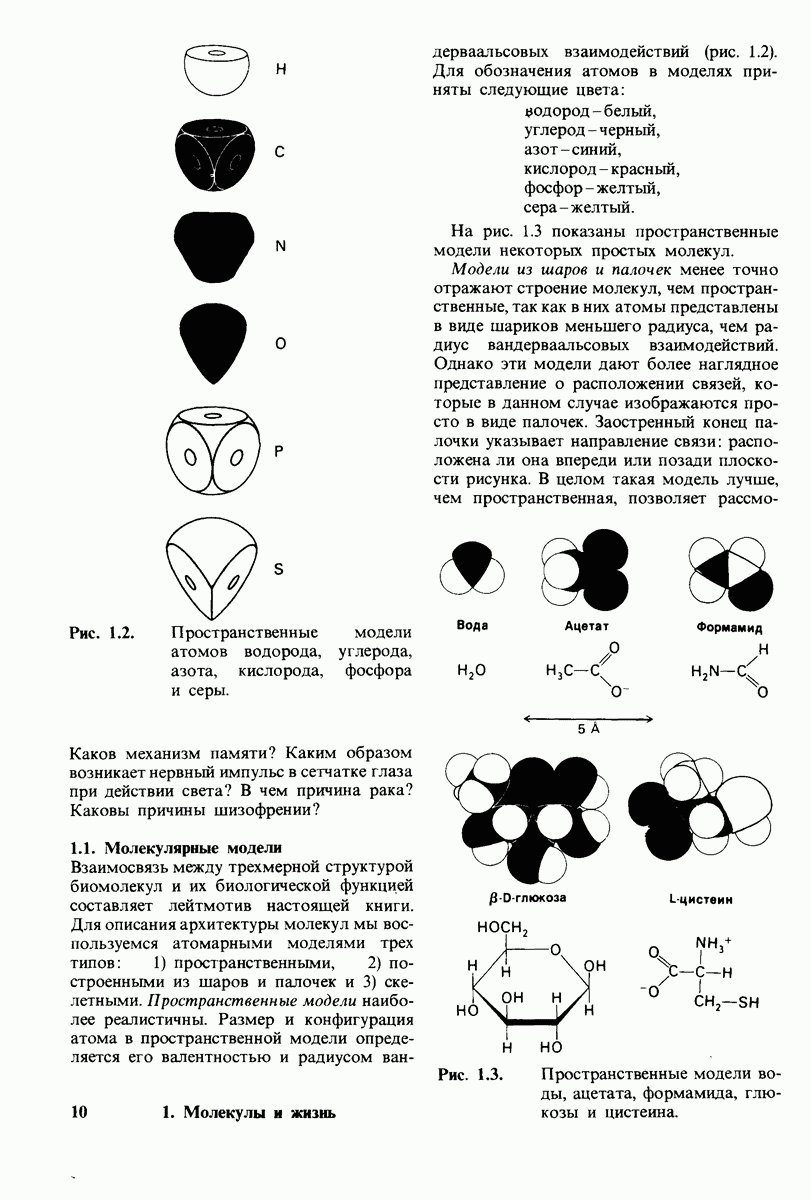
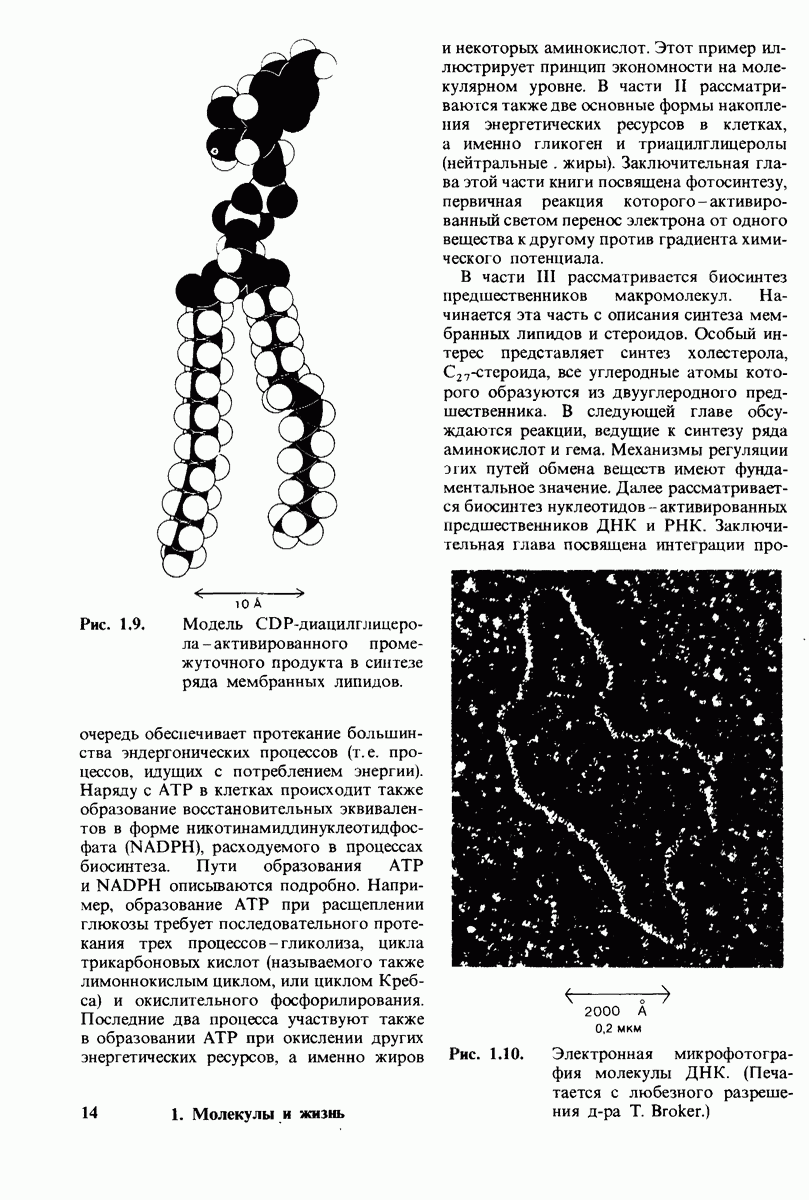
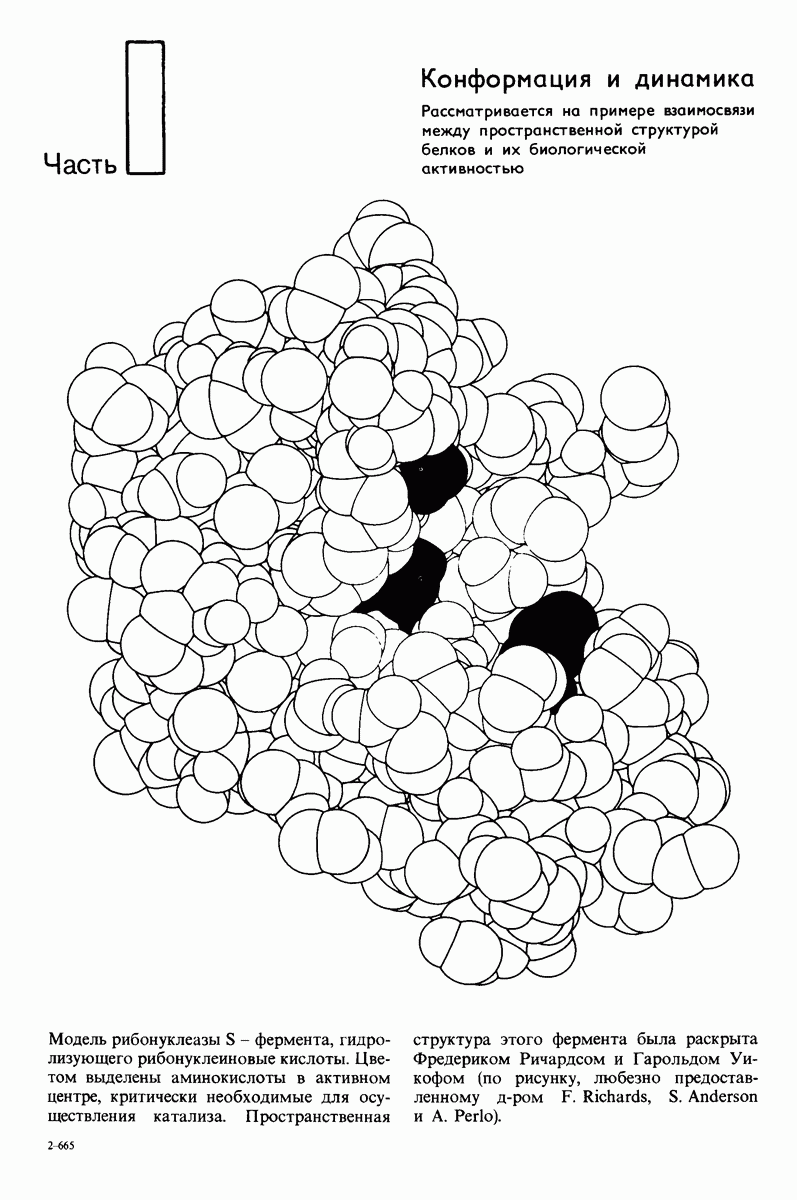
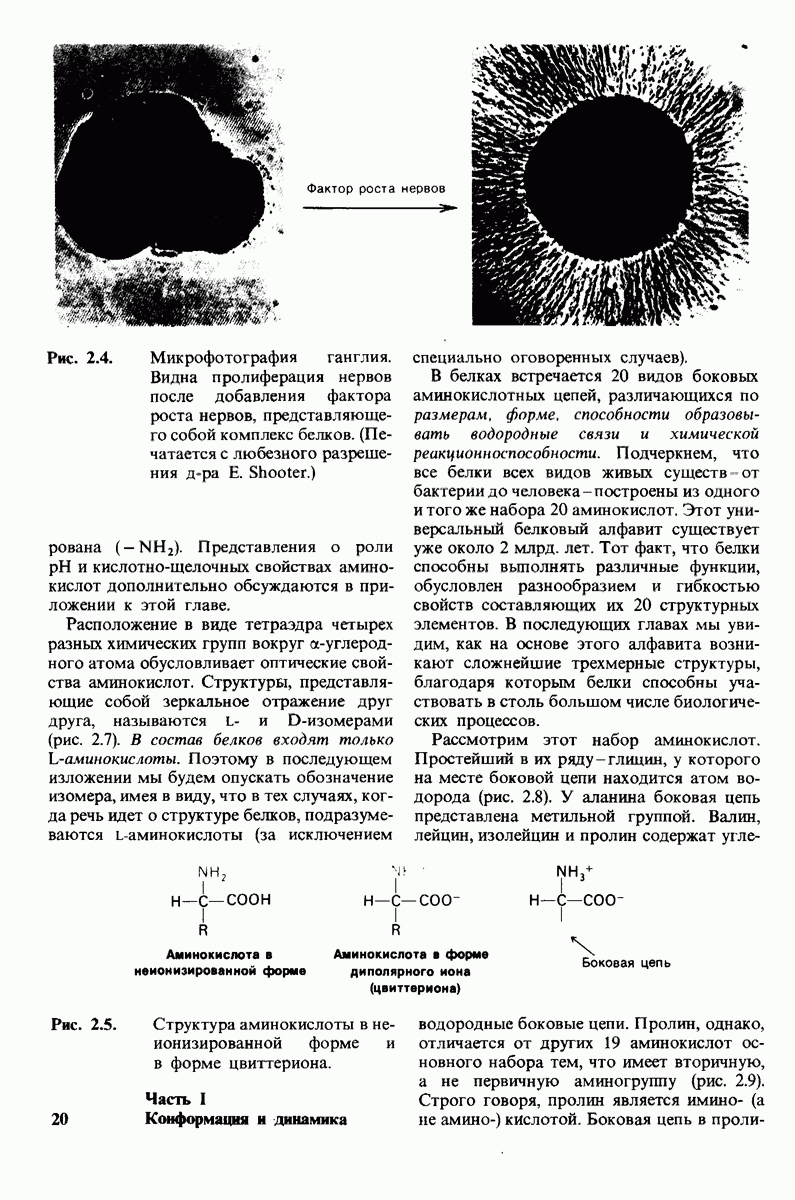
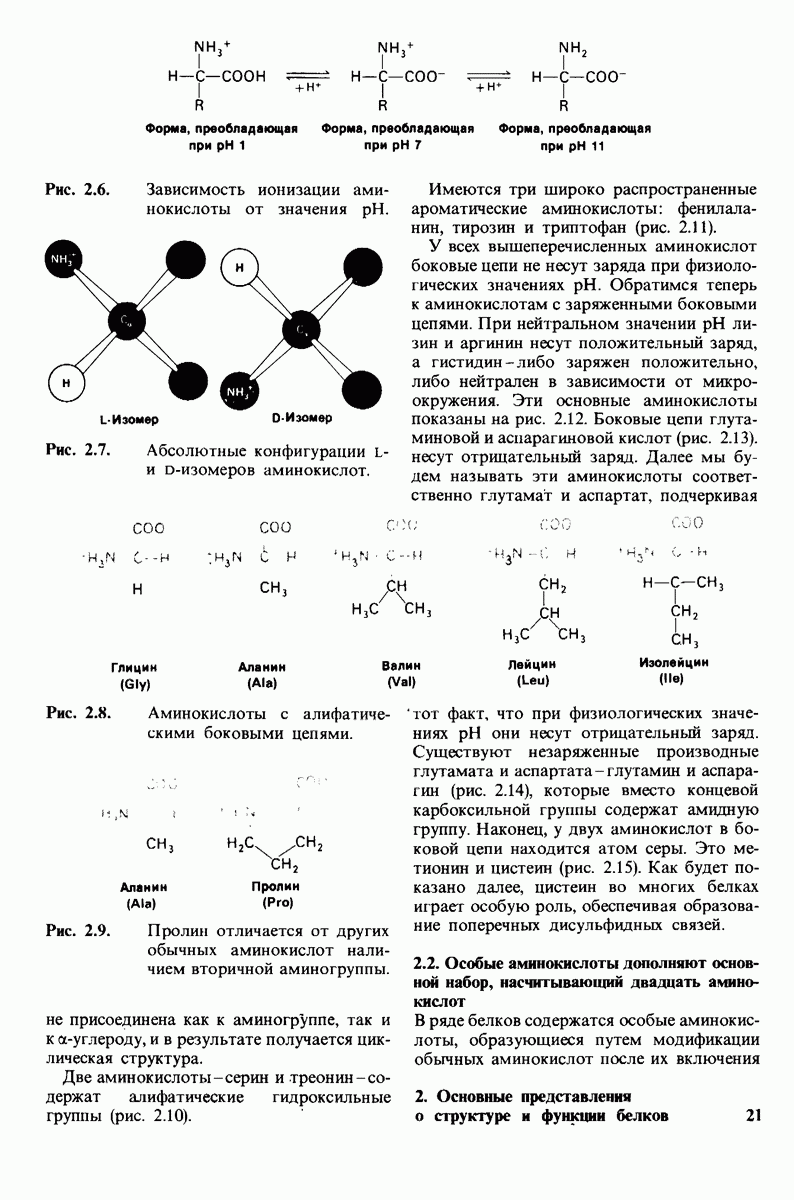
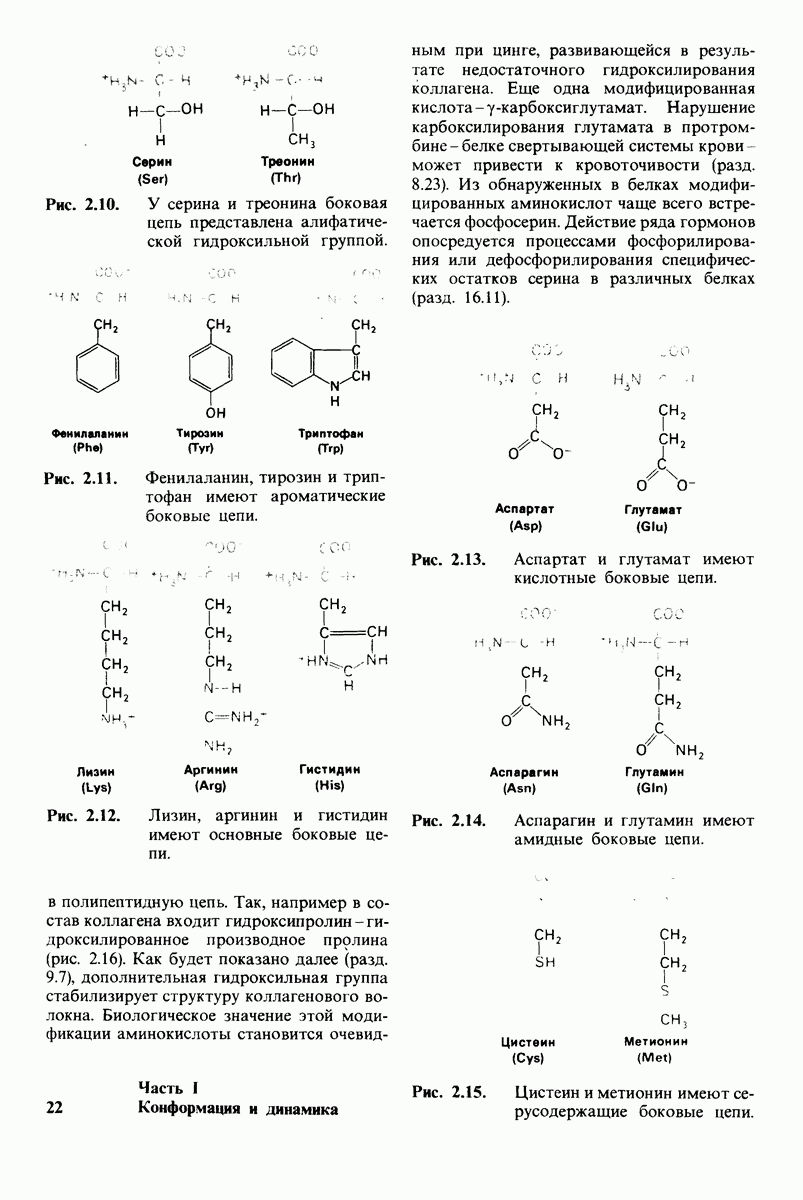
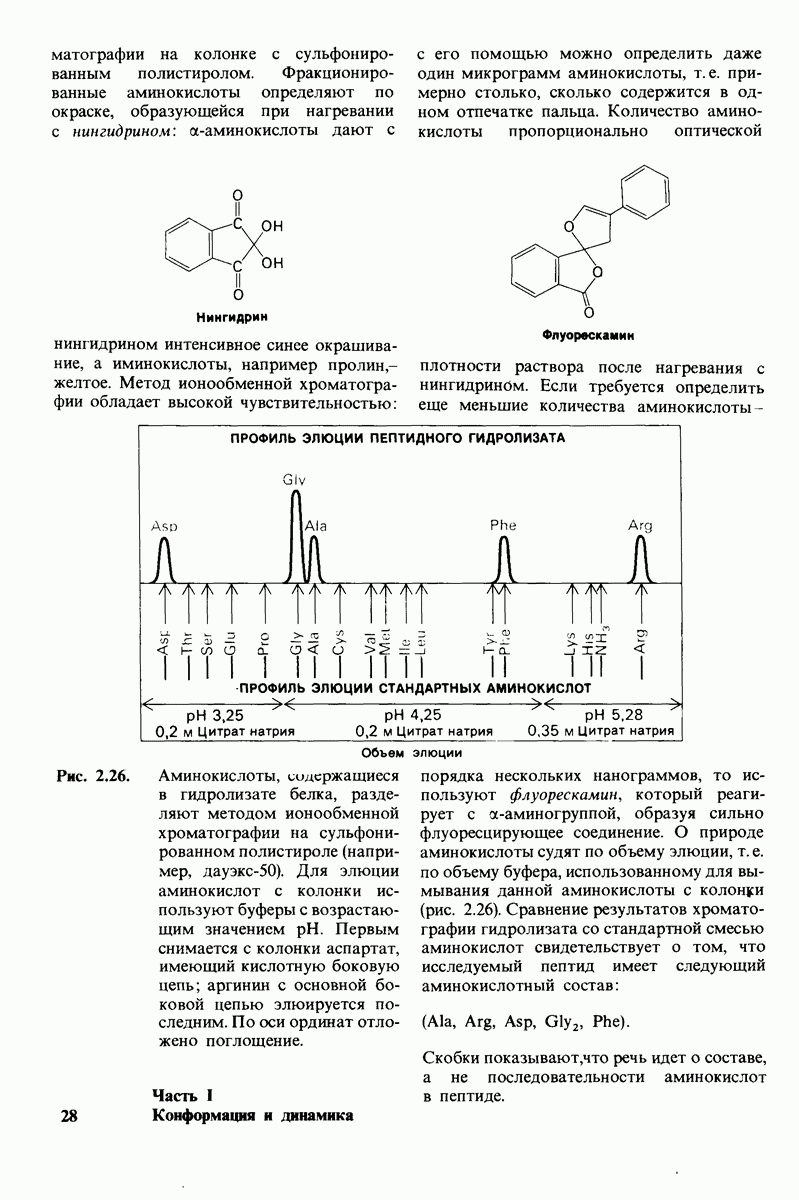
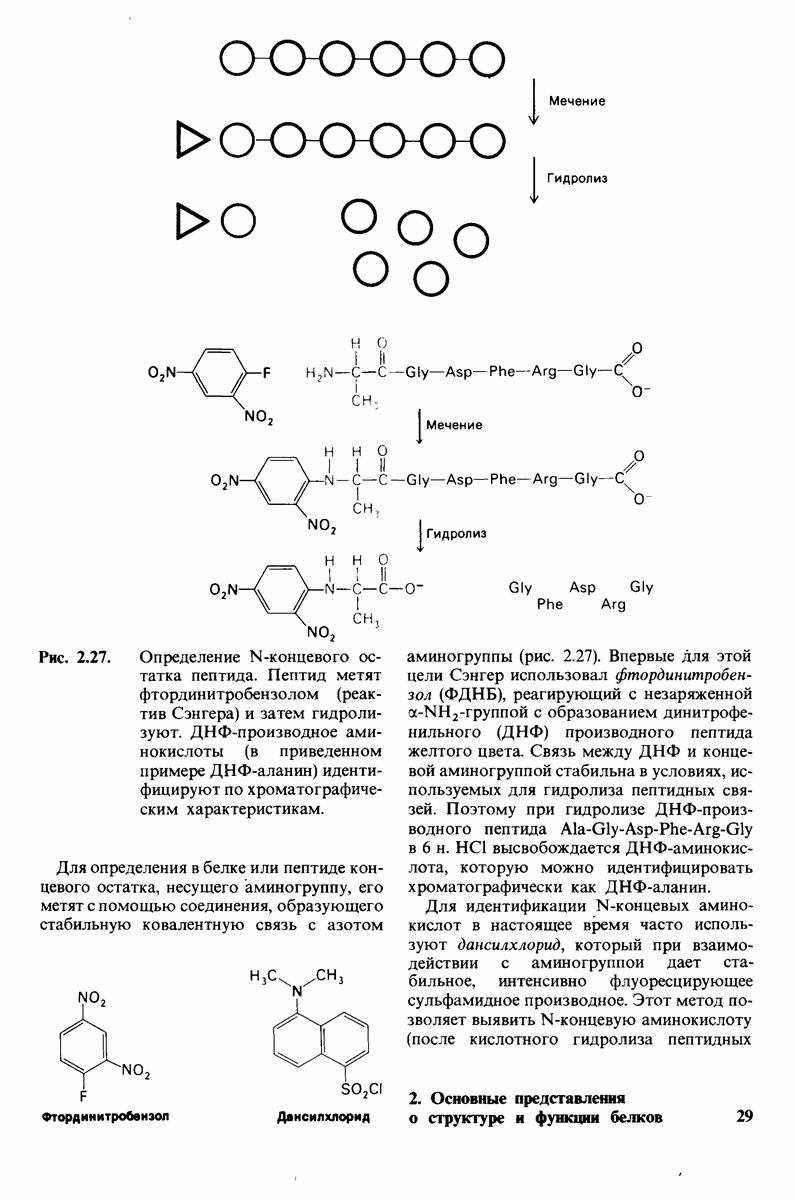
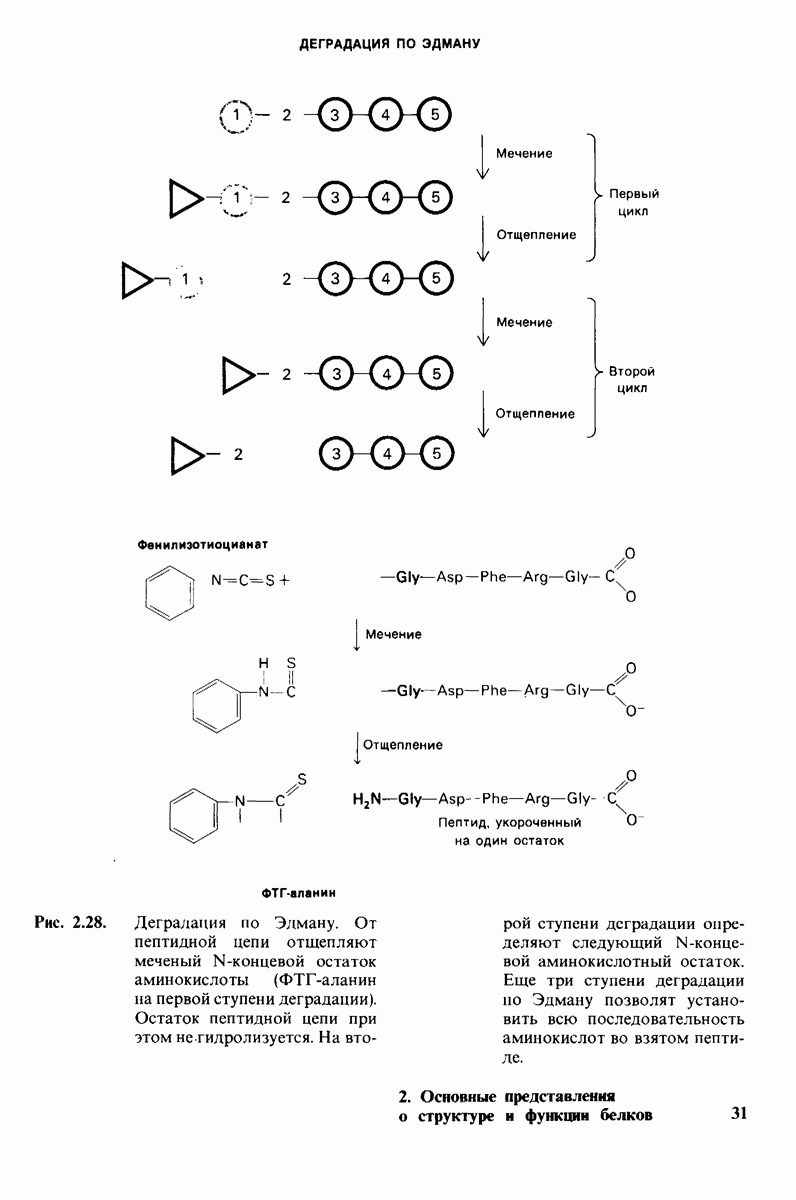
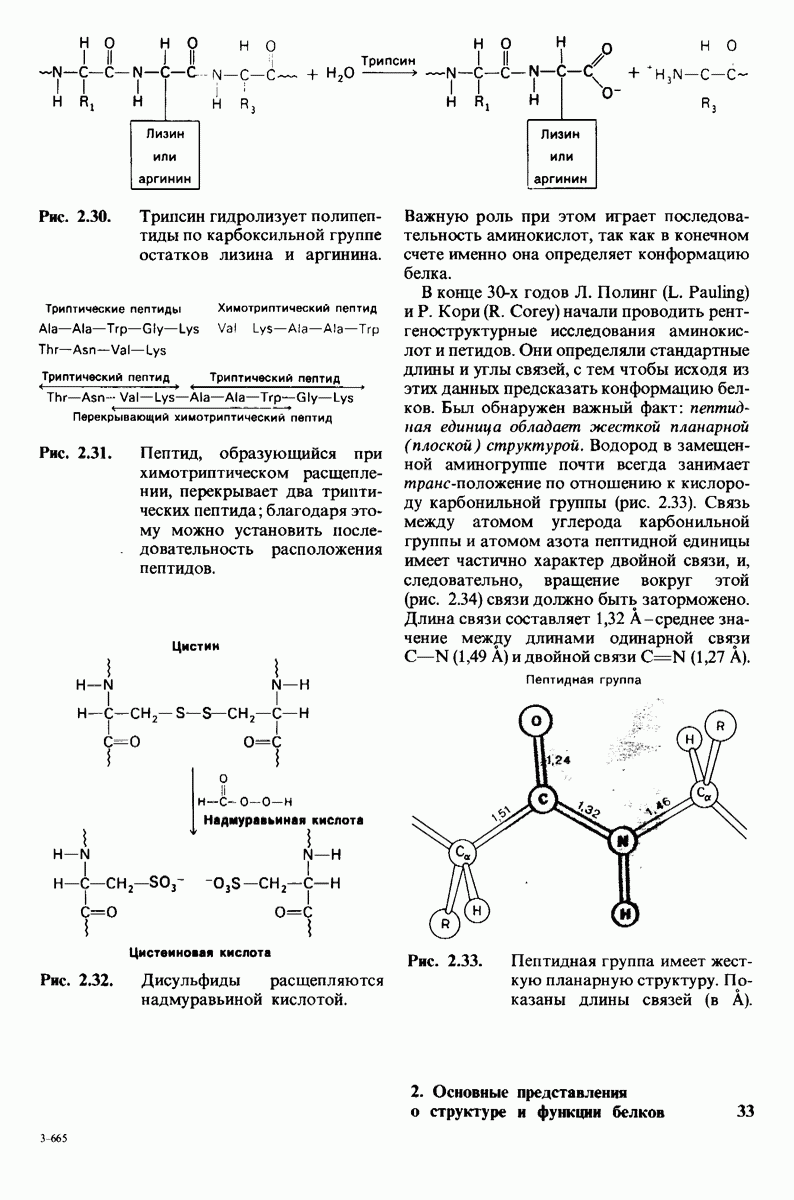
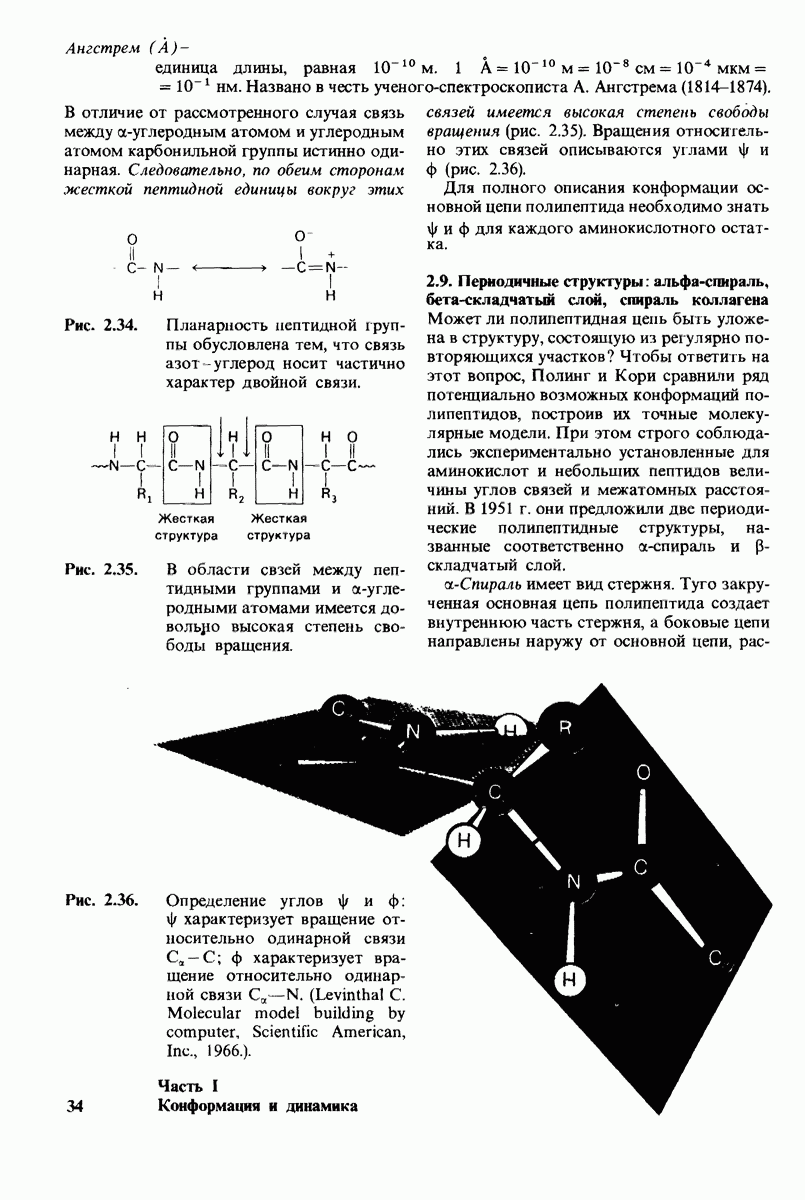
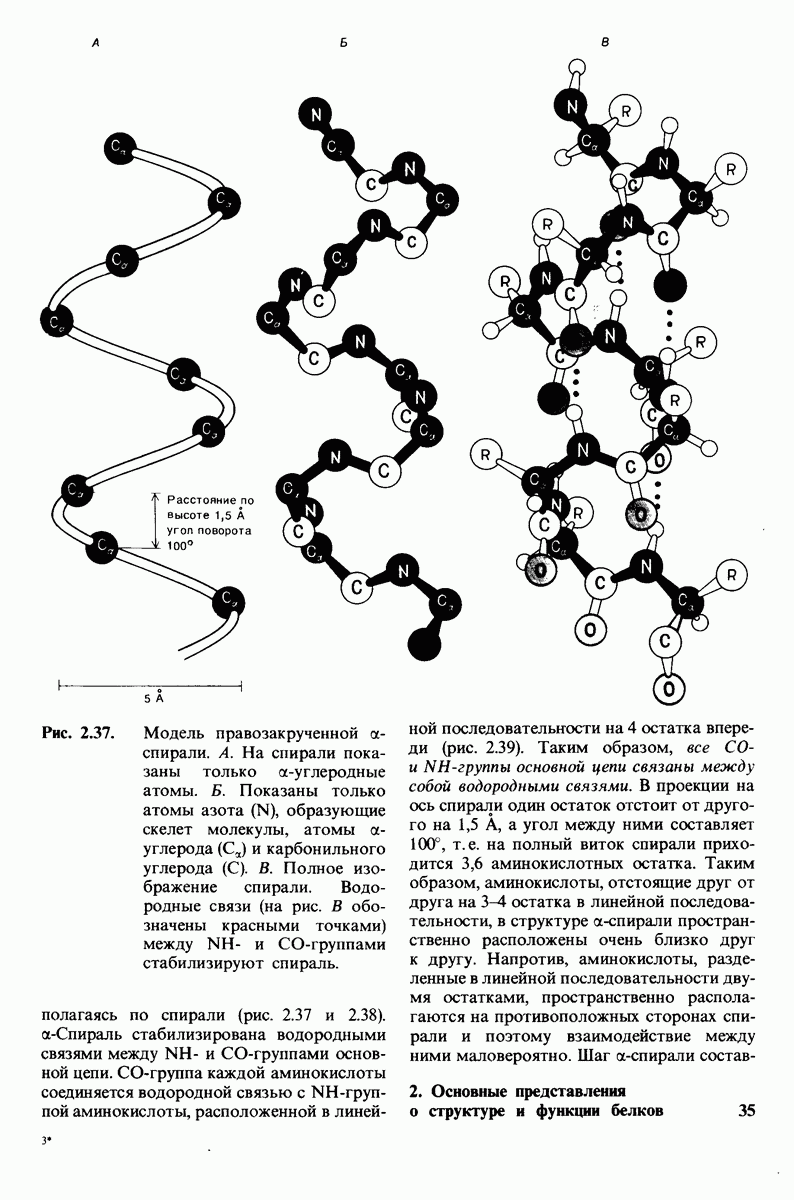
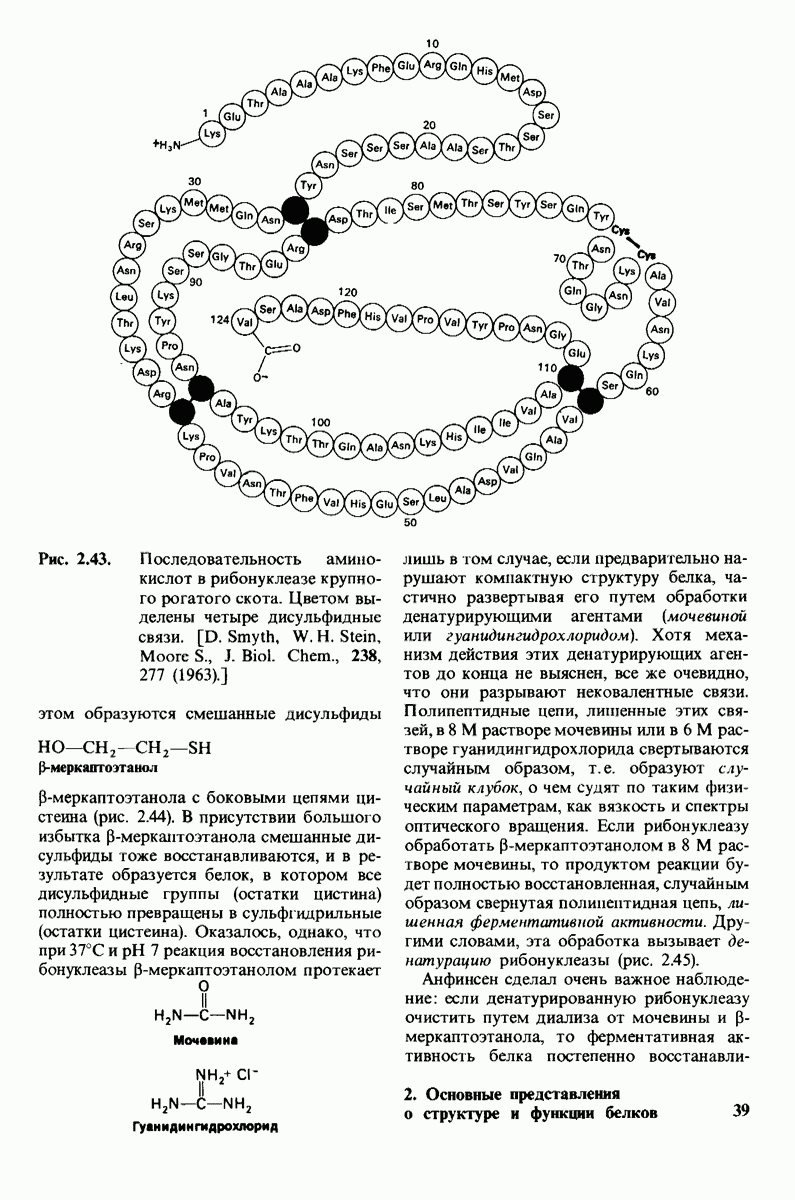
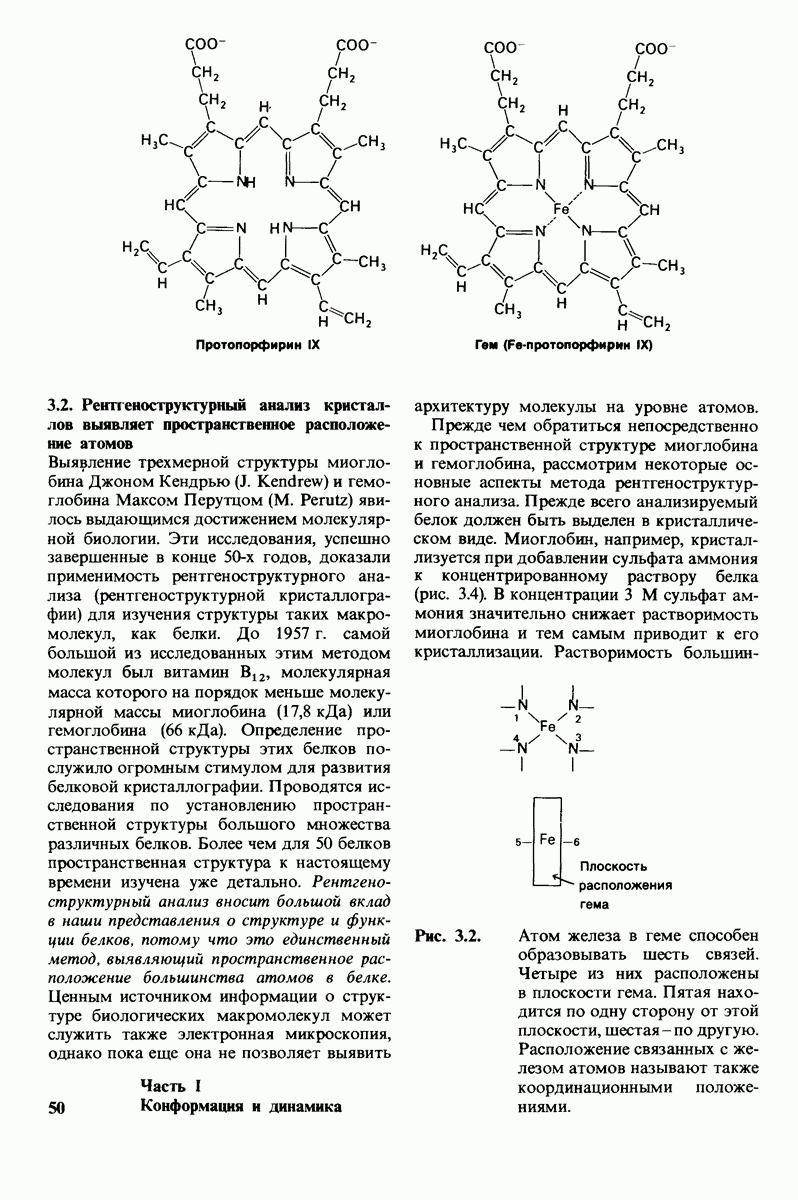
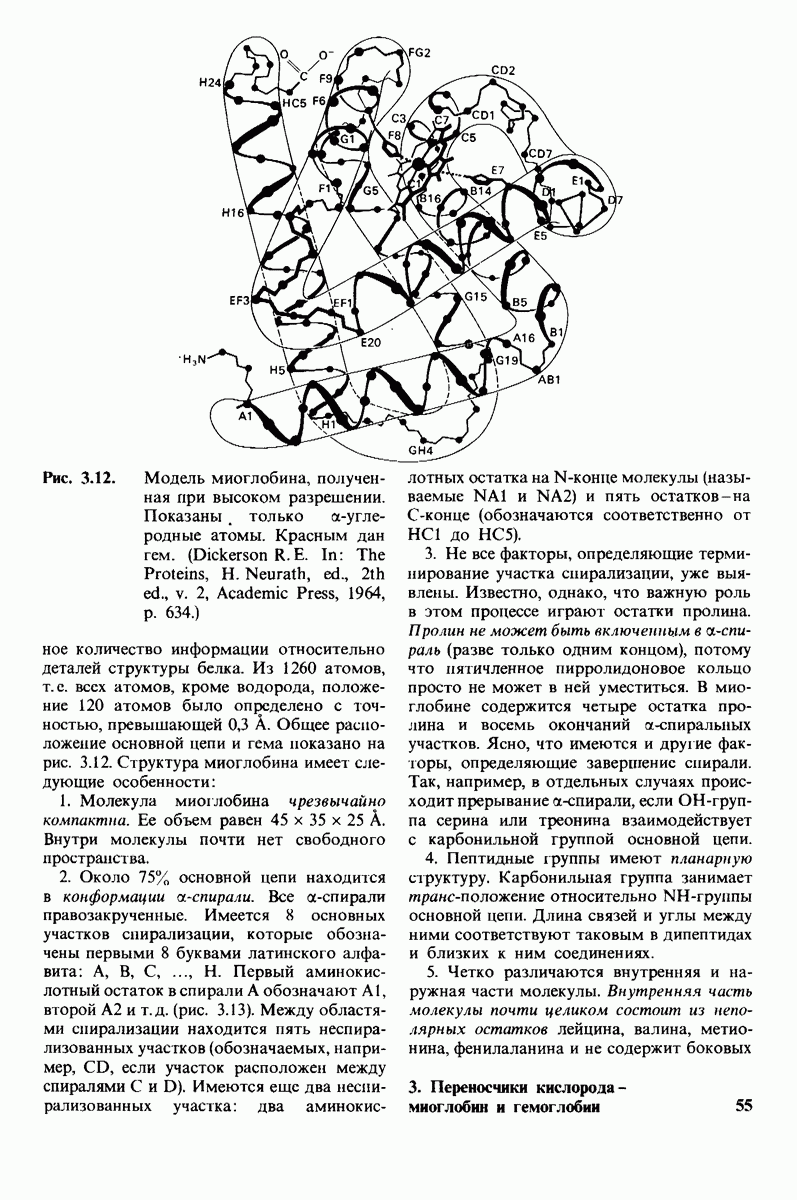
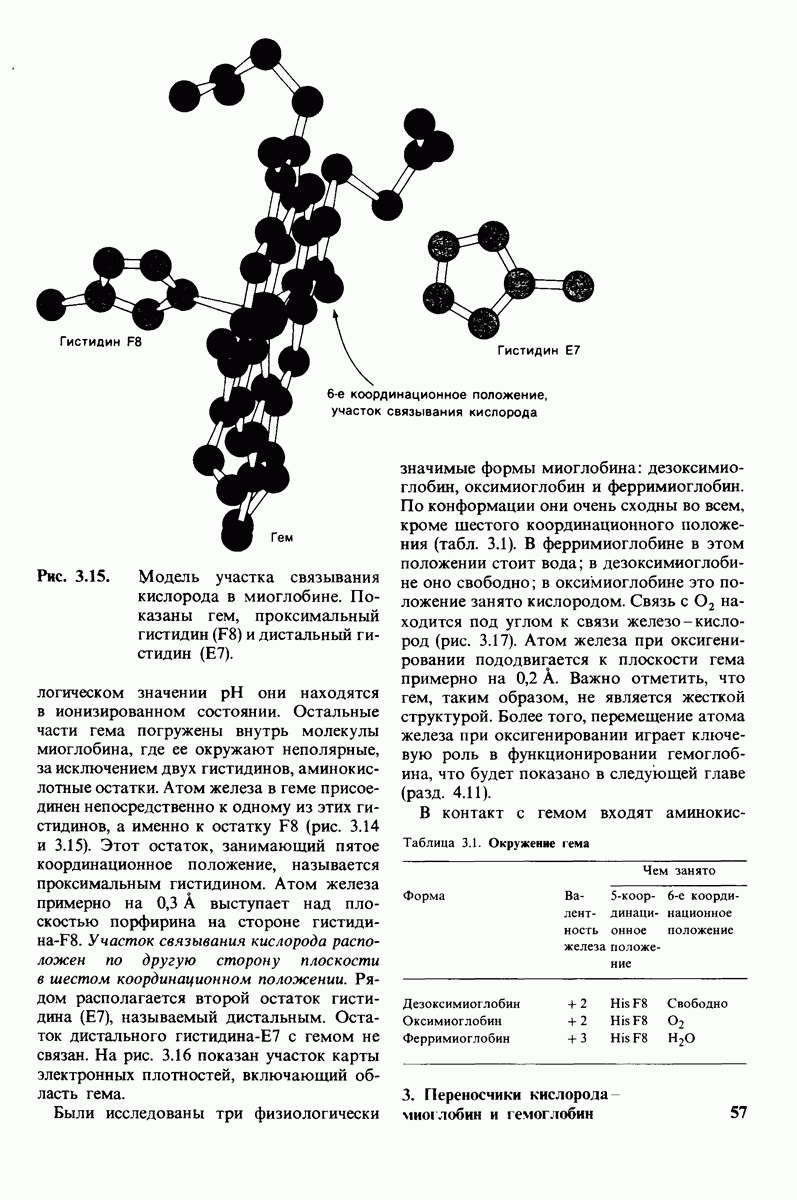
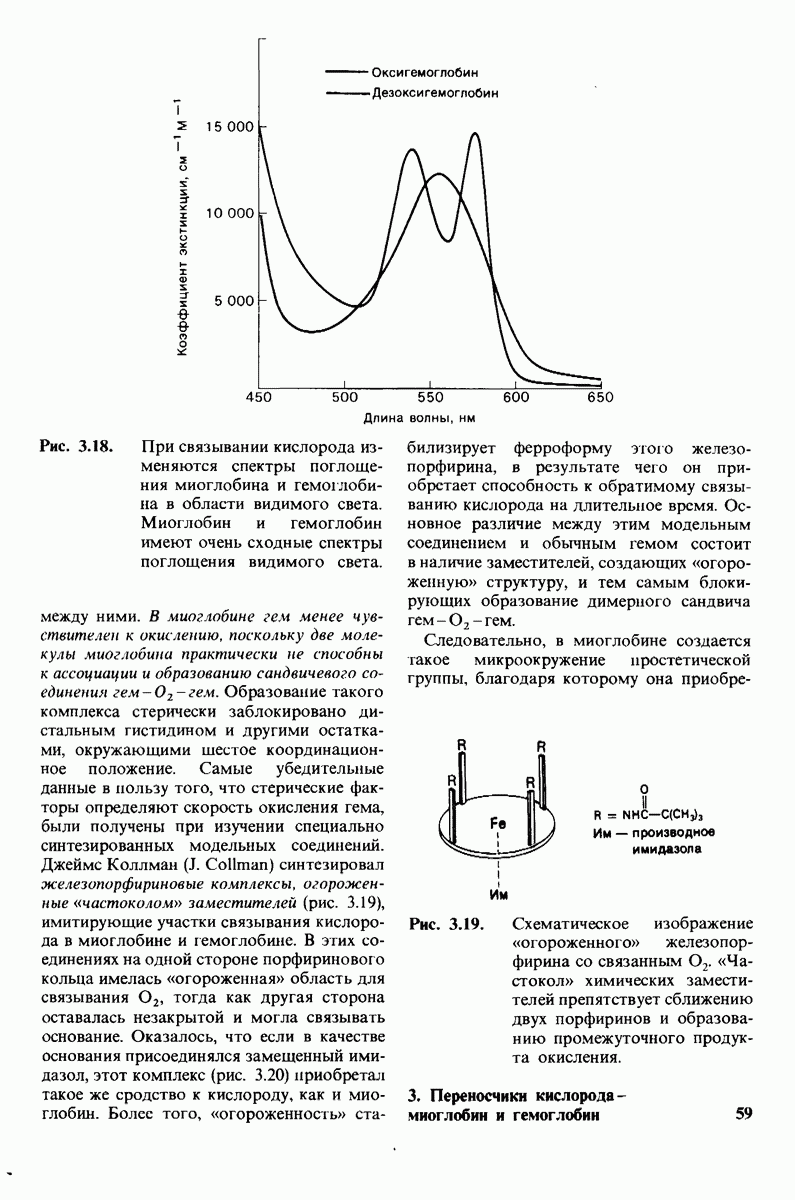
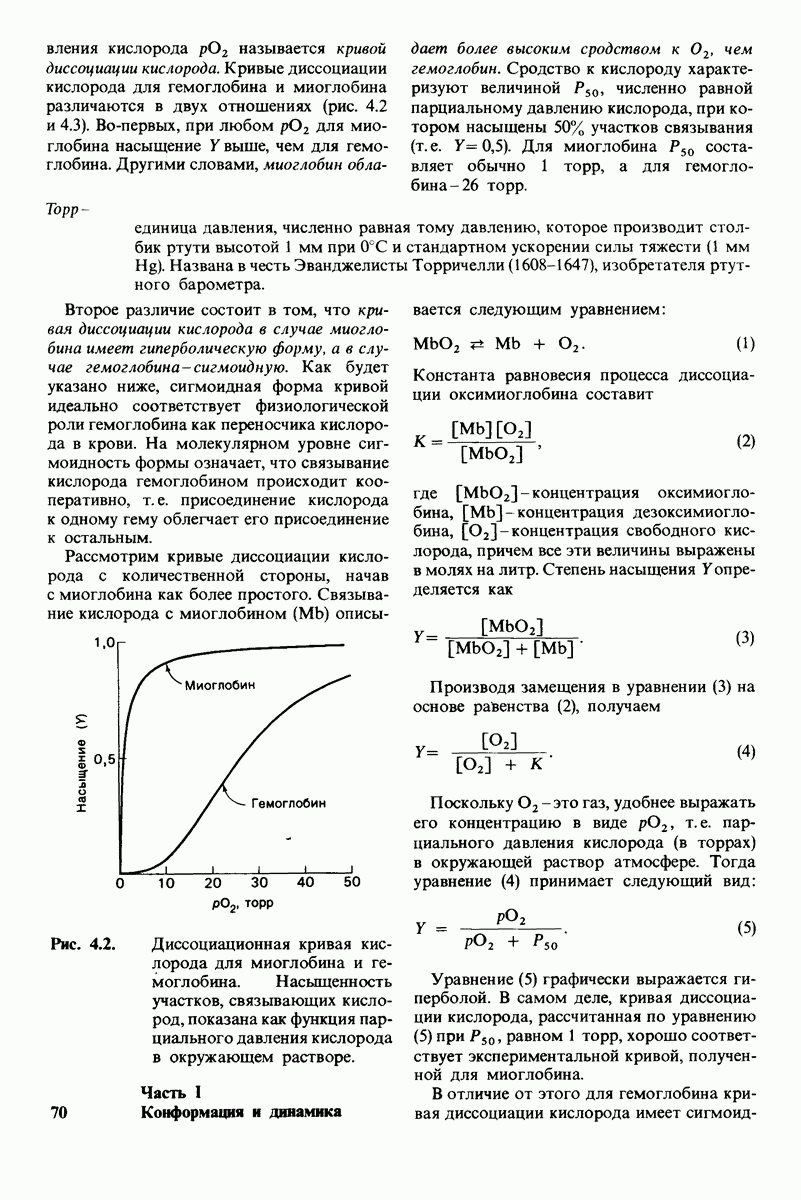
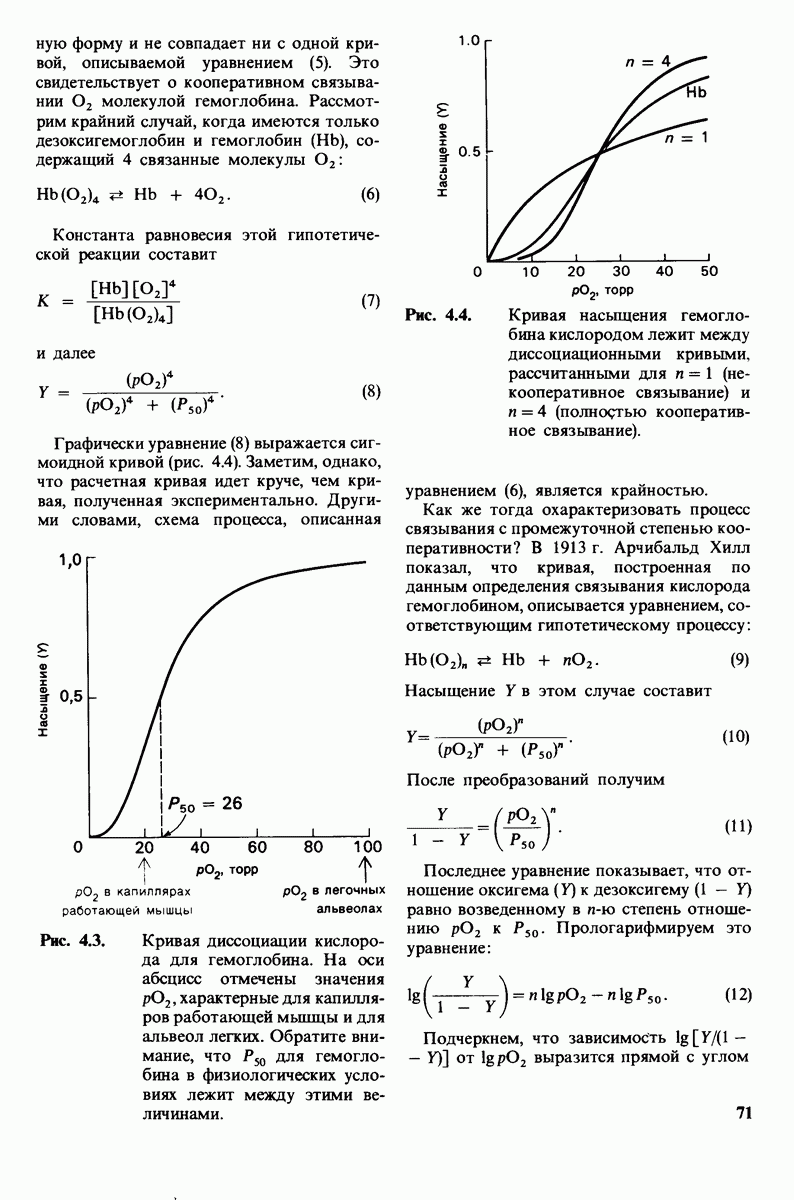
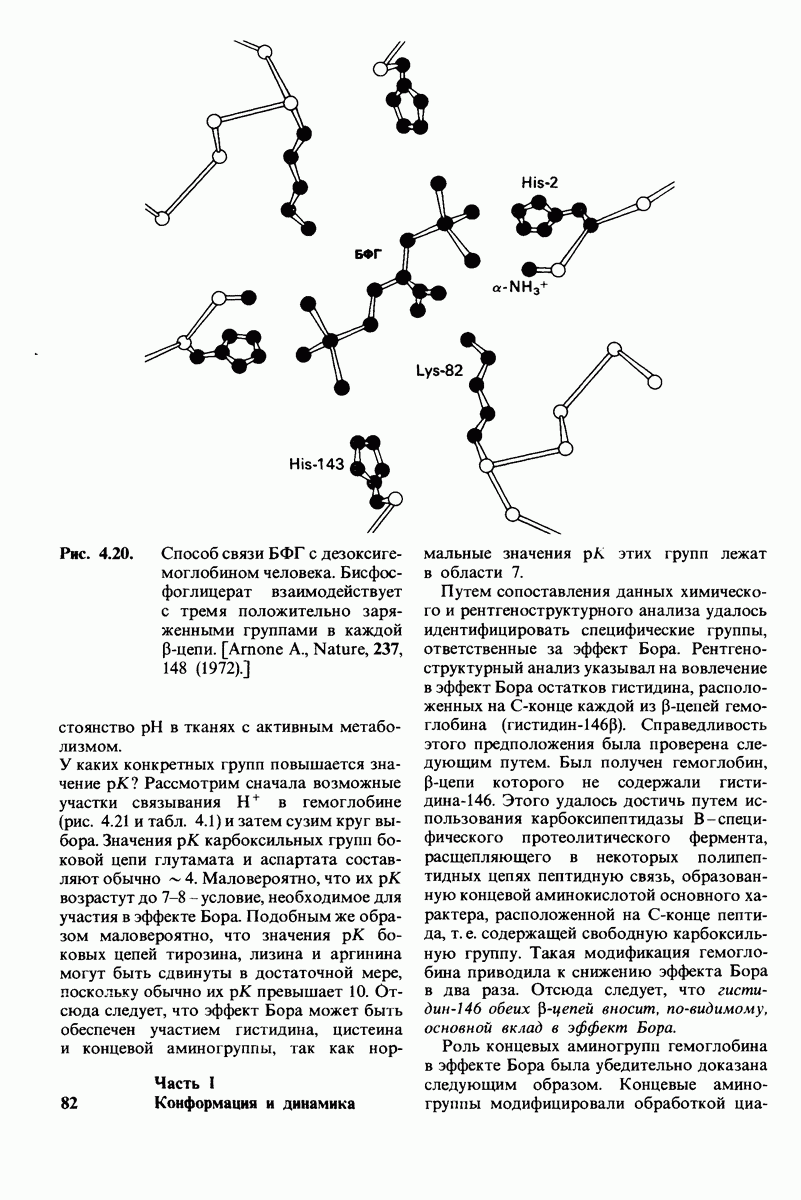
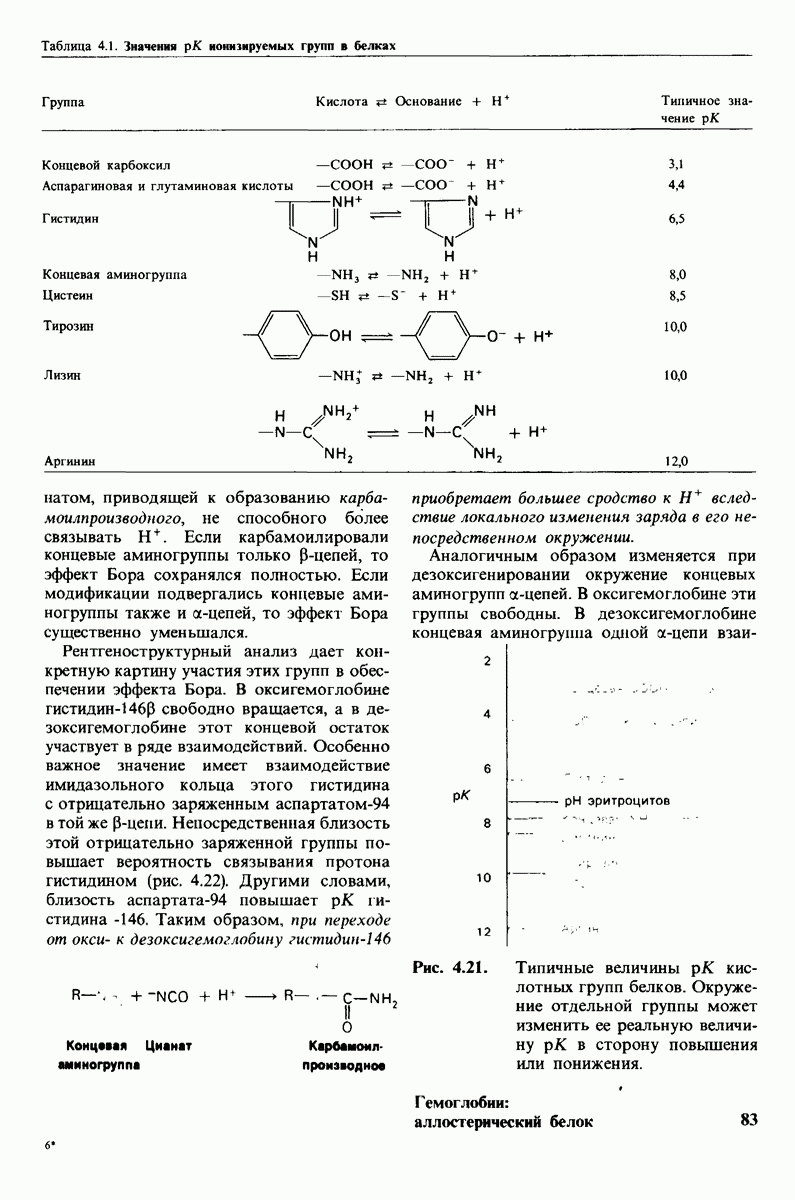
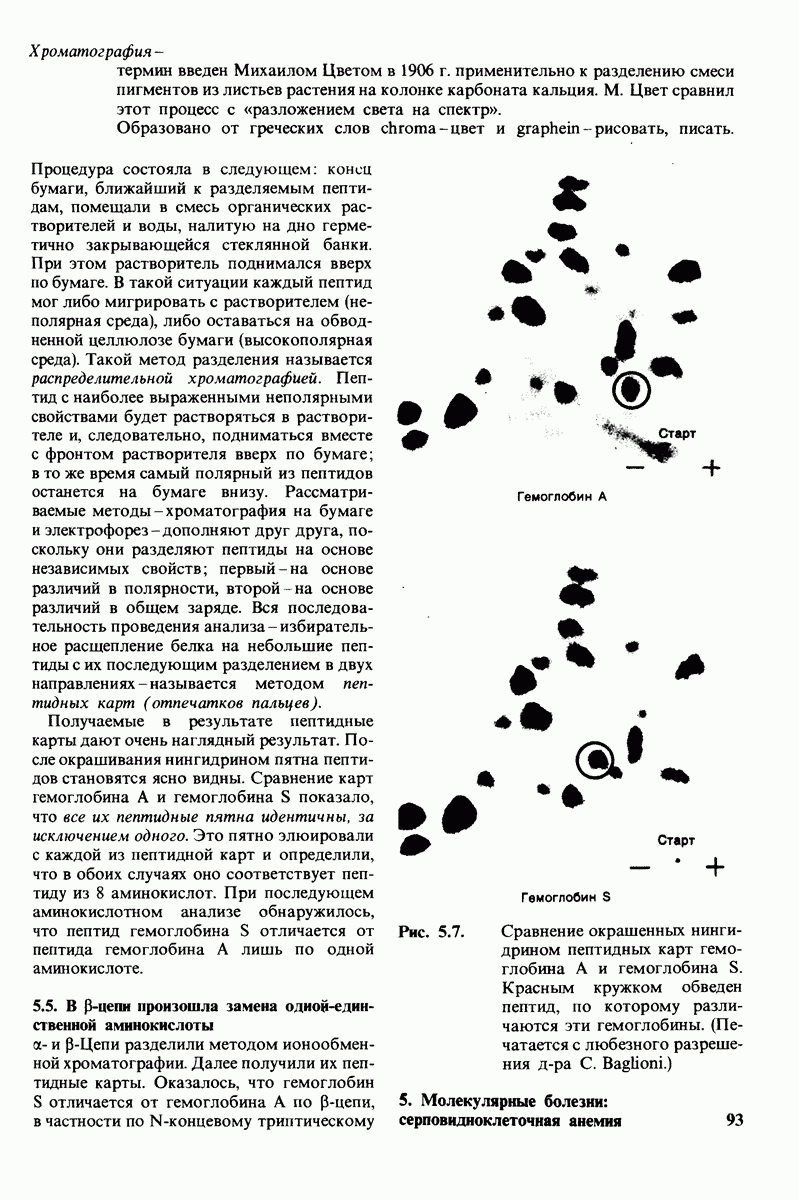
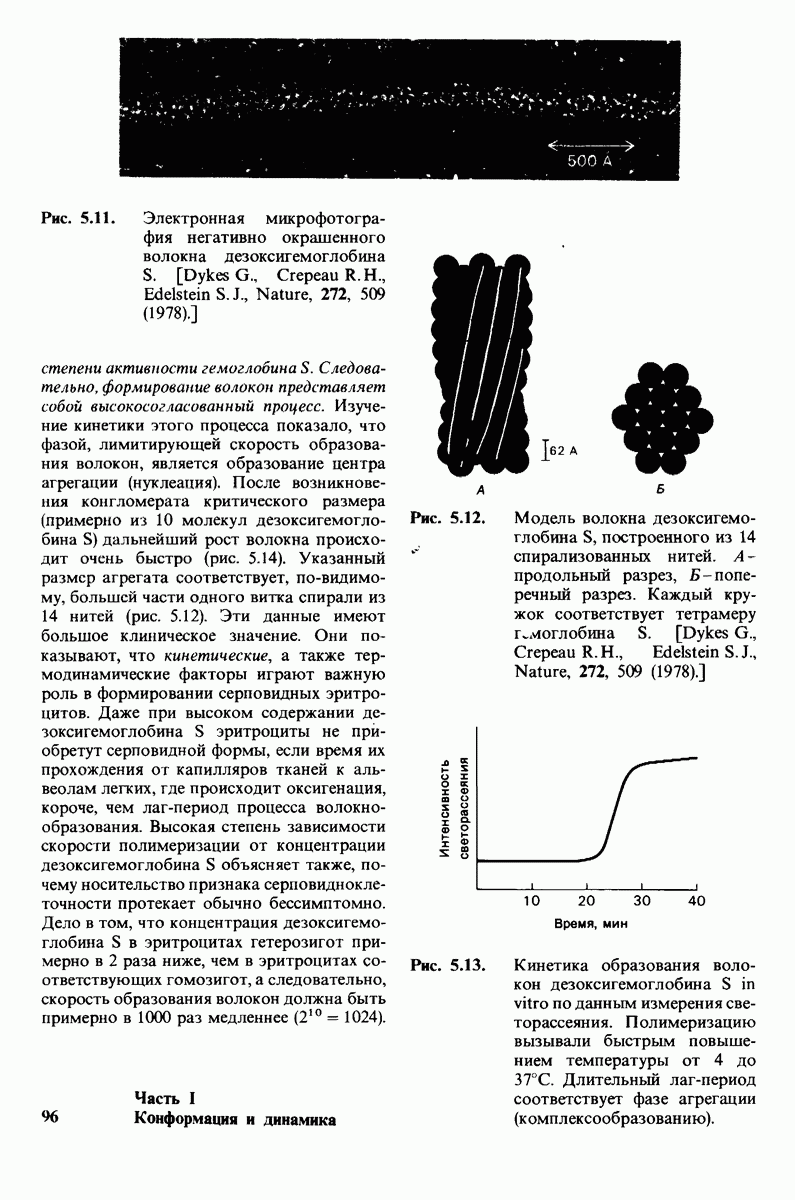
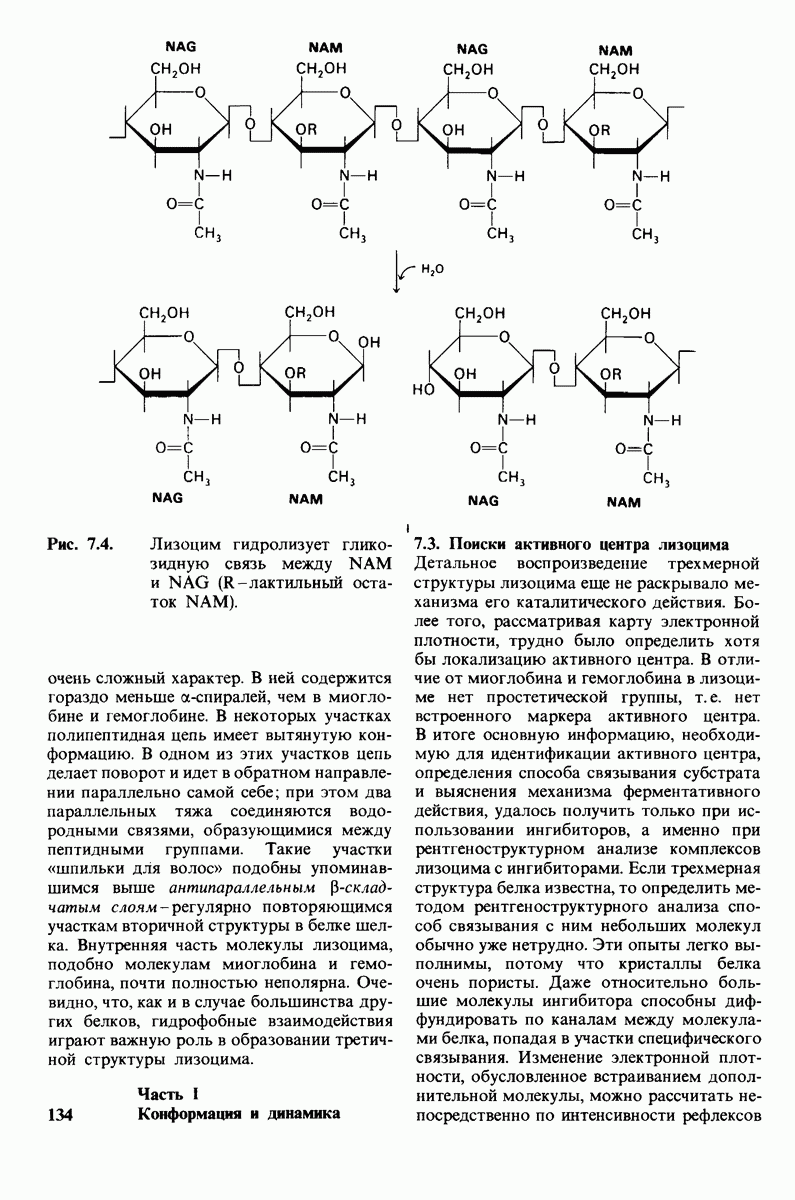
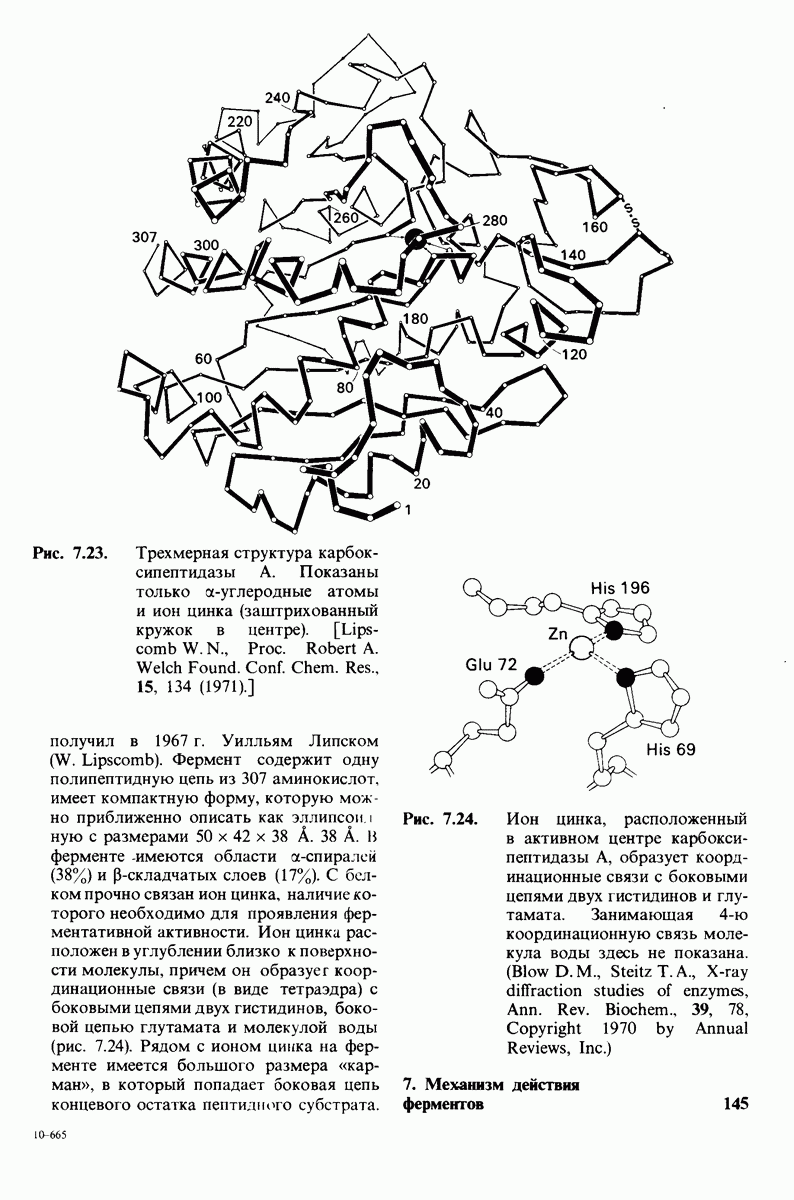
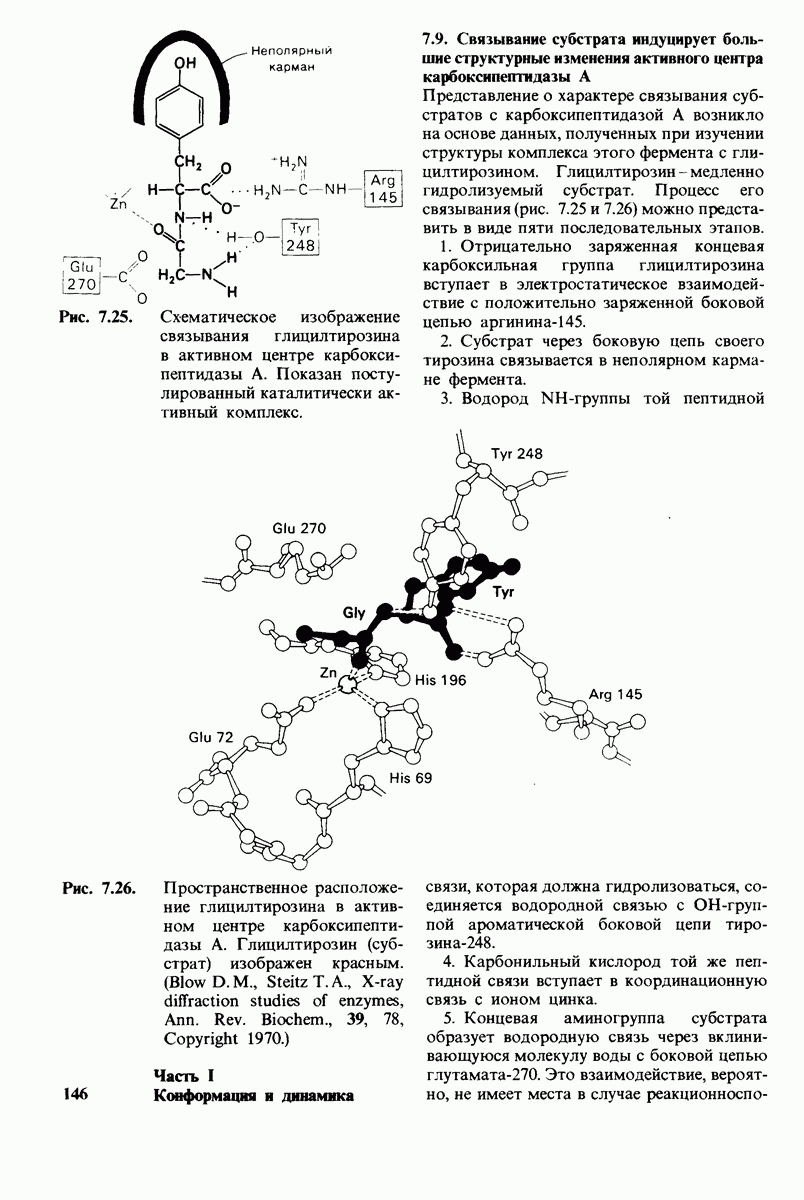
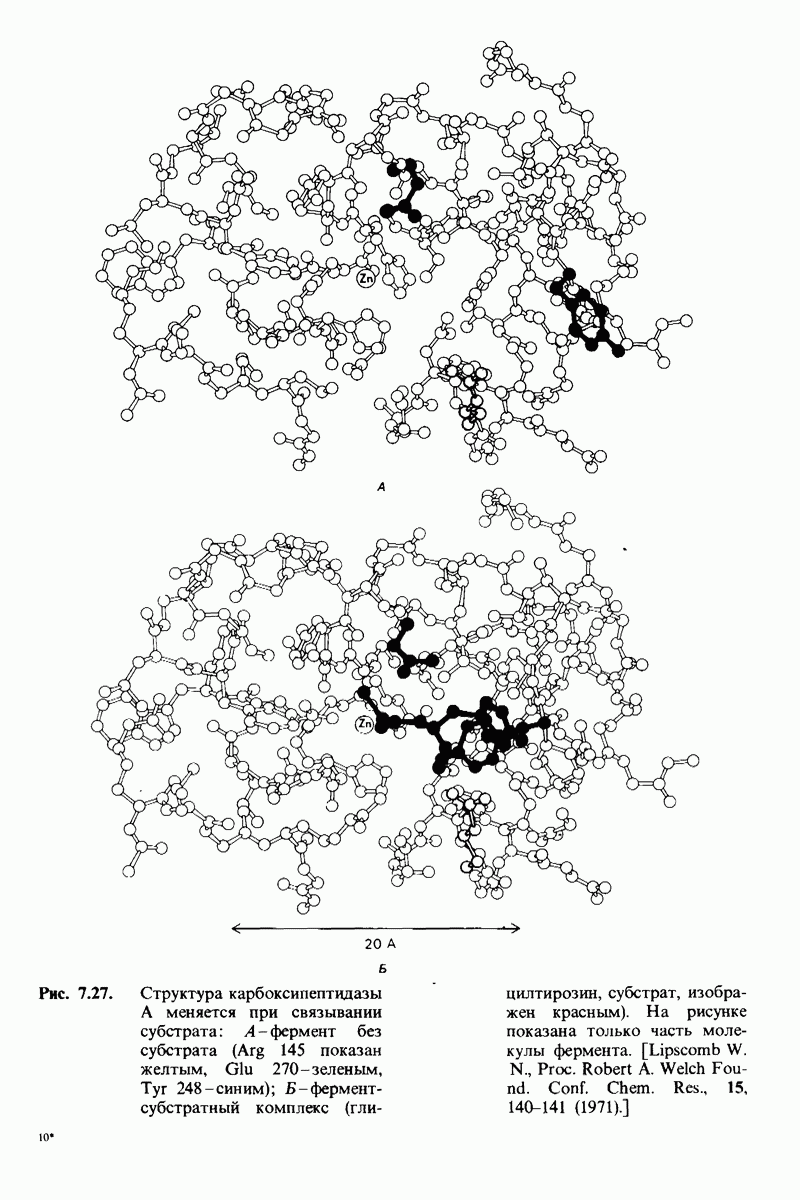
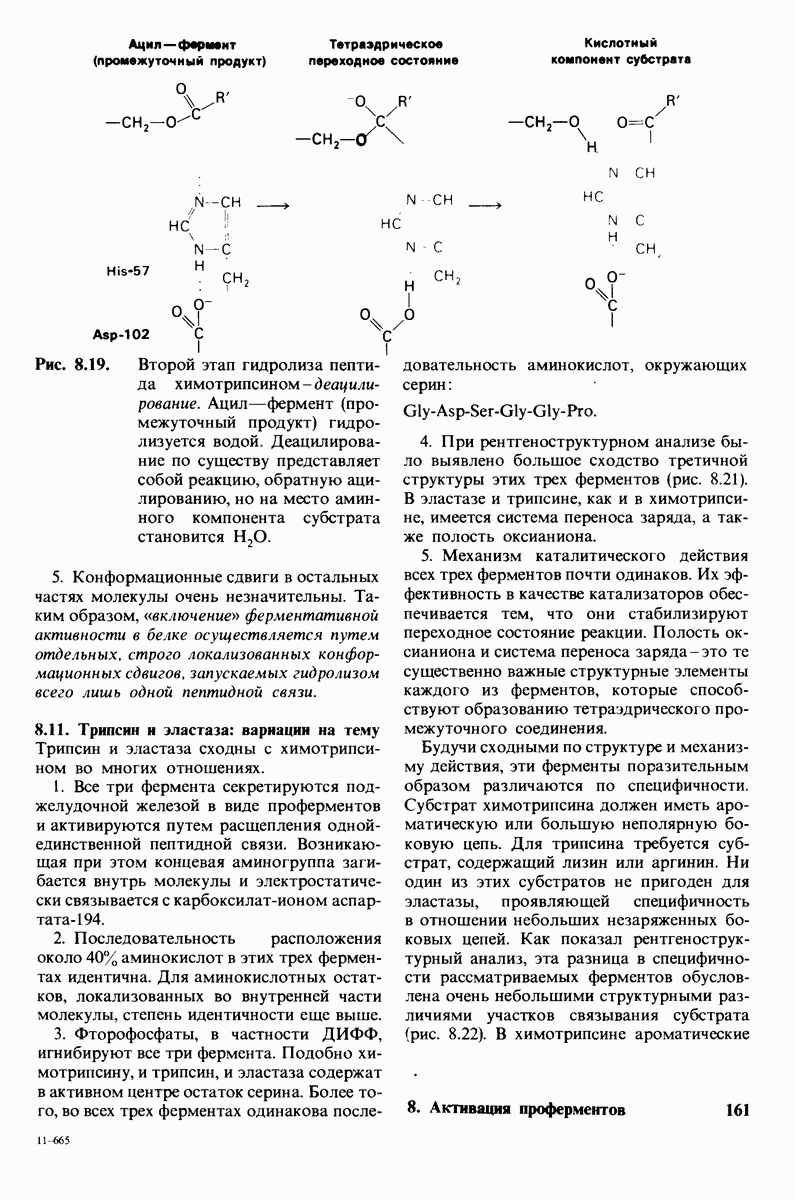
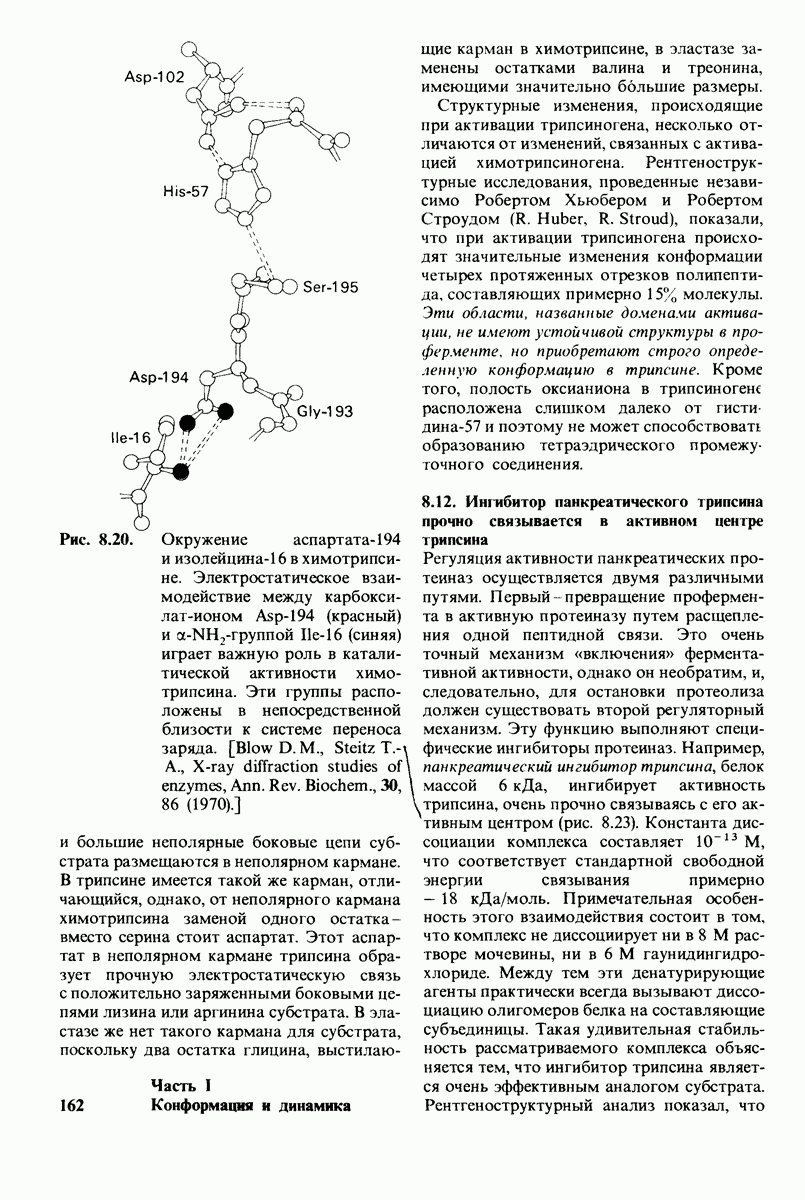
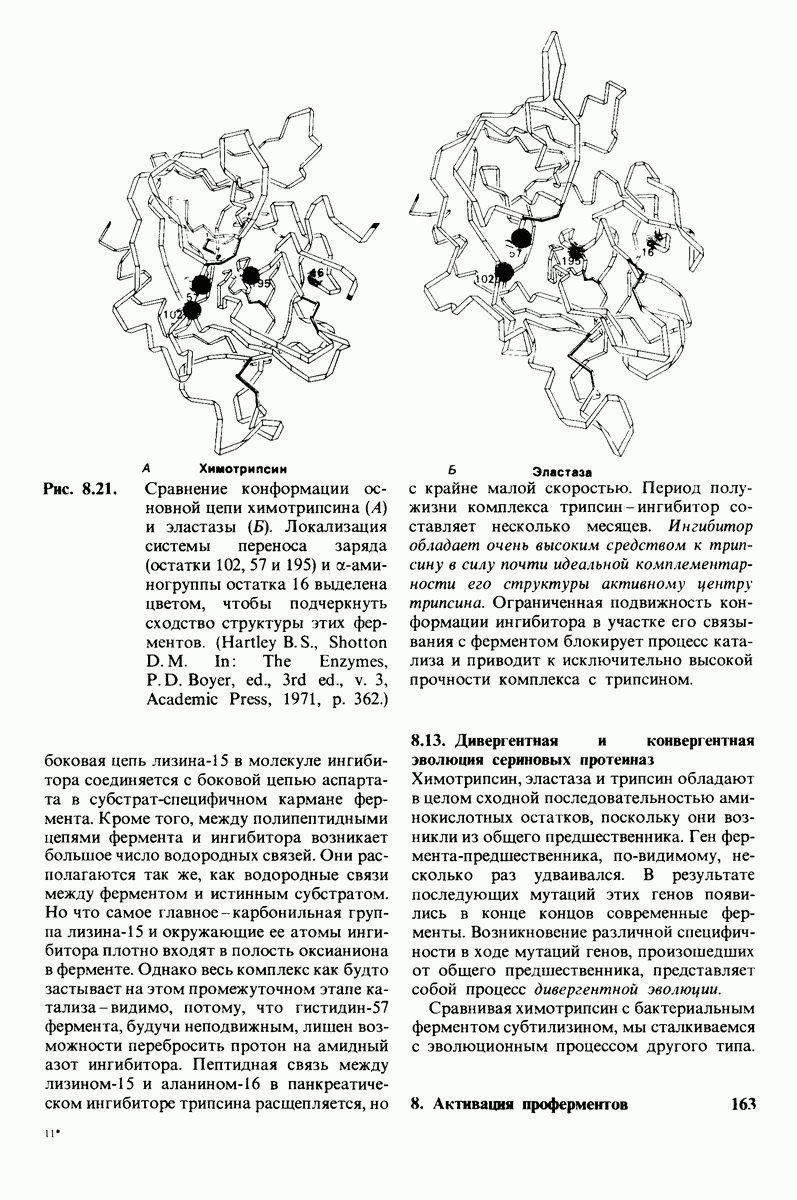
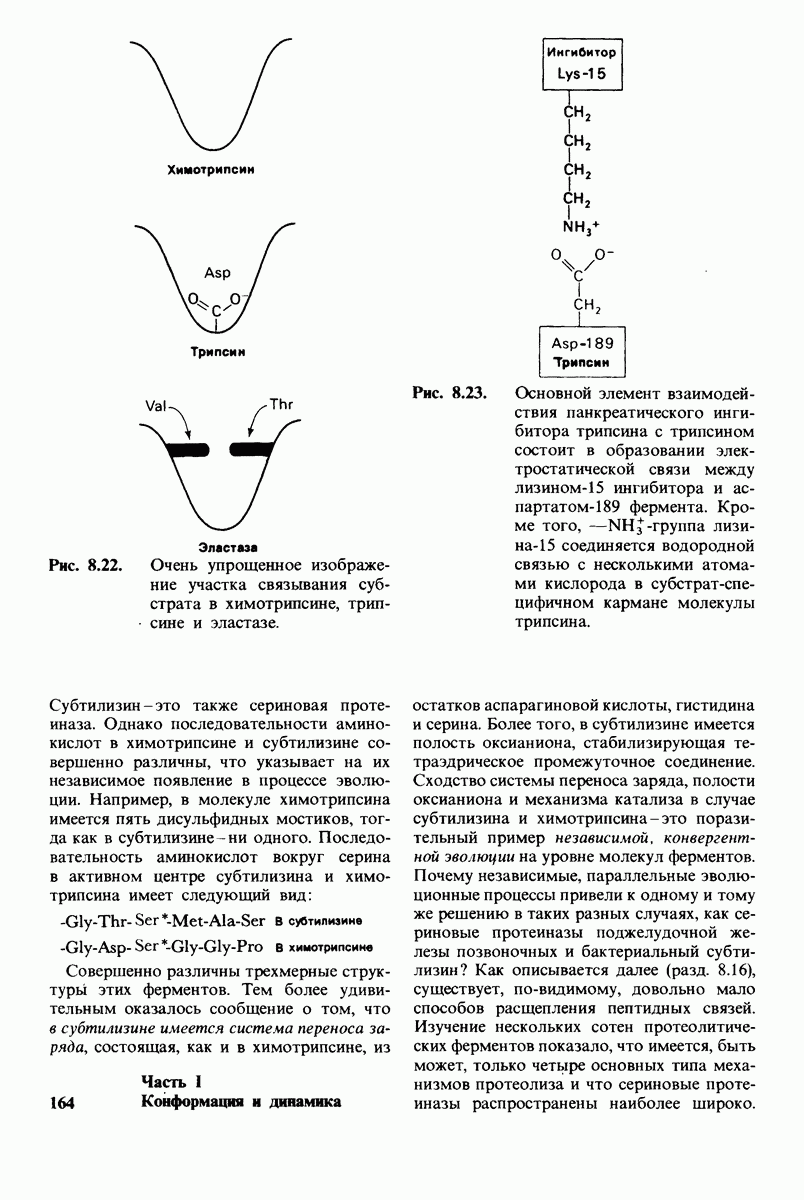
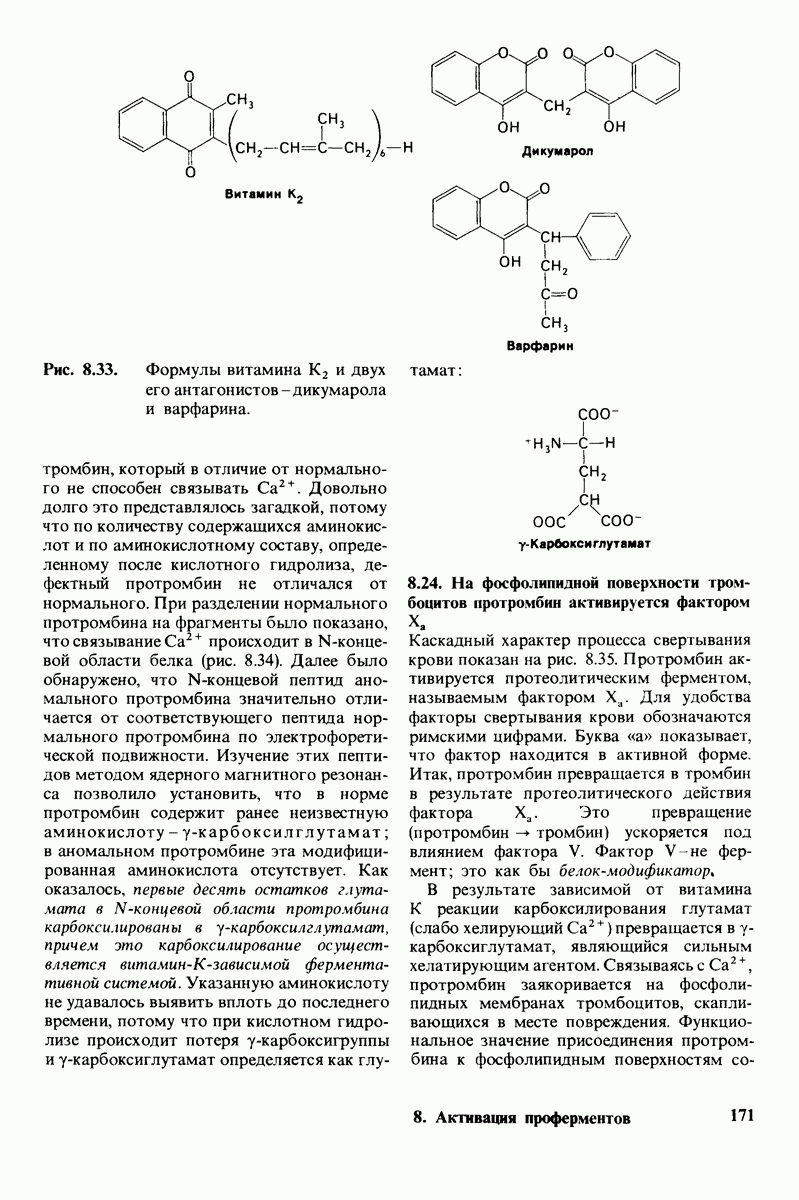
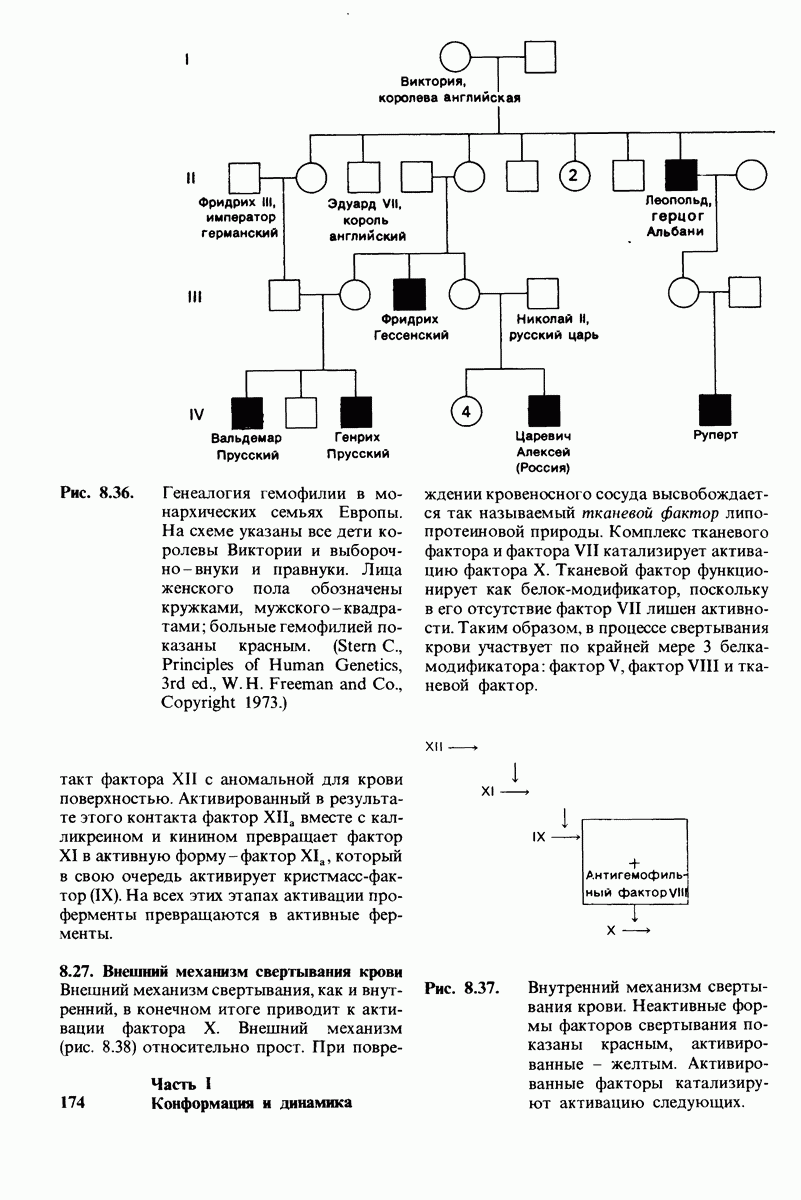
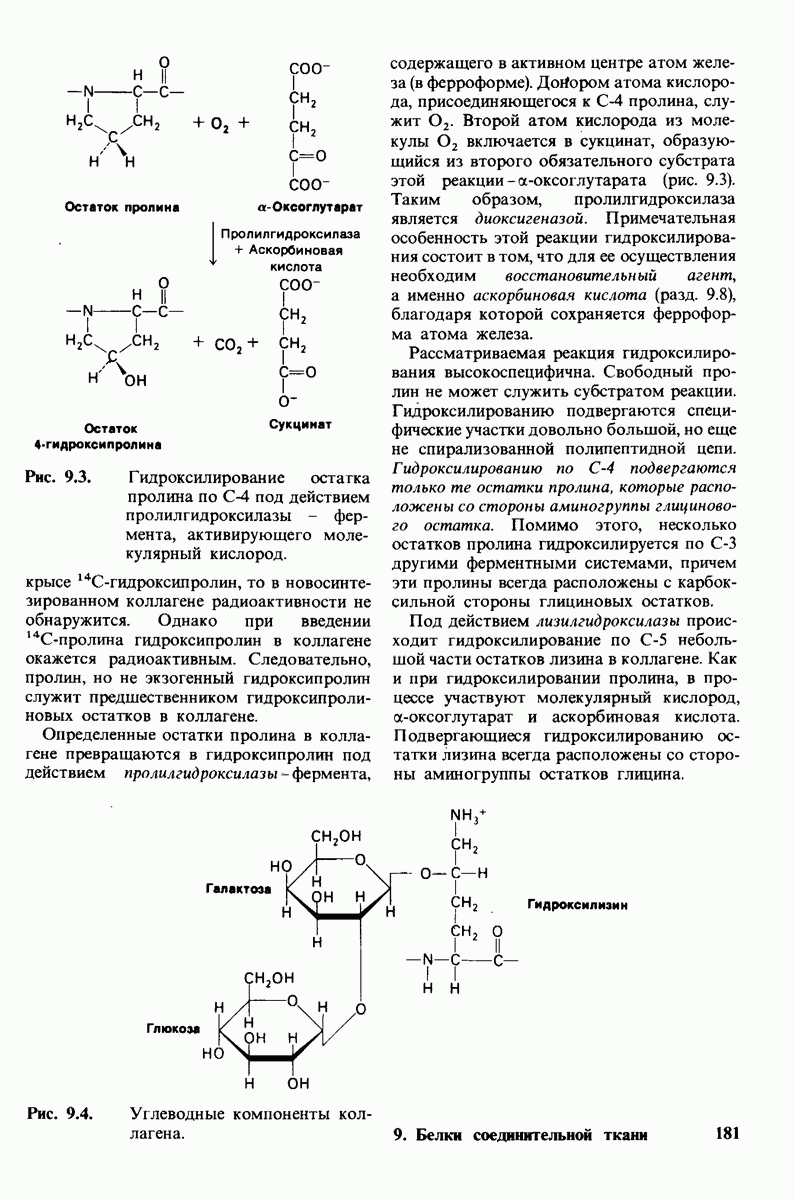
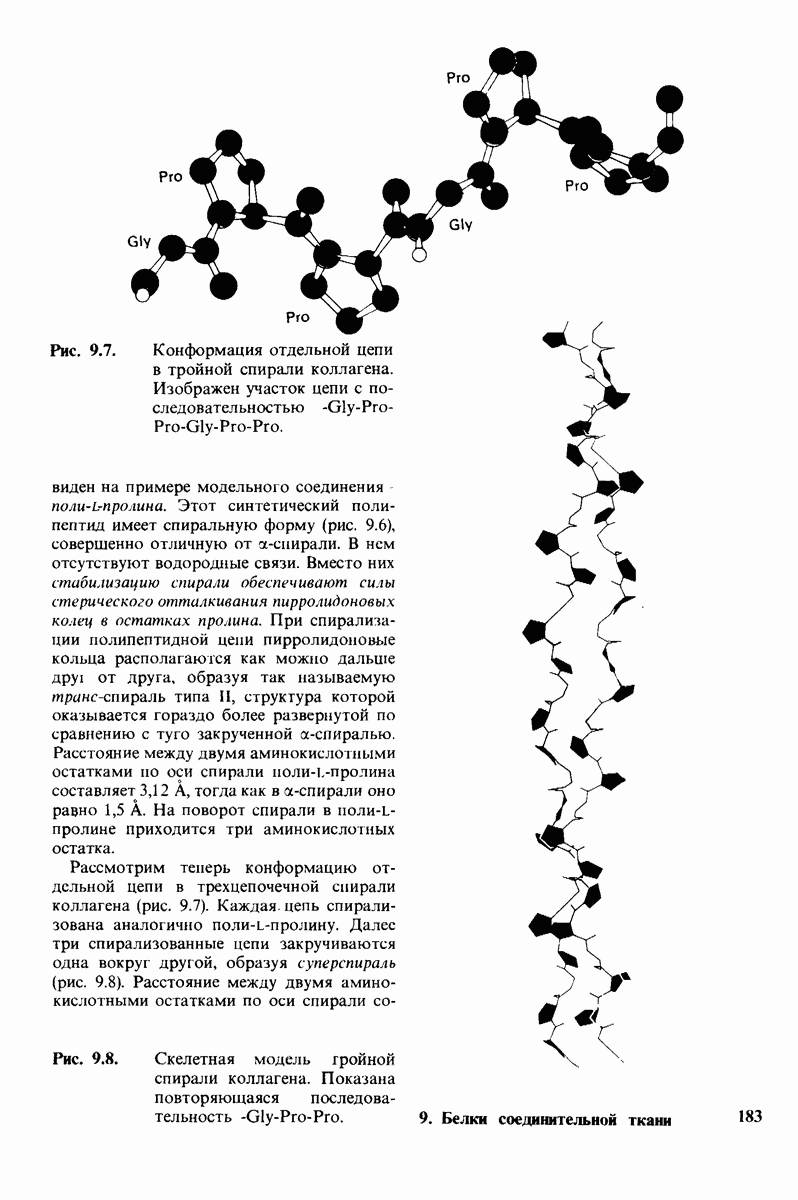
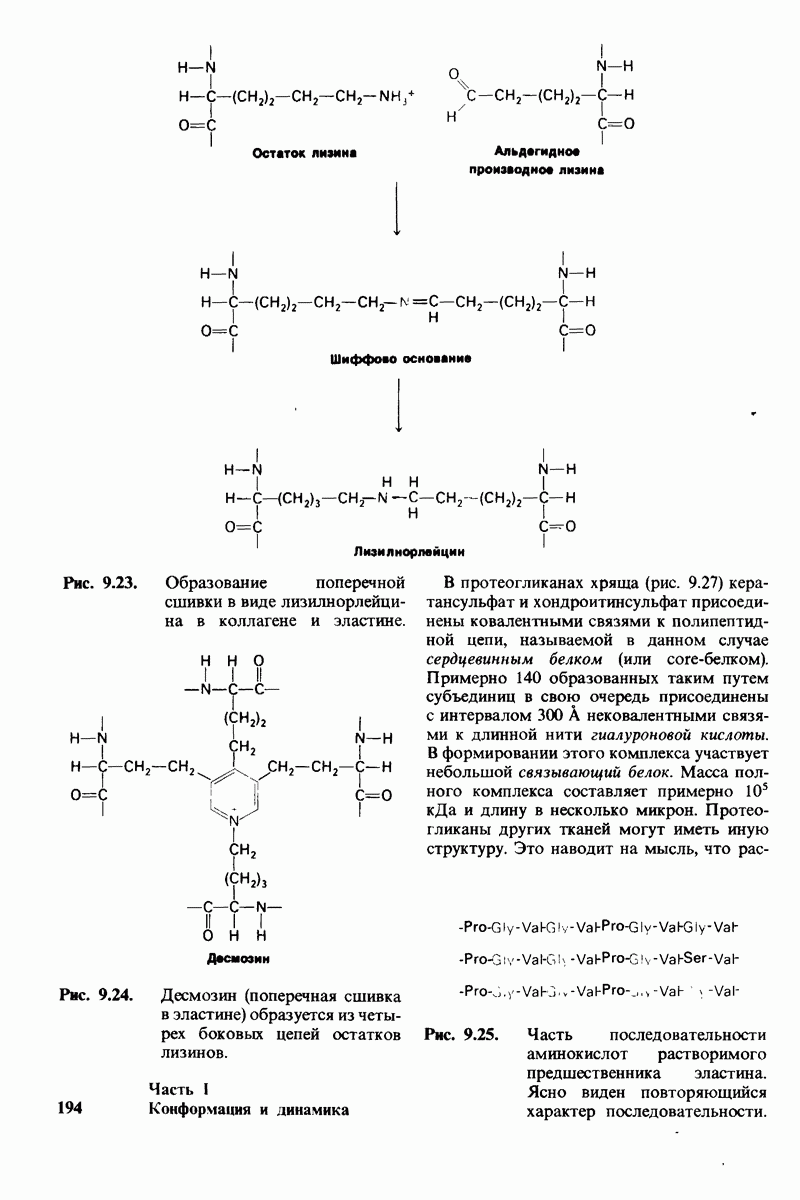
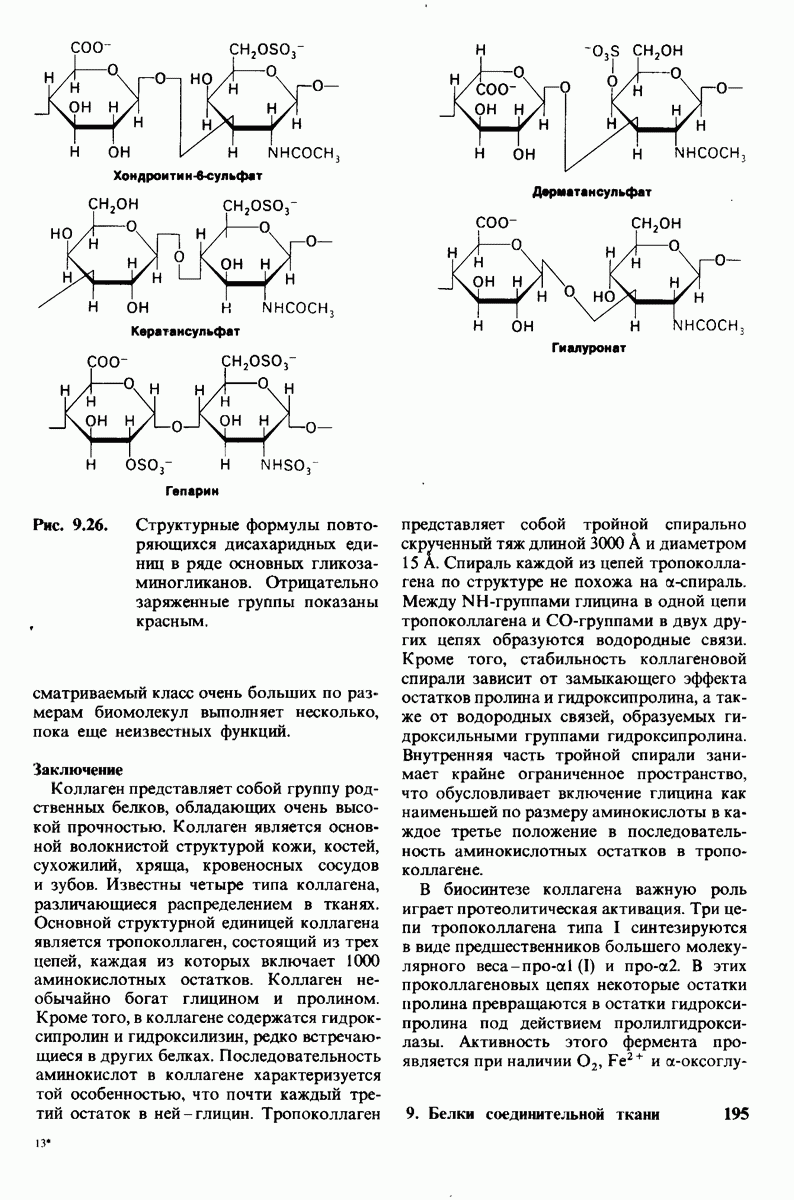
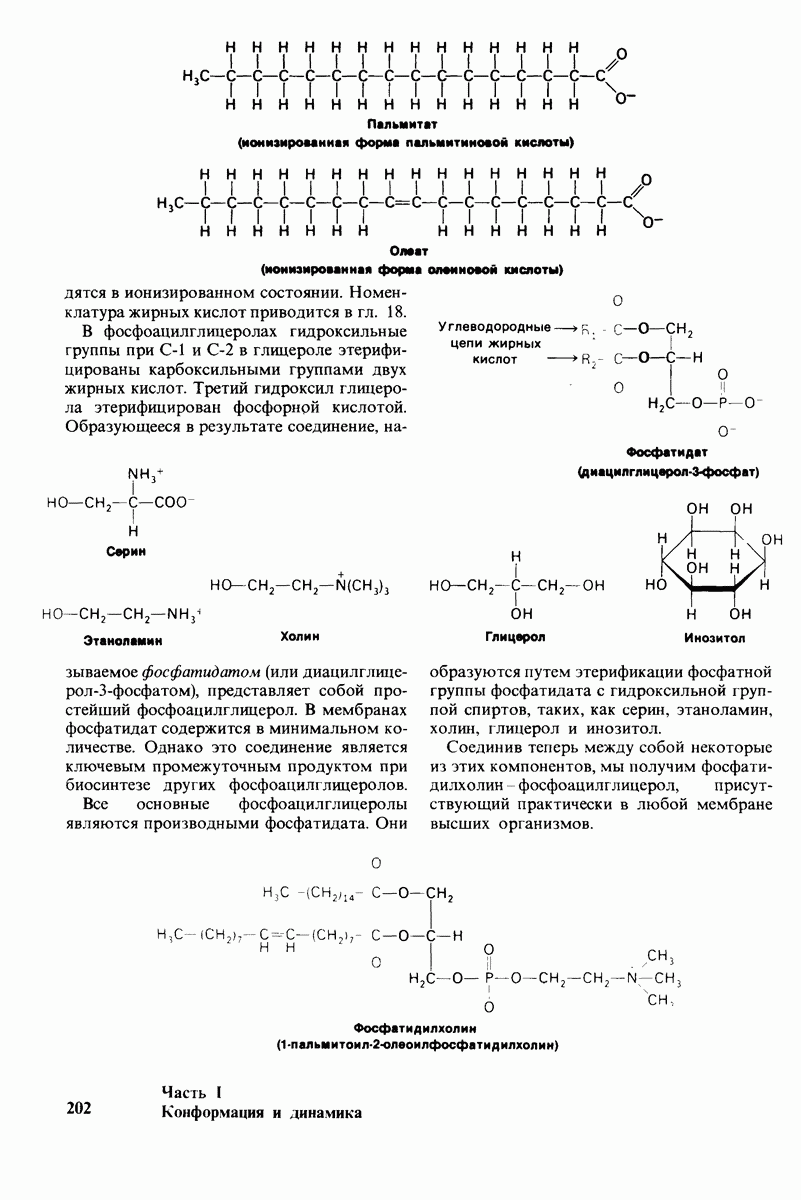
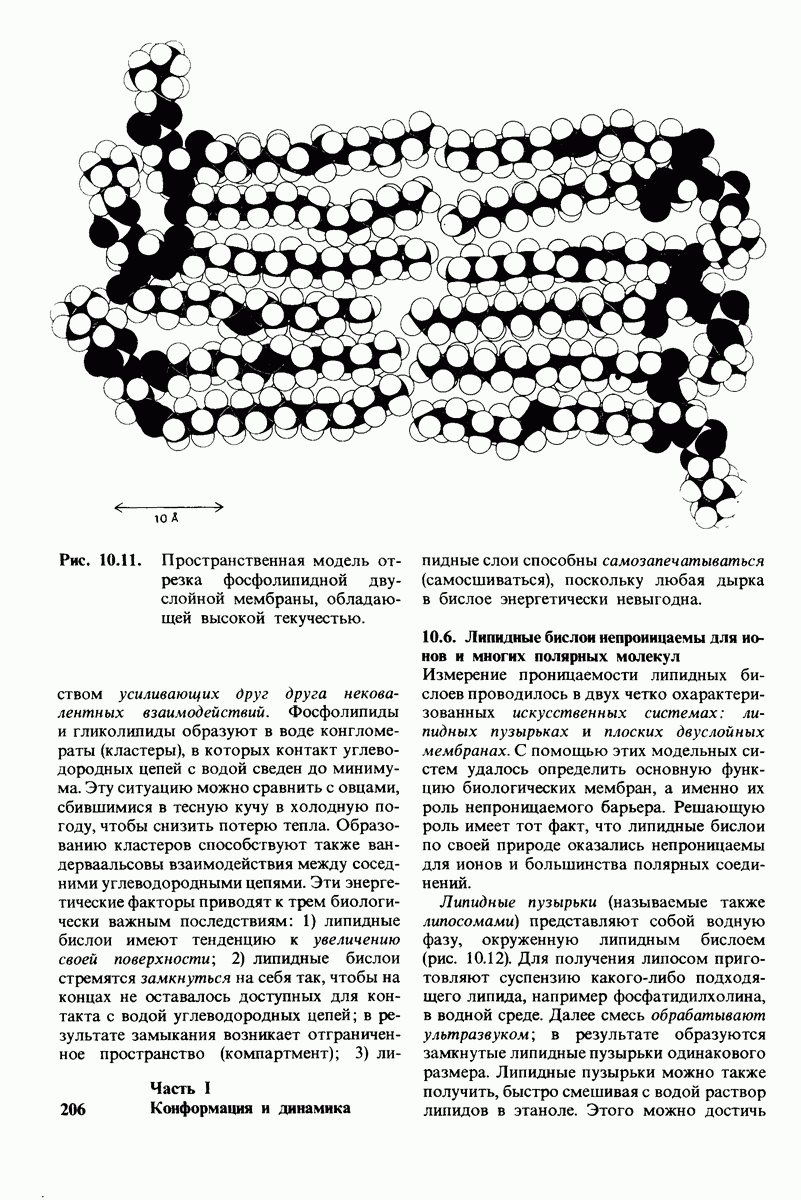
7.7. Экспериментальное доказательство предложенного механизма ферментативного катализа
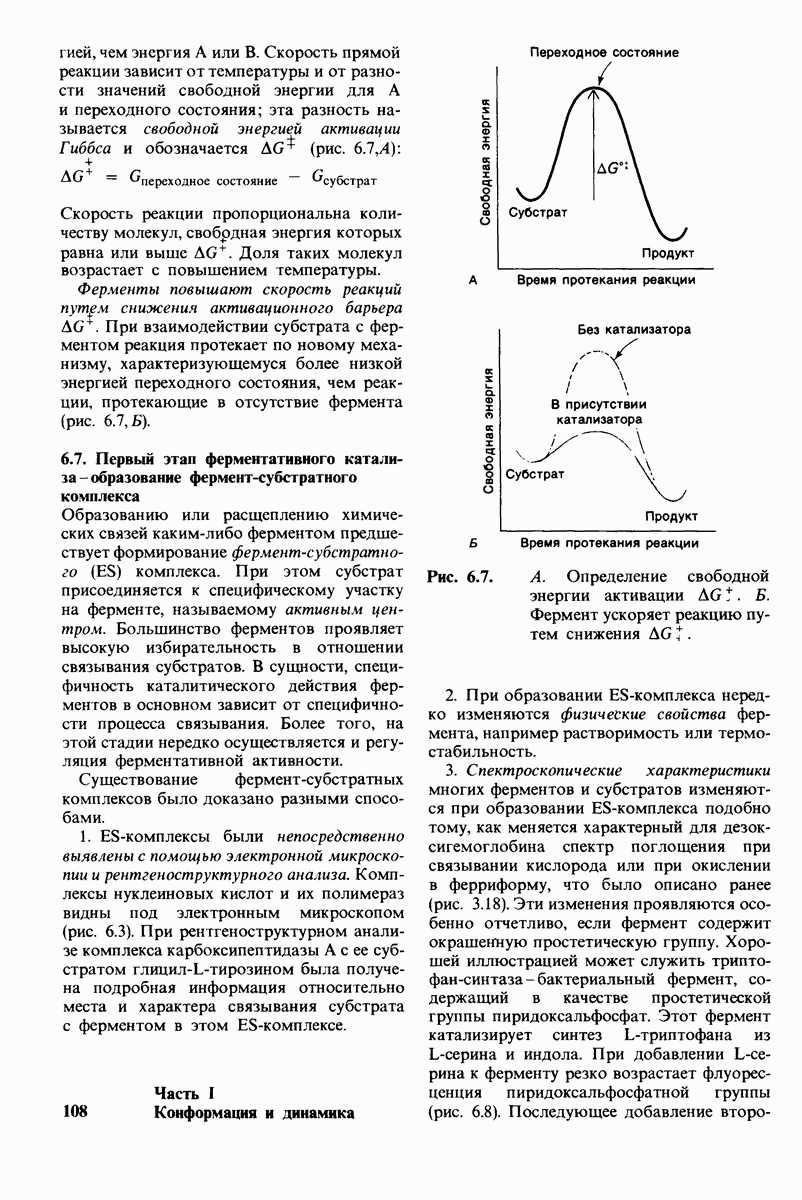
Основанная на рентгеноструктурных данных гипотеза о характере связывания субстрата и механизме катализа была проверена постановкой многочисленных химических экспериментов. Все полученные результаты подтверждают правильность выдвинутой гипотезы. Приведем ряд наиболее существенных экспериментальных доказательств.
1. Способ расщепления субстрата. В соответствии с предположением о том, что расщепление гексамера происходит между  остатками, оказалось, что действительно
остатками, оказалось, что действительно  расщепляется на
расщепляется на

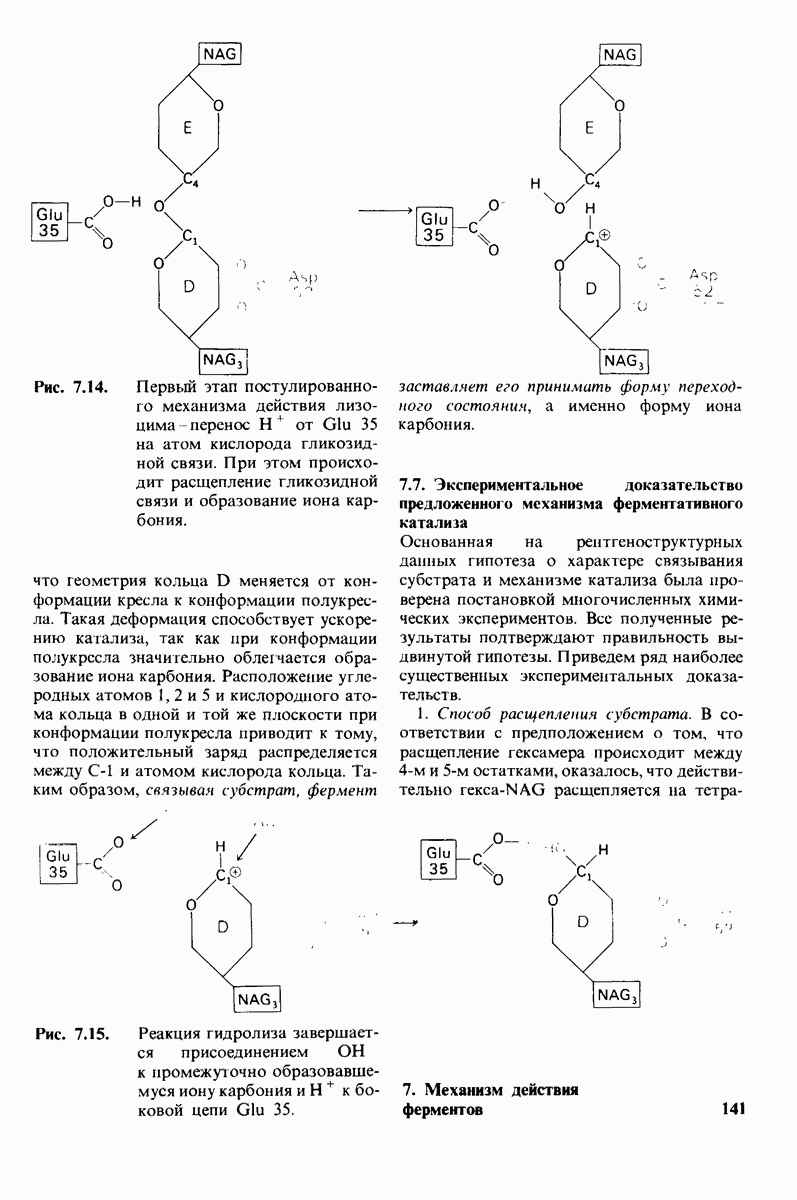
Рис. 7.15. Реакция гидролиза завершается присоединением  к промежуточно образовавшемуся иону карбония и
к промежуточно образовавшемуся иону карбония и  к боковой цепи
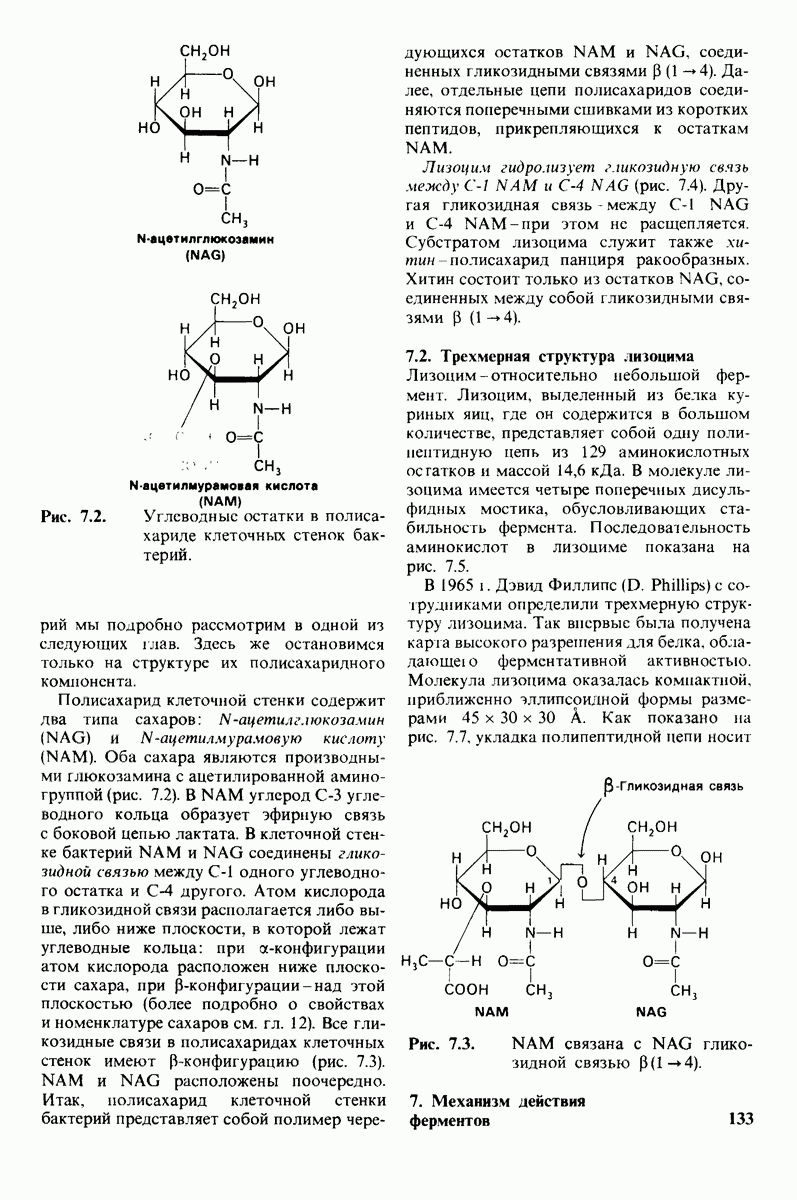
к боковой цепи 

Рис. 7.16. Изменение конформации кольца D субстрата лизоцима в конформацию полукресла. А - углеводный остаток в обычной конформации кресла;  -при связывании с лизоцимом атом кислорода кольца и
-при связывании с лизоцимом атом кислорода кольца и  в углеводном остатке D перемещаются так, что
в углеводном остатке D перемещаются так, что  и О оказываются в одной плоскости, как это показано на рис. В. (Phillips D. С., The three-dimensional structure of an enzyme molecule, Copyright 1966 by Scientific American, Inc.)
и О оказываются в одной плоскости, как это показано на рис. В. (Phillips D. С., The three-dimensional structure of an enzyme molecule, Copyright 1966 by Scientific American, Inc.)
NAG и ди-NAG (рис. 7.17).
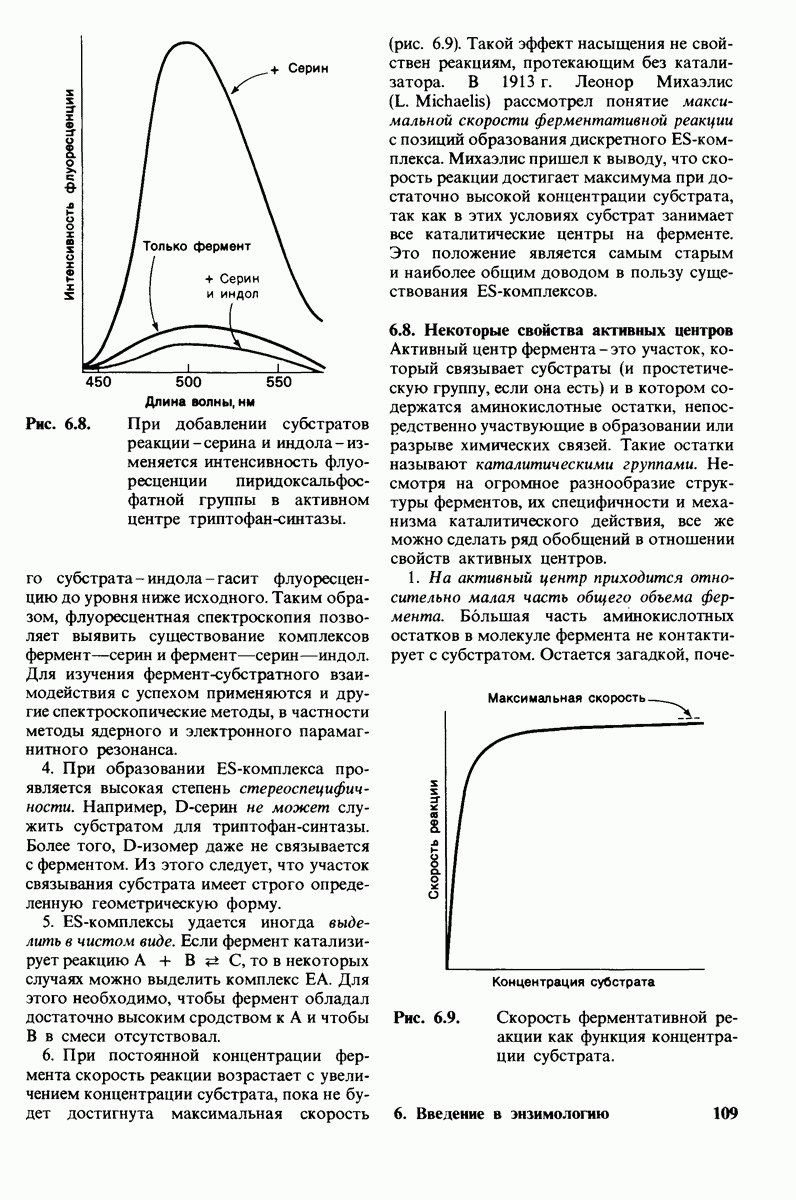
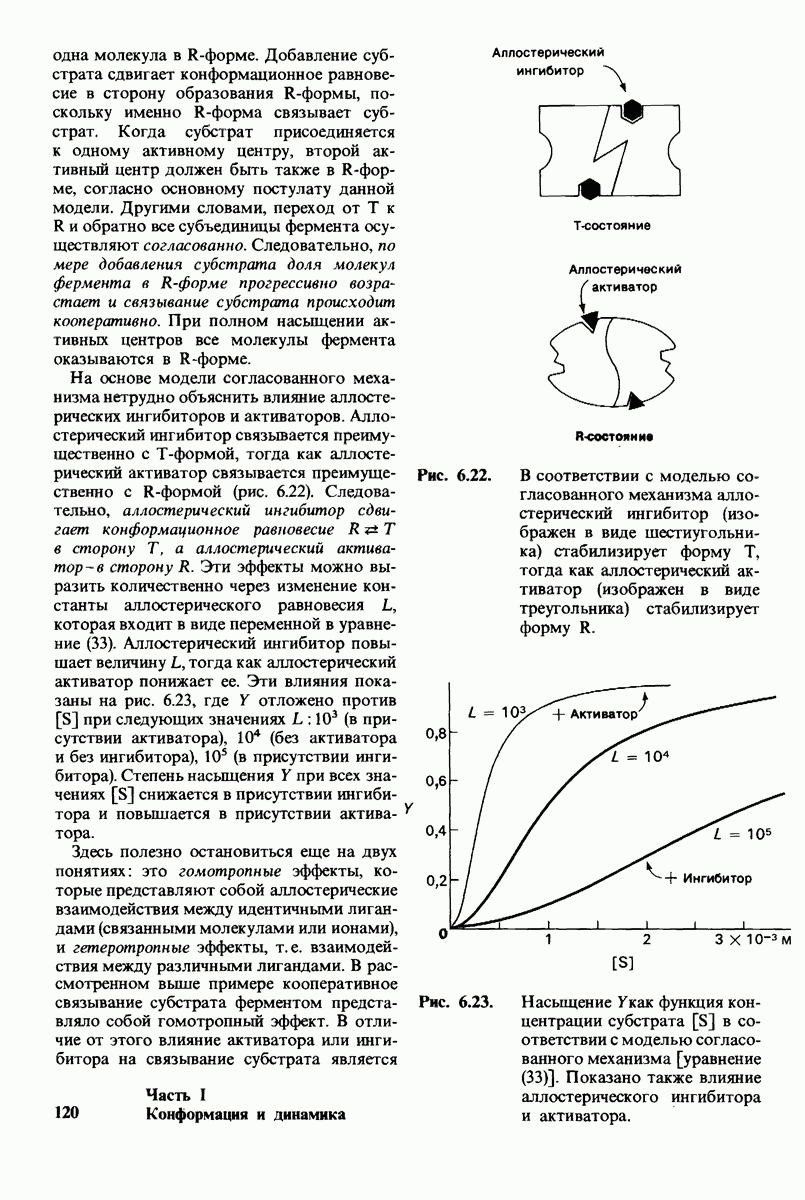
2. Сродство связывания. Путем измерения равновесия при реакции связывания с ферментом каждого из шести Сахаров определяли их вклад в общую величину свободной энергии связей, представленных в гексамере (рис. 7.18). Обнаружилось поразительное явление: вклад углеводного остатка D был отрицательным. На связывание остатка D расходовалось около 4 ккал/моль. Этот результат подтверждает гипотезу о деформации остатка D при связывании с ферментом: изменение от конформации кресла к конформации полукресла требует энергии. Любопытно также, что остаток С вносит наибольший положительный вклад в сродство связывания. Действительно, рентгеноструктурные данные показывают, что остаток С образует большое число водородных связей и вандерваальсовых взаимодействий.
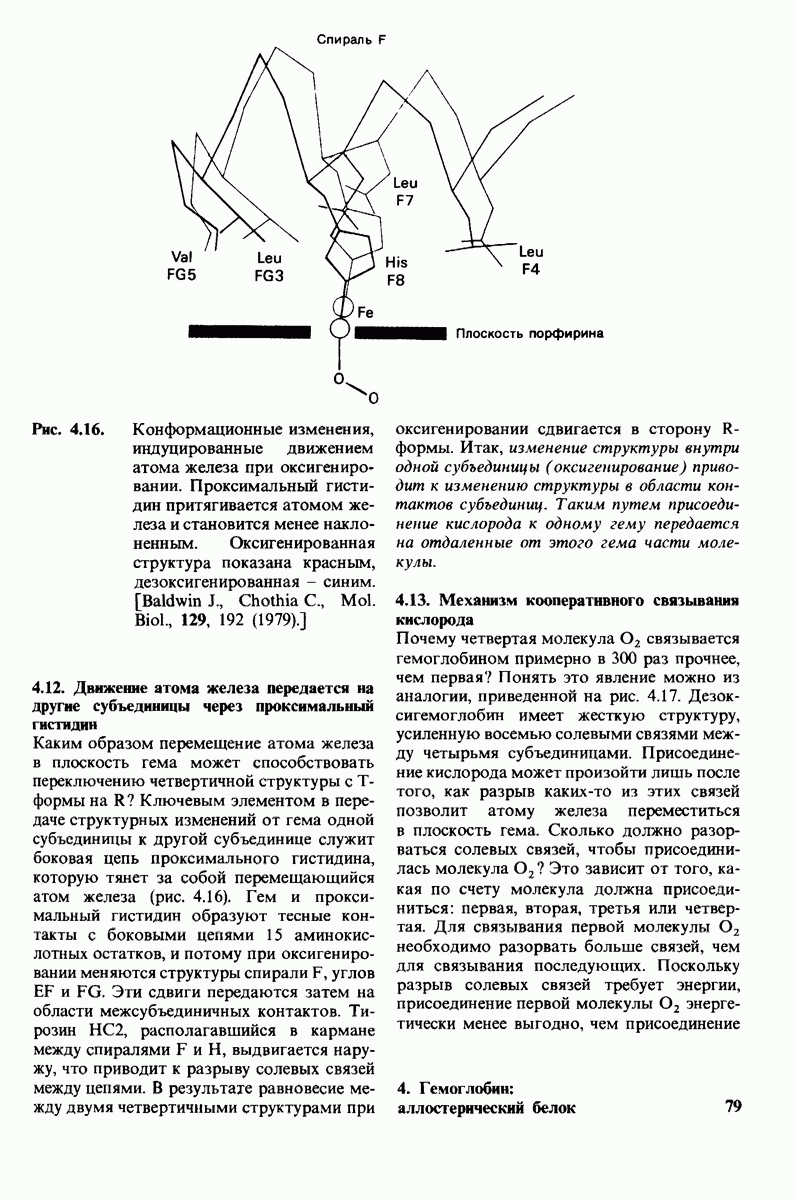
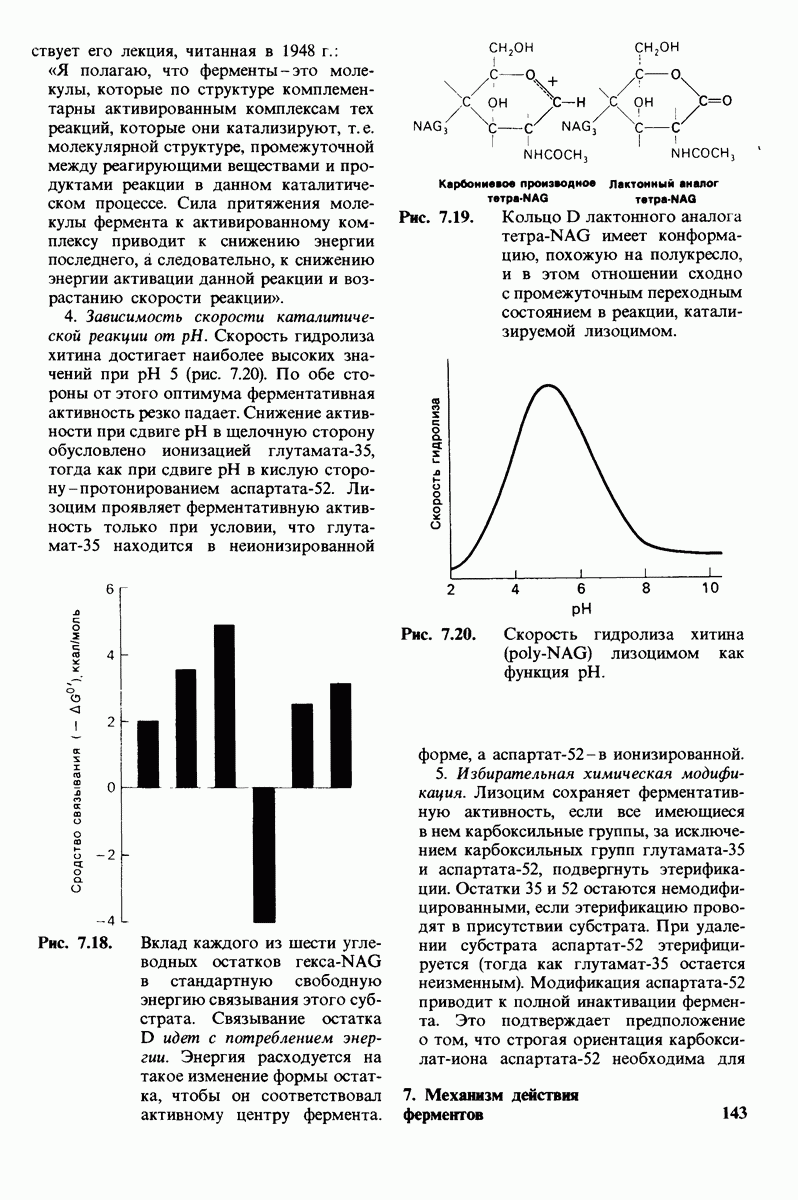
3. Аналоги переходного состояния. Представление о деформации углеводного остатка D в конформацию полукресла - важный аспект постулированного механизма катализа, поскольку конформация полукресла характерна для переходного состояния. Как уже отмечено выше, это представление подтверждается данными о затрате энергии на присоединение остатка D-энергии, расходуемой на деформацию. Другое подтверждение было получено при изучении аналога переходного состояния субстрата, т.е. соединения, имеющего как до, так и после связывания с ферментом такую же геометрию, как субстрат в переходном состоянии. Кольцо D лактонного аналога тетра-NAG (рис. 7.19) в кристалле тетрасахарида имеет конформацию полукресла. При связывании с лизоцимом атомы  и кислородный атом кольца D этого аналога располагаются в одной плоскости. Такая конформация «софы» сходна с постулированной для переходного состояния конформацией полукресла; это означает, что лактонный аналог в отличие от тетра-NAG при связывании с лизоцимом деформируется очень мало. Оказалось, что лактонный аналог связывается с лизоцимом (в участках связывания от А до
и кислородный атом кольца D этого аналога располагаются в одной плоскости. Такая конформация «софы» сходна с постулированной для переходного состояния конформацией полукресла; это означает, что лактонный аналог в отличие от тетра-NAG при связывании с лизоцимом деформируется очень мало. Оказалось, что лактонный аналог связывается с лизоцимом (в участках связывания от А до  в 3600 раз сильнее, чем тетра-NAG. Отсюда вытекает, что при деформации кольца D нормального субстрата скорость расщепления может возрасти примерно в 3600 раз.
в 3600 раз сильнее, чем тетра-NAG. Отсюда вытекает, что при деформации кольца D нормального субстрата скорость расщепления может возрасти примерно в 3600 раз.
Роль этого фактора в катализе ясно предвидел Полинг (Pauling), о чем

Рис.  гидролизуется с образованием
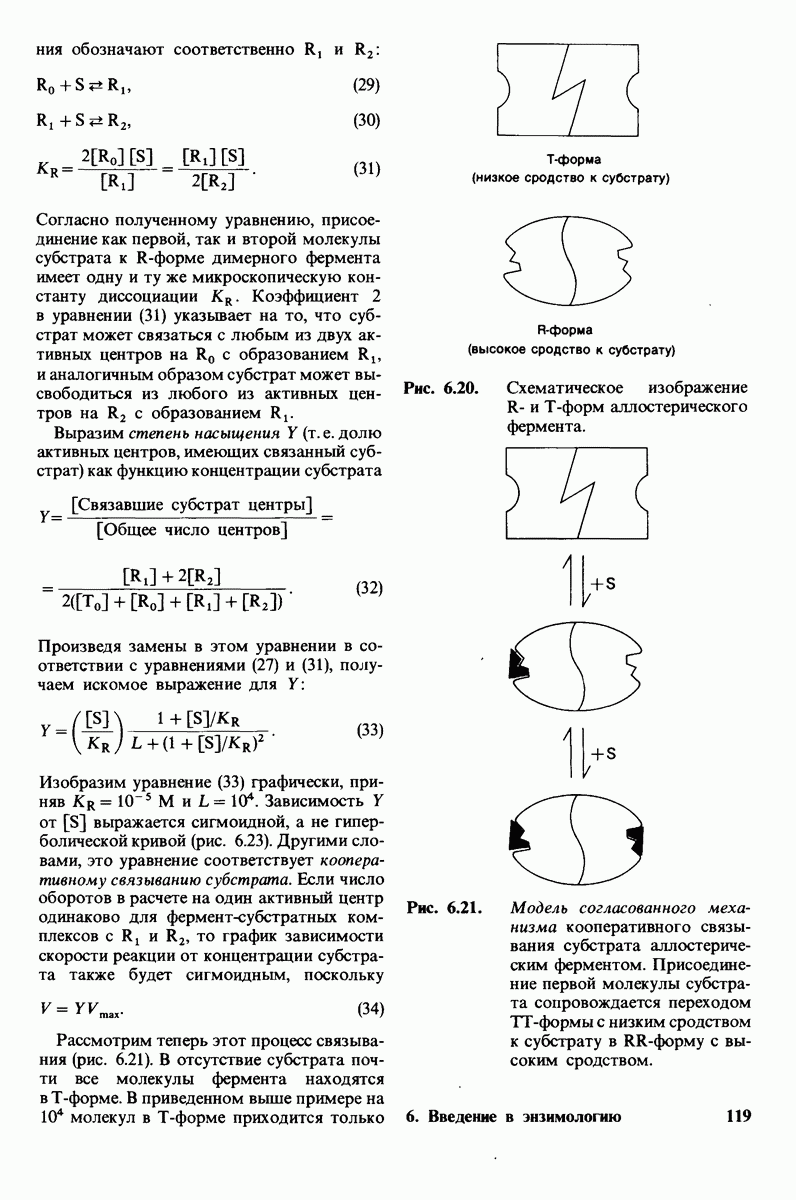
гидролизуется с образованием 
свидетельствует его лекция, читанная в 1948 г.: «Я полагаю, что ферменты - это молекулы, которые по структуре комплементарны активированным комплексам тех реакций, которые они катализируют, т.е. молекулярной структуре, промежуточной между реагирующими веществами и продуктами реакции в данном каталитическом процессе. Сила притяжения молекулы фермента к активированному комплексу приводит к снижению энергии последнего, а следовательно, к снижению энергии активации данной реакции и возрастанию скорости реакции».
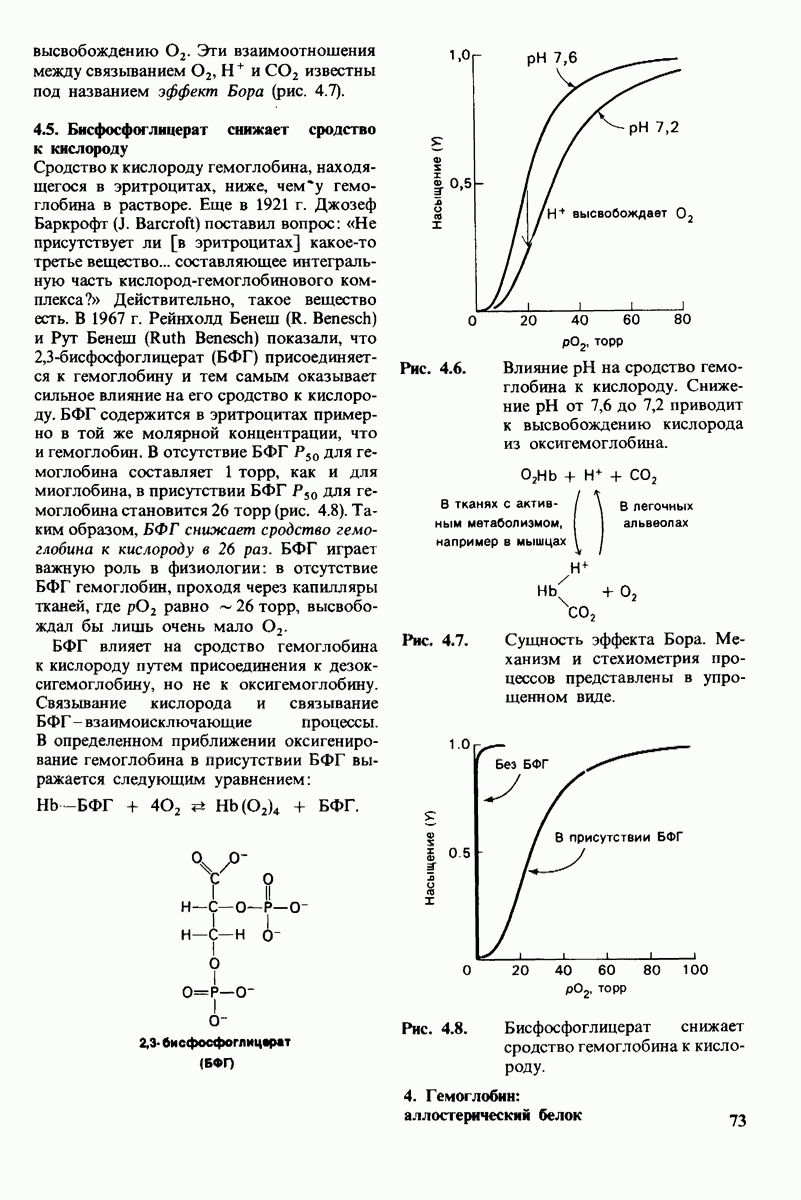
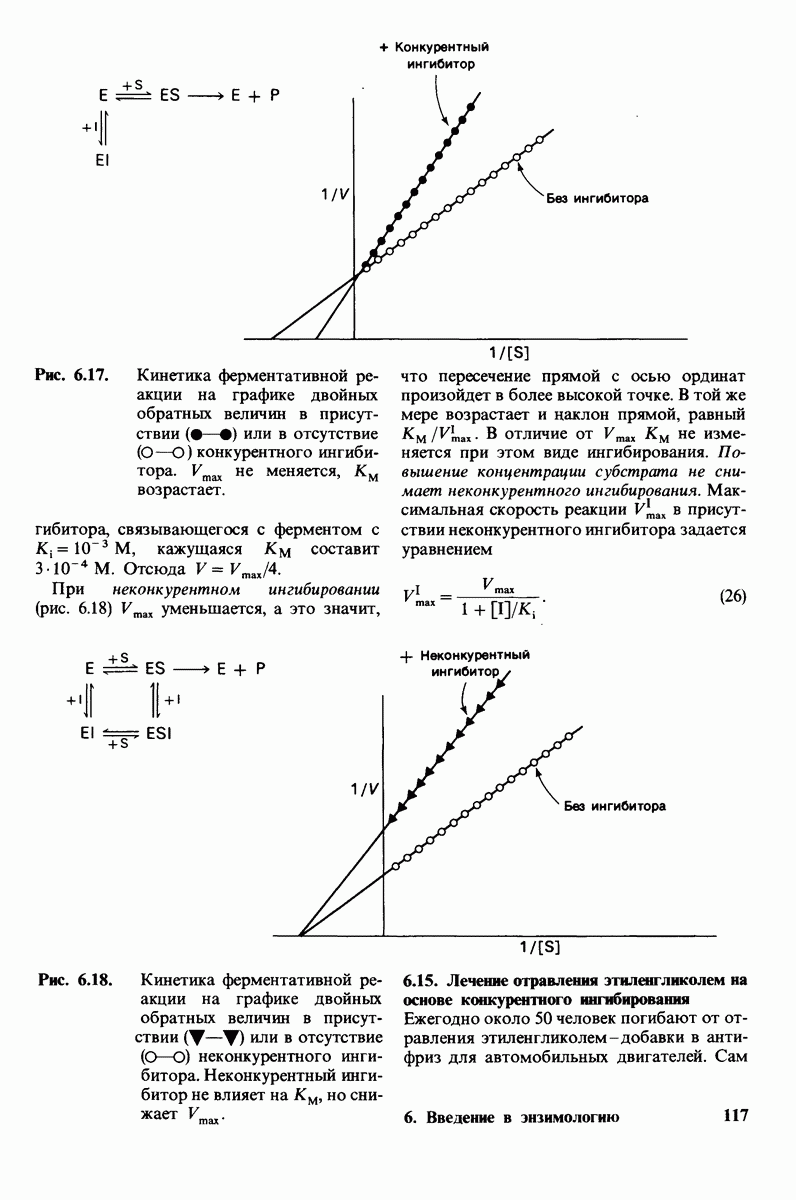
4. Зависимость скорости каталитической реакции от рН. Скорость гидролиза хитина достигает наиболее высоких значений при  (рис. 7.20). По обе стороны от этого оптимума ферментативная активность резко падает. Снижение активности при сдвиге рН в щелочную сторону обусловлено ионизацией глутамата-35, тогда как при сдвиге рН в кислую сторону - протонированием аспартата-52. Лизоцим проявляет ферментативную активность только при условии, что глутамат-35 находится в неионизированной форме, а аспартат-52 - в ионизированной.
(рис. 7.20). По обе стороны от этого оптимума ферментативная активность резко падает. Снижение активности при сдвиге рН в щелочную сторону обусловлено ионизацией глутамата-35, тогда как при сдвиге рН в кислую сторону - протонированием аспартата-52. Лизоцим проявляет ферментативную активность только при условии, что глутамат-35 находится в неионизированной форме, а аспартат-52 - в ионизированной.

Рис. 7.18.
Вклад каждого из шести углеводных остатков  в стандартную свободную энергию связывания этого субстрата. Связывание остатка D идет с потреблением энергии. Энергия расходуется на такое изменение формы остатка, чтобы он соответствовал активному центру фермента.
в стандартную свободную энергию связывания этого субстрата. Связывание остатка D идет с потреблением энергии. Энергия расходуется на такое изменение формы остатка, чтобы он соответствовал активному центру фермента.

Рис. 7.19. Кольцо D лактонного аналога тетра-NAG имеет конформацию, похожую на полукресло, и в этом отношении сходно с промежуточным переходным состоянием в реакции, катализируемой лизоцимом.

Рис. 7.20. Скорость гидролиза хитина (poly-NAG) лизоцимом как функция рН.
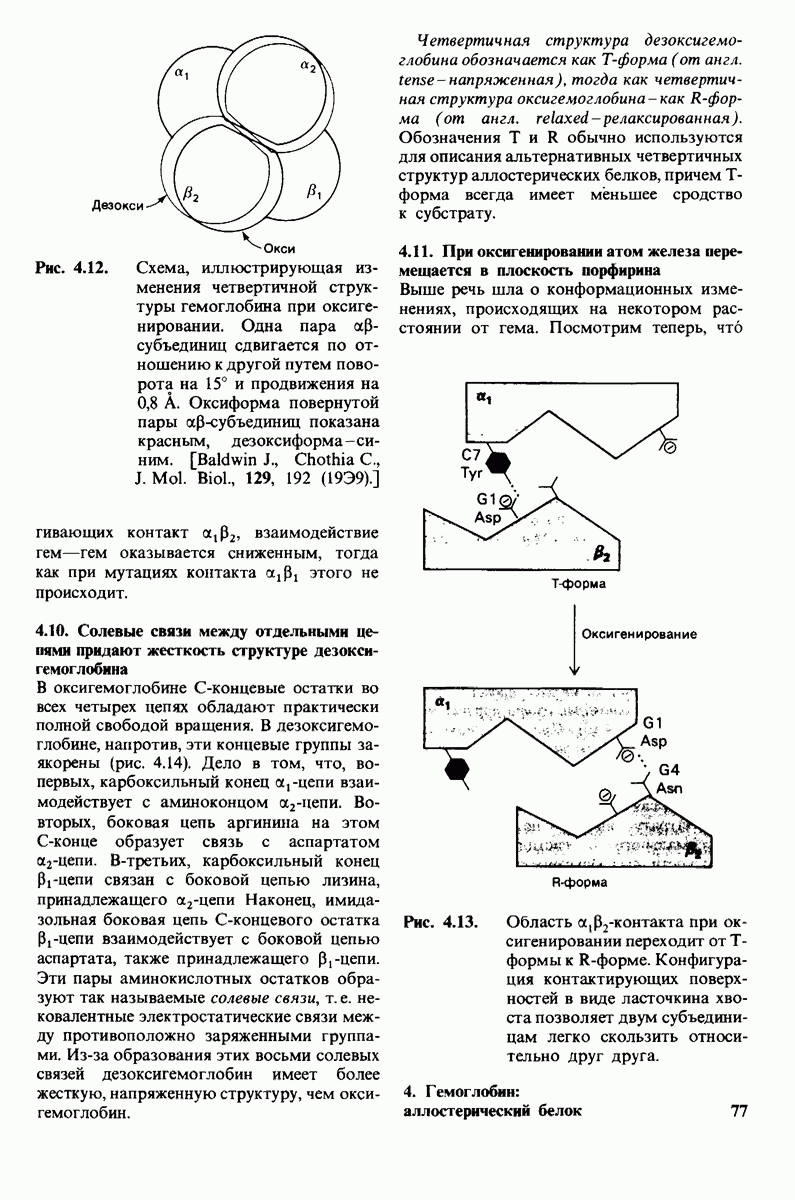
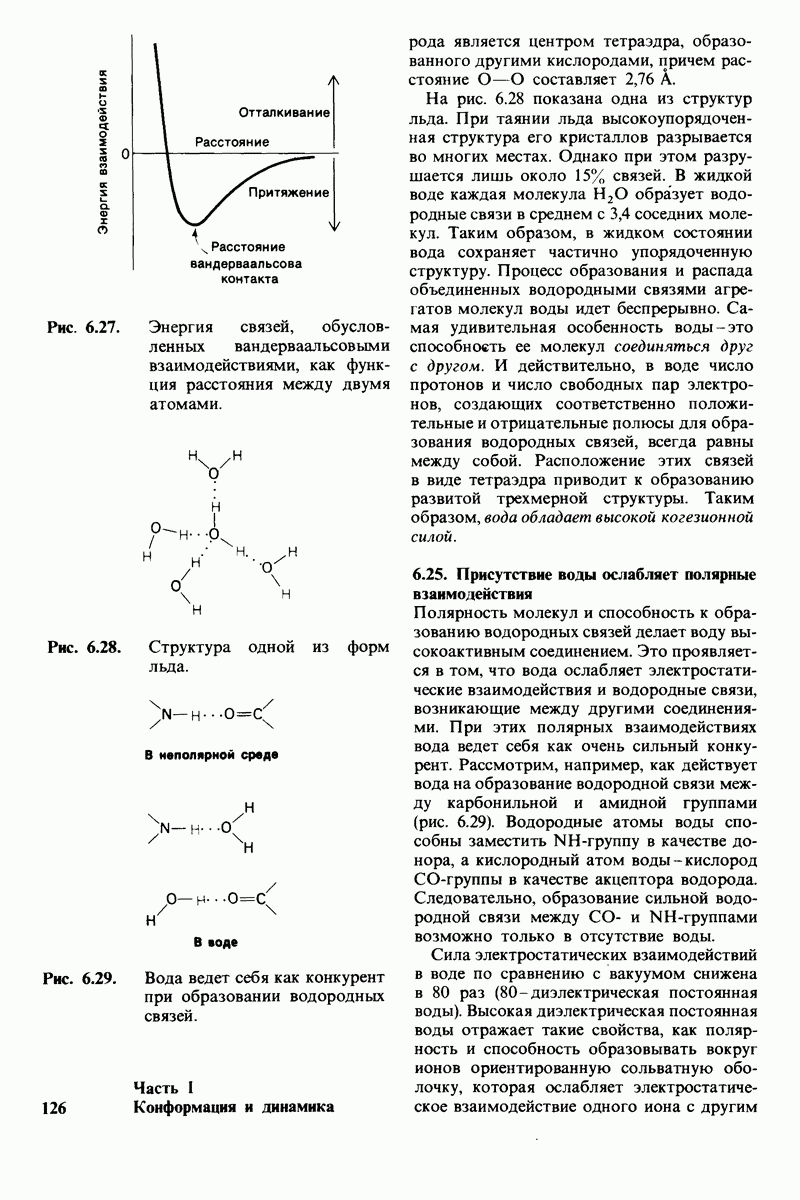
5. Избирательная химическая модификация. Лизоцим сохраняет ферментативную активность, если все имеющиеся в нем карбоксильные группы, за исключением карбоксильных групп глутамата-35 и аспартата-52, подвергнуть этерифика-ции. Остатки 35 и 52 остаются модифицированными, если этерификацию проводят в присутствии субстрата. При удалении субстрата аспартат-52 этерифици-руется (тогда как глутамат-35 остается неизменным). Модификация аспартата-52 приводит к полной инактивации фермента. Это подтверждает предположение о том, что строгая ориентация карбокси-лат-иона аспартата-52 необходима для

Рис. 7.21, Существование промежуточного продукта гликозил - фермент подтверждается способностью лизоцима катализировать, хотя и медленно, реакцию транс! ликозилирования.  (показан красным) соединяется с промежуточным продуктом гликозил фермент (показан синим на рис.
(показан красным) соединяется с промежуточным продуктом гликозил фермент (показан синим на рис.  и в результате образуется
и в результате образуется 
стабилизации образующегося в качестве промежуточного продукта иона карбония.
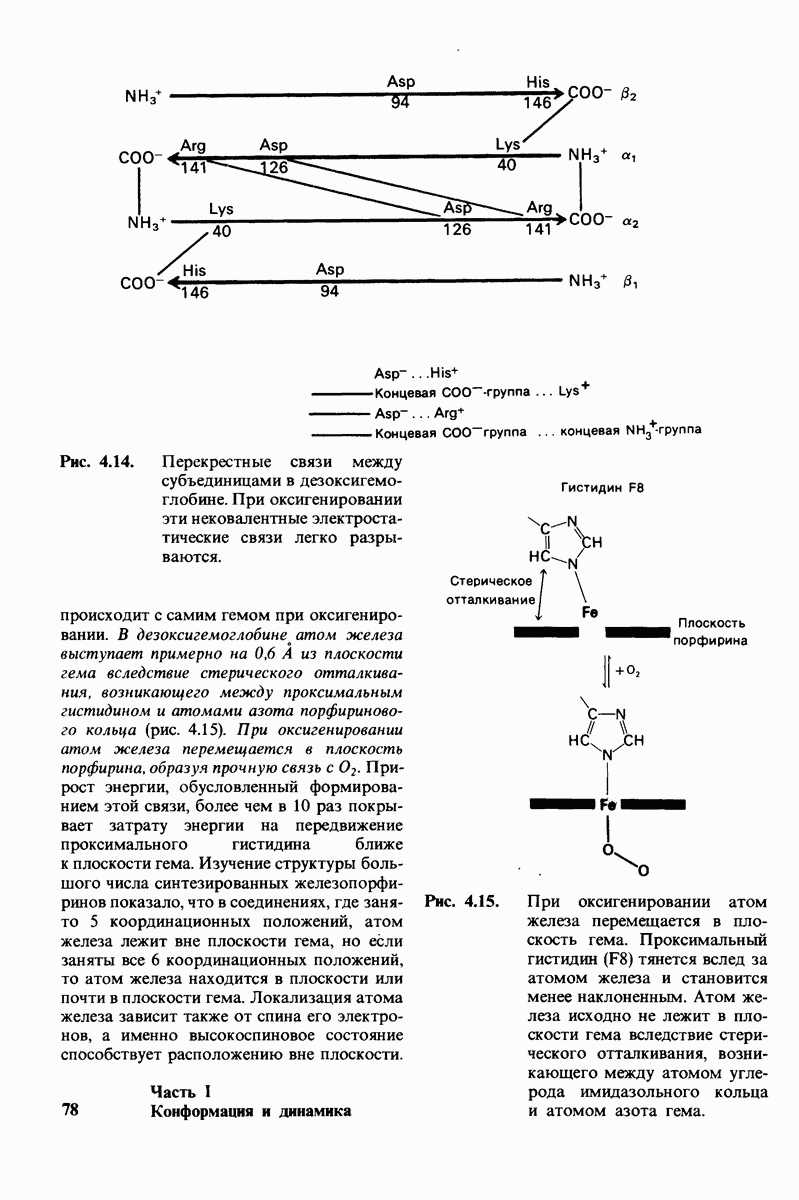
6. Трансгликозилирование. При добавлении к лизоциму тетра-NAG происходит медленное образование гекса-NAG и ди-NAG (рис. 7.21). Существование этой реакции, называемой трансгликозилированием, подтверждает наличие одного из важных элементов предложенного механизма ферментативной реакции, а именно образования в качестве промежуточного продукта комплекса гликозил — фермент.

Рис. 7.22. Реакция, катализируемая карбоксипептидазой А.
При обычных гидролитических реакциях этот промежуточный продукт реагирует с  При трансгликозилировании в реакции используется вторая молекула углевода:
При трансгликозилировании в реакции используется вторая молекула углевода:  Реакция трансгликозилирования специфична, так как акцептор связывается в участках
Реакция трансгликозилирования специфична, так как акцептор связывается в участках  в щели активного центра. Более того, образующаяся гликозидная связь имеет
в щели активного центра. Более того, образующаяся гликозидная связь имеет  -конфигурацию, как и в субстрате. Таким образом, эти данные подтверждают правильность предполагаемой формы промежуточного продукта катализа.
-конфигурацию, как и в субстрате. Таким образом, эти данные подтверждают правильность предполагаемой формы промежуточного продукта катализа.