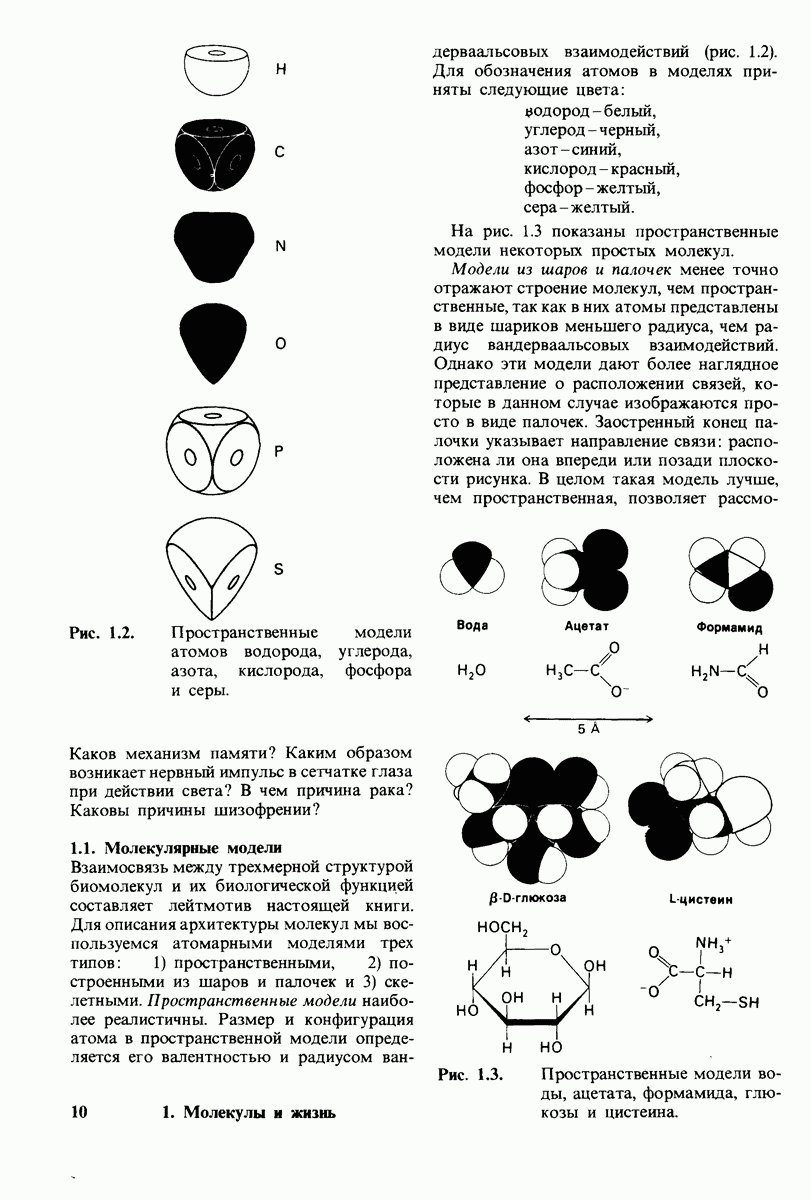
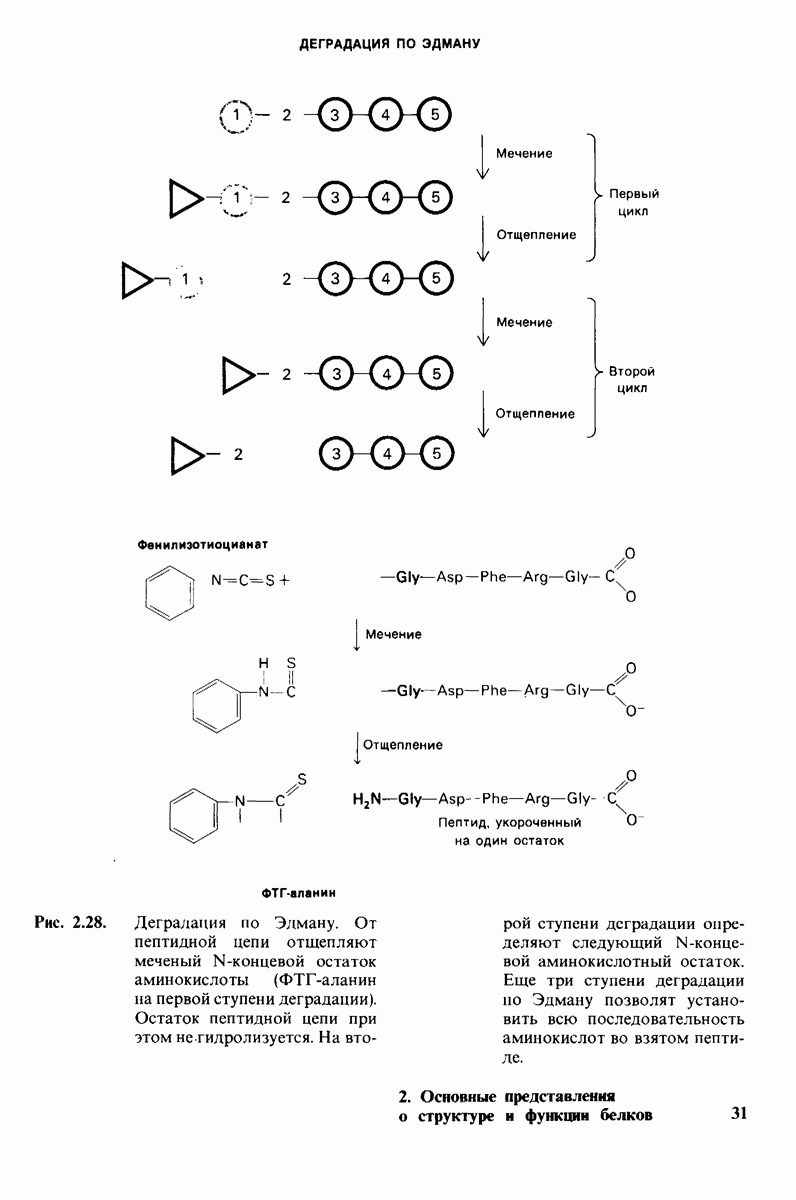
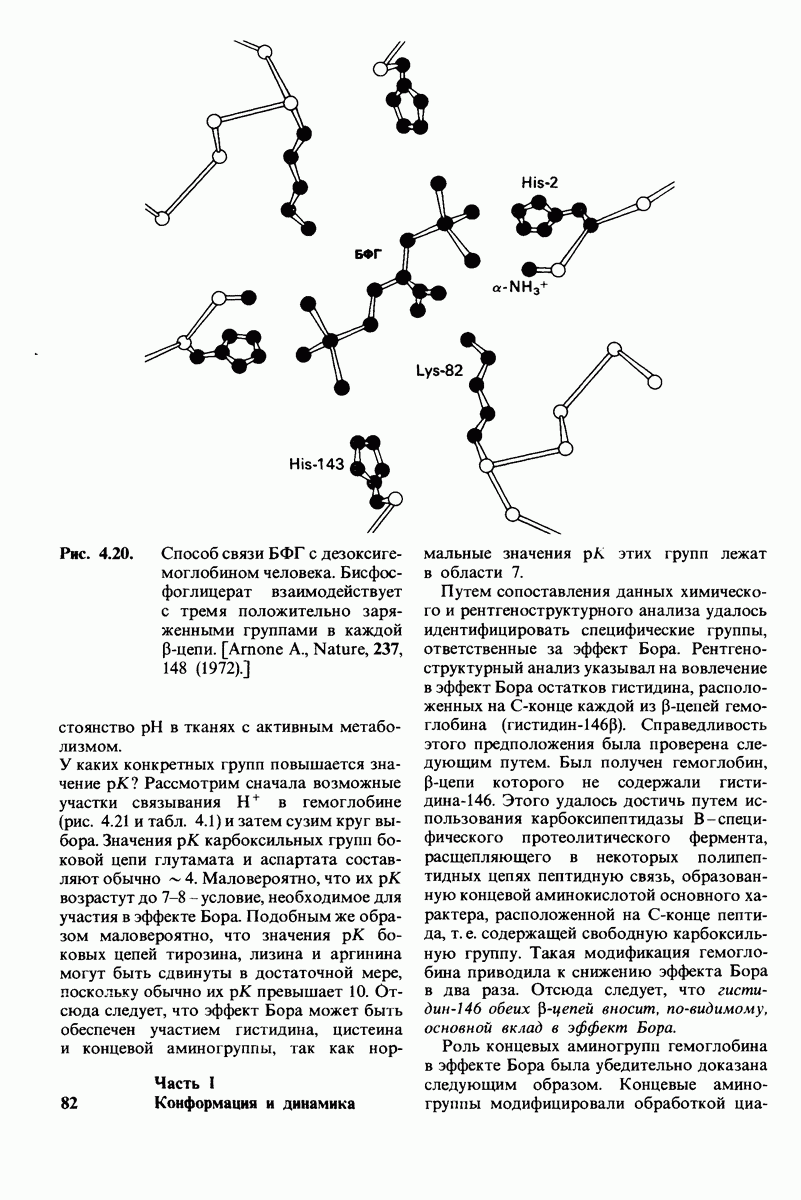
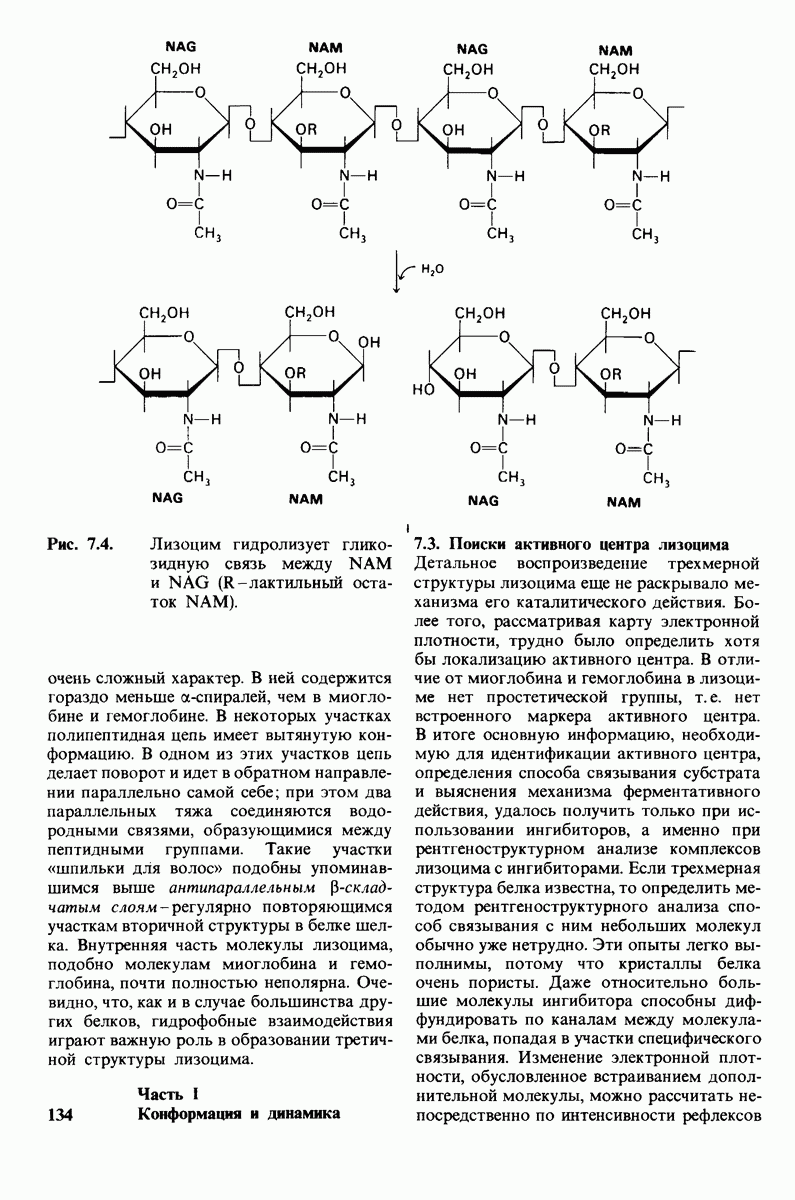
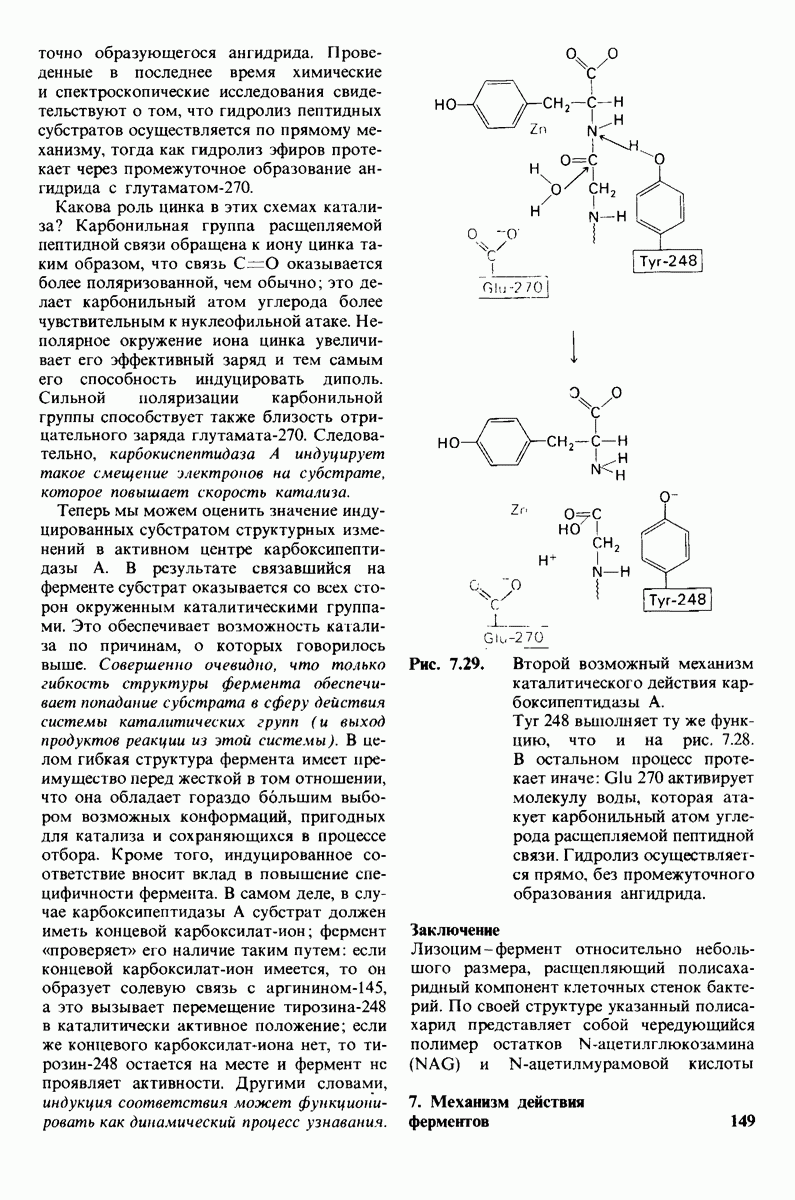
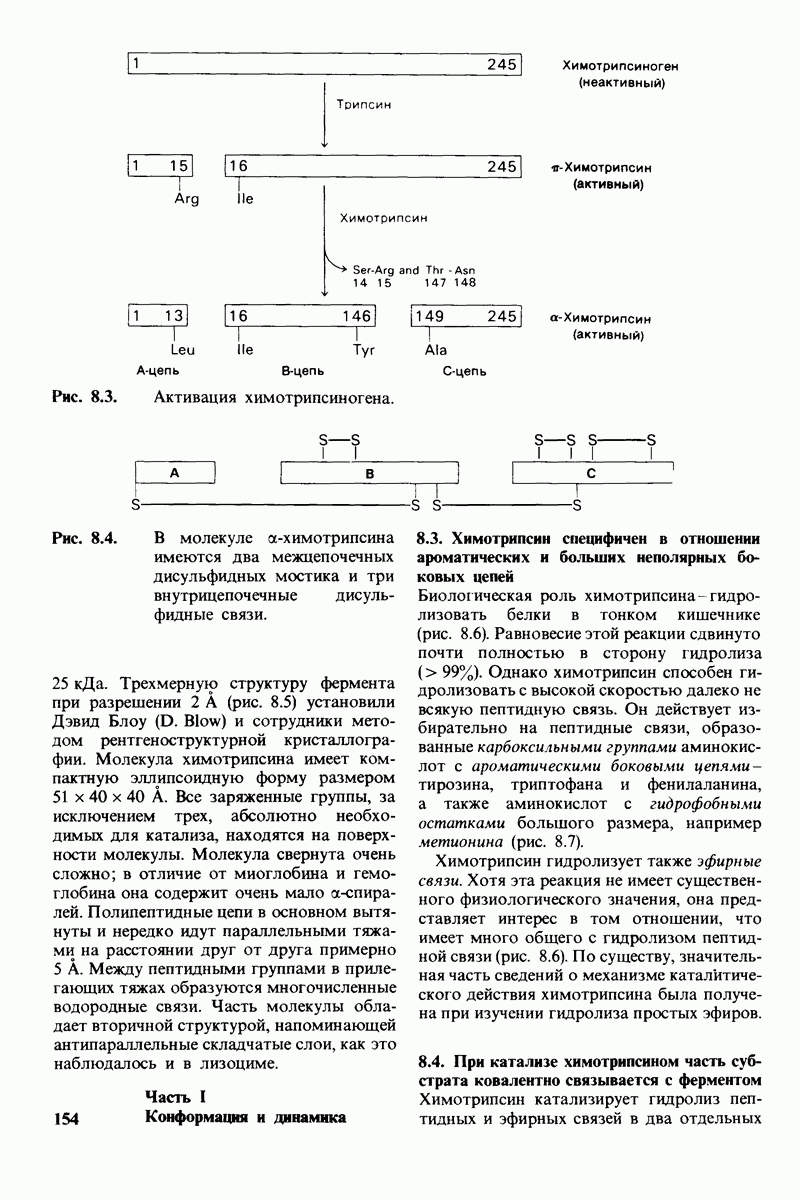
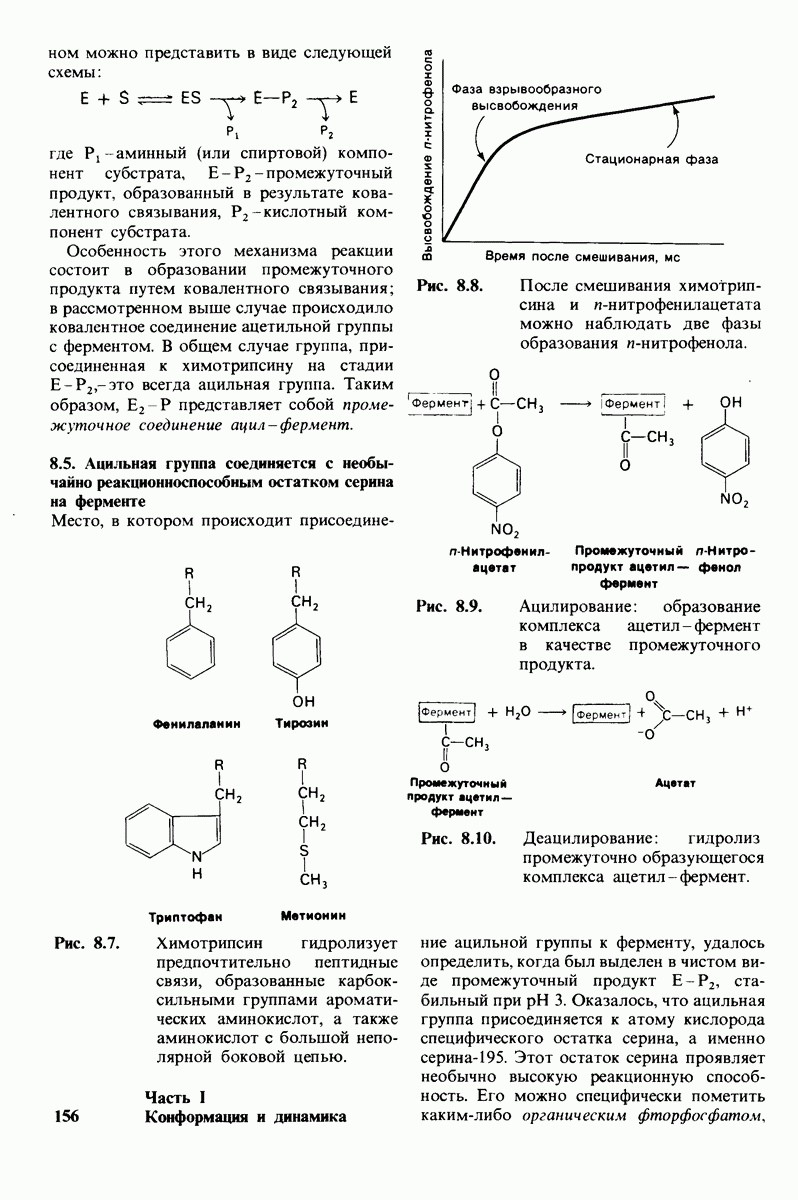
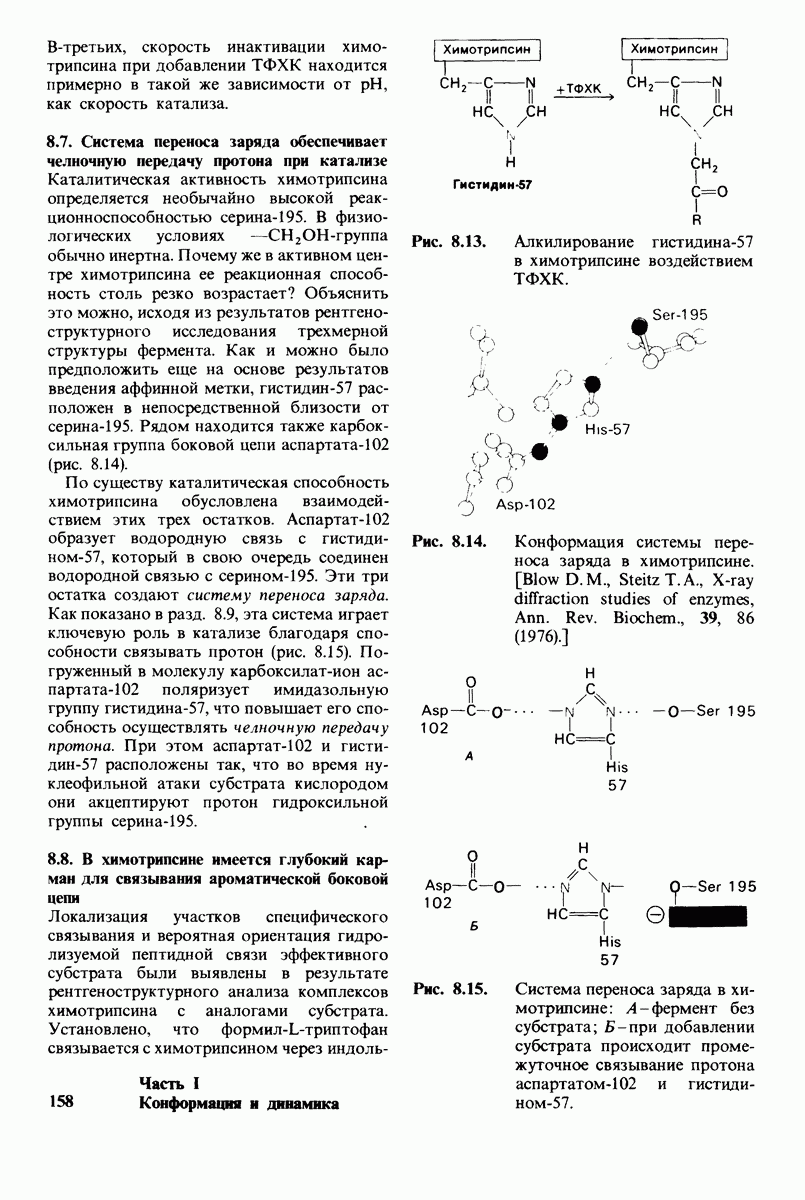
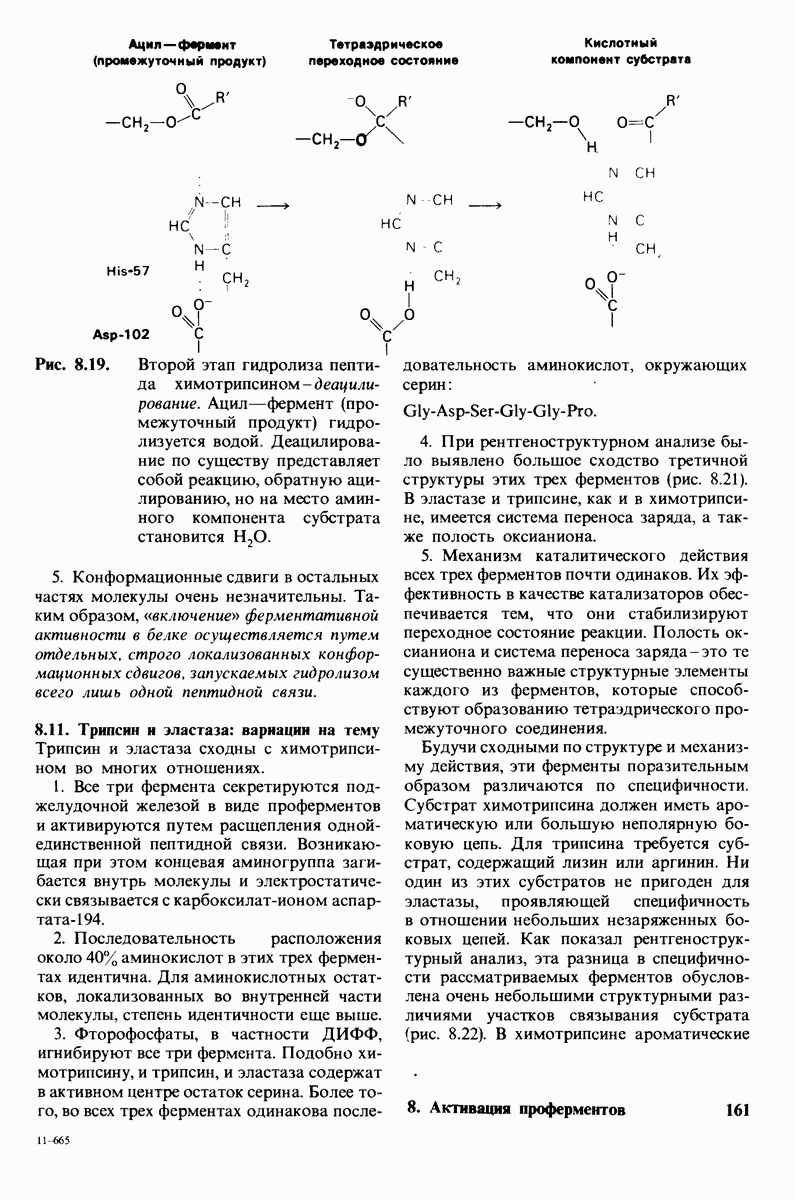
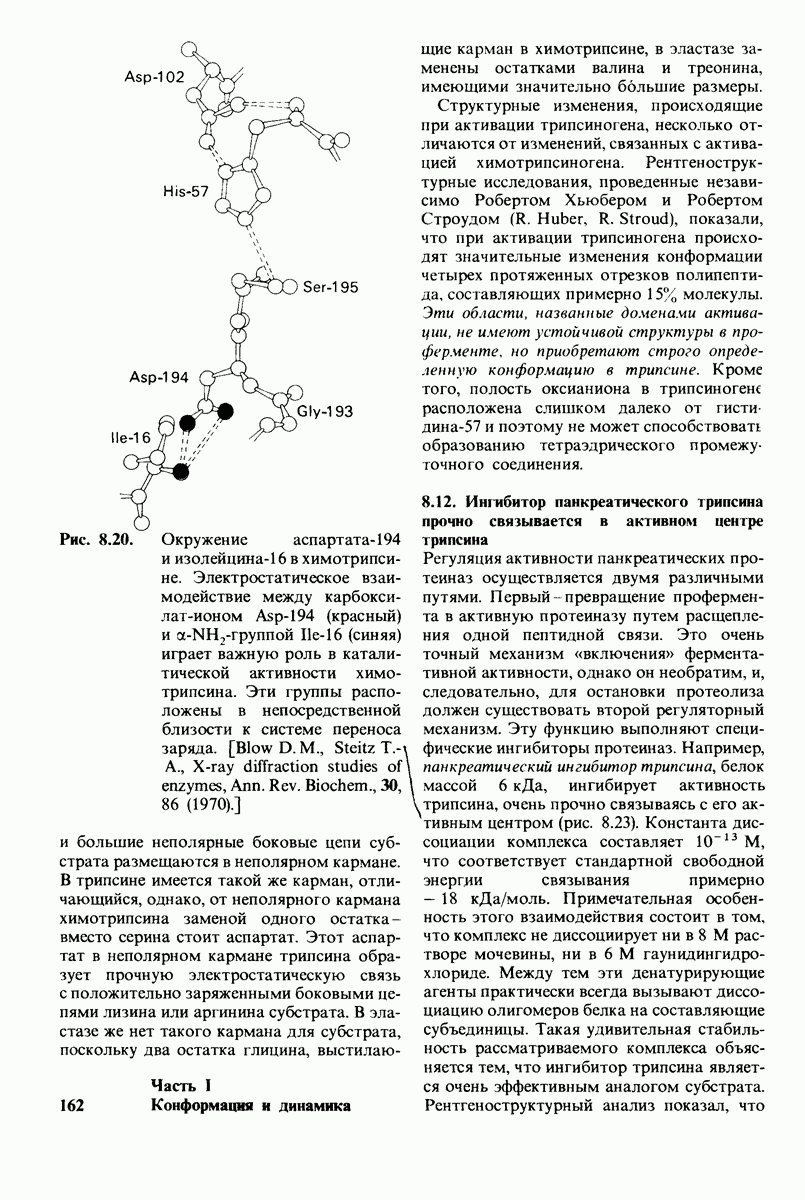
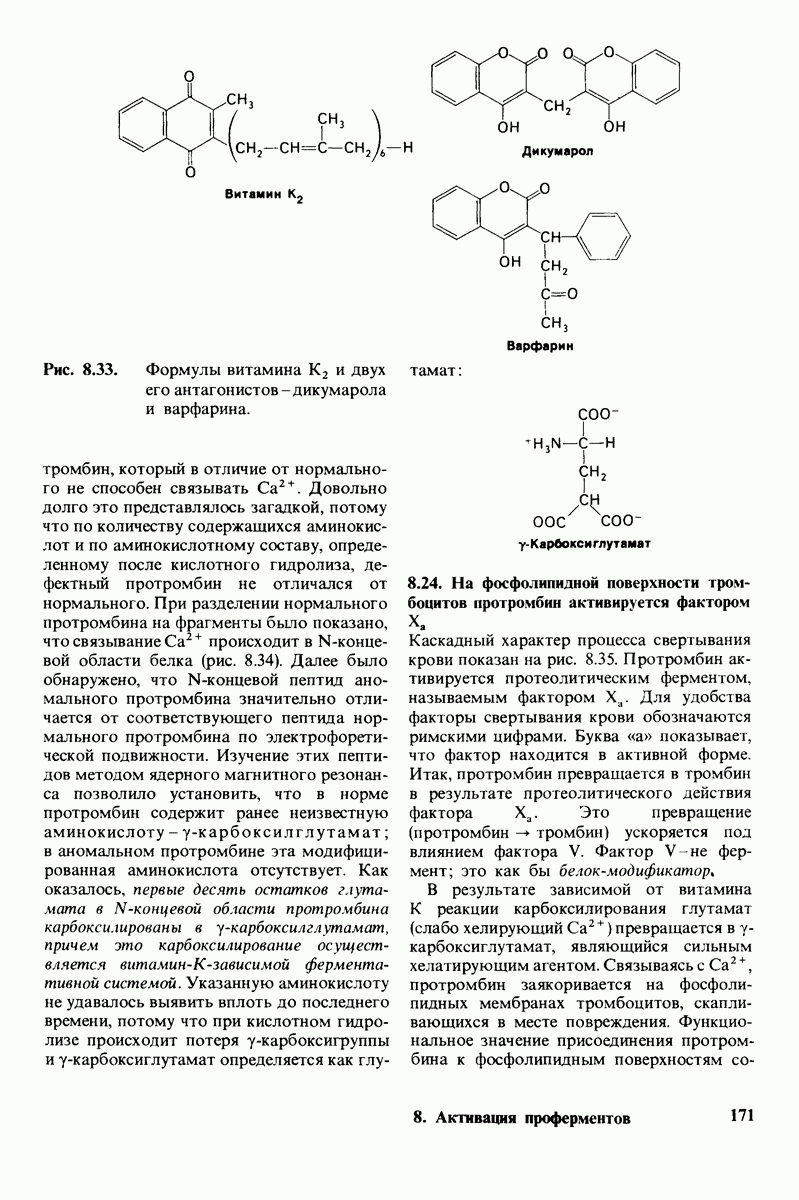
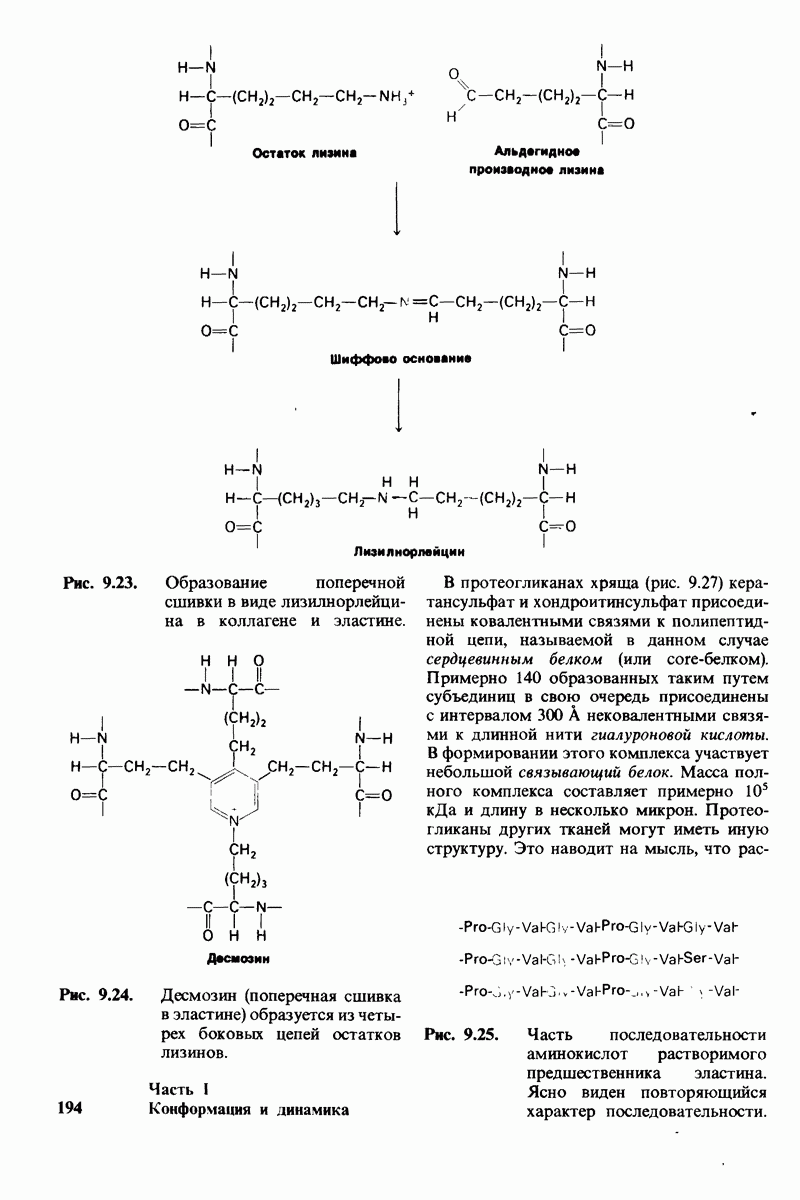
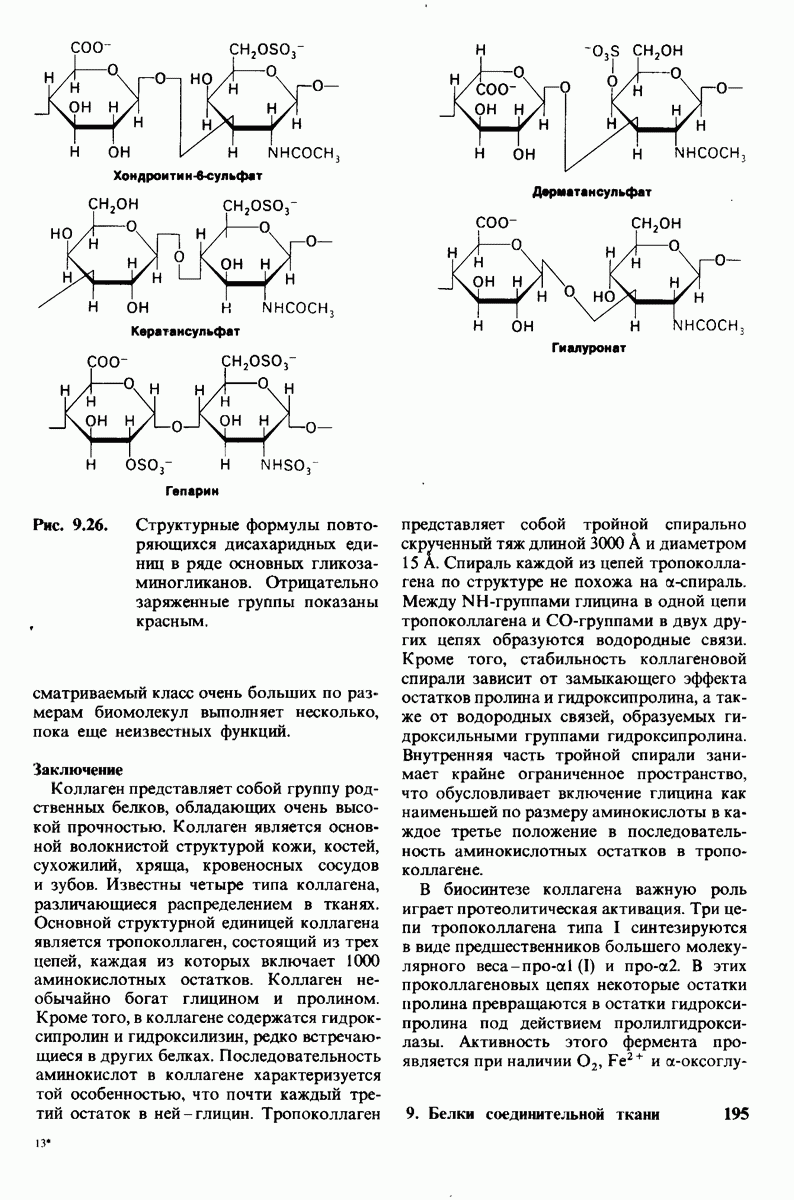
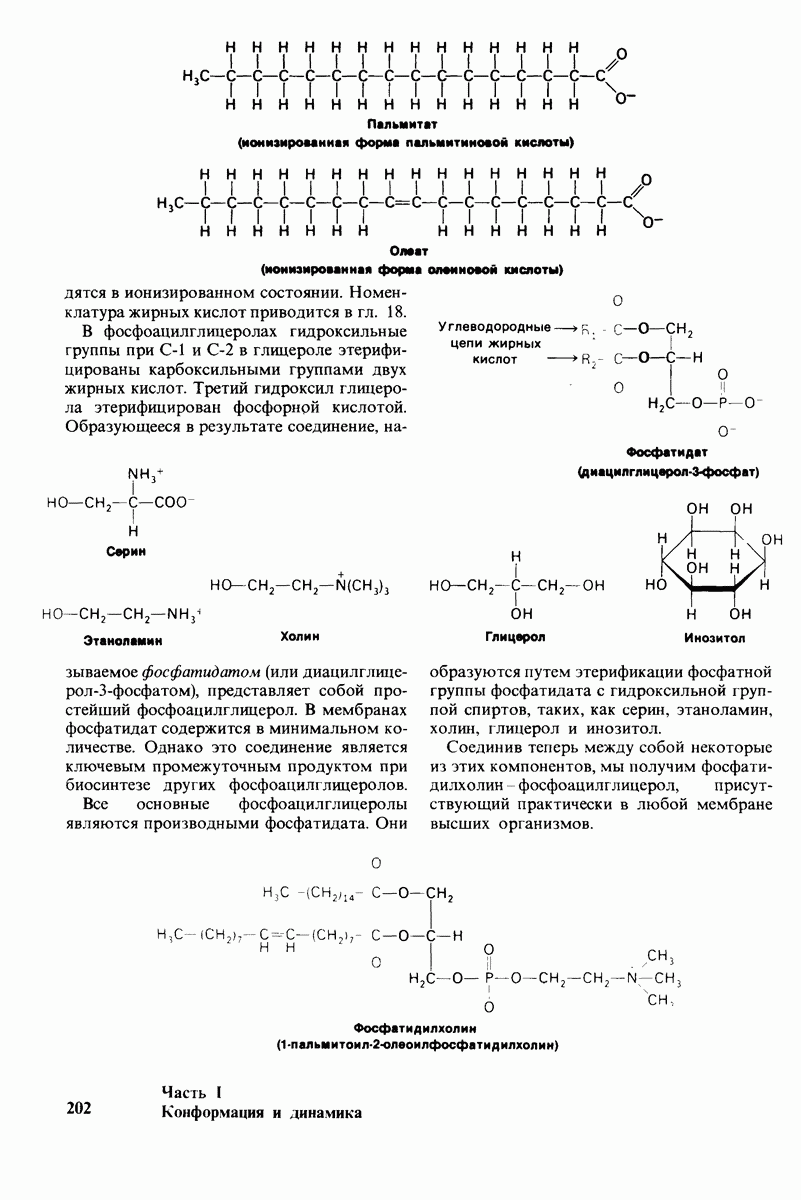
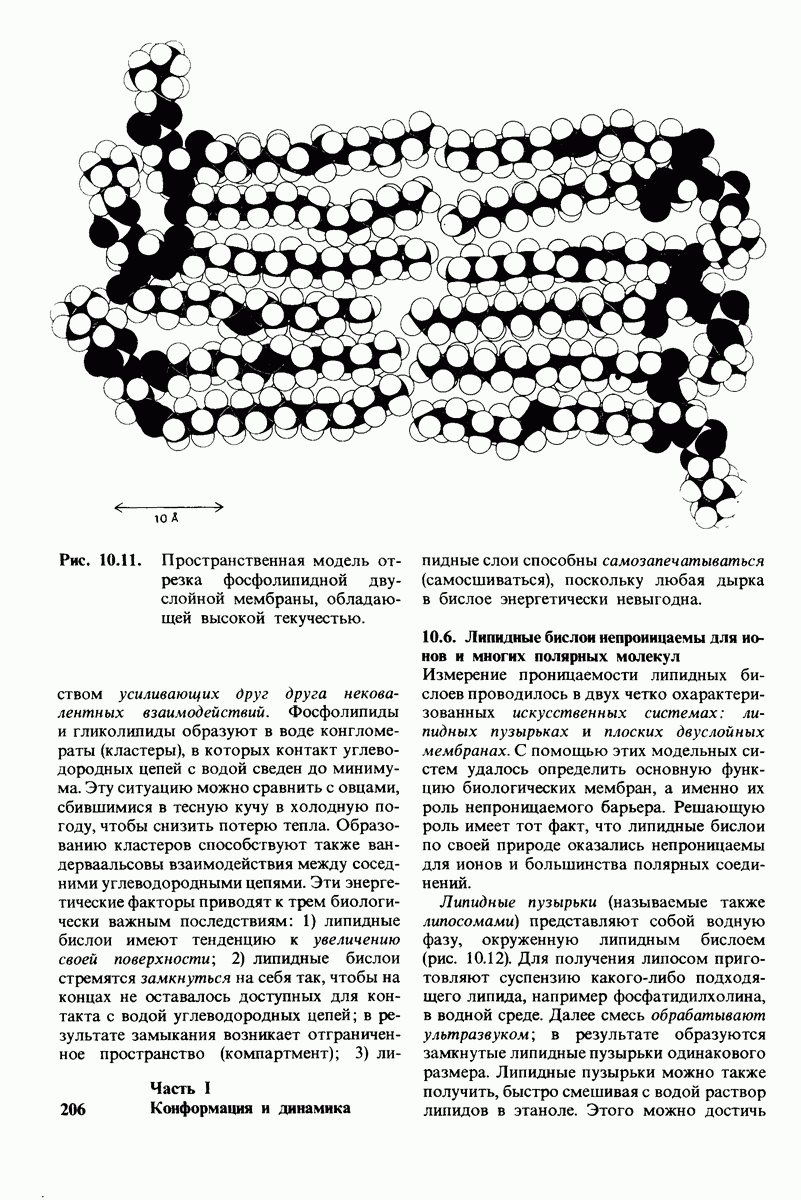
6.26. Гидрофобные взаимодействия; в водной среде неполярные группы стремятся ассоциировать
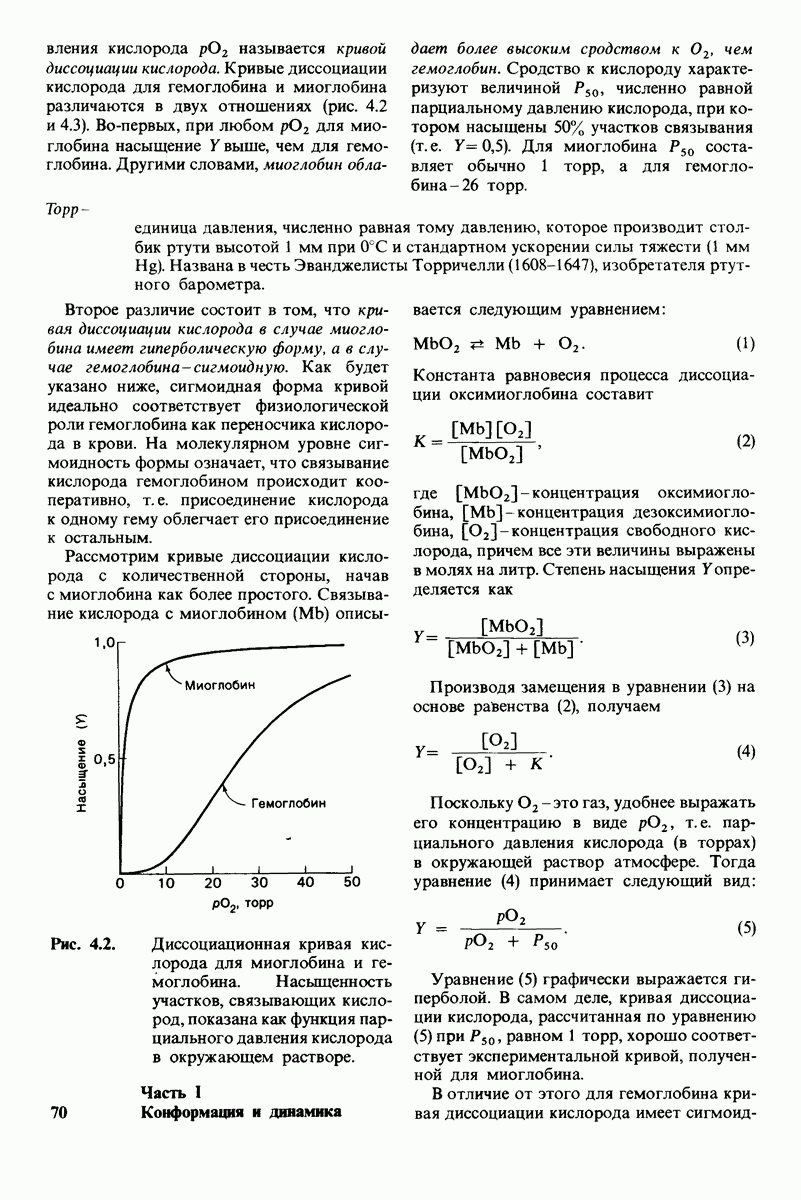
Каждый знает, как мелкие капельки масла в воде соединяются в одну большую каплю. Аналогичный процесс происходит и на уровне атомов, а именно неполярные молекулы или группы в водной среде объединяются в кластеры. Этот процесс ассоциации обусловлен гидрофобными взаимодействиями. Фигурально выражаясь, вода прижимает неиолярные соединения друг к другу.
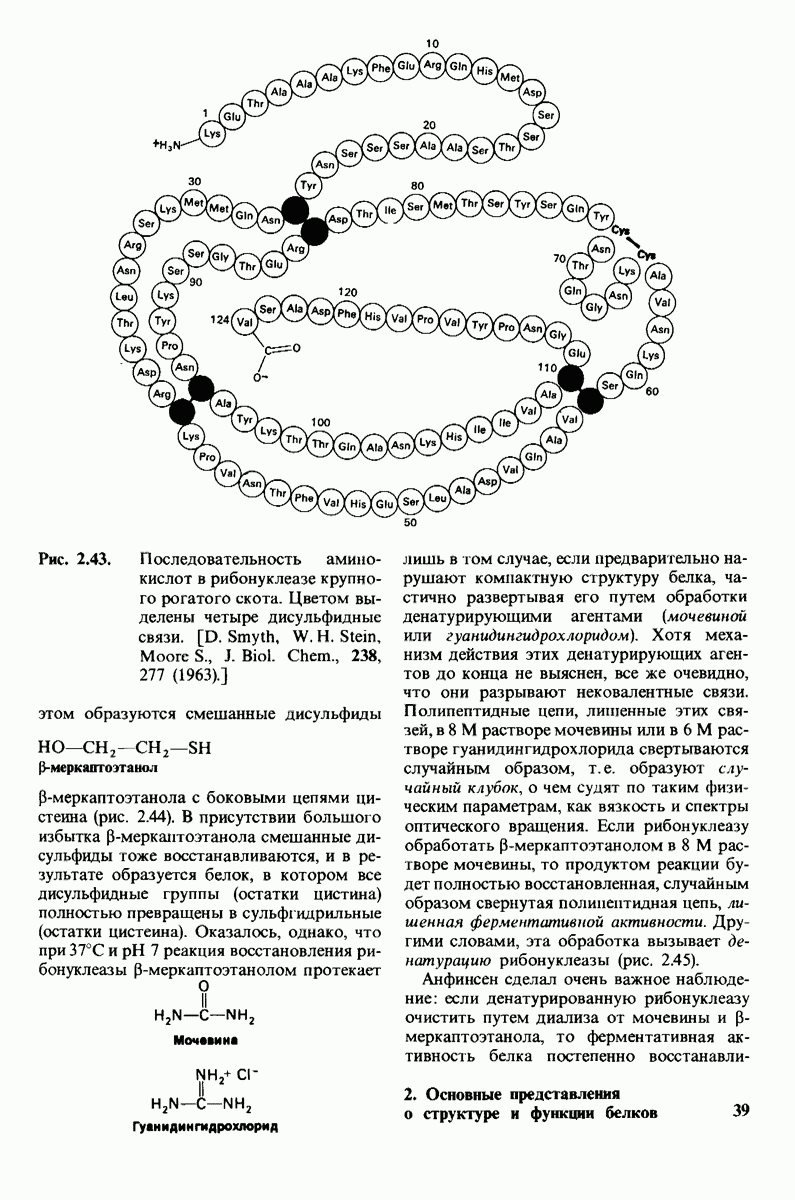
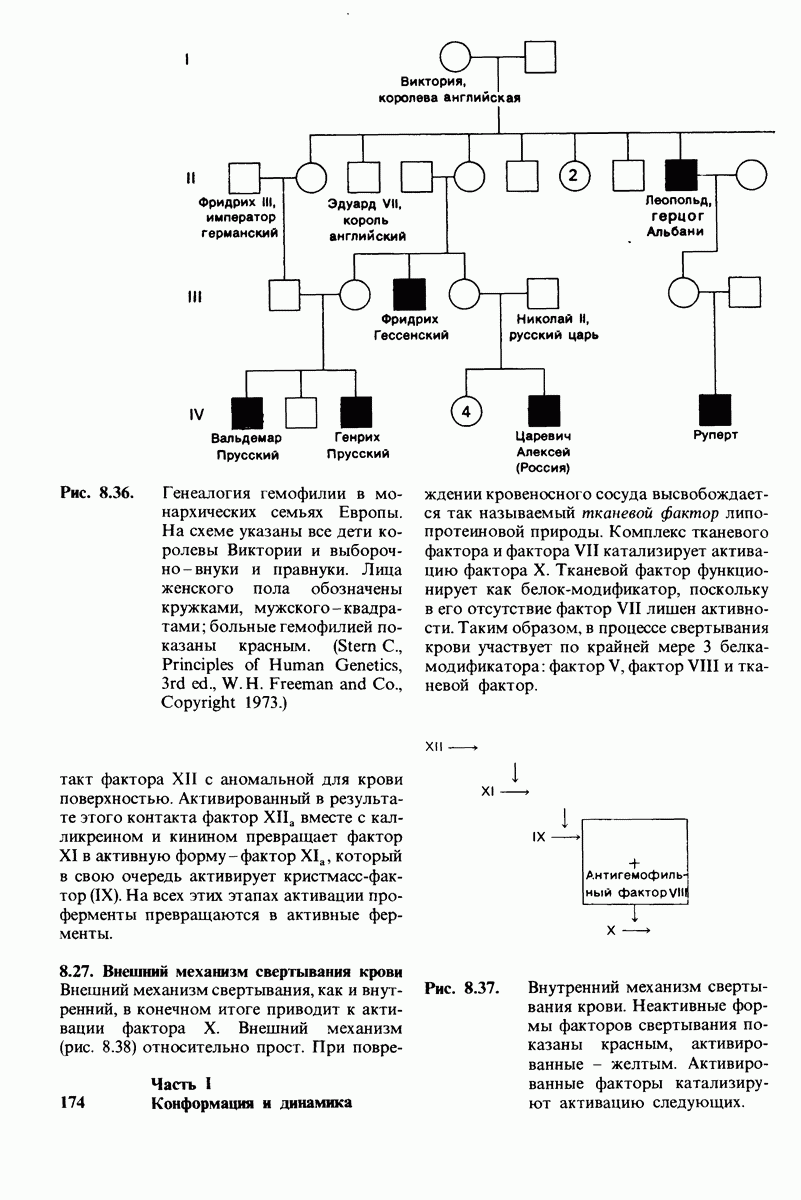
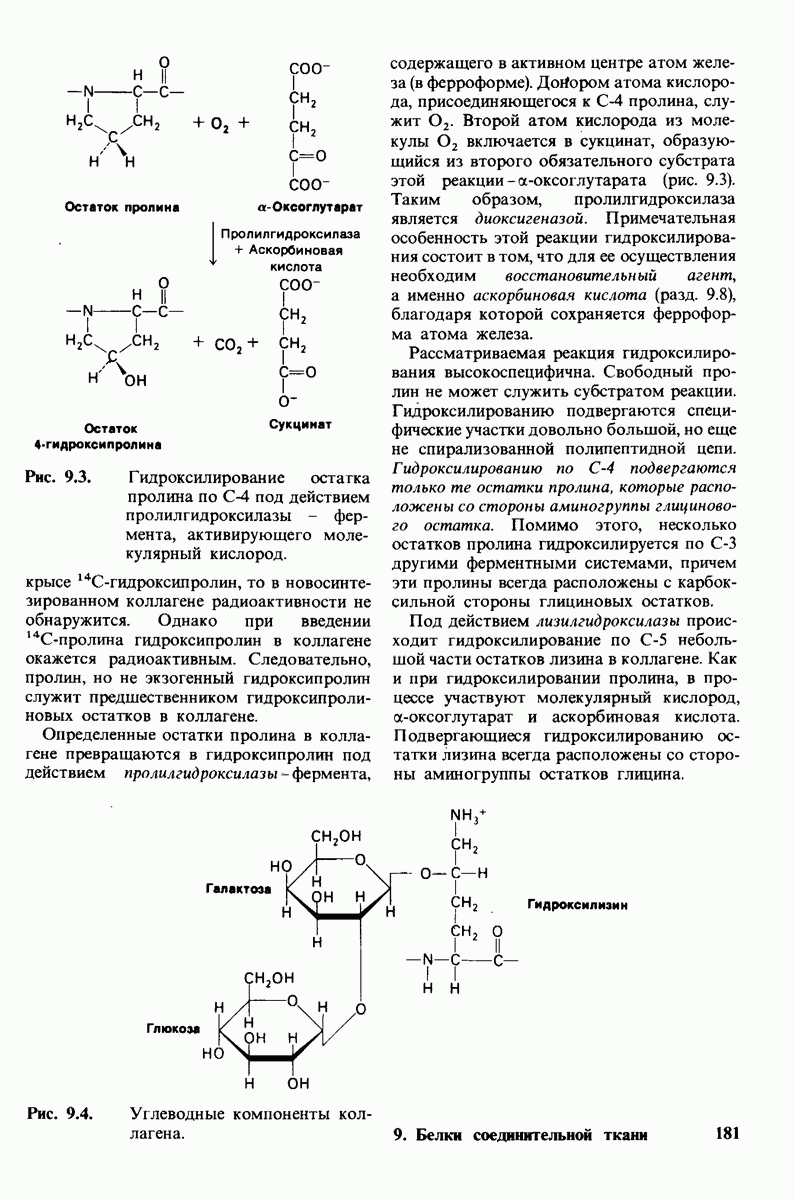
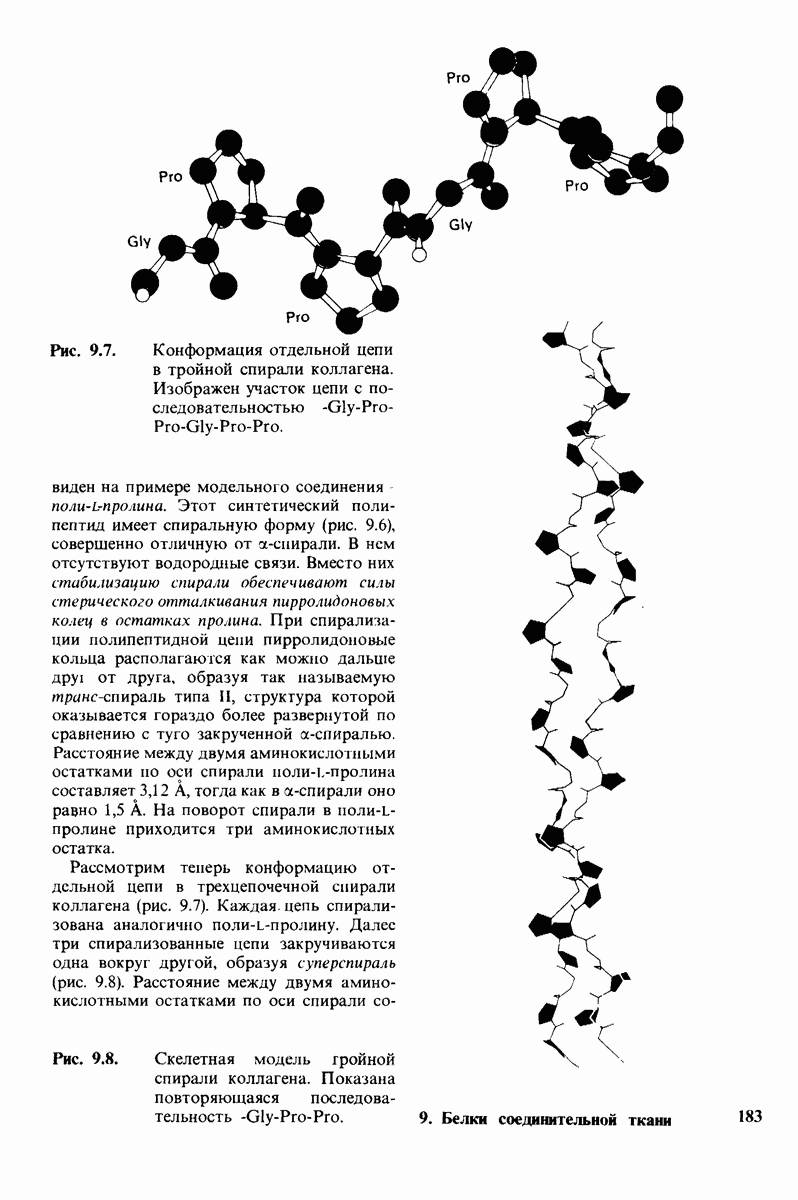
Гидрофобные взаимодействия играют решающую роль в складывании макромолекул, в связывании субстратов с ферментами и во многих других молекулярных процессах. Рассмотрим, что лежит в их основе. Допустим, что в воду вносится молекула неполярного соединения, например гексана. В воде при этом образуется полость, из-за чего на какой-то момент разрываются водородные связи между молекулами воды. Далее сдвинутые со своих мест молекулы воды переориентируются и образуют максимально возможное число новых водородных связей. Этого, однако, удается достичь лишь за определенную «плату»: возможностей образования благоприятных водородных связей в решетке воды вокруг молекул гексана значительно меньше, чем в чистой воде. В результате молекулы воды вокруг молекулы гексана располагаются гораздо более упорядоченным образом, чем в остальных частях раствора, а это означает, что энтропия раствора снижается. Посмотрим теперь, что произойдет, если в воде окажутся две молекулы гексана. Будут ли они находиться в двух малых полостях (рис. 6.31, А) или же в одной большой полости (рис. 6.31, Б)? Опыт показывает, что две молекулы гексана объединяются и занимают одну большую полость. Это объединение происходит в результате высвобождения отдельных ориентированных молекул воды, окружающих разделенные молекулы гексана. Следовательно, в основе гидрофобного взаимодействия лежит увеличение энтропии, обусловленное возрастанием степеней свободы высвобожденных молекул воды. Таким образом, неполярные молекулы в воде объединяются друг с другом не в силу высокого взаимного сродства, а прежде всего вследствие существования прочных связей между молекулами воды.
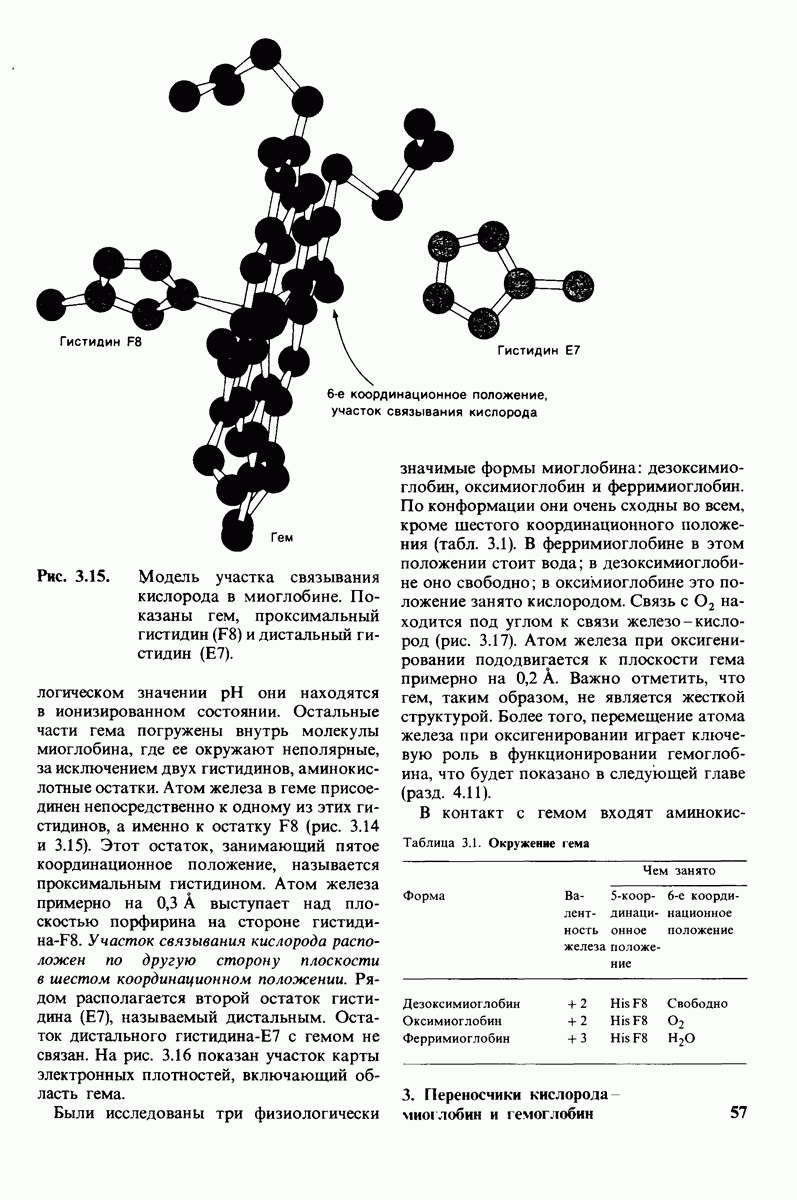
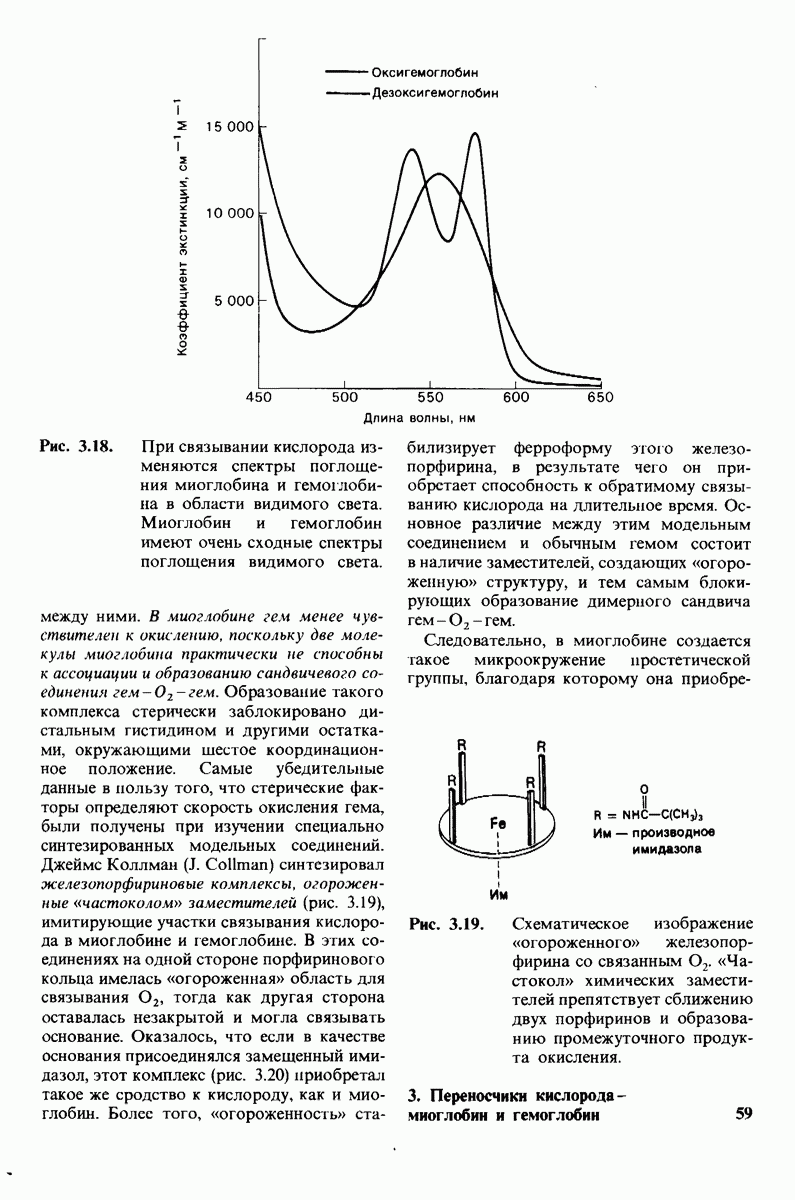
Заключение
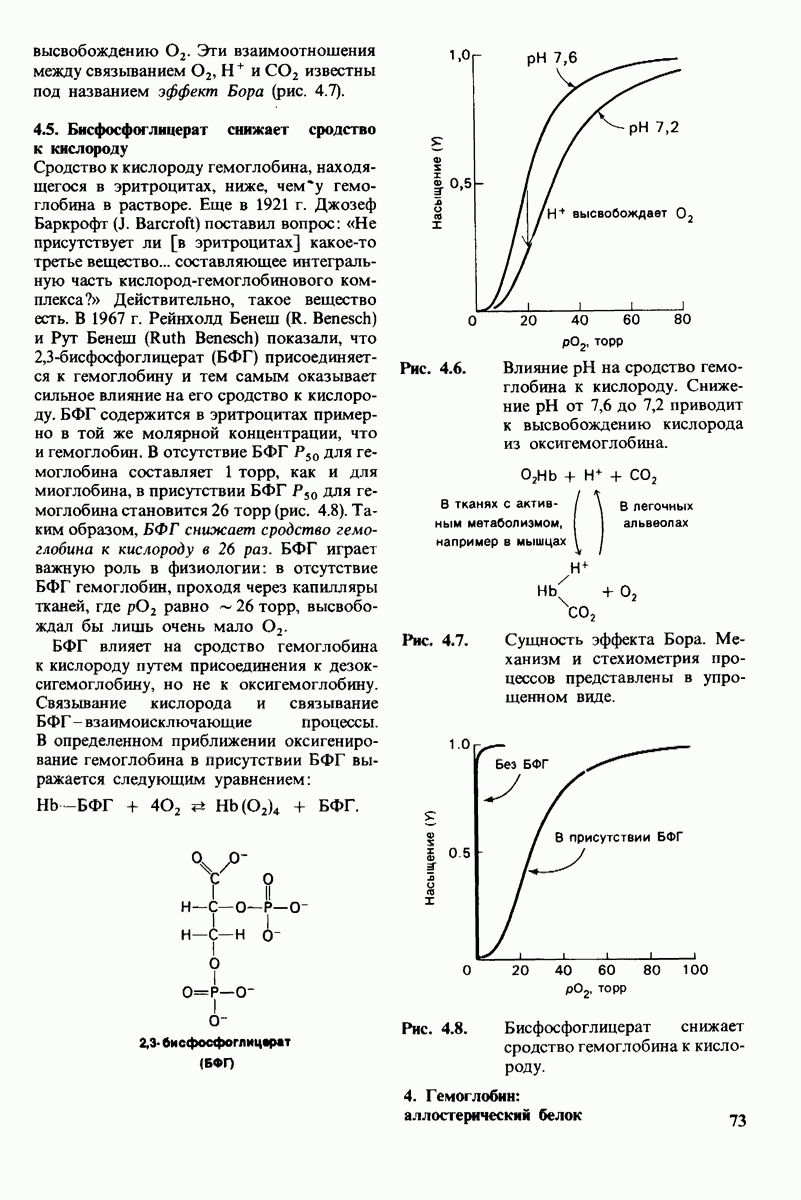
Катализаторами в биологических системах служат ферменты; все ферменты - белки. Ферменты высокоспецифичны и обладают огромной каталитической силой. Обычно они повышают скорость реакции по крайней мере в 107 раз, Ферменты не сдвигают равновесия реакции, а выполняют функцию катализаторов путем снижения энергии активации химических реакций. Кинетические
Таблица 6.5. (см. скан) Диэлектрические постоянные ряда растворителей

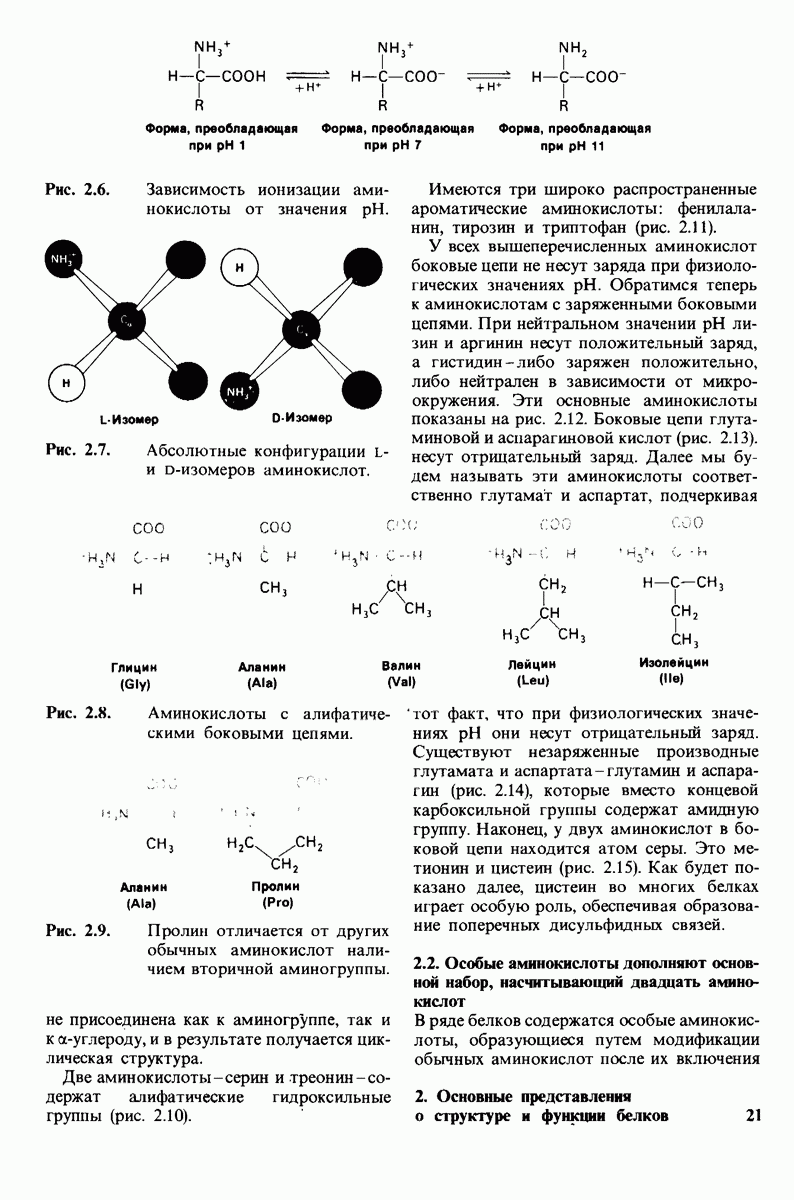
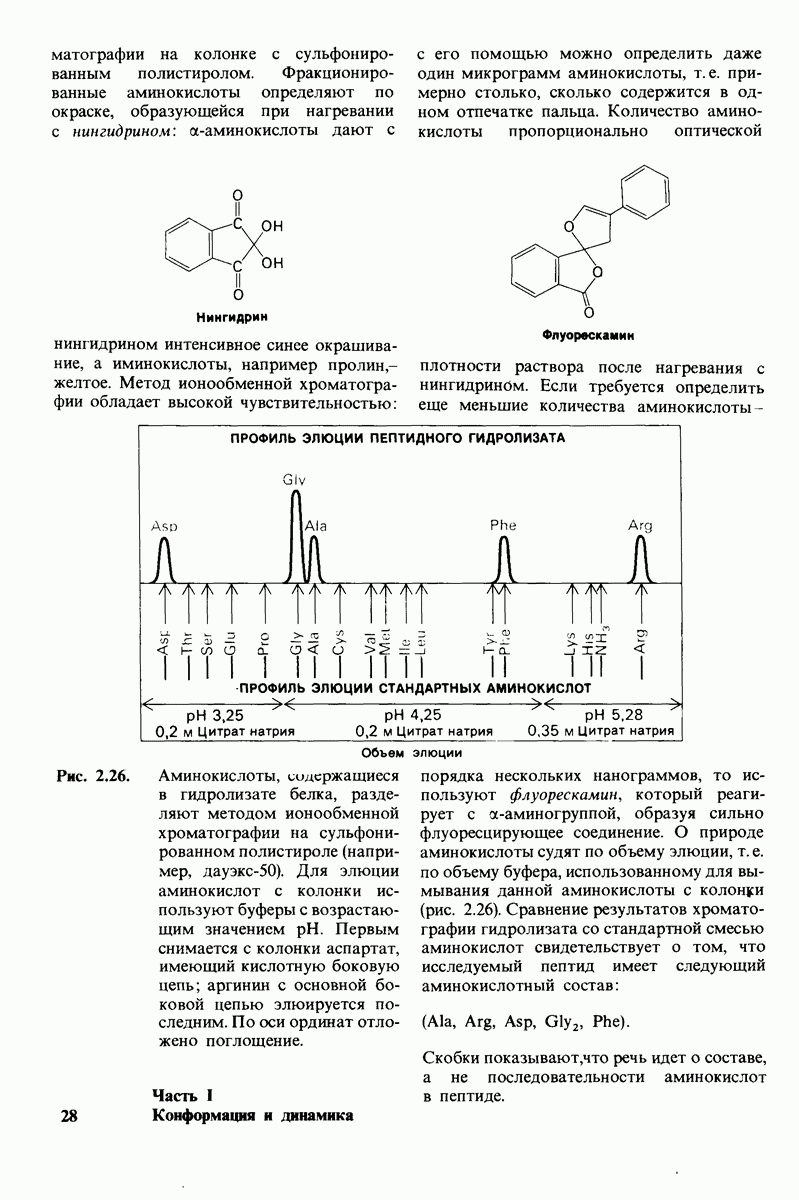
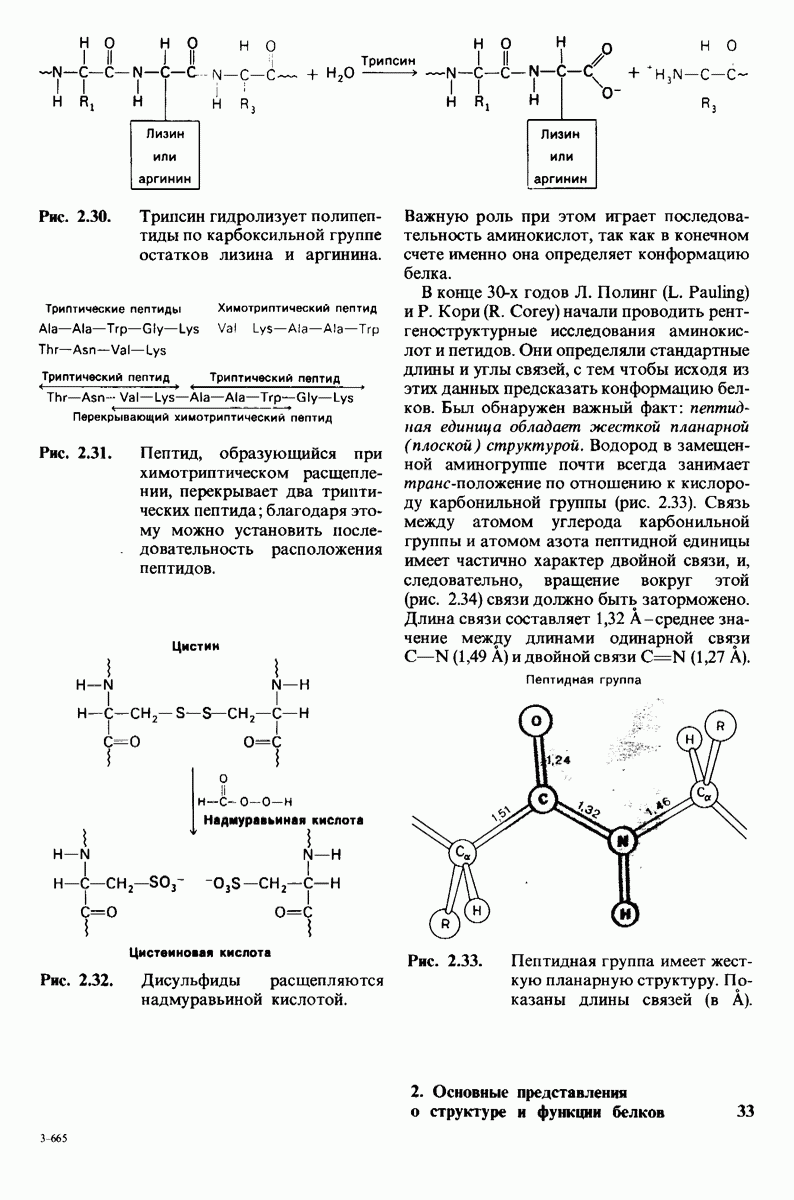
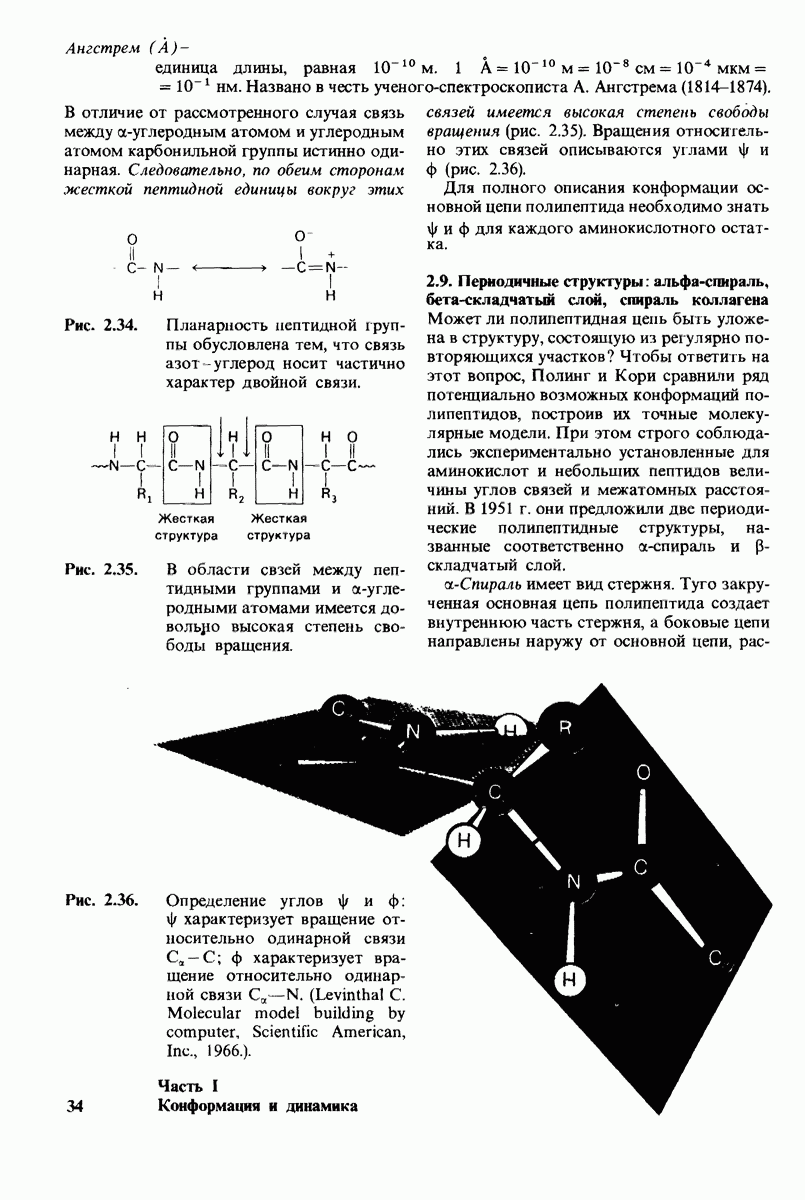
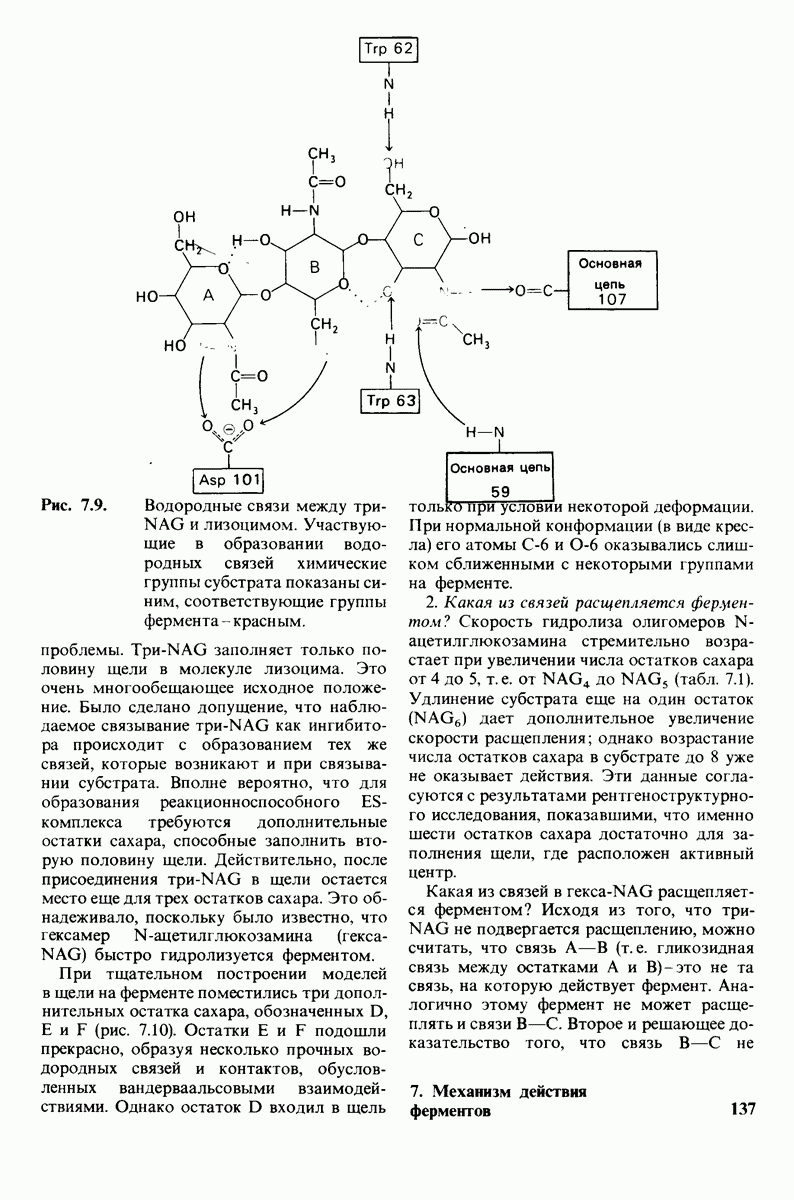
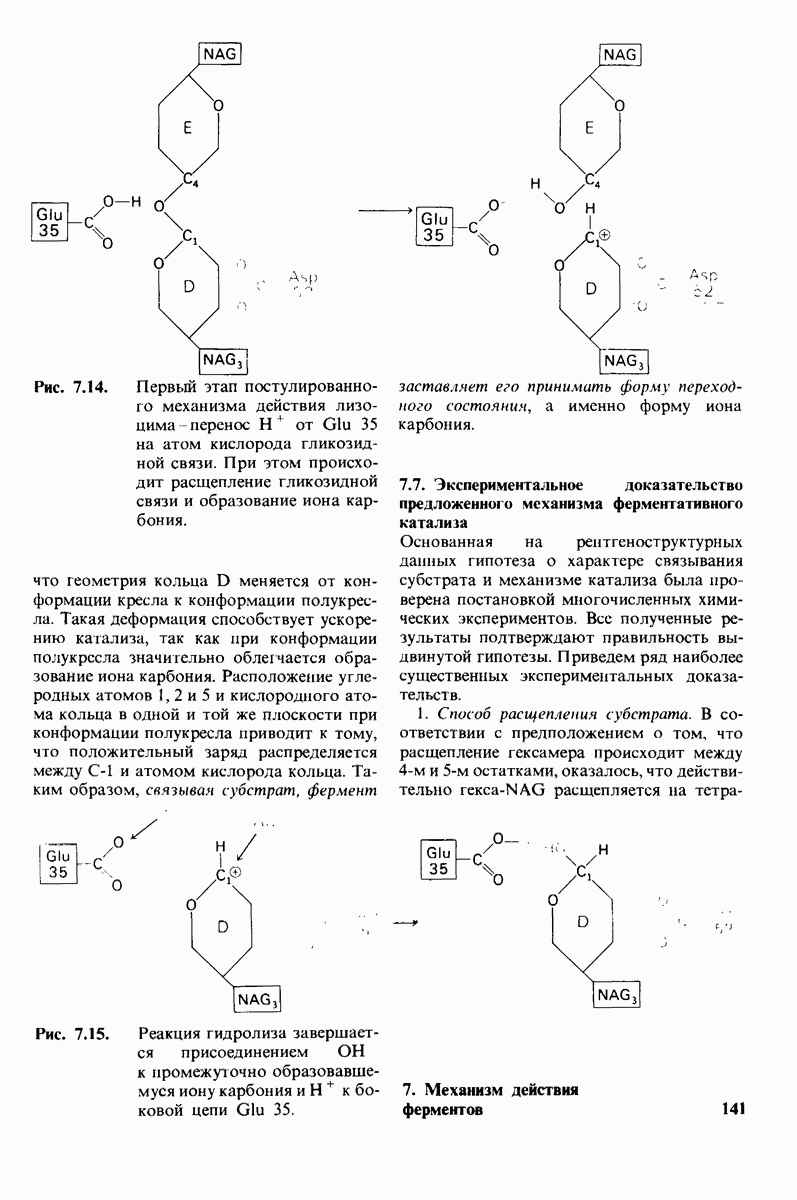
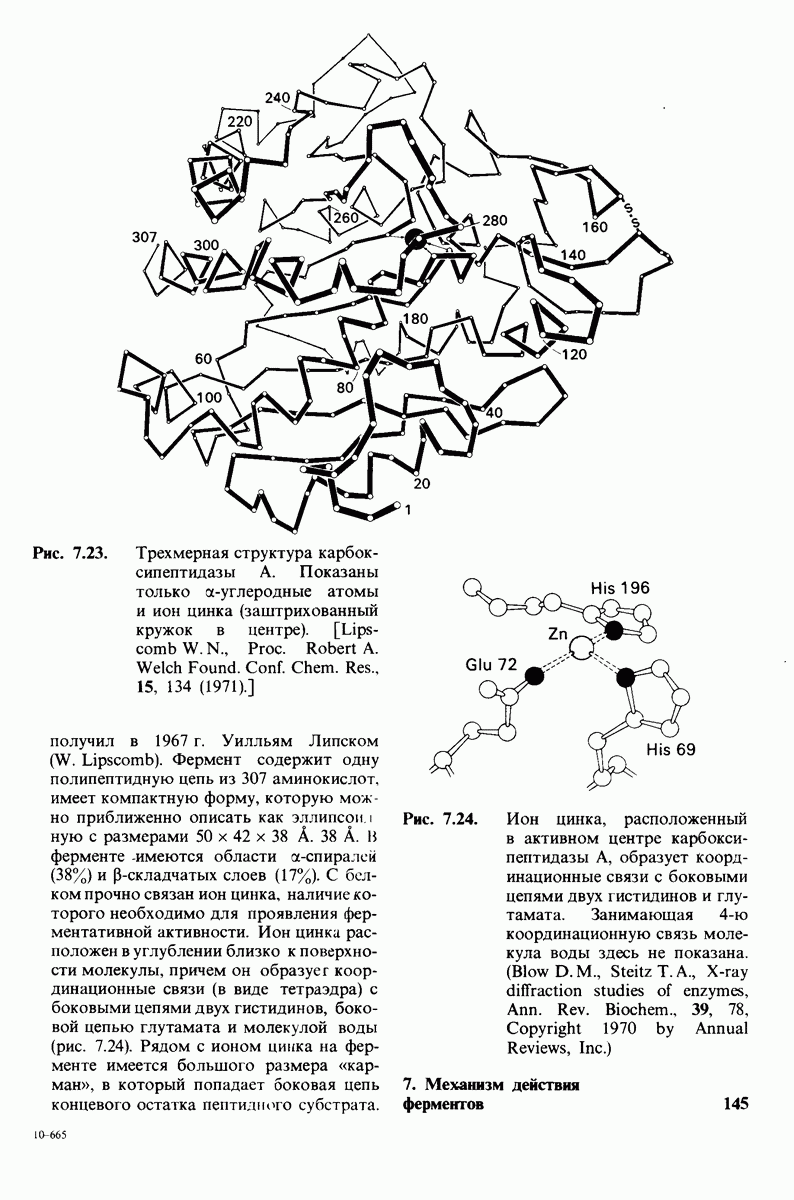
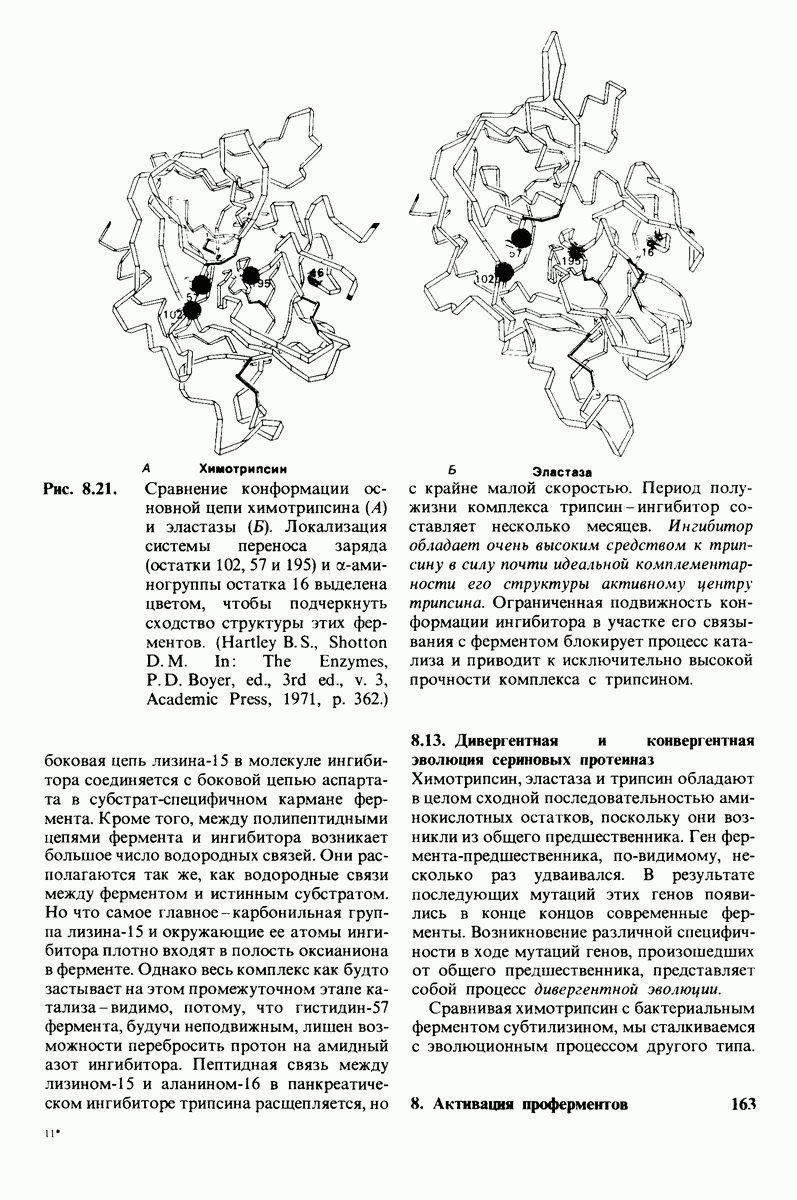
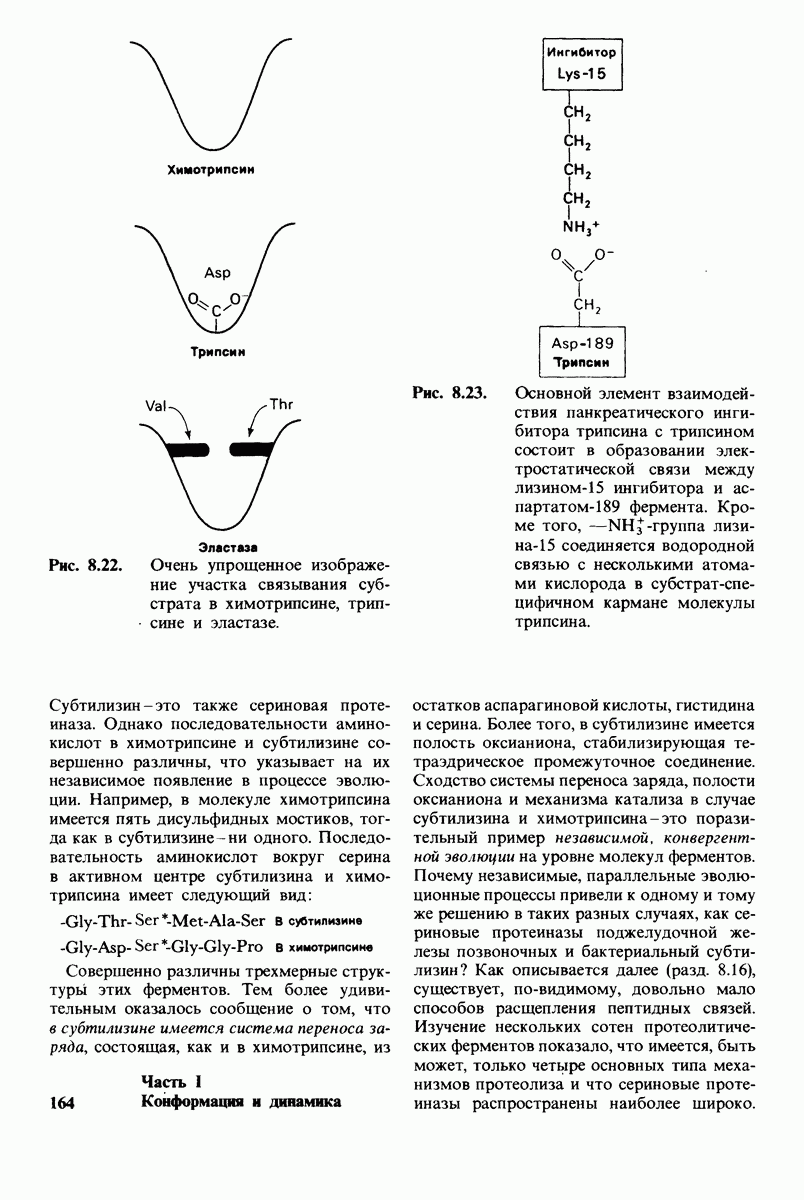
Рис. 6.31.
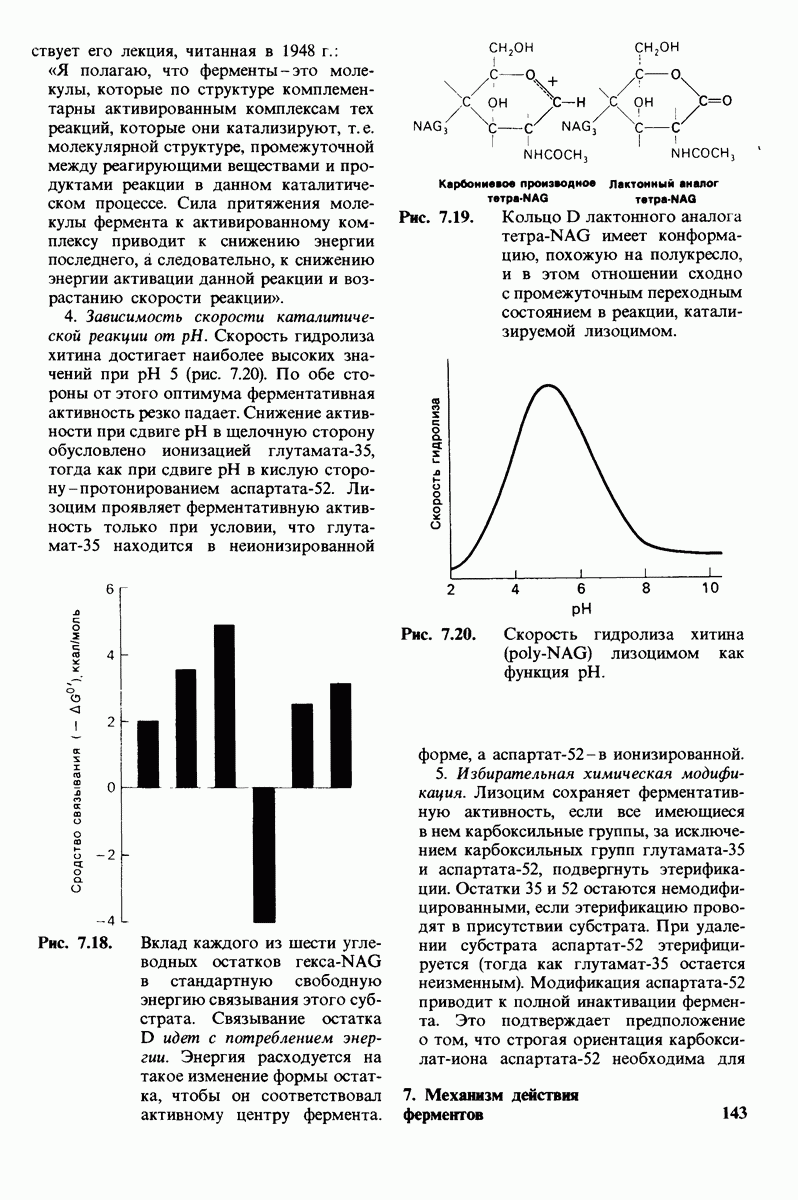
Схематическое изображение двух молекул гексана в небольшом объеме воды.  -молекулы гексана занимают различные полости в структуре воды.
-молекулы гексана занимают различные полости в структуре воды.  -молекулы гексана занимают одну общую полость, что энергетически более выгодно.
-молекулы гексана занимают одну общую полость, что энергетически более выгодно.
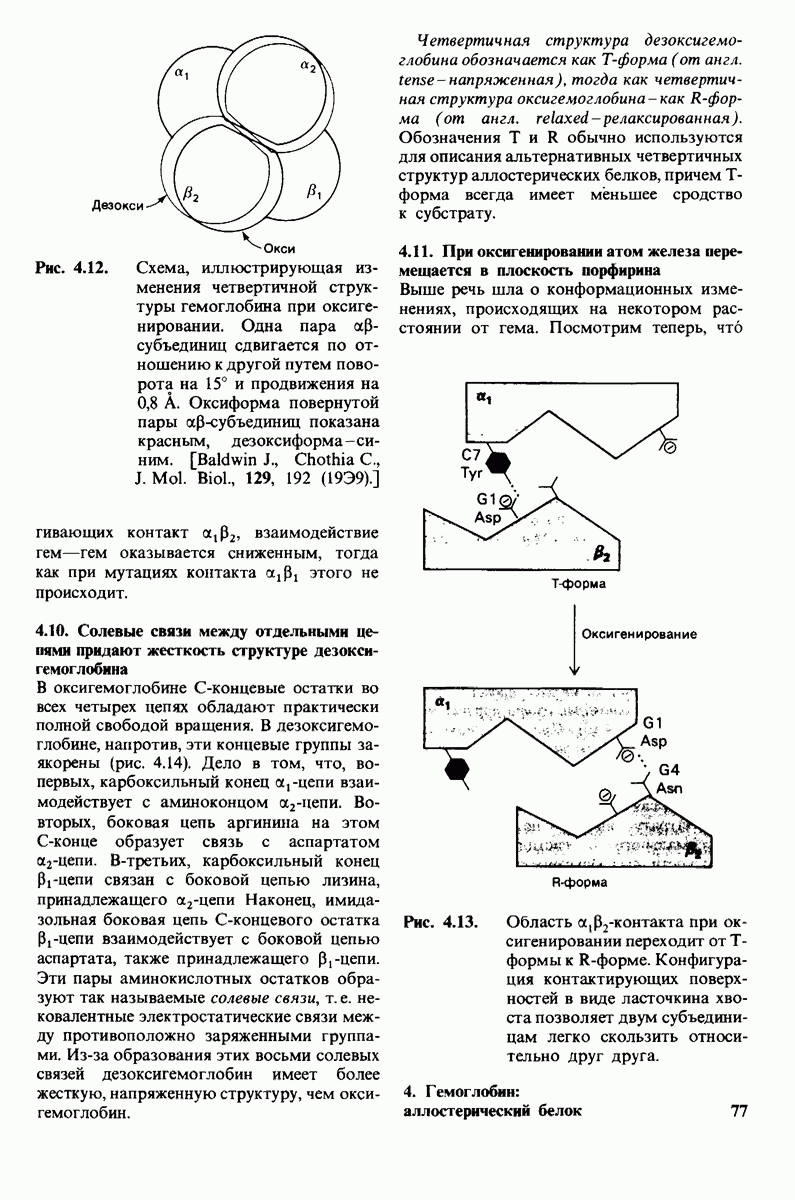
параметры некоторых ферментов описываются моделью Михаэлиса-Ментен. Согласно этой модели, фермент  соединяется с субстратом
соединяется с субстратом  образуя фермент-субстратный комплекс
образуя фермент-субстратный комплекс  который либо превращается далее в продукт
который либо превращается далее в продукт  реакции, либо диссоциирует на
реакции, либо диссоциирует на 

Скорость  образования продуктов описывается уравнением Михаэлиса-Ментен:
образования продуктов описывается уравнением Михаэлиса-Ментен:

где  скорость реакции при полном насыщении фермента субстратом, а
скорость реакции при полном насыщении фермента субстратом, а  константа Михаэлиса, равная концентрации субстрата, при которой скорость реакции составляет половину от максимальной. Максимальная скорость
константа Михаэлиса, равная концентрации субстрата, при которой скорость реакции составляет половину от максимальной. Максимальная скорость  равна произведению
равна произведению  на общую концентрацию фермента. Кинетическая константа называемая числом оборотов фермента, показывает, сколько молекул субстрата превратилось в продукт реакции за единицу времени в одном каталитическом центре при полном насыщении фермента субстратом. Для большинства ферментов число оборотов лежит в пределах от 1 до 104 в 1 с.
на общую концентрацию фермента. Кинетическая константа называемая числом оборотов фермента, показывает, сколько молекул субстрата превратилось в продукт реакции за единицу времени в одном каталитическом центре при полном насыщении фермента субстратом. Для большинства ферментов число оборотов лежит в пределах от 1 до 104 в 1 с.
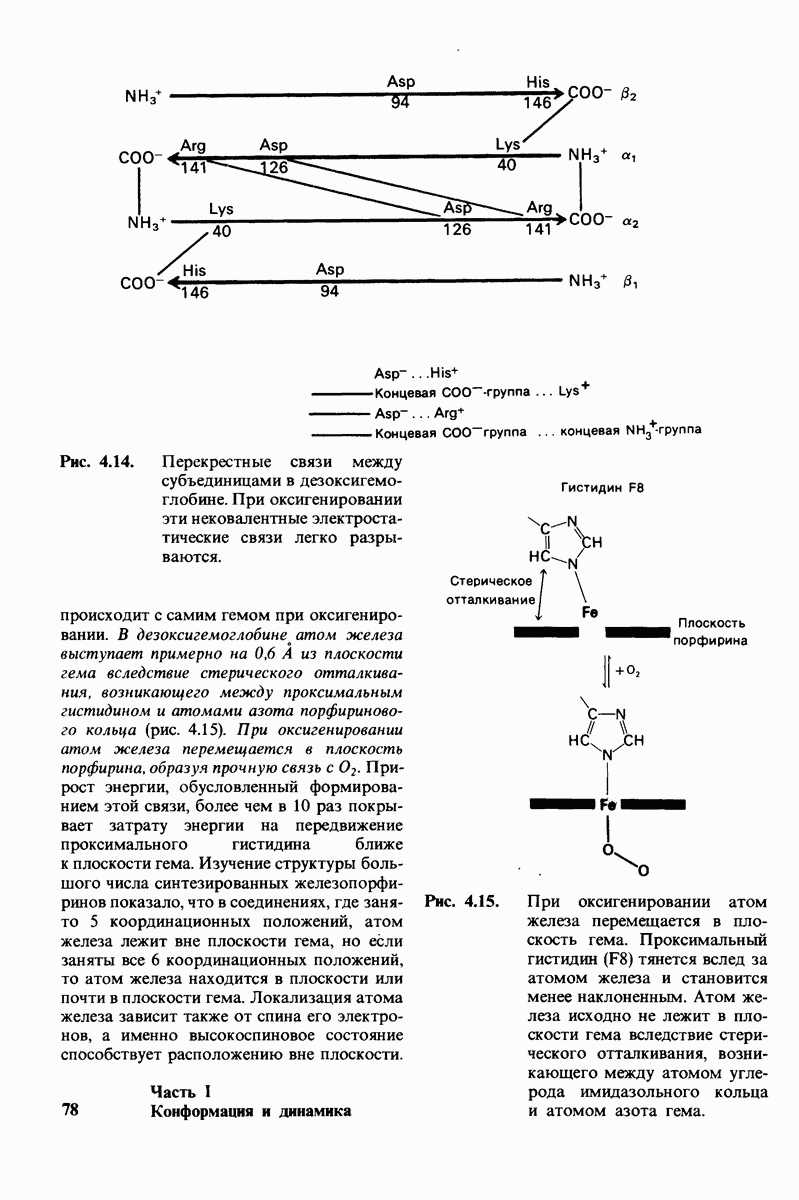
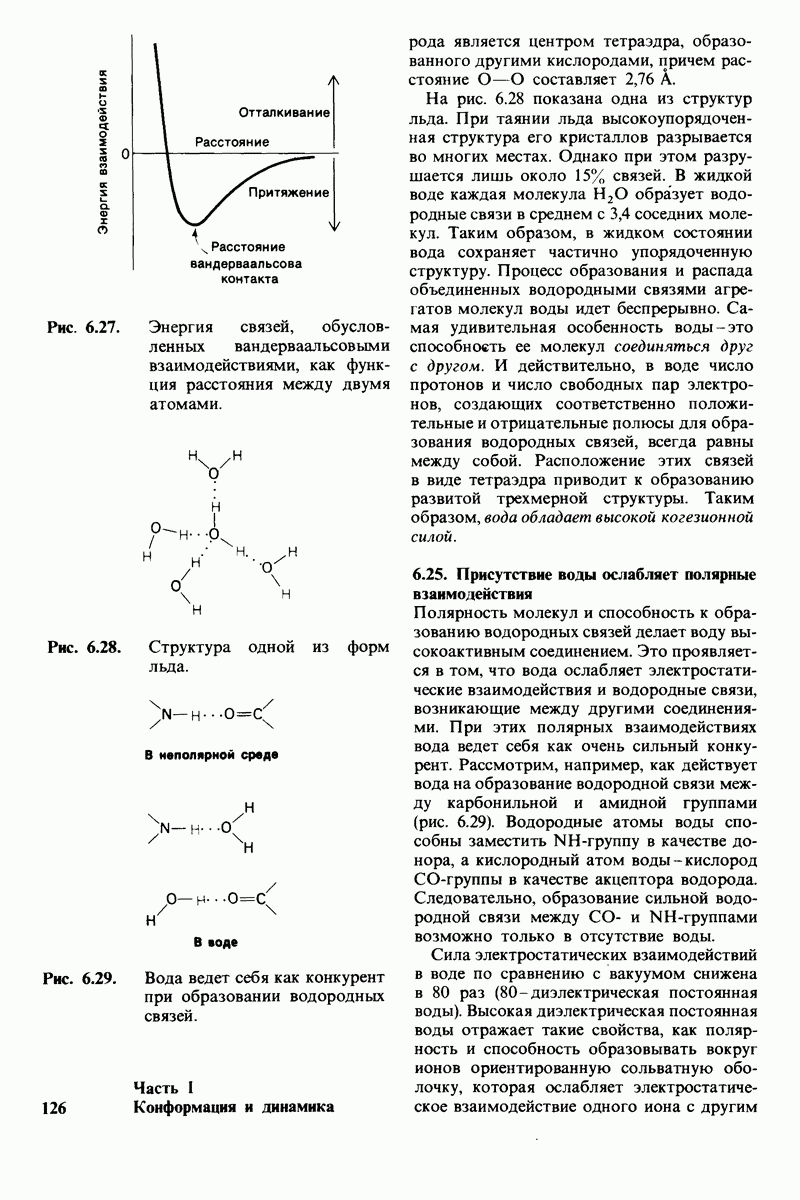
Некоторые специфические низкомолекулярные вещества и ионы способны ингибировать ферменты. При необратимом ингибировании ингибитор ковалентно соединяется с ферментом или же связывается с ним настолько прочно, что его диссоциация идет очень медленно. В отличие от этого обратимое ингибирование характеризуется тем, что равновесие между ферментом и ингибитором устанавливается быстро. Конкурентный ингибитор препятствует связыванию субстрата в активном центре. Он уменьшает скорость реакции путем снижения относительного количества молекул фермента, связавших субстрат. При неконкурентном ингибировании ингибитор снижает число оборотов фермента. Конкурентное ингибирование в отличие от неконкурентного снимается при повышении концентрации субстрата; таким путем можно различить эти два вида ингибирования.
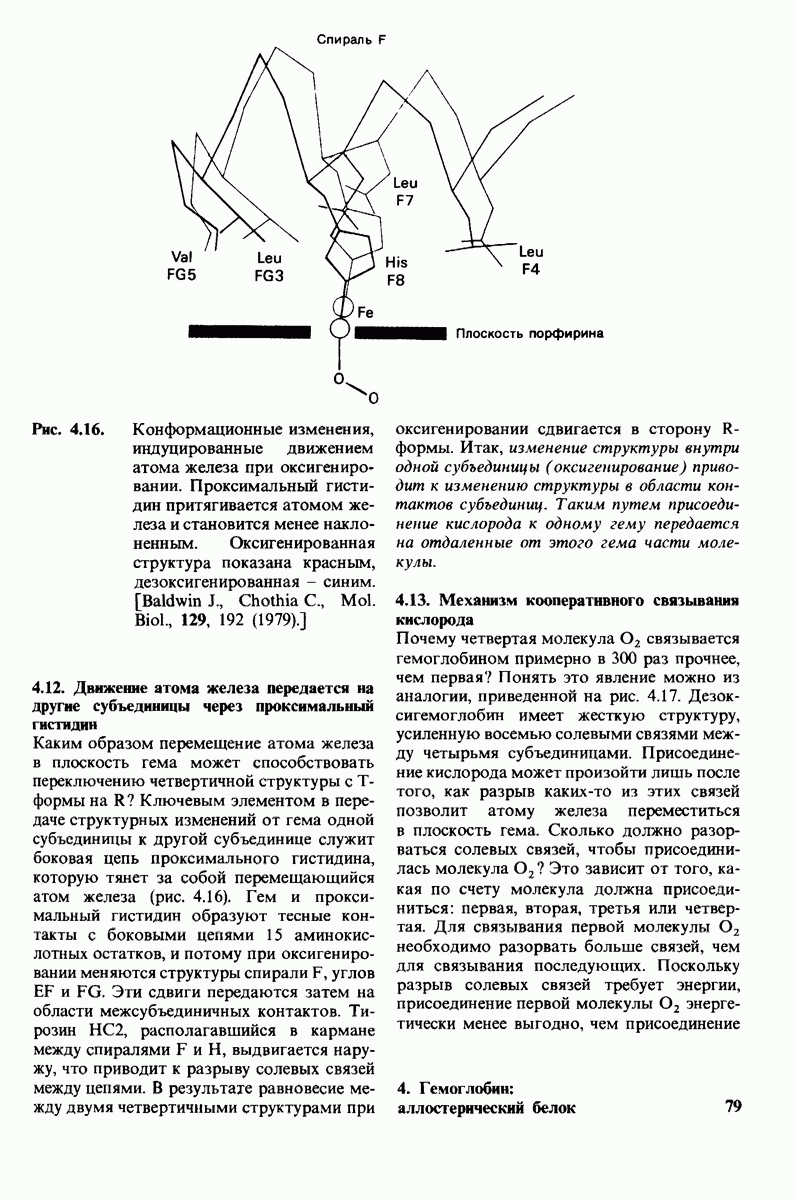
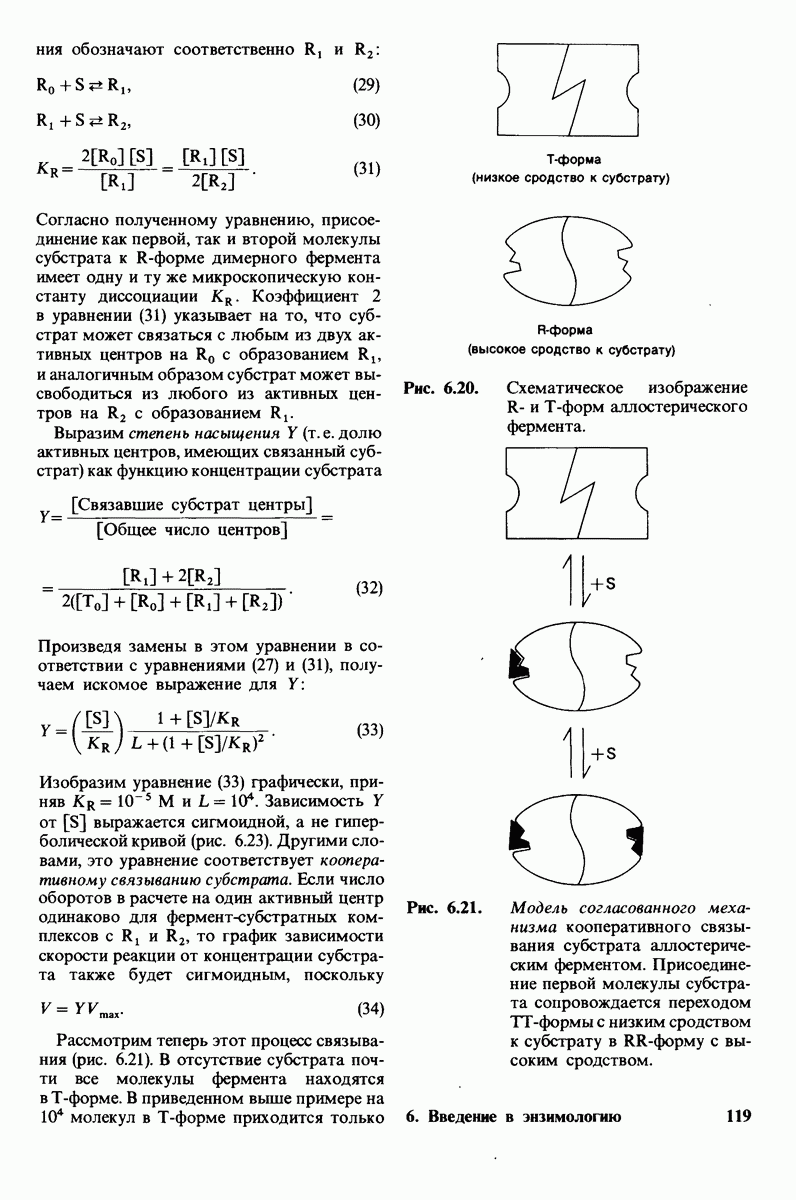
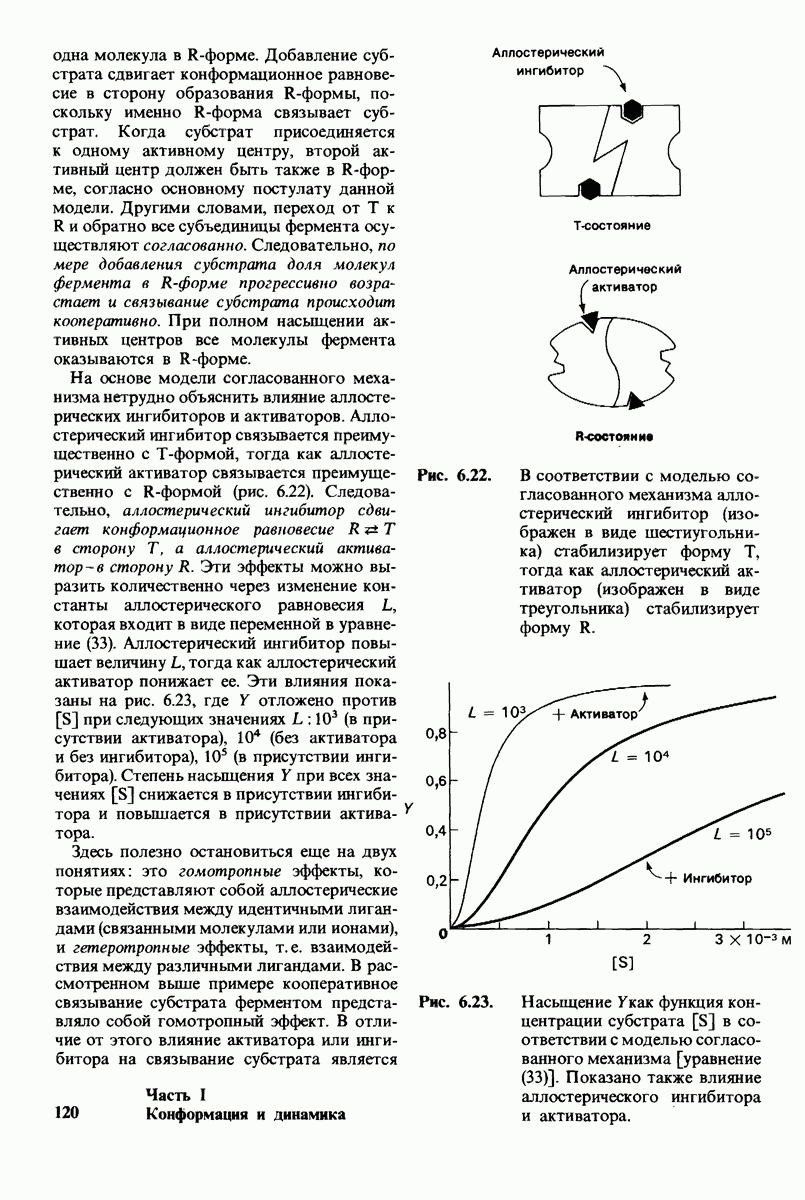
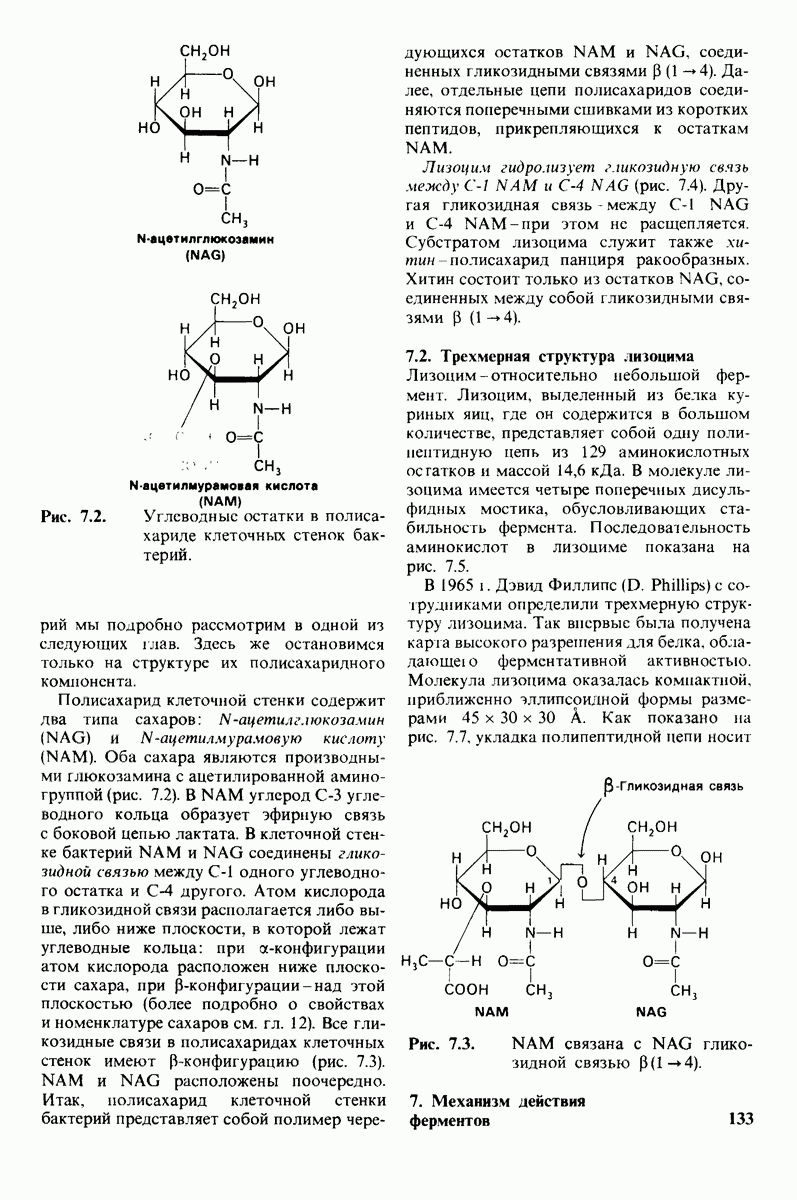
Каталитическая активность многих ферментов подвержена регуляции in vivo. В этом отношении особенно важную роль играют аллостерические взаимодействия, т. е. взаимодействия между пространственно разделенными участками фермента. Все известные в настоящее время аллостерические ферменты состоят из двух и более субъединиц. Аллостерические взаимодействия опосредованы конформационными изменениями, которые передаются с одной субъединицы на другую. Кривая зависимости скорости реакции V от концентрации субстрата  для аллостерических ферментов имеет сигмоидную, а не гиперболическую форму. Для объяснения некоторых свойств этих ферментов предложены две модели - модель согласованного механизма и модель последовательного механизма.
для аллостерических ферментов имеет сигмоидную, а не гиперболическую форму. Для объяснения некоторых свойств этих ферментов предложены две модели - модель согласованного механизма и модель последовательного механизма.
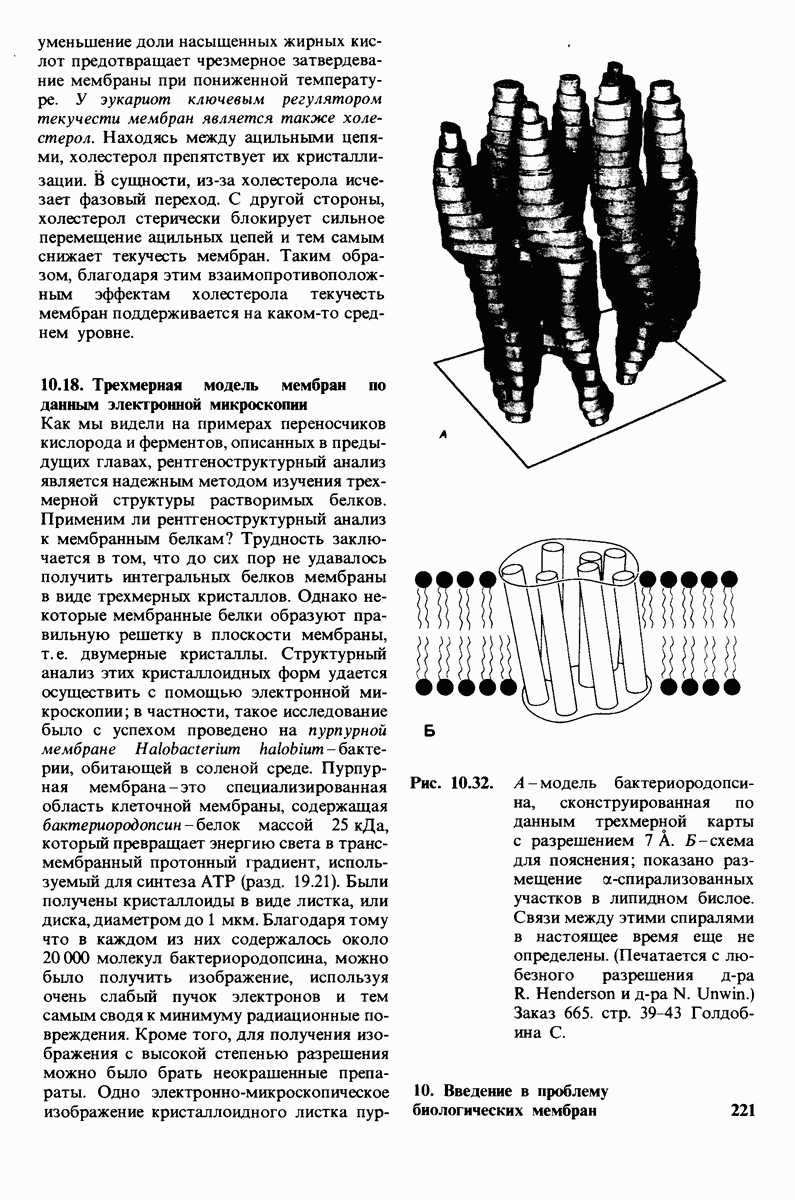
Обратимые молекулярные взаимодействия в биологических системах обусловлены возникновением водородных связей, а также электростатических и вандерваальсовых взаимодействий. Сильнейшее влияние на эти взаимодействия оказывает вода благодаря таким своим свойствам, как полярность, когезионная сила и способность к образованию водородных связей в качестве и донора, и акцептора водорода. В присутствии воды ослабевают электростатические взаимодействия и водородные связи между другими молекулами и ионами. С другой стороны, в присутствии воды усиливается взаимодействие неполярных молекул. Так, при связывании субстрата с активным центром, лежащим в щели на ферменте, происходит исключение воды из этой щели. Отсутствие воды усиливает электростатические взаимодействия и водородные связи между ферментом и субстратом. Ассоциация неполярных групп субстрата и активного центра фермента обеспечивает значительную часть энергии, необходимой для связывания. Основой специфичности фермент-субстратного взаимодействия служат, во-первых, водородные связи, имеющие резко выраженный направленный характер, и, во-вторых, форма активного центра фермента, которая препятствует связыванию некомплементарных ей молекул. Узнавание субстрата ферментами - это во многих случаях динамический процесс, сопровождающийся конформационными изменениями в активном центре фермента.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
(см. скан)