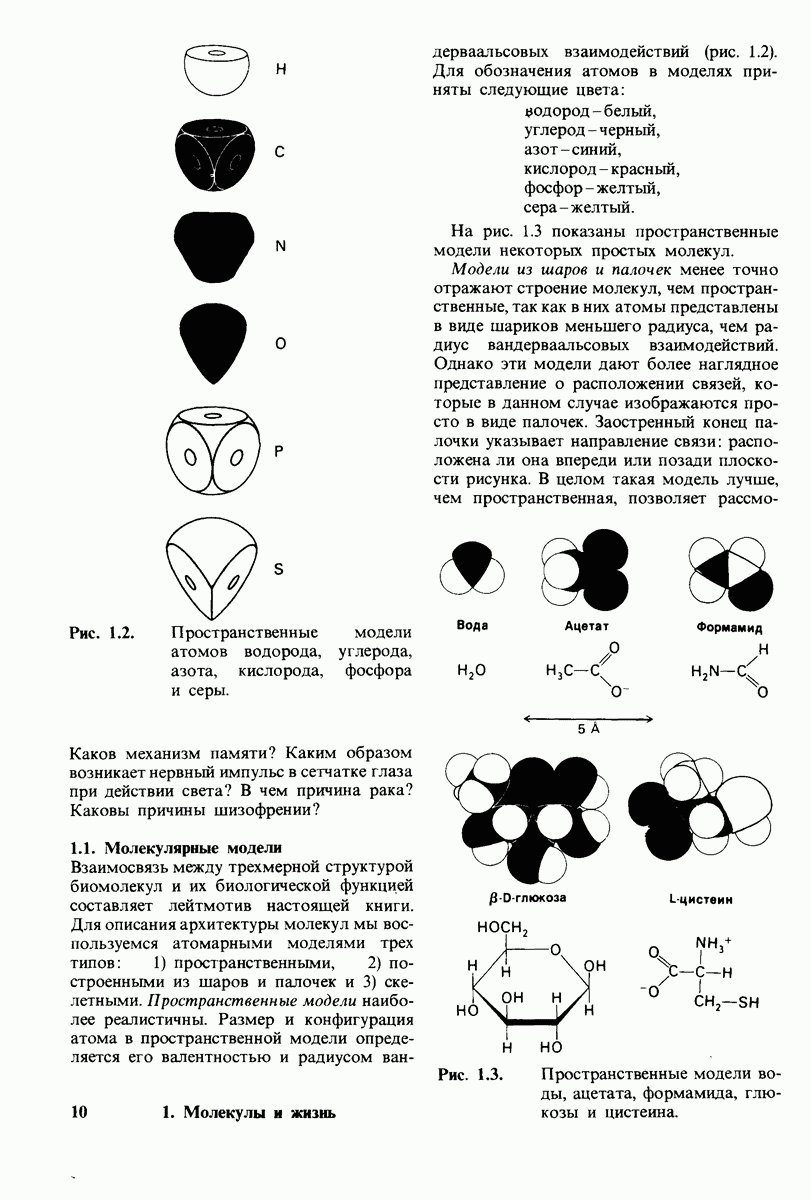
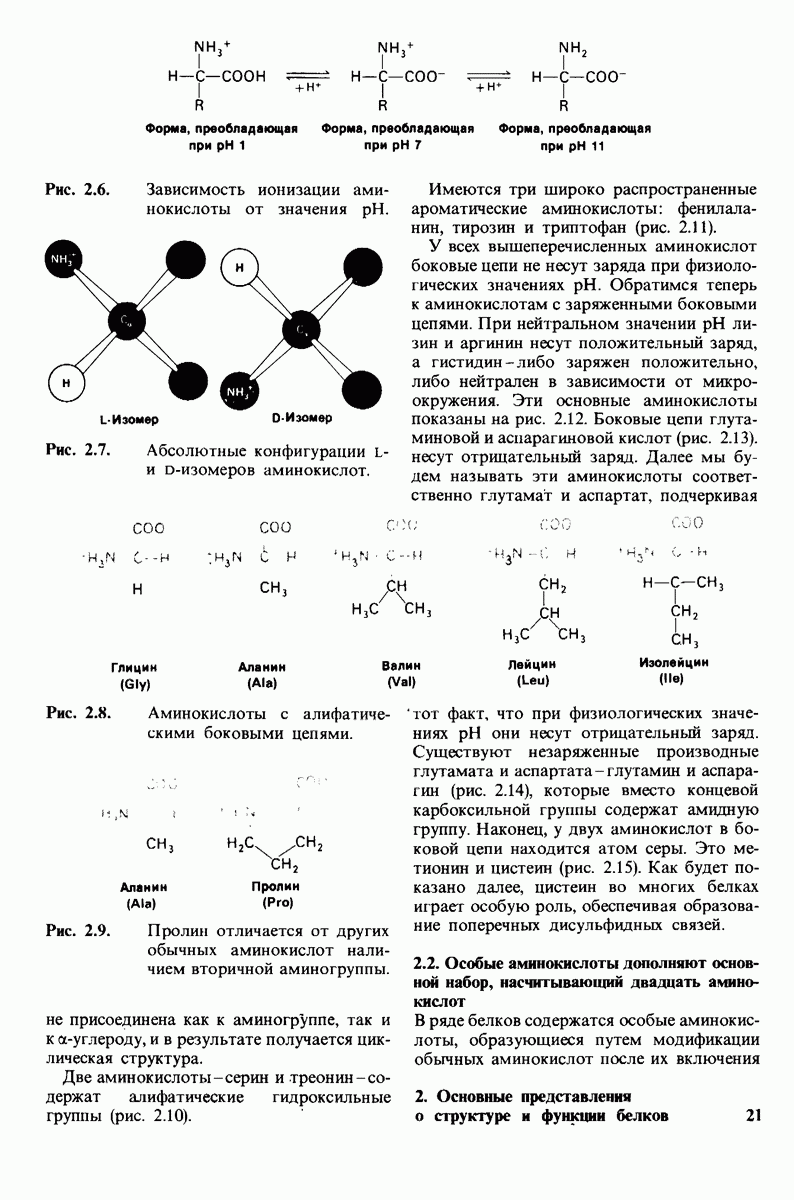
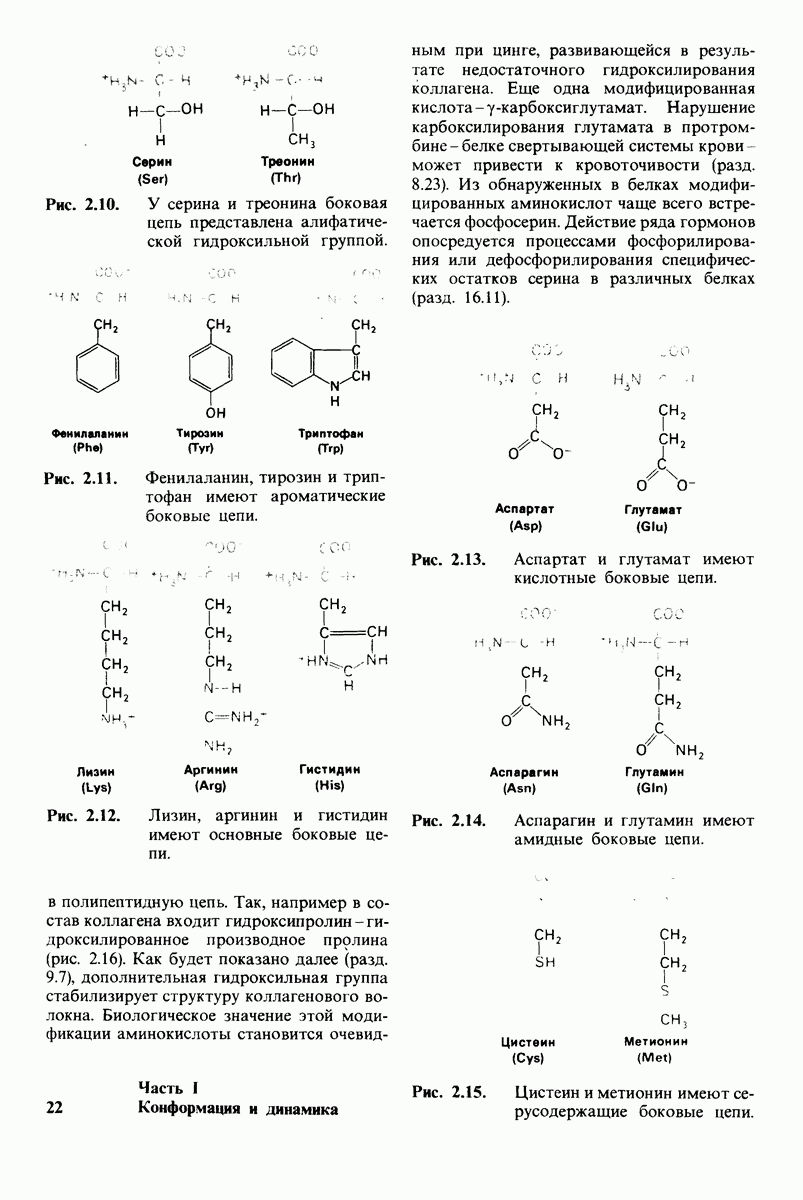
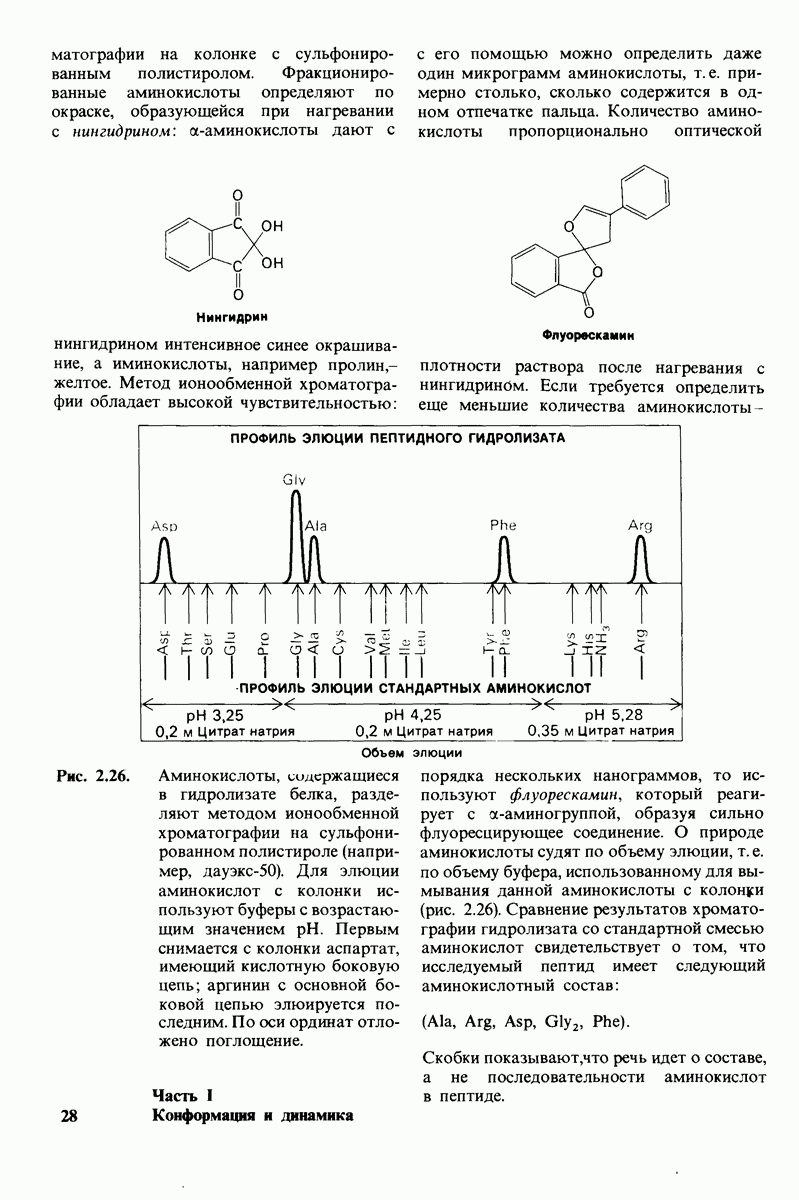
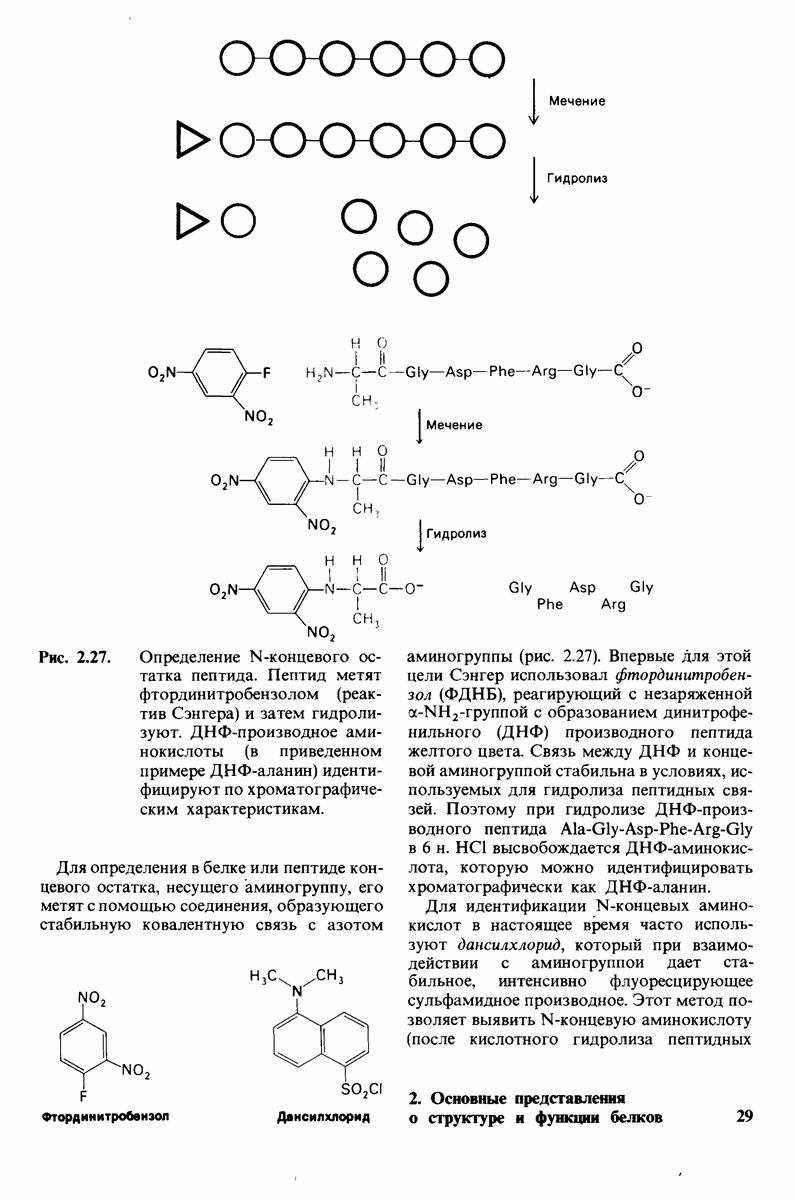
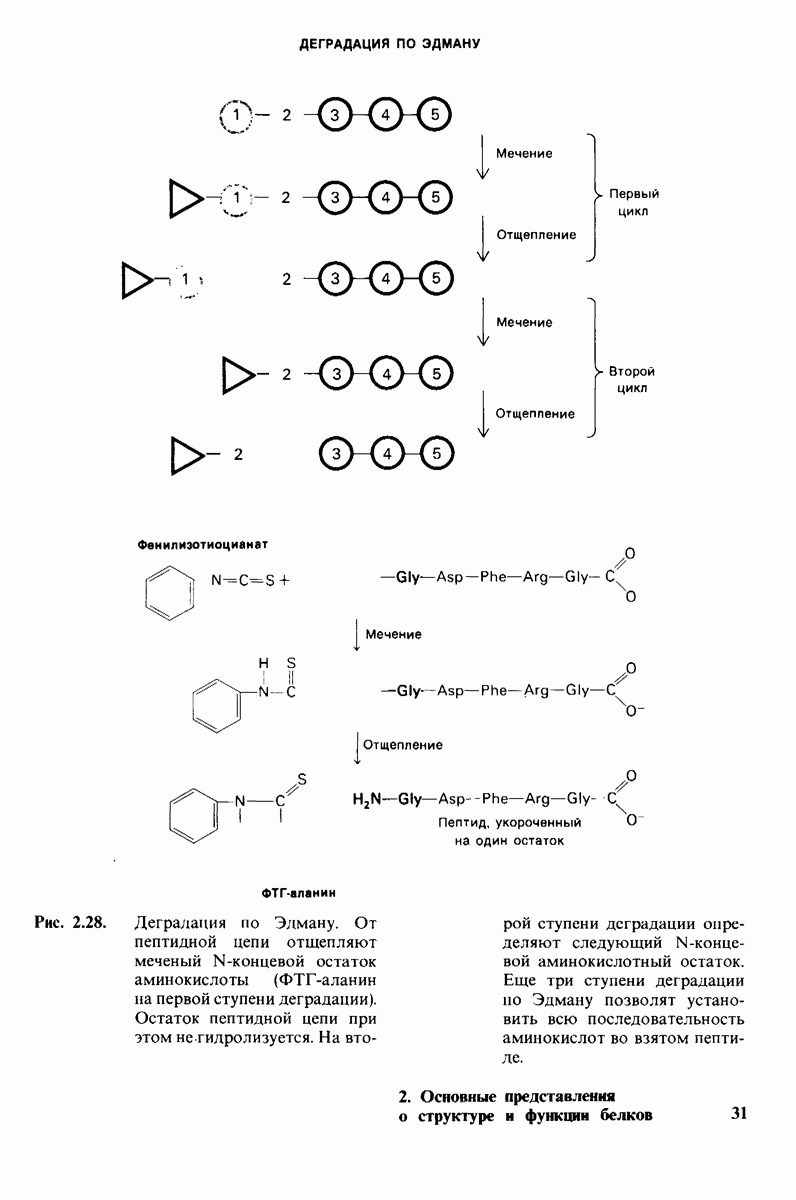
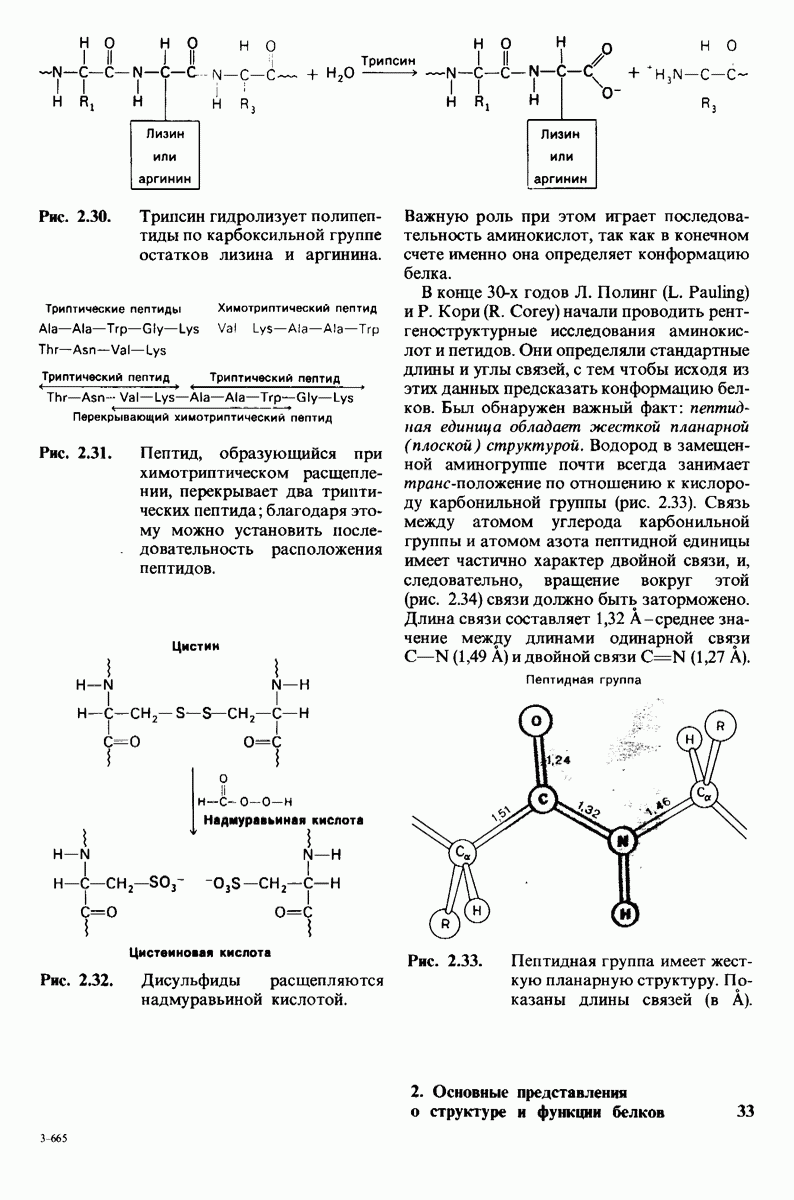
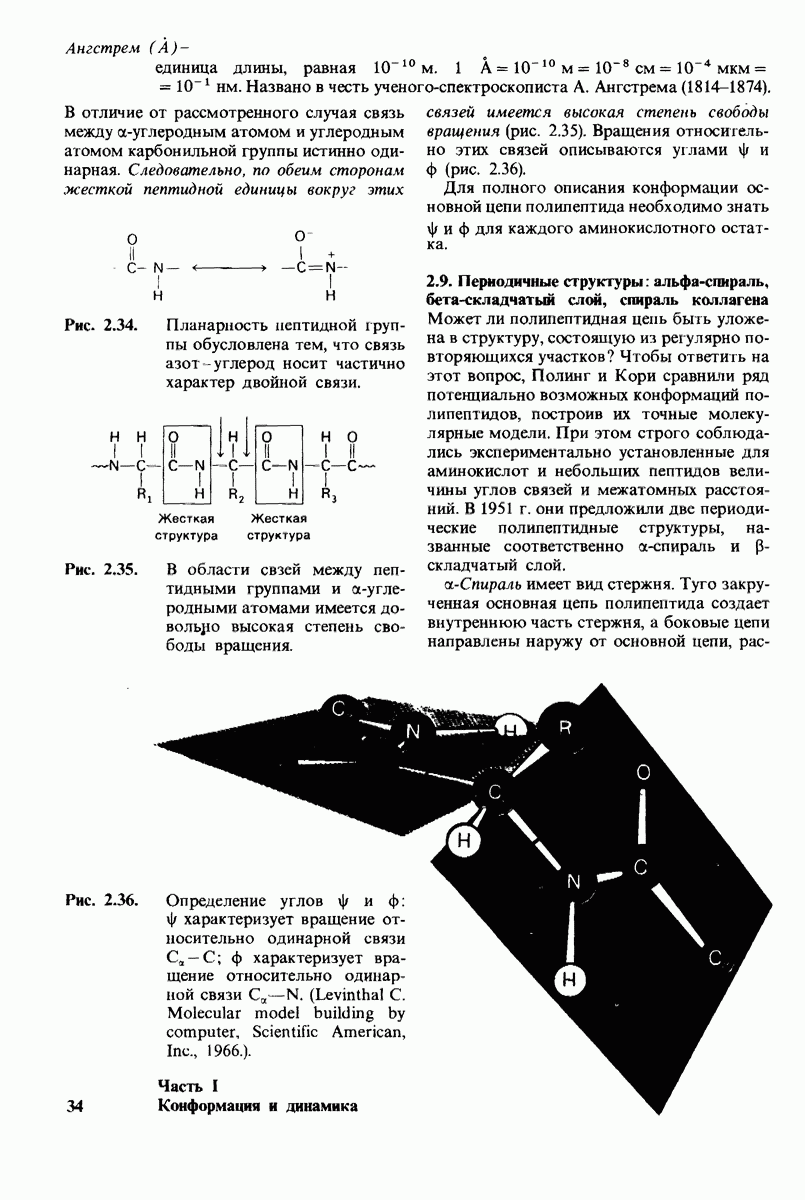
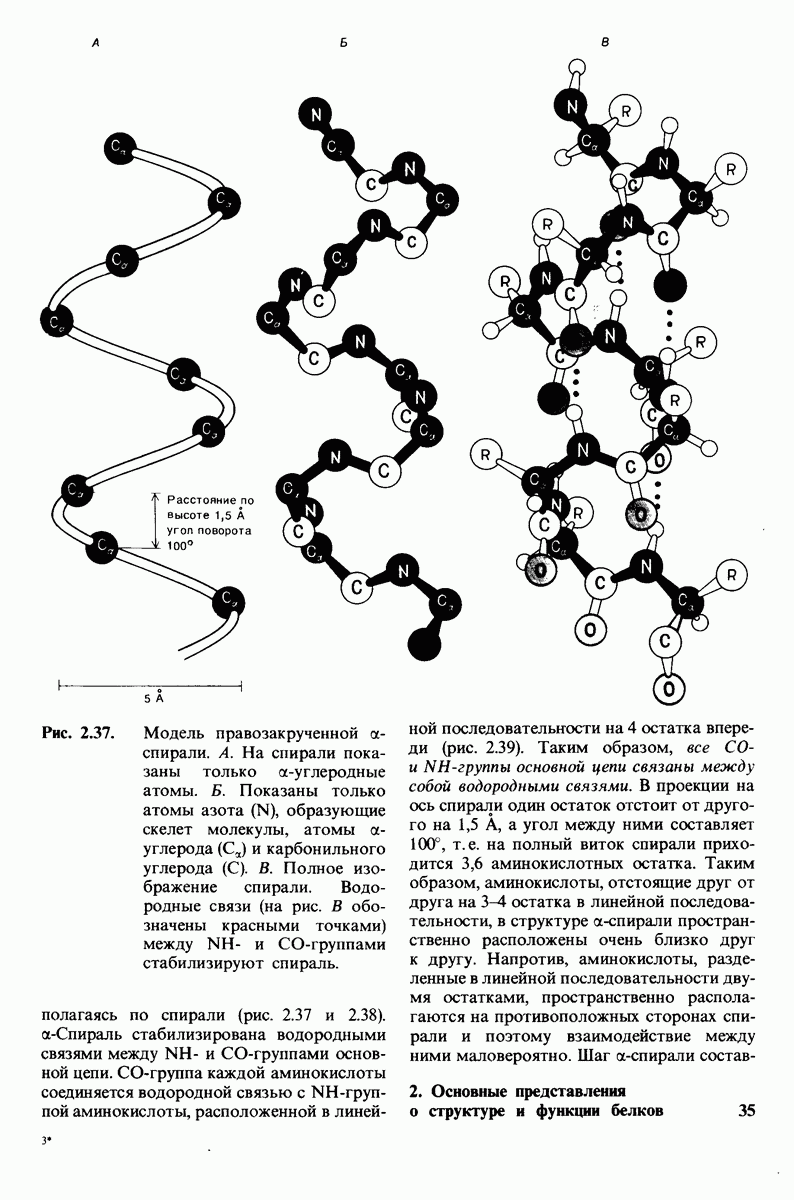
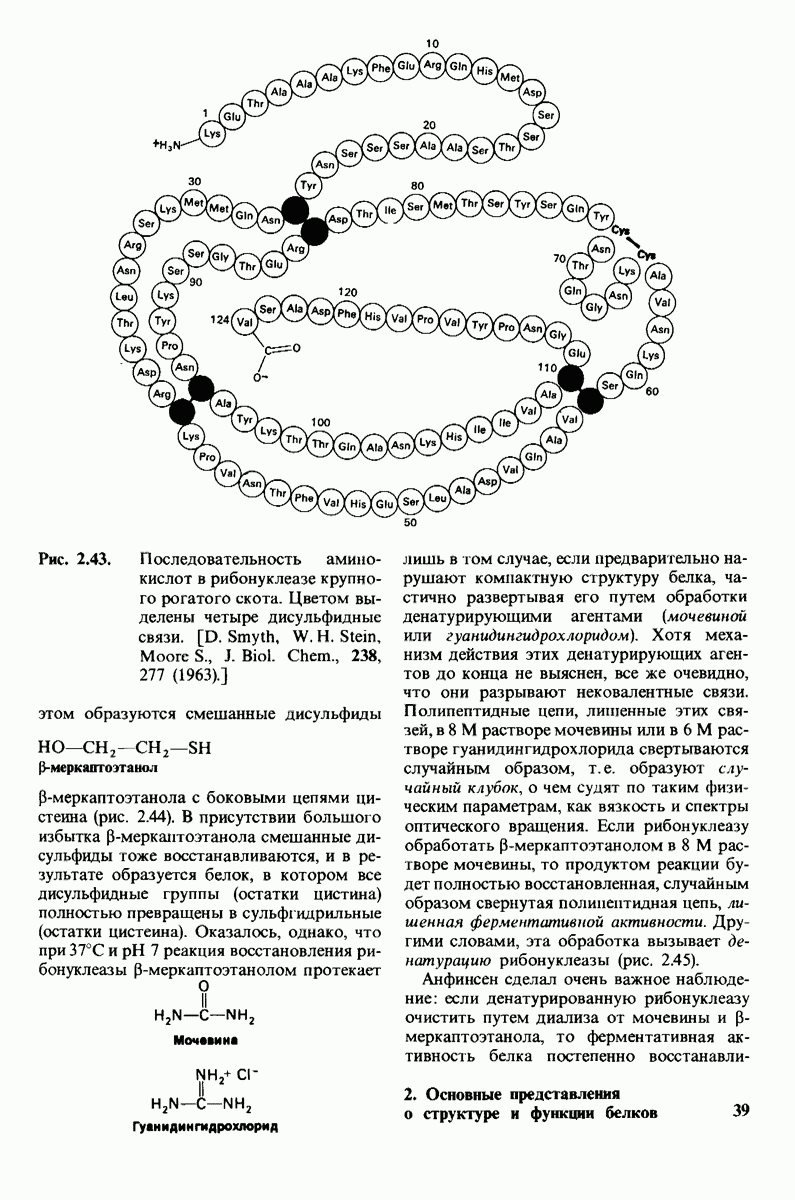
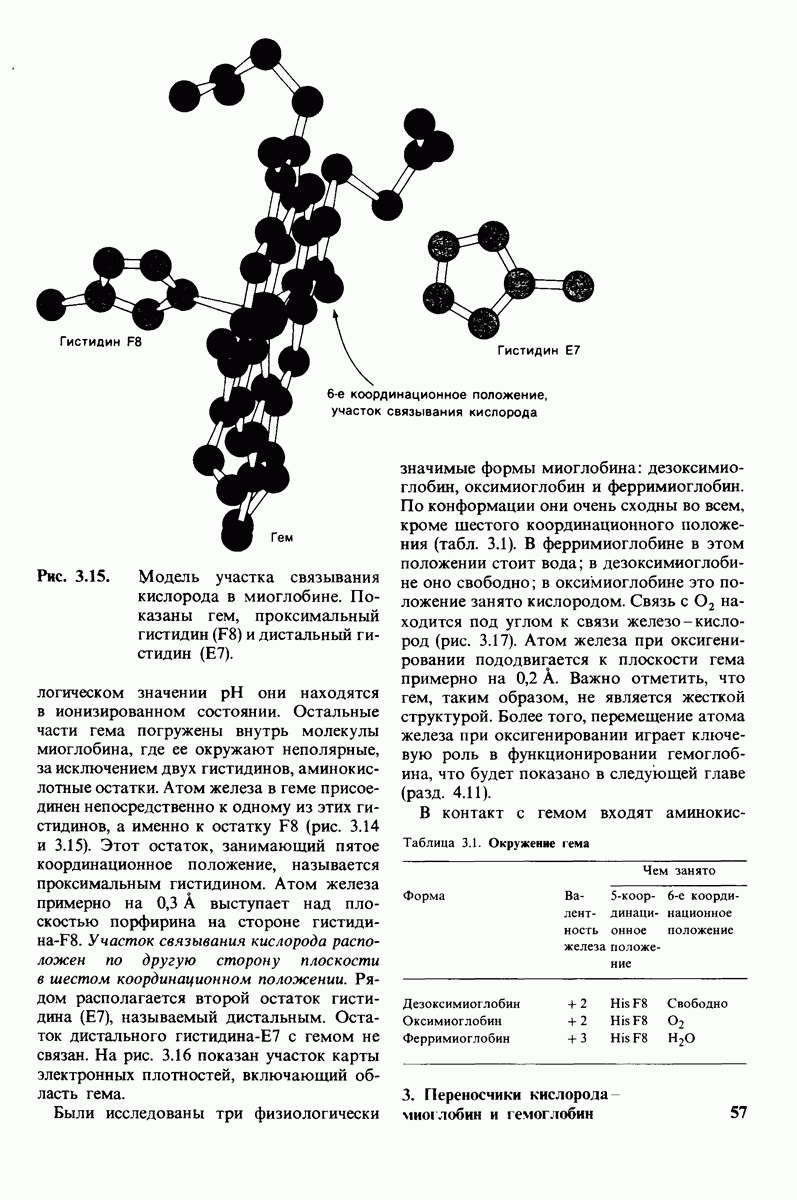
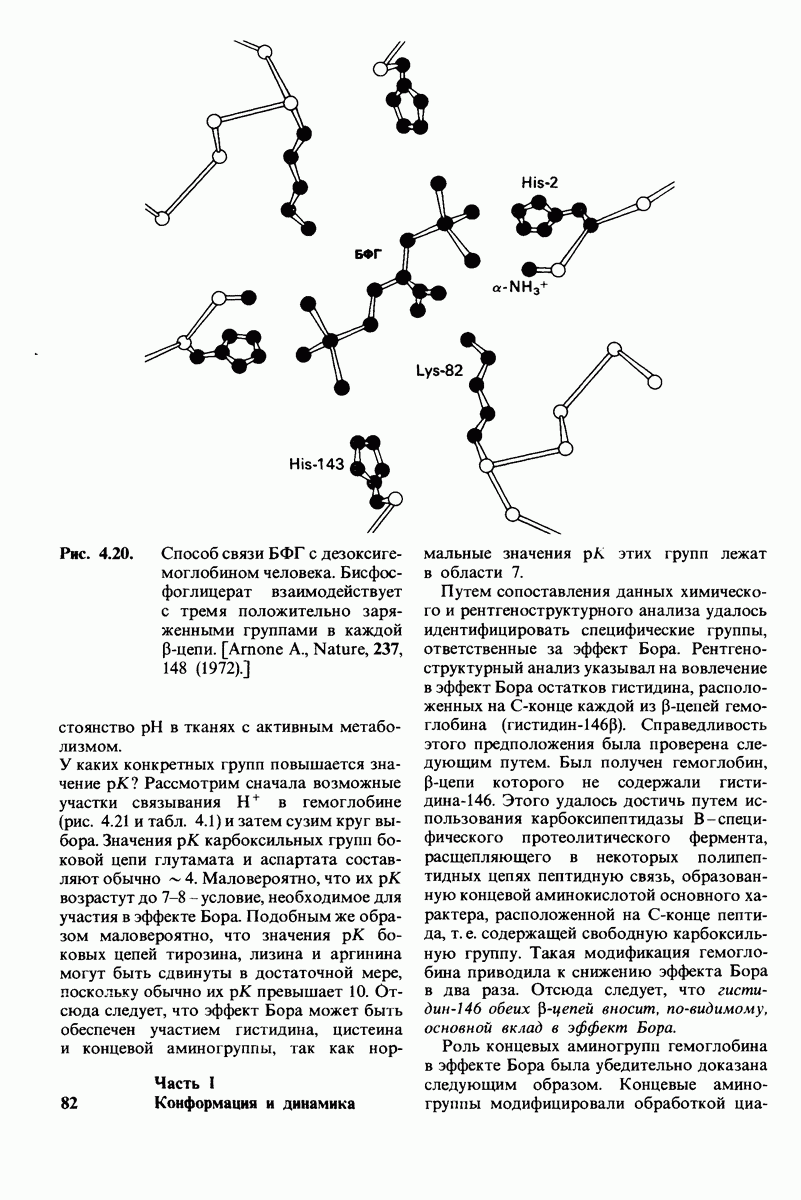
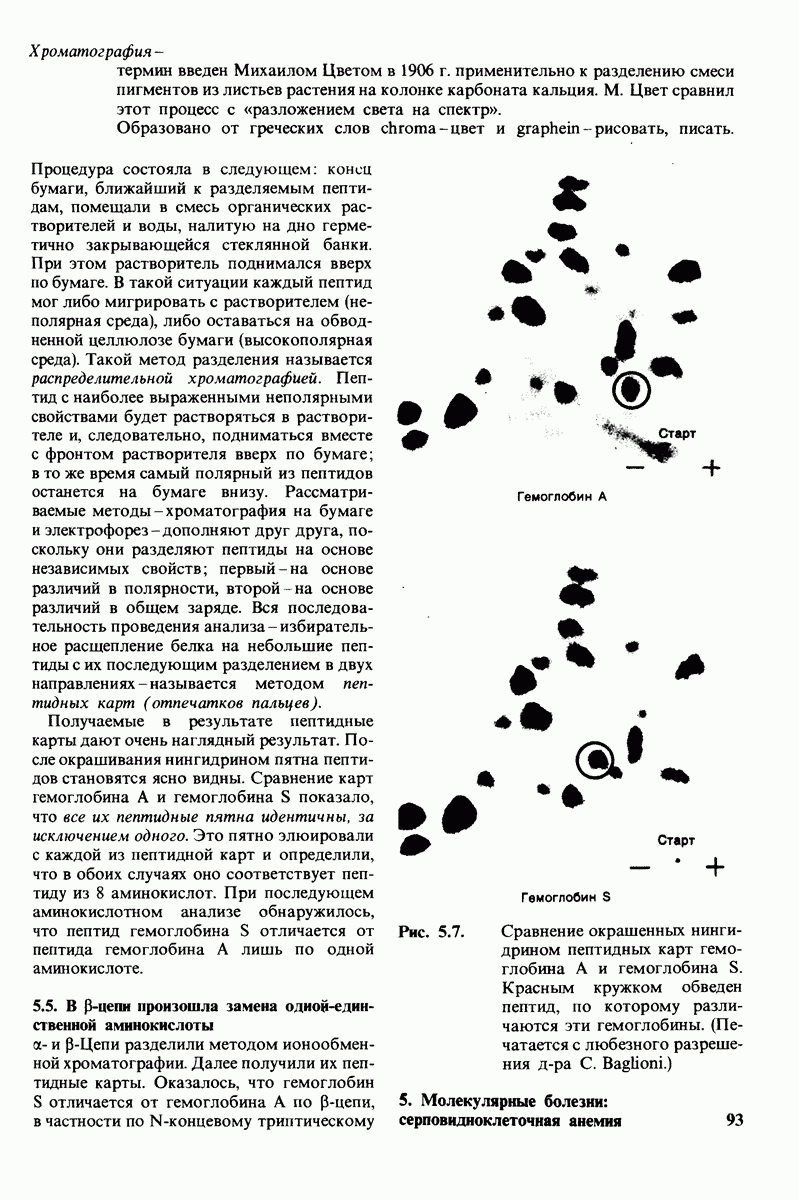
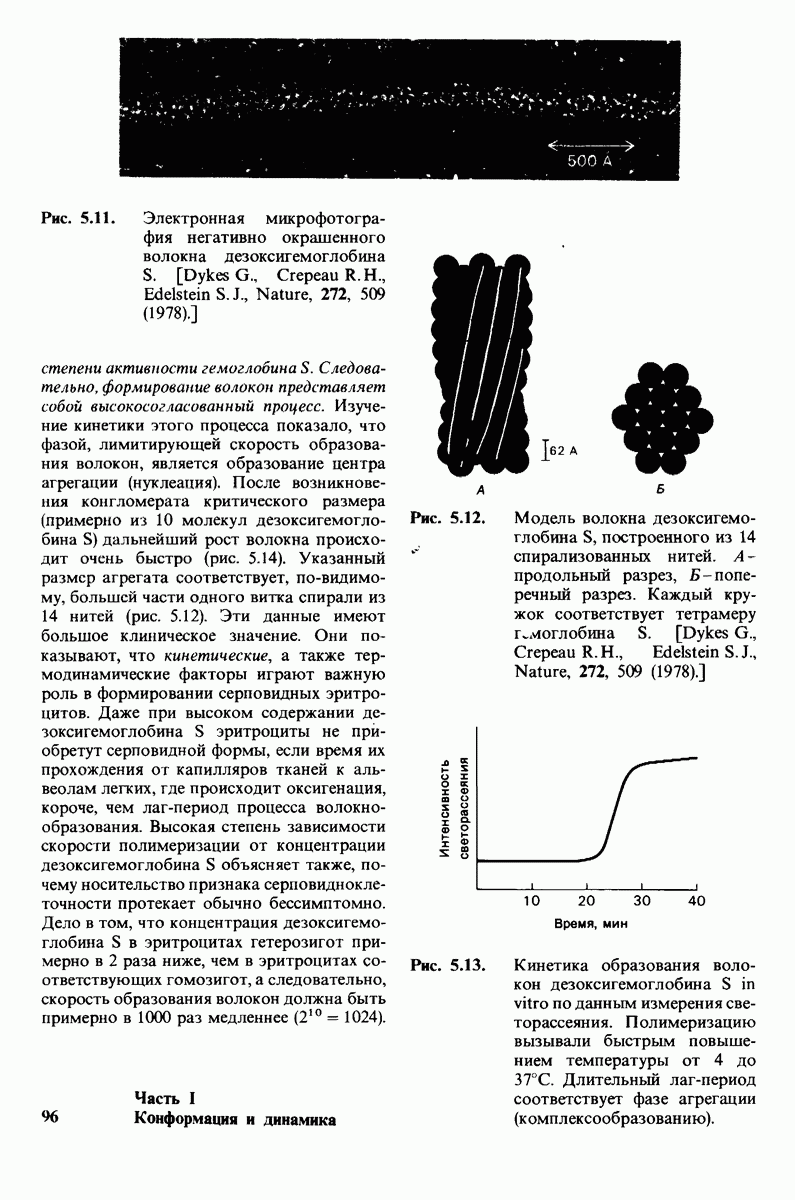
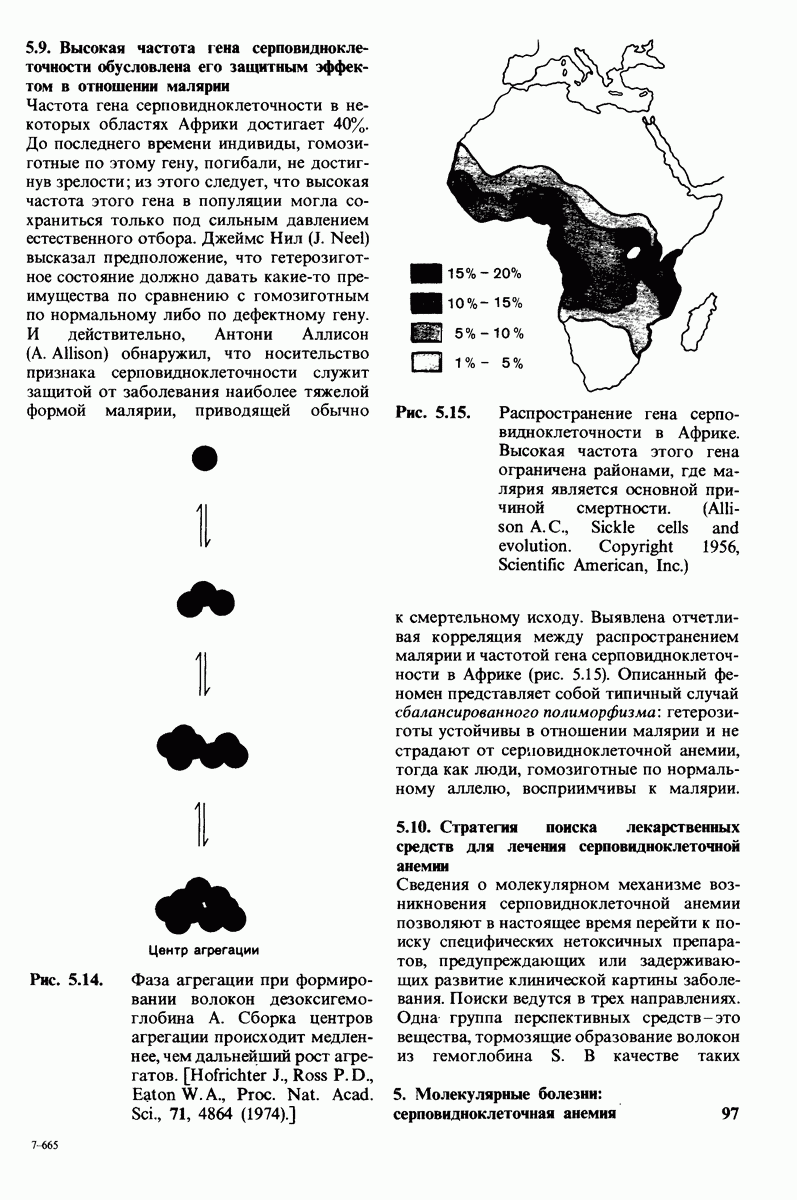
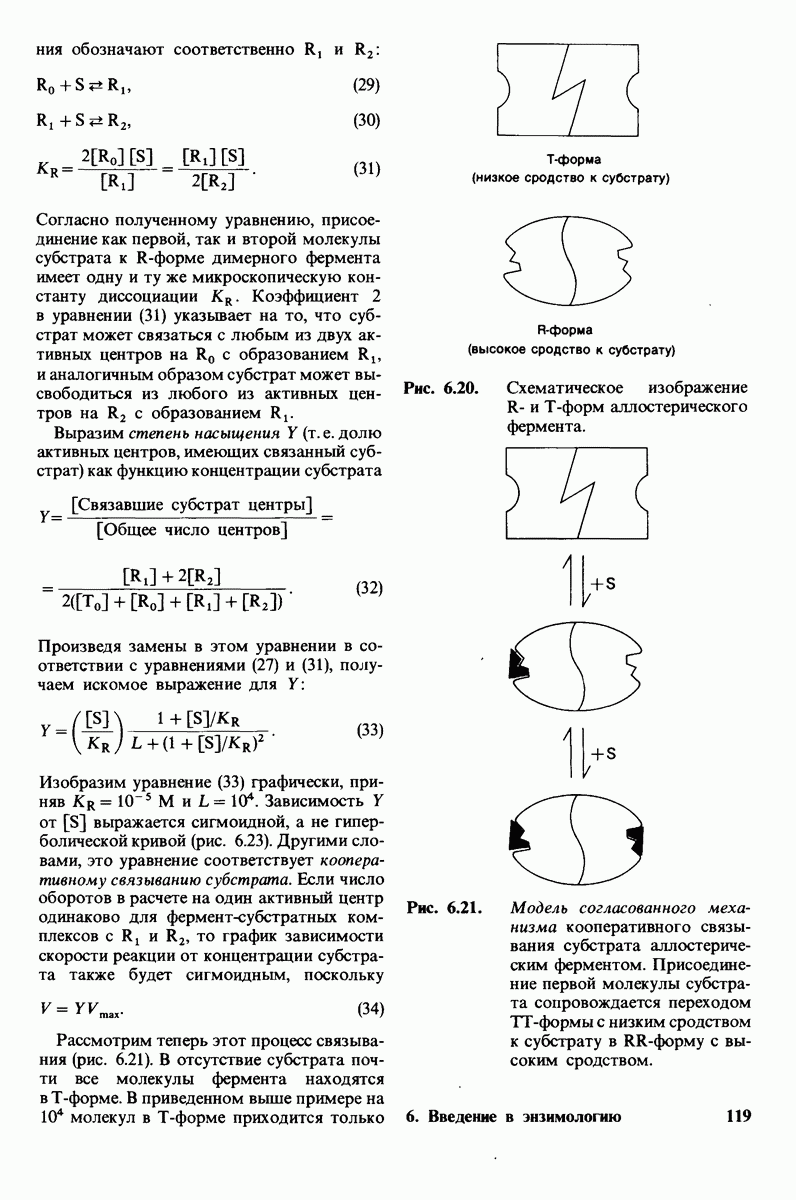
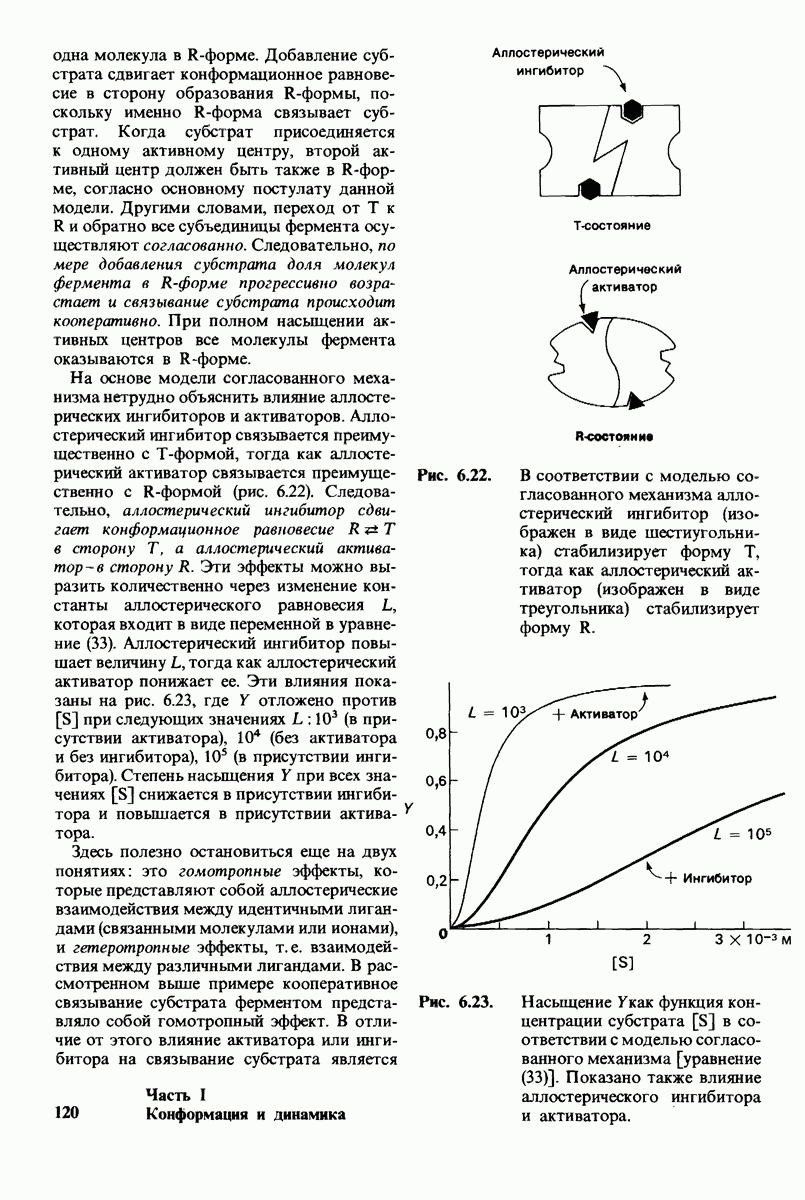
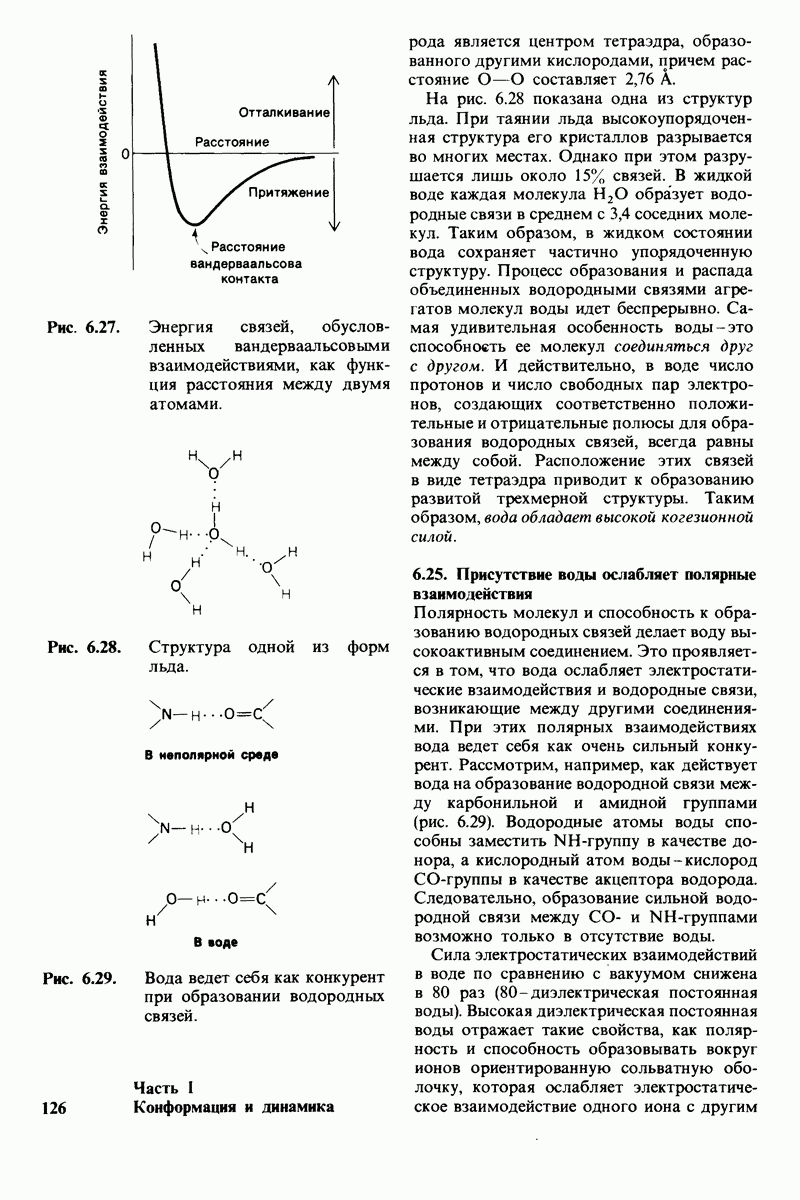
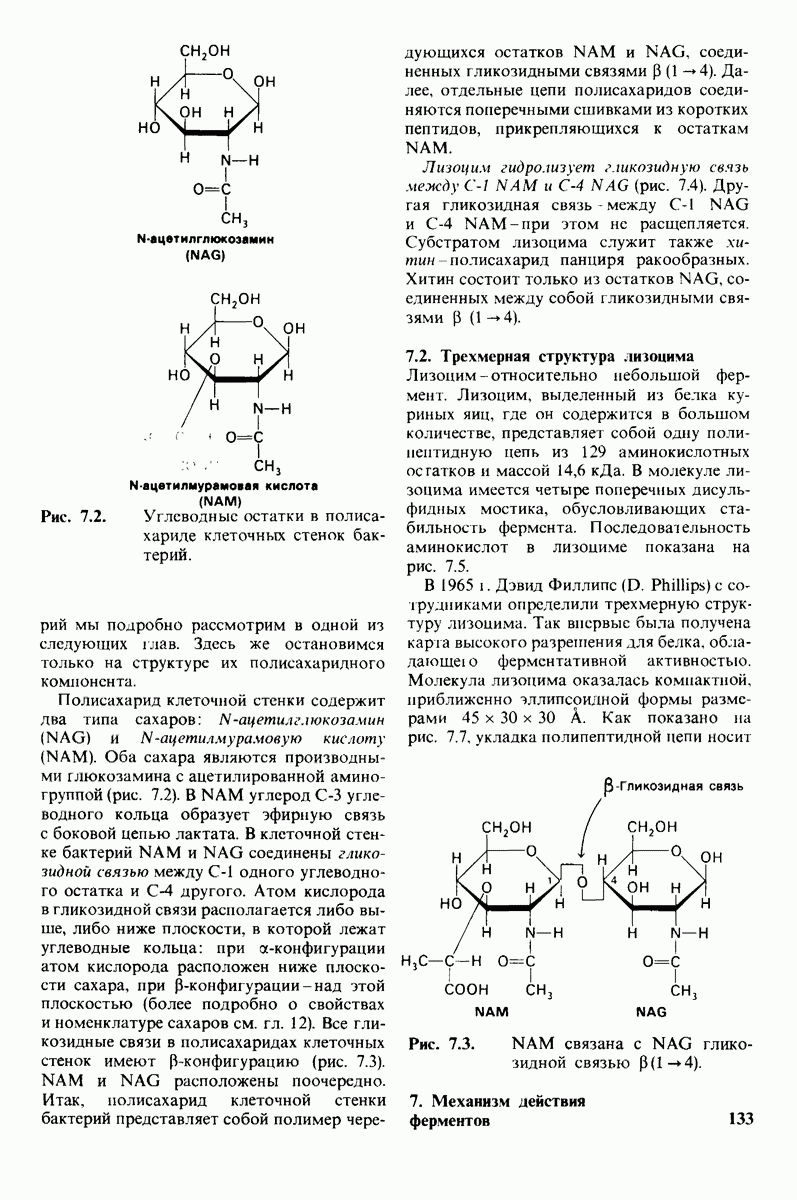
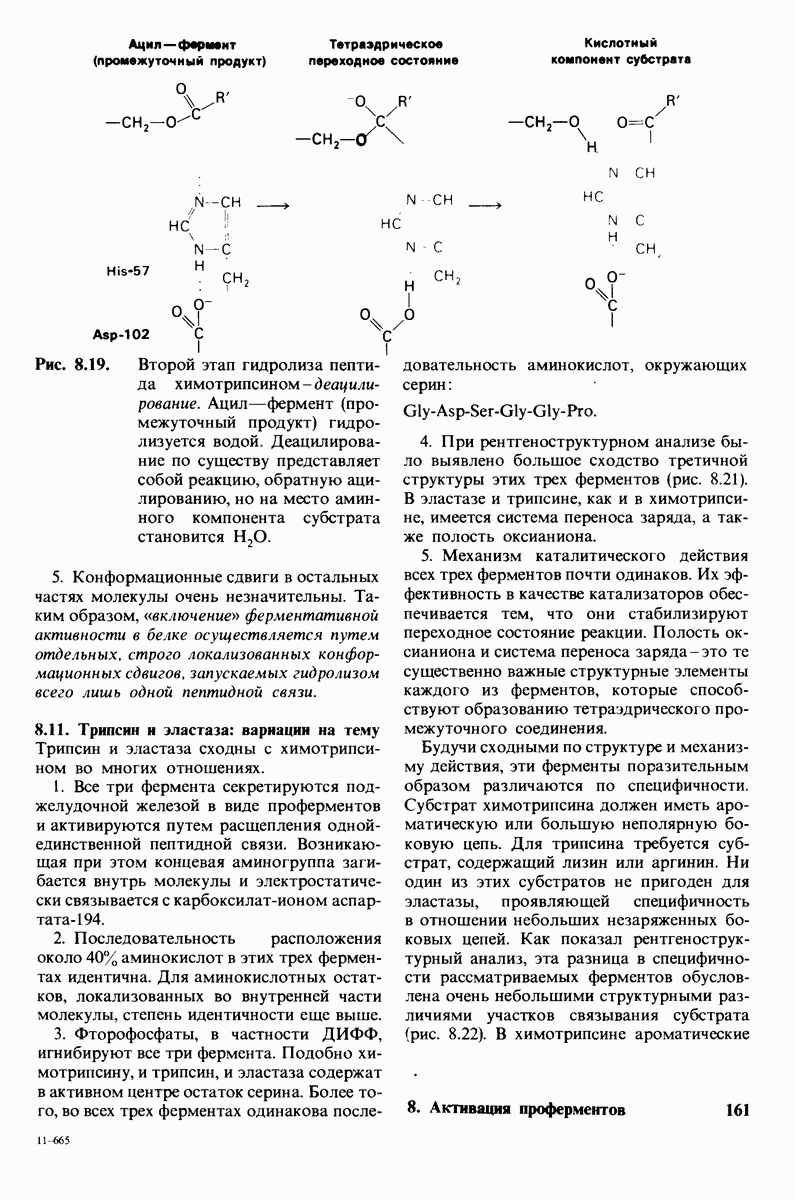
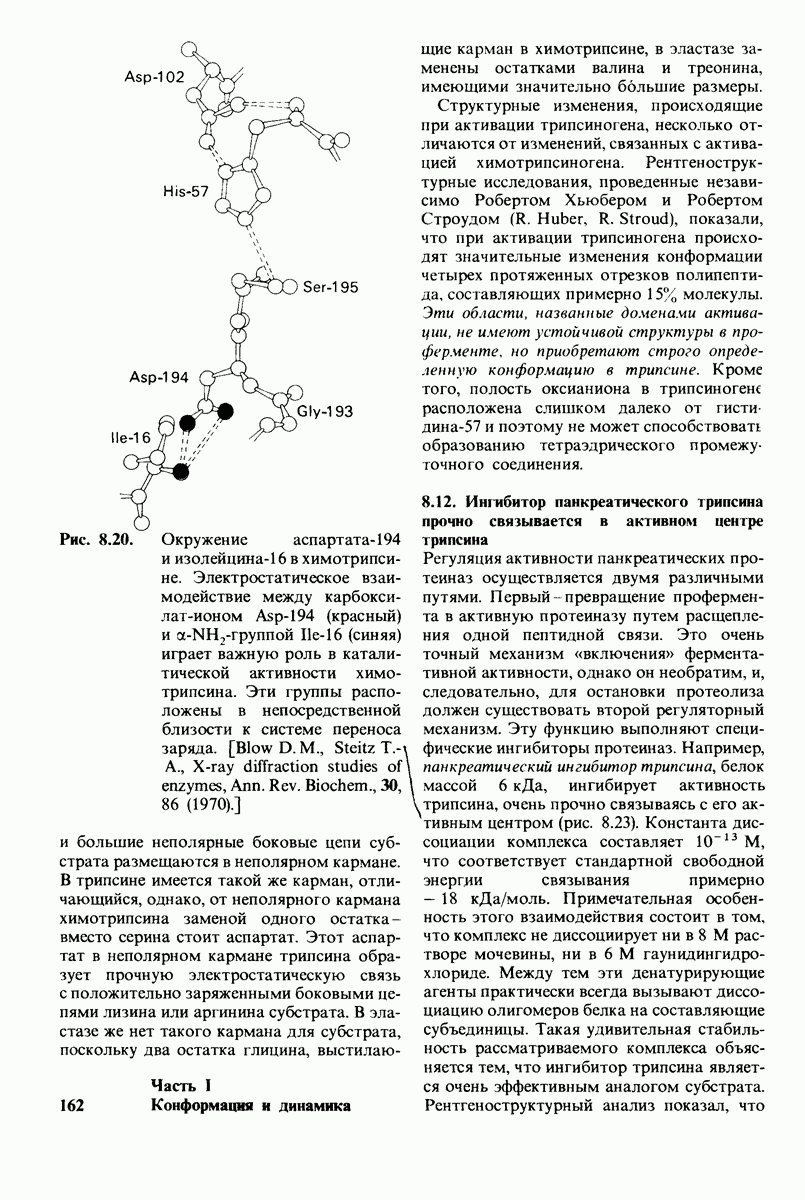
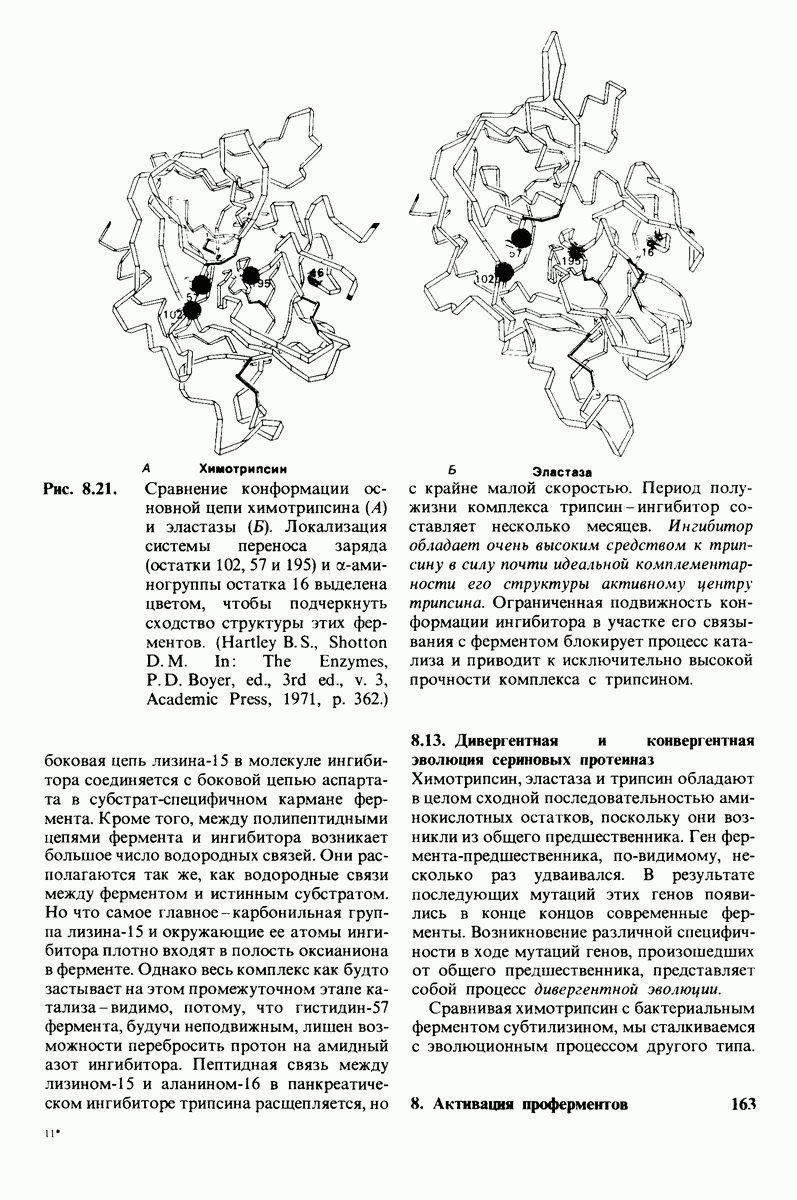
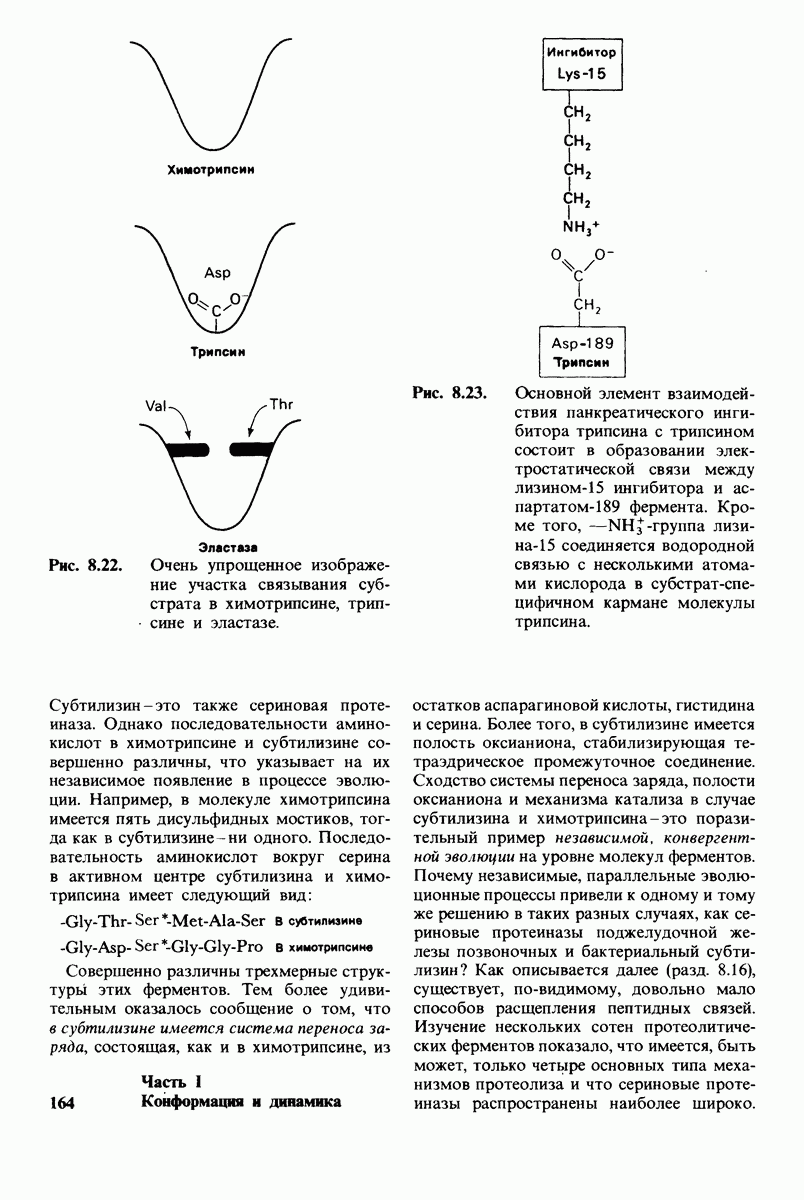
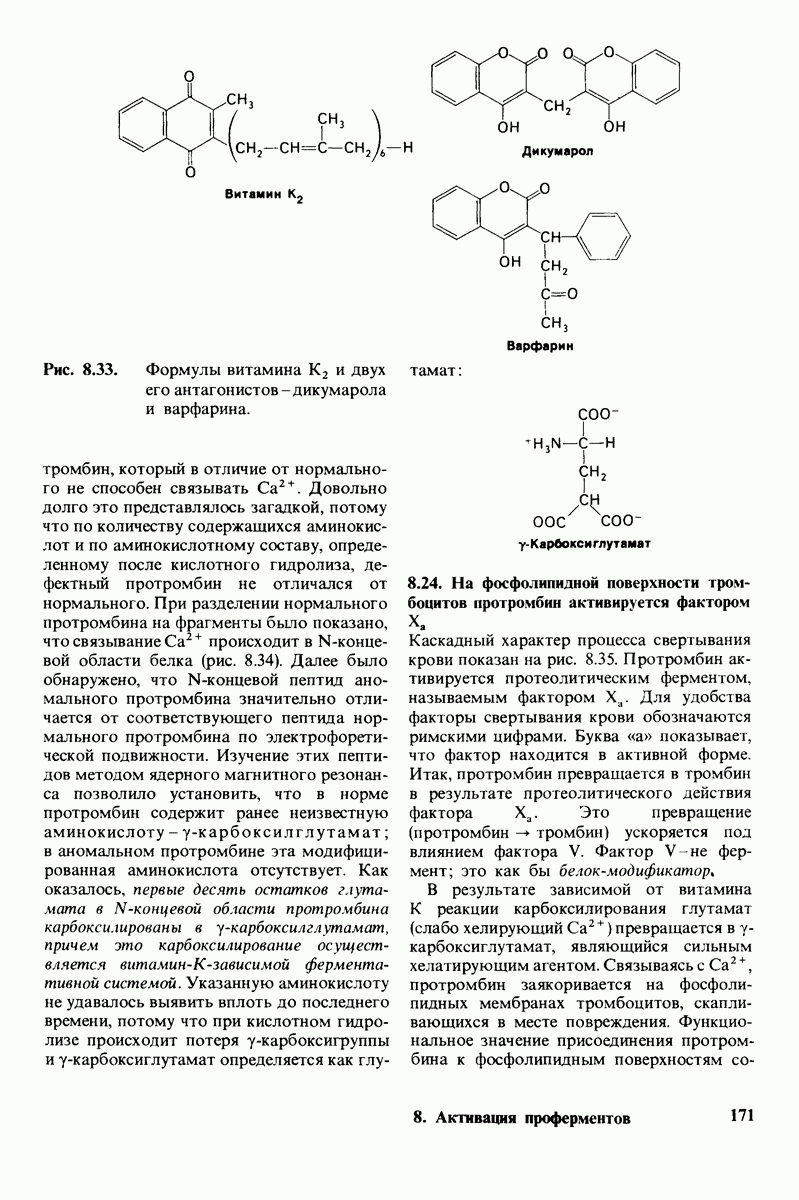
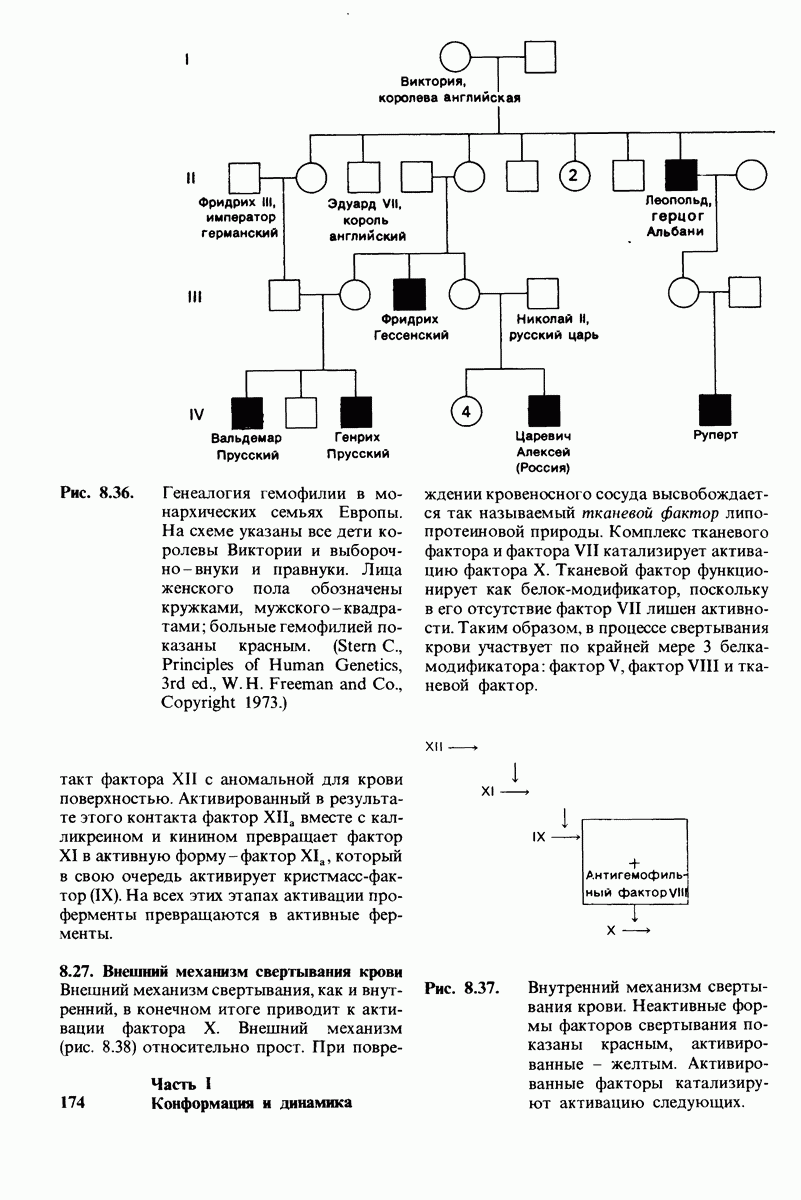
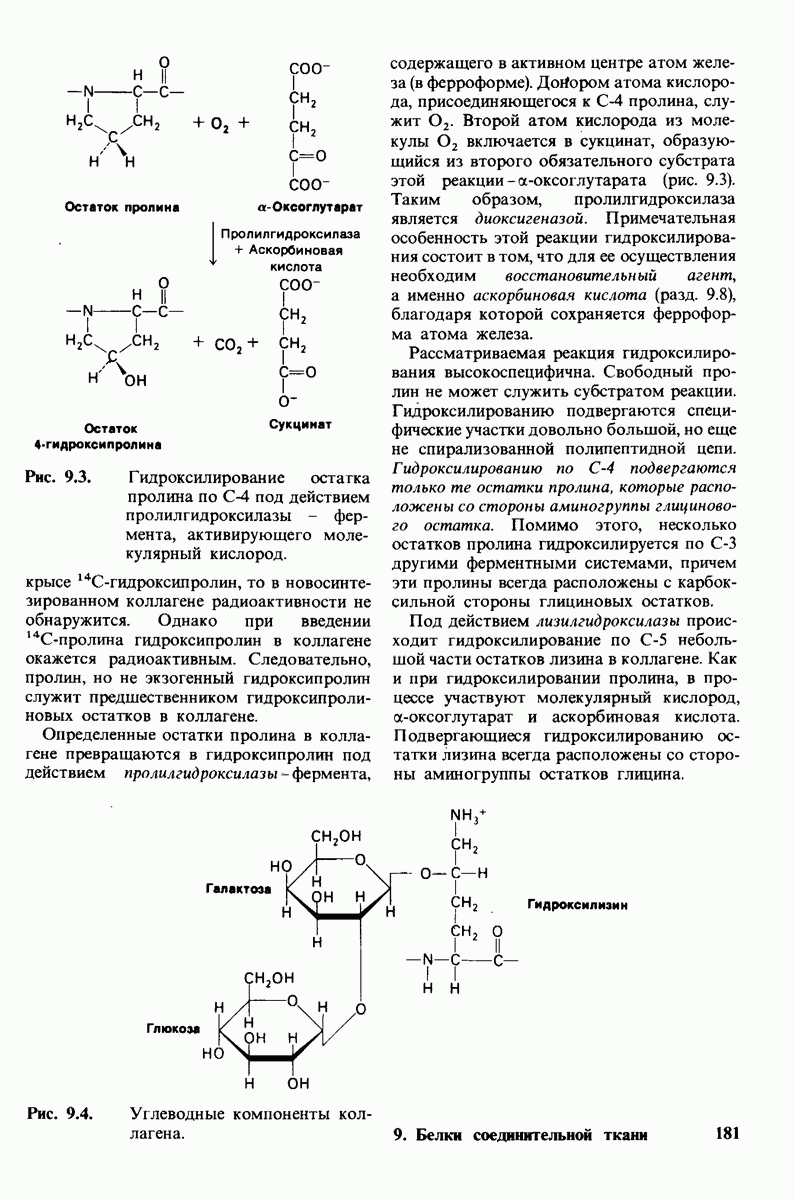
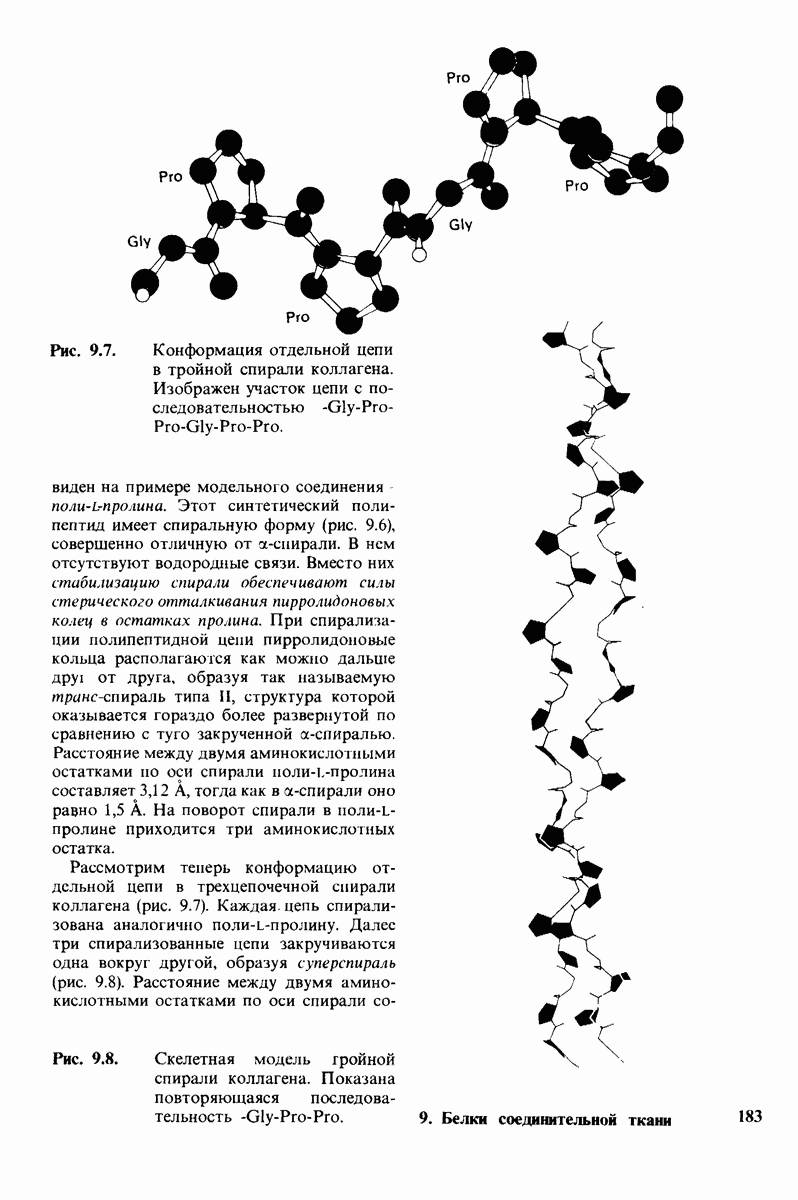
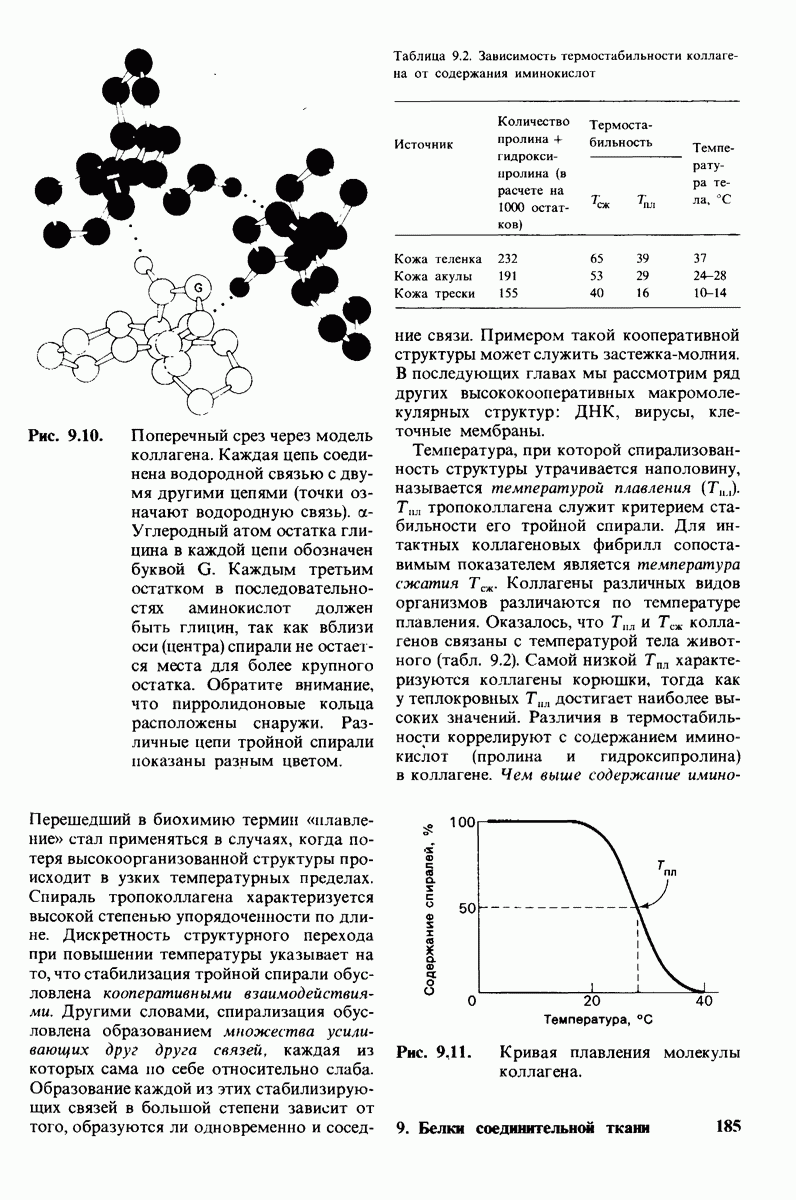
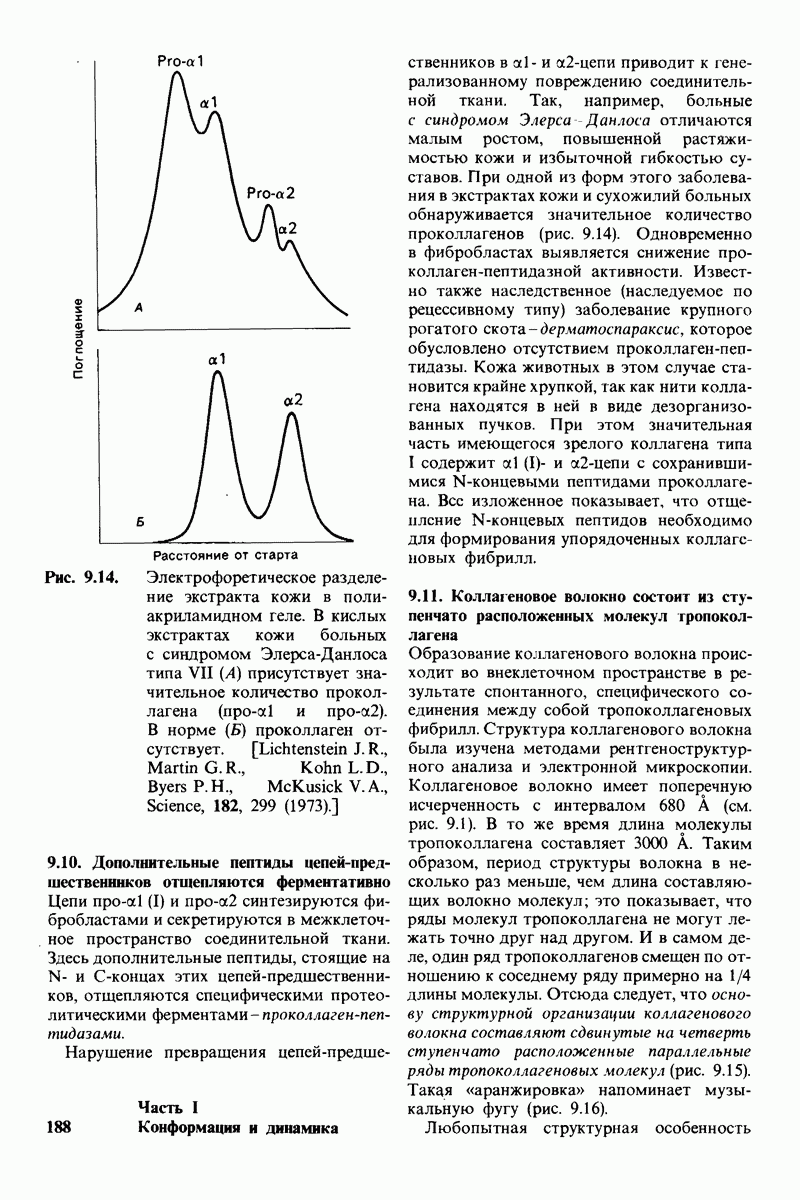
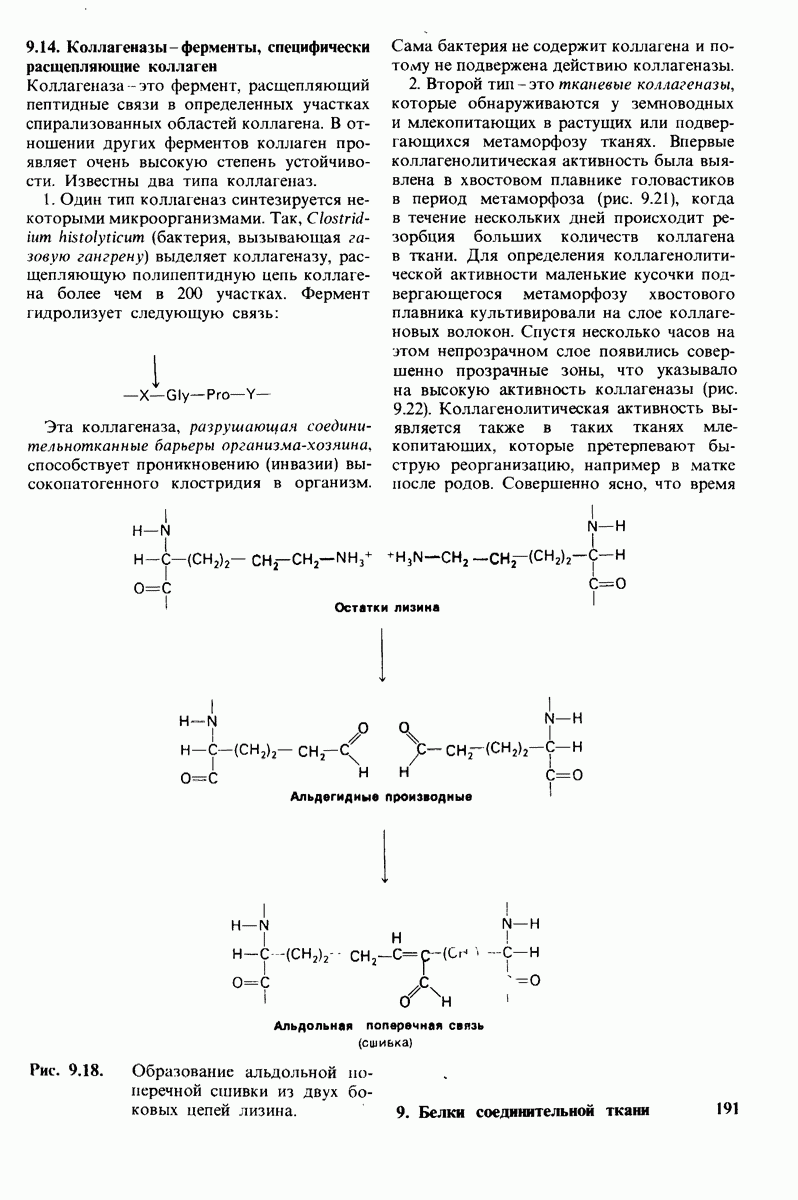
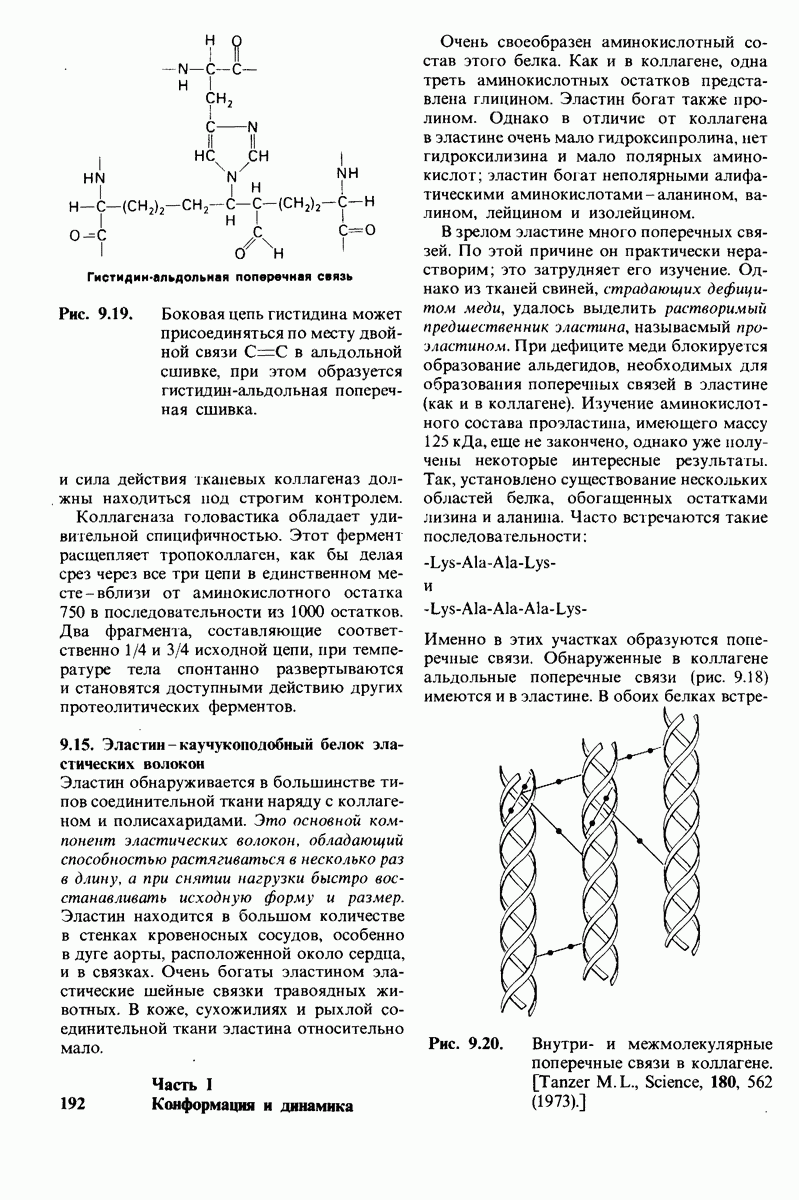
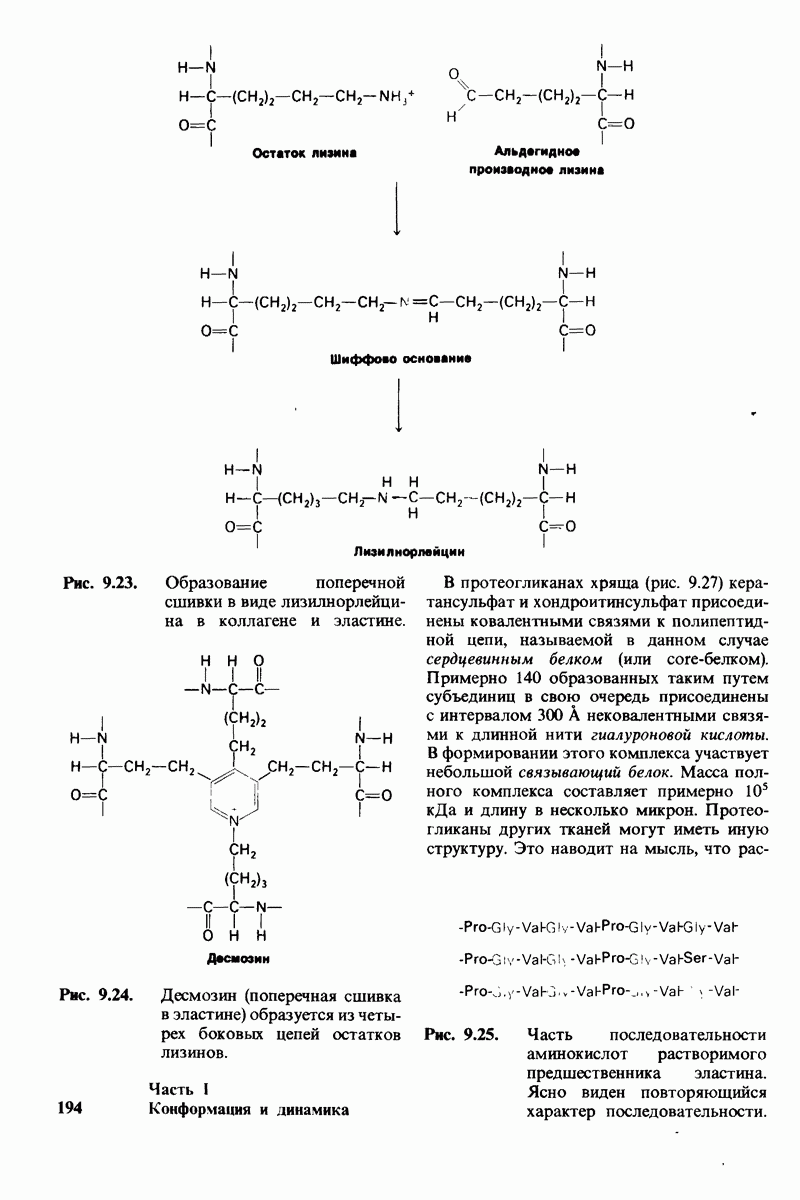
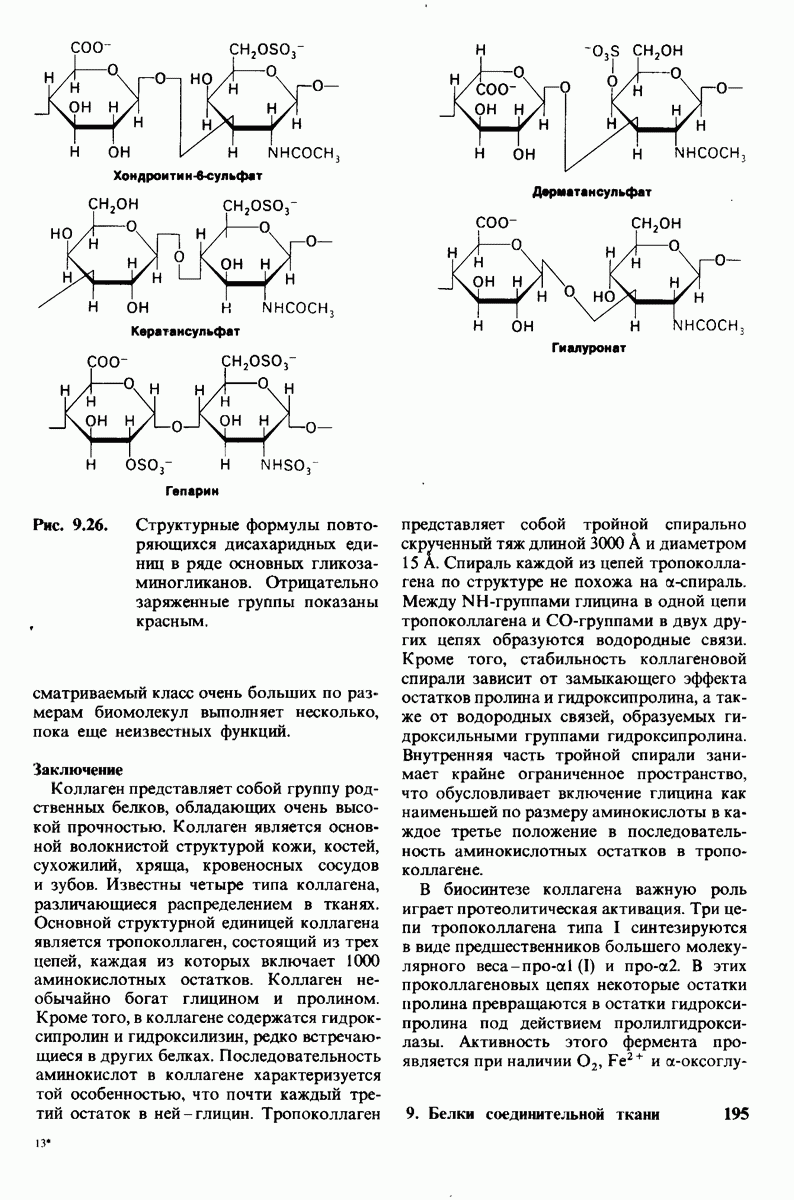
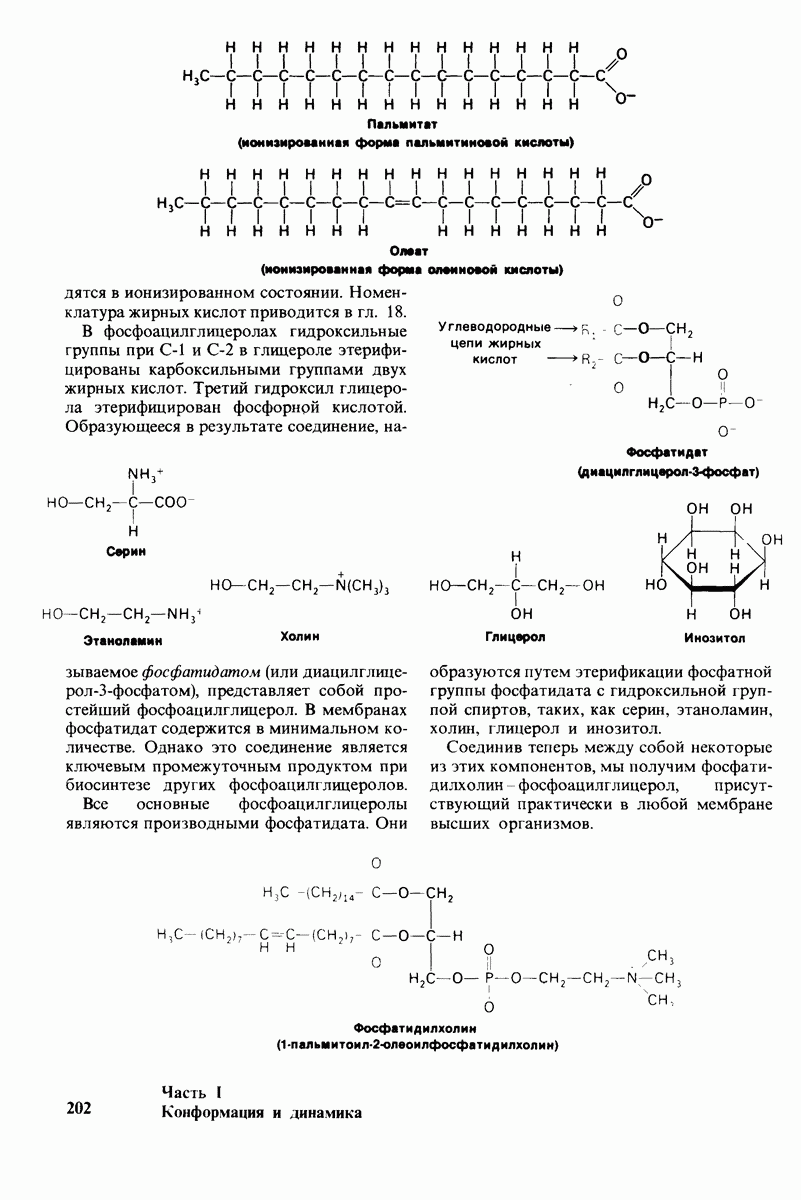
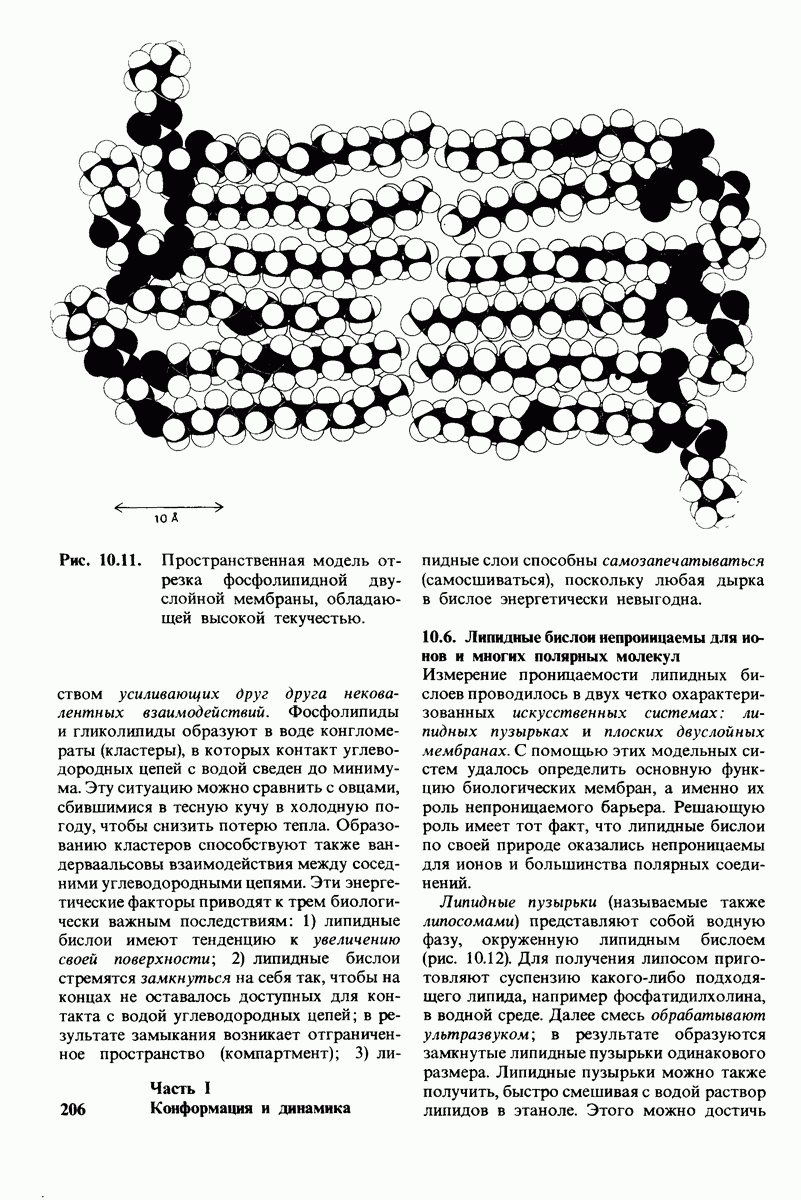
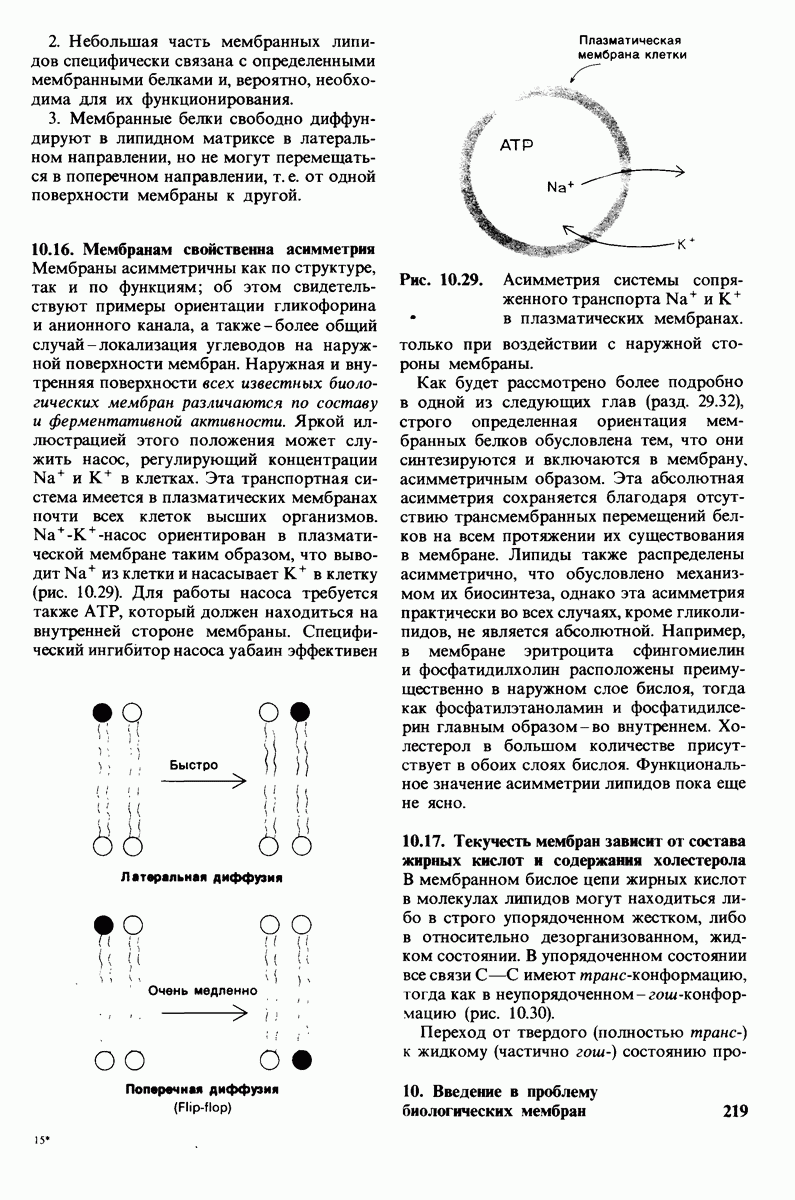
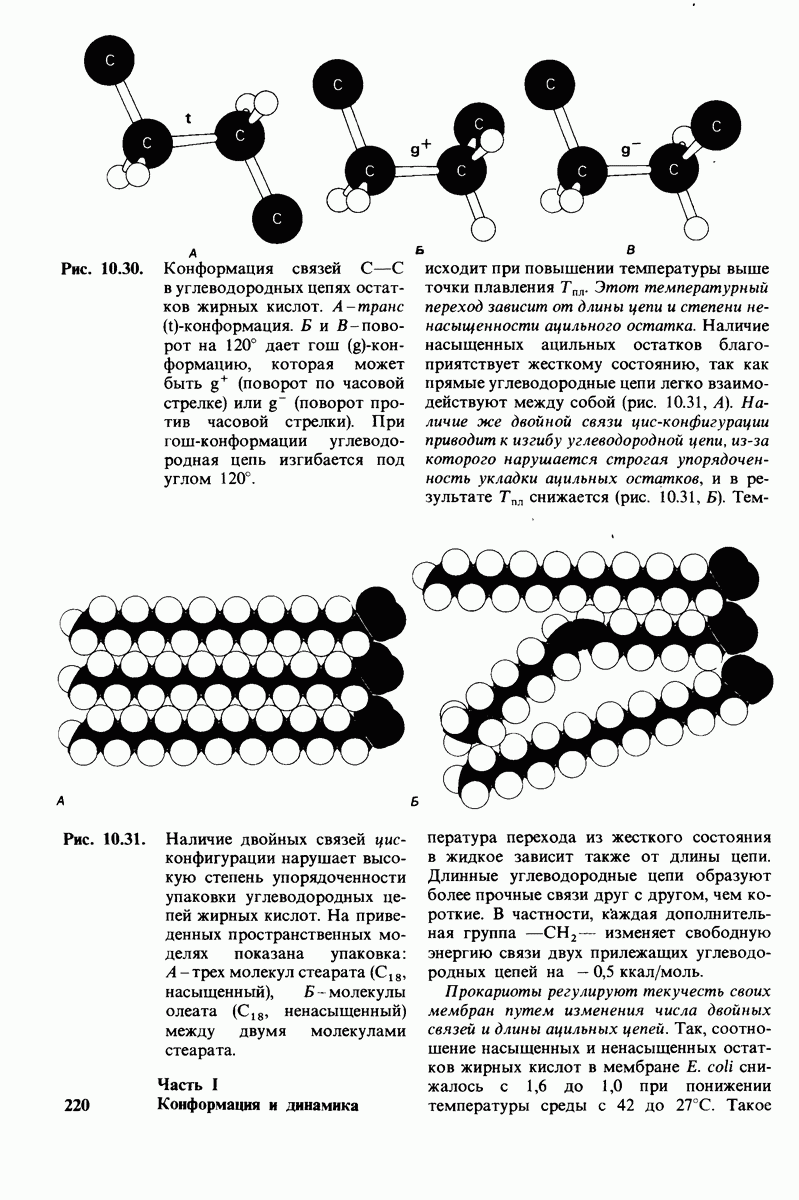
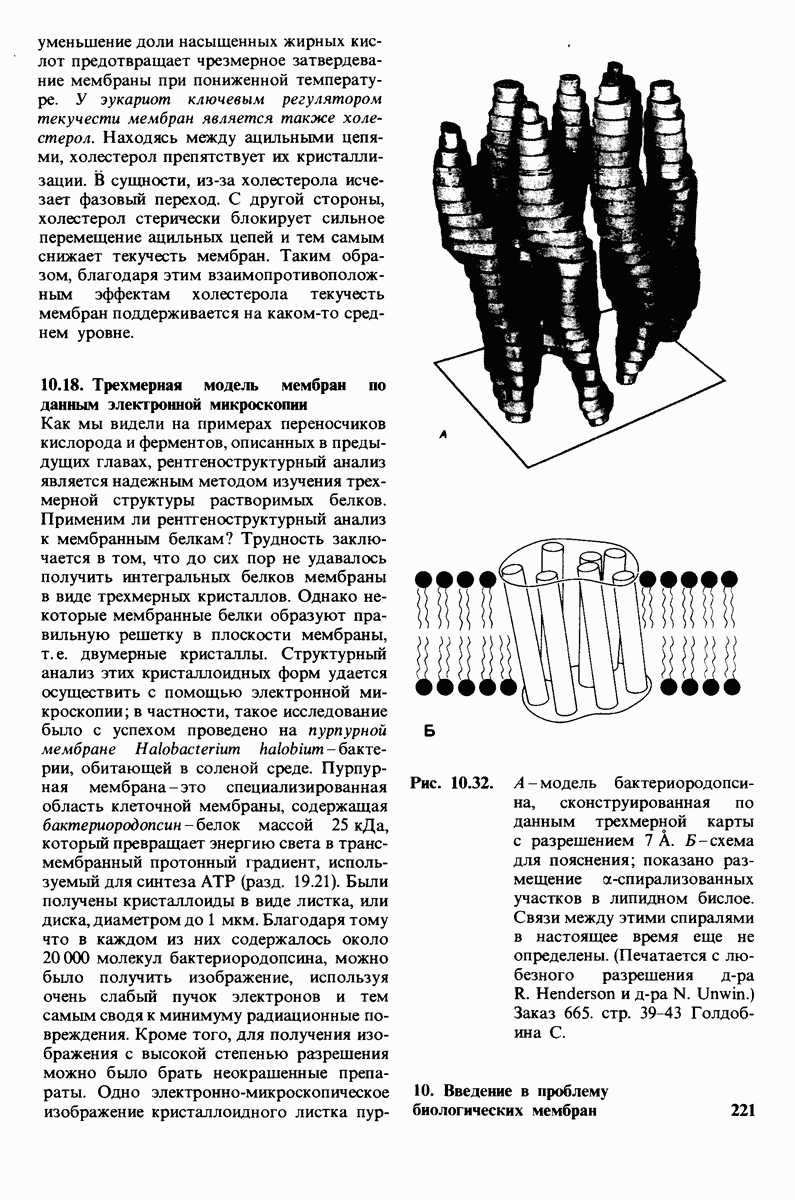
6.7. Первый этап ферментативного катализа - образование фермент-субстратного комплекса
Образованию или расщеплению химических связей каким-либо ферментом предшествует формирование фермент-субстратного  комплекса. При этом субстрат присоединяется к специфическому участку на ферменте, называемому активным центром. Большинство ферментов проявляет высокую избирательность в отношении связывания субстратов. В сущности, специфичность каталитического действия ферментов в основном зависит от специфичности процесса связывания. Более того, на этой стадии нередко осуществляется и регуляция ферментативной активности.
комплекса. При этом субстрат присоединяется к специфическому участку на ферменте, называемому активным центром. Большинство ферментов проявляет высокую избирательность в отношении связывания субстратов. В сущности, специфичность каталитического действия ферментов в основном зависит от специфичности процесса связывания. Более того, на этой стадии нередко осуществляется и регуляция ферментативной активности.
Существование фермент-субстратных комплексов было доказано разными способами.
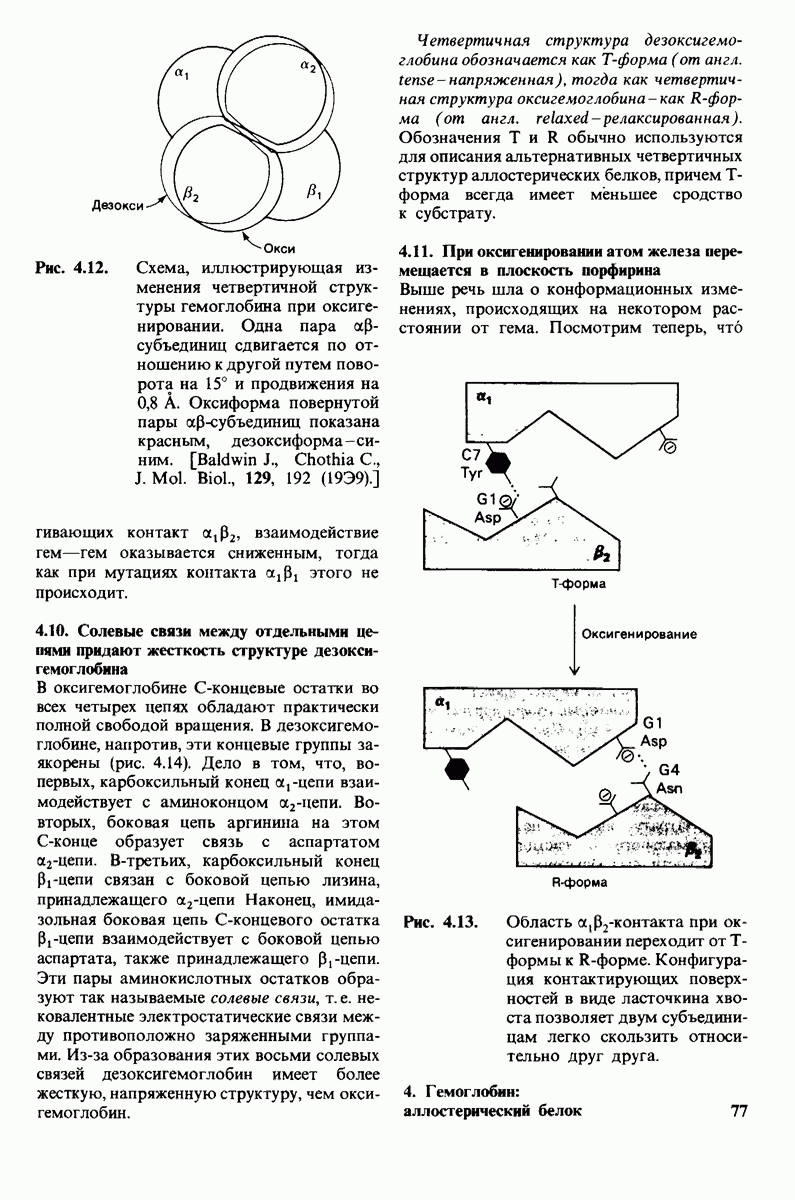
1. ES-комплексы были непосредственно выявлены с помощью электронной микроскопии и рентгеноструктурного анализа. Комплексы нуклеиновых кислот и их полимераз видны под электронным микроскопом (рис. 6.3). При рентгеноструктурном анализе комплекса карбоксипептидазы А с ее субстратом  была получена подробная информация относительно места и характера связывания субстрата с ферментом в этом
была получена подробная информация относительно места и характера связывания субстрата с ферментом в этом  -комплексе.
-комплексе.

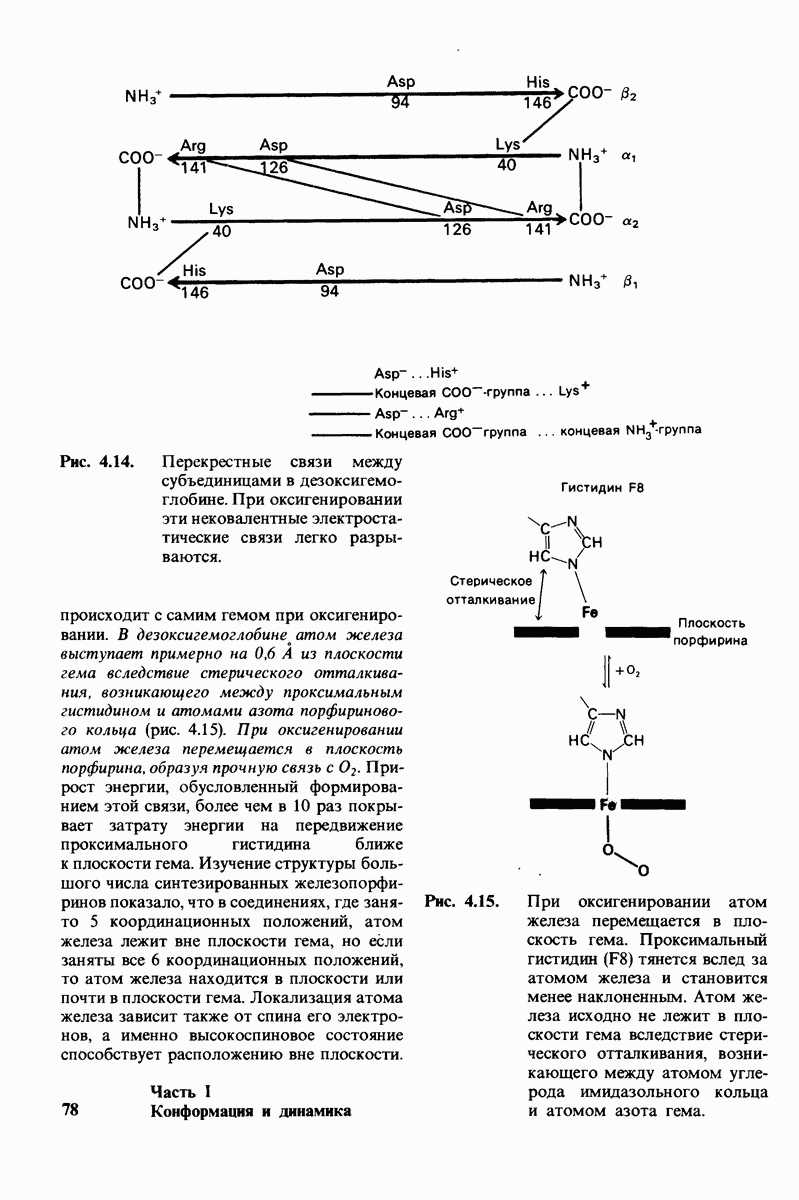
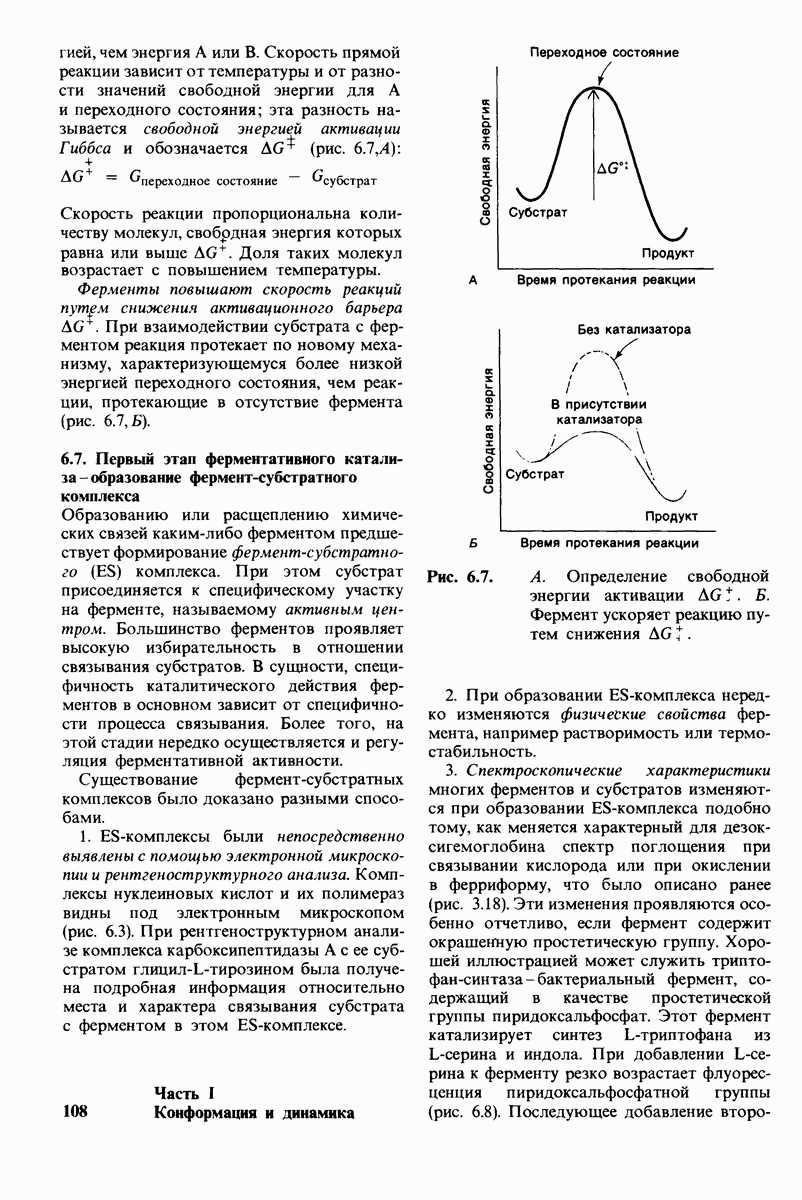
Рис. 6.7. А. Определение свободной энергии активации  Фермент ускоряет реакцию путем снижения
Фермент ускоряет реакцию путем снижения 
2. При образовании  -комплекса нередко изменяются физические свойства фермента, например растворимость или термостабильность.
-комплекса нередко изменяются физические свойства фермента, например растворимость или термостабильность.
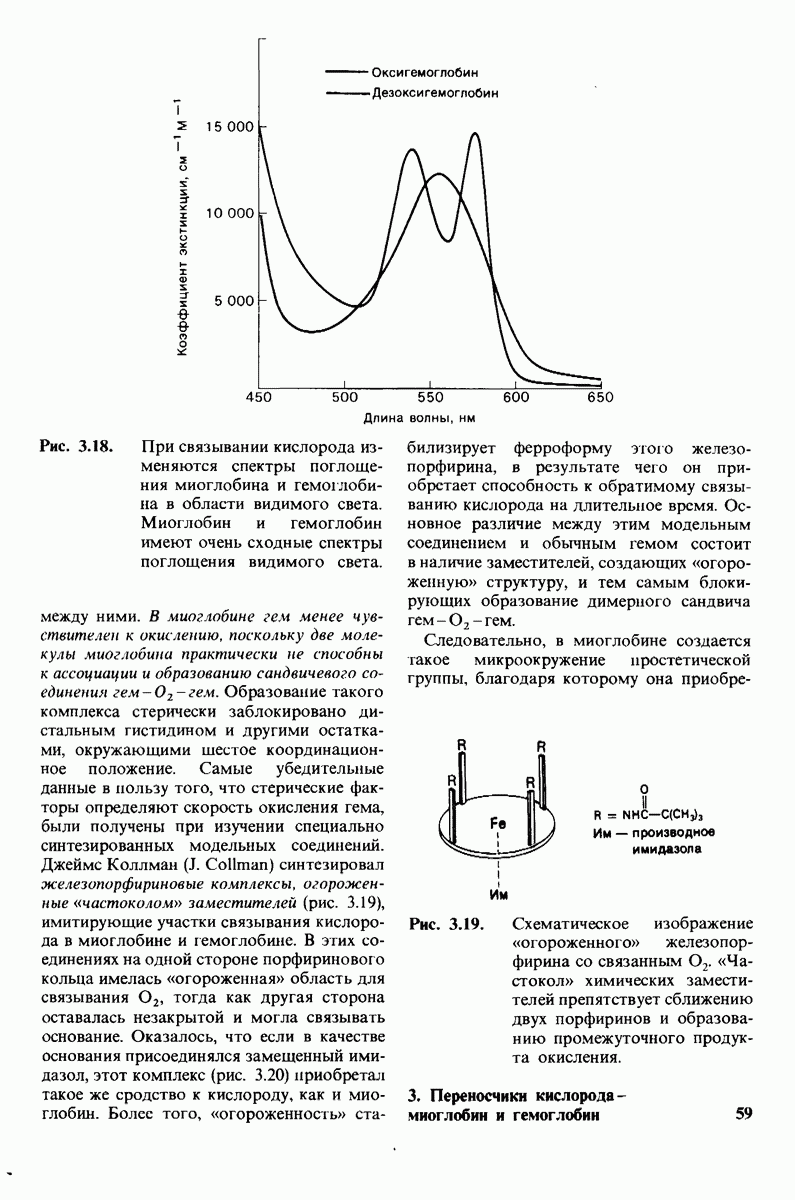
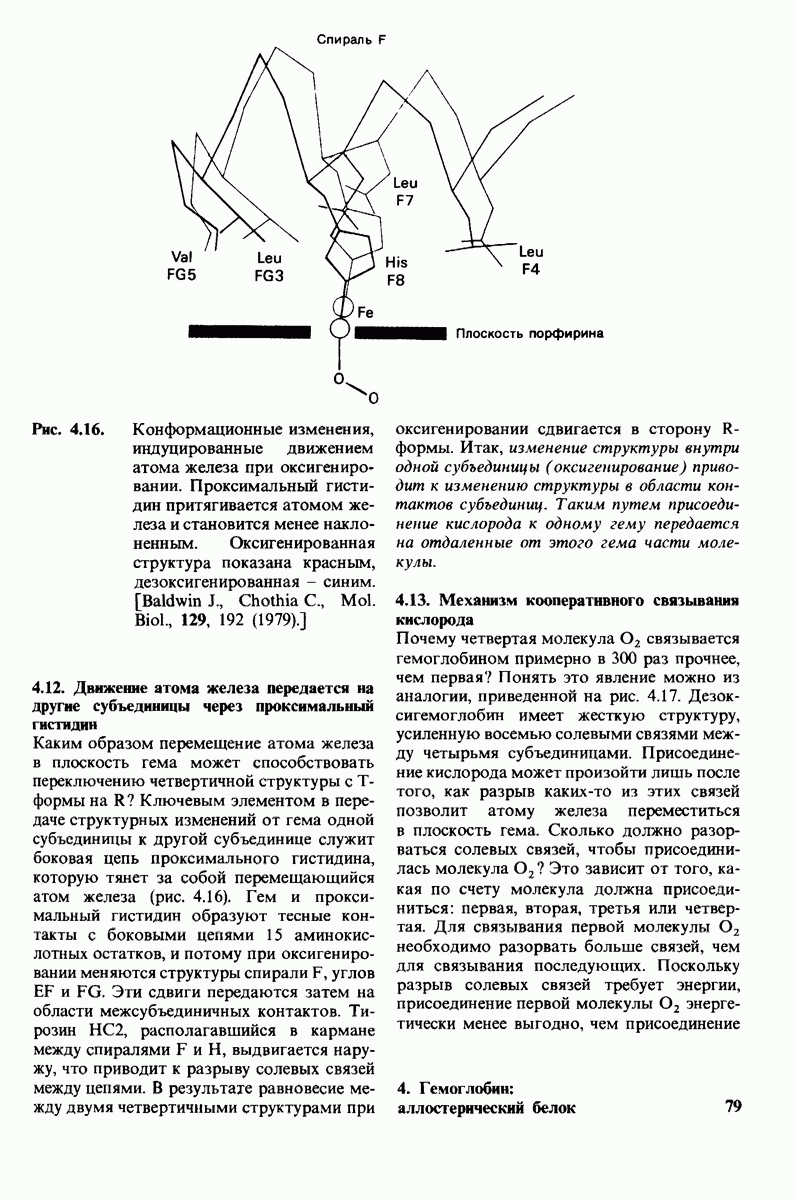
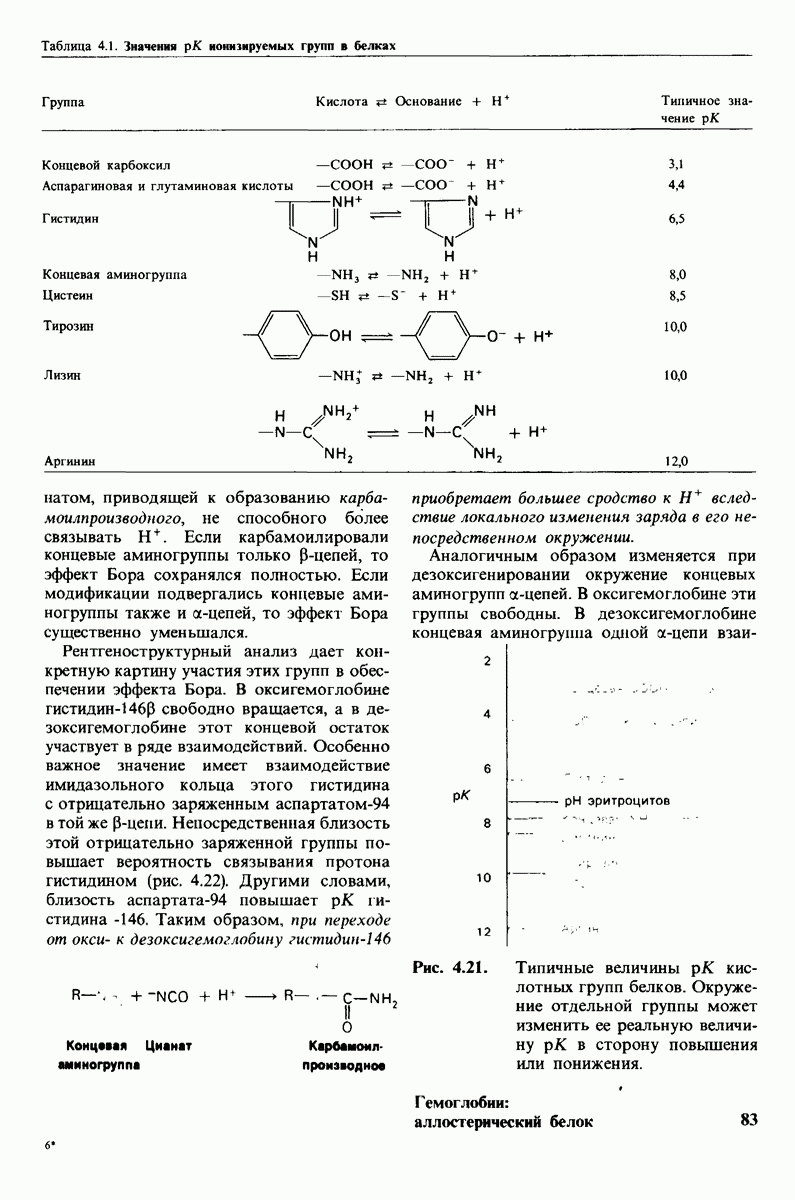
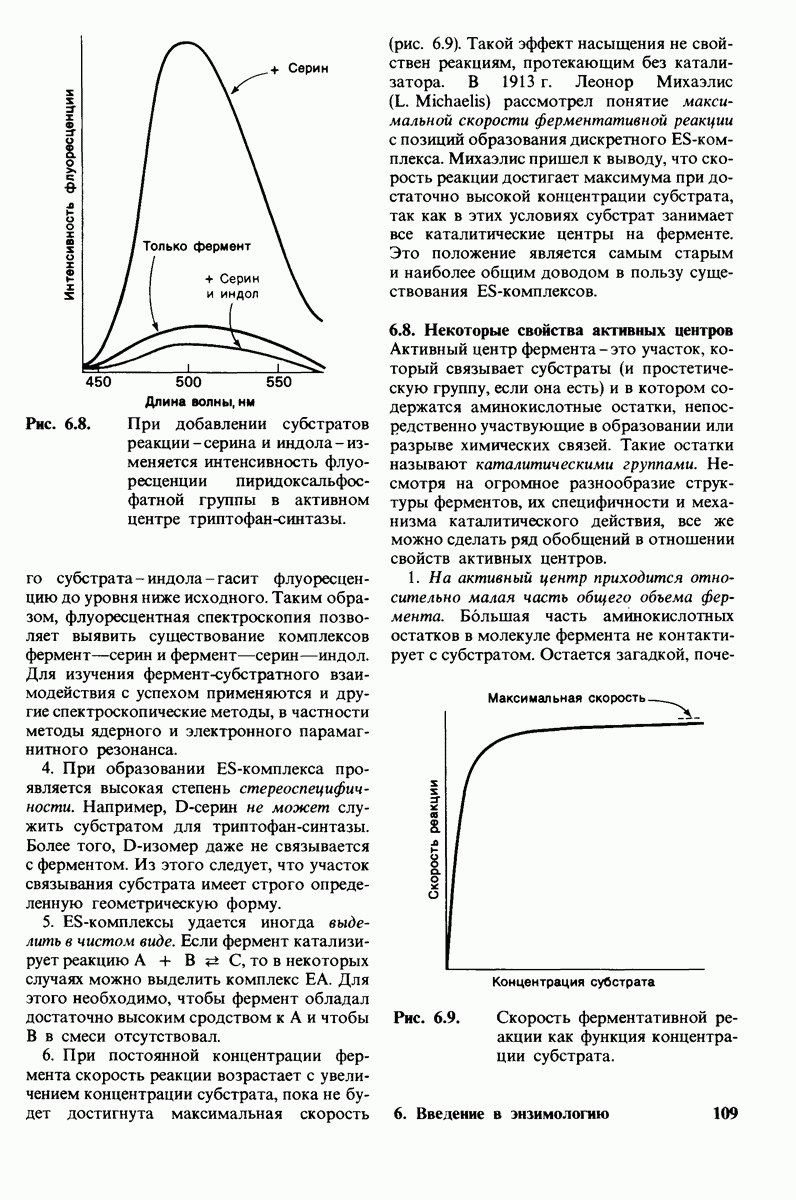
3. Спектроскопические характеристики многих ферментов и субстратов изменяются при образовании  -комплекса подобно тому, как меняется характерный для дезоксигемоглобина спектр поглощения при связывании кислорода или при окислении в ферриформу, что было описано ранее (рис. 3.18). Эти изменения проявляются особенно отчетливо, если фермент содержит окрашенную простетическую группу. Хорошей иллюстрацией может служить триптофан-синтаза-бактериальный фермент, содержащий в качестве простетической группы пиридоксальфосфат. Этот фермент катализирует синтез L-триптофана из L-серина и индола. При добавлении L-ceрина к ферменту резко возрастает флуоресценция пиридоксальфосфатной группы (рис. 6.8). Последующее добавление
-комплекса подобно тому, как меняется характерный для дезоксигемоглобина спектр поглощения при связывании кислорода или при окислении в ферриформу, что было описано ранее (рис. 3.18). Эти изменения проявляются особенно отчетливо, если фермент содержит окрашенную простетическую группу. Хорошей иллюстрацией может служить триптофан-синтаза-бактериальный фермент, содержащий в качестве простетической группы пиридоксальфосфат. Этот фермент катализирует синтез L-триптофана из L-серина и индола. При добавлении L-ceрина к ферменту резко возрастает флуоресценция пиридоксальфосфатной группы (рис. 6.8). Последующее добавление

Рис. 6.8. При добавлении субстратов реакции - серина и индола - изменяется интенсивность флуоресценции пиридоксальфосфатной группы в активном центре триптофан-синтазы.
второго субстрата - индола - гасит флуоресценцию до уровня ниже исходного. Таким образом, флуоресцентная спектроскопия позволяет выявить существование комплексов фермент—серин и фермент—серин—индол. Для изучения фермент-субстратного взаимодействия с успехом применяются и другие спектроскопические методы, в частности методы ядерного и электронного парамагнитного резонанса.
4. При образовании  -комплекса проявляется высокая степень стереоспецифичности. Например, D-серин не может служить субстратом для триптофан-синтазы. Более того, D-изомер даже не связывается с ферментом. Из этого следует, что участок связывания субстрата имеет строго определенную геометрическую форму.
-комплекса проявляется высокая степень стереоспецифичности. Например, D-серин не может служить субстратом для триптофан-синтазы. Более того, D-изомер даже не связывается с ферментом. Из этого следует, что участок связывания субстрата имеет строго определенную геометрическую форму.
 -комплексы удается иногда выделить в чистом виде. Если фермент катализирует реакцию
-комплексы удается иногда выделить в чистом виде. Если фермент катализирует реакцию  , то в некоторых случаях можно выделить комплекс
, то в некоторых случаях можно выделить комплекс  Для этого необходимо, чтобы фермент обладал достаточно высоким сродством к А и чтобы В в смеси отсутствовал.
Для этого необходимо, чтобы фермент обладал достаточно высоким сродством к А и чтобы В в смеси отсутствовал.
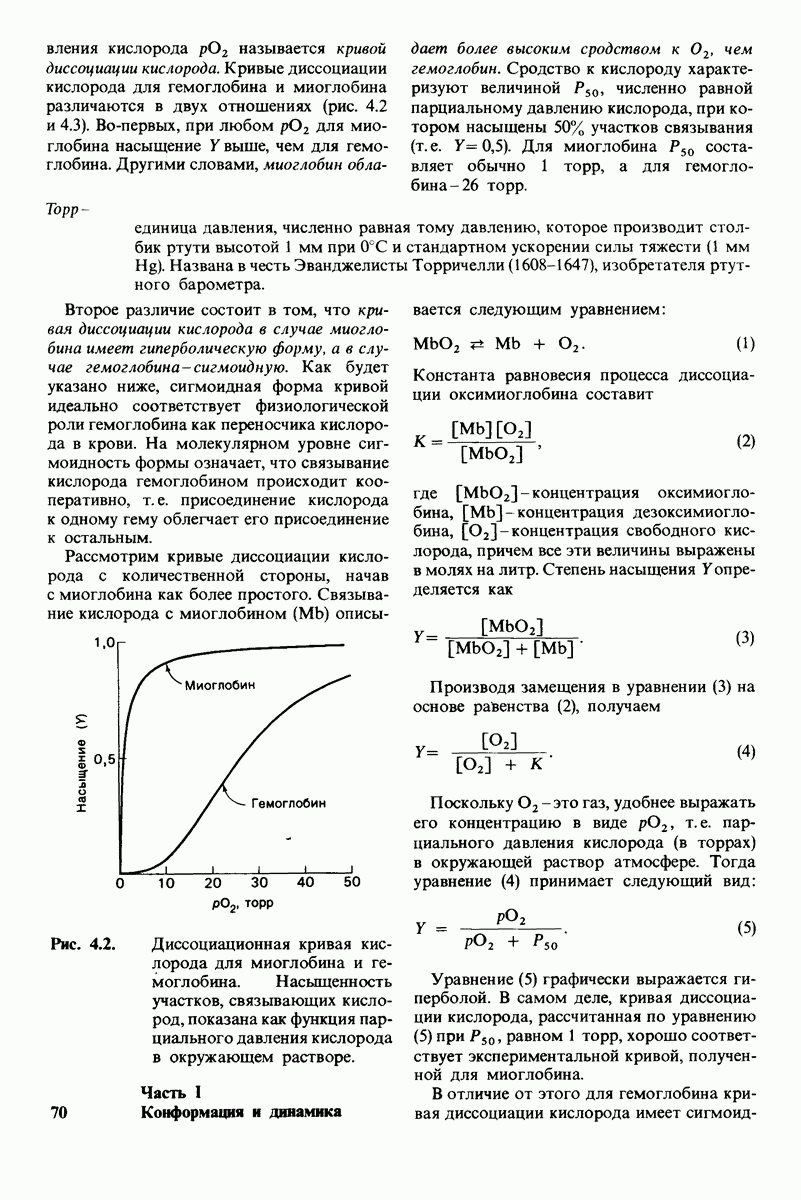
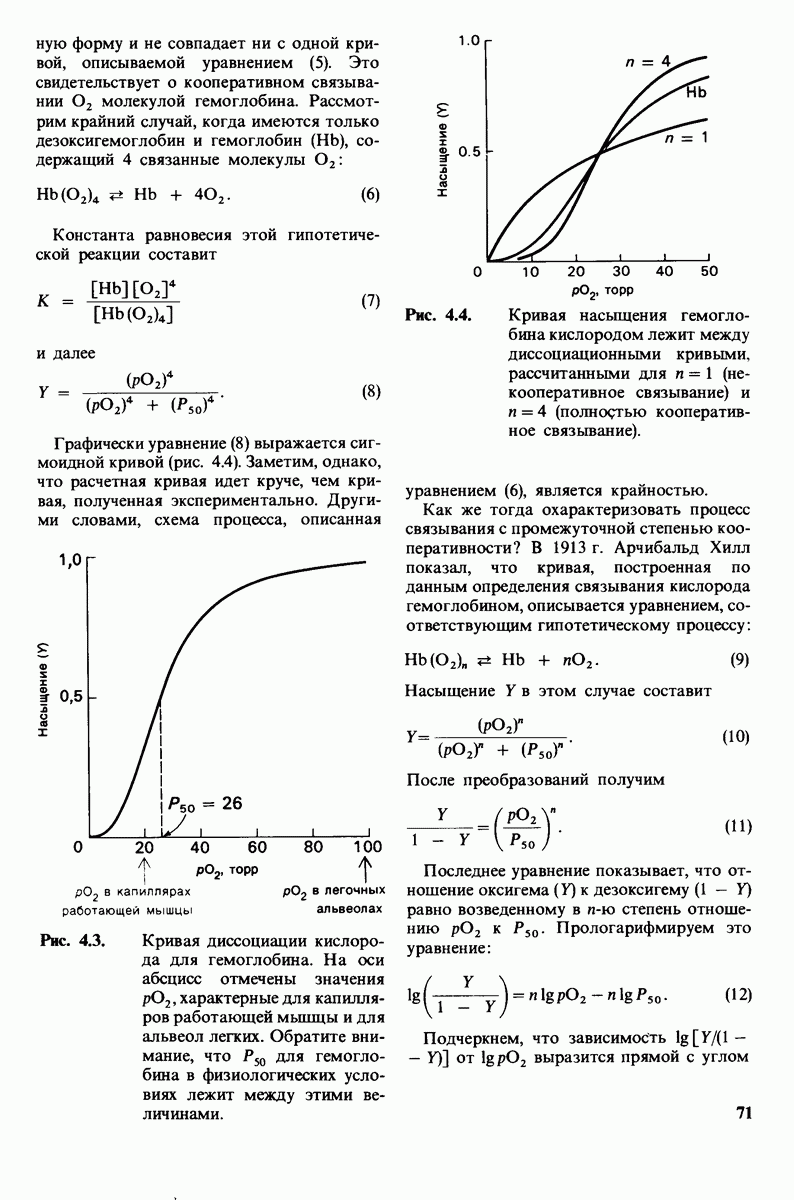
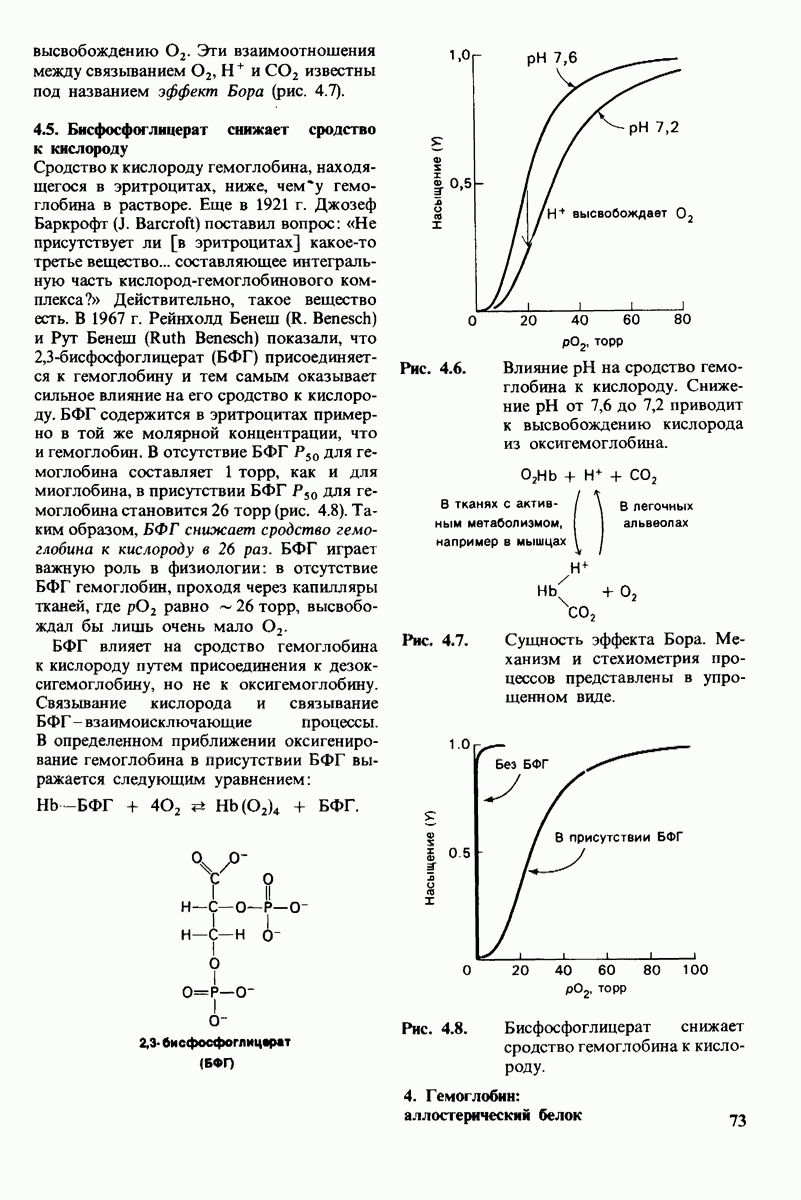
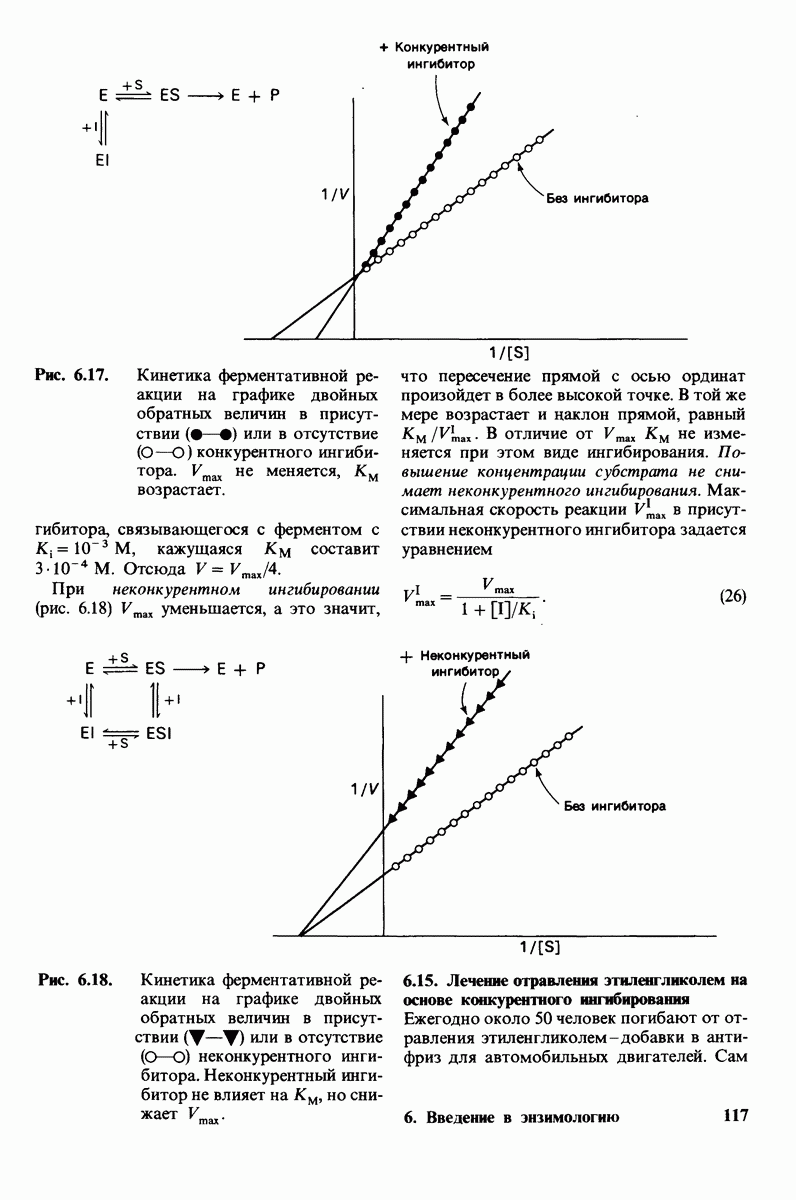
6. При постоянной концентрации фермента скорость реакции возрастает с увеличением концентрации субстрата, пока не будет достигнута максимальная скорость (рис. 6.9). Такой эффект насыщения не свойствен реакциям, протекающим без катализатора. В 1913 г. Леонор Михаэлис (L. Michaelis) рассмотрел понятие максимальной скорости ферментативной реакции с позиций образования дискретного  -комплекса. Михаэлис пришел к выводу, что скорость реакции достигает максимума при достаточно высокой концентрации субстрата, так как в этих условиях субстрат занимает все каталитические центры на ферменте. Это положение является самым старым и наиболее общим доводом в пользу существования
-комплекса. Михаэлис пришел к выводу, что скорость реакции достигает максимума при достаточно высокой концентрации субстрата, так как в этих условиях субстрат занимает все каталитические центры на ферменте. Это положение является самым старым и наиболее общим доводом в пользу существования  -комплексов.
-комплексов.