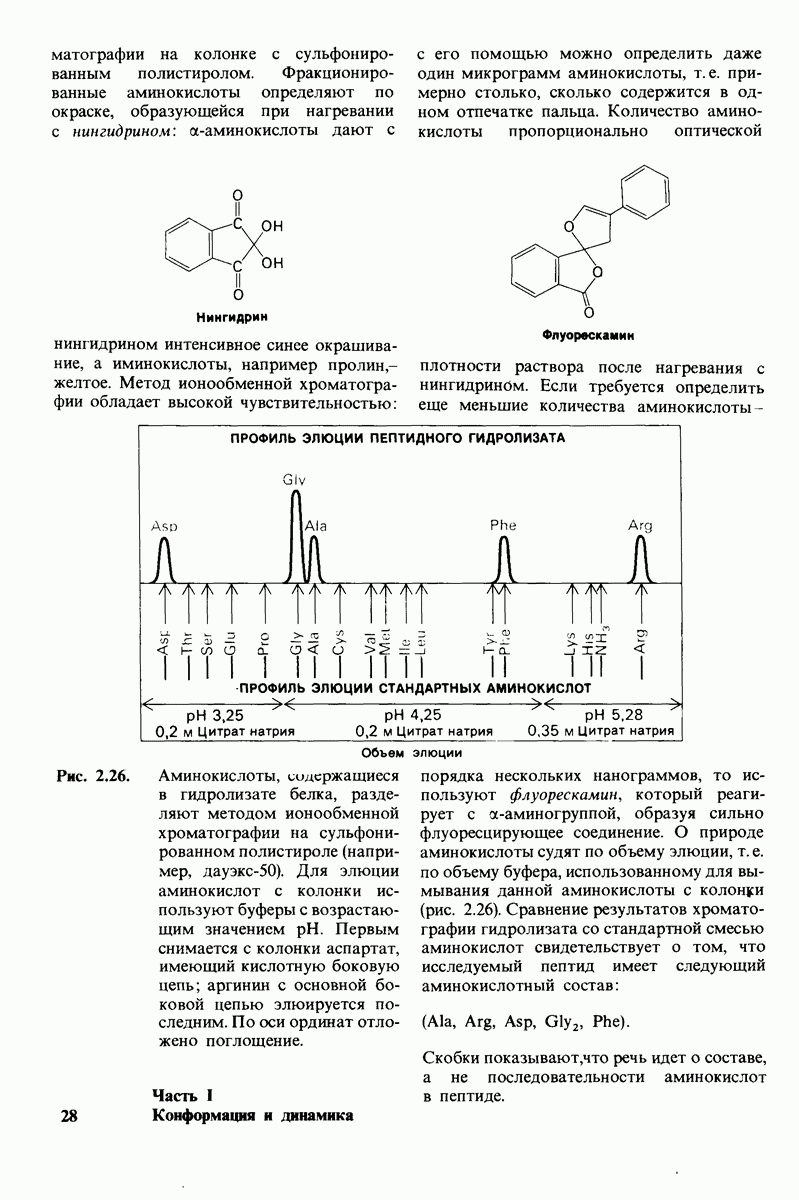
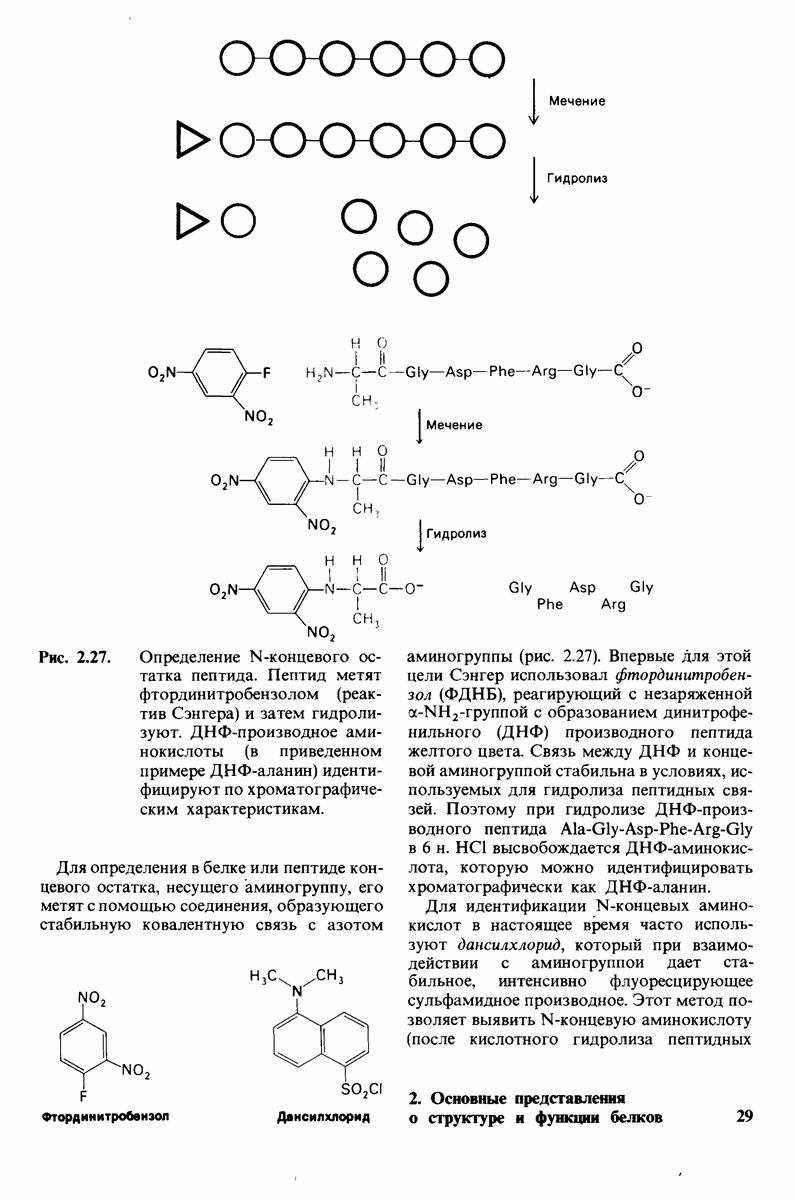
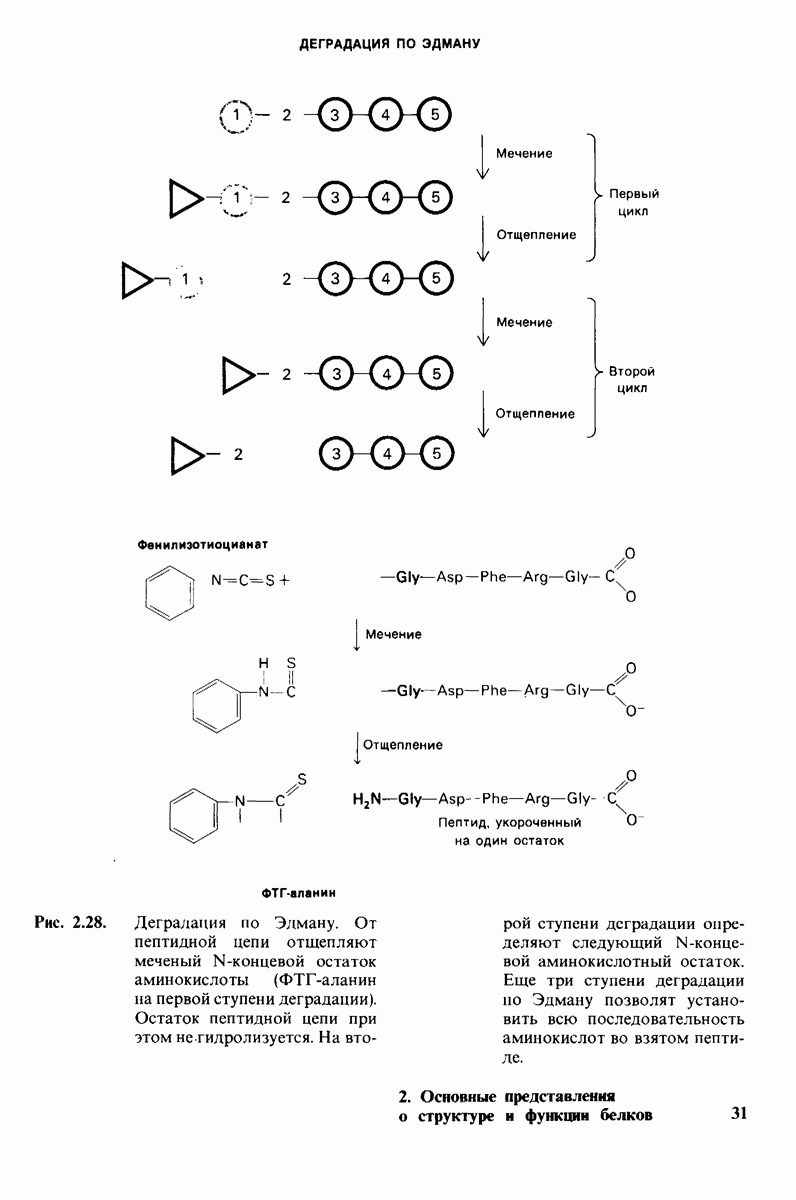
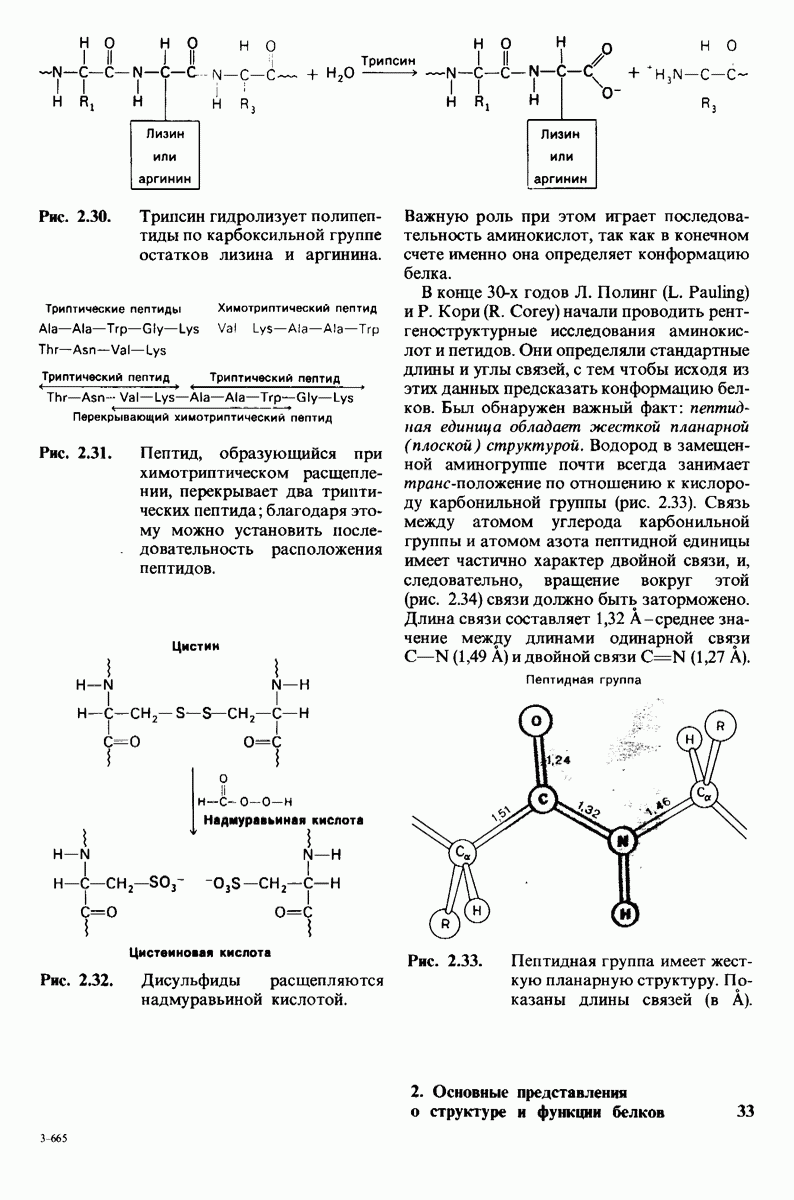
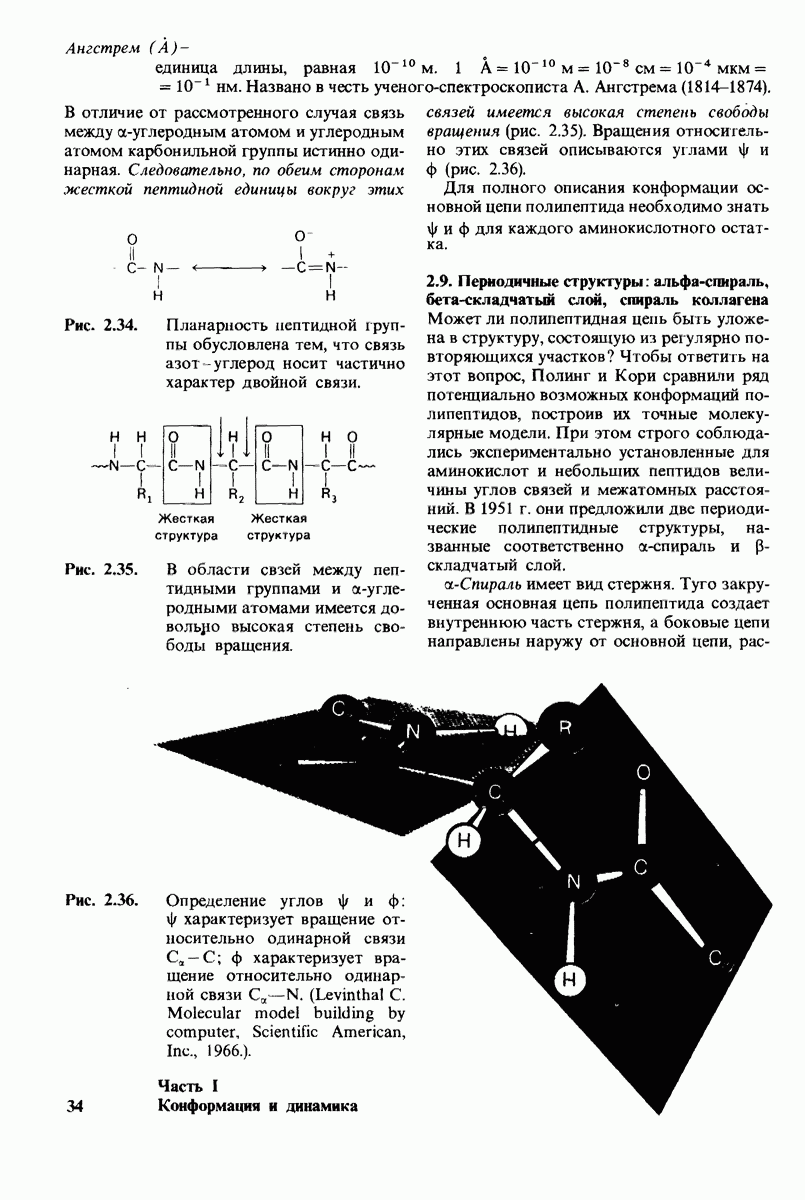
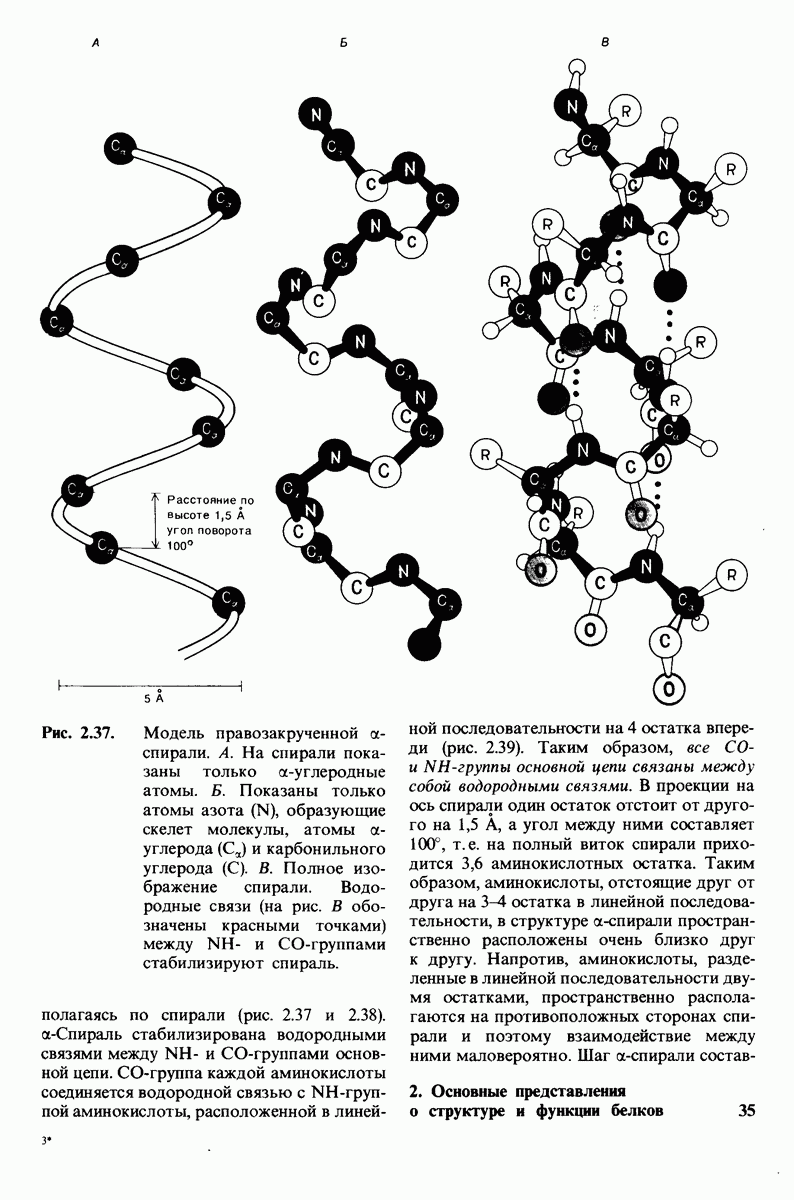
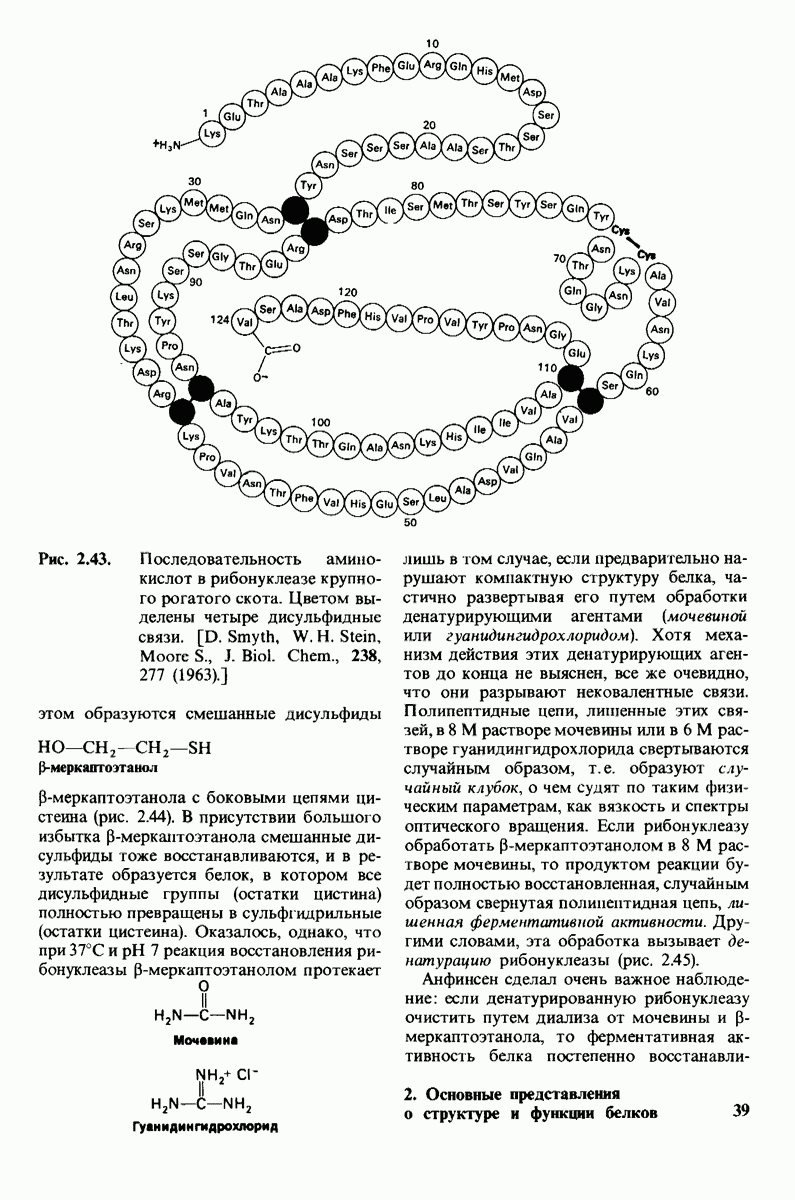
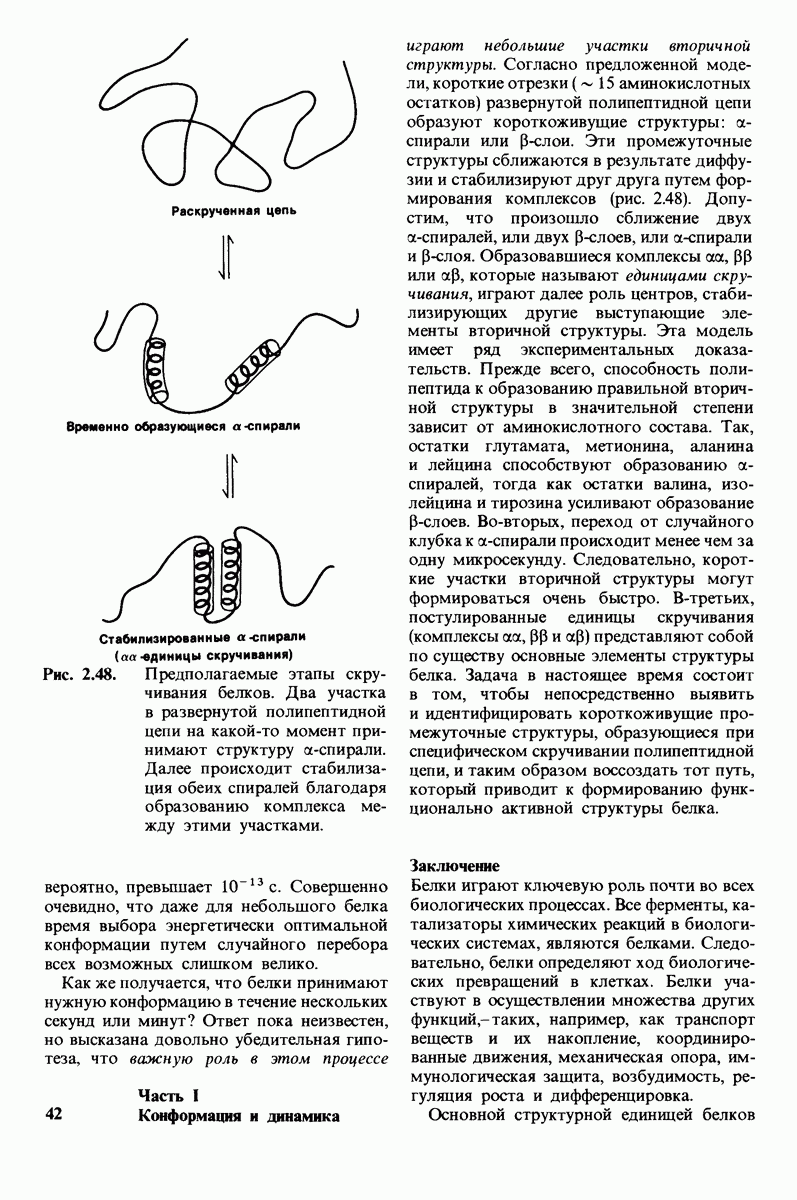
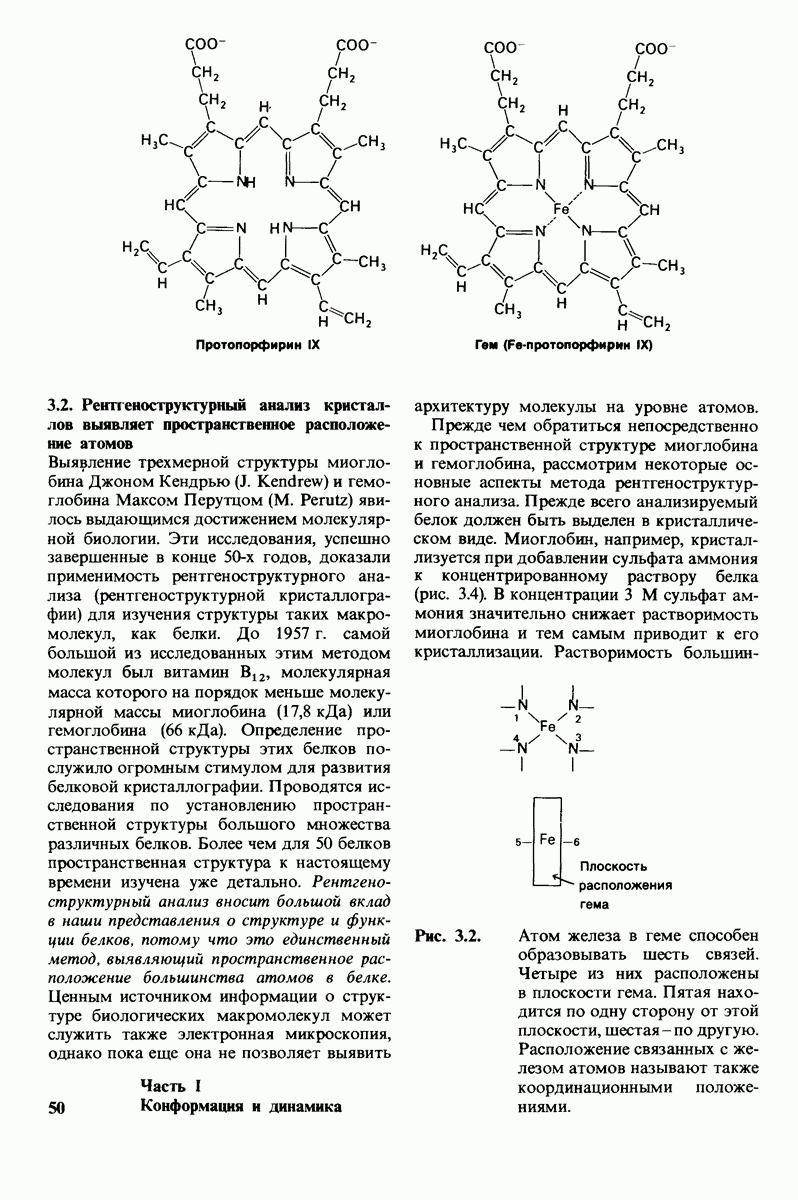
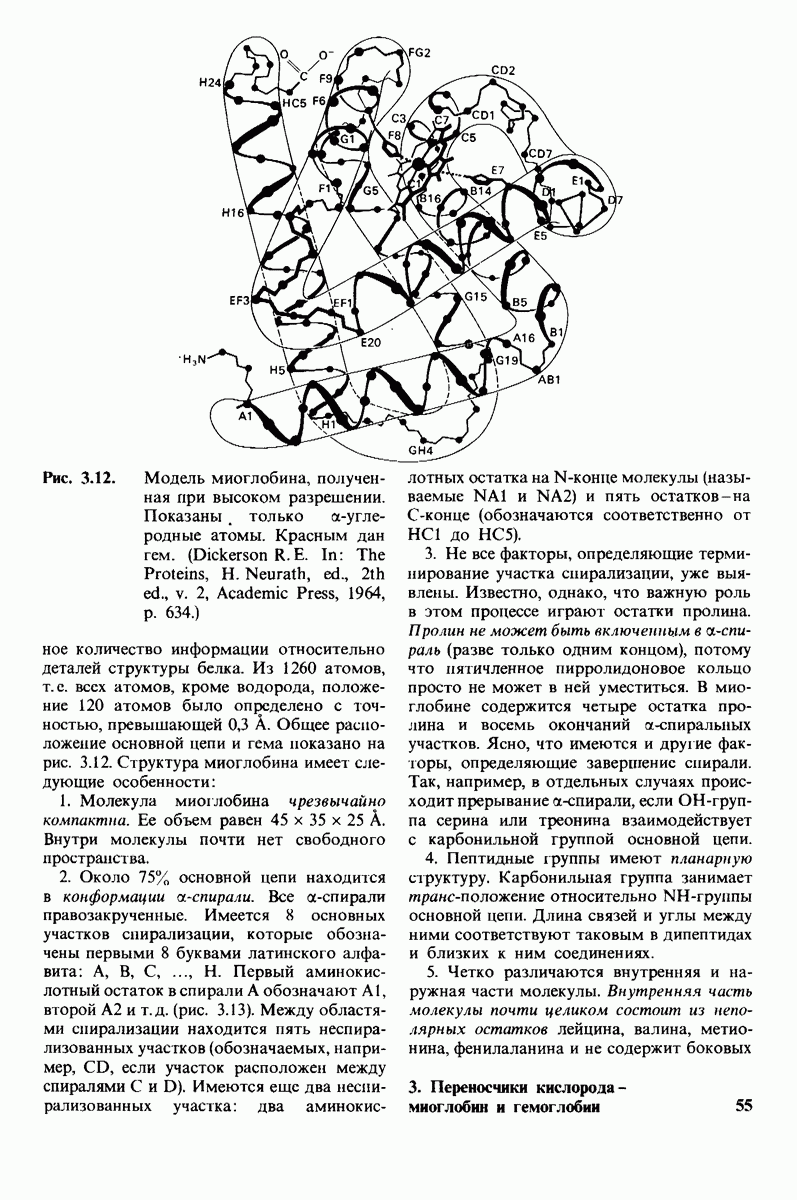
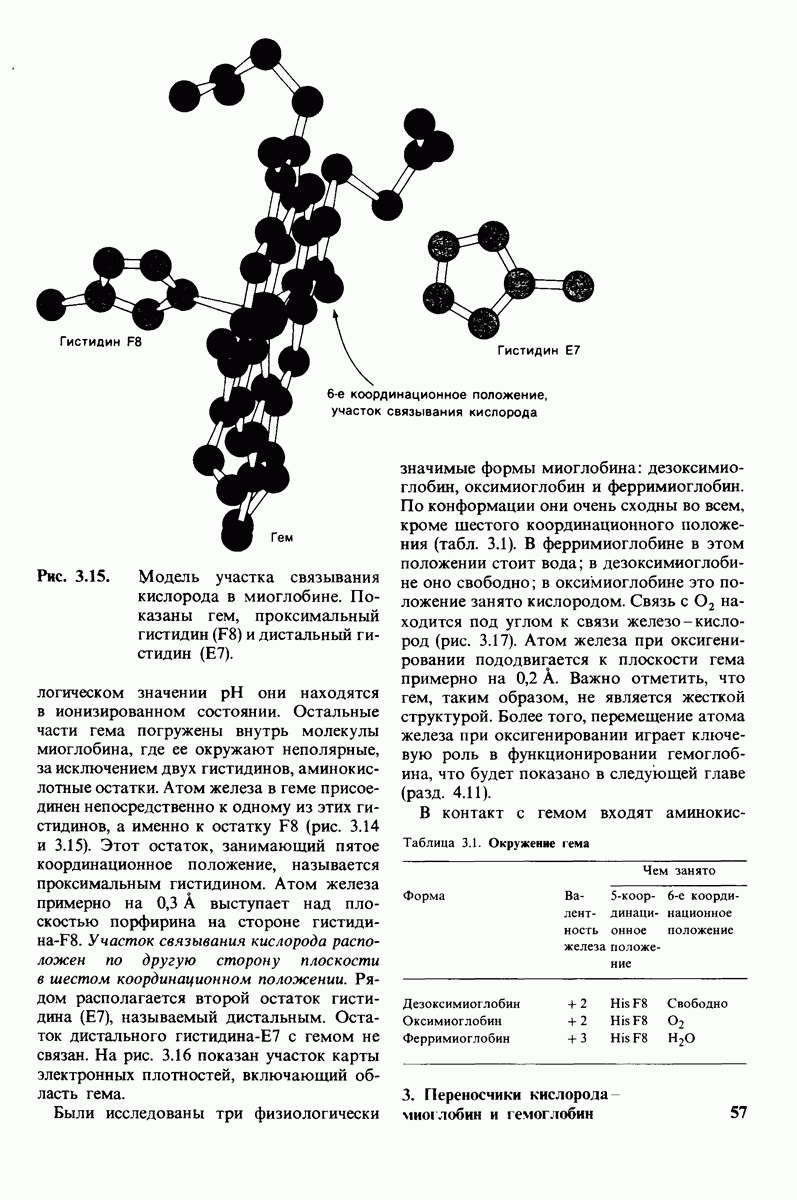
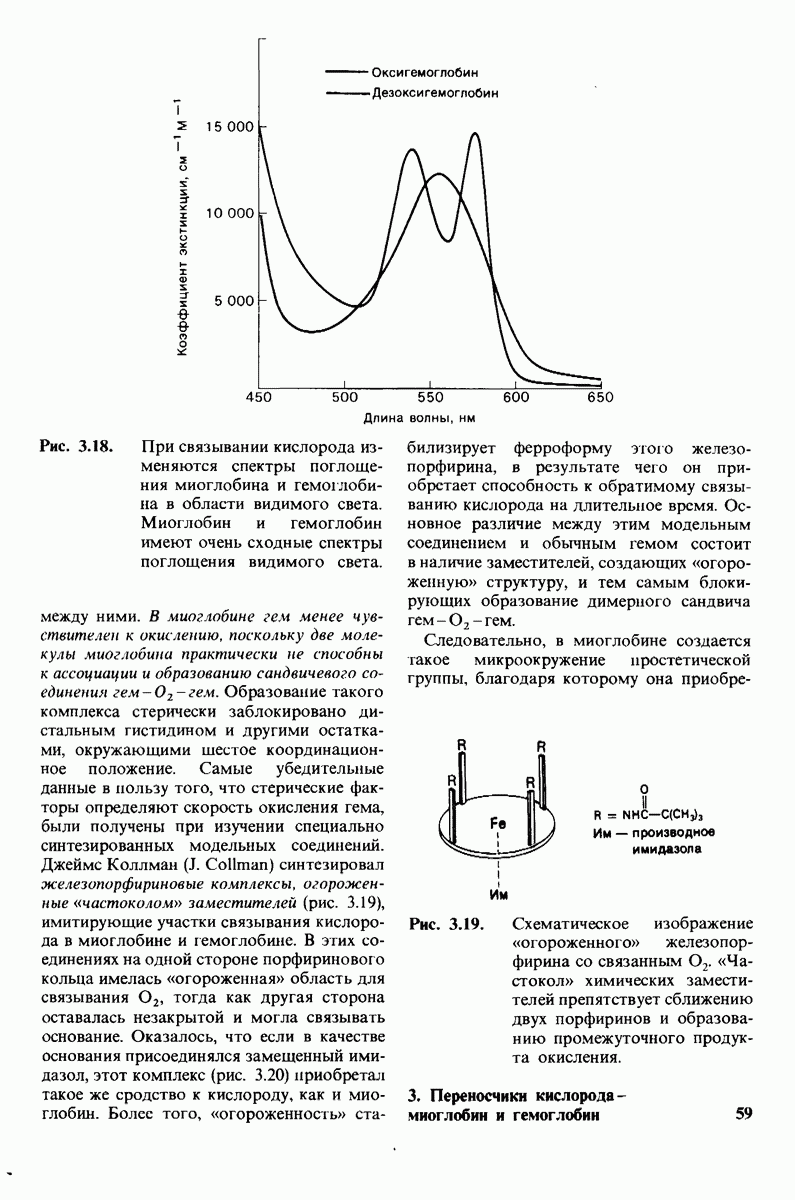
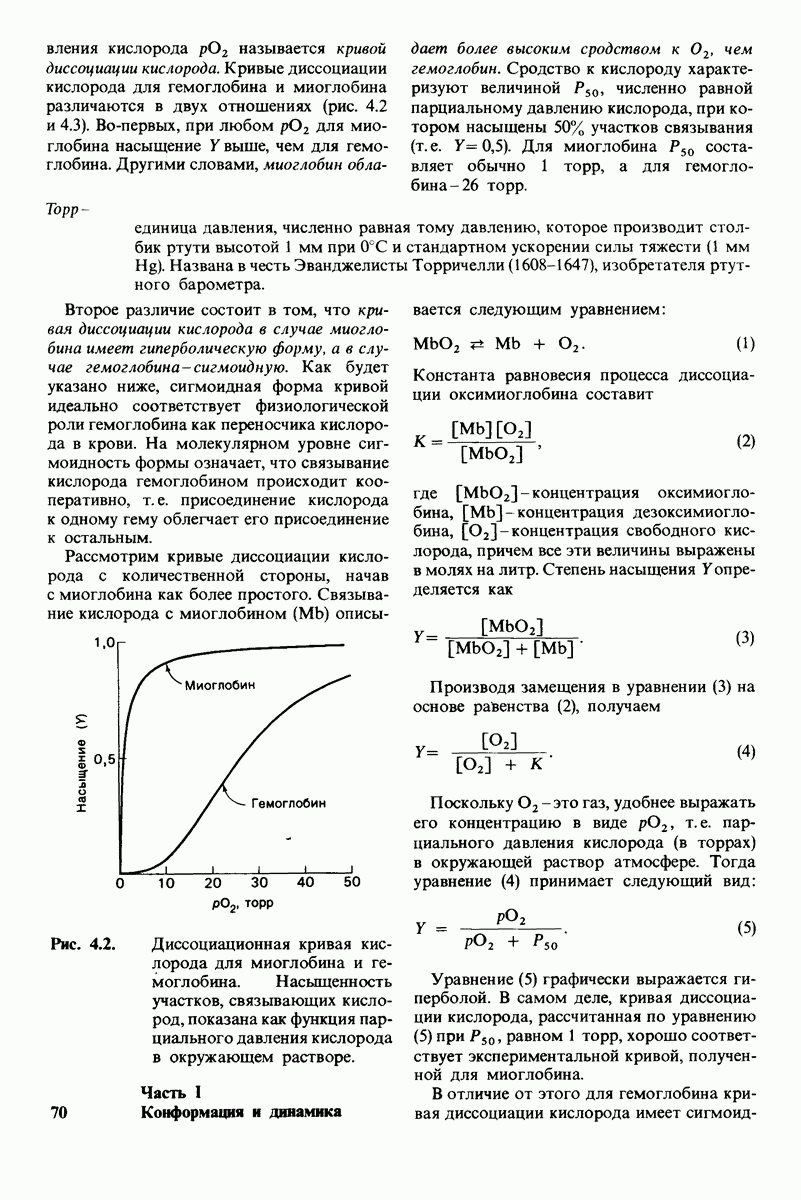
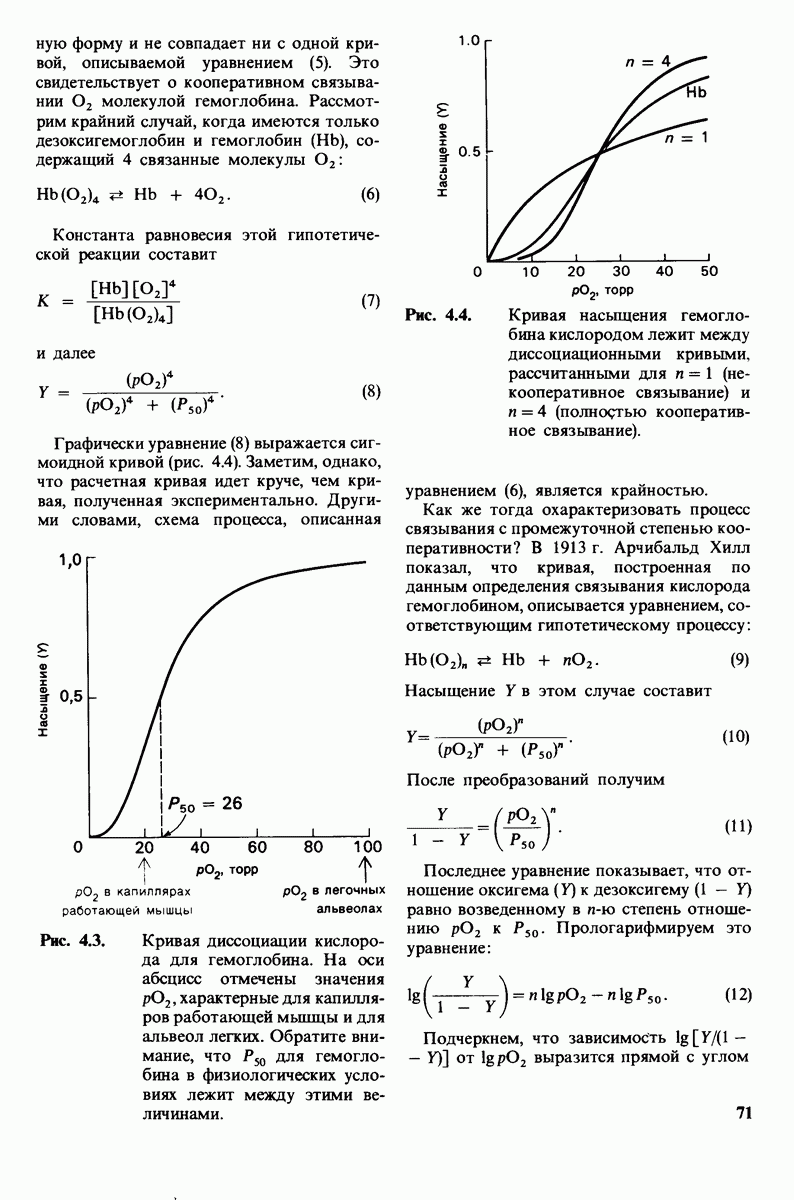
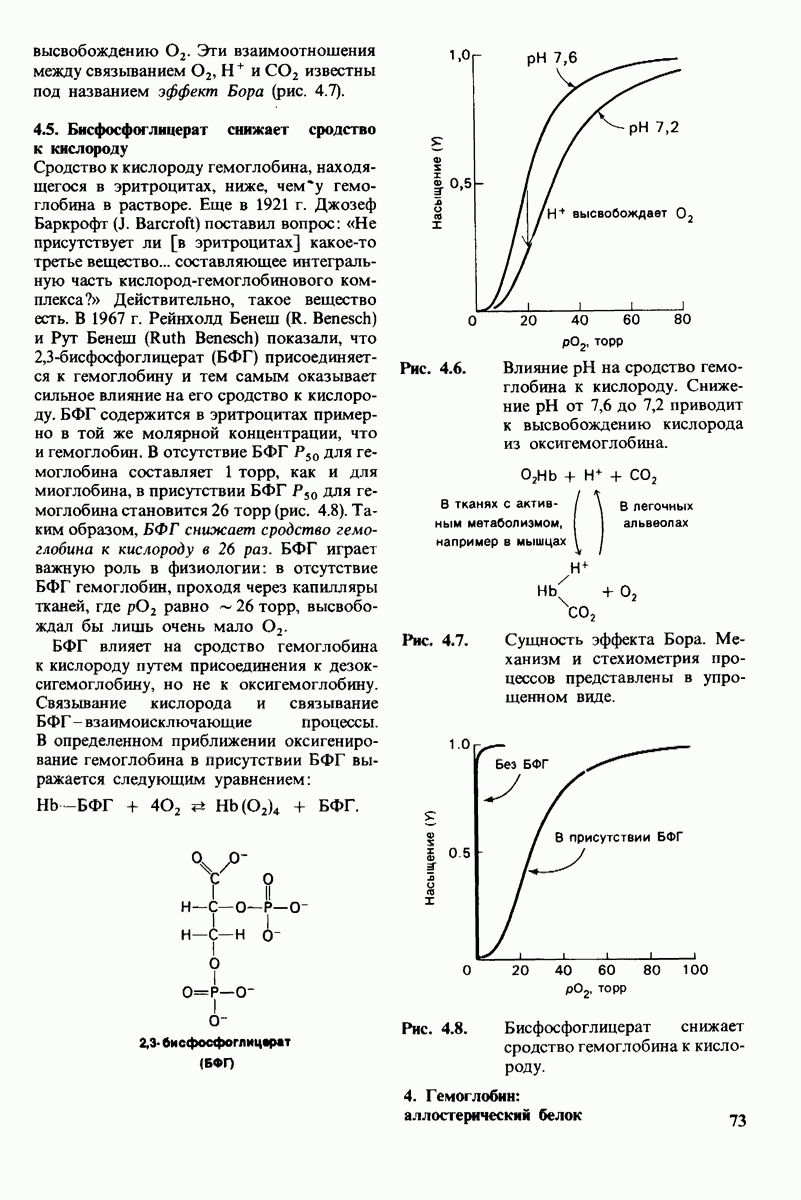
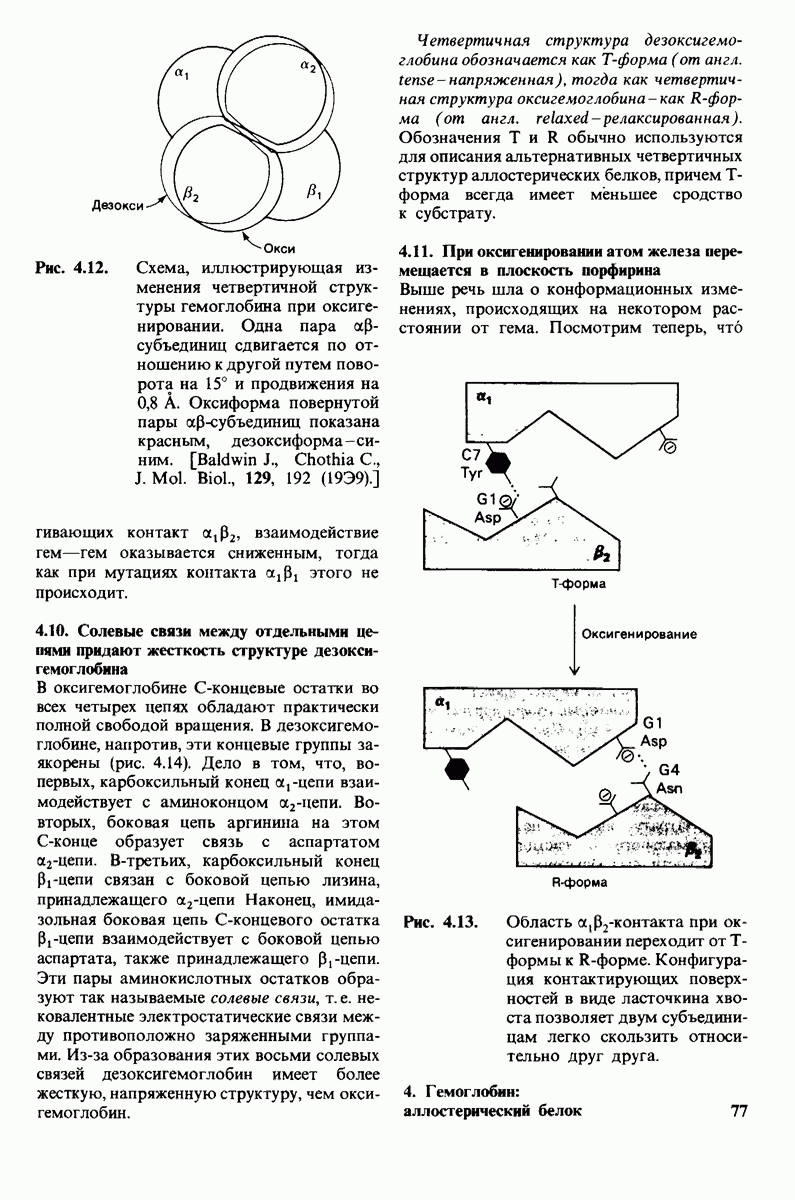
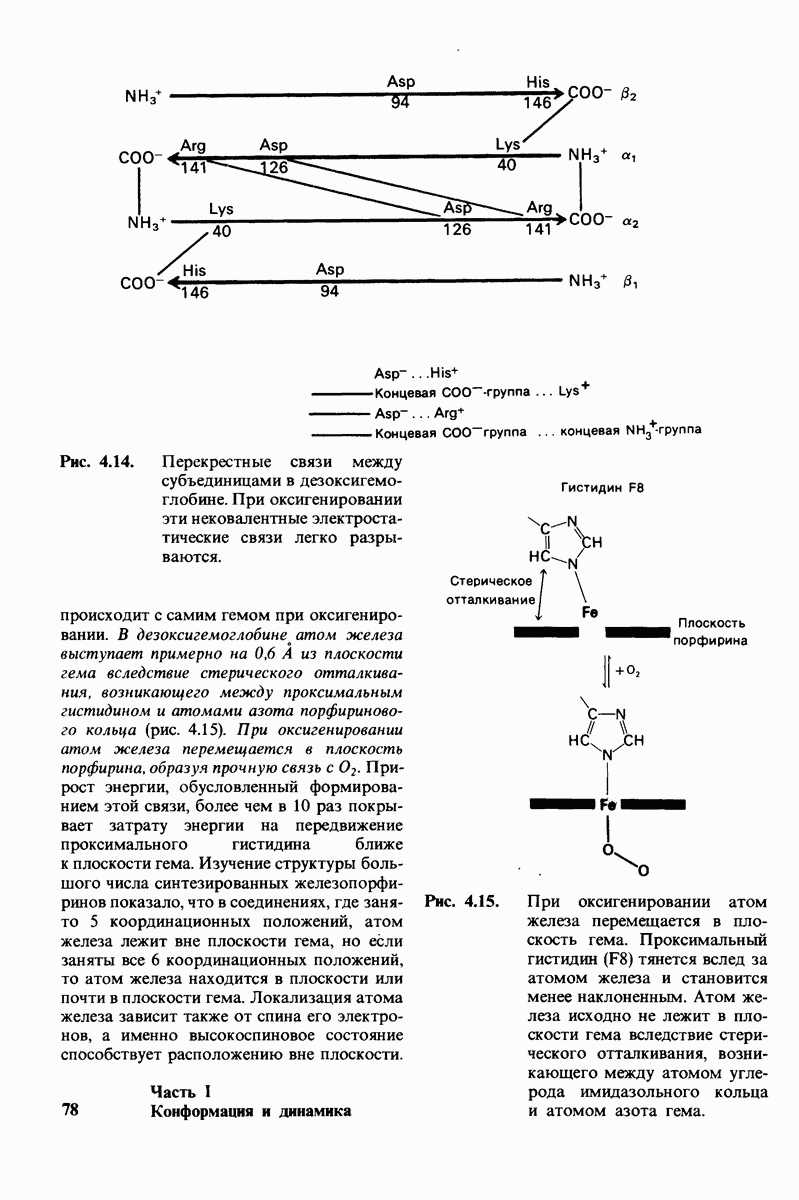
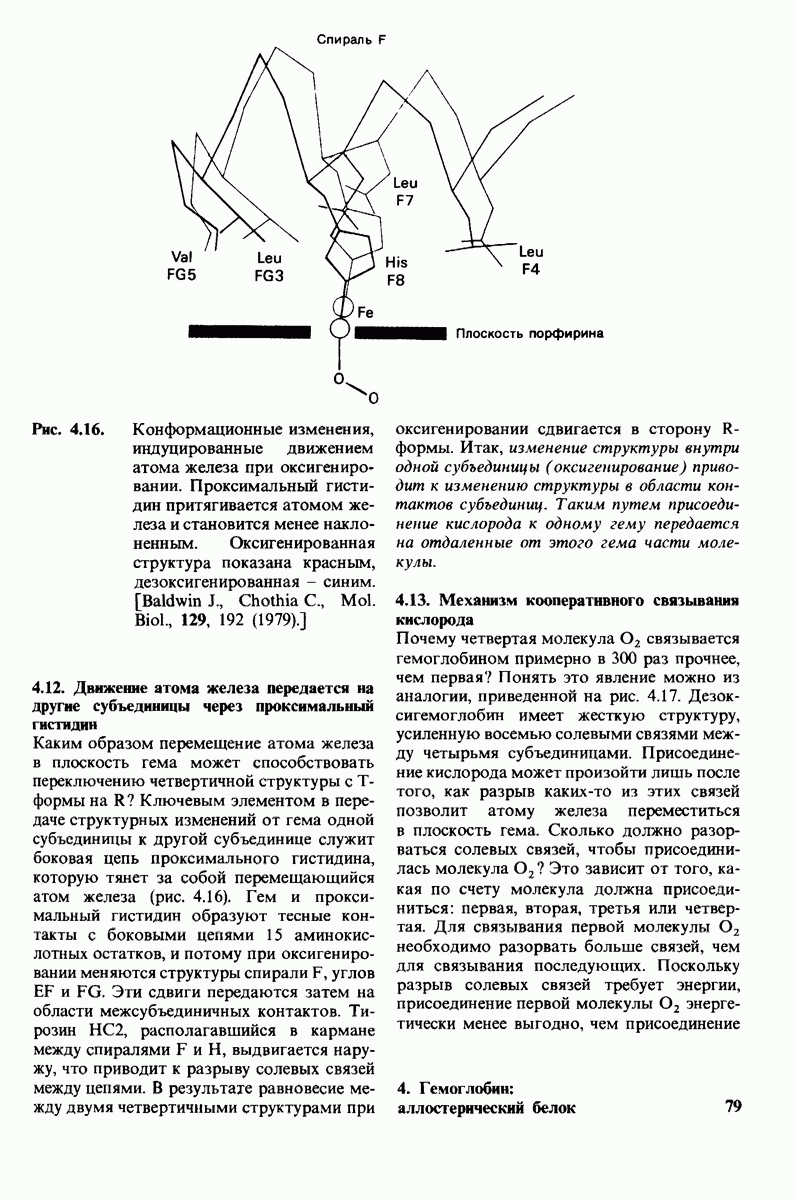
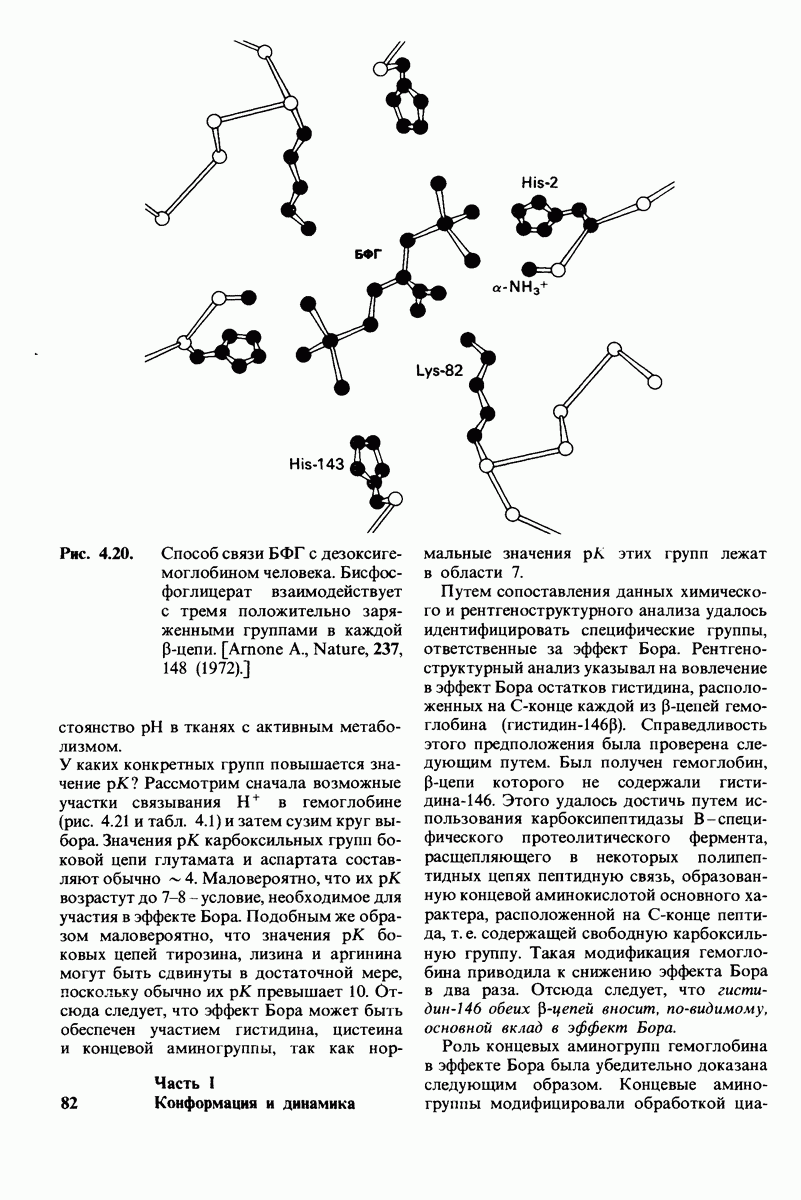
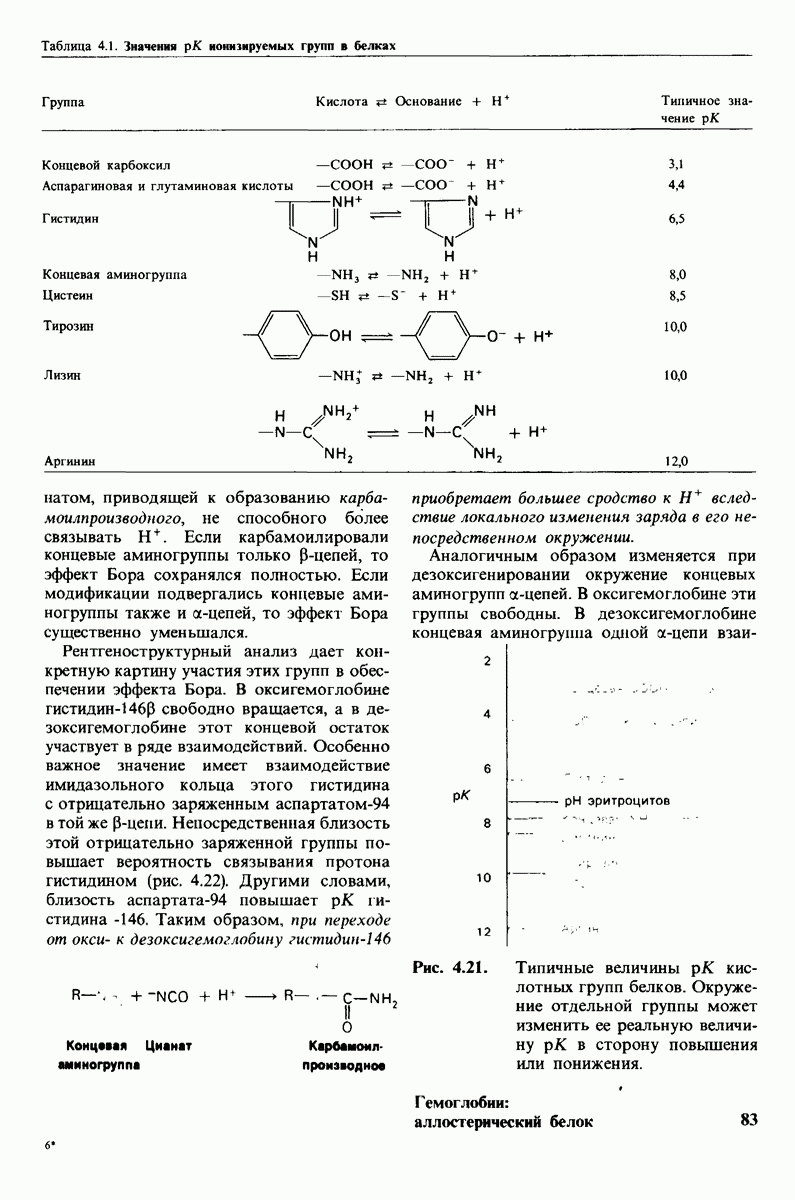
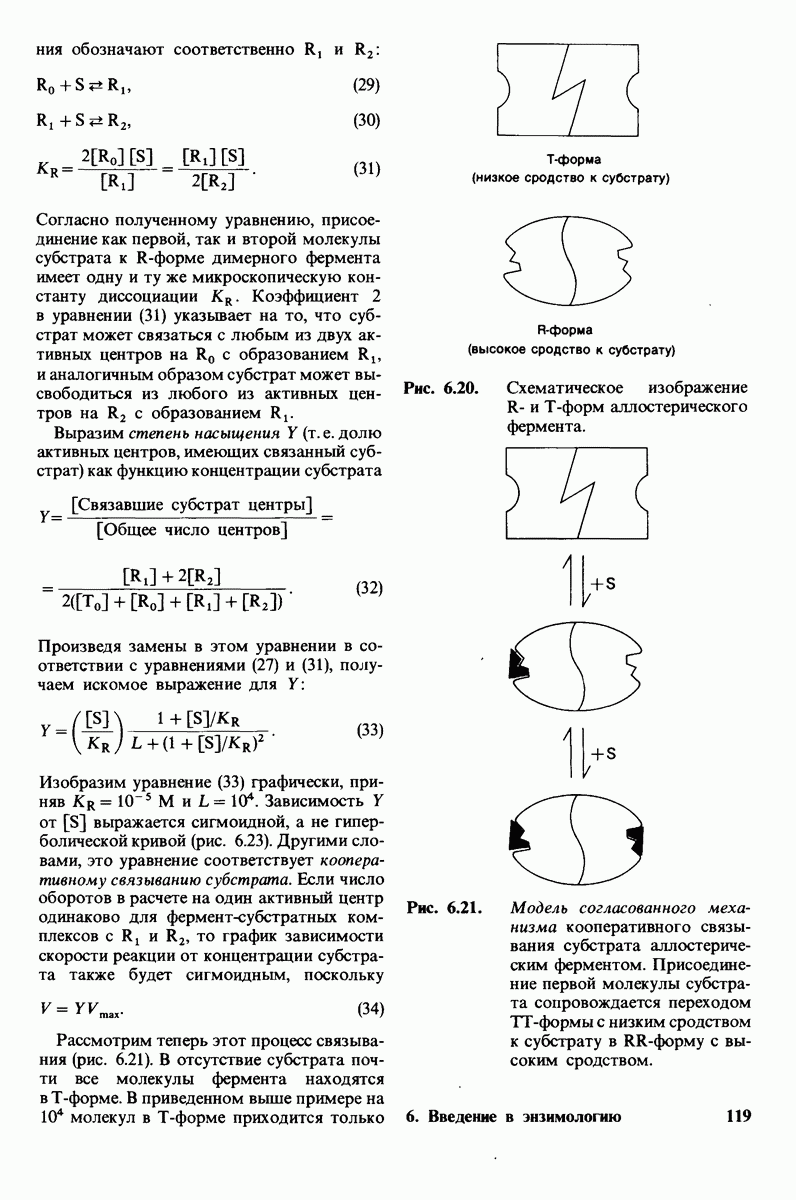
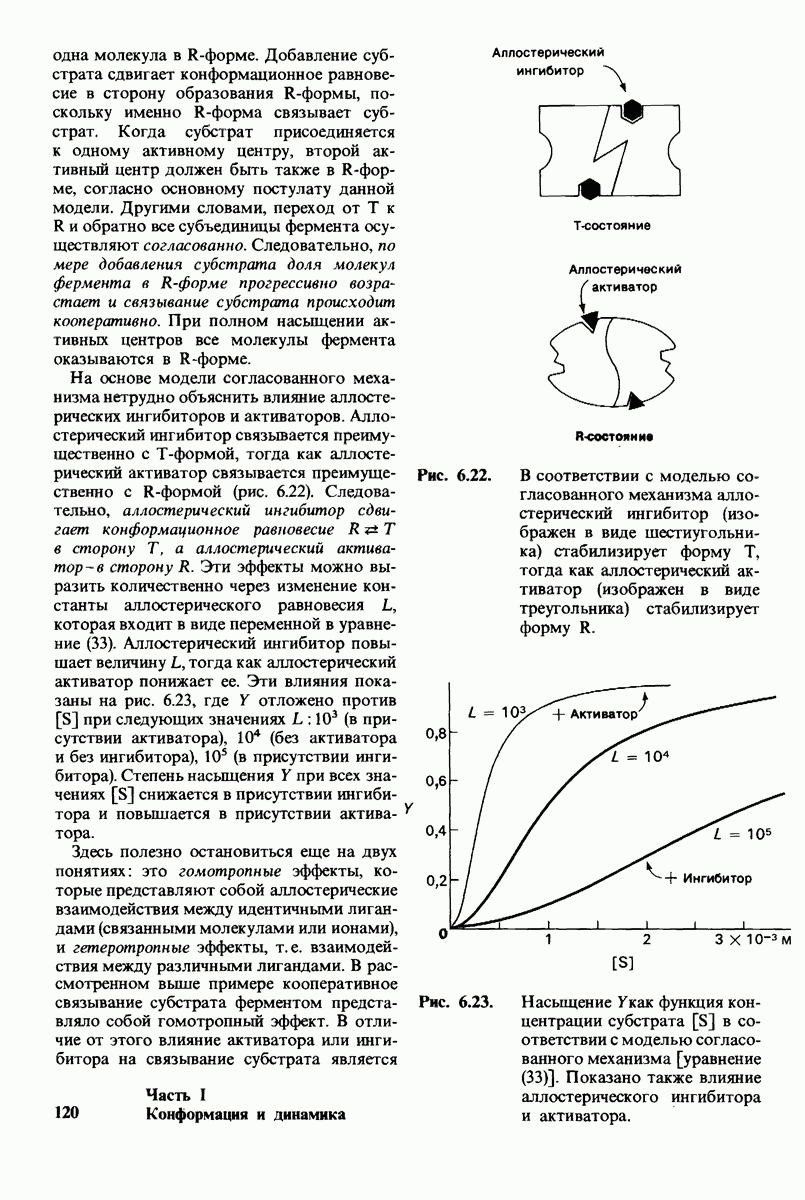
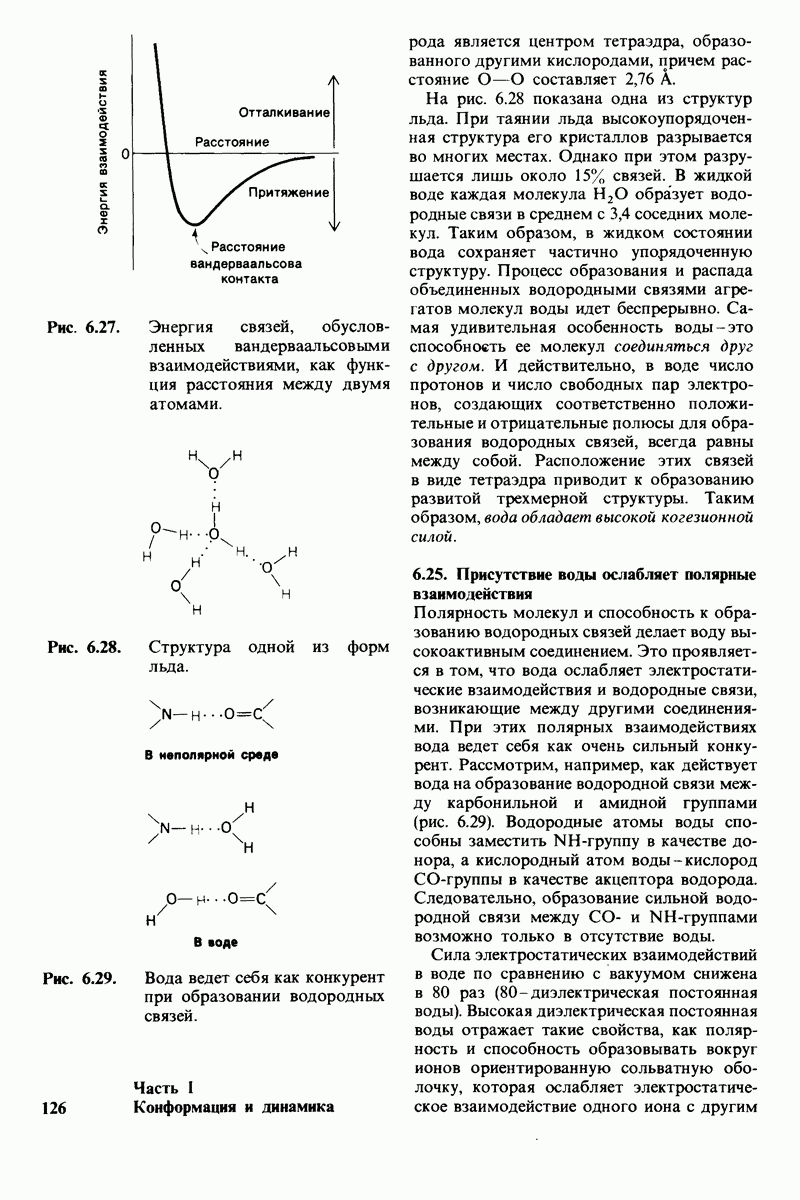
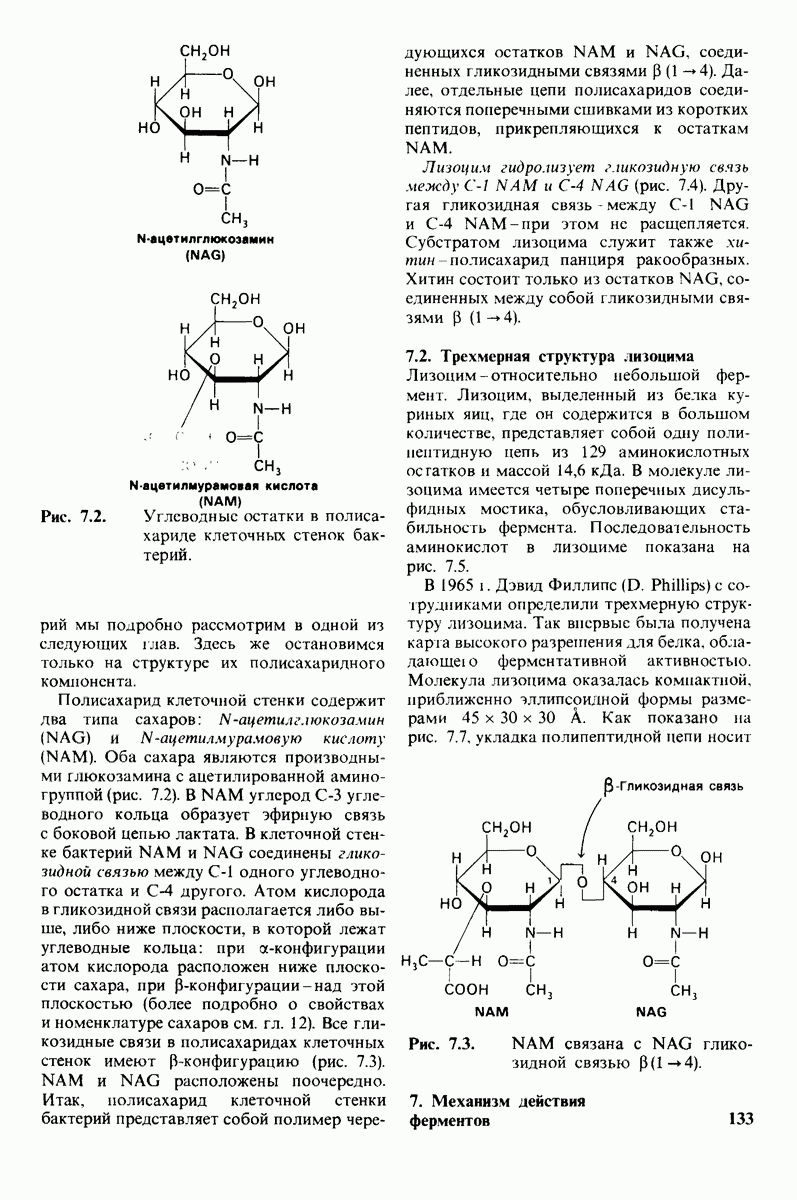
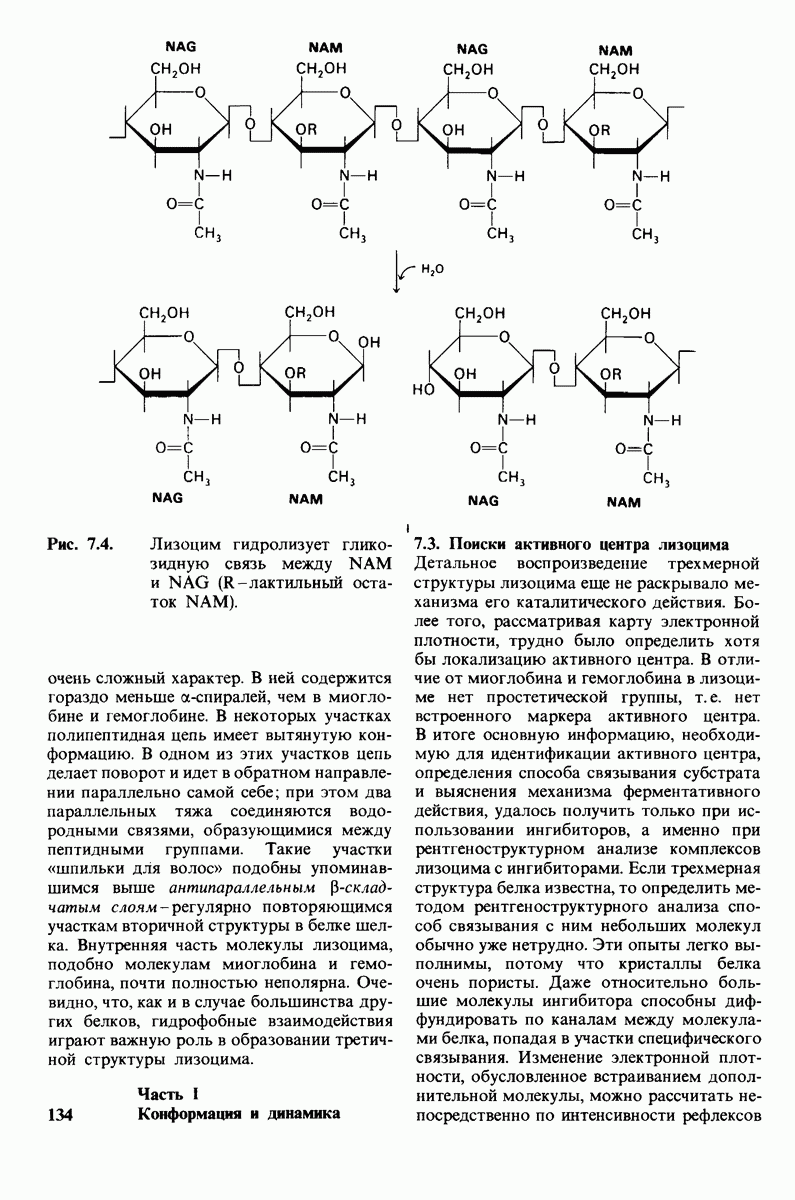
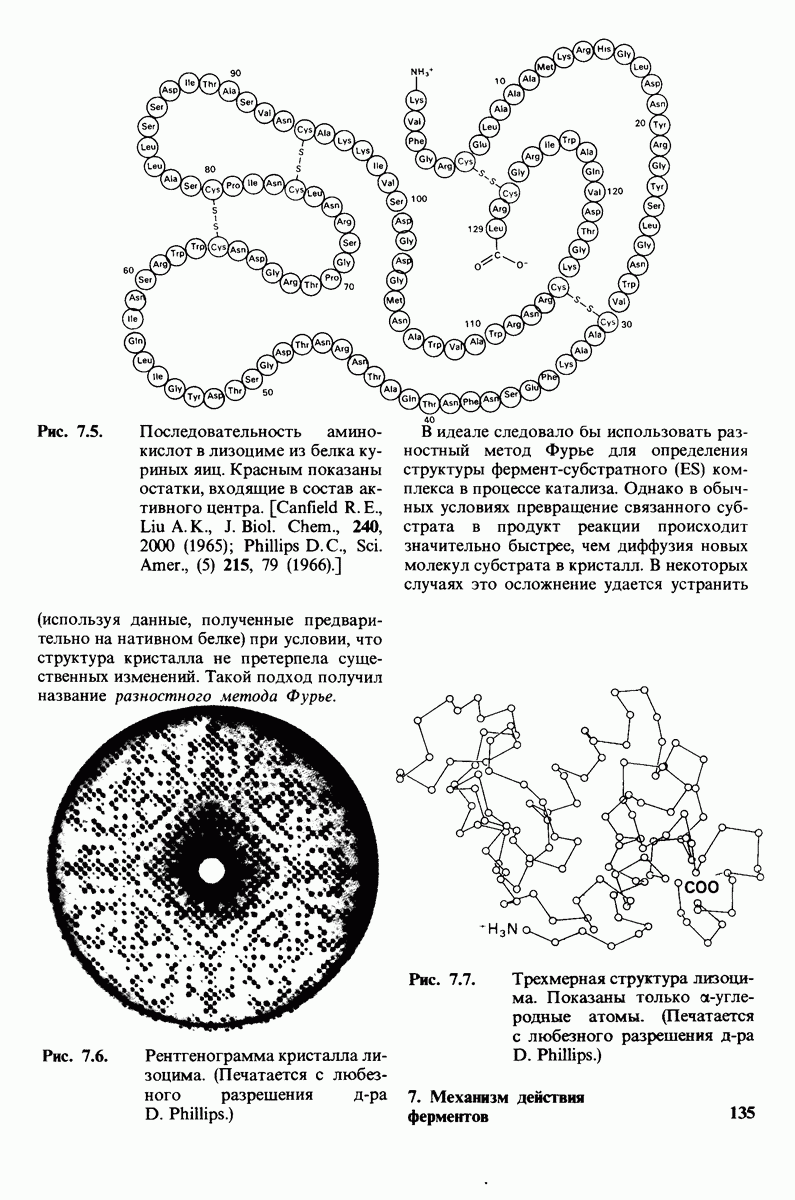
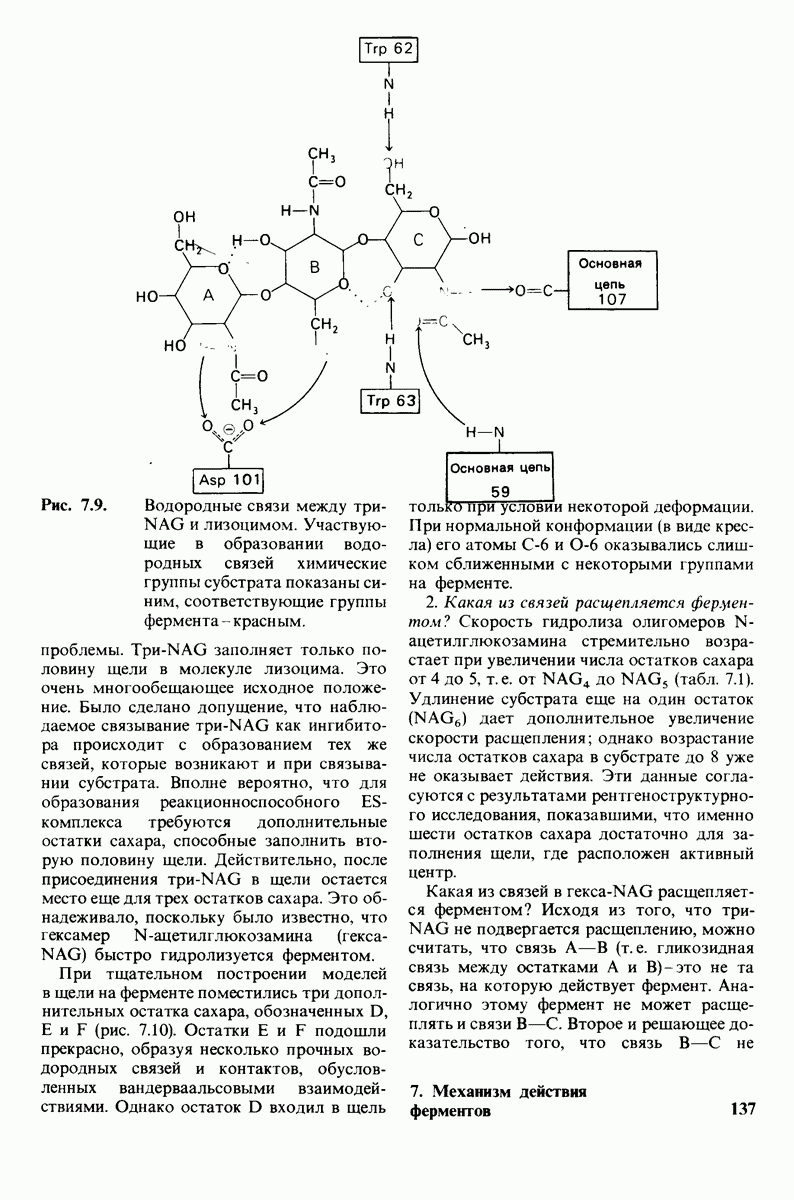
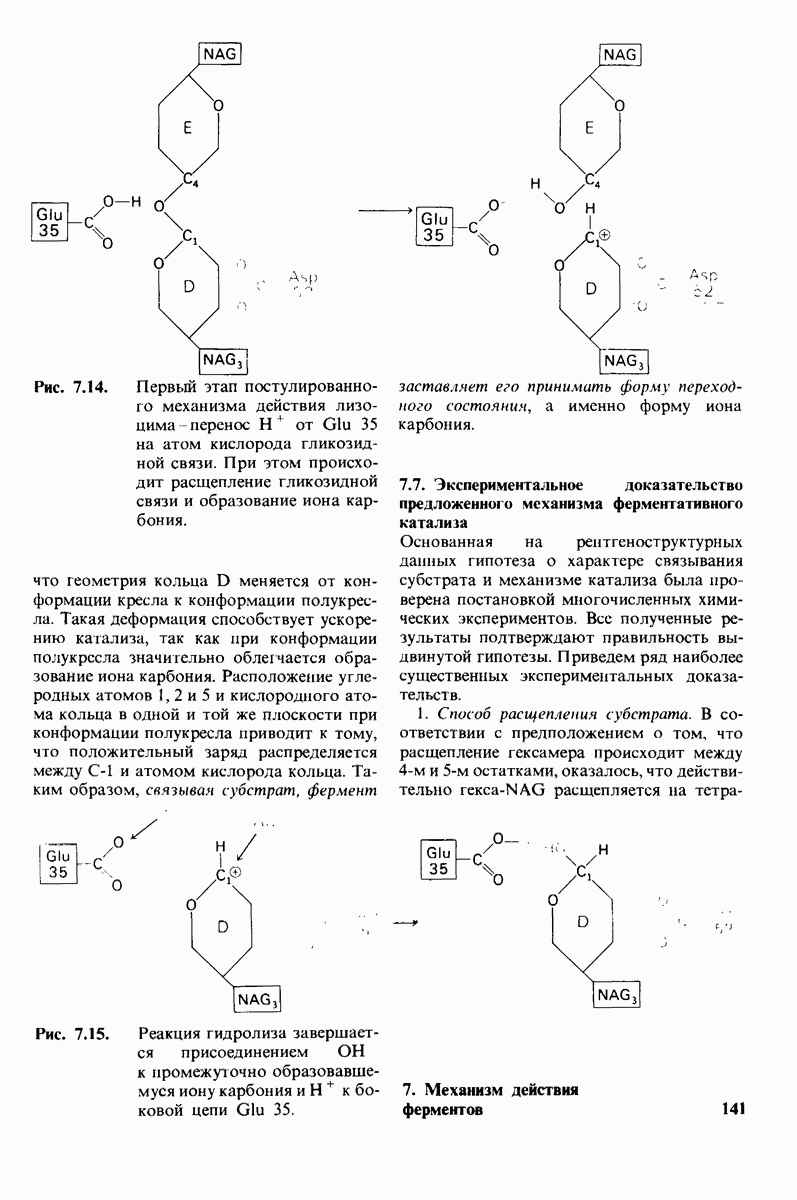
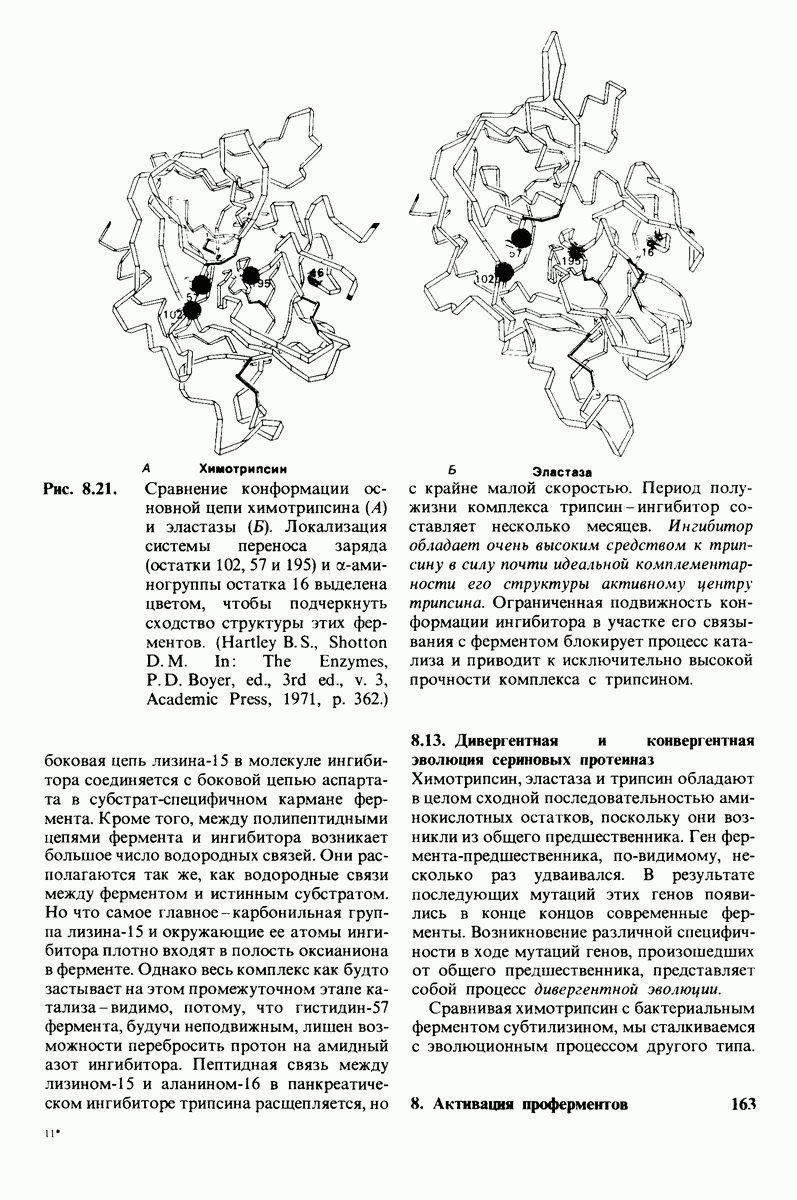
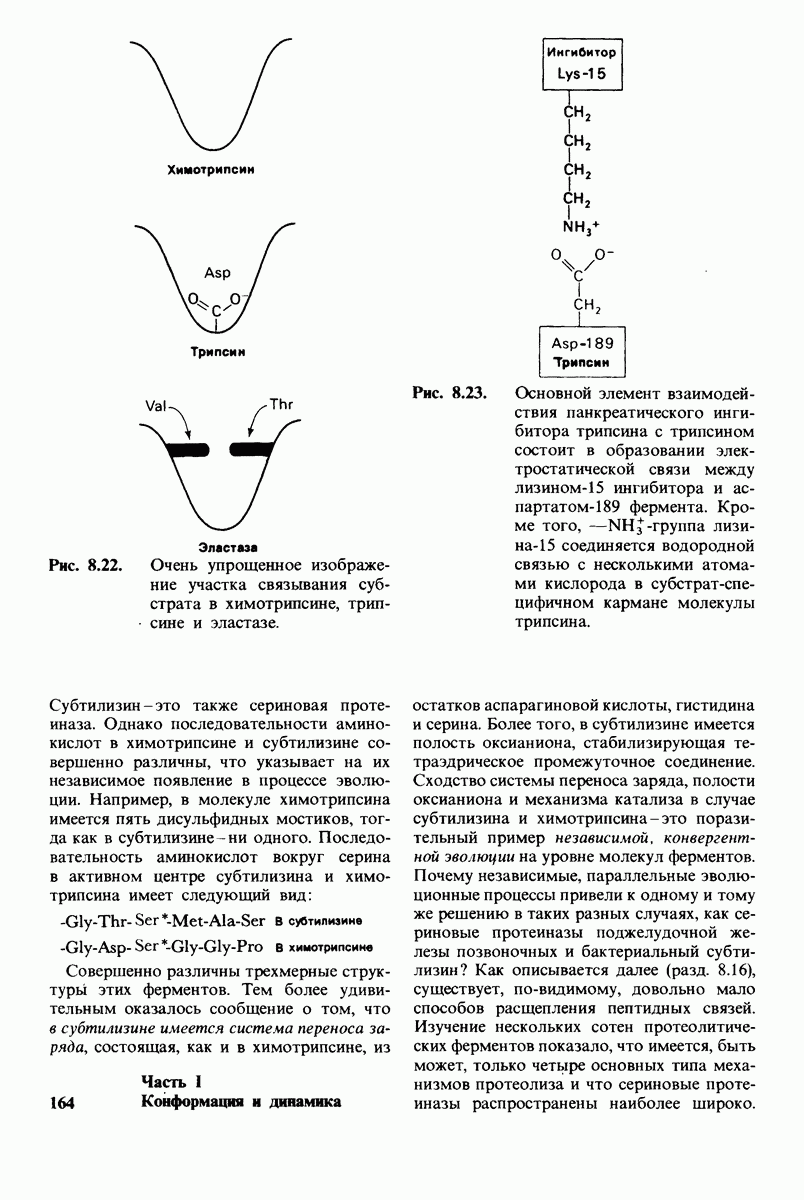
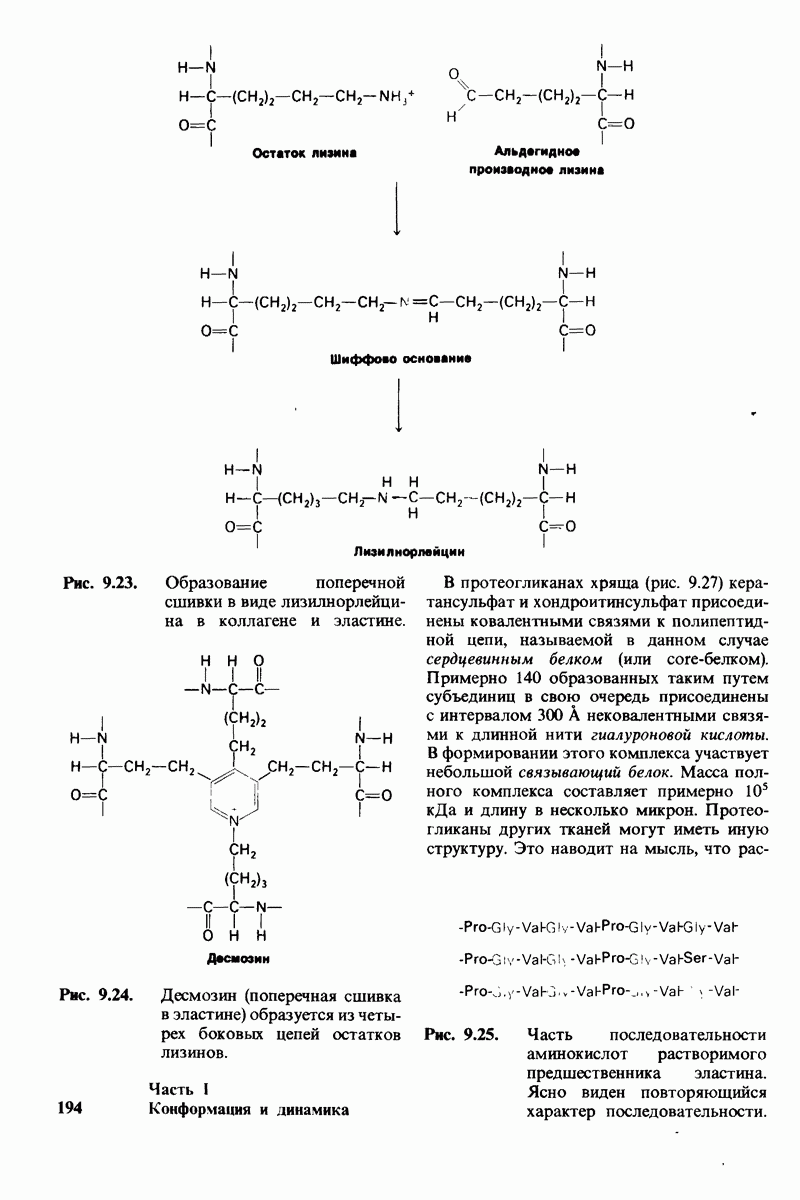
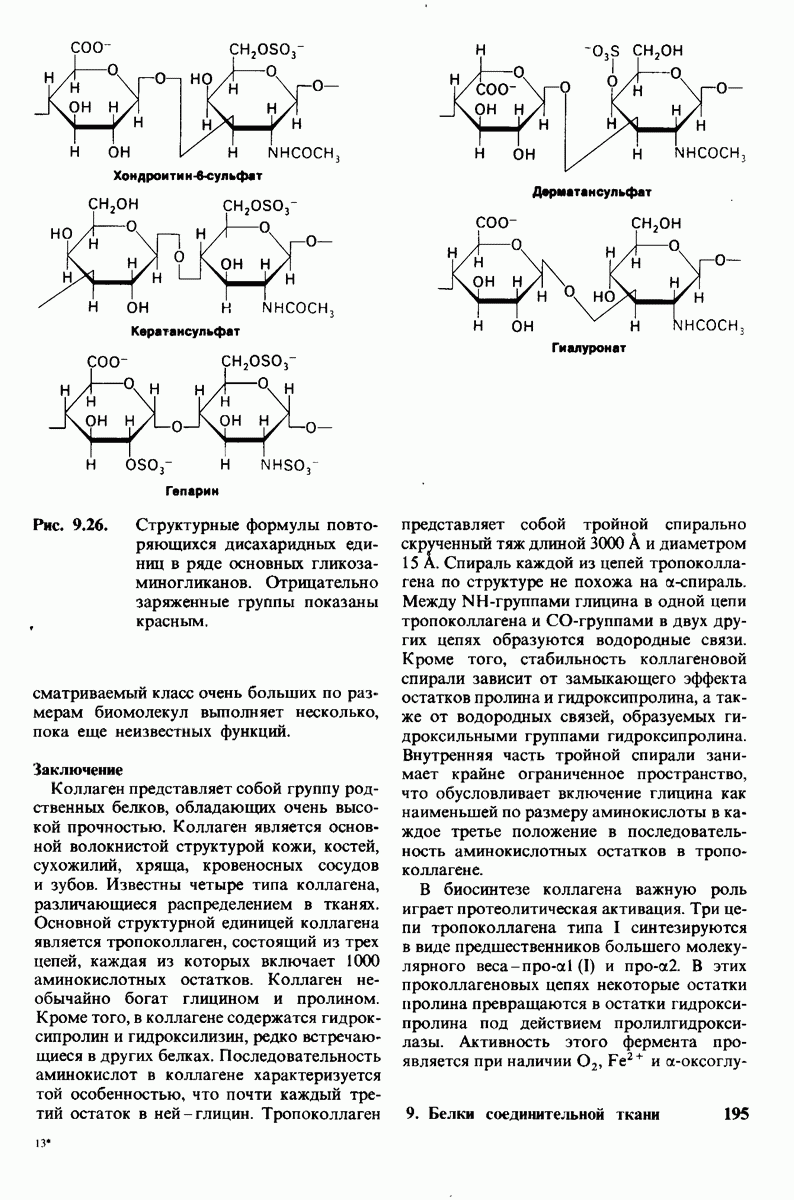
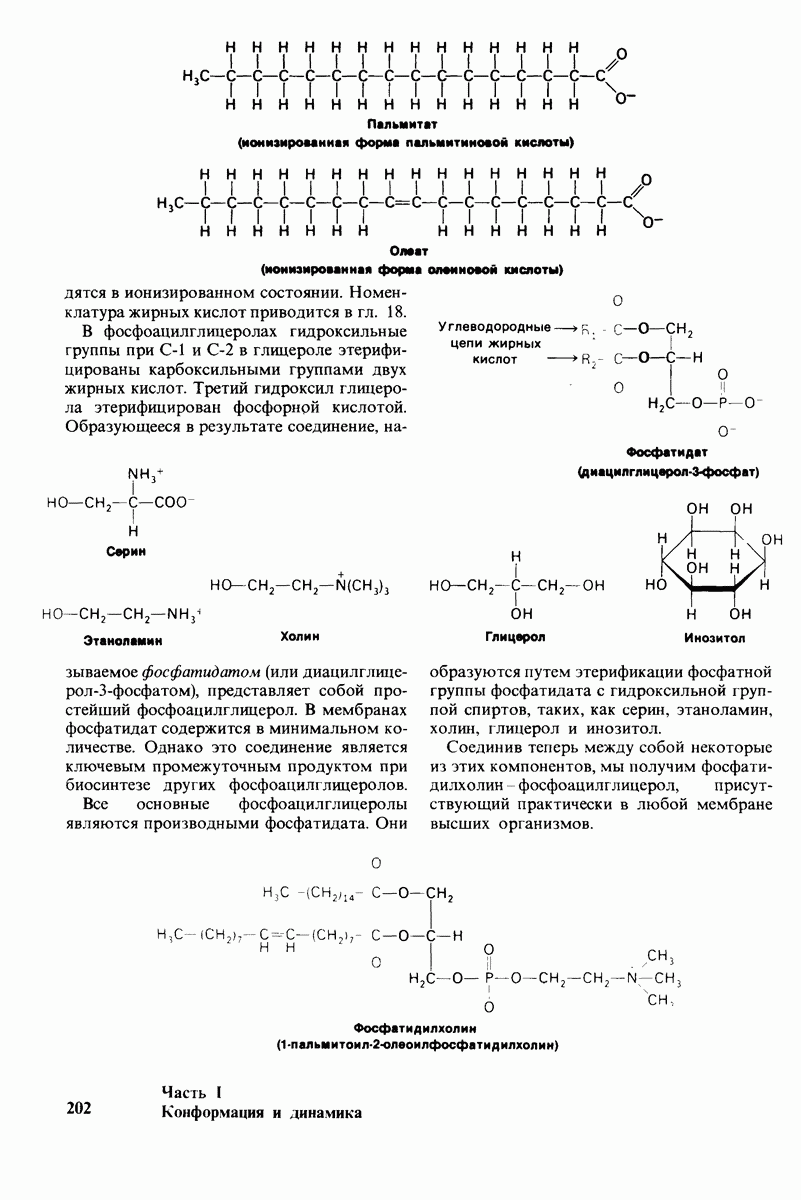
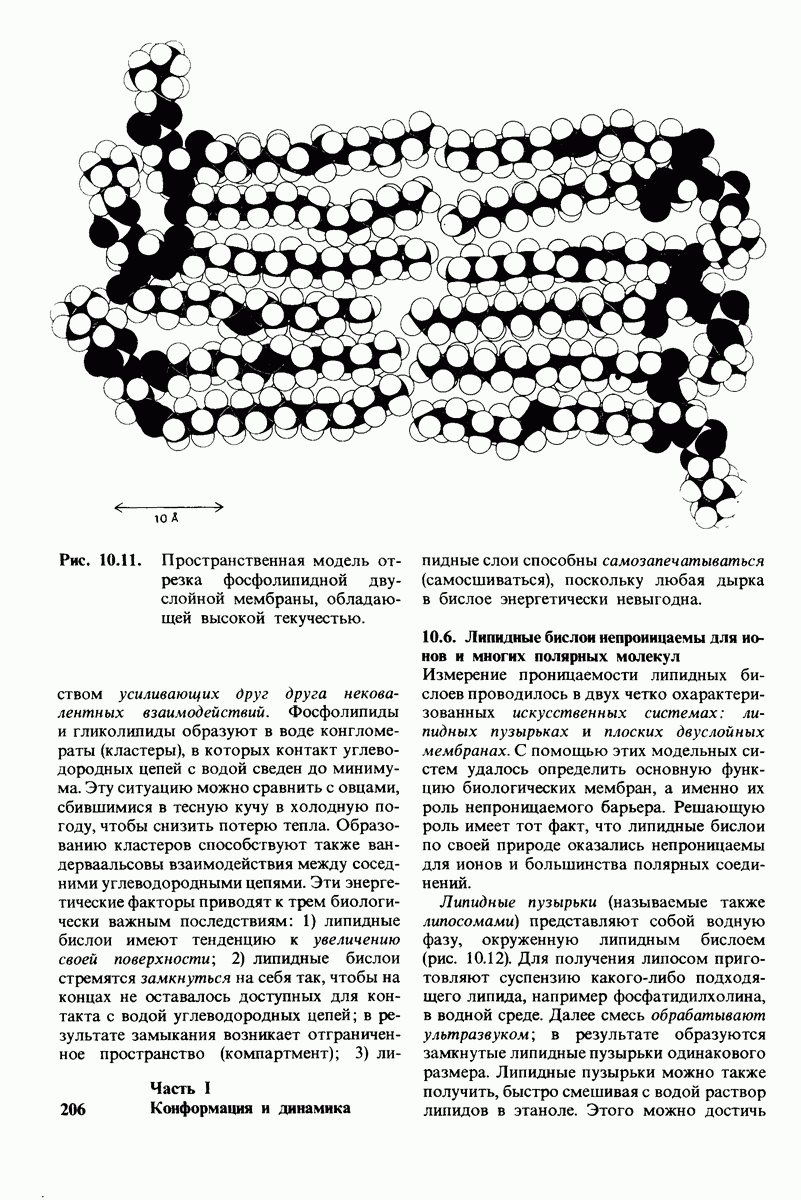
8.10. Механизм активации профермента
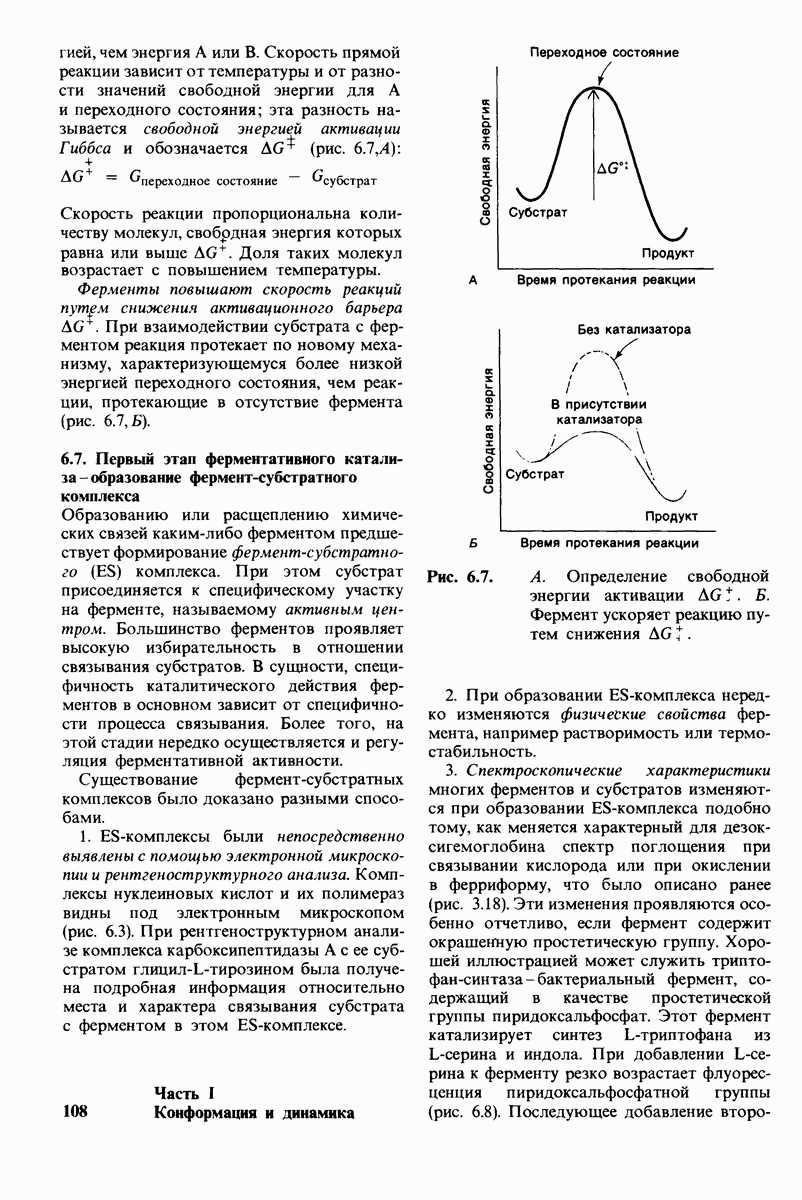
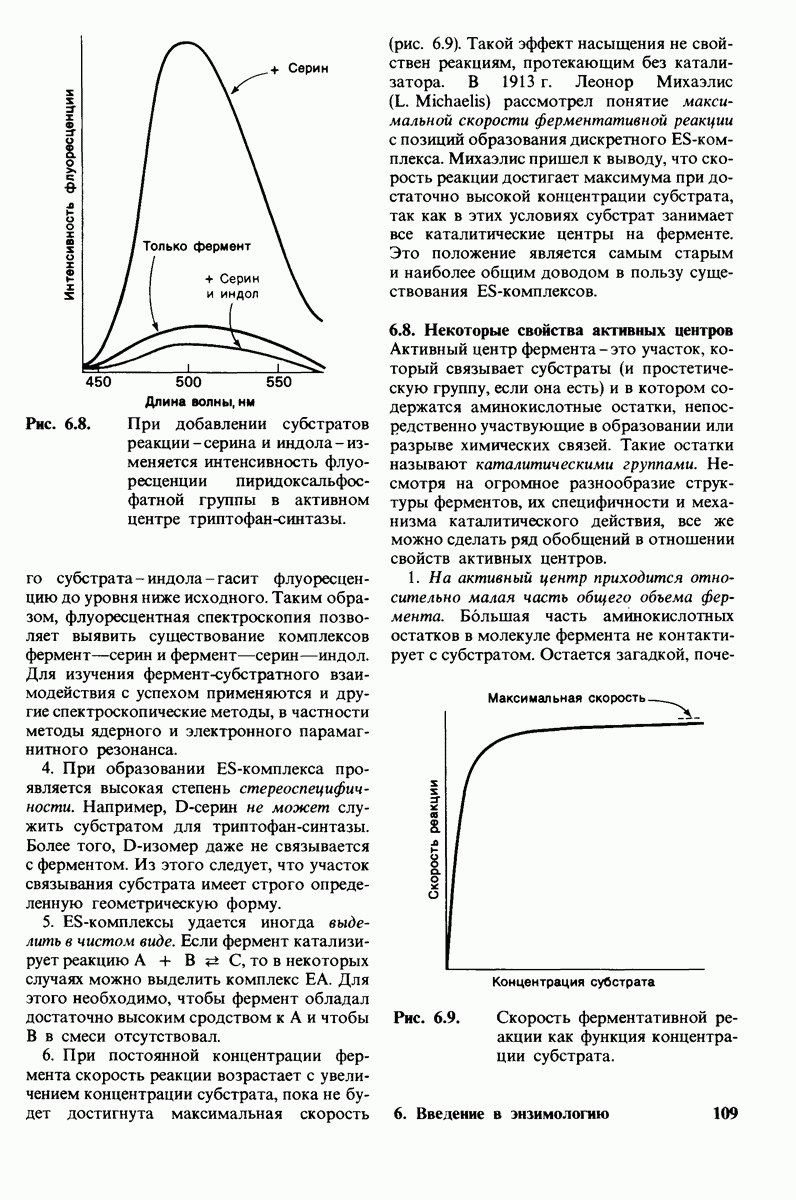
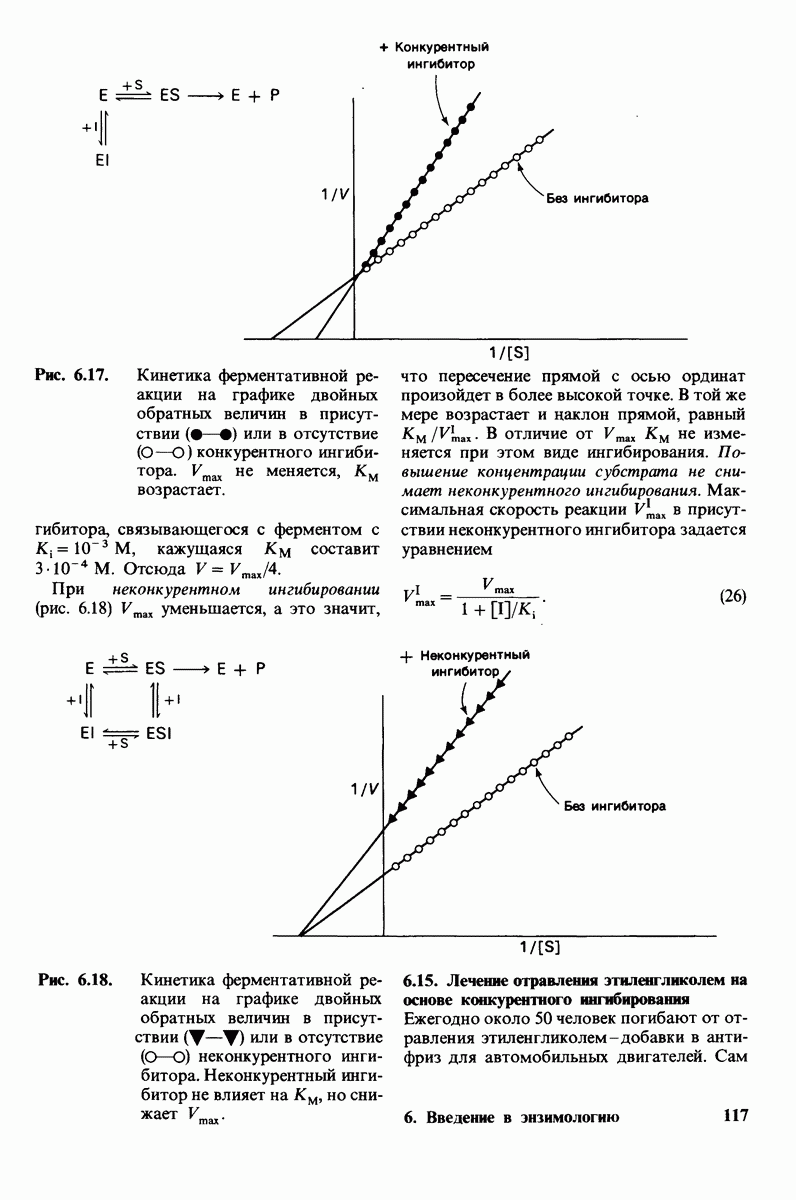
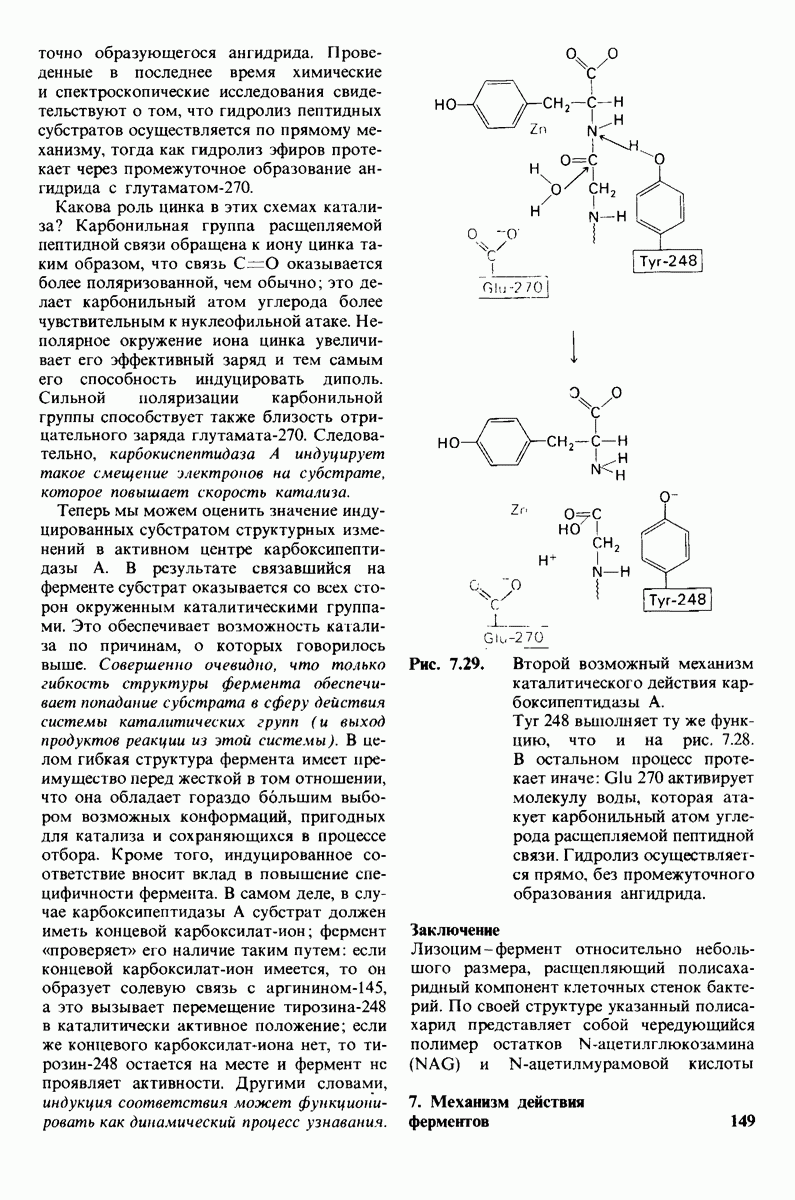
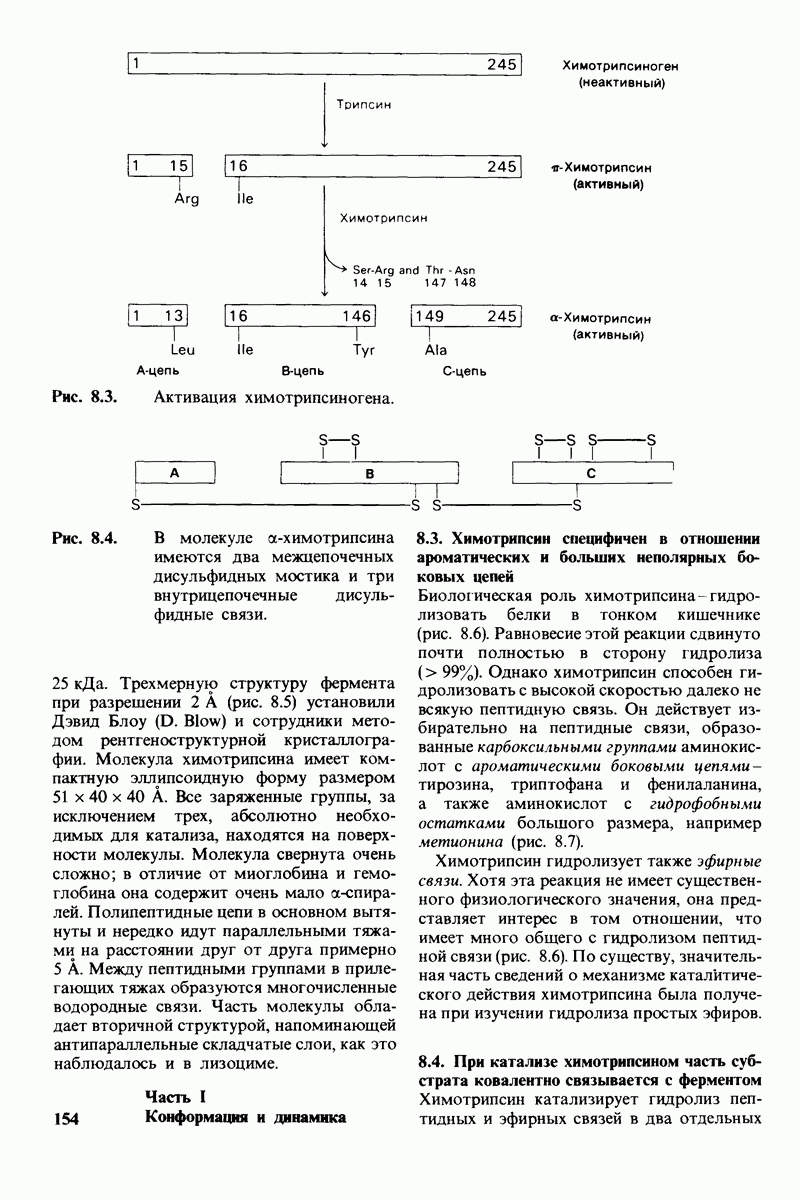
Обратимся теперь к вопросу о том, каким образом расщепление всего одной пептидной связи в химотрипсиногене превращает его в активный фермент. Трехмерную структуру химотрипсиногема исследовал Джозеф Краут (J. Kraut), установивший, что в процессе активации фермента его конформация притерпевает ряд изменений.
1. Гидролиз пептидной связи между аргинином-15 и изо лейцином-16 создает новые С- и N-концевые группы.
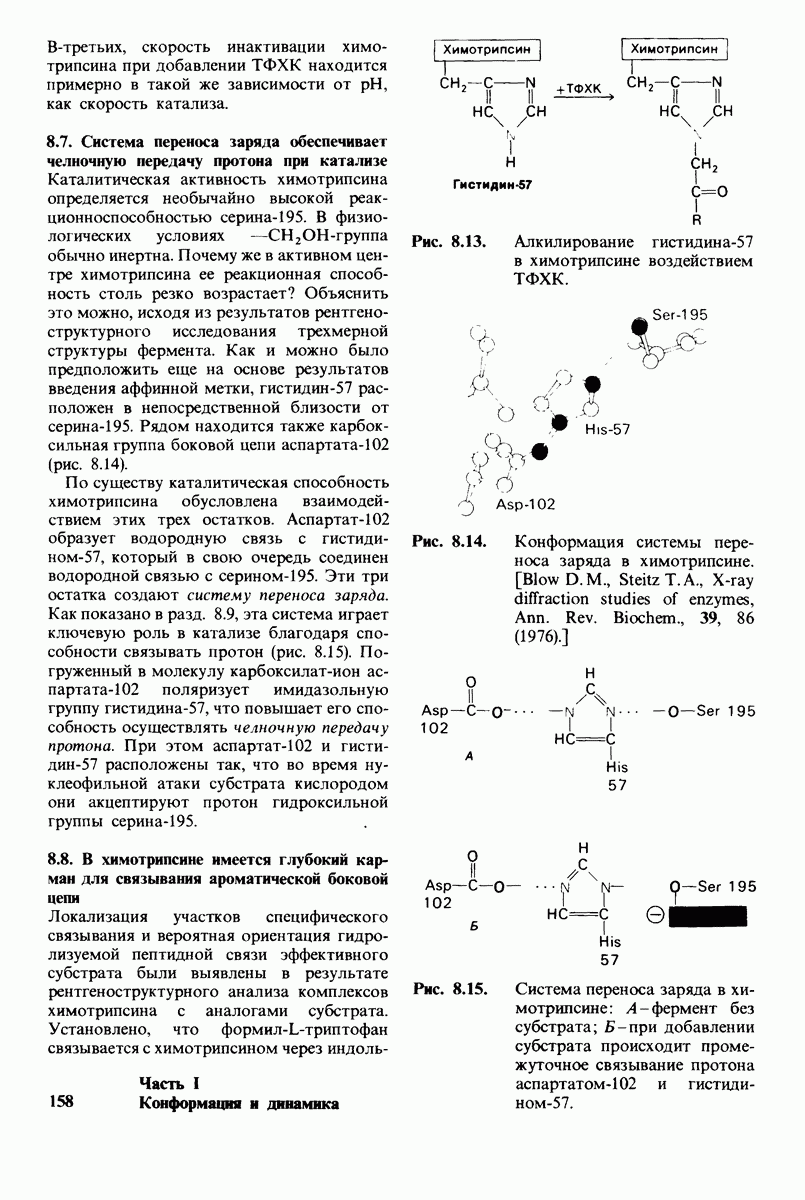
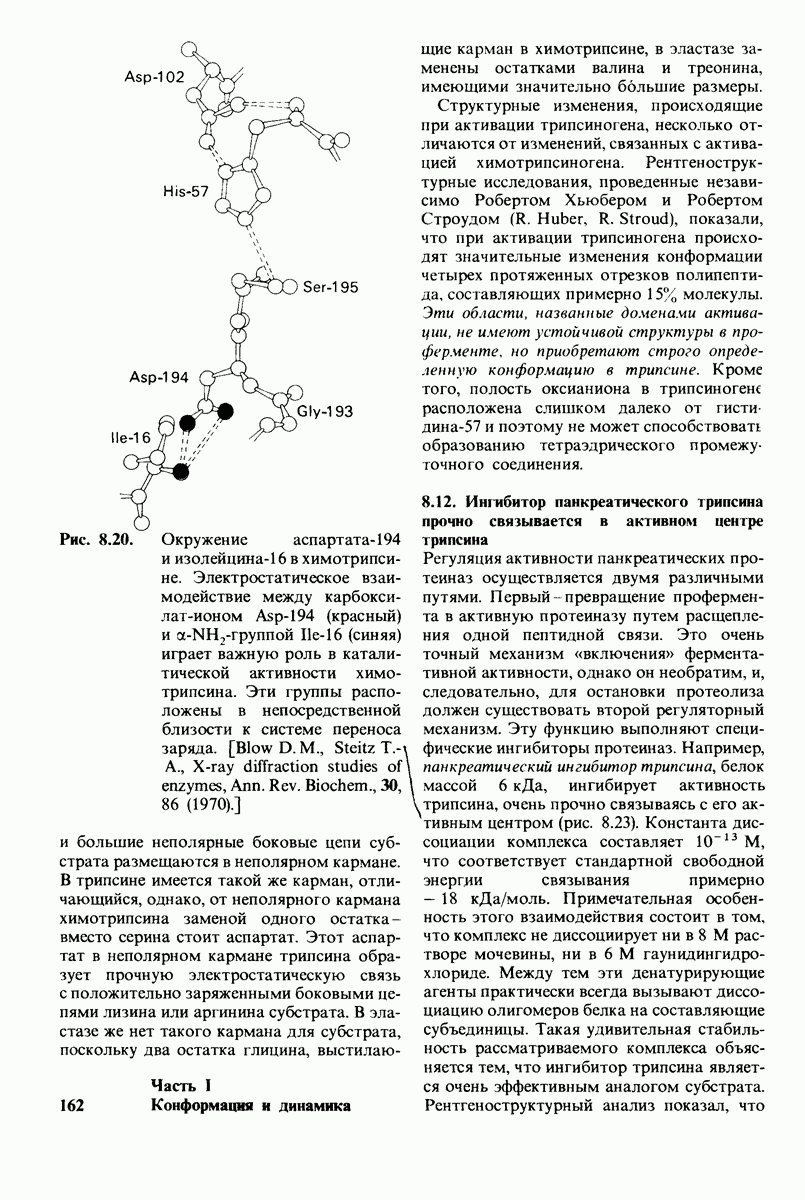
2. Вновь образованная N-концевая группа изолейцина-16 загибается внутрь и взаимодействует с аспартатом-194 внутри молекулы химотрипсина (рис. 8.20). Протонирование этой аминогруппы стабилизирует активную форму химотрипсина, о чем свидетельствует характер зависимости ферментативной активности от рН.
3. Это электростатическое взаимодействие между положительно заряженной аминогруппой и отрицательно заряженным карбоксилат-ионом, происходящее в неполярной области фермента, запускает ряд конформационных сдвигов. Метионин-192 из глубины молекулы перемещается ближе к поверхности, а остатки 187 и 193 становятся более вытянутыми. В результате образуется субстрат-специфичный участок для ароматических и больших неполярных групп. Одну сторону этого участка составляют остатки от 189 до 192. В проферменте эта полость для связывания части субстрата сформирована не полностью.
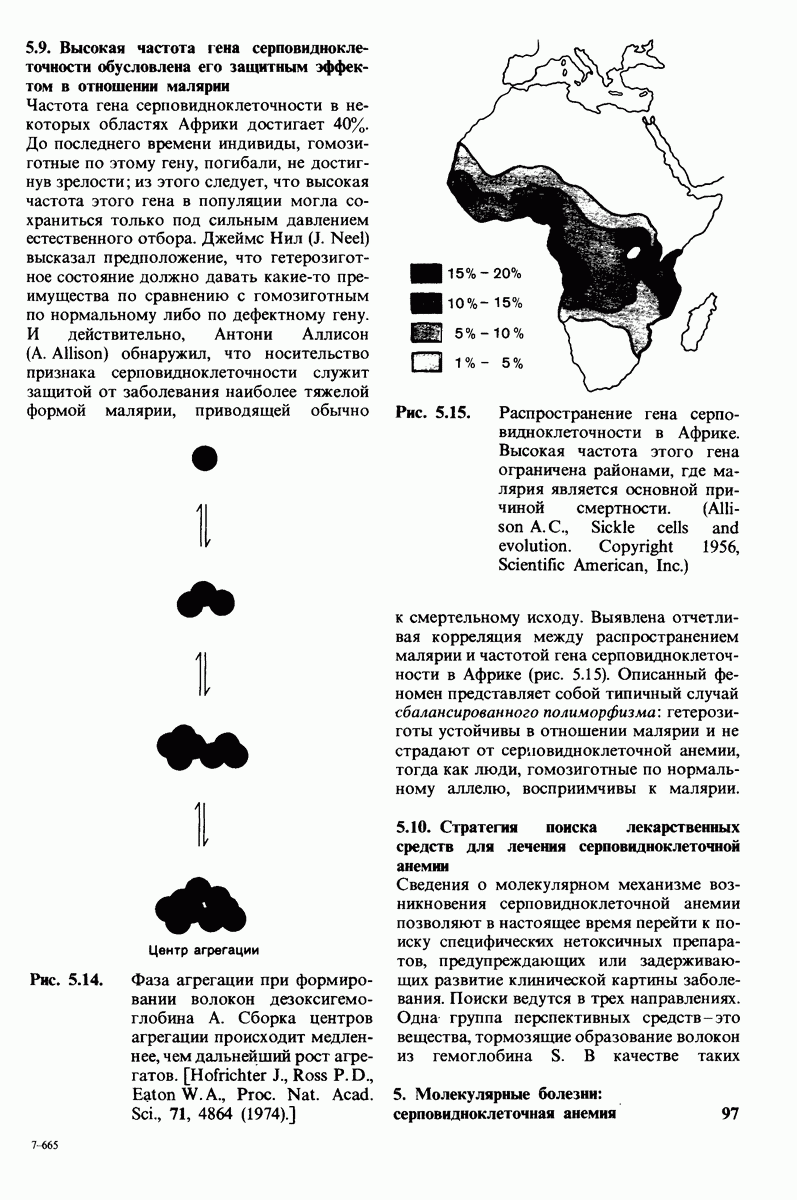
4. Тетраэдрическое переходное состояние, возникающее при катализе химотрипсином, стабилизируется водородными связями между отрицательно заряженным атомом кислорода карбоксильной группы и двумя NH-группами самой полипептидной цепи (рис. 8.17). В химотрипсиногене одна из этих NH-групп расположена так, что недоступна для связывания. Следовательно, в зимогене и полость оксианиона до конца не сформирована.

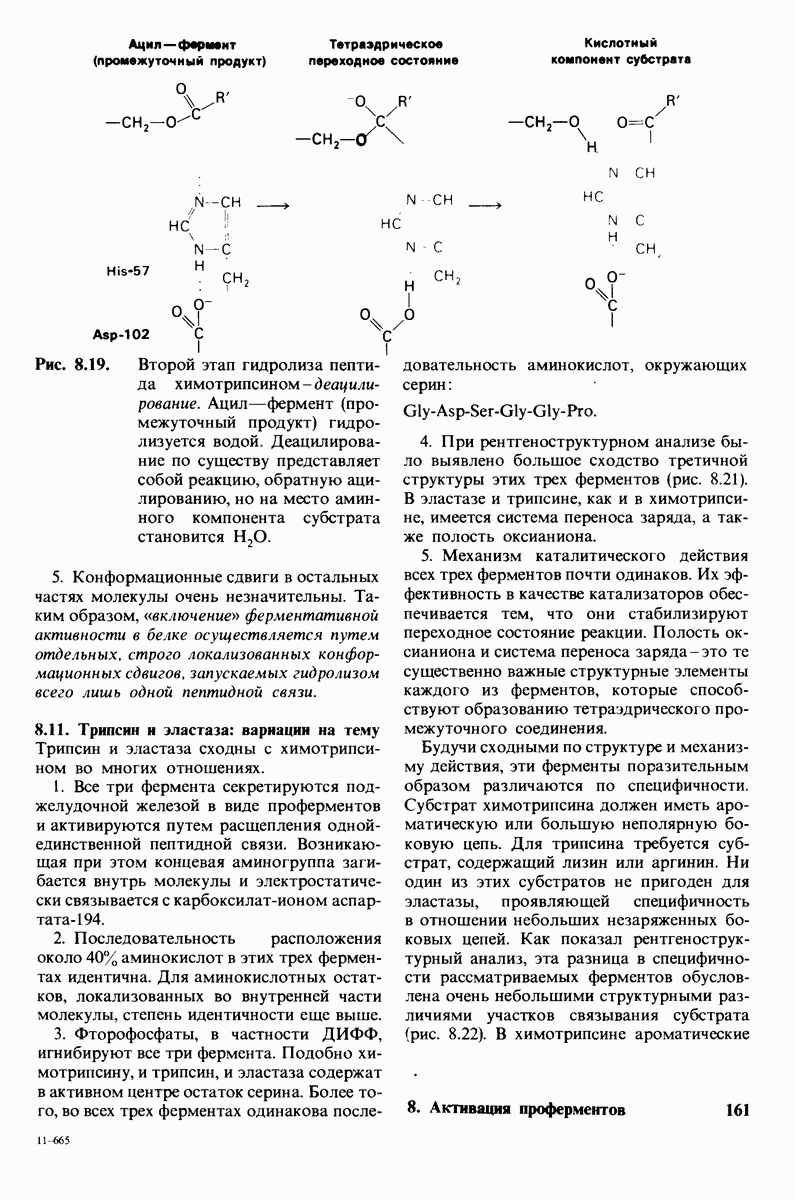
Рис. 8.19.
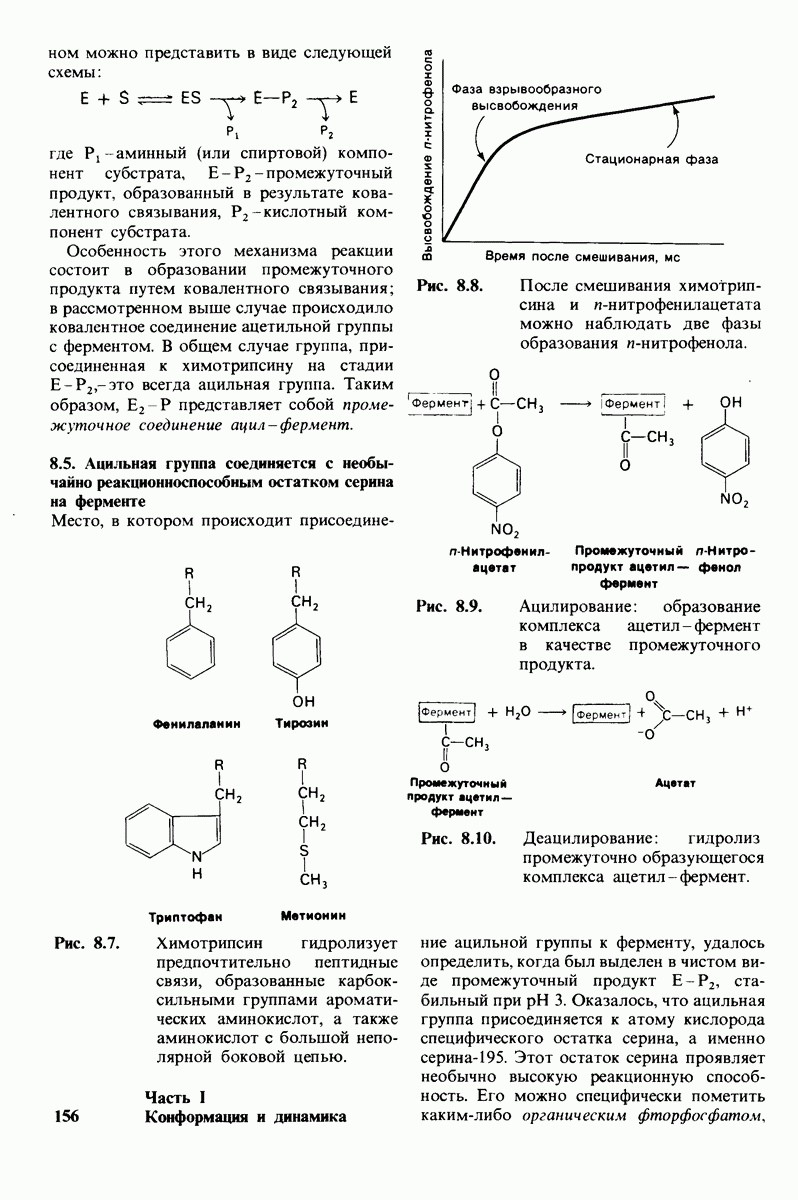
Второй этап гидролиза пептида химотрипсин ом - деацилирование. Ацил—фермент (промежуточный продукт) гидролиз уется водой. Деацилирование по существу представляет собой реакцию, обратную ацилированию, но на место аминного компонента субстрата становится 
5. Конформационные сдвиги в остальных частях молекулы очень незначительны. Таким образом, «включение» ферментативной активности в белке осуществляется путем отдельных, строго локализованных конформационных сдвигов, запускаемых гидролизом всего лишь одной пептидной связи.