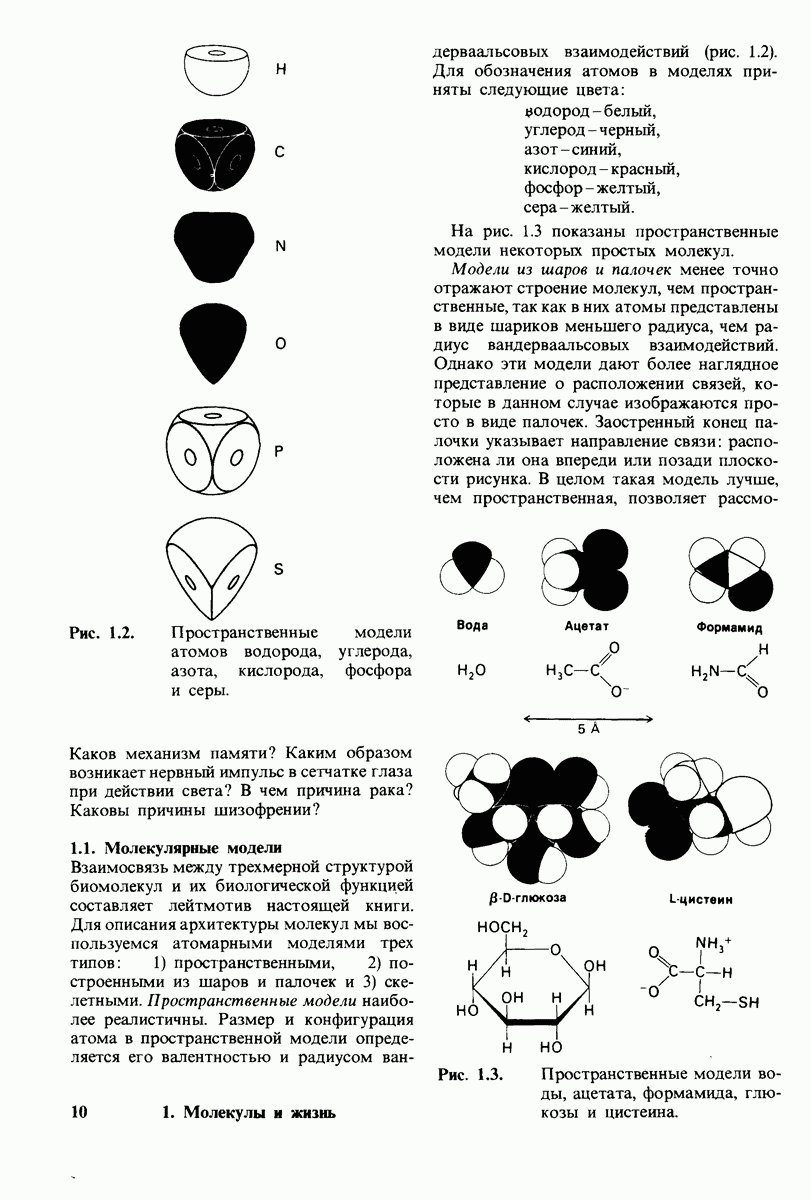
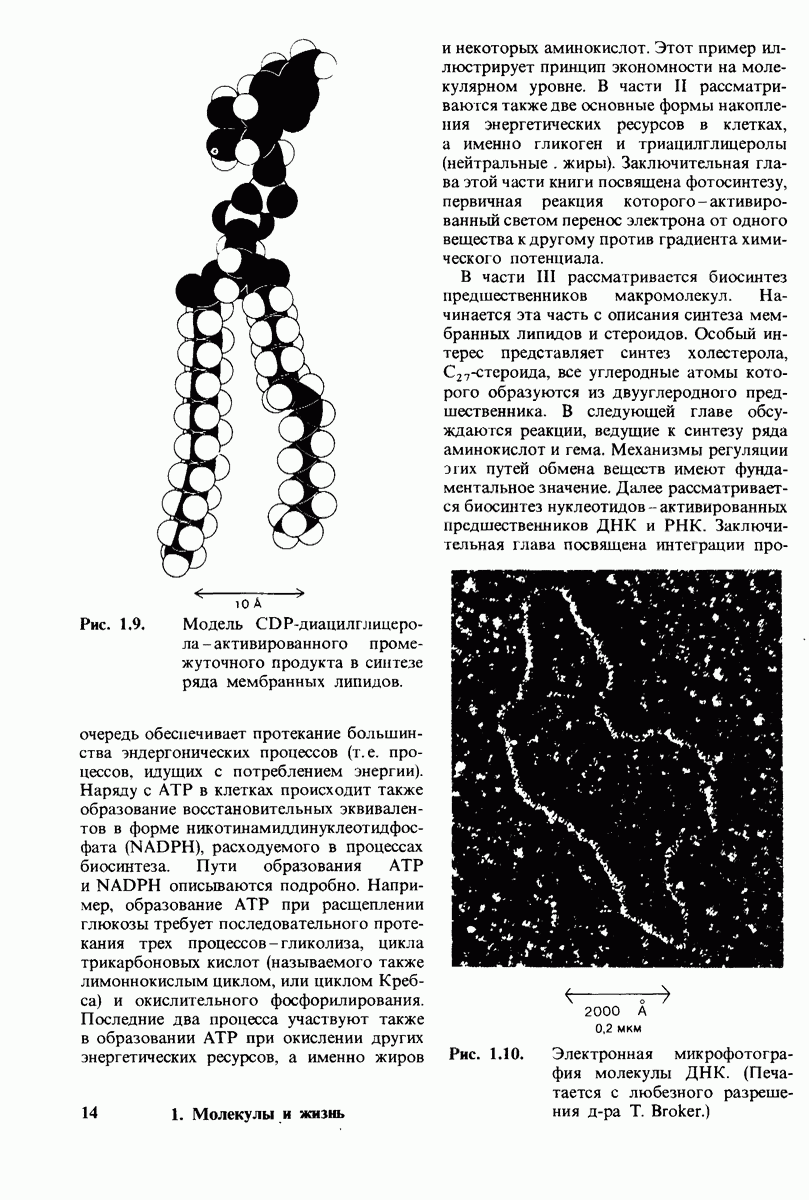
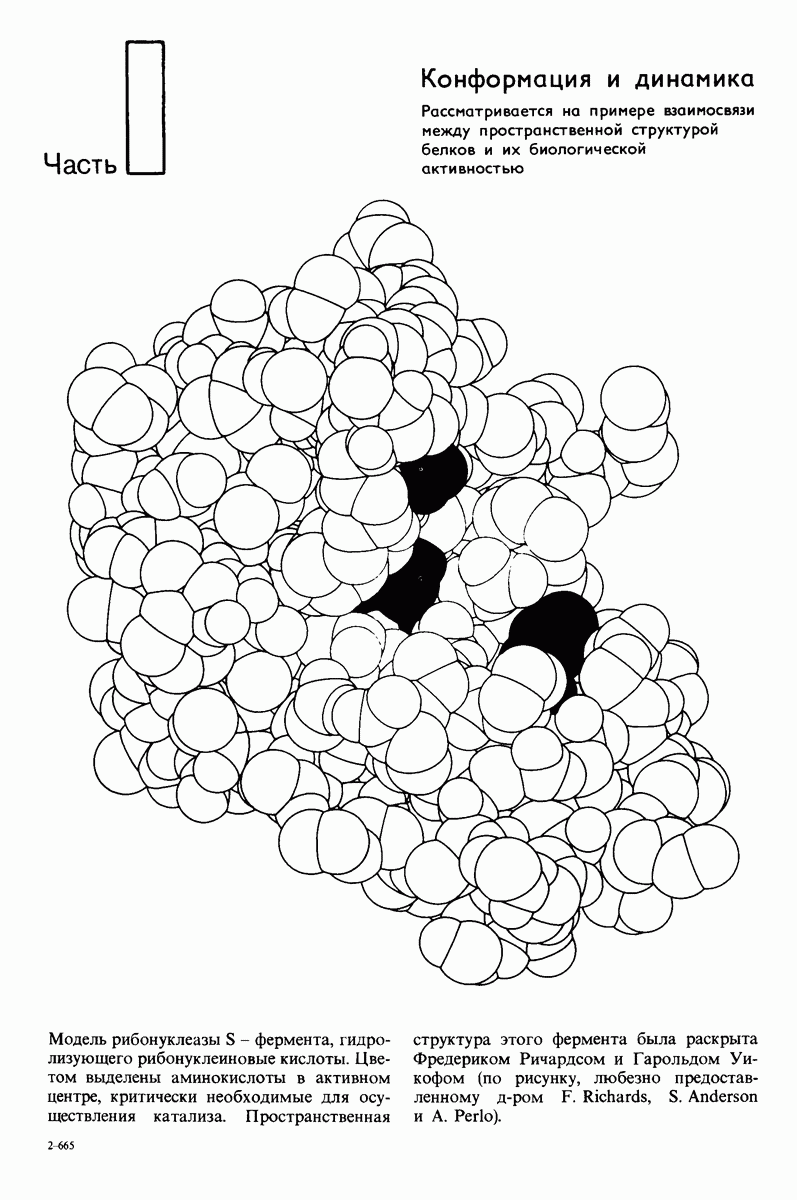
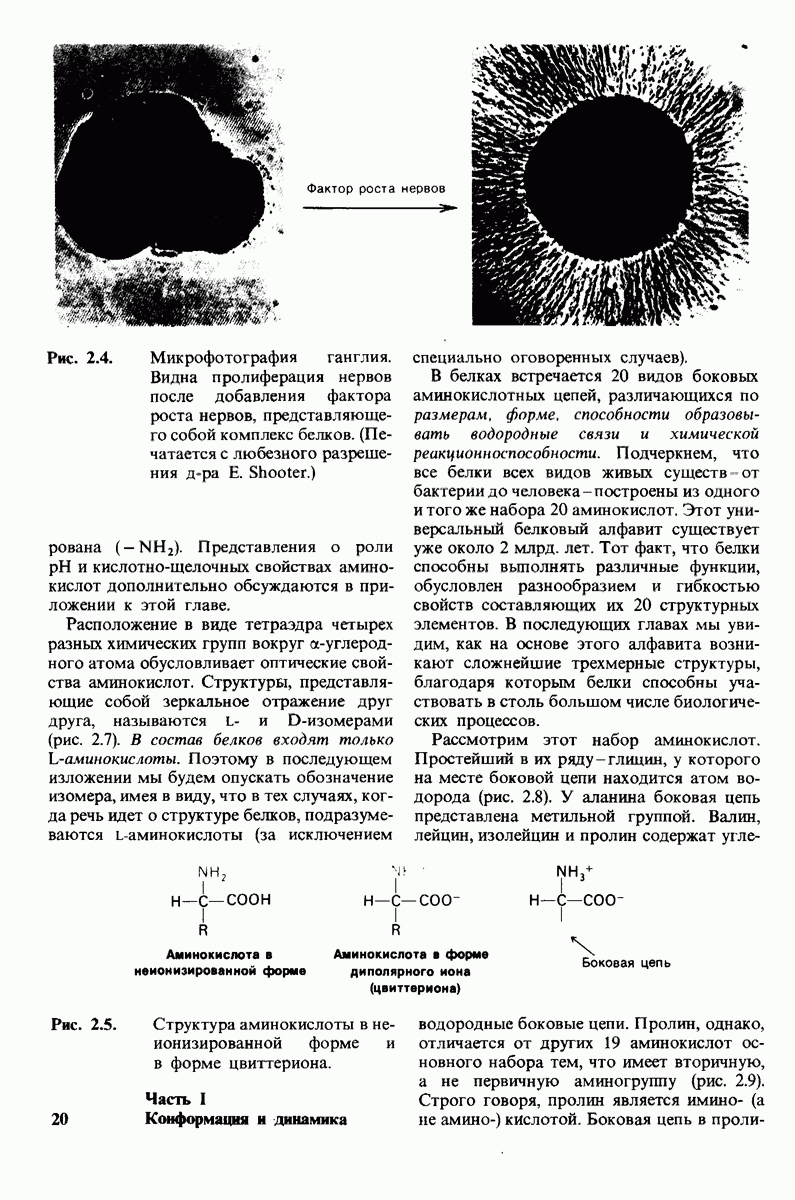
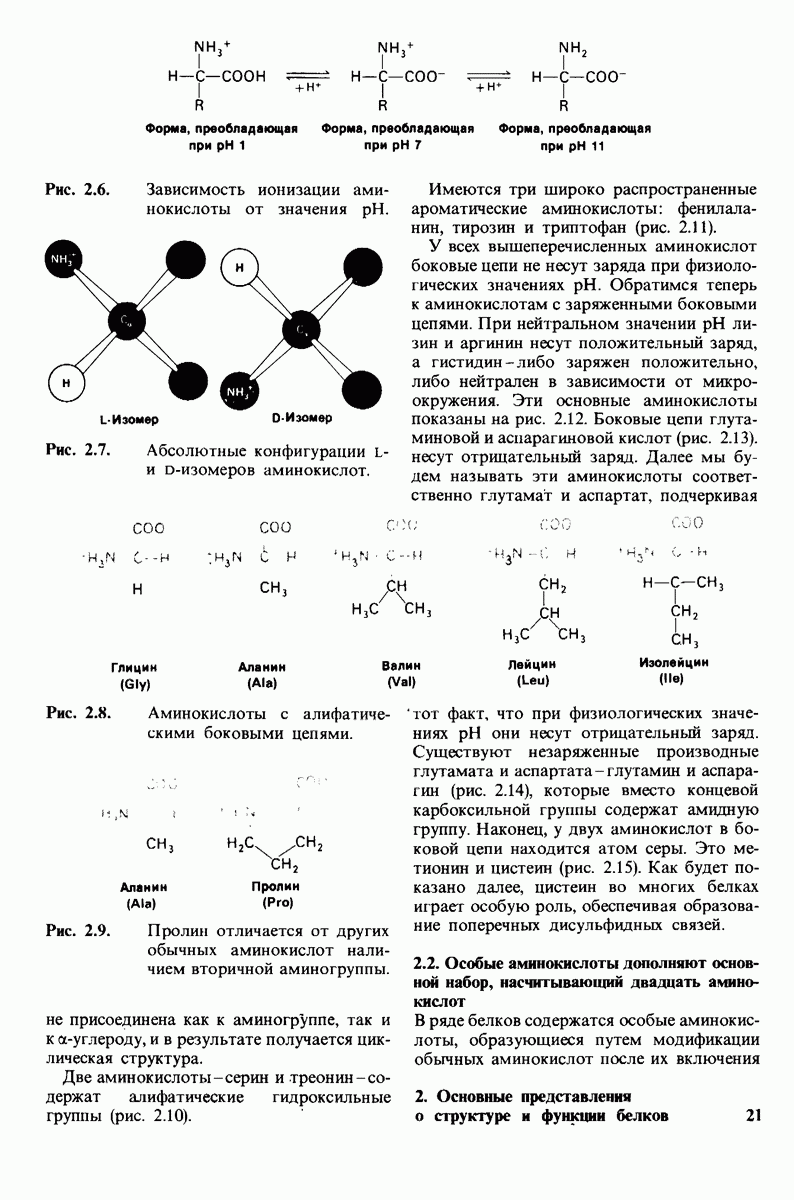
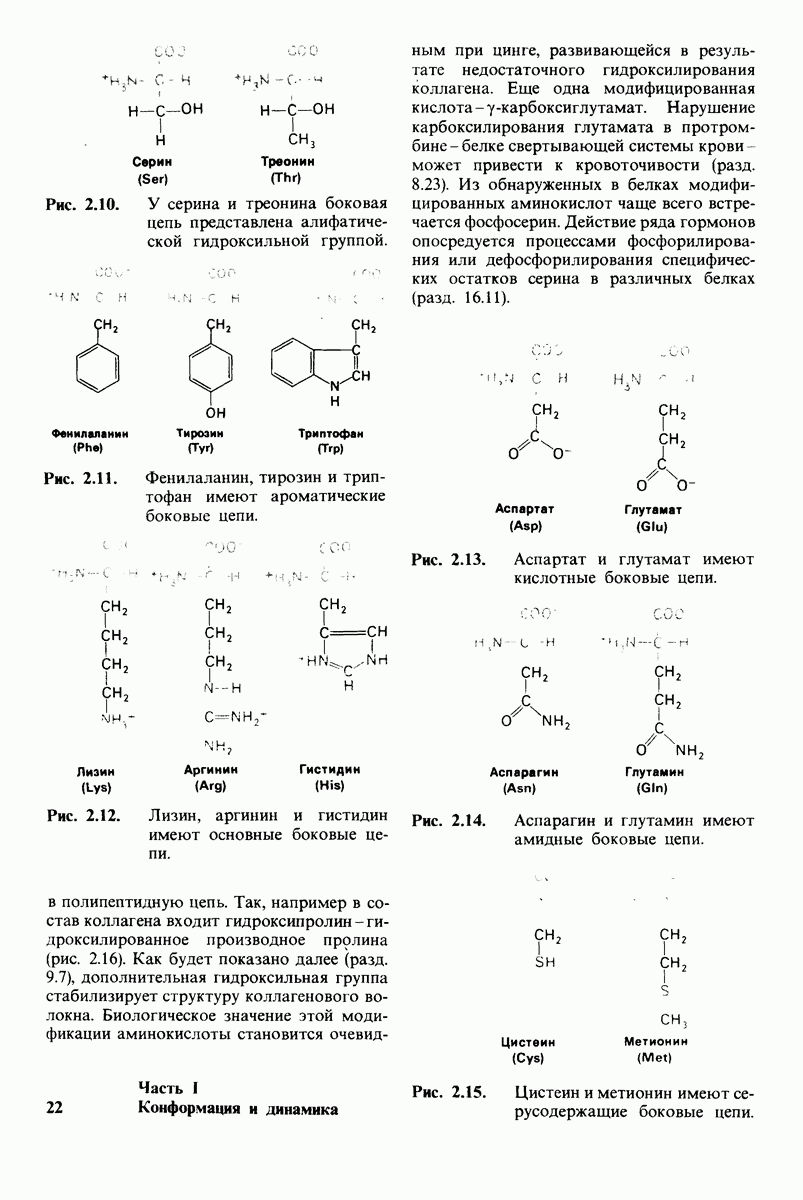
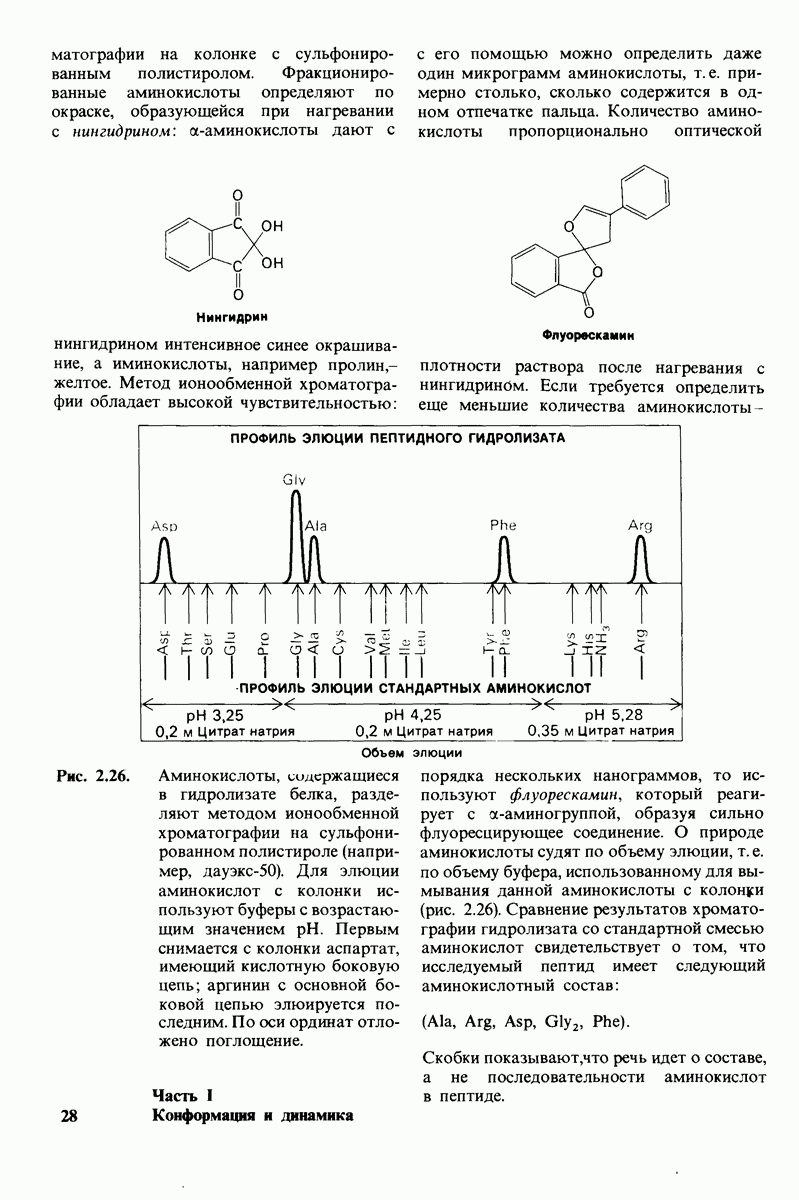
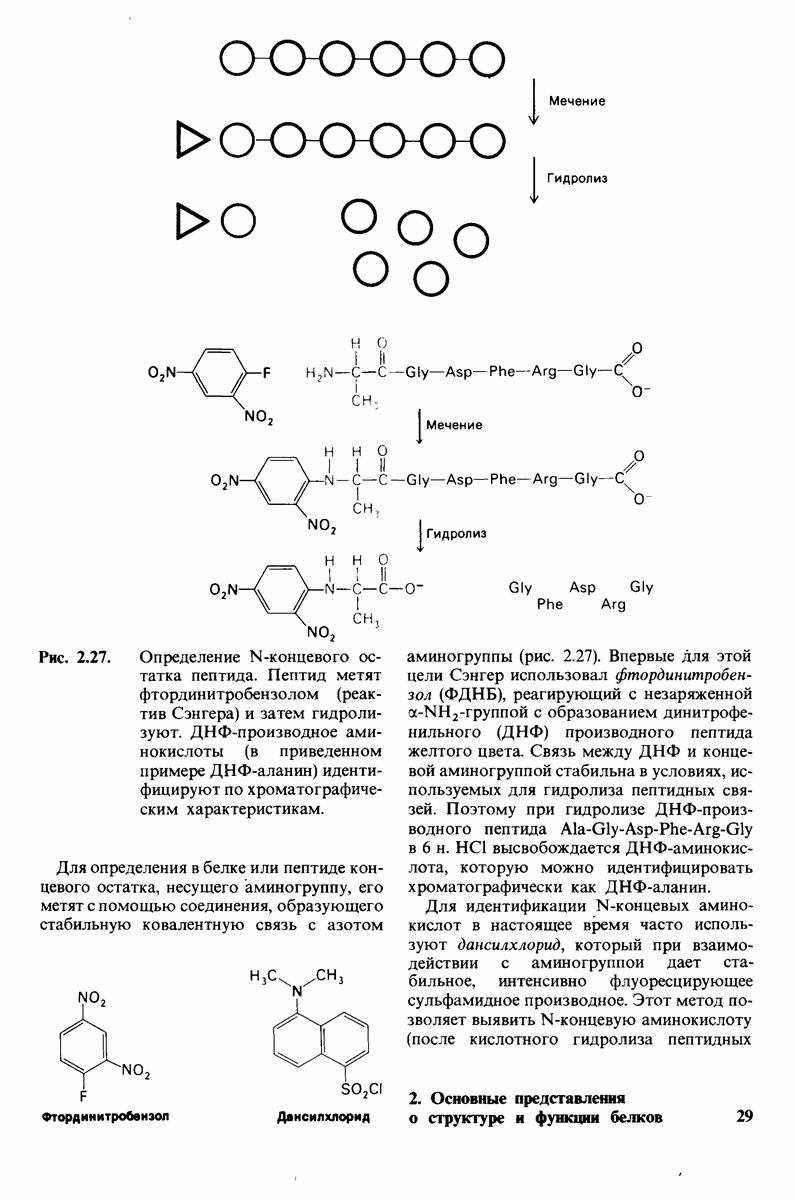
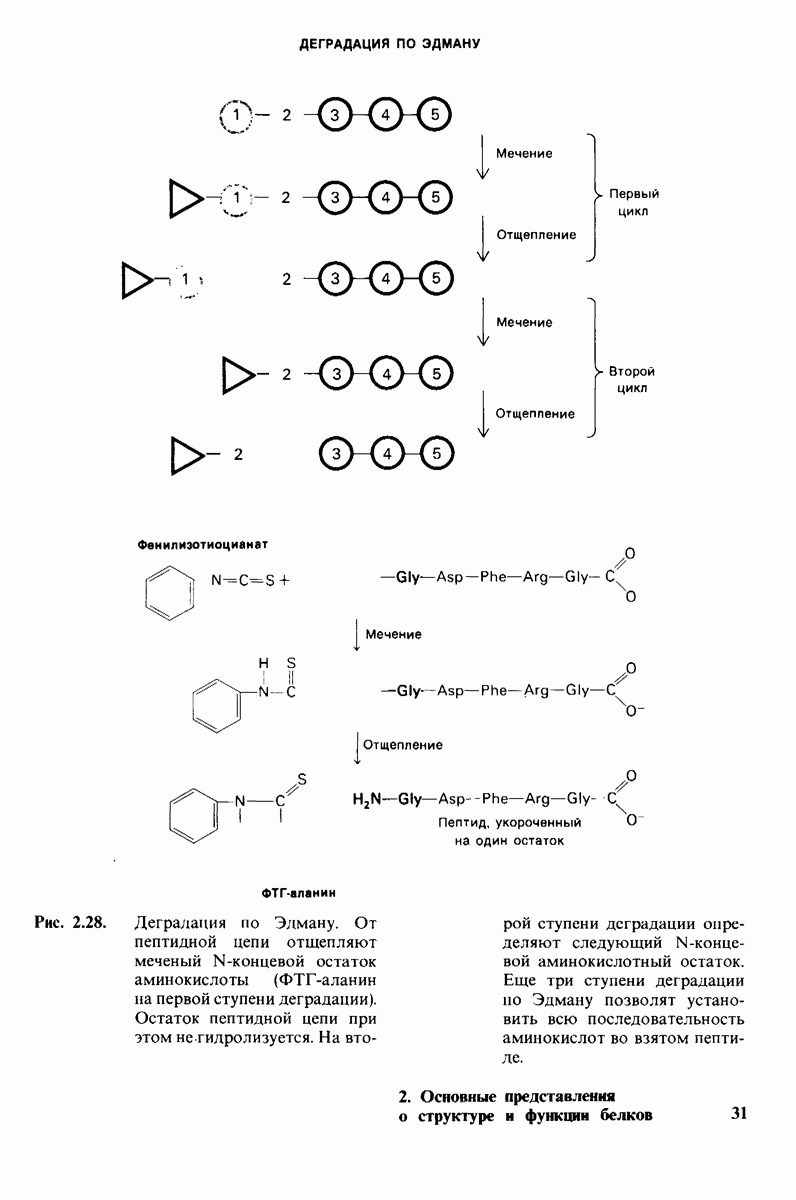
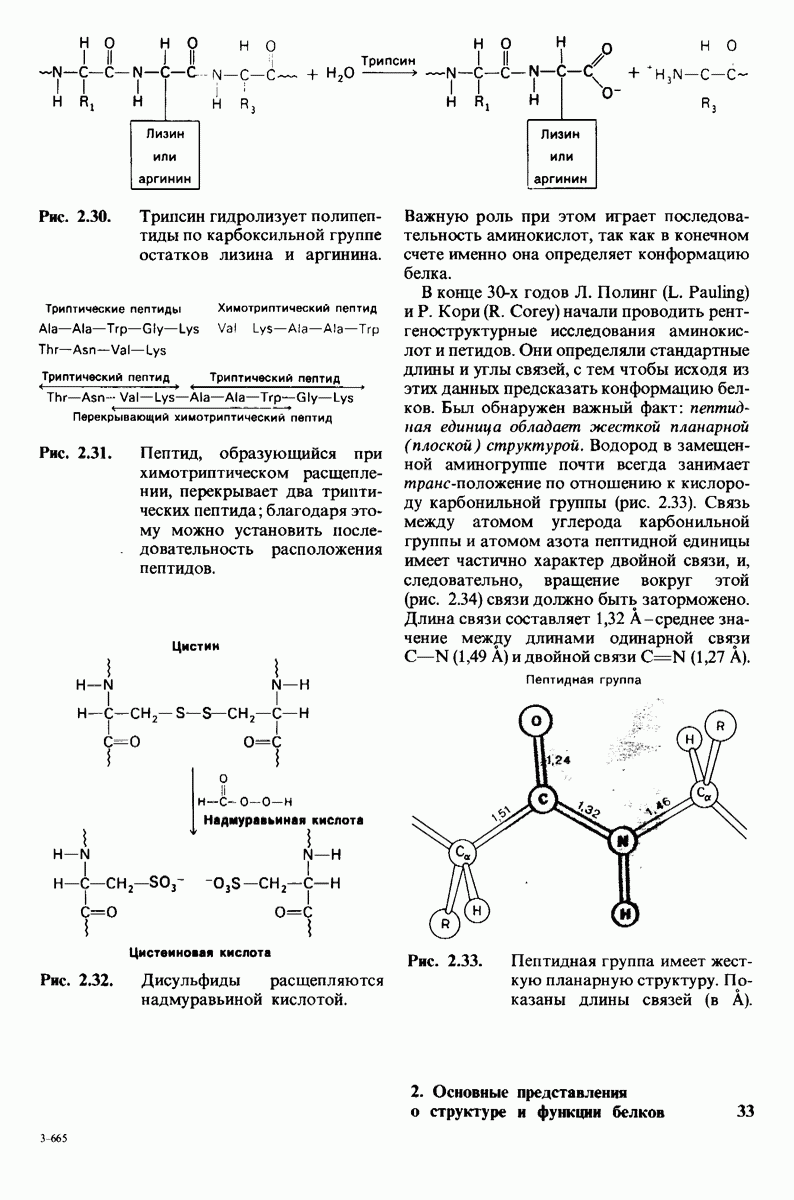
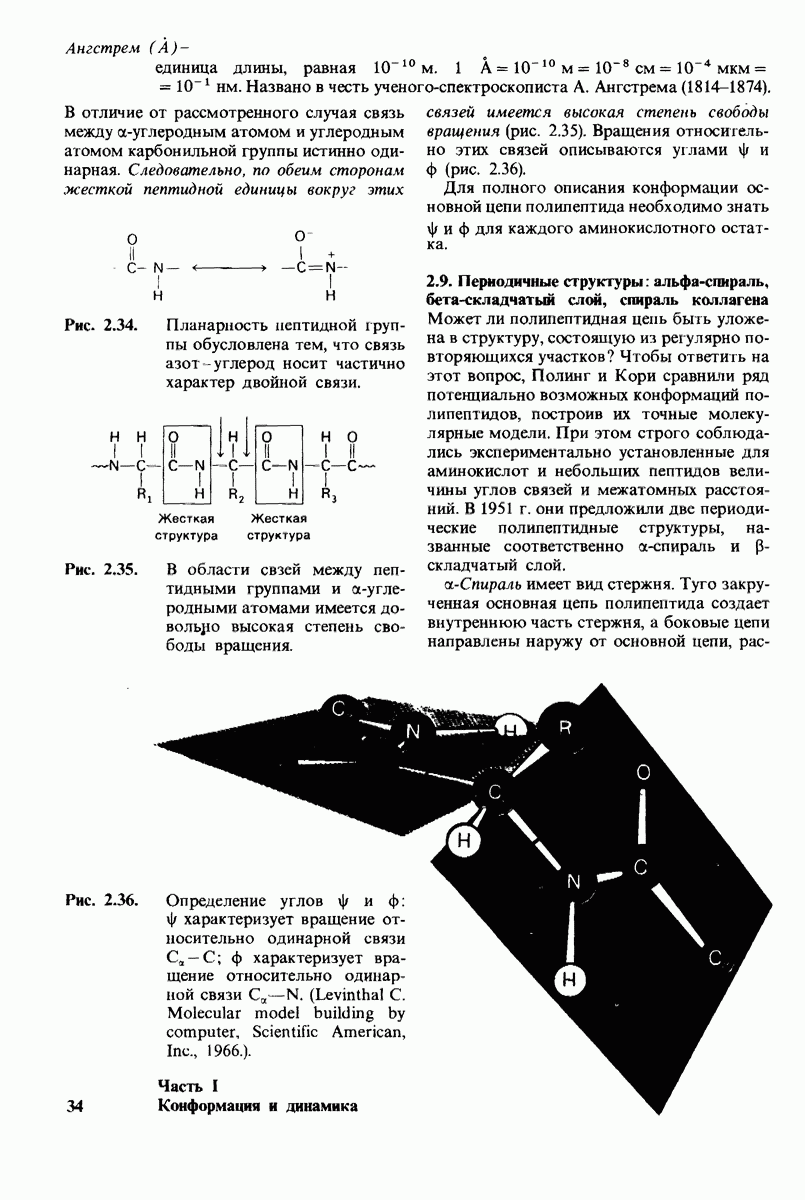
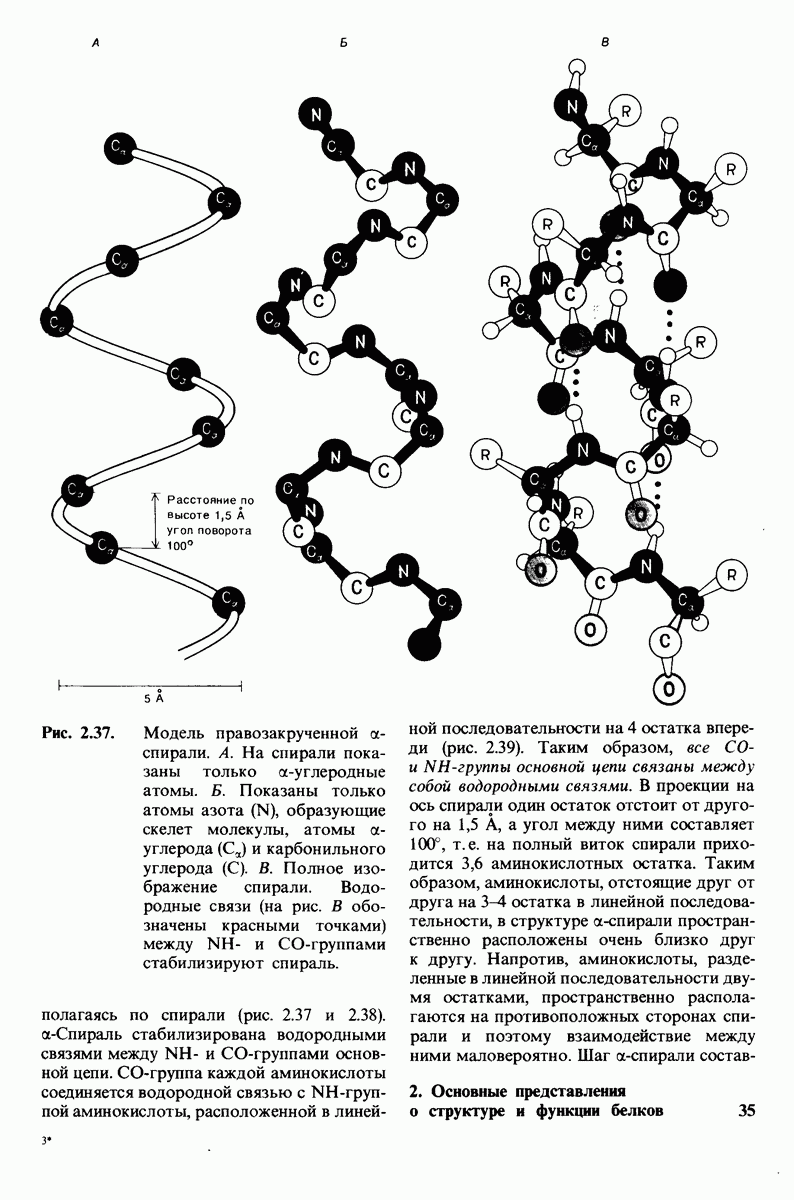
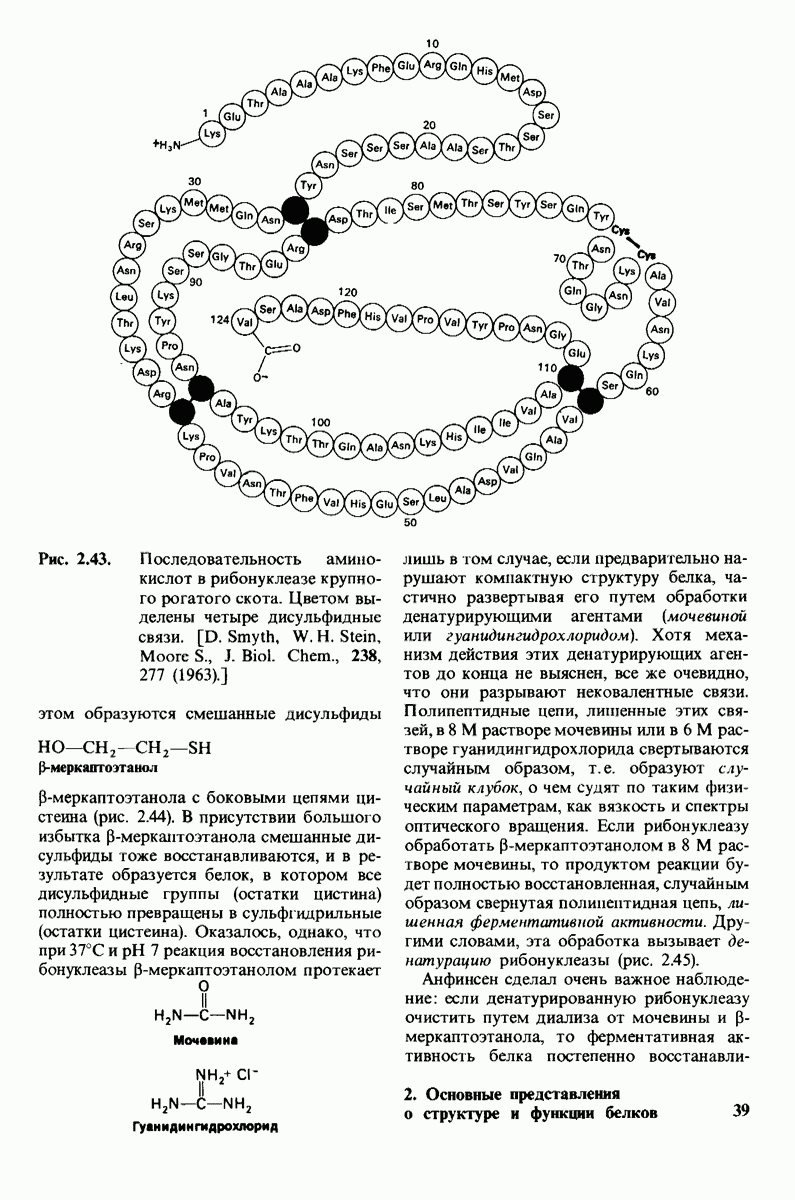
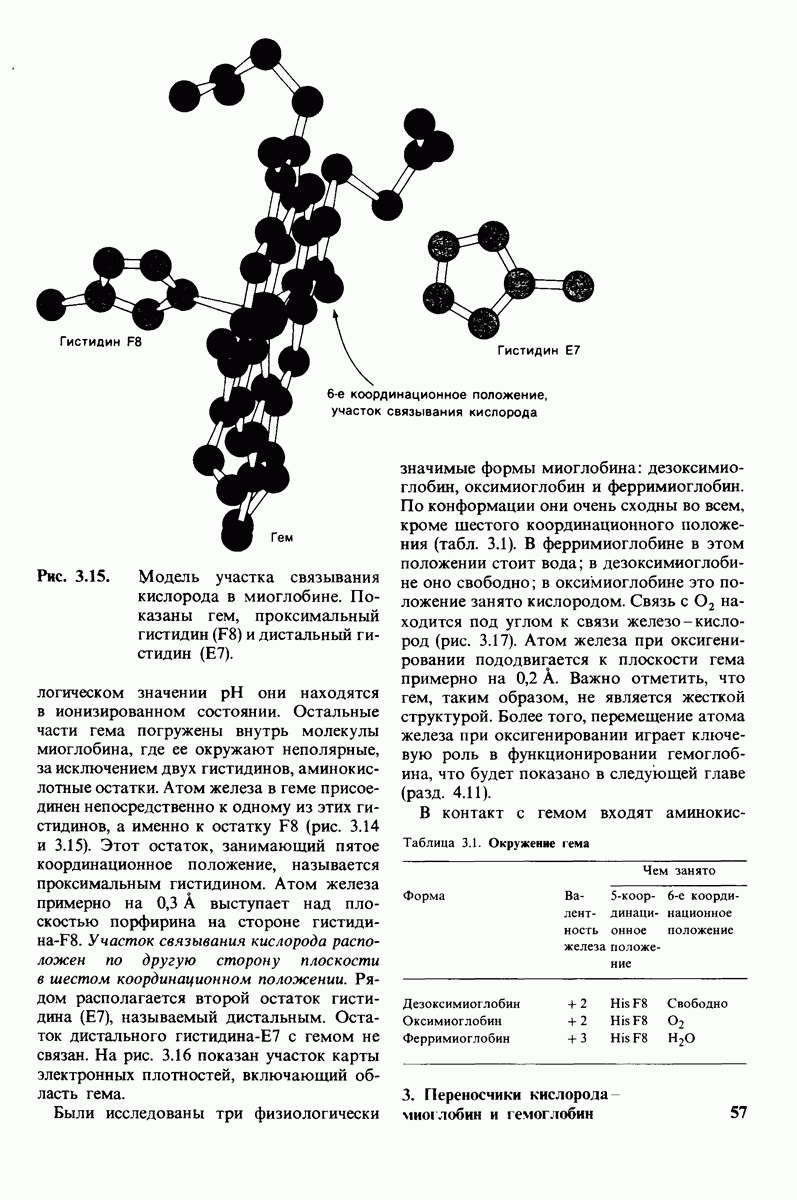
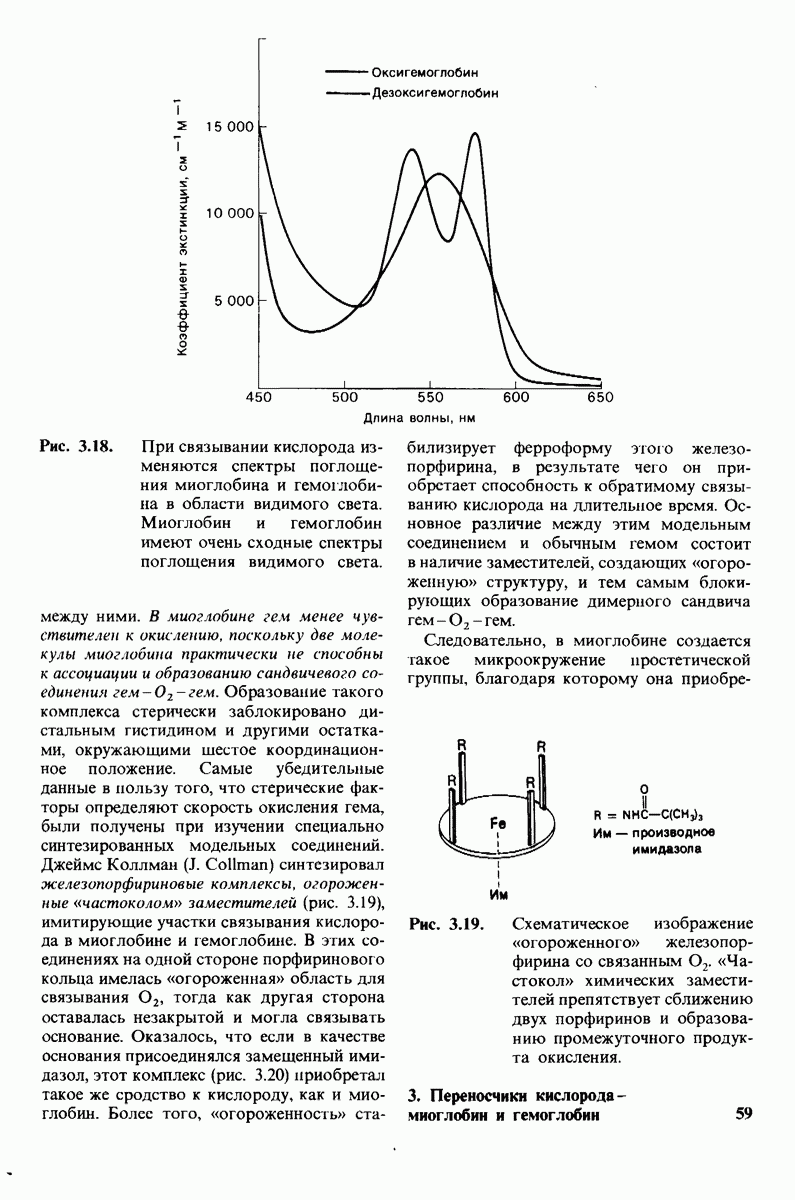
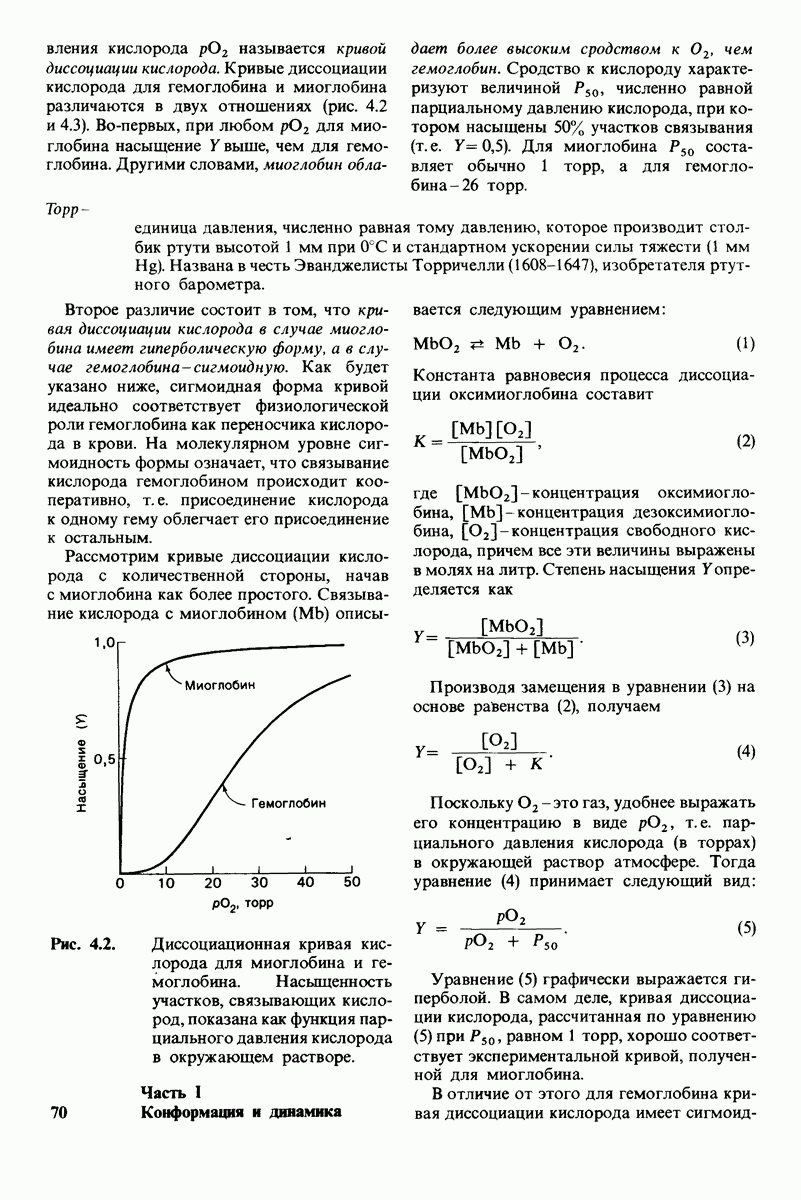
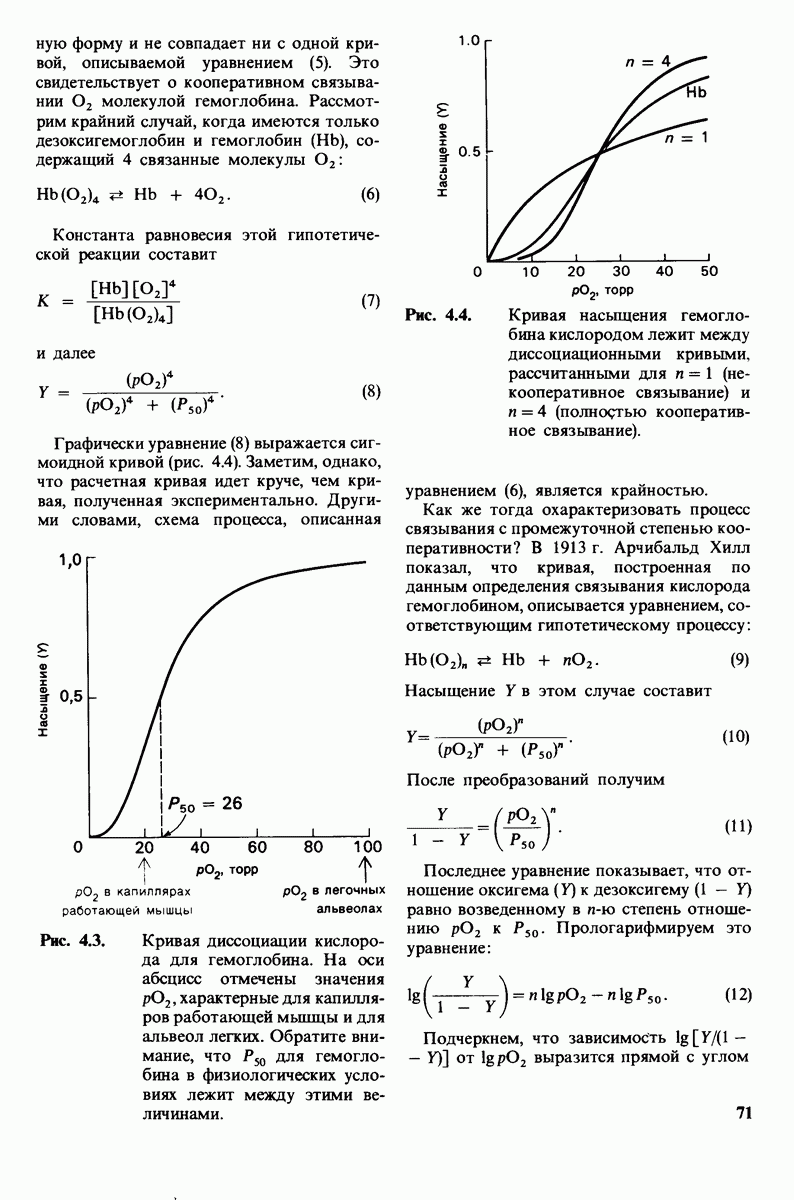
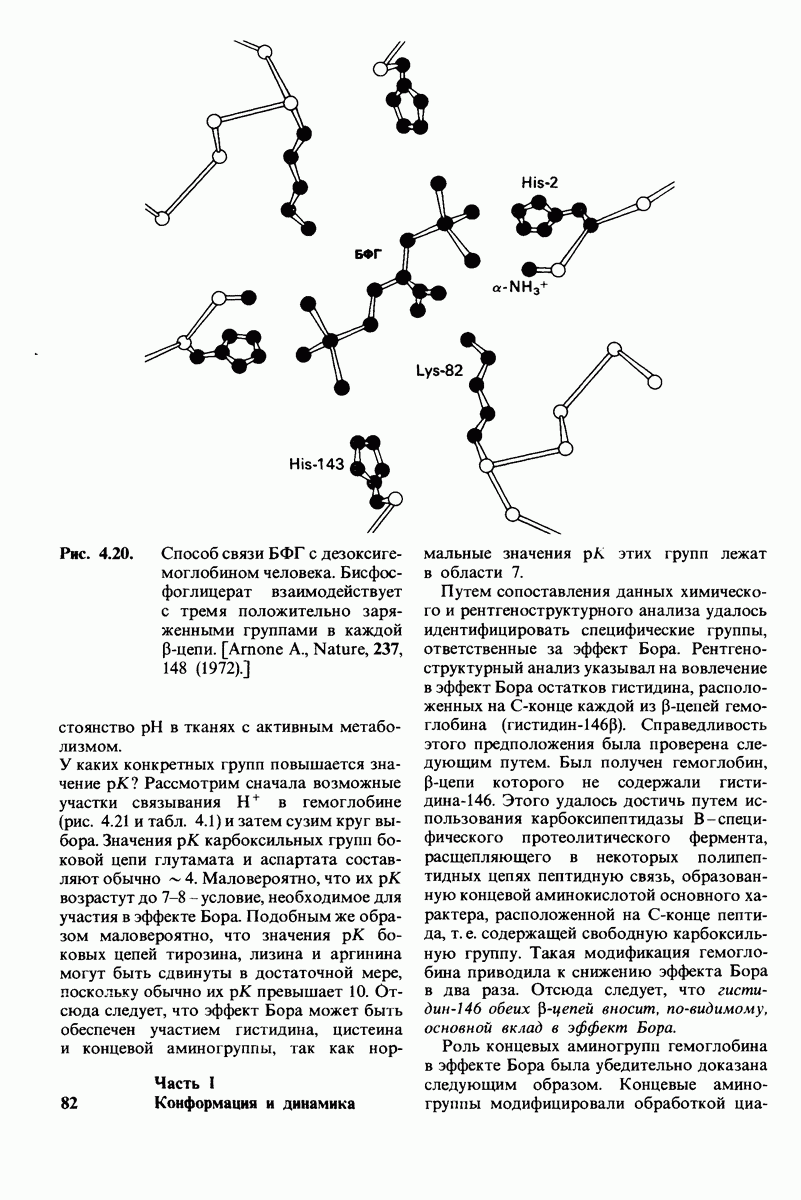
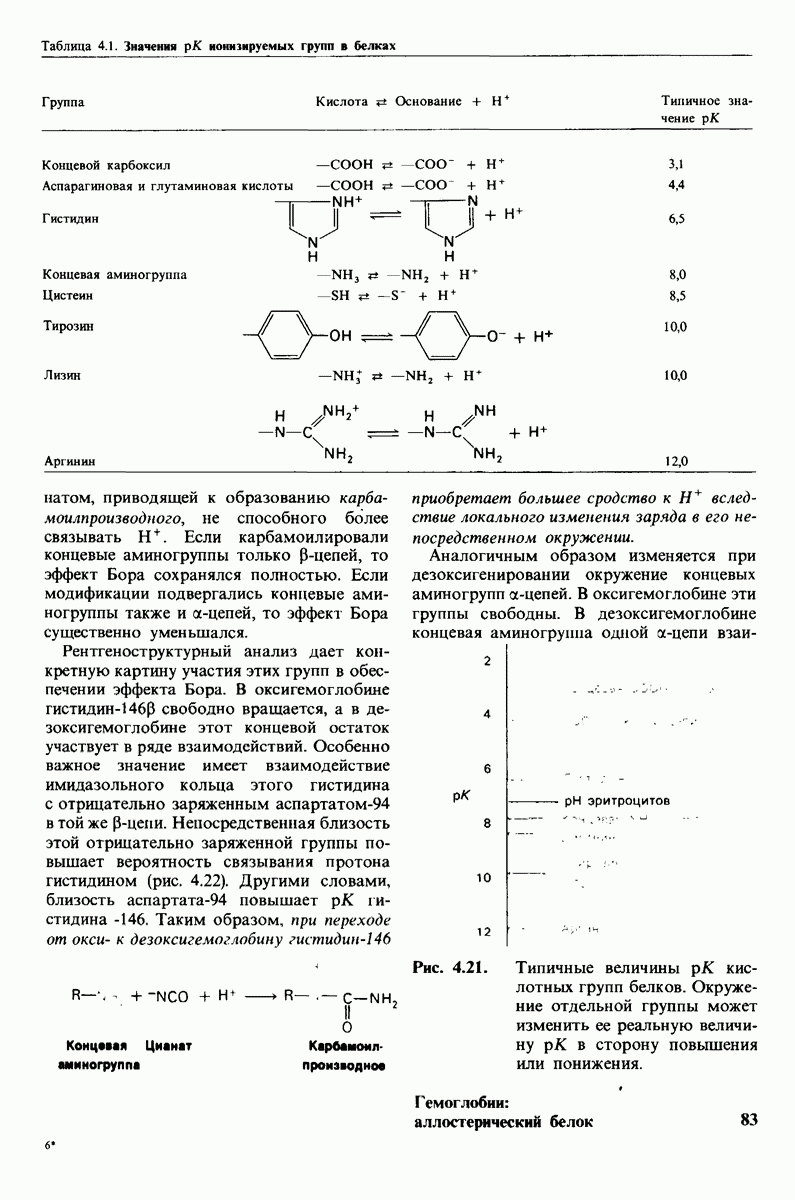
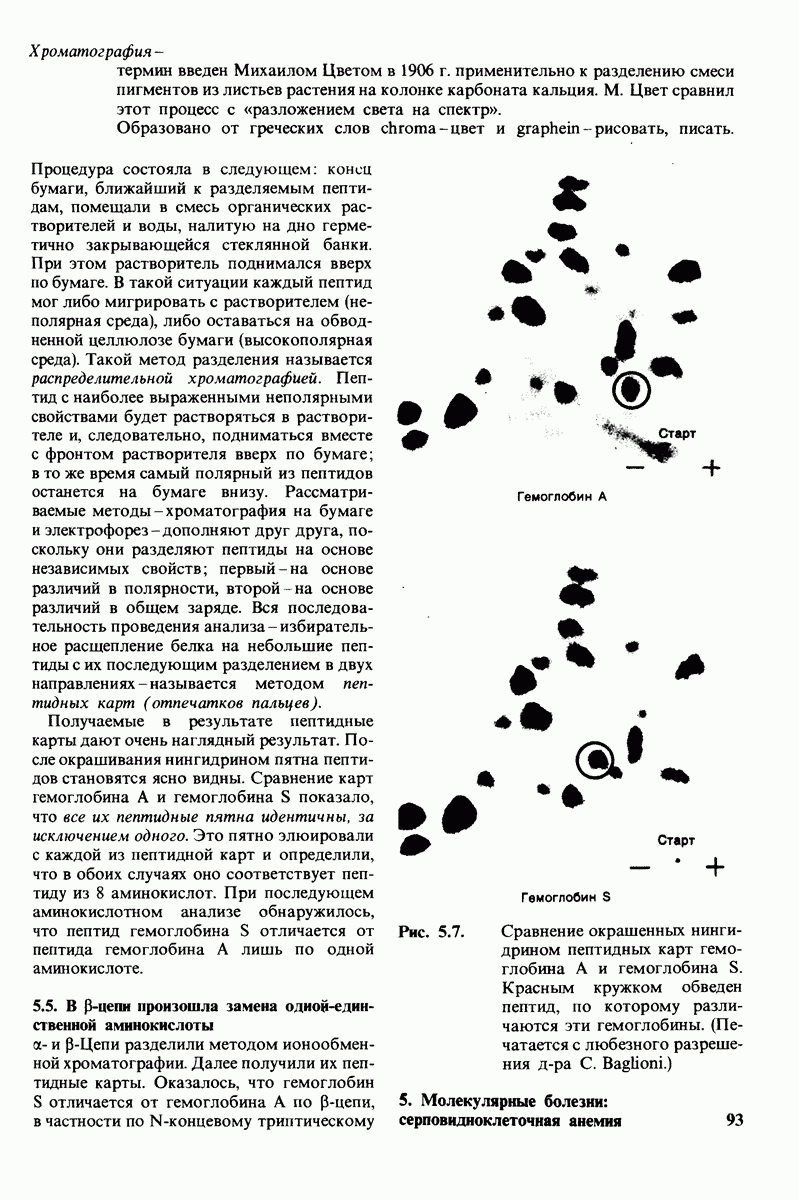
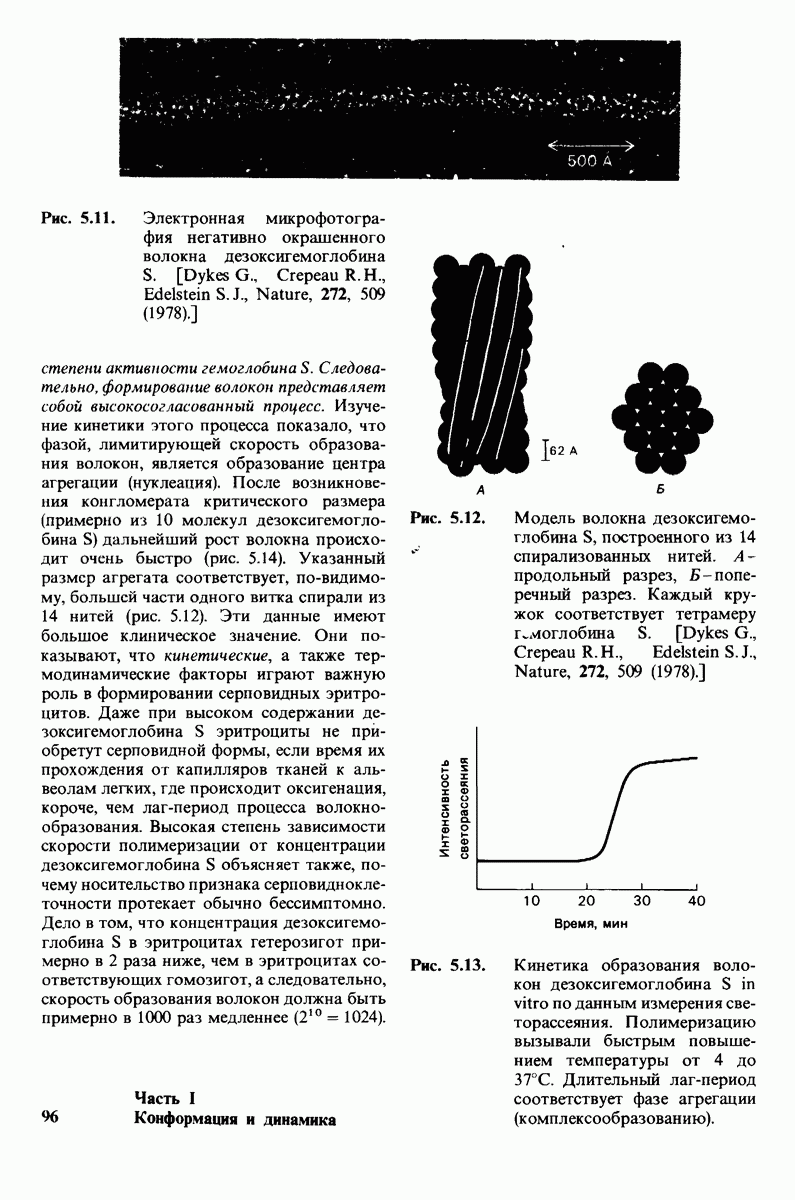
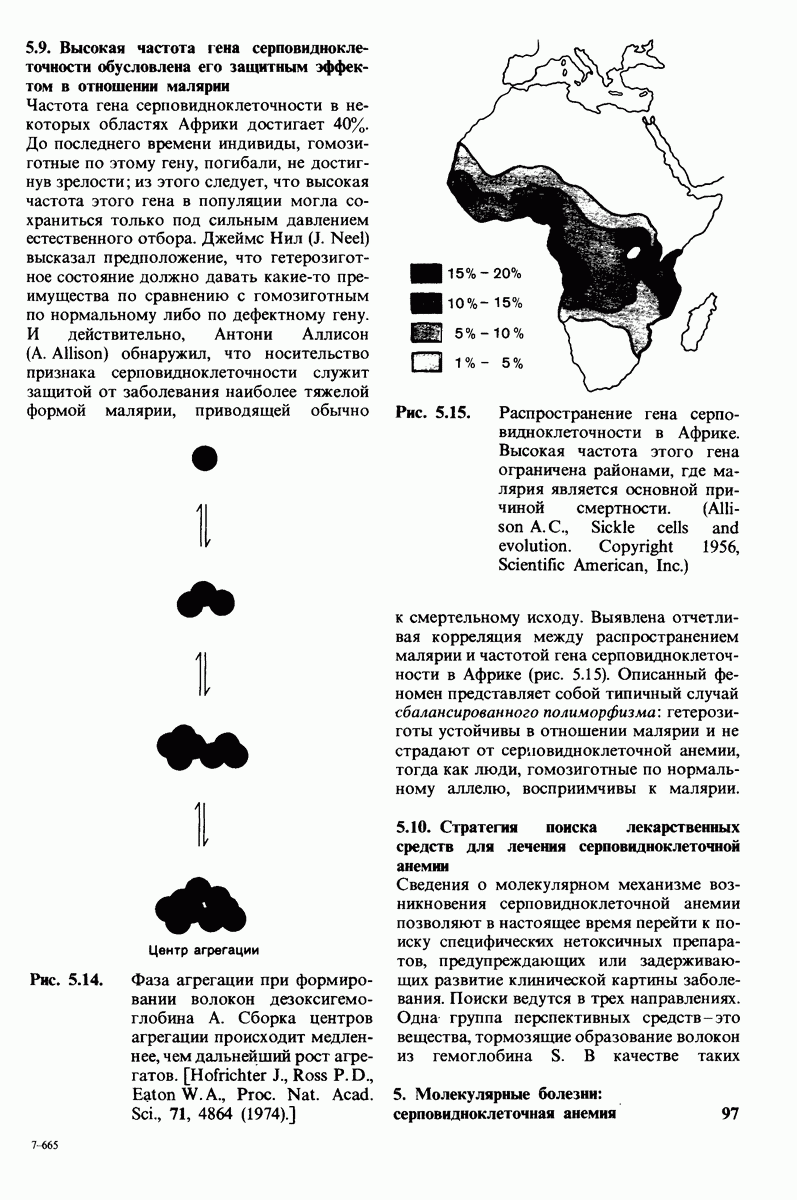
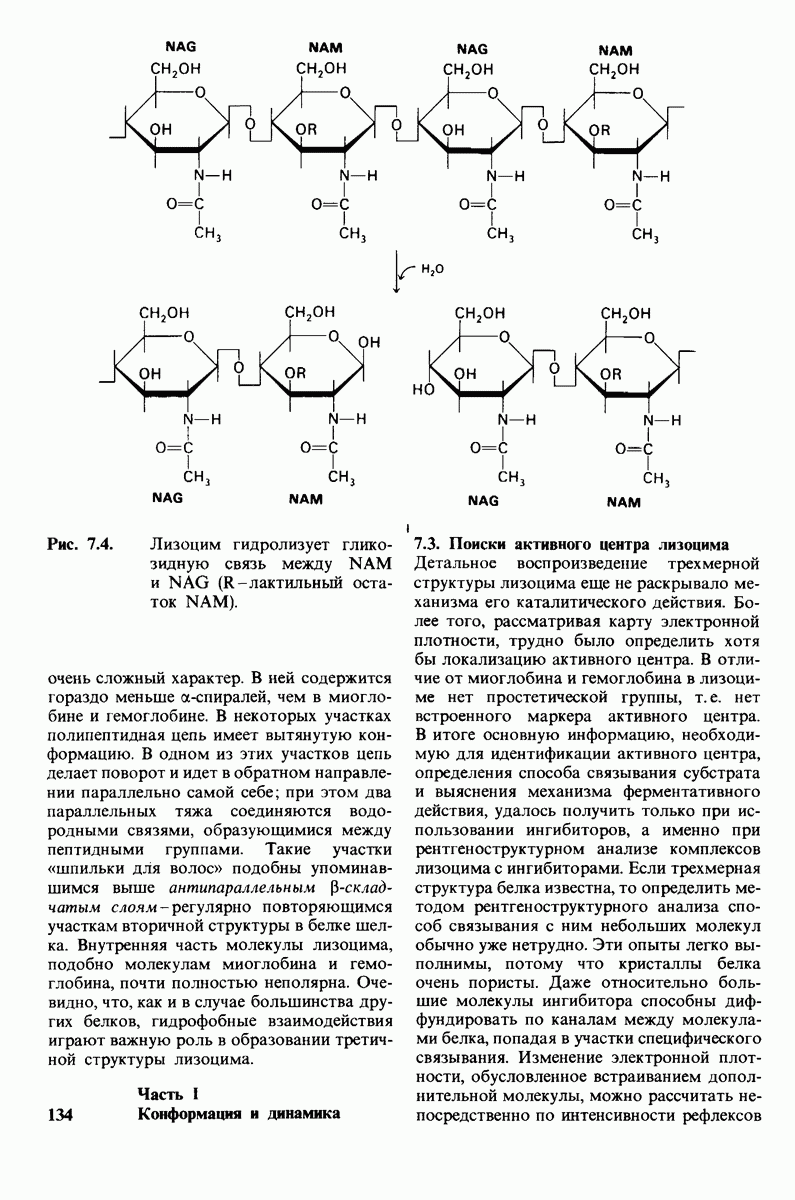
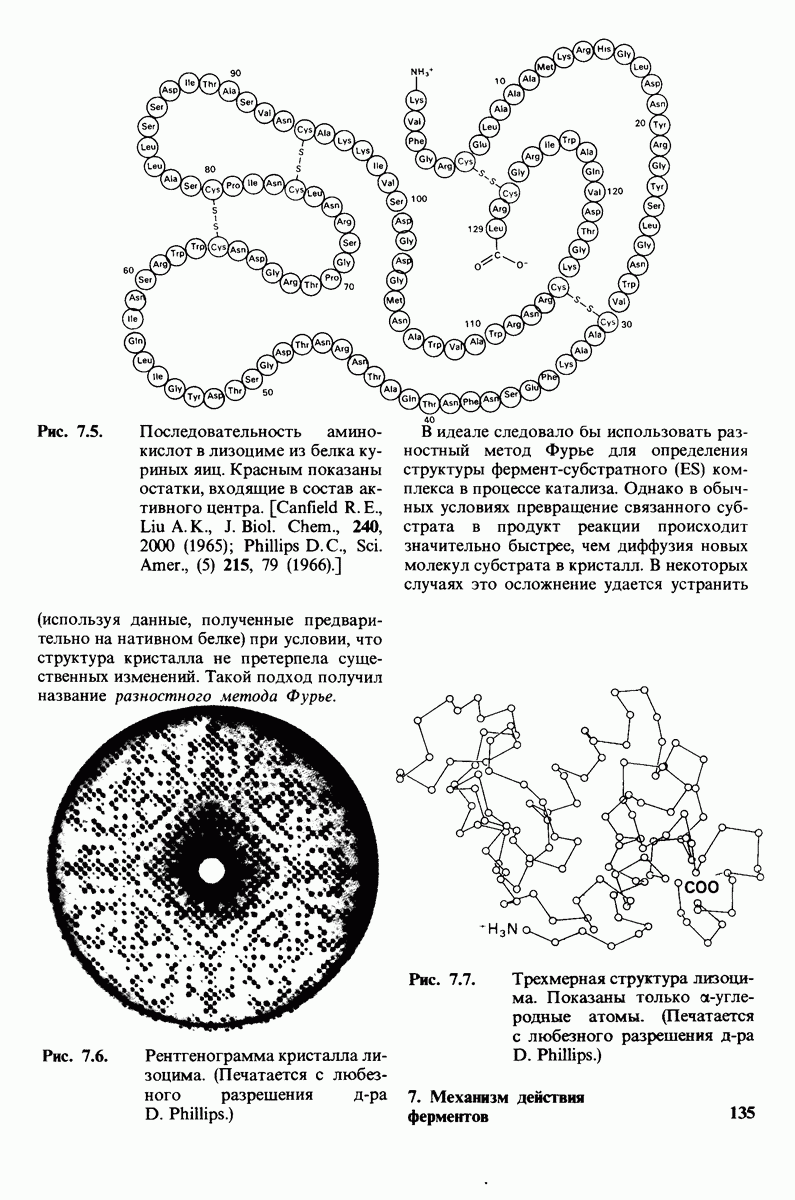
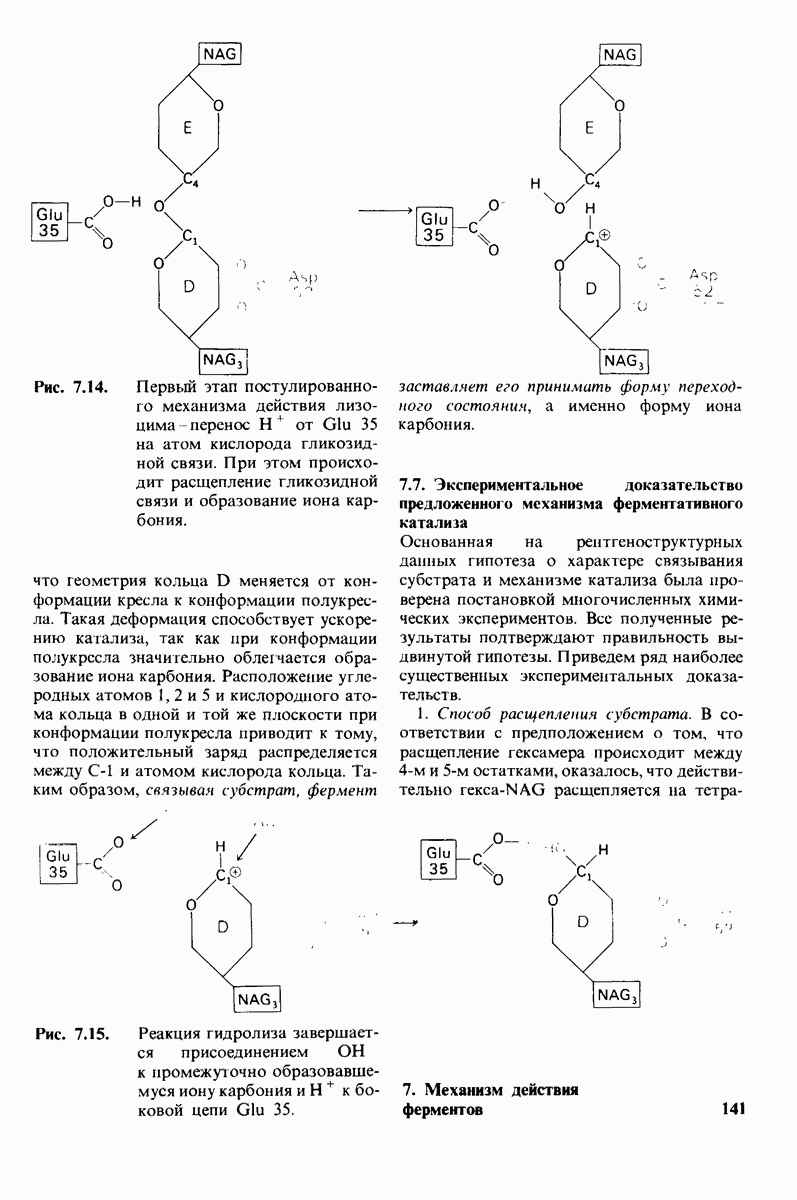
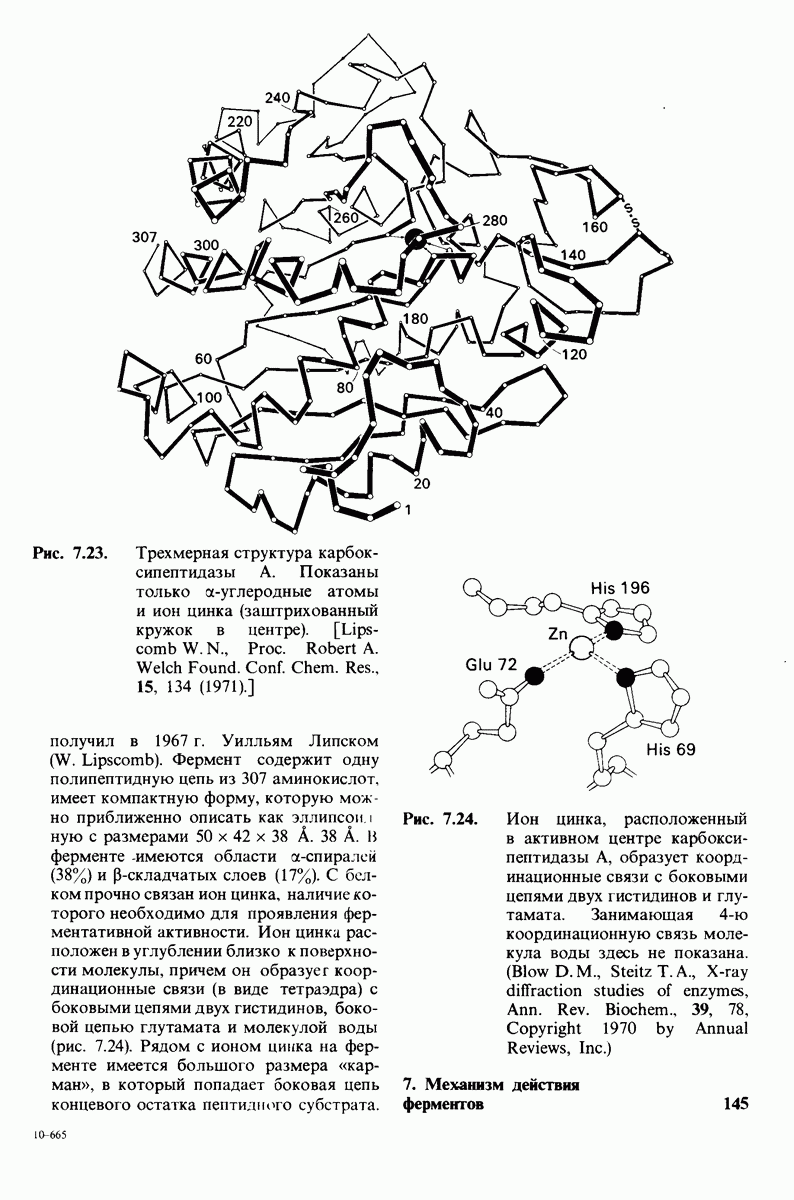
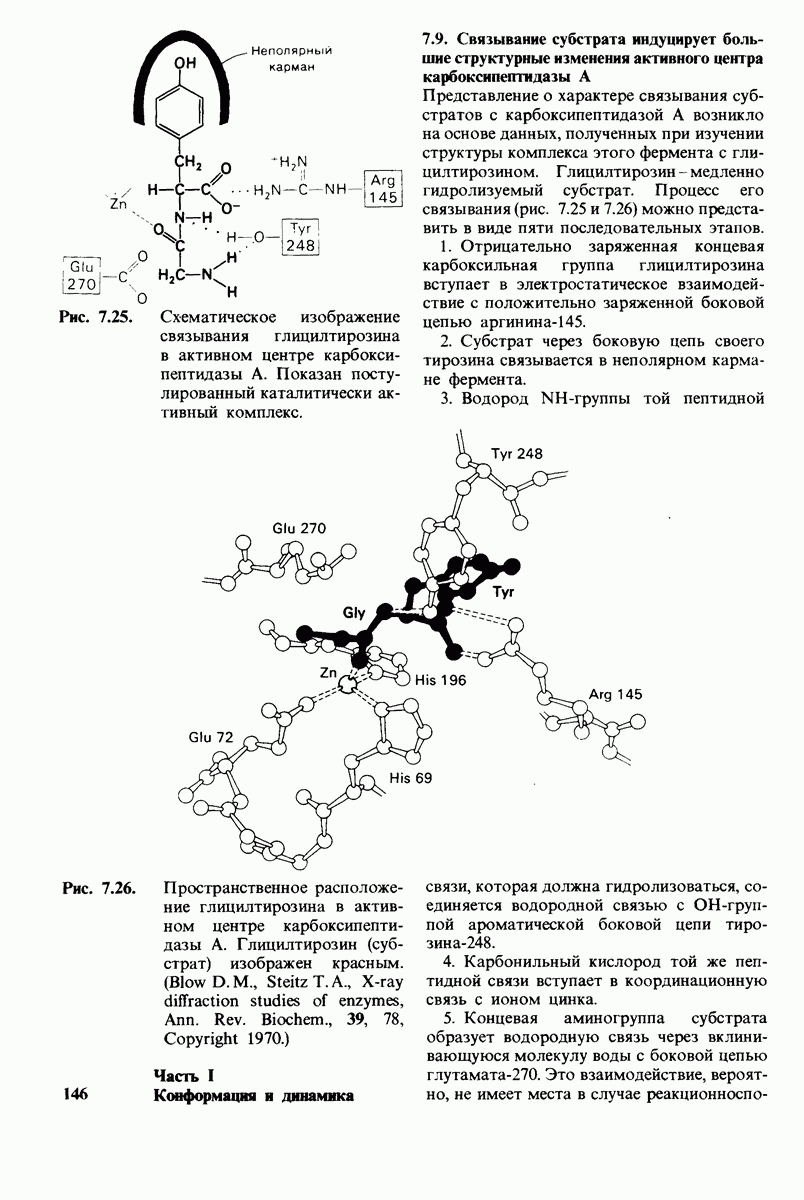
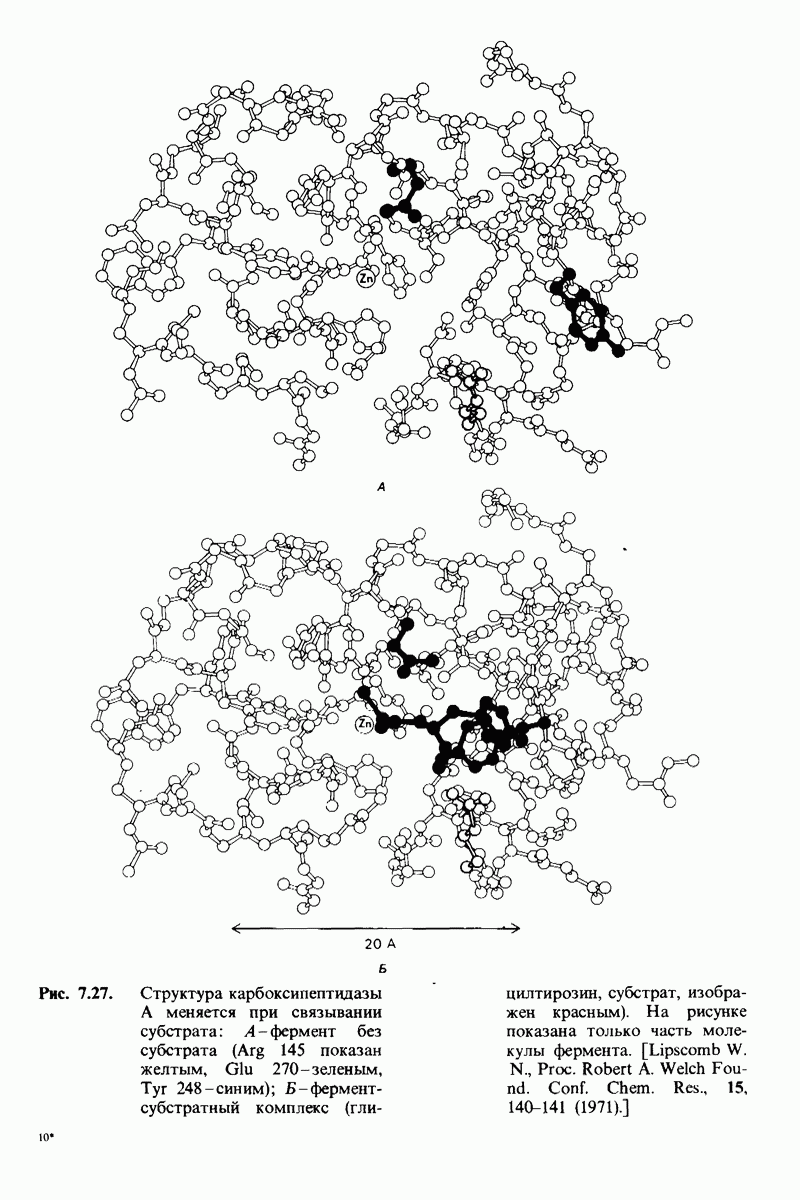
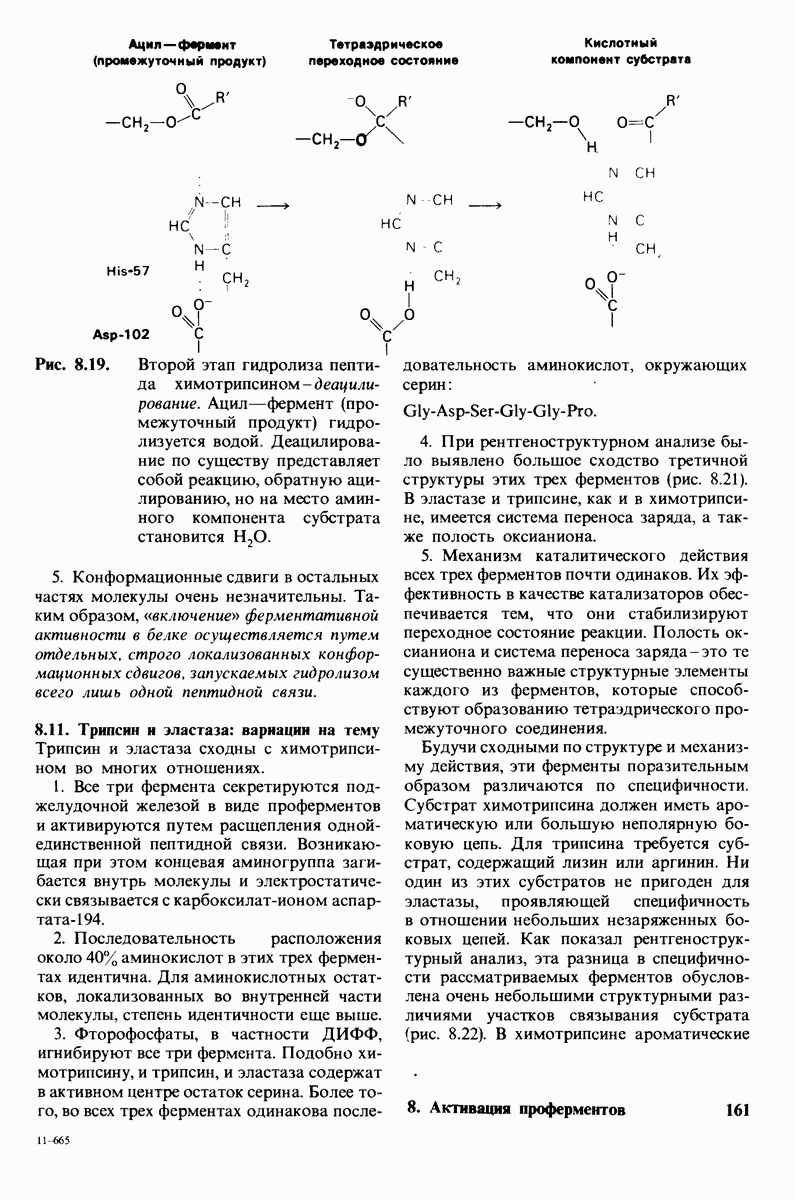
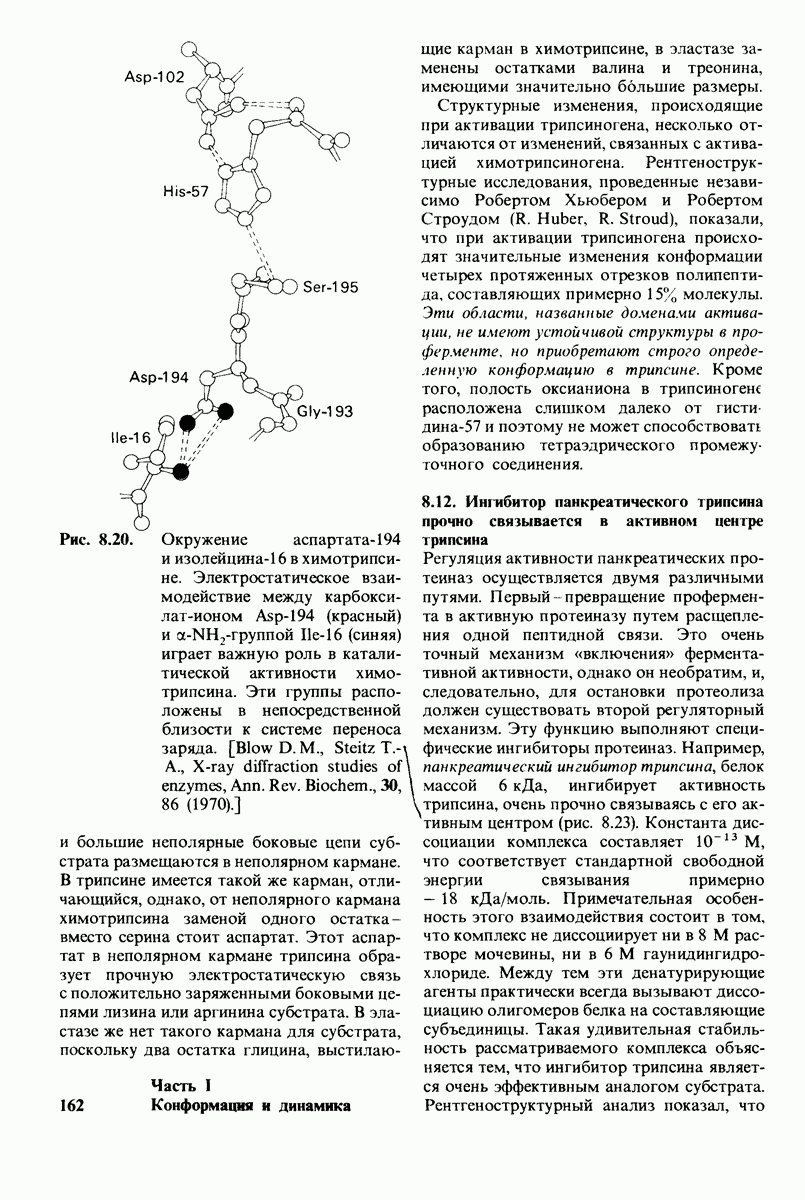
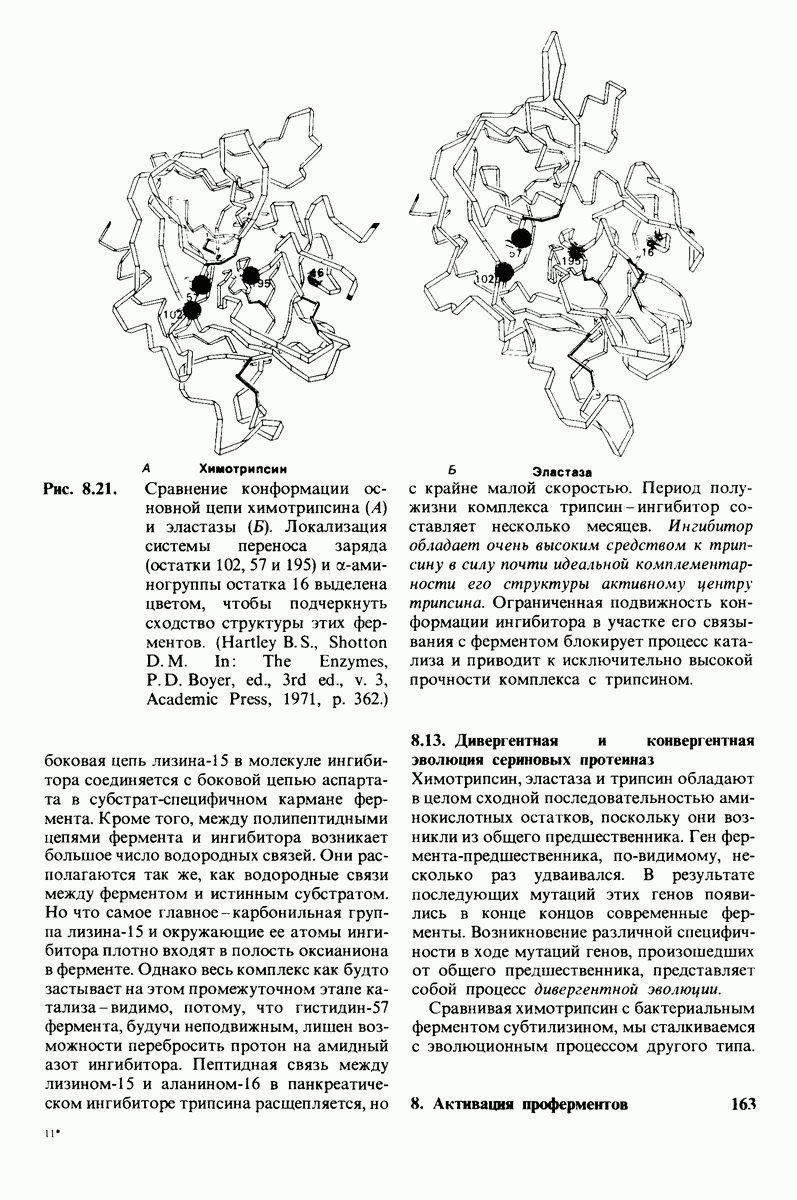
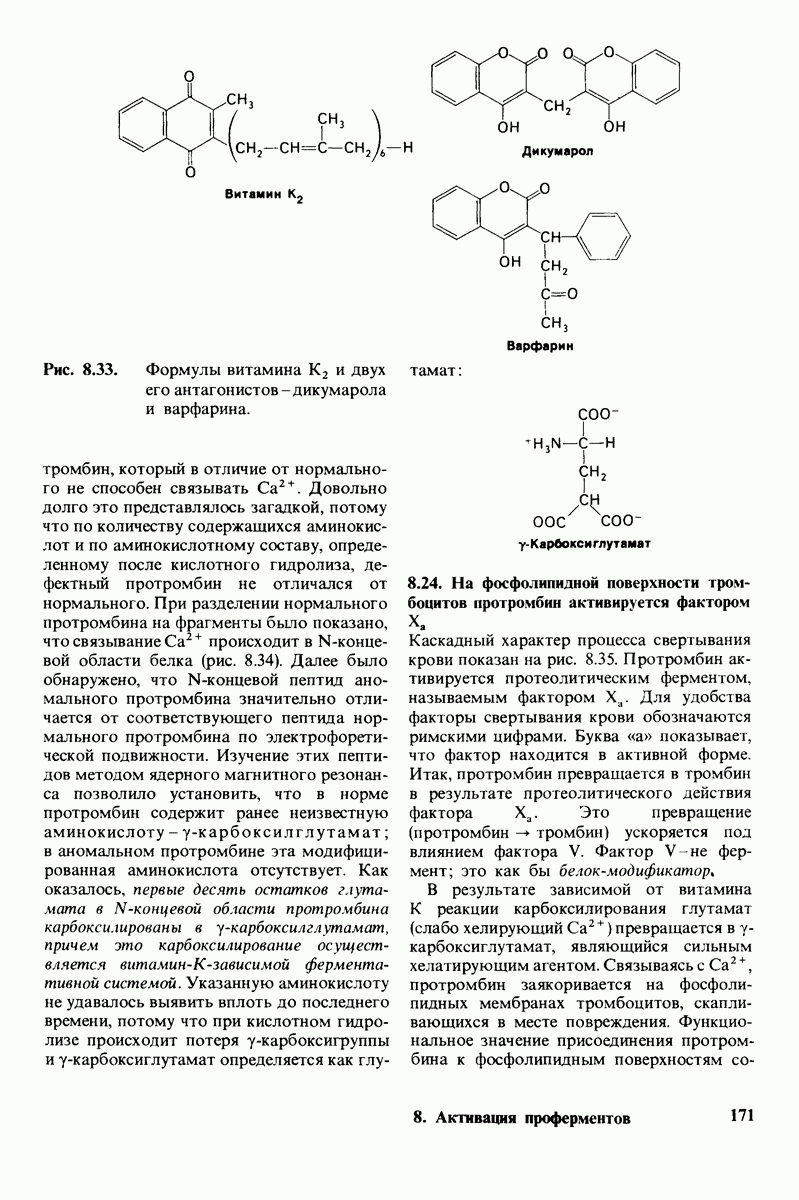
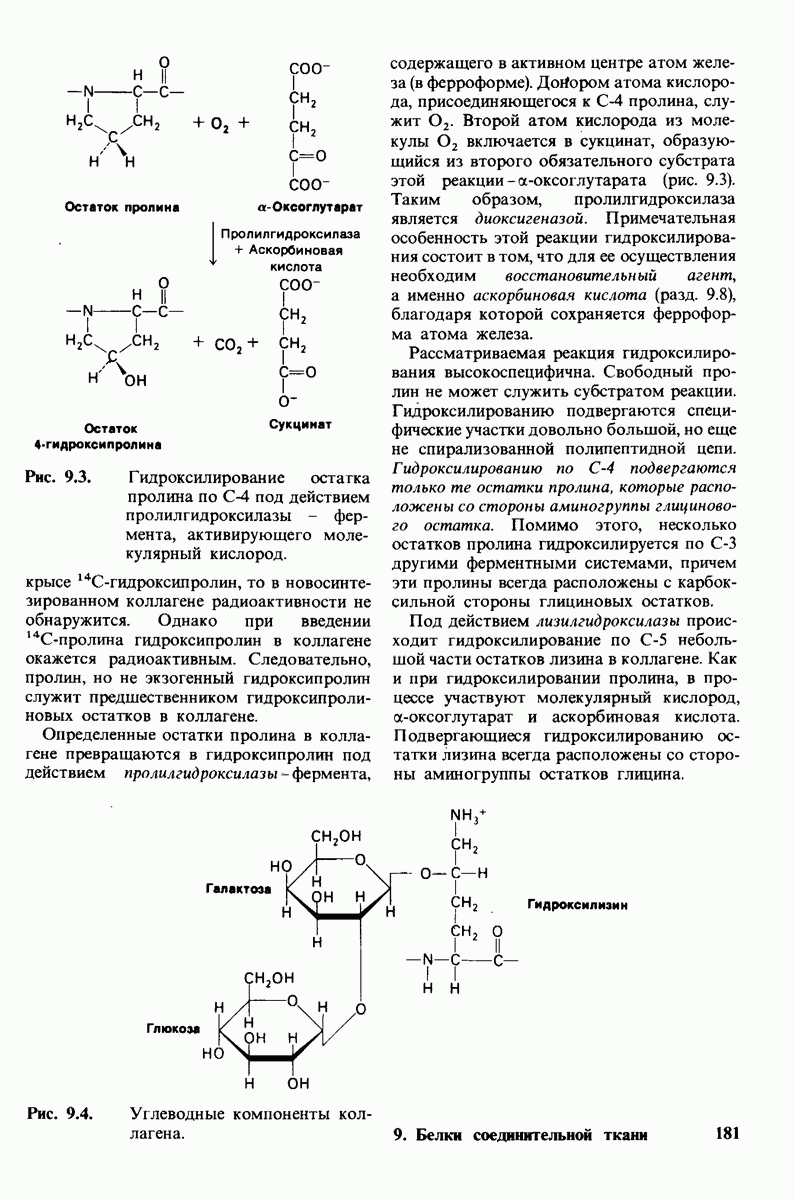
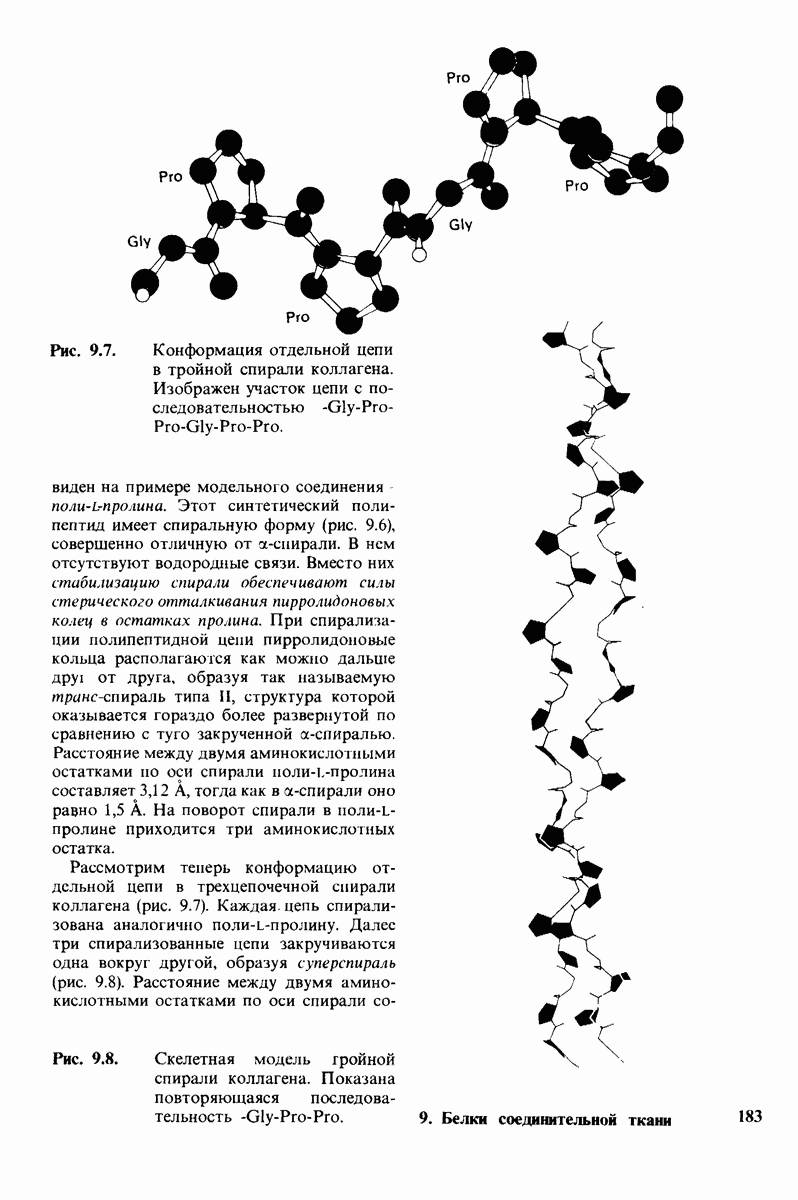
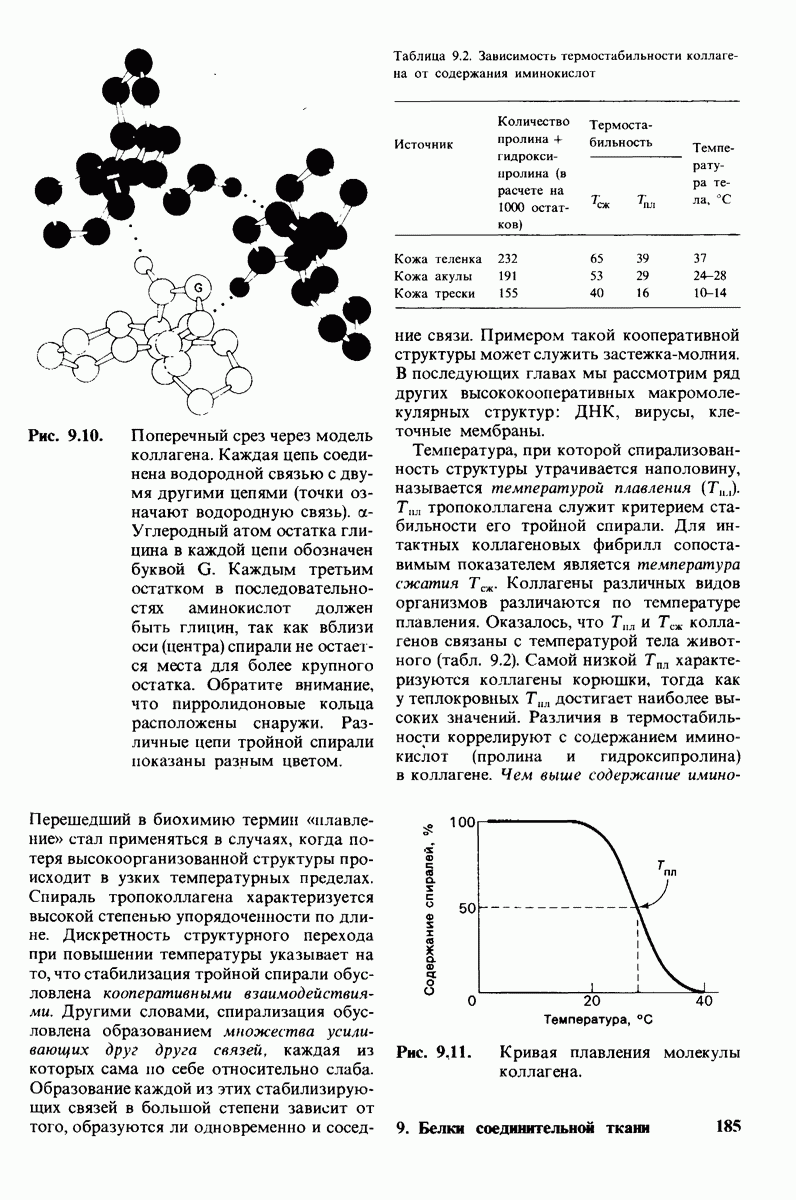
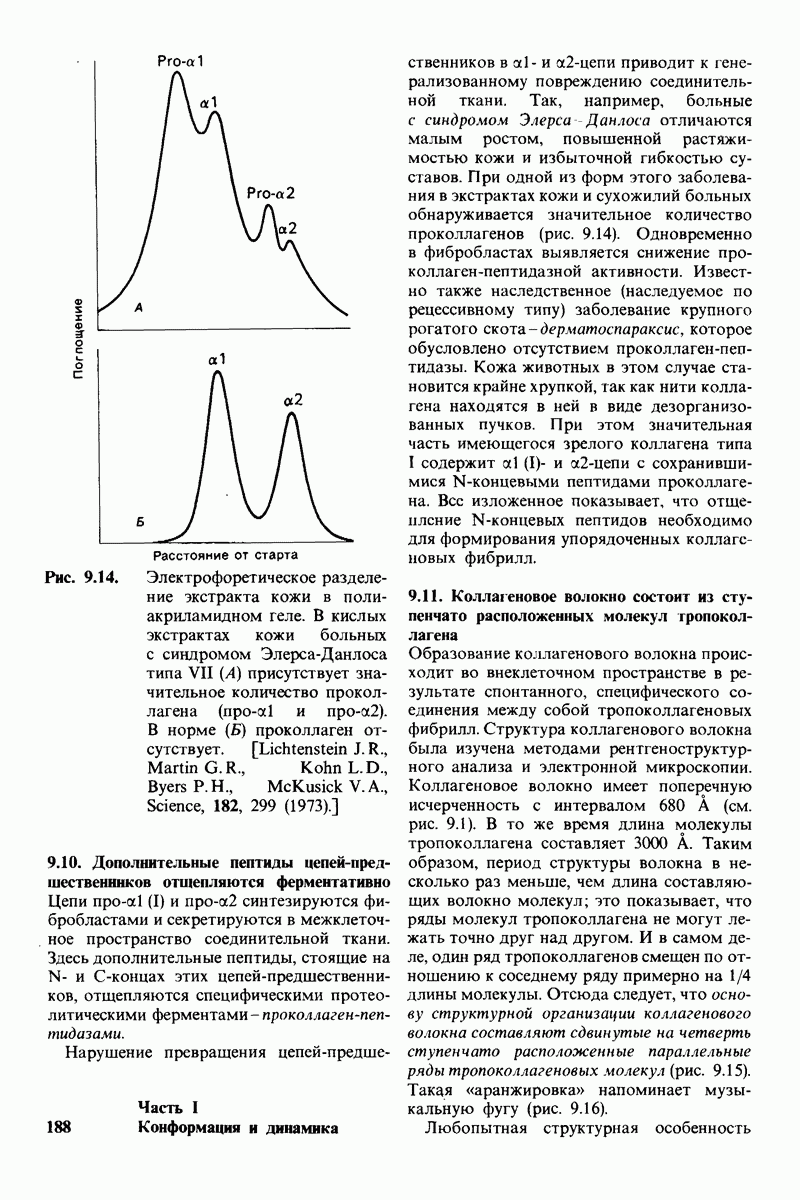
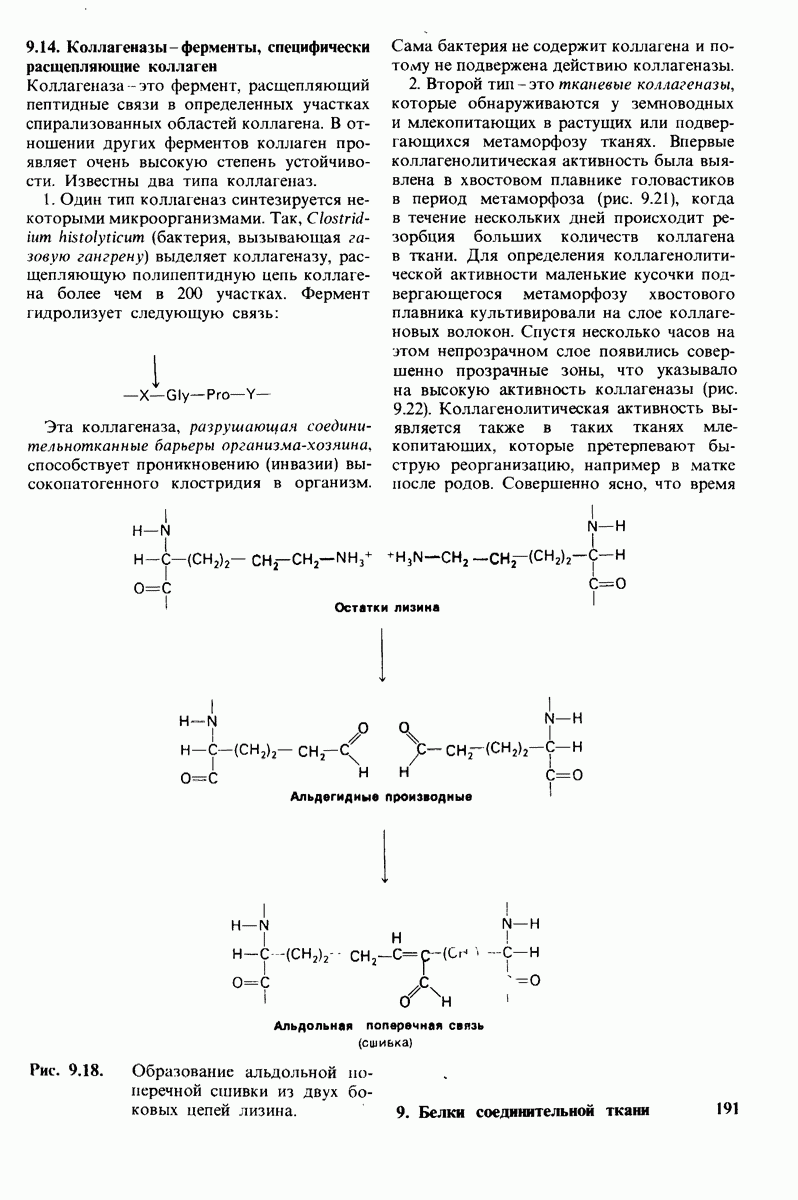
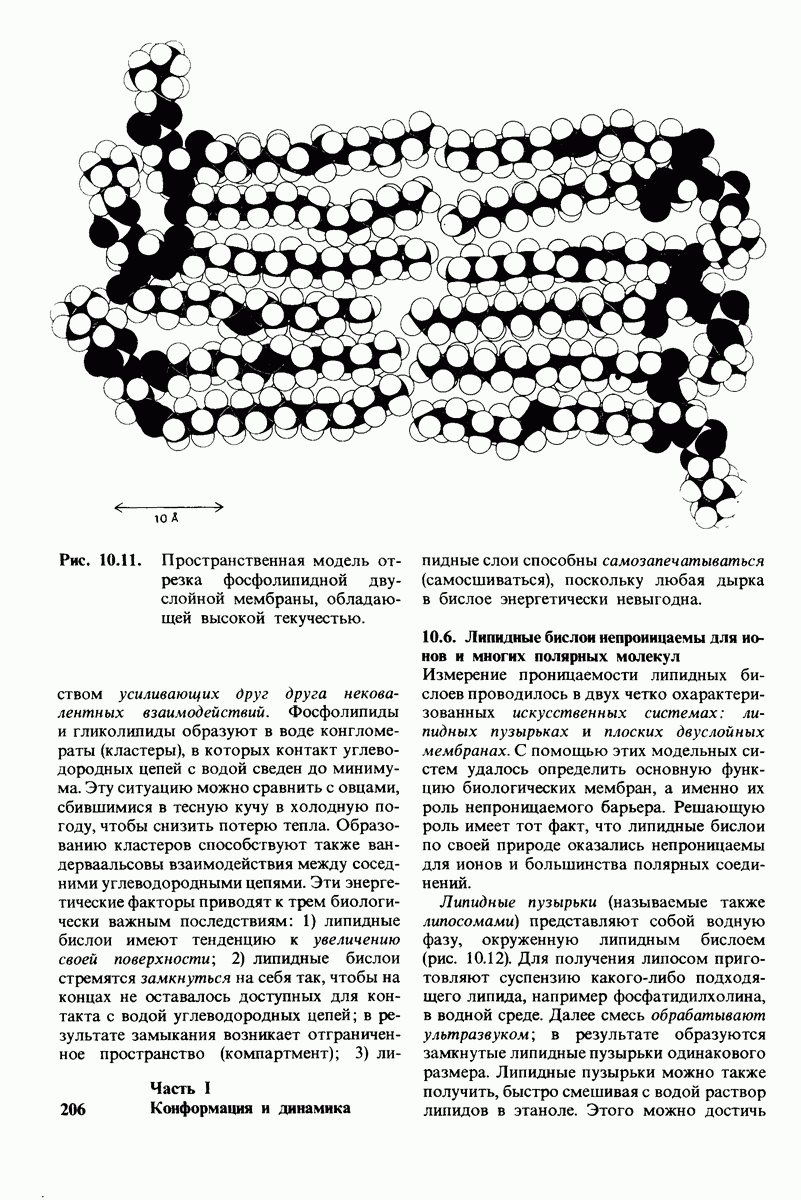
10.17. Текучесть мембран зависит от состава жирных кислот и содержания холестерола
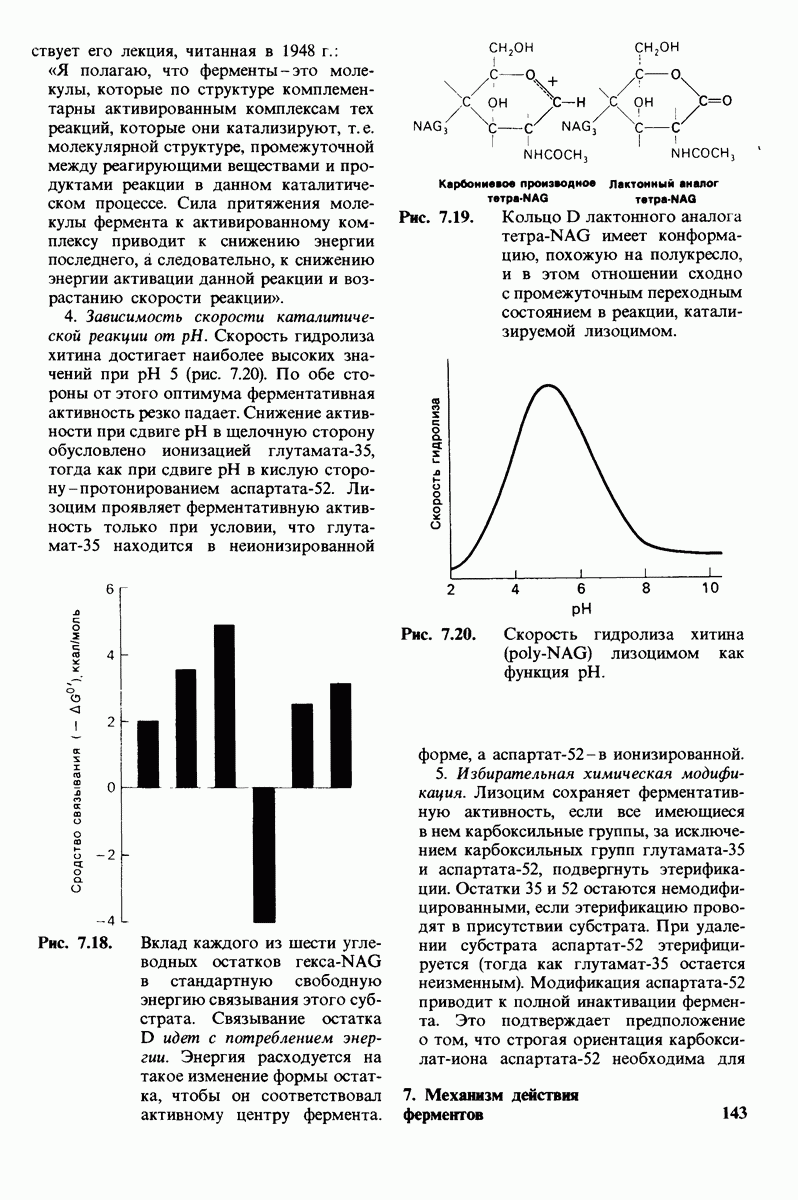
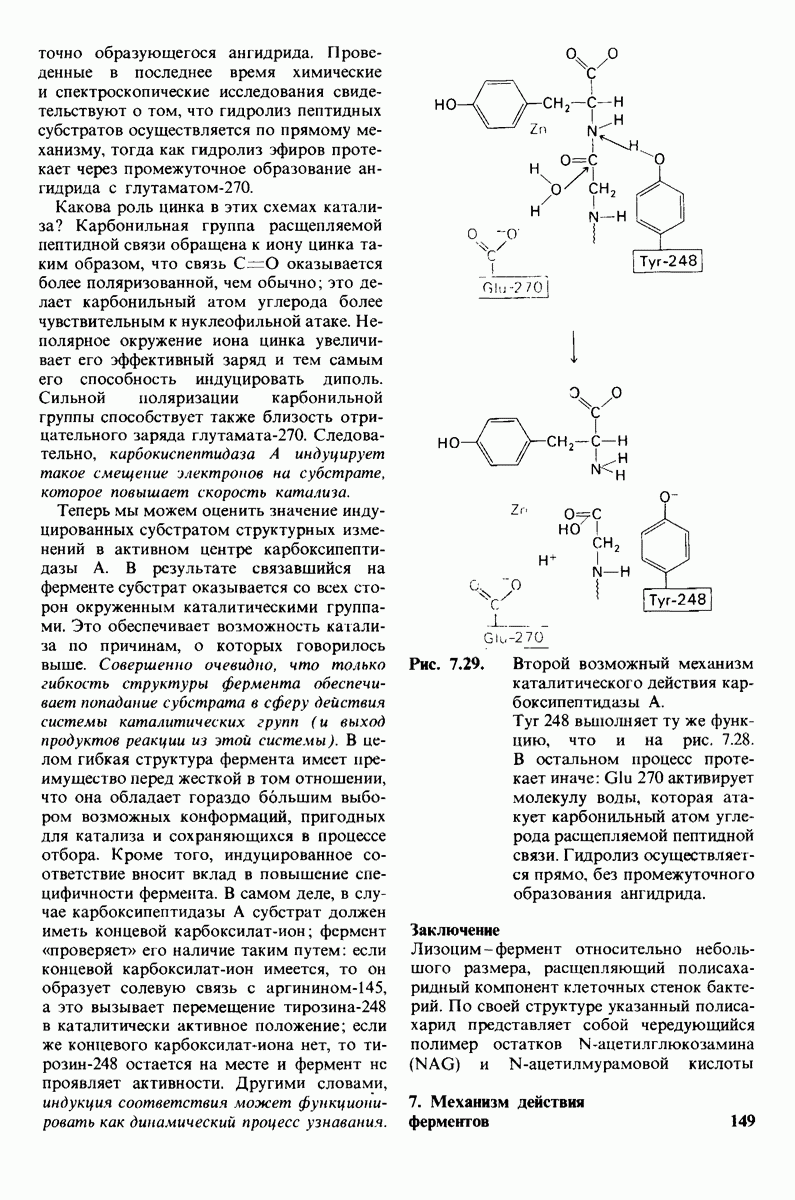
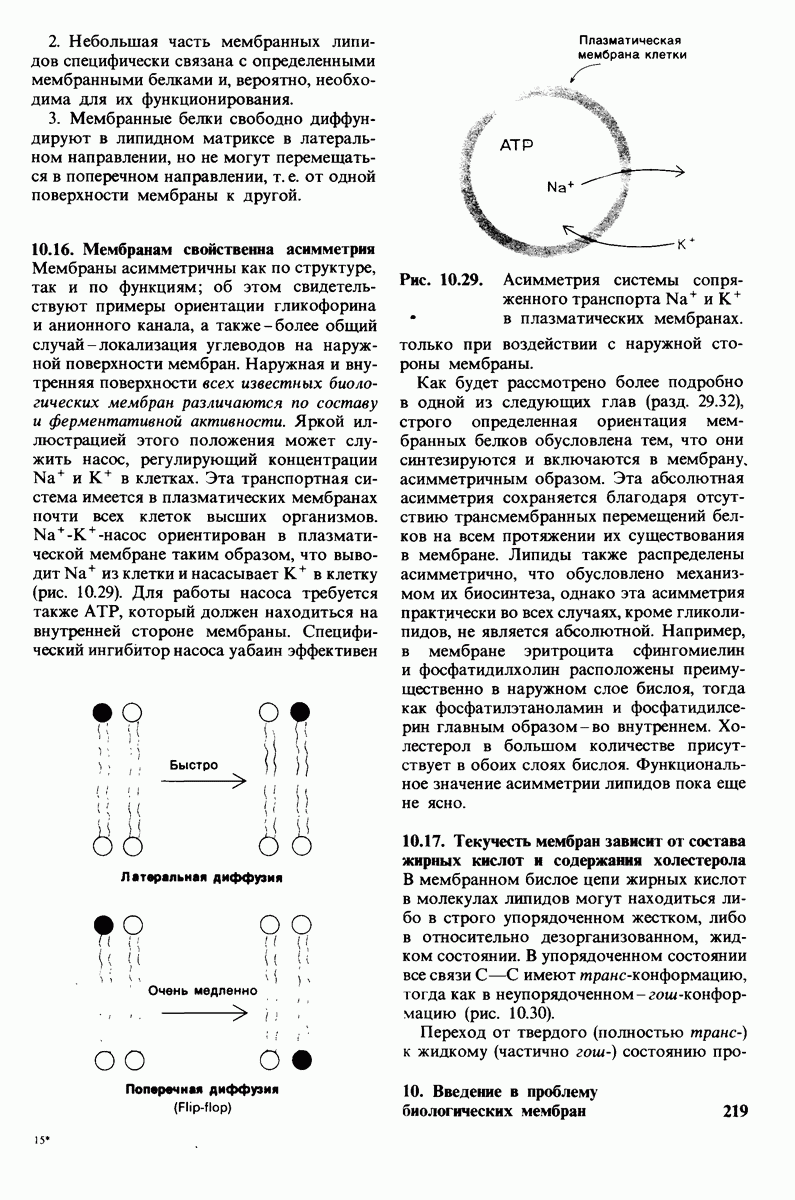
В мембранном бислое цепи жирных кислот в молекулах липидов могут находиться либо в строго упорядоченном жестком, либо в относительно дезорганизованном, жидком состоянии. В упорядоченном состоянии все связи  имеют трансконформацию, тогда как в неупорядоченном — гош-конформацию (рис. 10.30).
имеют трансконформацию, тогда как в неупорядоченном — гош-конформацию (рис. 10.30).
Переход от твердого (полностью транс) к жидкому (частично гош) состоянию

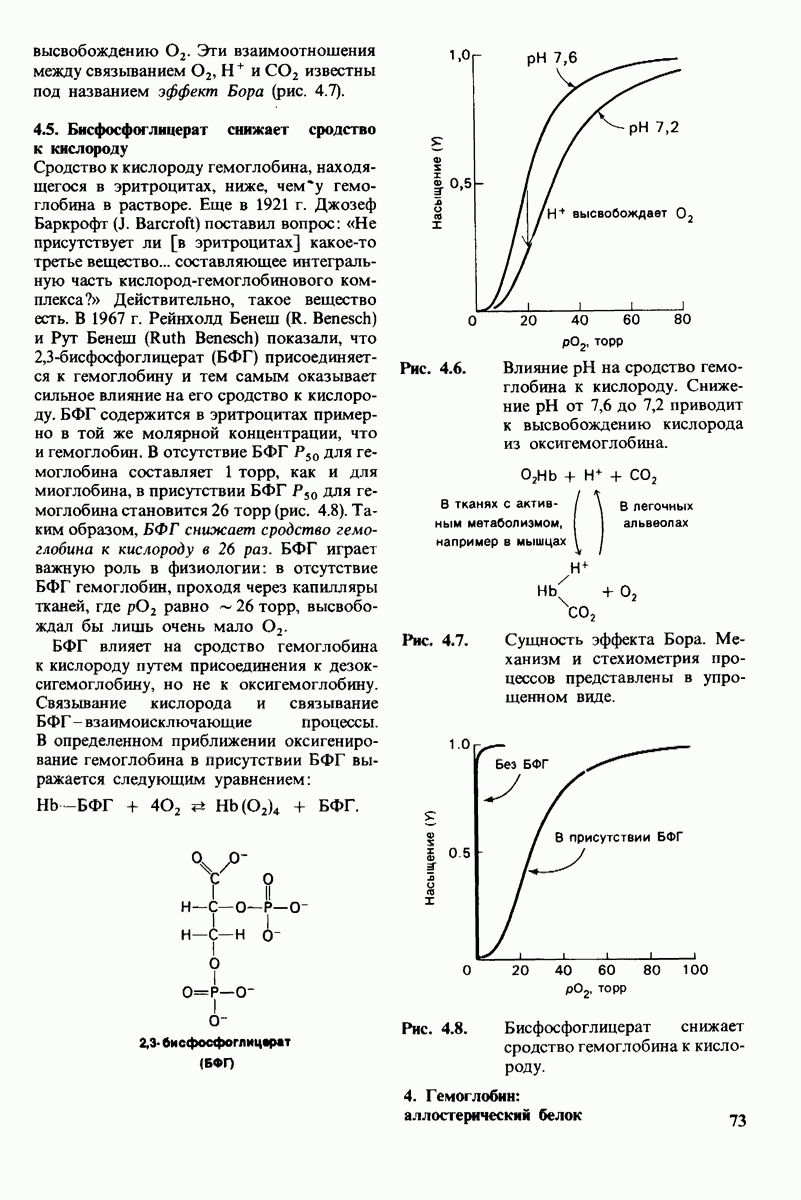
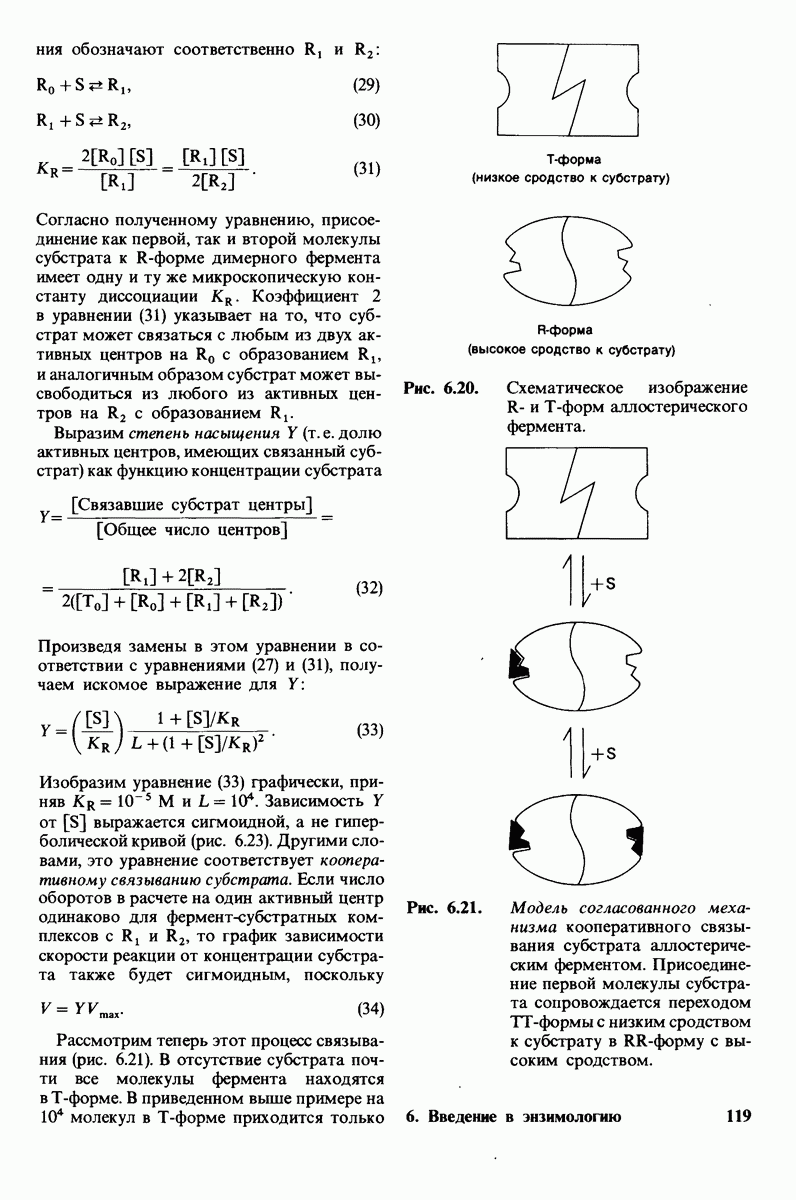
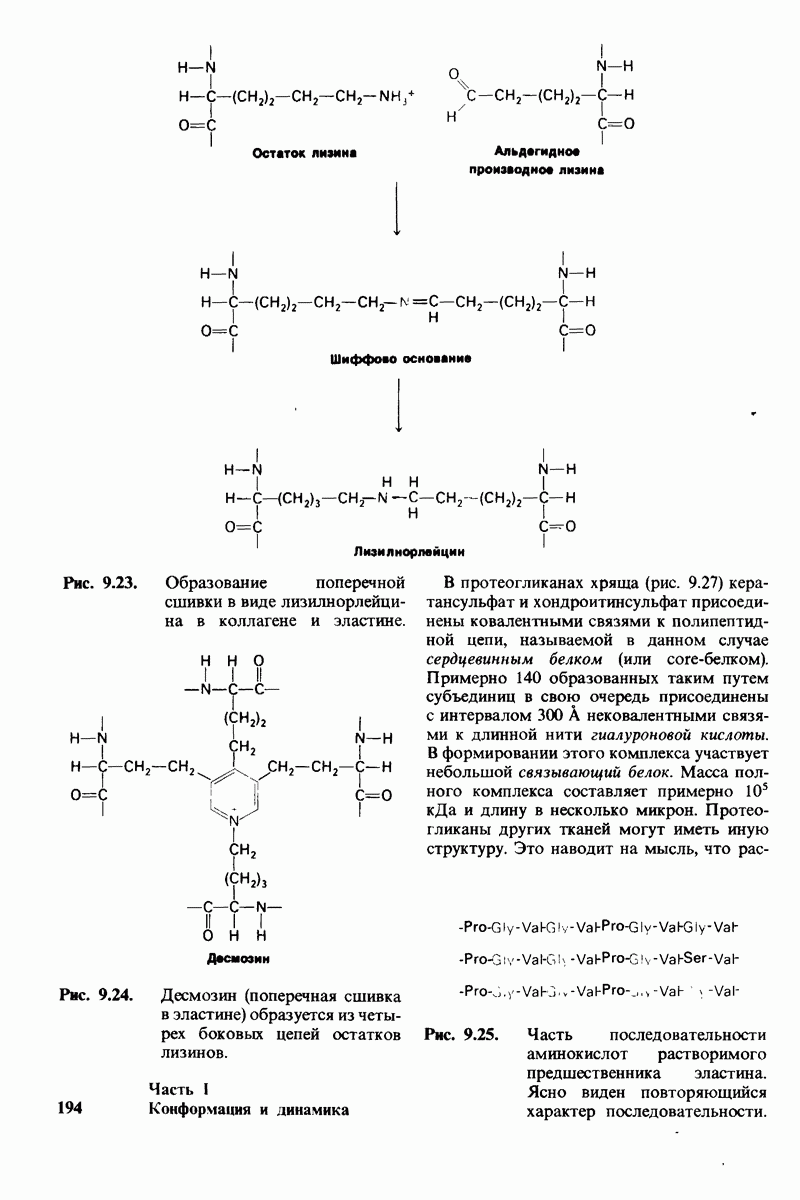
Рис. 10.30. Конформация связей  в углеводородных цепях остатков жирных кислот.
в углеводородных цепях остатков жирных кислот.  - транс
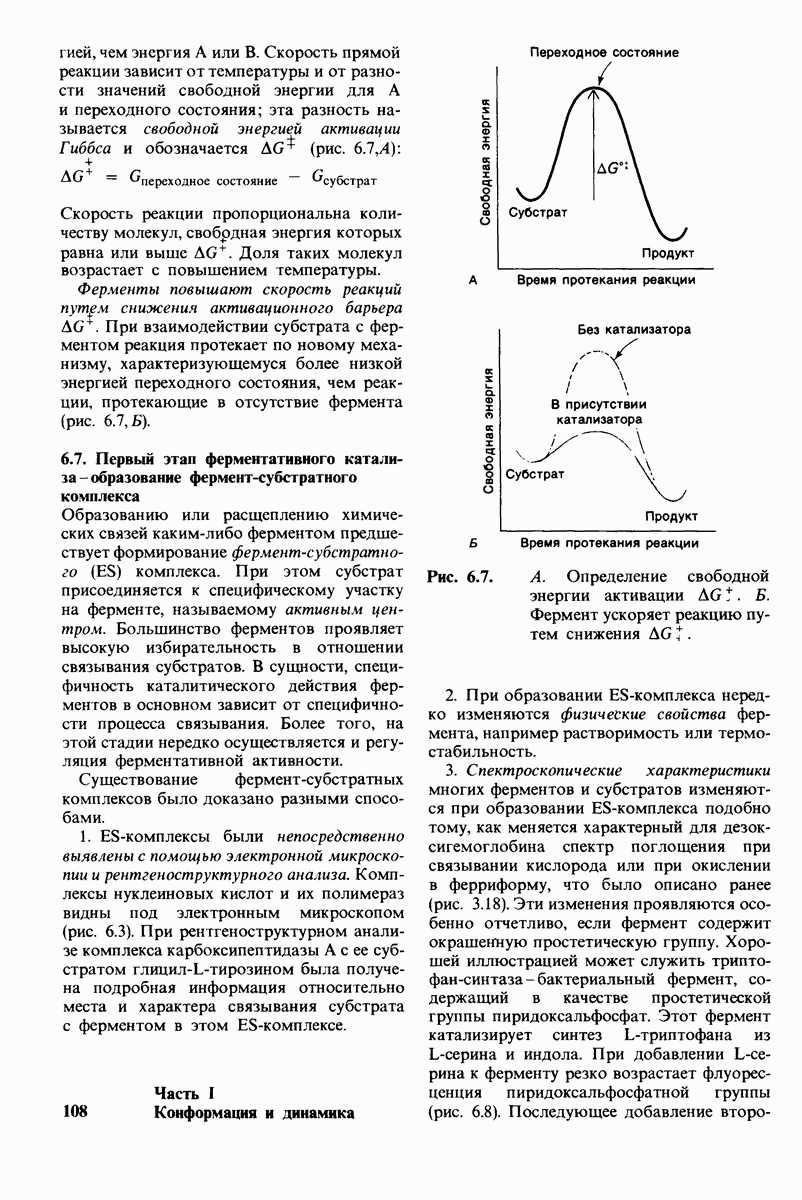
- транс  -конформация.
-конформация.  -поворот на 120° дает гош -конформацию, которая может быть
-поворот на 120° дает гош -конформацию, которая может быть  по часовой стрелке) или
по часовой стрелке) или  против часовой стрелки). При гош-конформации углеводородная цепь изгибается под углом 120°.
против часовой стрелки). При гош-конформации углеводородная цепь изгибается под углом 120°.

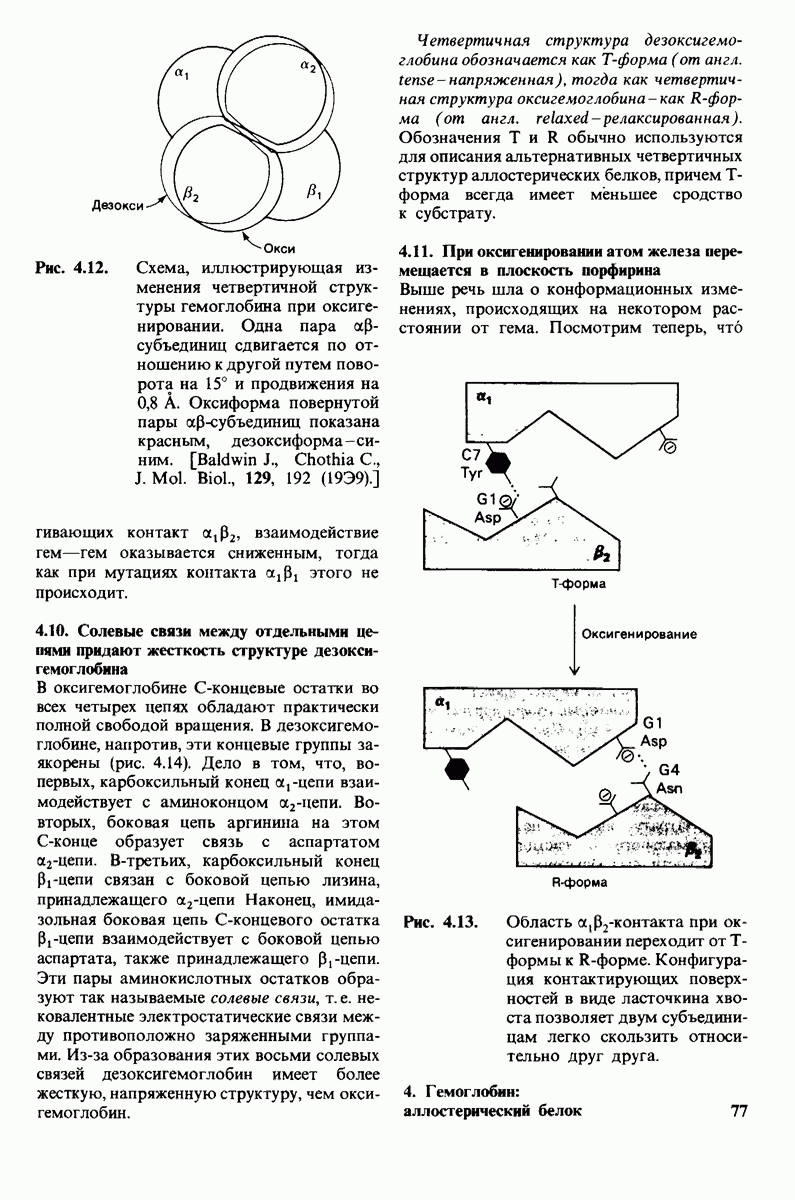
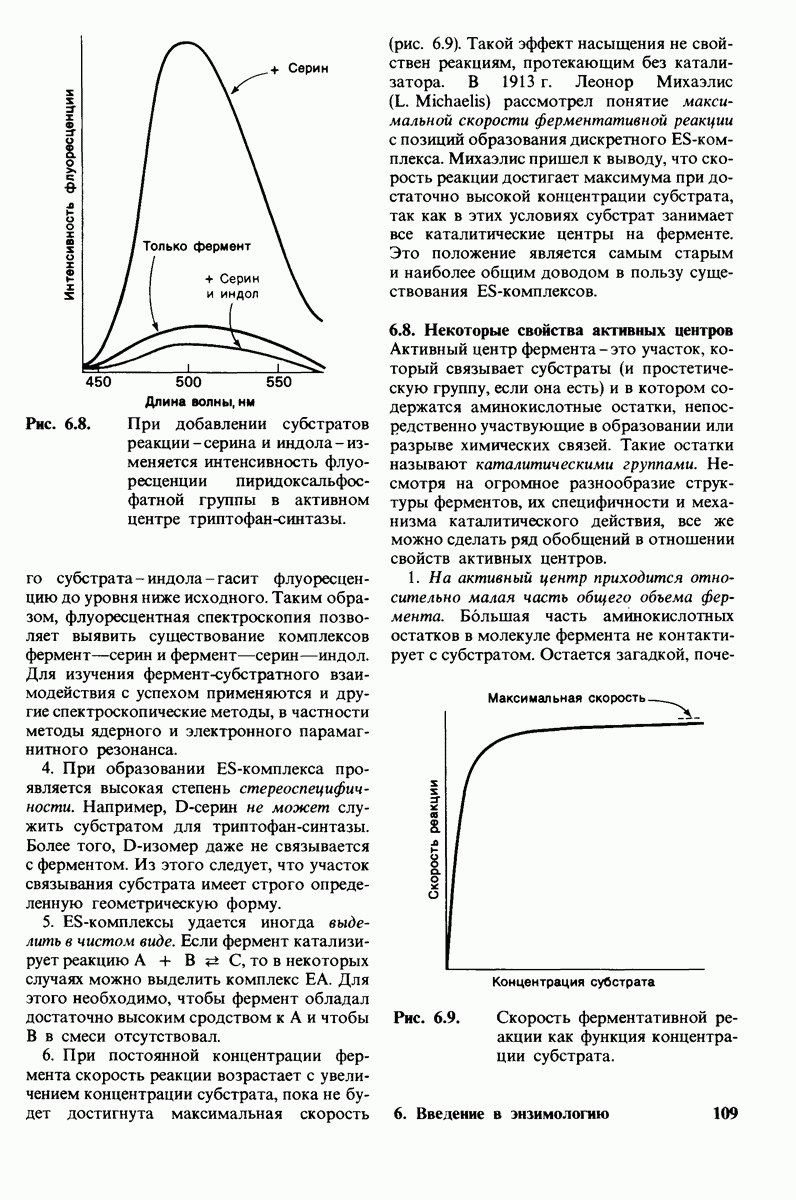
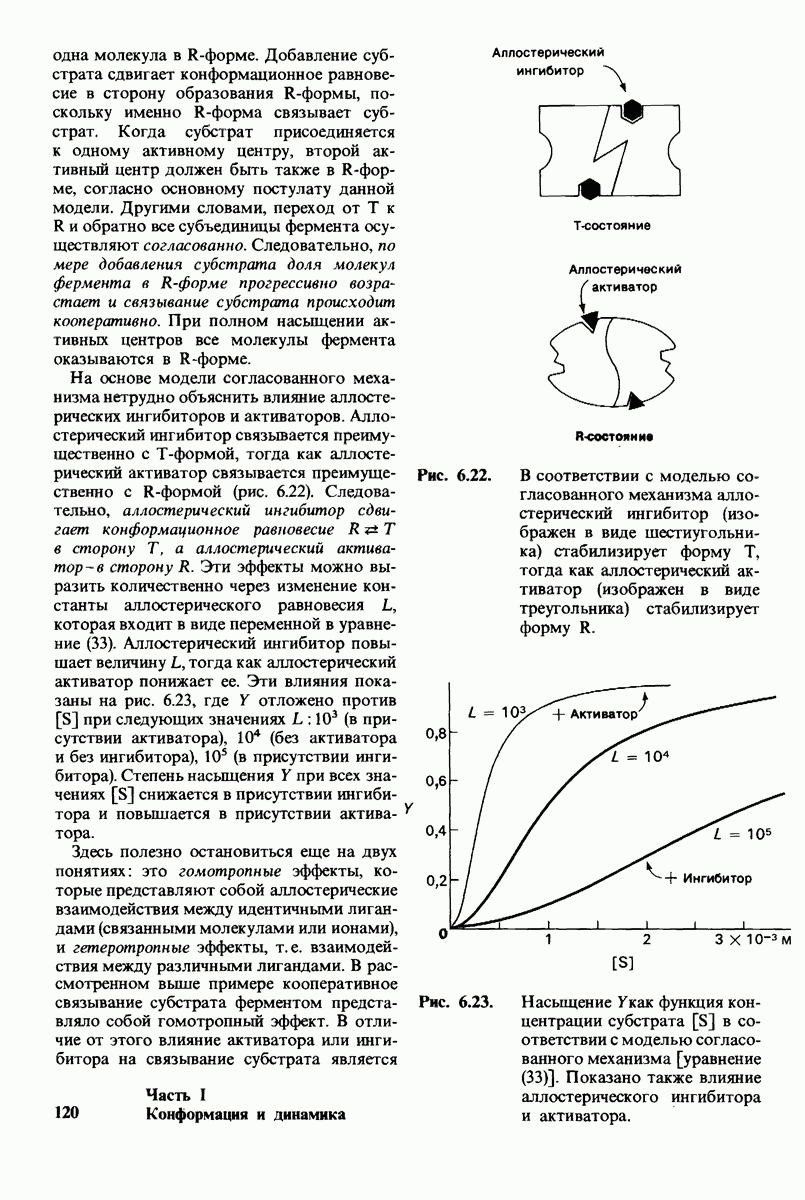
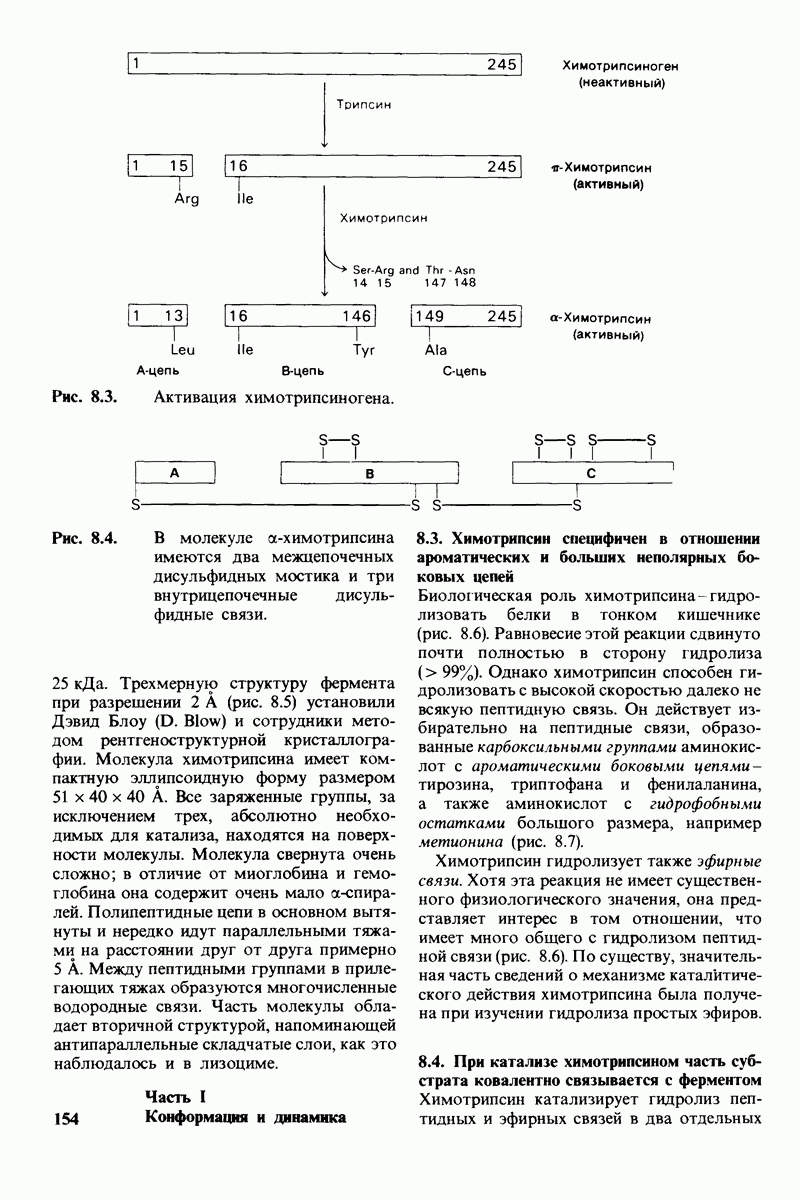
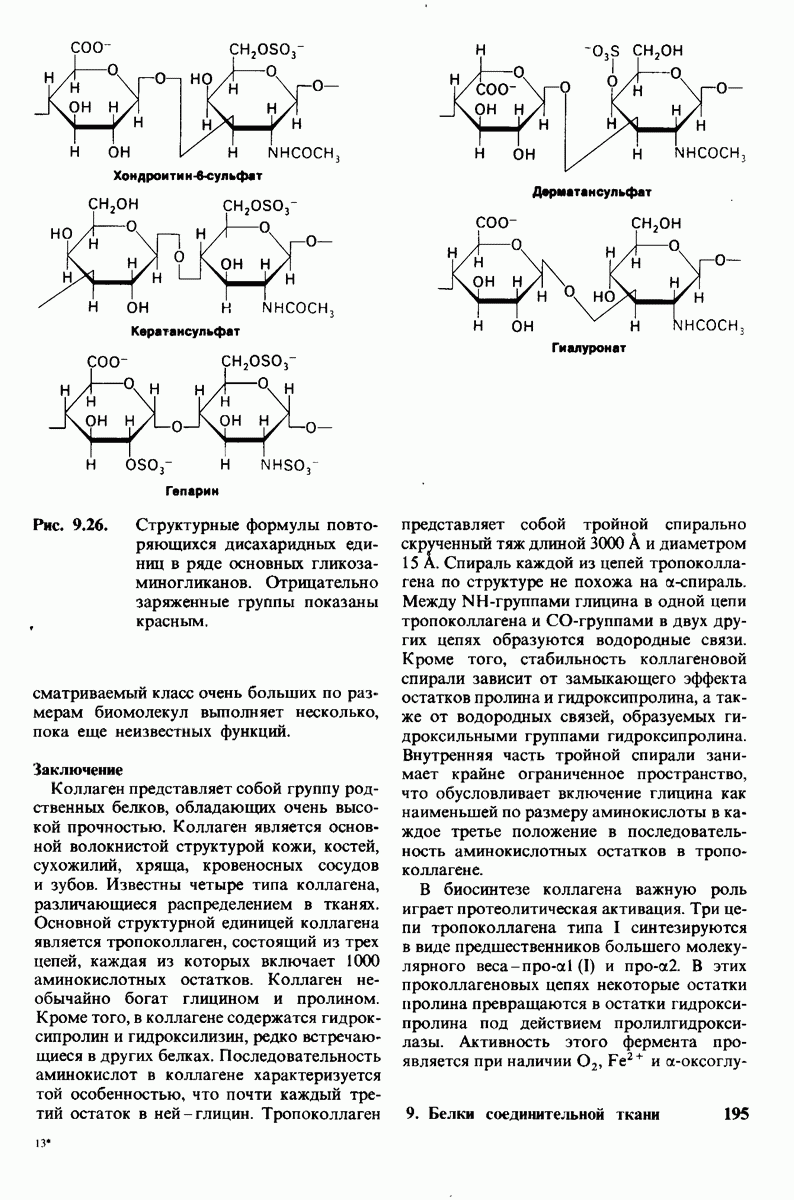
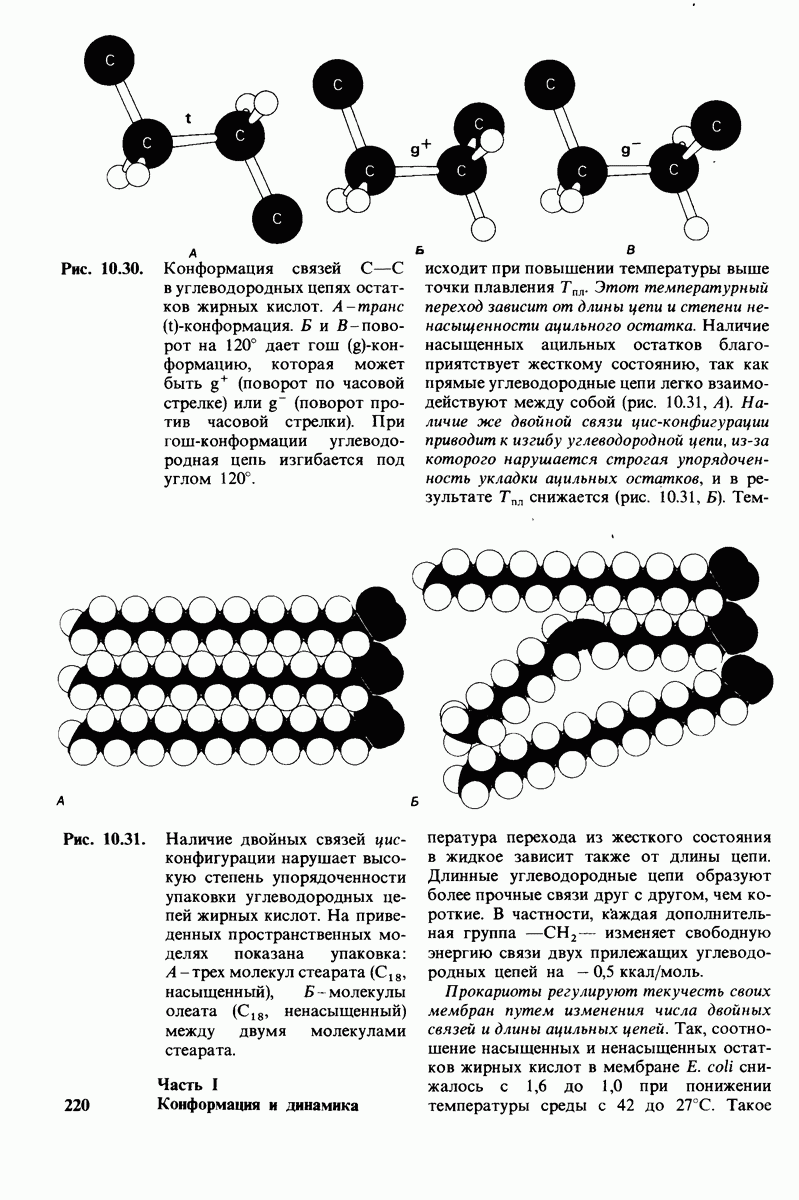
Рис. 10.31. Наличие двойных связей цис-конфигурации нарушает высокую степень упорядоченности упаковки углеводородных цепей жирных кислот. На приведенных пространственных моделях показана упаковка: А - трех молекул стеарата  насыщенный),
насыщенный),  молекулы олеата
молекулы олеата  ненасыщенный) между двумя молекулами стеарата.
ненасыщенный) между двумя молекулами стеарата.
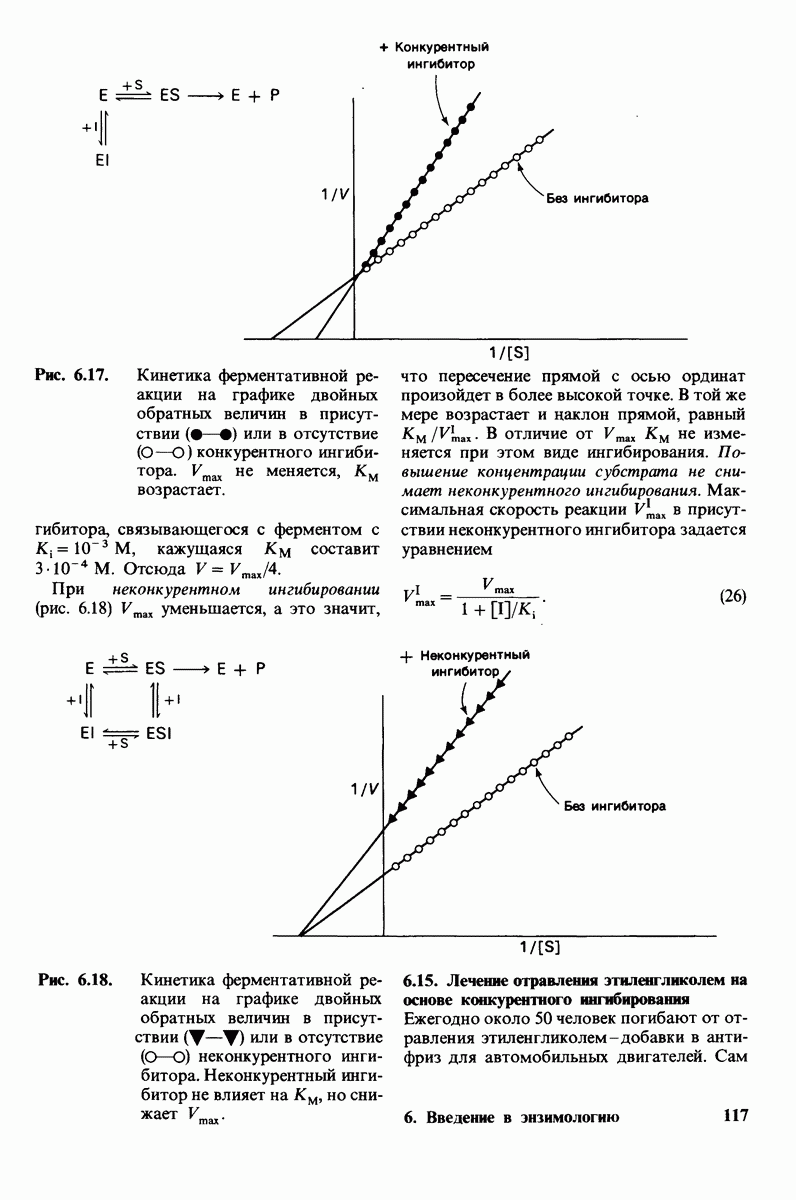
происходит при повышении температуры выше точки плавления  Этот температурный переход зависит от длины цепи и степени ненасыщенности ацильного остатка. Наличие насыщенных ацильных остатков благоприятствует жесткому состоянию, так как прямые углеводородные цепи легко взаимодействуют между собой (рис. 10.31, А). Наличие же двойной связи цис-конфигурации приводит к изгибу углеводородной цепи, из-за которого нарушается строгая упорядоченность укладки ацильных остатков, и в результате
Этот температурный переход зависит от длины цепи и степени ненасыщенности ацильного остатка. Наличие насыщенных ацильных остатков благоприятствует жесткому состоянию, так как прямые углеводородные цепи легко взаимодействуют между собой (рис. 10.31, А). Наличие же двойной связи цис-конфигурации приводит к изгибу углеводородной цепи, из-за которого нарушается строгая упорядоченность укладки ацильных остатков, и в результате  снижается (рис.
снижается (рис.  Температура перехода из жесткого состояния в жидкое зависит также от длины цепи. Длинные углеводородные цепи образуют более прочные связи друг с другом, чем короткие. В частности, каждая дополнительная группа
Температура перехода из жесткого состояния в жидкое зависит также от длины цепи. Длинные углеводородные цепи образуют более прочные связи друг с другом, чем короткие. В частности, каждая дополнительная группа  изменяет свободную энергию связи двух прилежащих углеводородных цепей на — 0,5 ккал/моль.
изменяет свободную энергию связи двух прилежащих углеводородных цепей на — 0,5 ккал/моль.
Прокариоты регулируют текучесть своих мембран путем изменения числа двойных связей и длины ацильных цепей. Так, соотношение насыщенных и ненасыщенных остатков жирных кислот в мембране Е. coli снижалось с 1,6 до 1,0 при понижении температуры среды с 42 до  Такое
Такое
уменьшение доли насыщенных жирных кислот предотвращает чрезмерное затвердевание мембраны при пониженной температуре. У эукариот ключевым регулятором текучести мембран является также холестерол. Находясь между ацильными цепями, холестерол препятствует их кристаллизации. В сущности, из-за холестерола исчезает фазовый переход. С другой стороны, холестерол стерически блокирует сильное перемещение ацильных цепей и тем самым снижает текучесть мембран. Таким образом, благодаря этим взаимопротивоположным эффектам холестерола текучесть мембран поддерживается на каком-то среднем уровне.