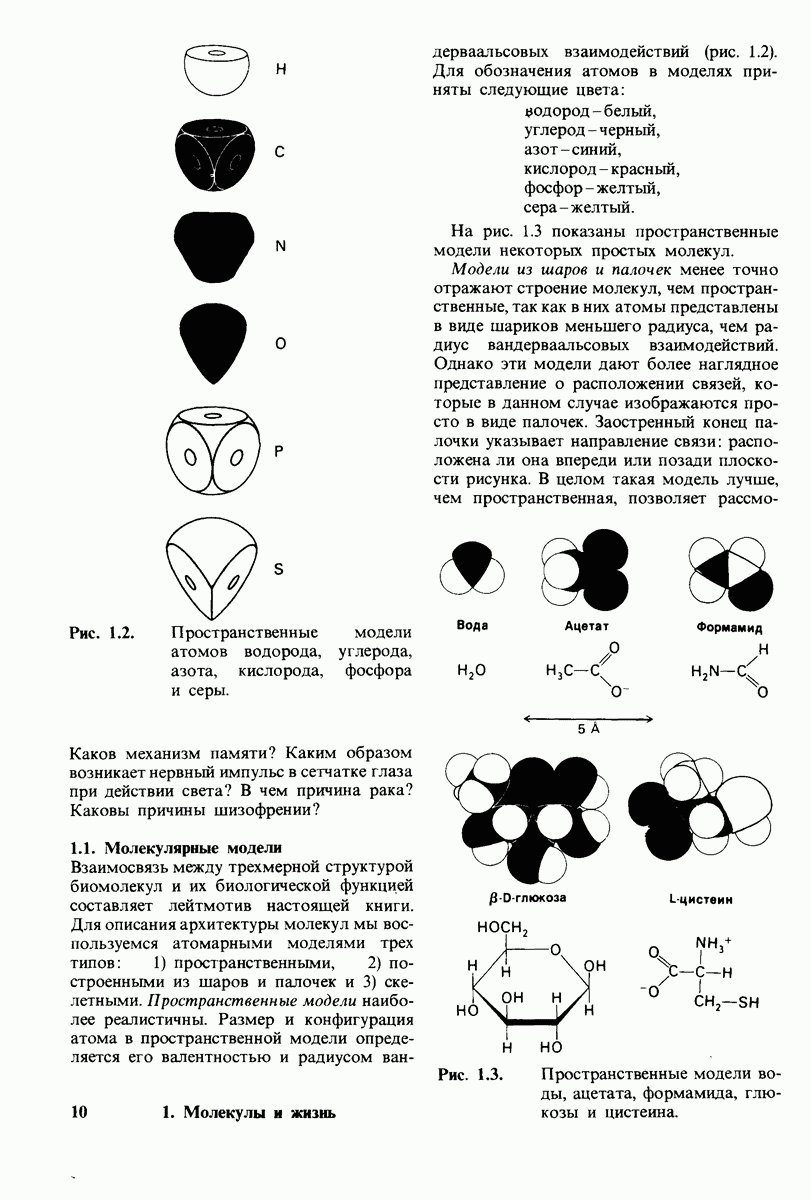
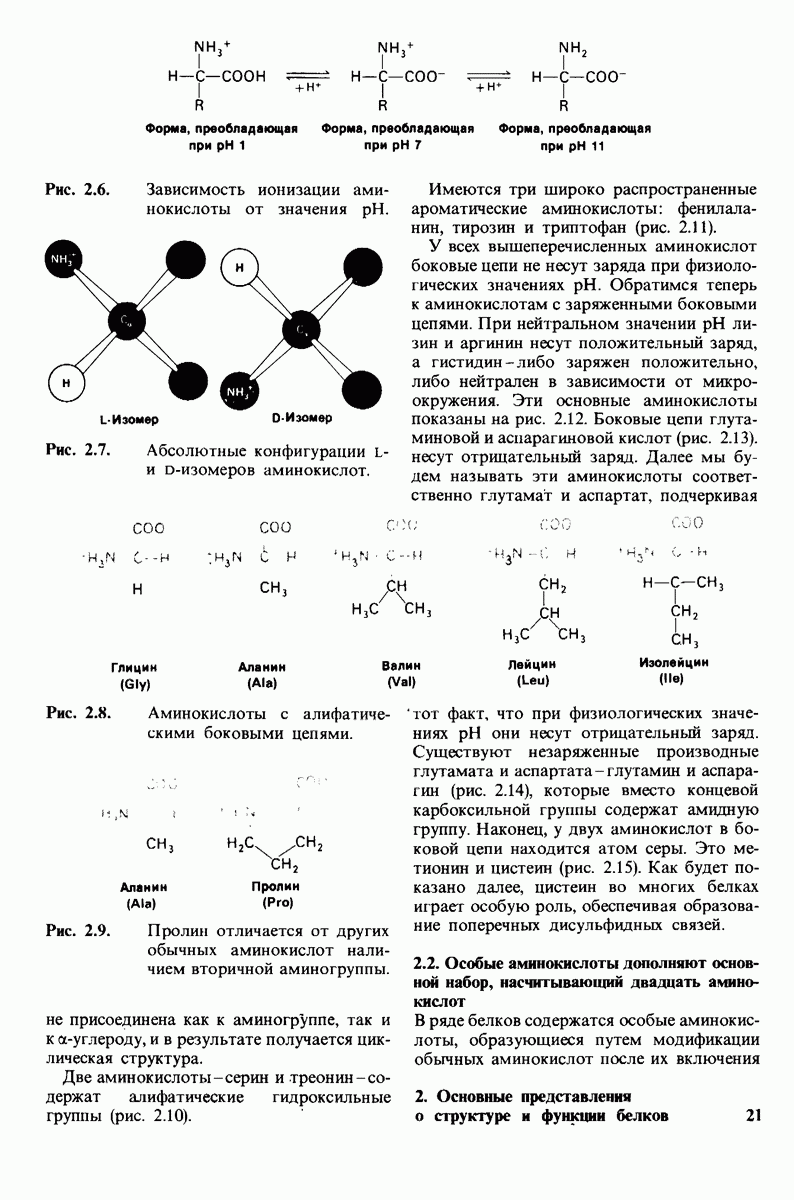
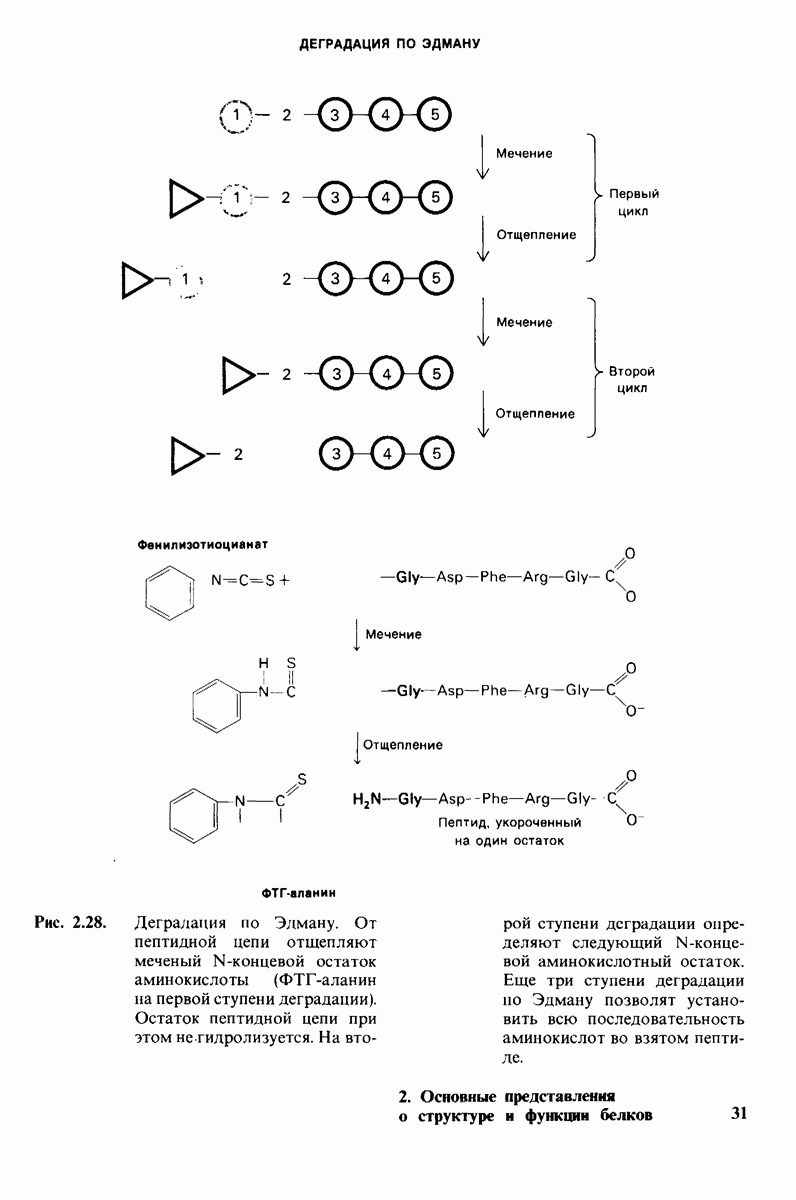
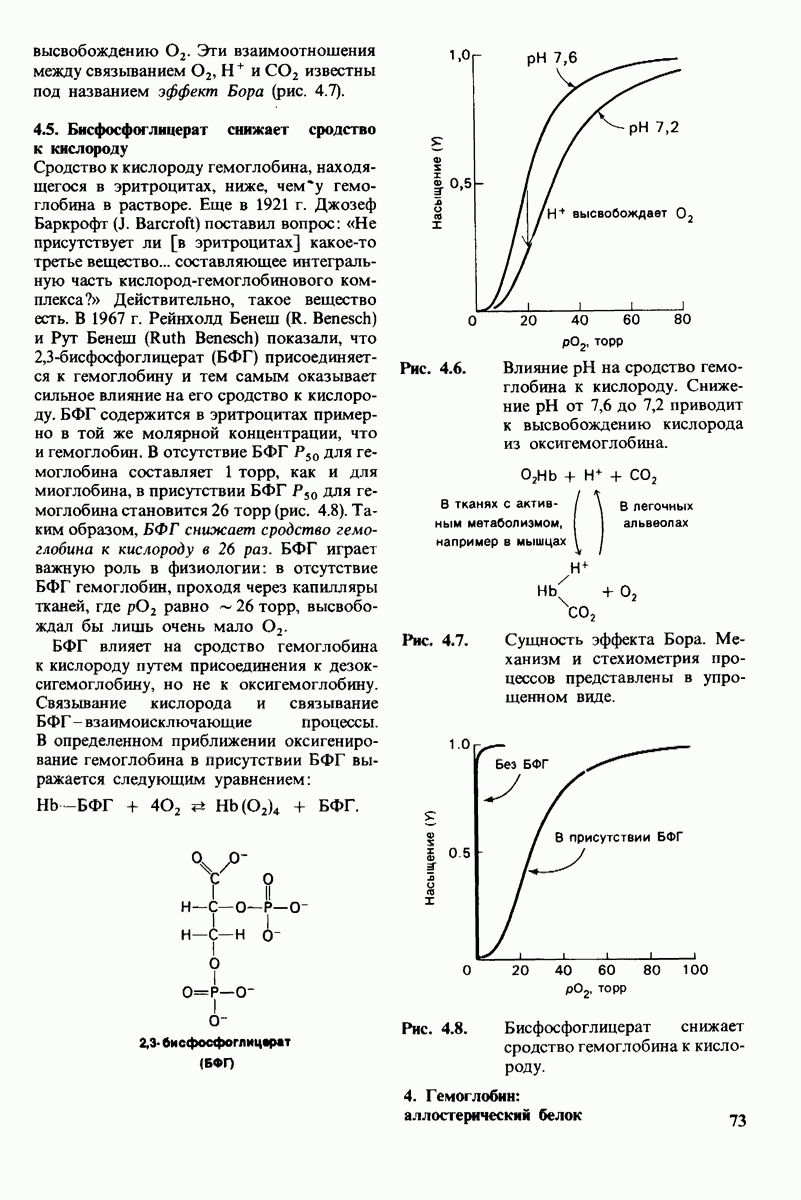
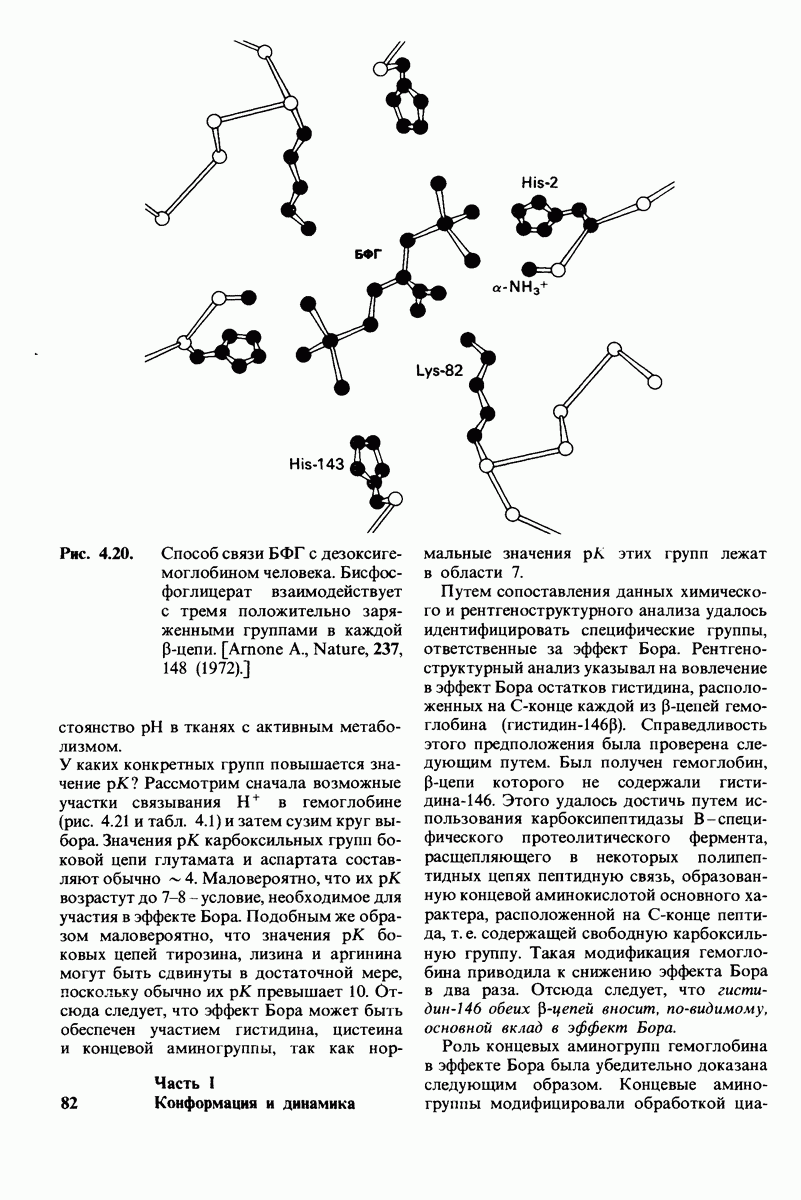
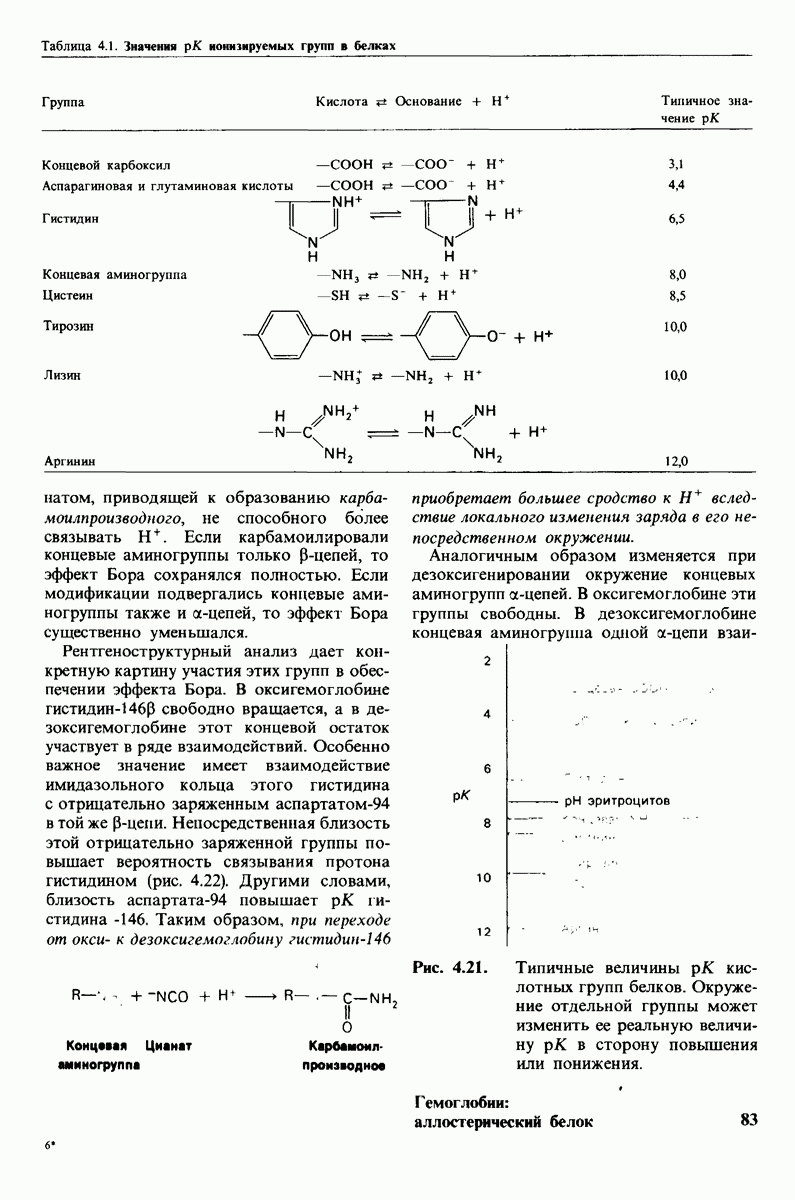
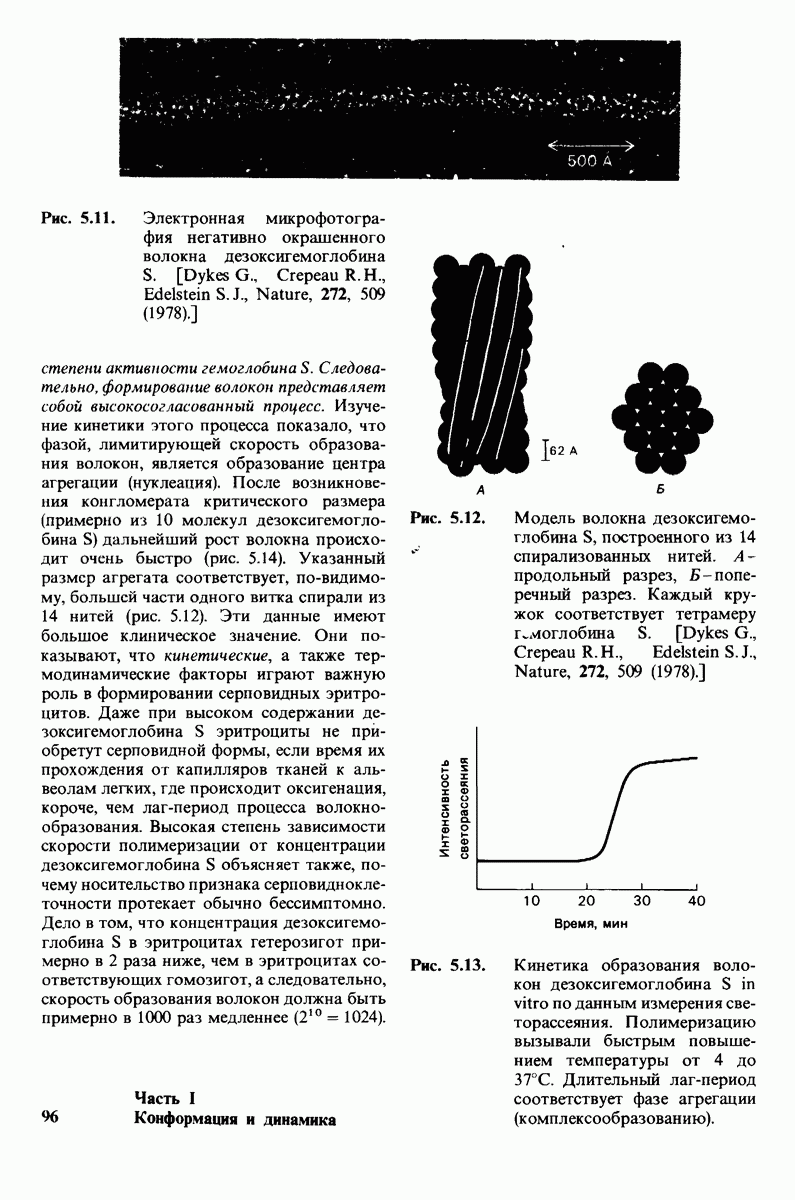
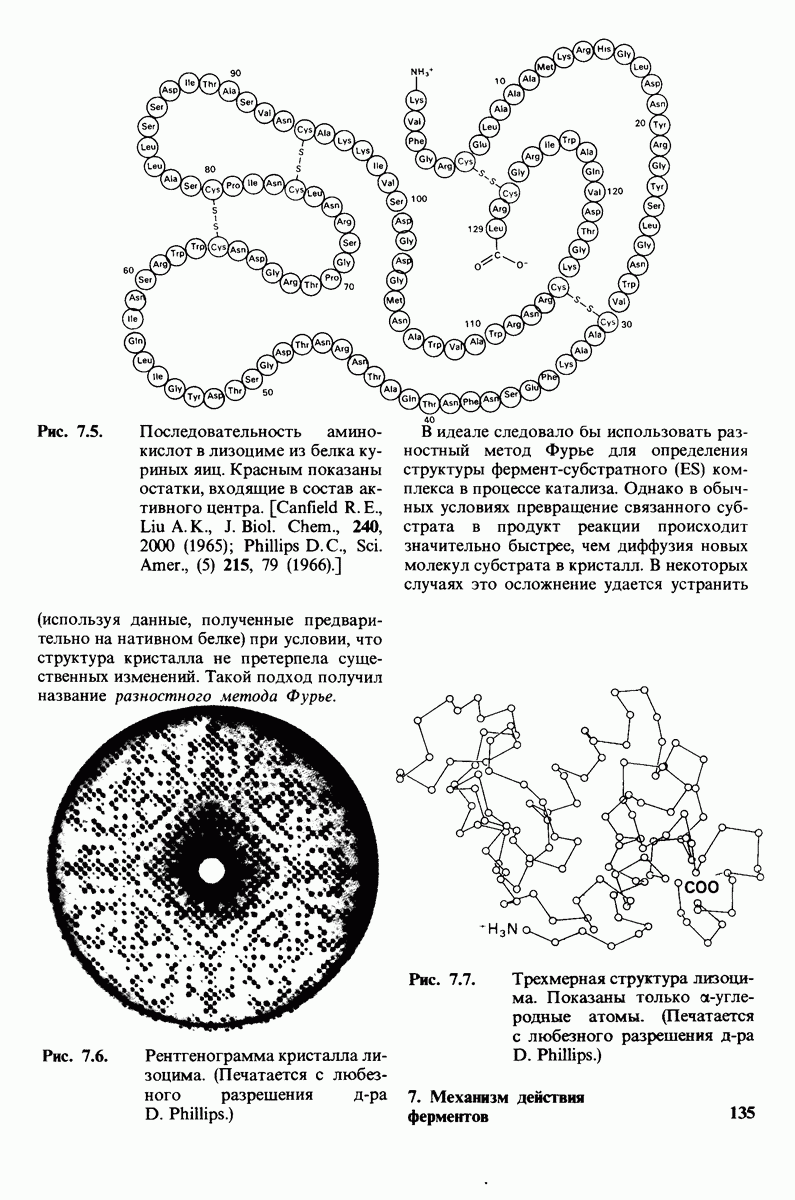
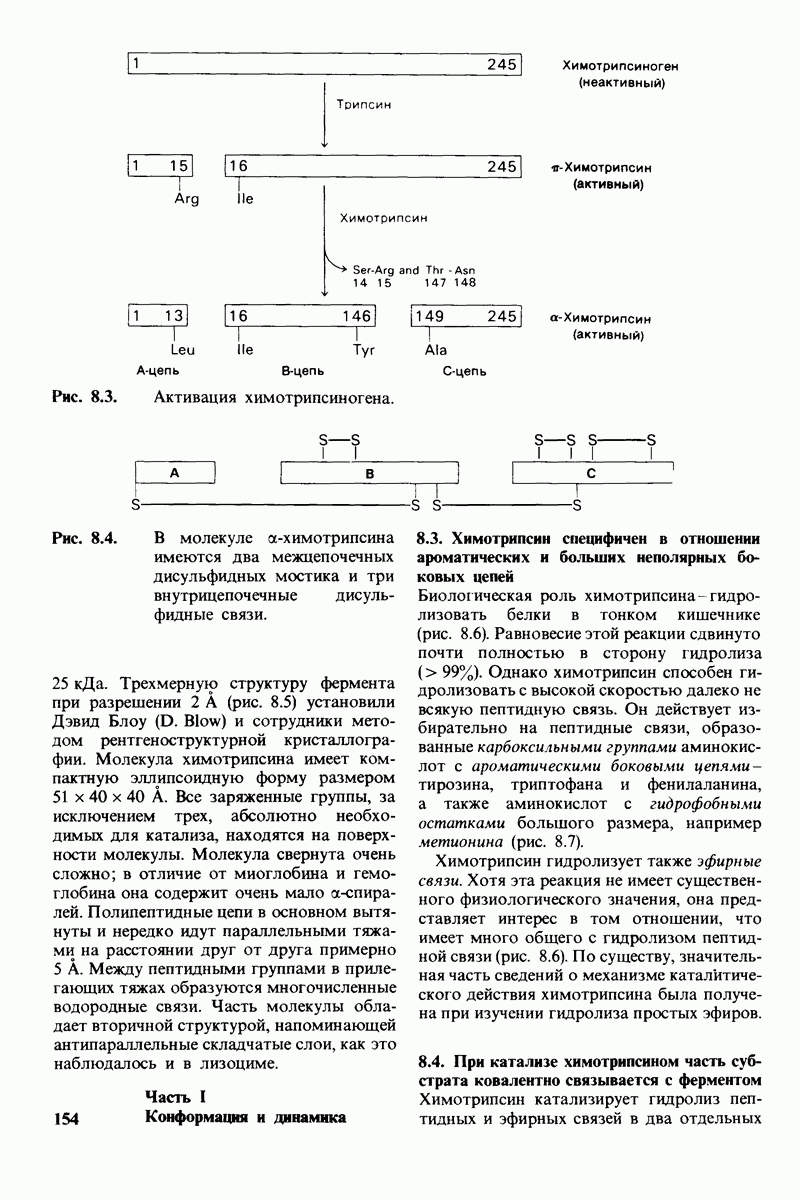
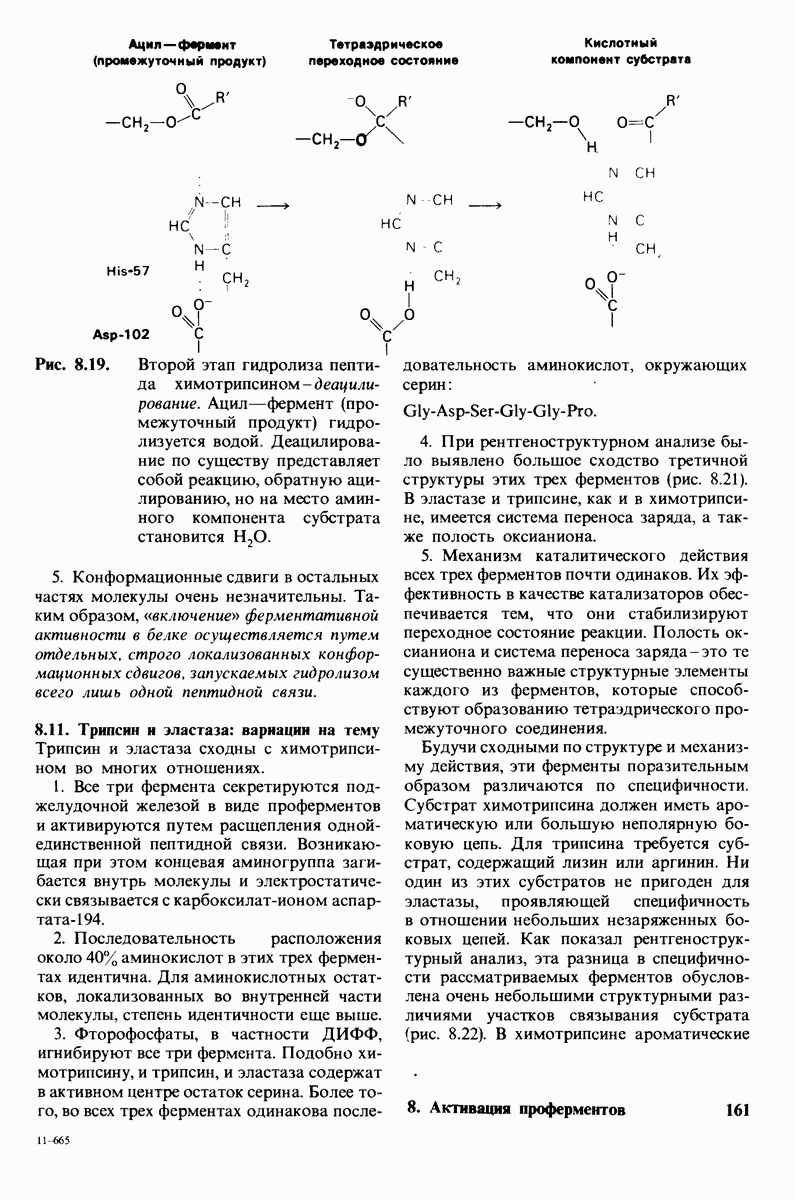
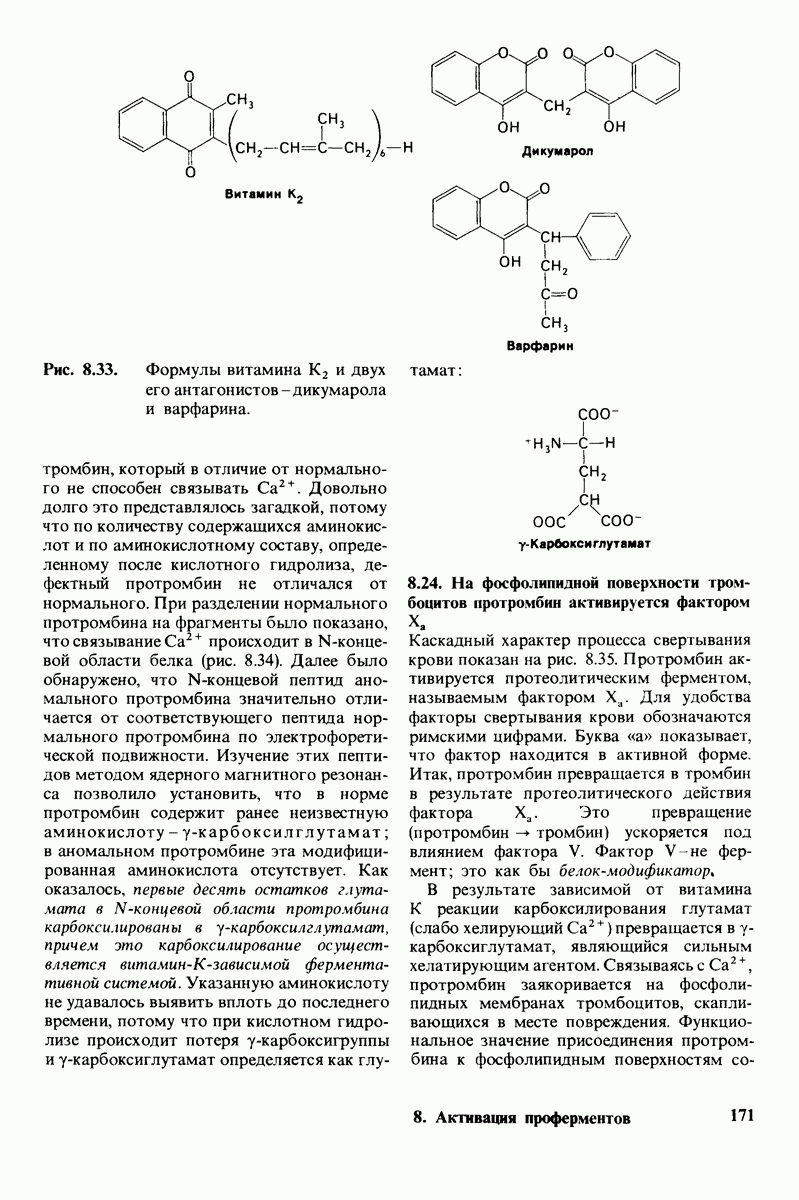
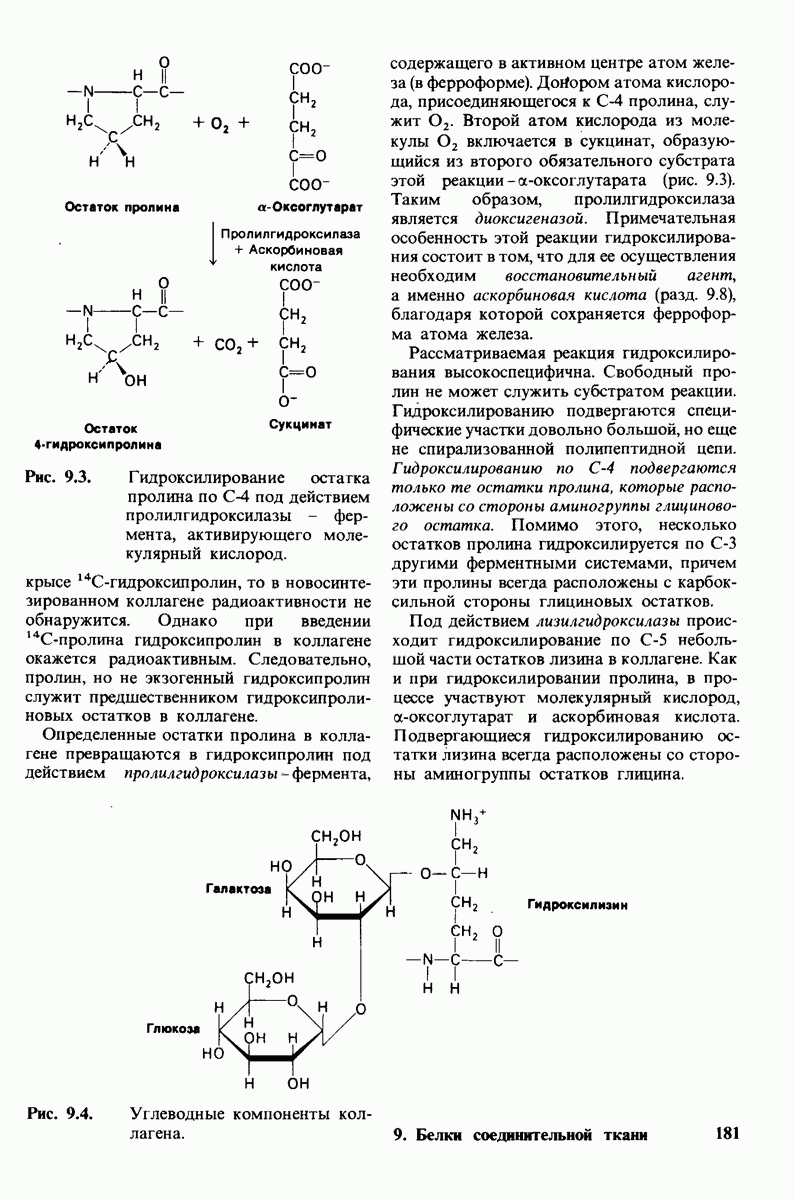
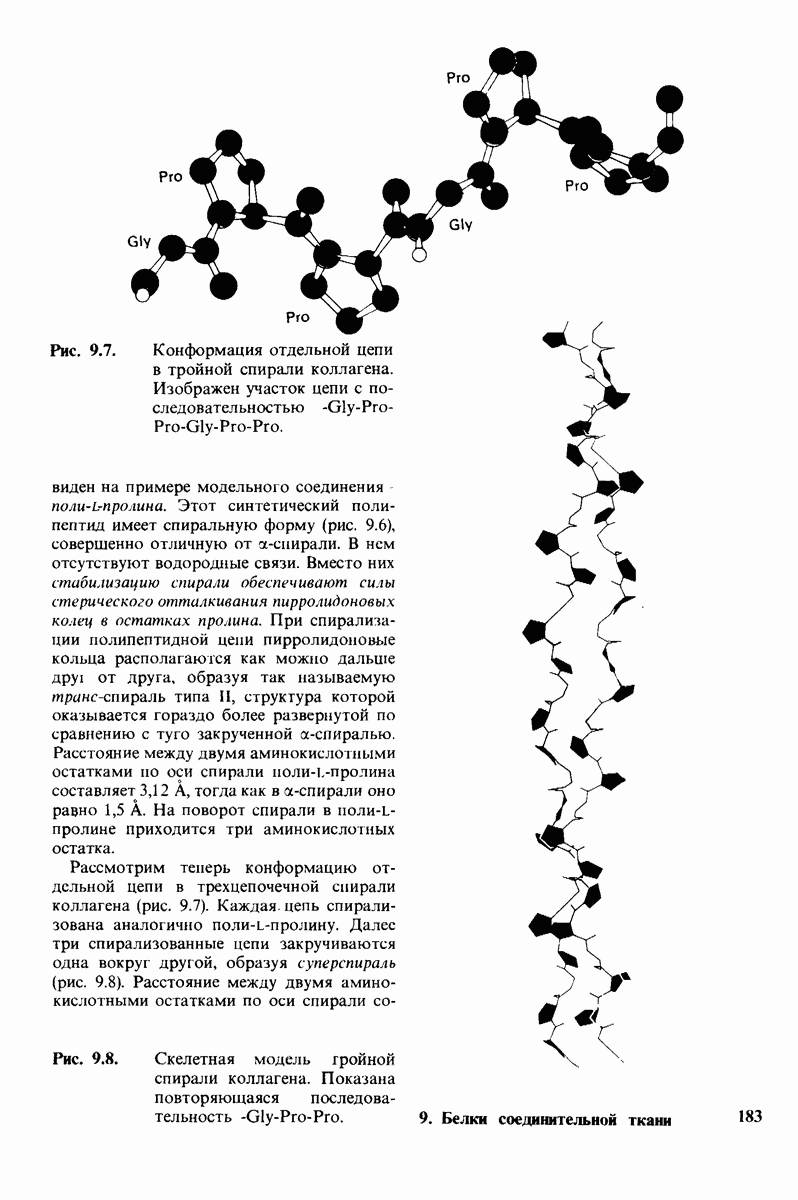
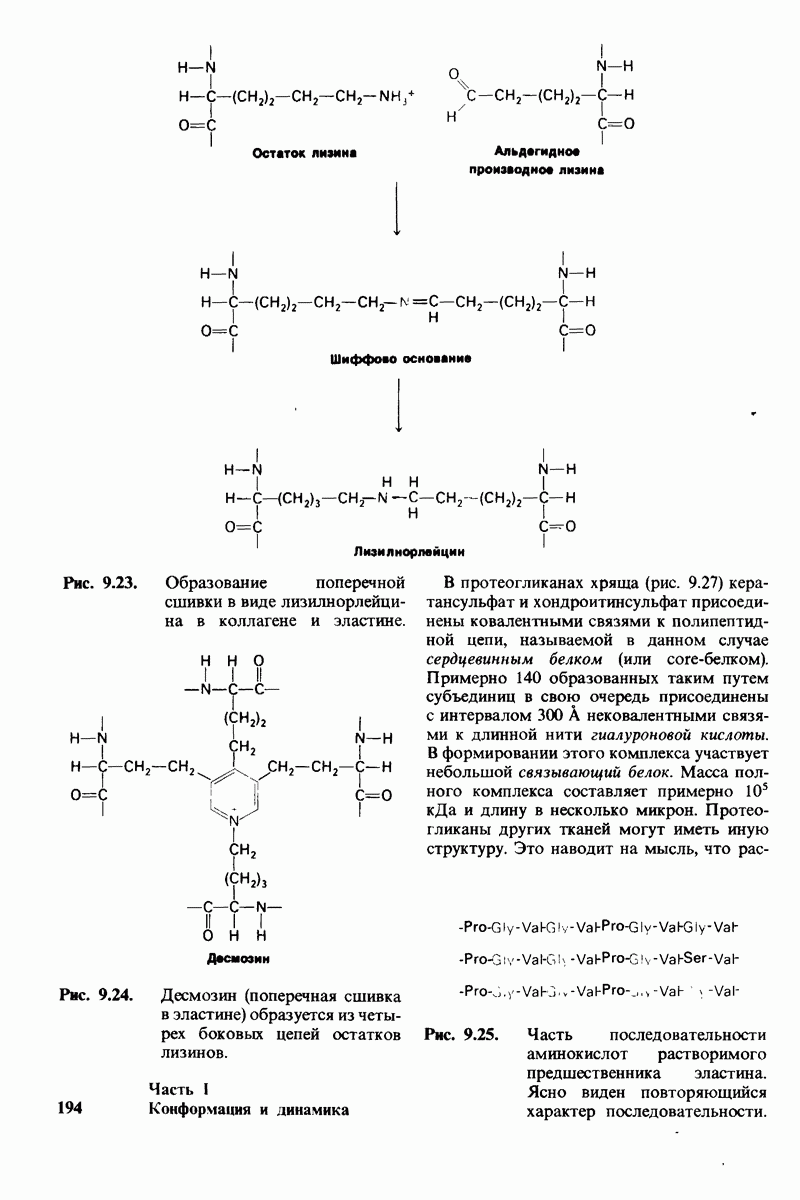
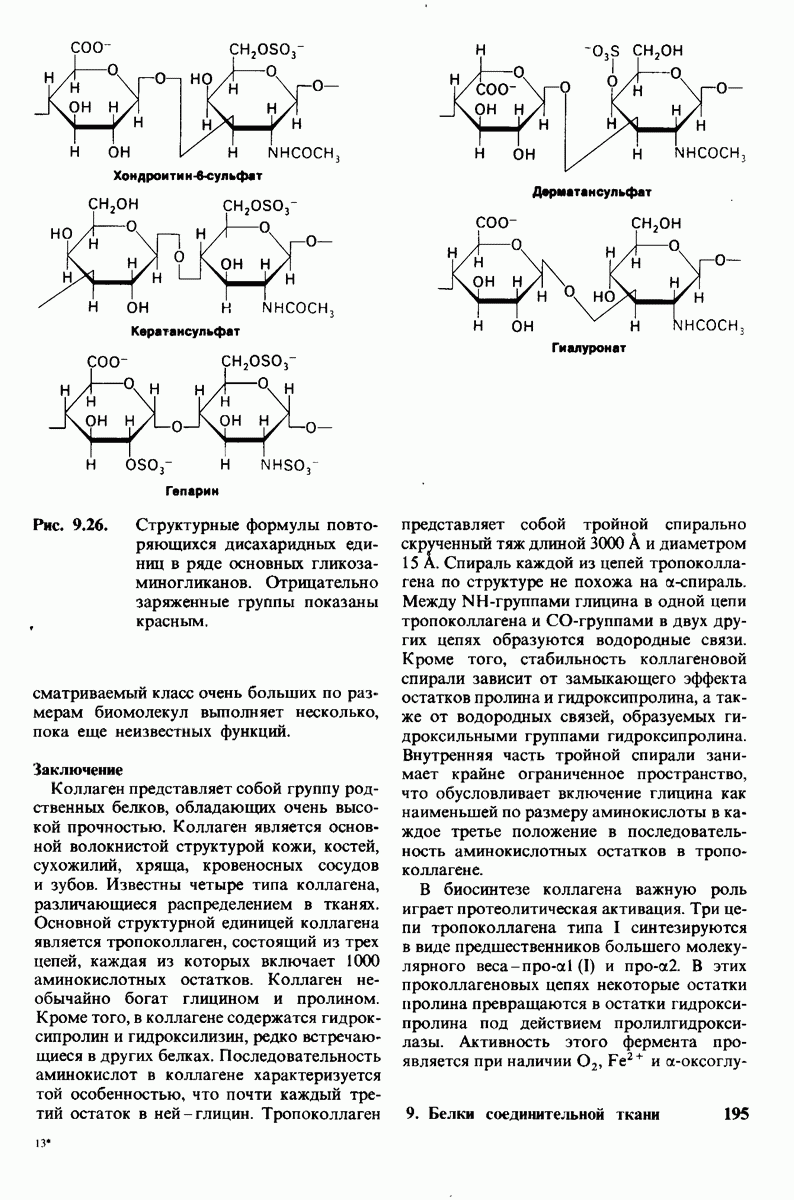
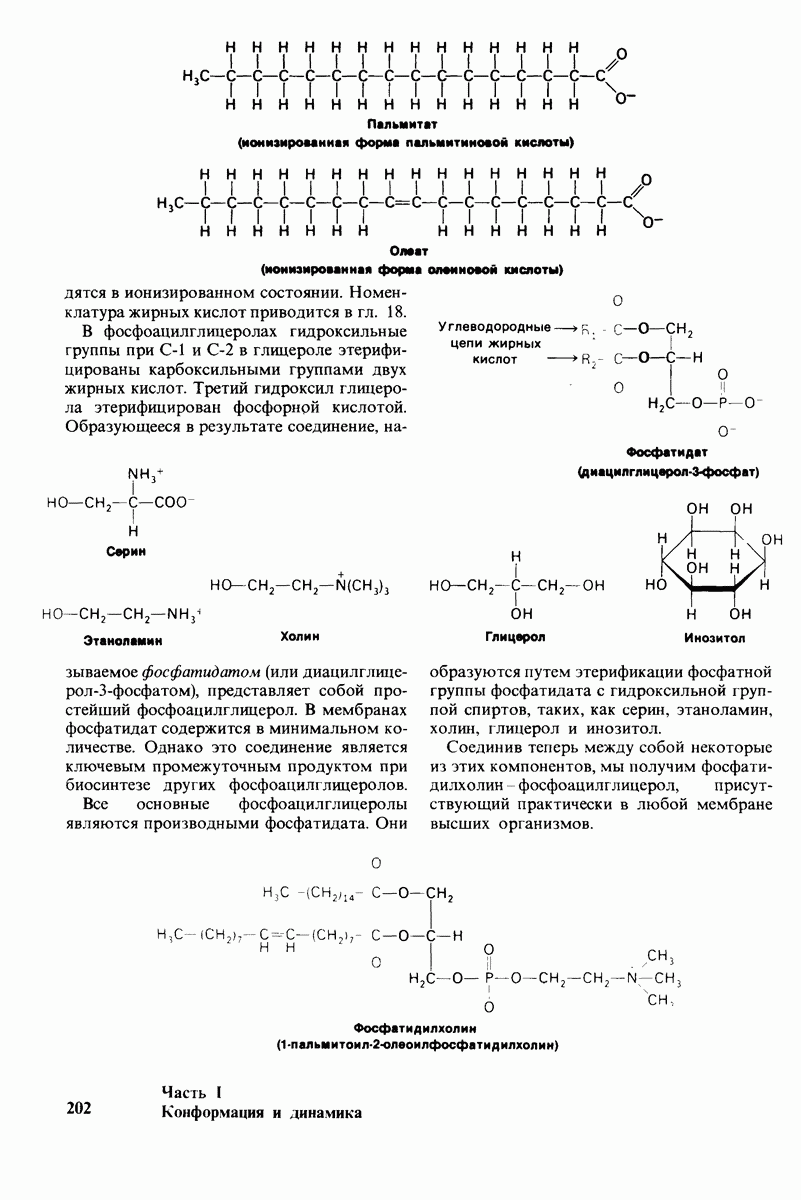
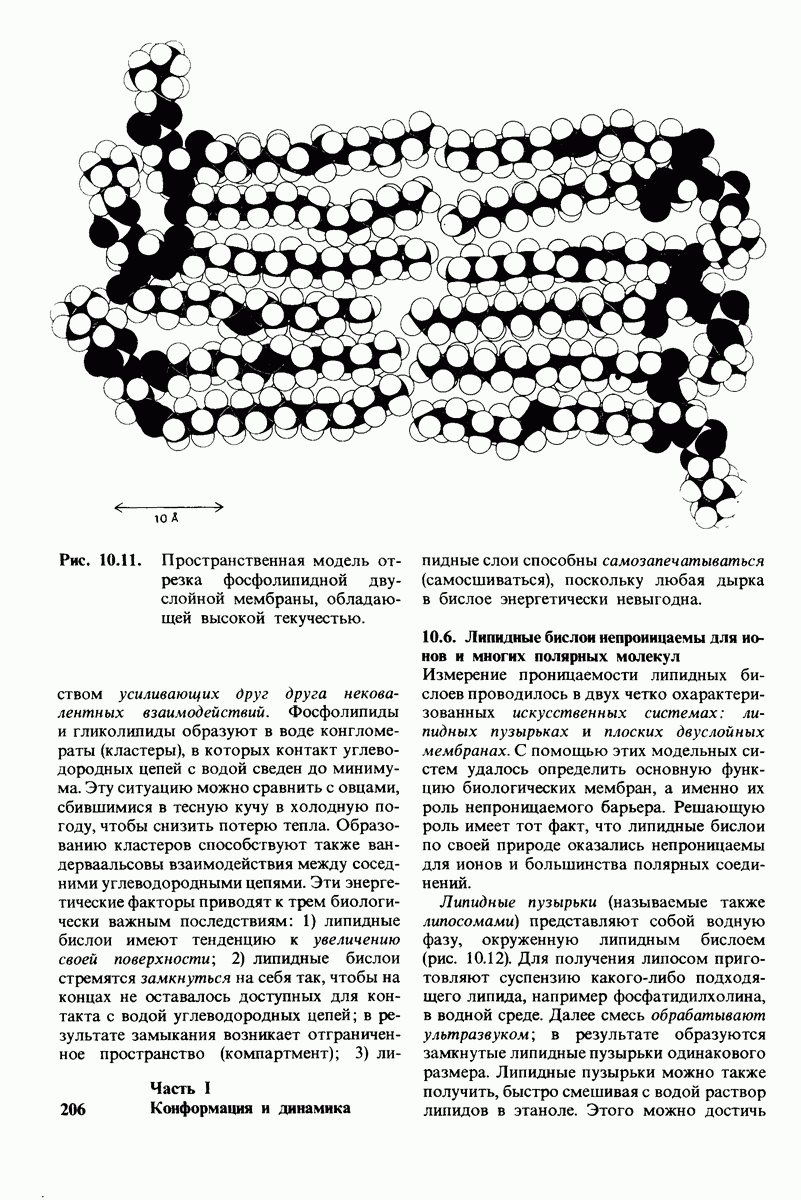
7.10. Скорость катализа карбоксипептидазой А возрастает благодаря смещению электронов
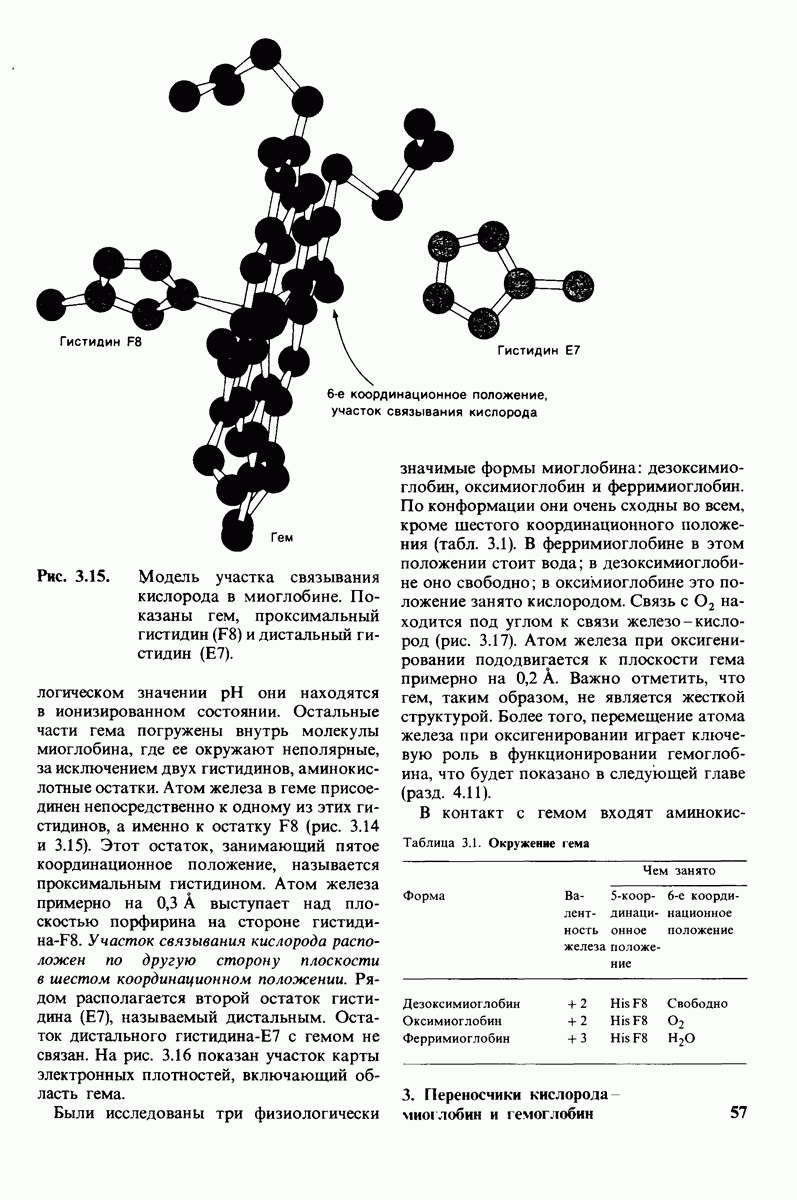
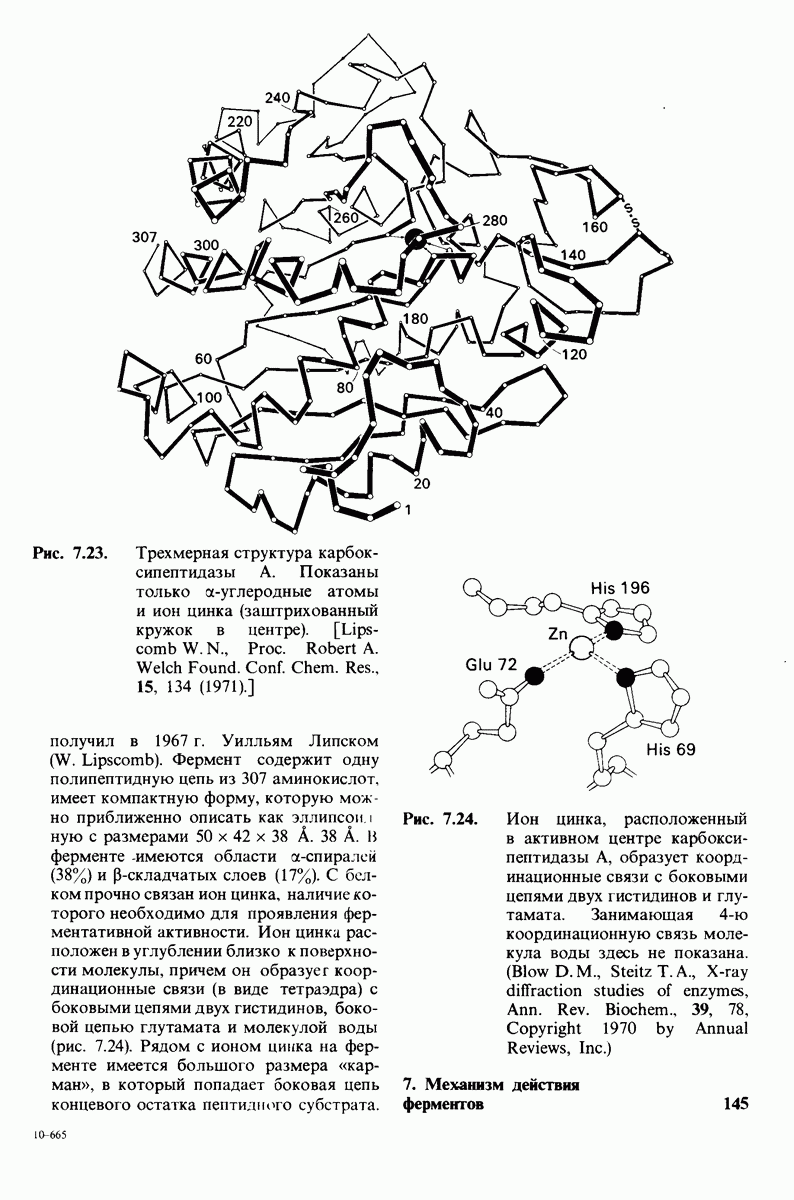
На основе данных рентгеноструктурного анализа Липском (Lipscomb) предложил механизм каталитического действия карбокси-пептидазы А. Постулированная структура реакционноспособного  -комплекса показана на рис. 7.28. Согласно предложенному механизму, ОН-группа тирозина-248 отдает протон на NH-группу расщепляемой пептидной связи. Карбонильный атом углерода этой пептидной связи подвергается атаке со стороны карбоксильной группы глутамата-270, которая выступает в данном случае как нуклеофильная группа. Образующийся в результате ангидрид глутамата-270 и кислотного компонента субстрата подвергается далее гидролизу.
-комплекса показана на рис. 7.28. Согласно предложенному механизму, ОН-группа тирозина-248 отдает протон на NH-группу расщепляемой пептидной связи. Карбонильный атом углерода этой пептидной связи подвергается атаке со стороны карбоксильной группы глутамата-270, которая выступает в данном случае как нуклеофильная группа. Образующийся в результате ангидрид глутамата-270 и кислотного компонента субстрата подвергается далее гидролизу.
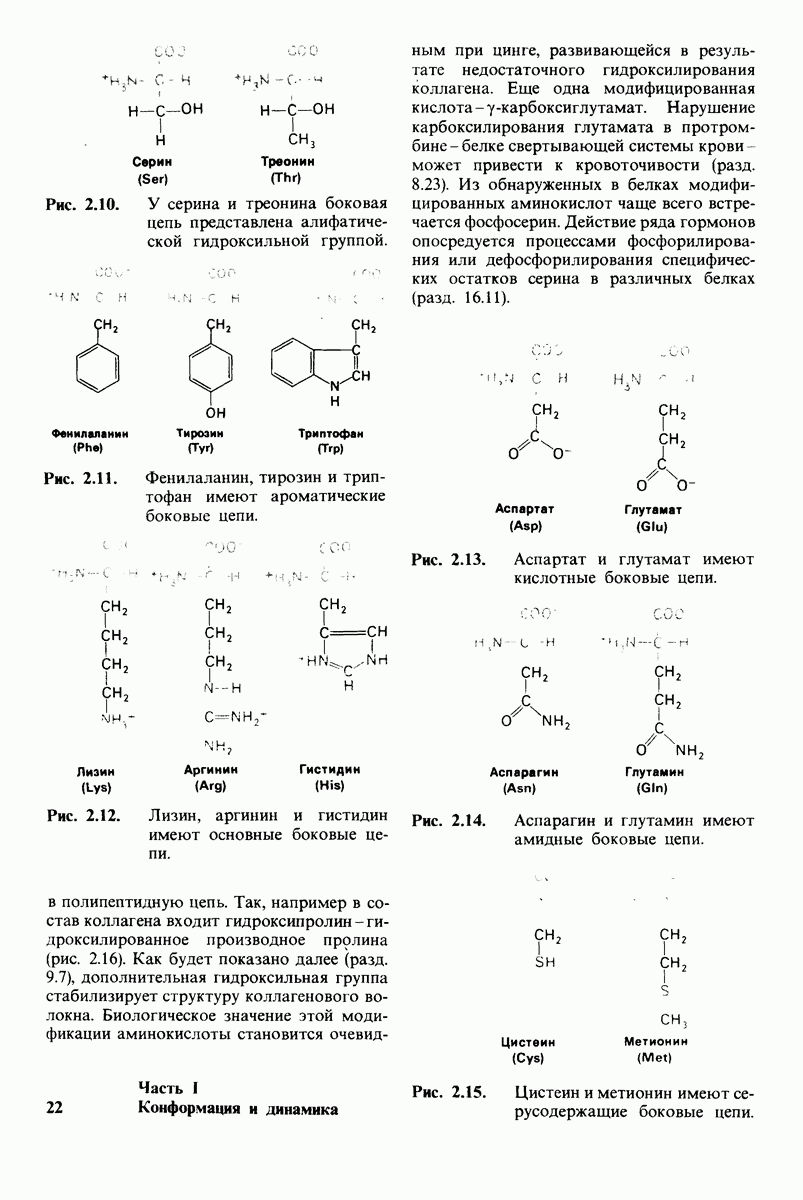
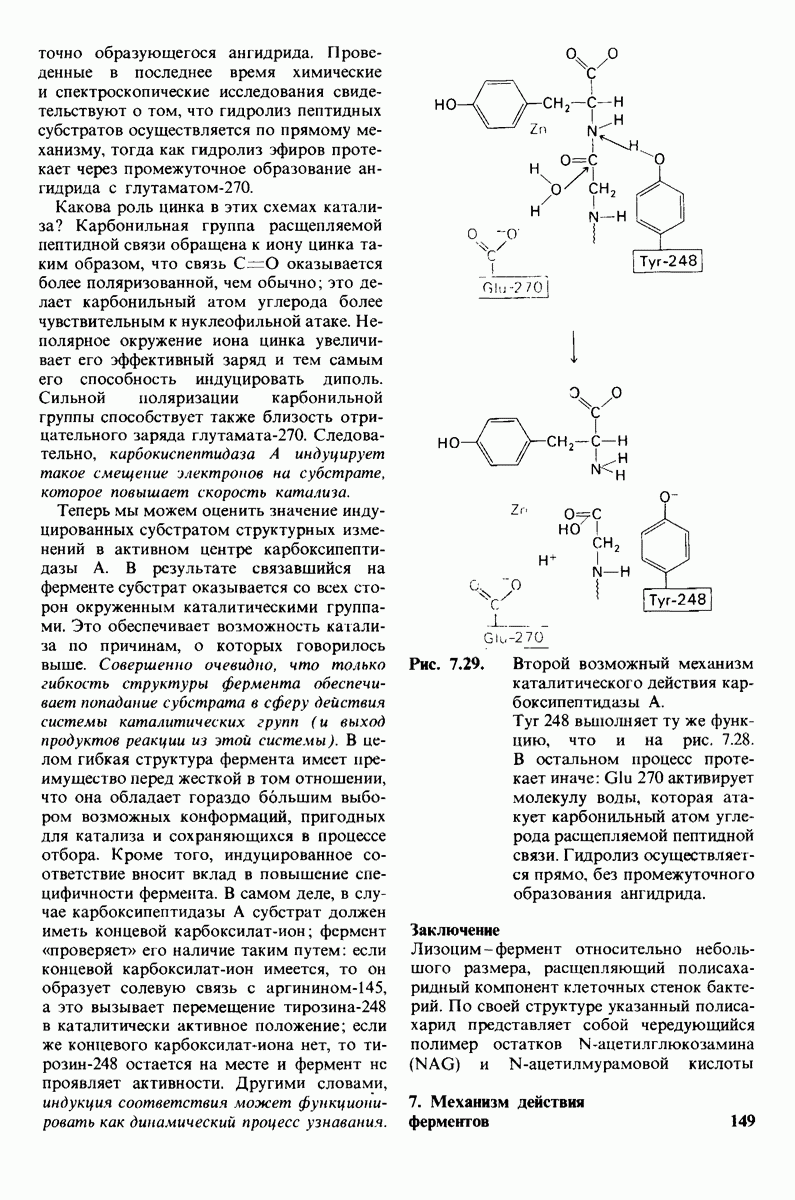
Возможен и другой механизм катализа, также согласующийся с данными рентгеноструктурного анализа; он показан на рис. 7.29. По этой схеме глутамат-270 активирует молекулу воды. Образующийся  непосредственно атакует карбонильный атом углерода расщепляемой пептидной связи. Одновременно тирозин-248 отдает протон на ее NH-группу, и в итоге пептидная связь гидролизуется. Этот механизм катализа отличается от механизма, приведенного на рис. 7.28, тем, что предполагает гидролиз водой непосредственно пептидной связи субстрата, а не
непосредственно атакует карбонильный атом углерода расщепляемой пептидной связи. Одновременно тирозин-248 отдает протон на ее NH-группу, и в итоге пептидная связь гидролизуется. Этот механизм катализа отличается от механизма, приведенного на рис. 7.28, тем, что предполагает гидролиз водой непосредственно пептидной связи субстрата, а не
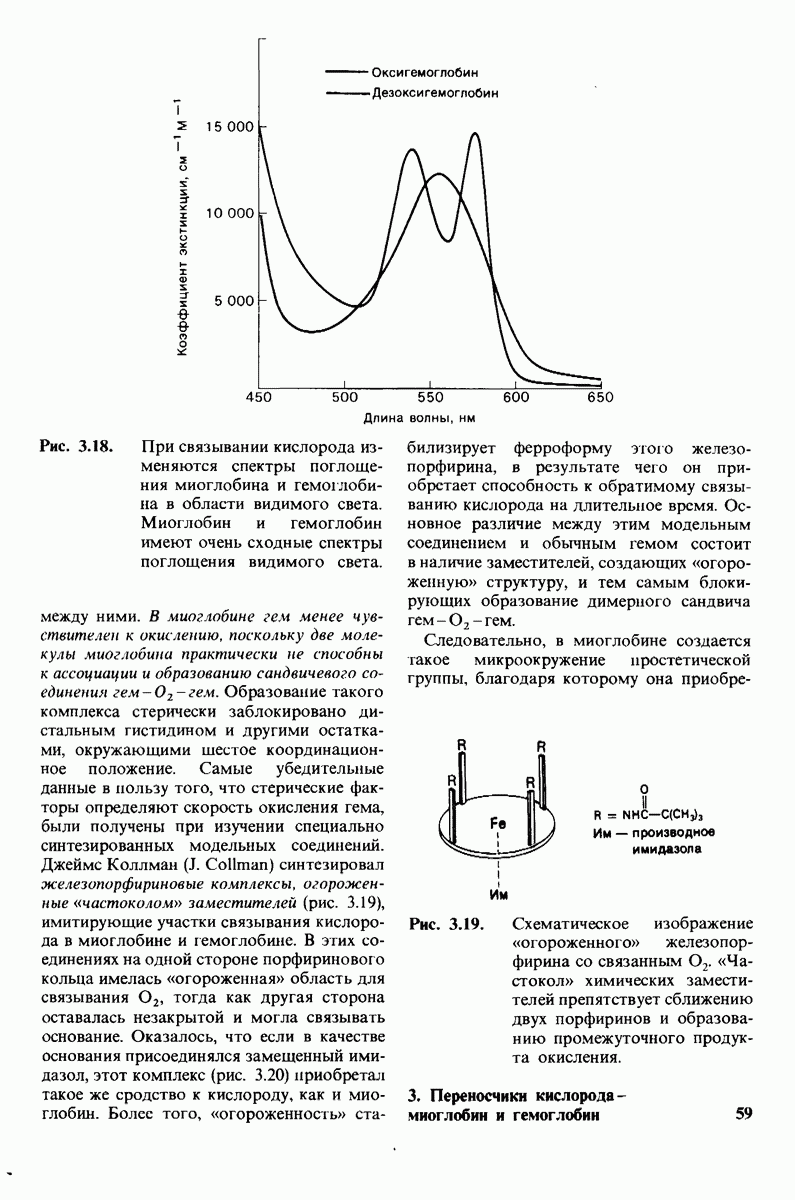
промежуточно образующегося ангидрида. Проведенные в последнее время химические и спектроскопические исследования свидетельствуют о том, что гидролиз пептидных субстратов осуществляется по прямому механизму, тогда как гидролиз эфиров протекает через промежуточное образование ангидрида с глутаматом-270.
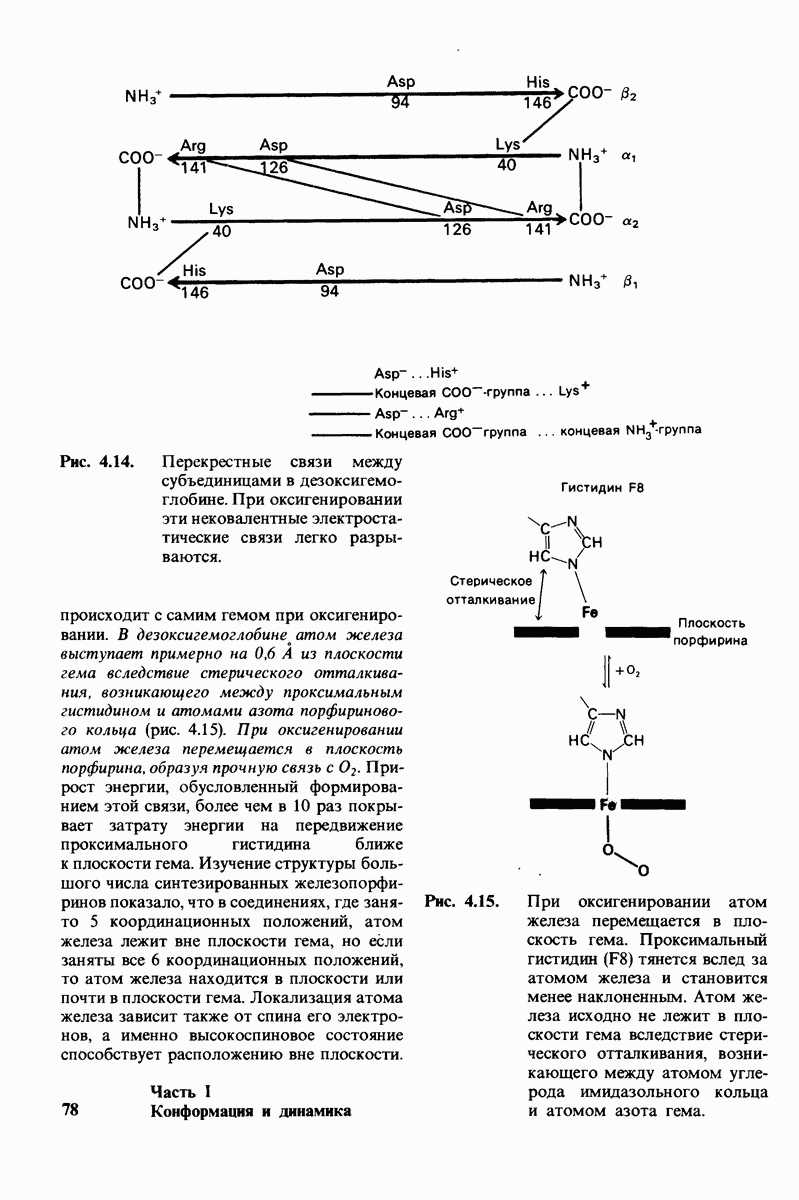
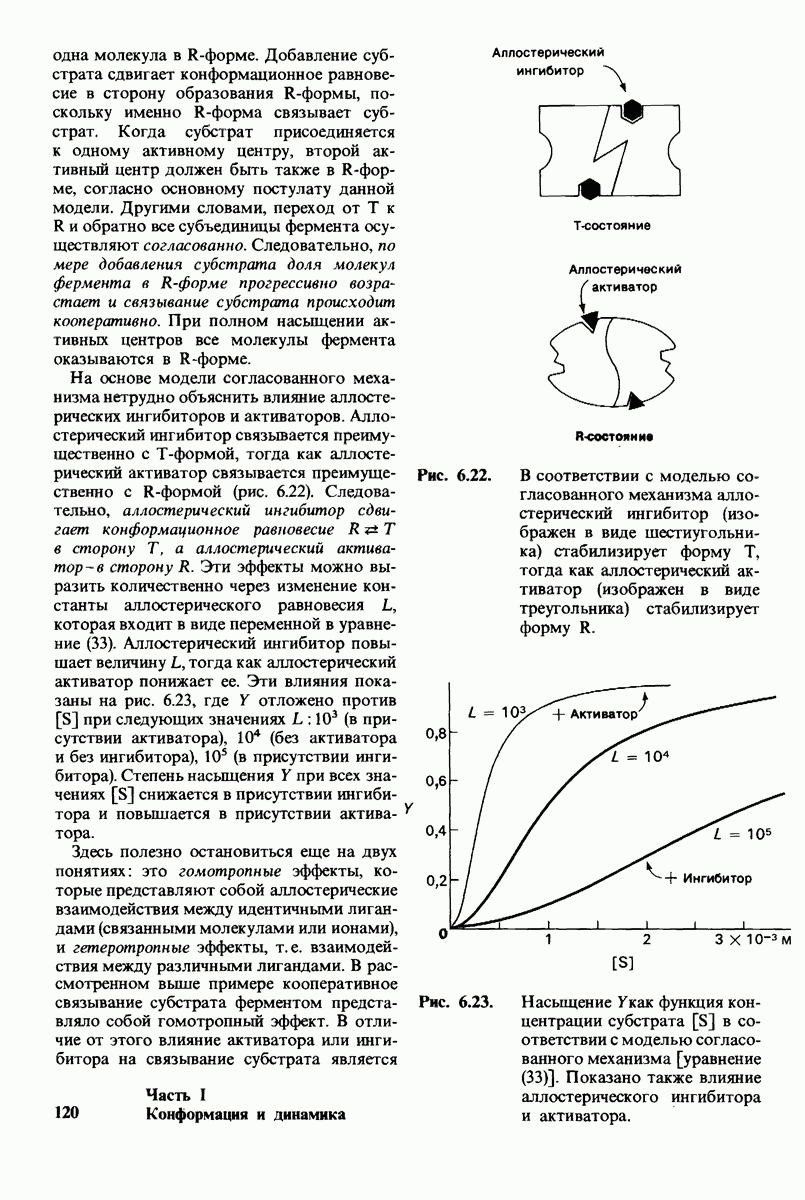
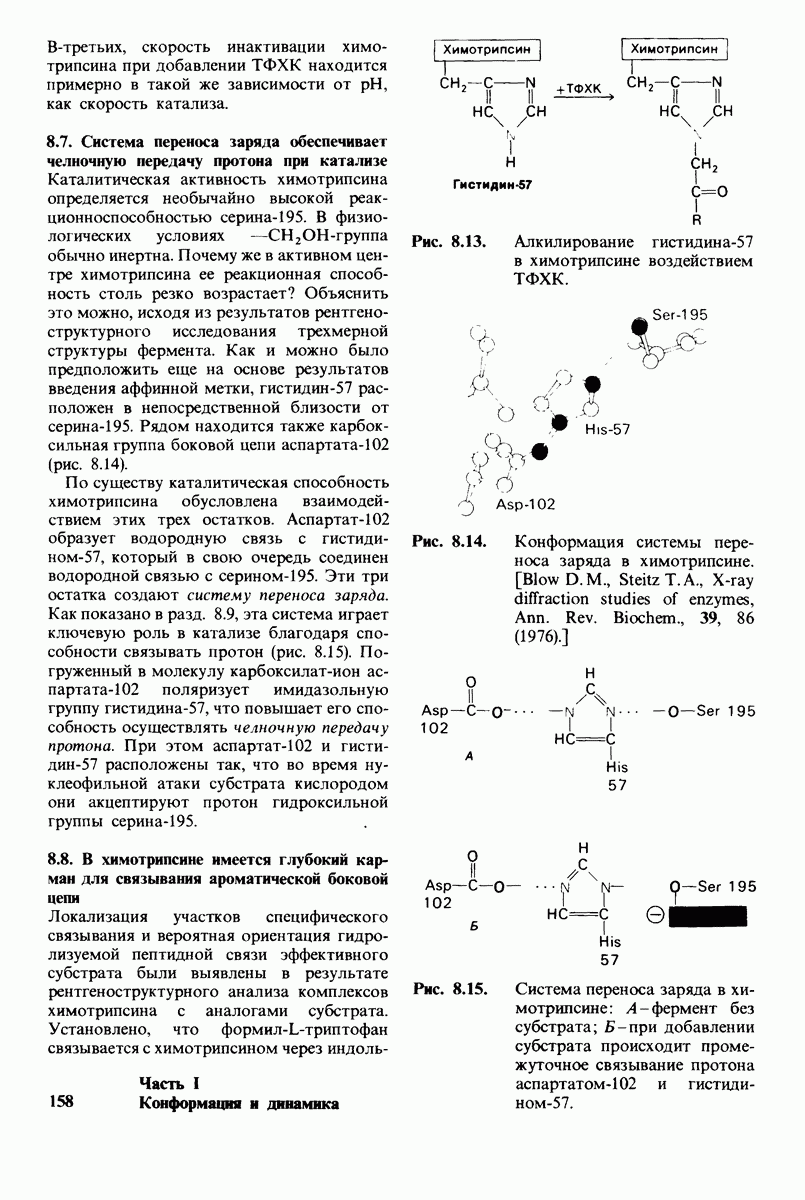
Какова роль цинка в этих схемах катализа? Карбонильная группа расщепляемой пептидной связи обращена к иону цинка таким образом, что связь  оказывается более поляризованной, чем обычно; это делает карбонильный атом углерода более чувствительным к нуклеофильной атаке. Неполярное окружение иона цинка увеличивает его эффективный заряд и тем самым его способность индуцировать диполь. Сильной поляризации карбонильной группы способствует также близость отрицательного заряда глутамата-270. Следовательно, карбокиспептидаза
оказывается более поляризованной, чем обычно; это делает карбонильный атом углерода более чувствительным к нуклеофильной атаке. Неполярное окружение иона цинка увеличивает его эффективный заряд и тем самым его способность индуцировать диполь. Сильной поляризации карбонильной группы способствует также близость отрицательного заряда глутамата-270. Следовательно, карбокиспептидаза  индуцирует такое смещение электронов на субстрате, которое повышает скорость катализа.
индуцирует такое смещение электронов на субстрате, которое повышает скорость катализа.
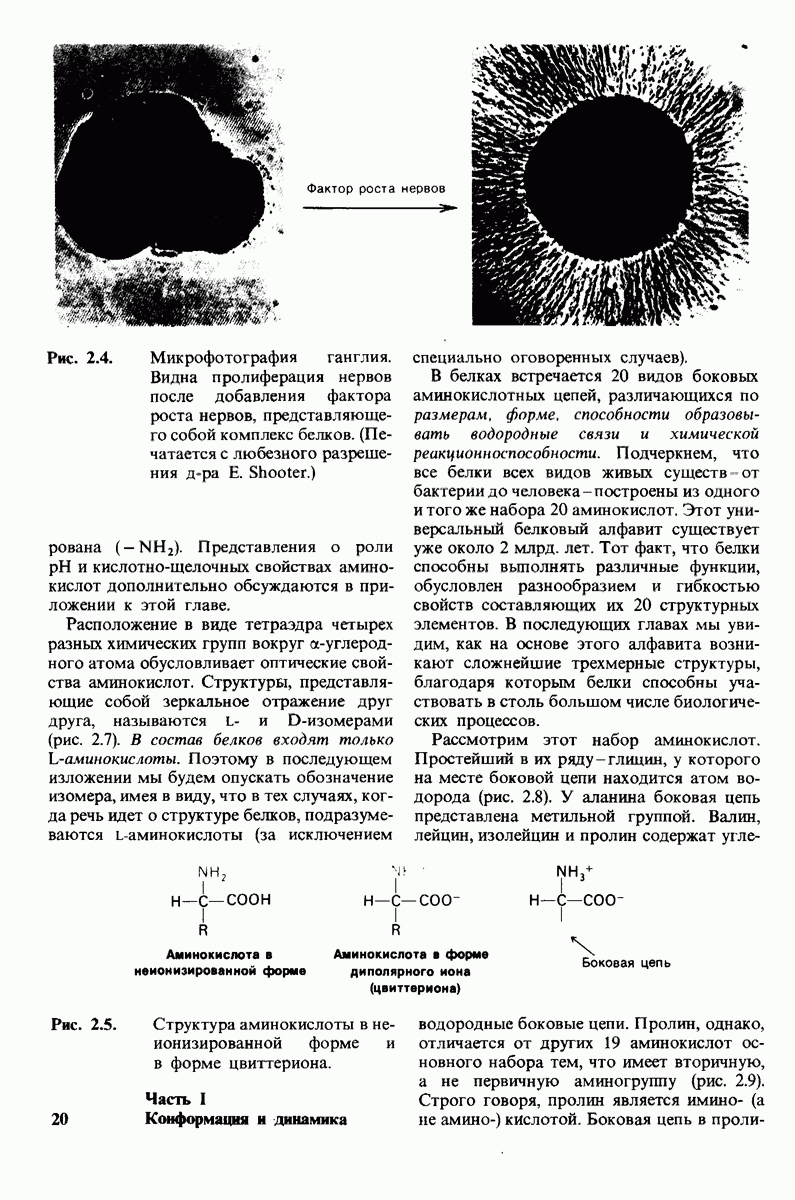
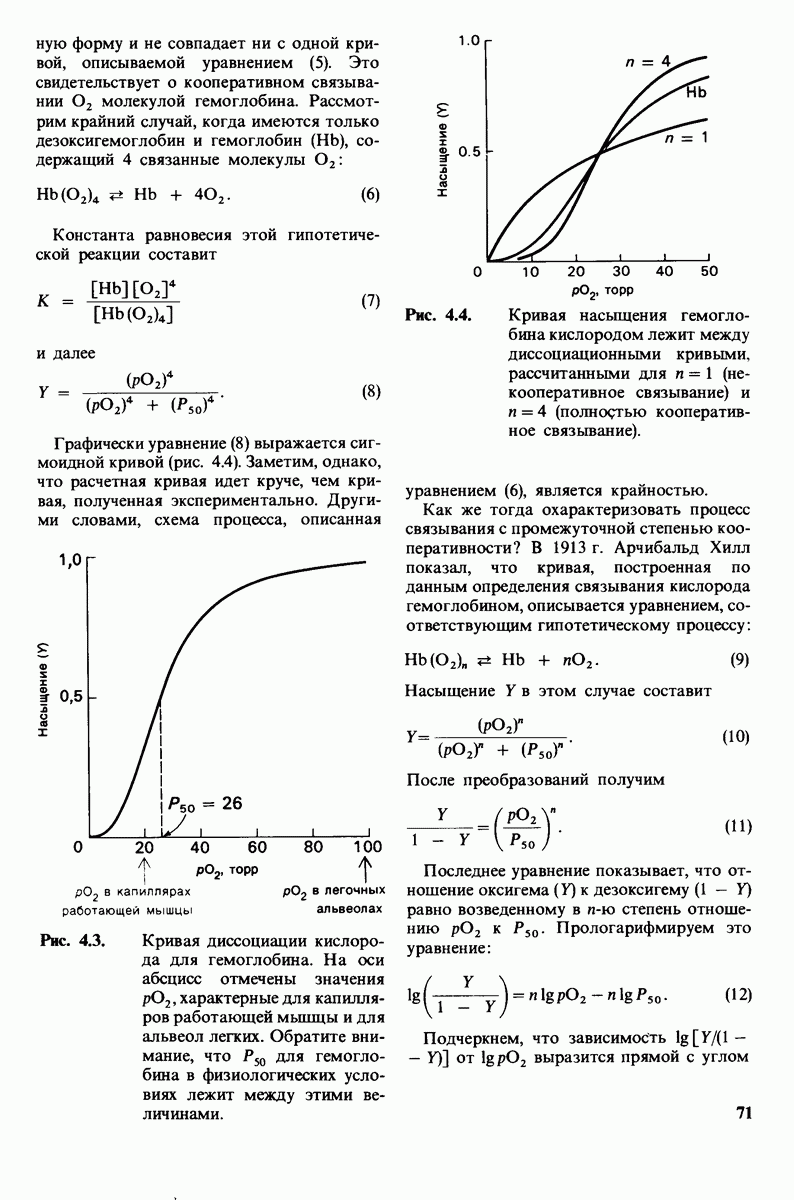
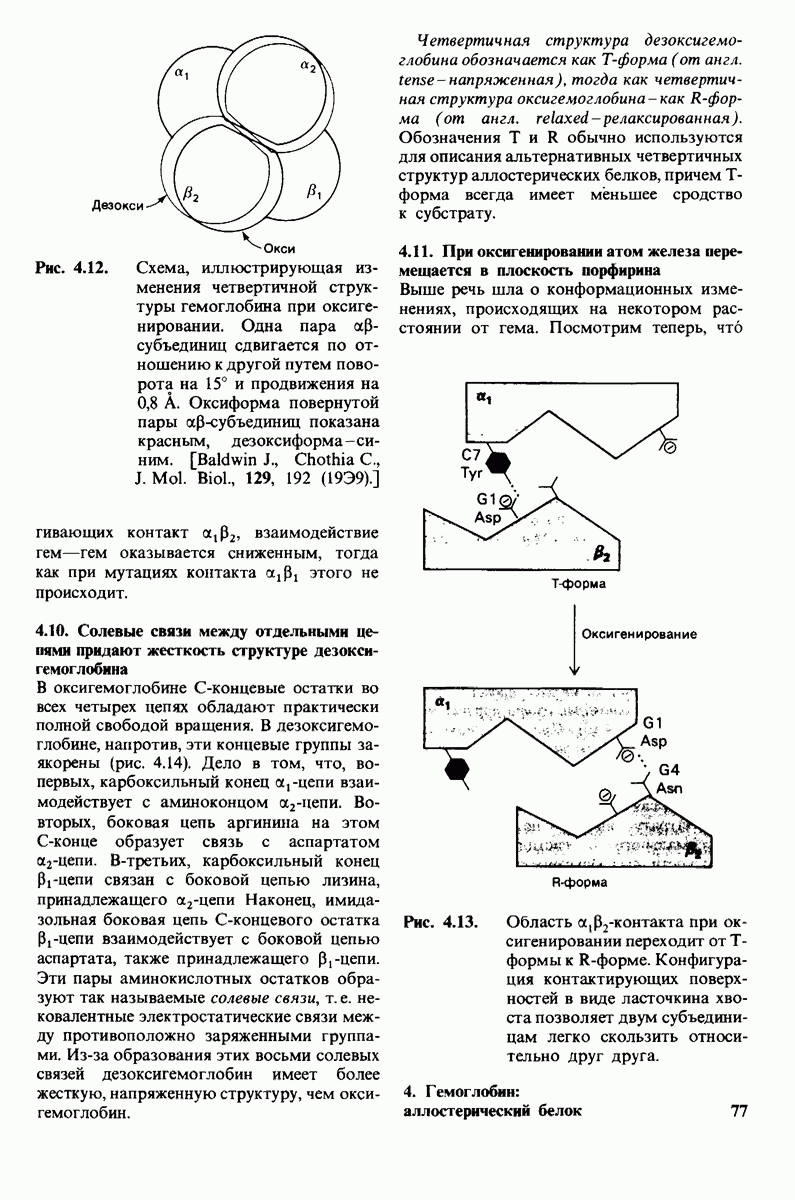
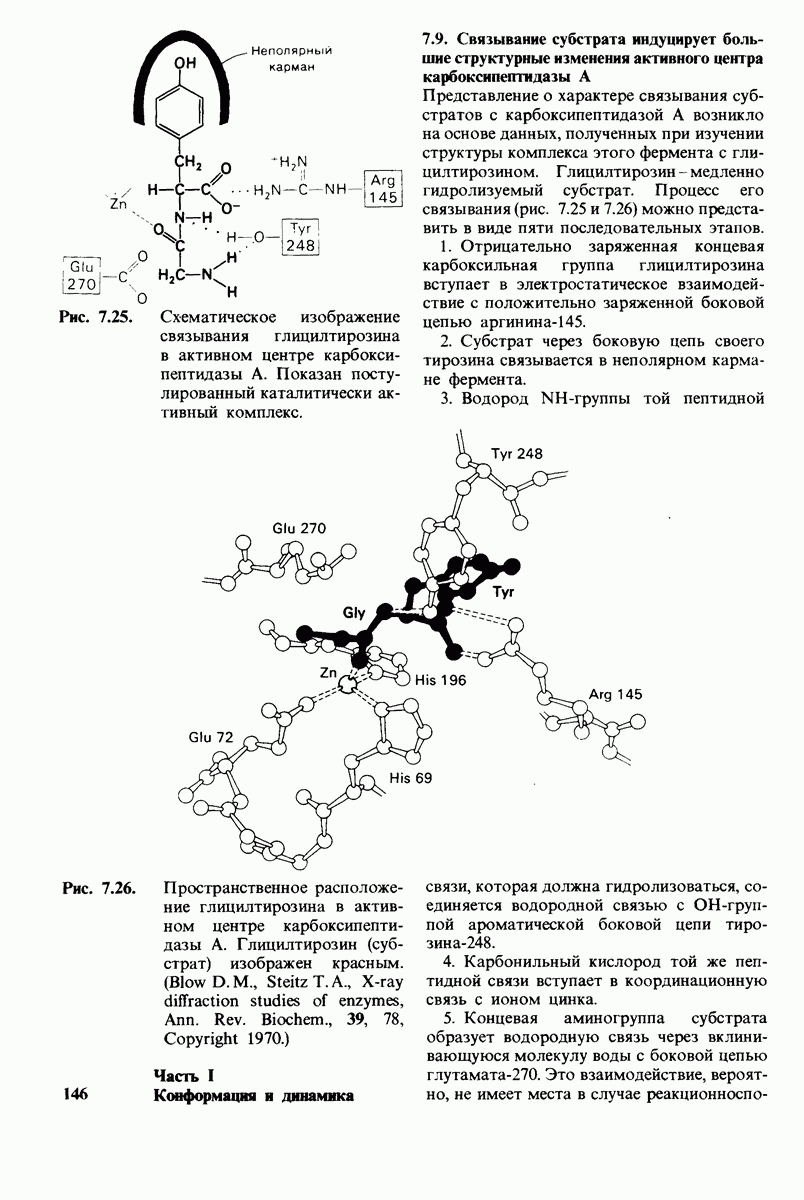
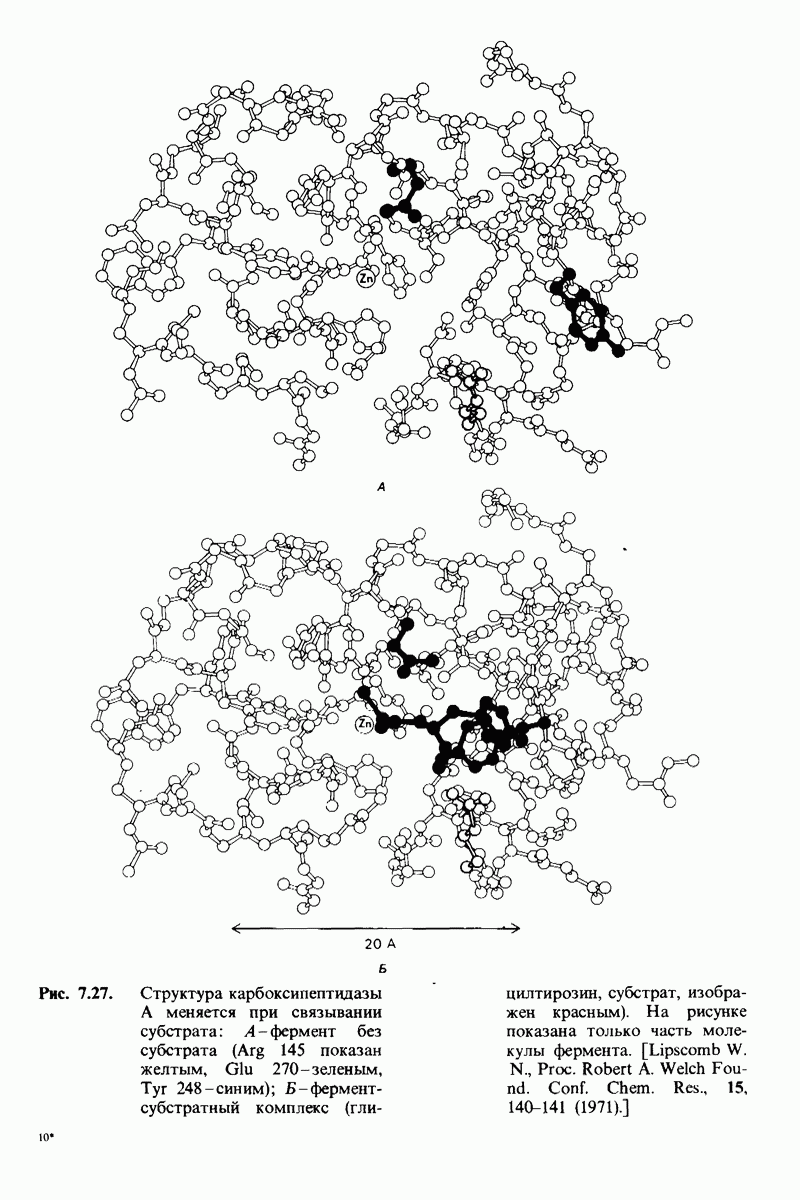
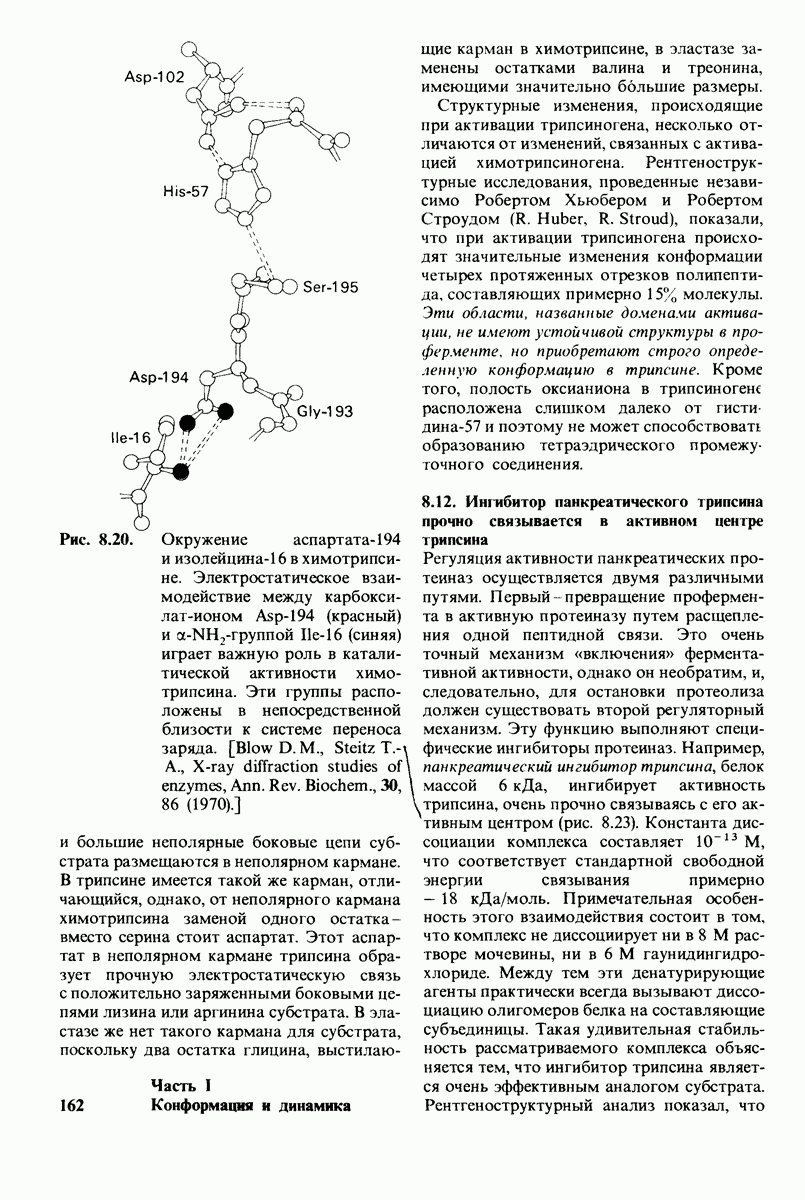
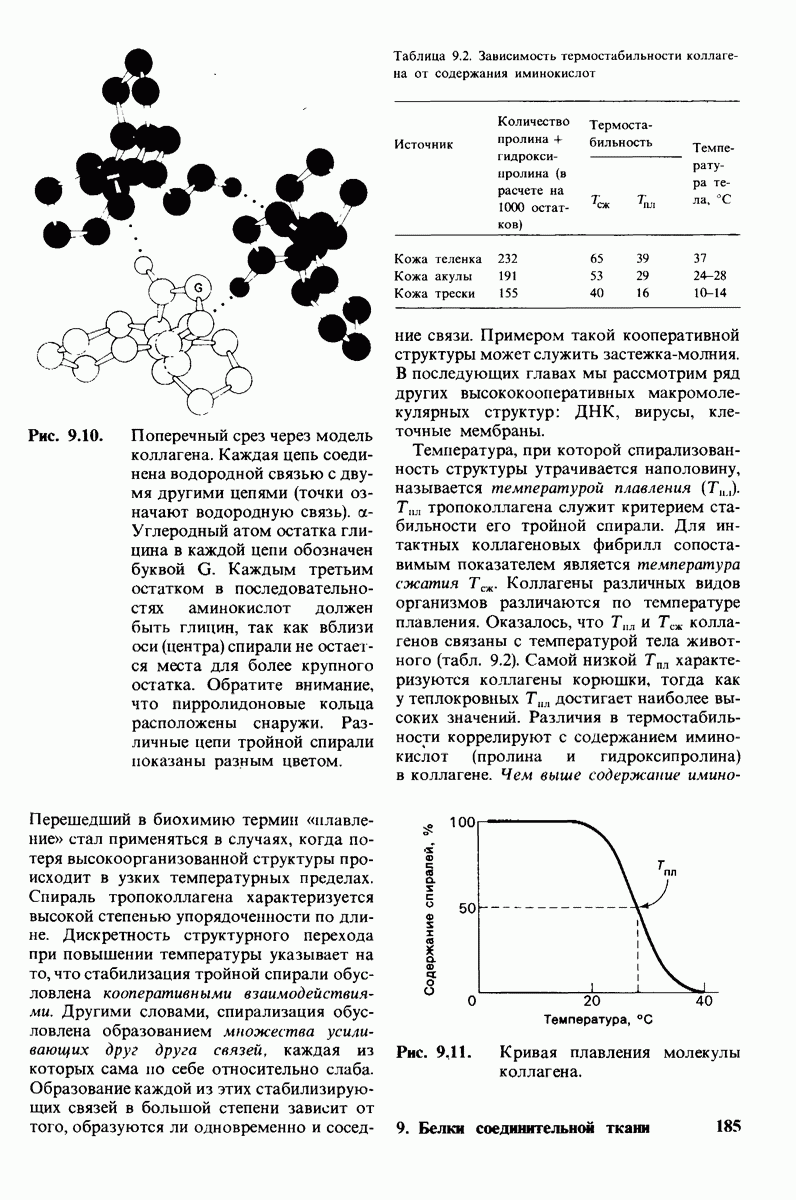
Теперь мы можем оценить значение индуцированных субстратом структурных изменений в активном центре карбоксипептидазы А. В результате связавшийся на ферменте субстрат оказывается со всех сторон окруженным каталитическими группами. Это обеспечивает возможность катализа по причинам, о которых говорилось выше. Совершенно очевидно, что только гибкость структуры фермента обеспечивает попадание субстрата в сферу действия системы каталитических групп (и выход продуктов реакции из этой системы). В целом гибкая структура фермента имеет преимущество перед жесткой в том отношении, что она обладает гораздо большим выбором возможных конформаций, пригодных для катализа и сохраняющихся в процессе отбора. Кроме того, индуцированное соответствие вносит вклад в повышение специфичности фермента. В самом деле, в случае карбоксипептидазы А субстрат должен иметь концевой карбоксилат-ион; фермент «проверяет» его наличие таким путем: если концевой карбоксилат-ион имеется, то он образует солевую связь с аргинином-145, а это вызывает перемещение тирозина-248 в каталитически активное положение; если же концевого карбоксилат-иона нет, то тирозин-248 остается на месте и фермент не проявляет активности. Другими словами, индукция соответствия может функционировать как дшшмический процесс узнавания.

Рис. 7.29. Второй возможный механизм каталитического действия карбоксипептидазы А. Туг 248 выполняет ту же функцию, что и на рис. 7.28. В остальном процесс протекает иначе:  активирует молекулу воды, которая атакует карбонильный атом углерода расщепляемой пептидной связи. Гидролиз осуществляется прямо, без промежуточного образования ангидрида.
активирует молекулу воды, которая атакует карбонильный атом углерода расщепляемой пептидной связи. Гидролиз осуществляется прямо, без промежуточного образования ангидрида.
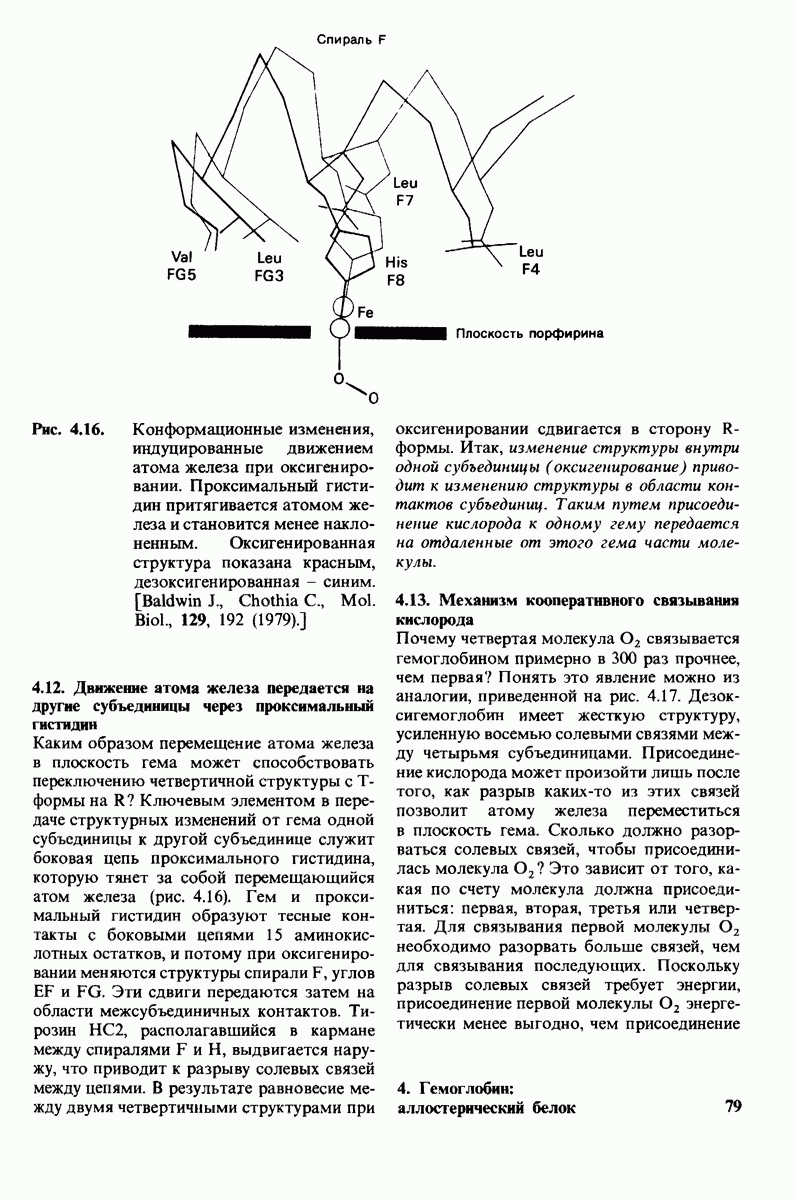
Заключение
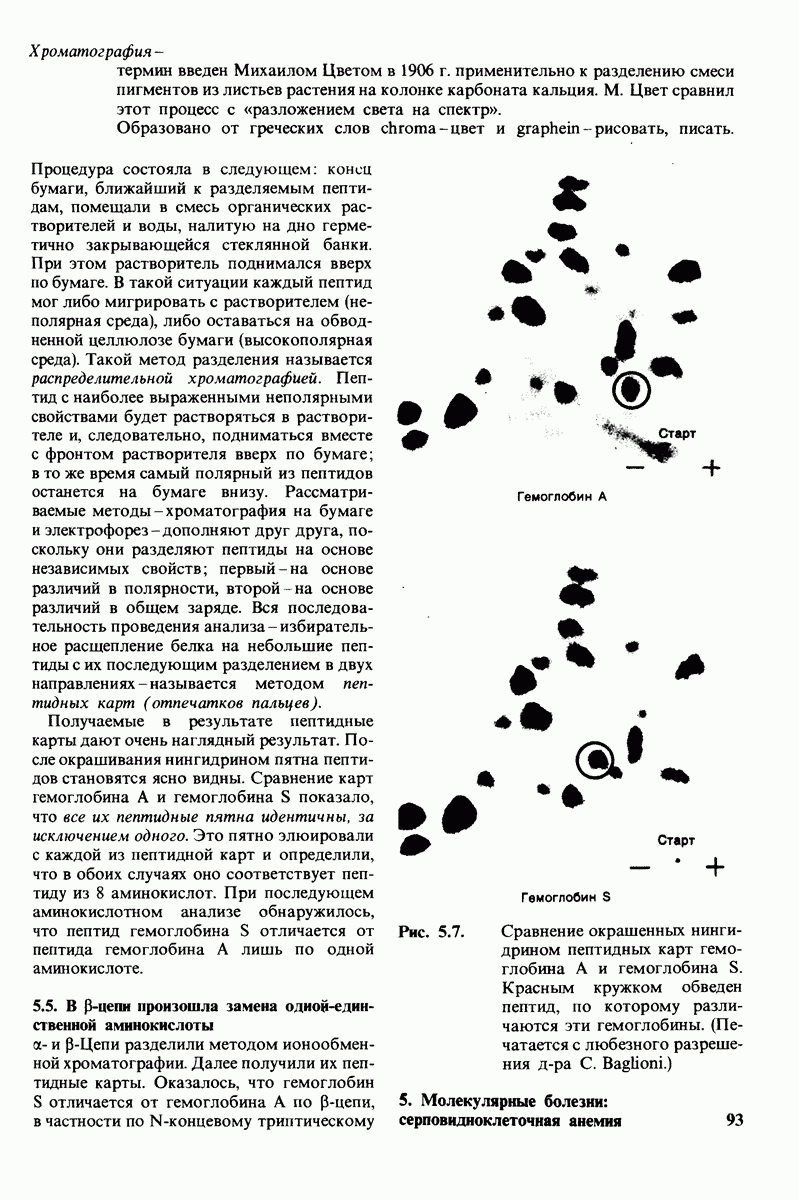
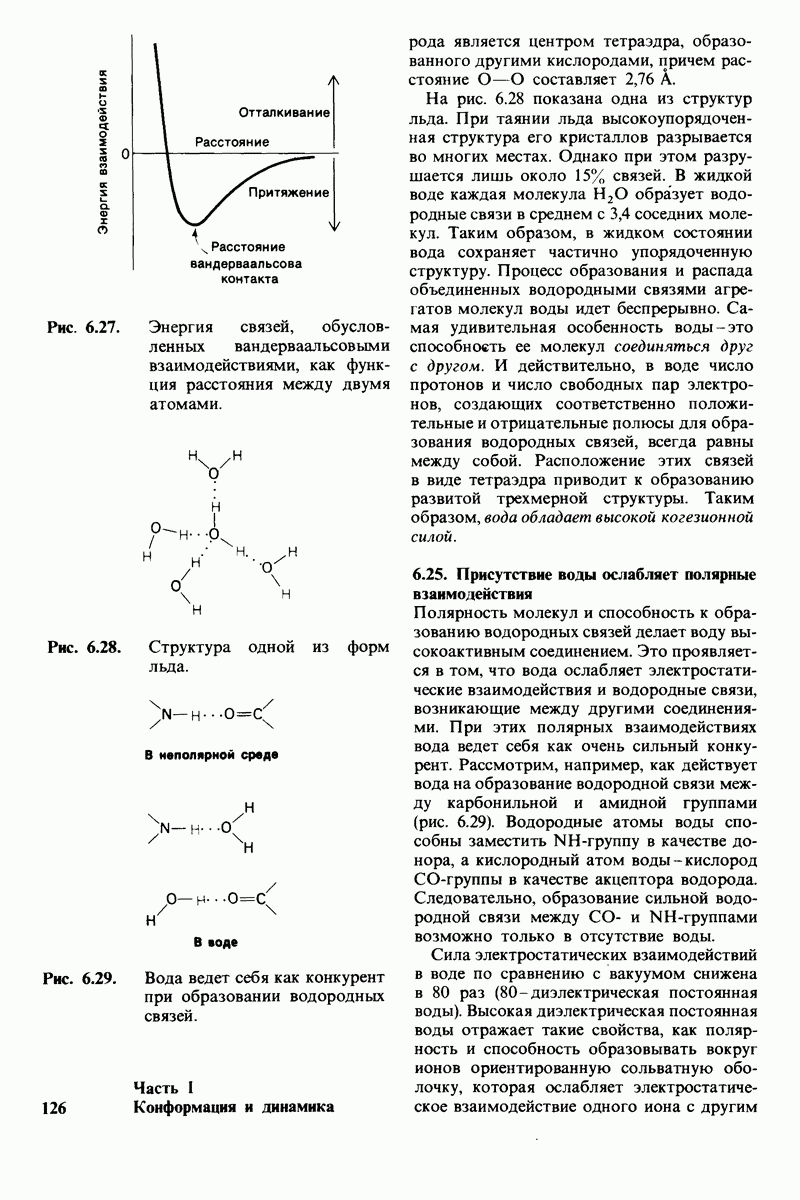
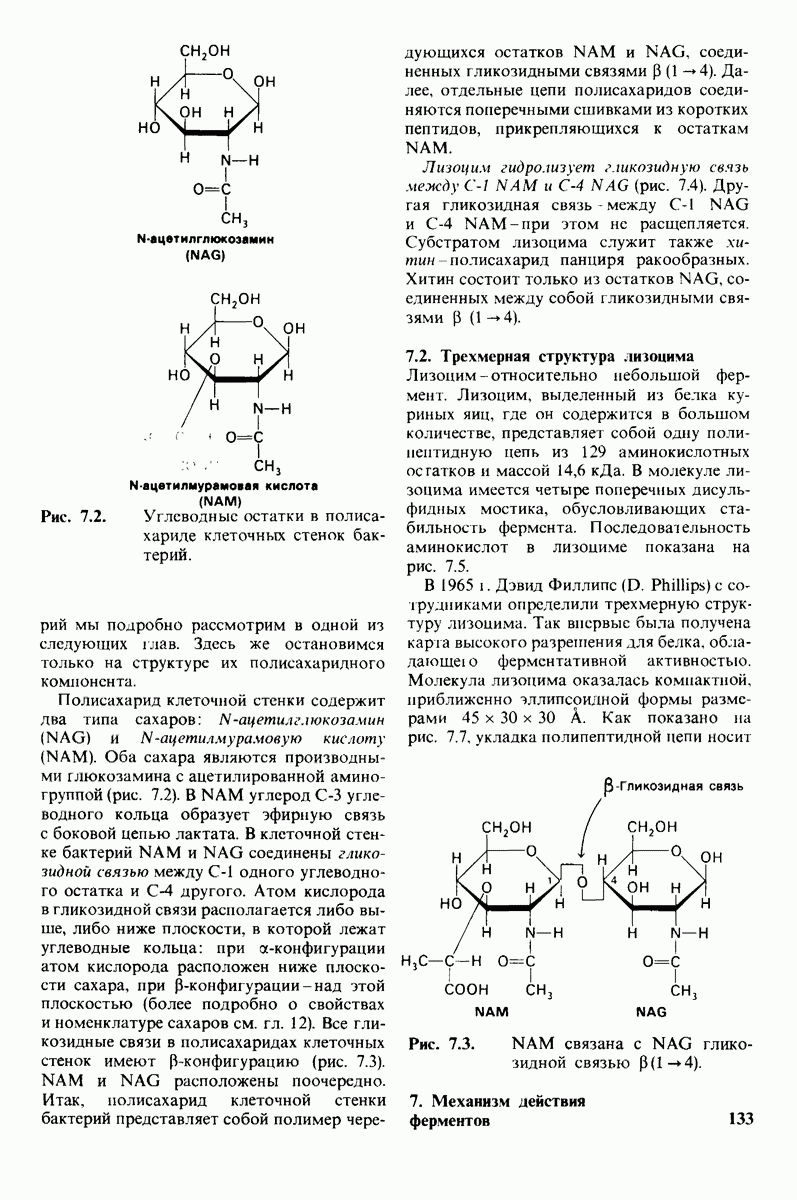
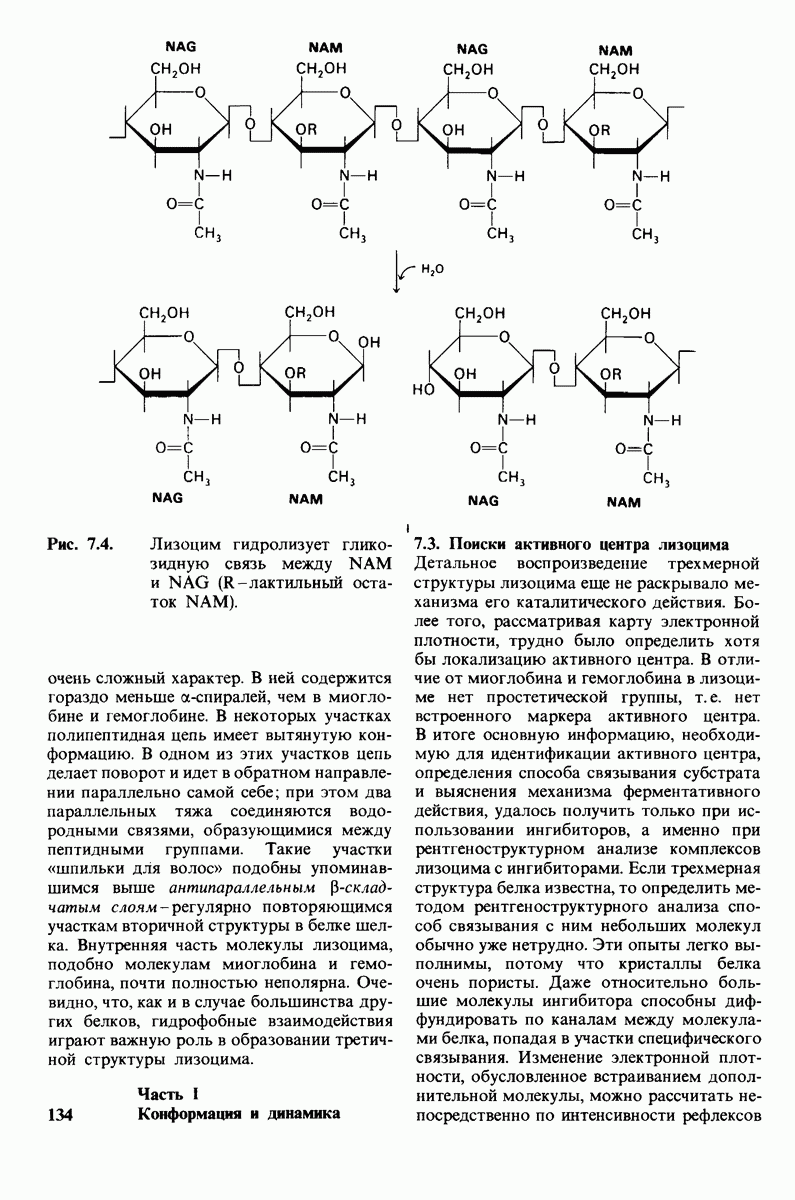
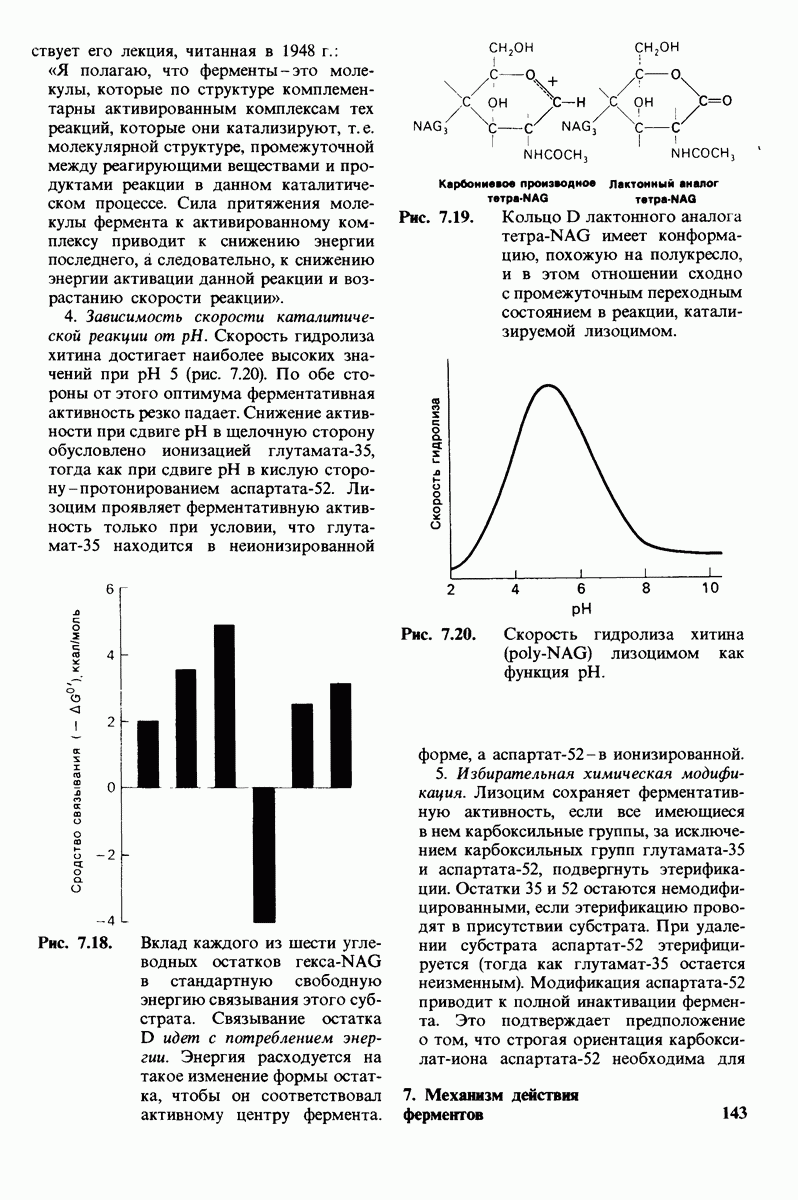
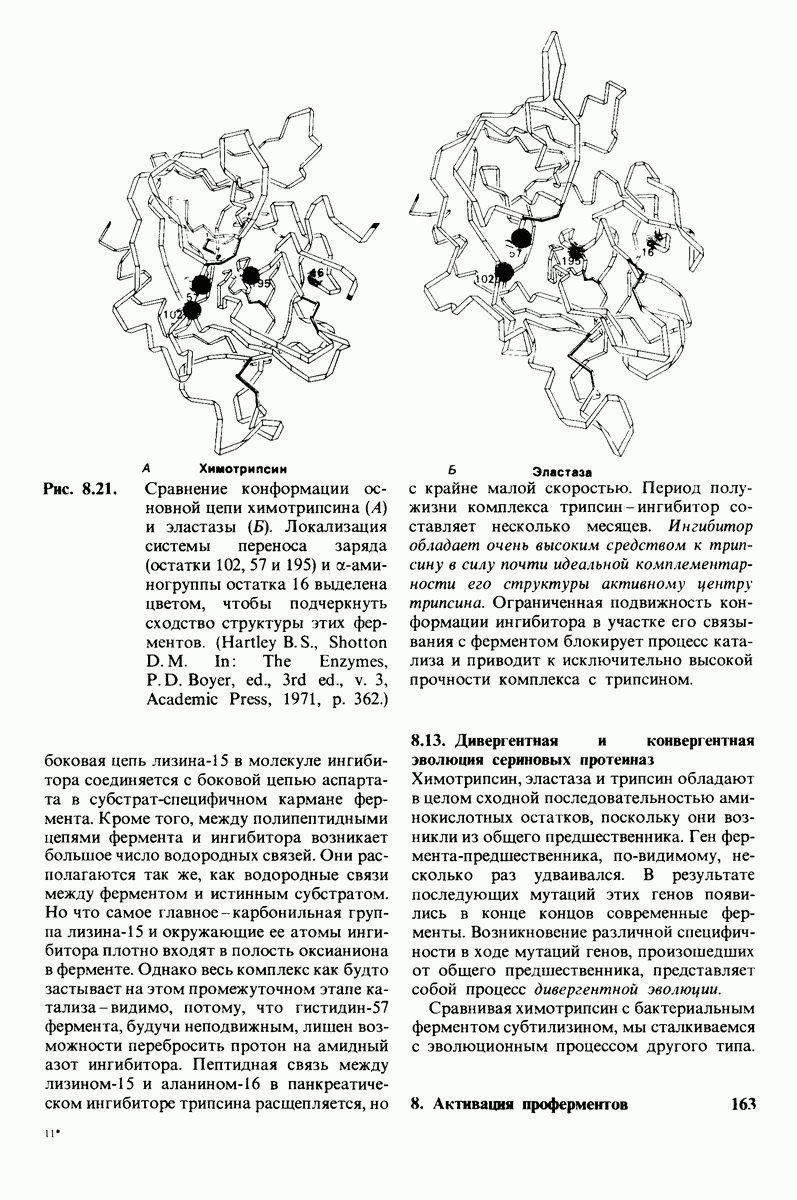
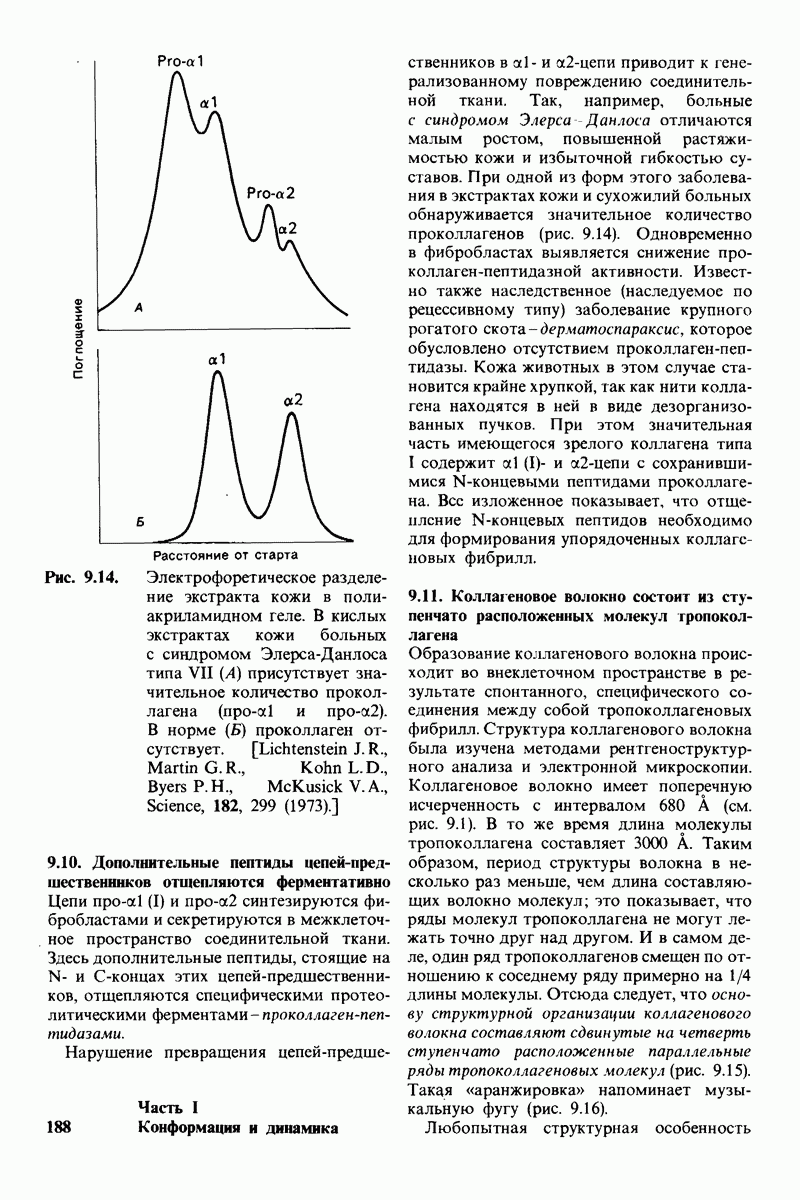
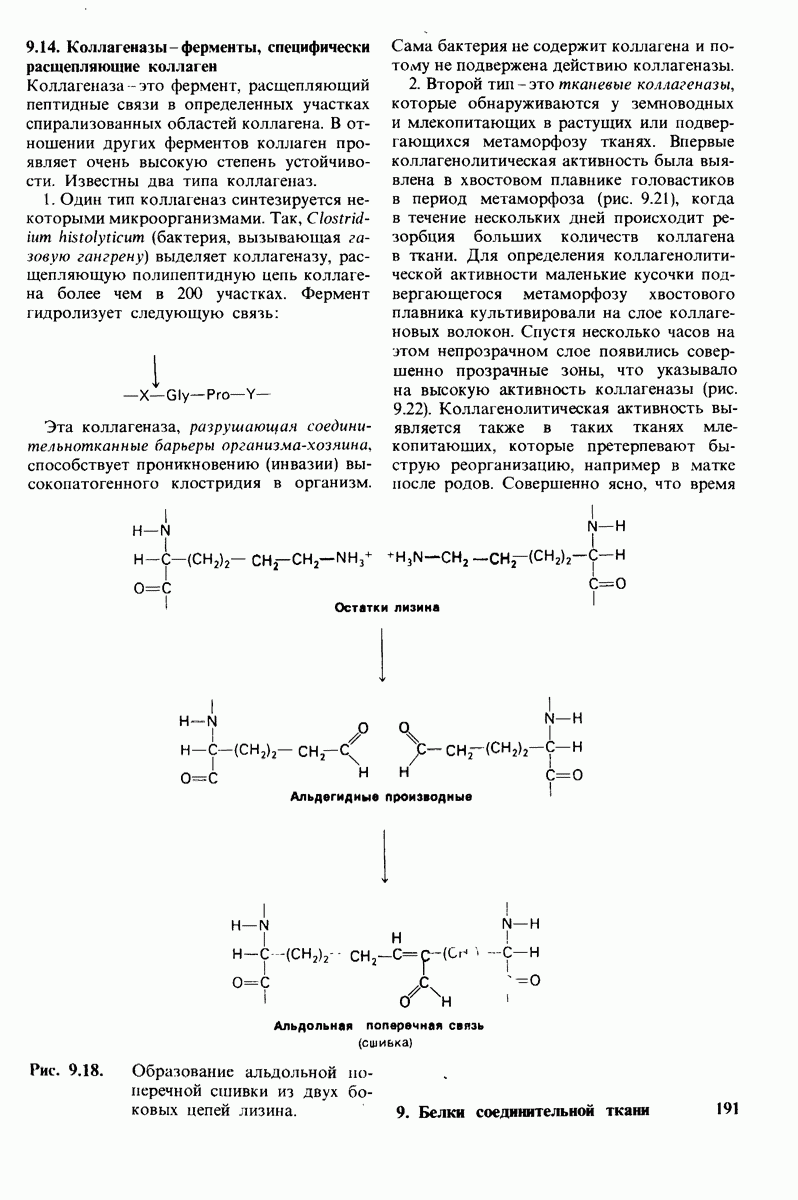
Лизоцим-фермент относительно небольшого размера, расщепляющий полисахаридный компонент клеточных стенок бактерий. По своей структуре указанный полисахарид представляет собой чередующийся полимер остатков N-ацетил глюкозамина (NAG) и N-ацетилмурамовой кислоты
 соединенных гликозидными связями
соединенных гликозидными связями  Лизоцим гидролизует глизозид-ную связь между
Лизоцим гидролизует глизозид-ную связь между  остатка NAM и
остатка NAM и  остатка NAG. Олигомеры N-ацетилглюко-замина также гидролизуются лизоцимом. При этом
остатка NAG. Олигомеры N-ацетилглюко-замина также гидролизуются лизоцимом. При этом  и более длинные полимеры легко расщепляются ферментом, тогда как
и более длинные полимеры легко расщепляются ферментом, тогда как  гидролизуются с крайне малой скоростью.
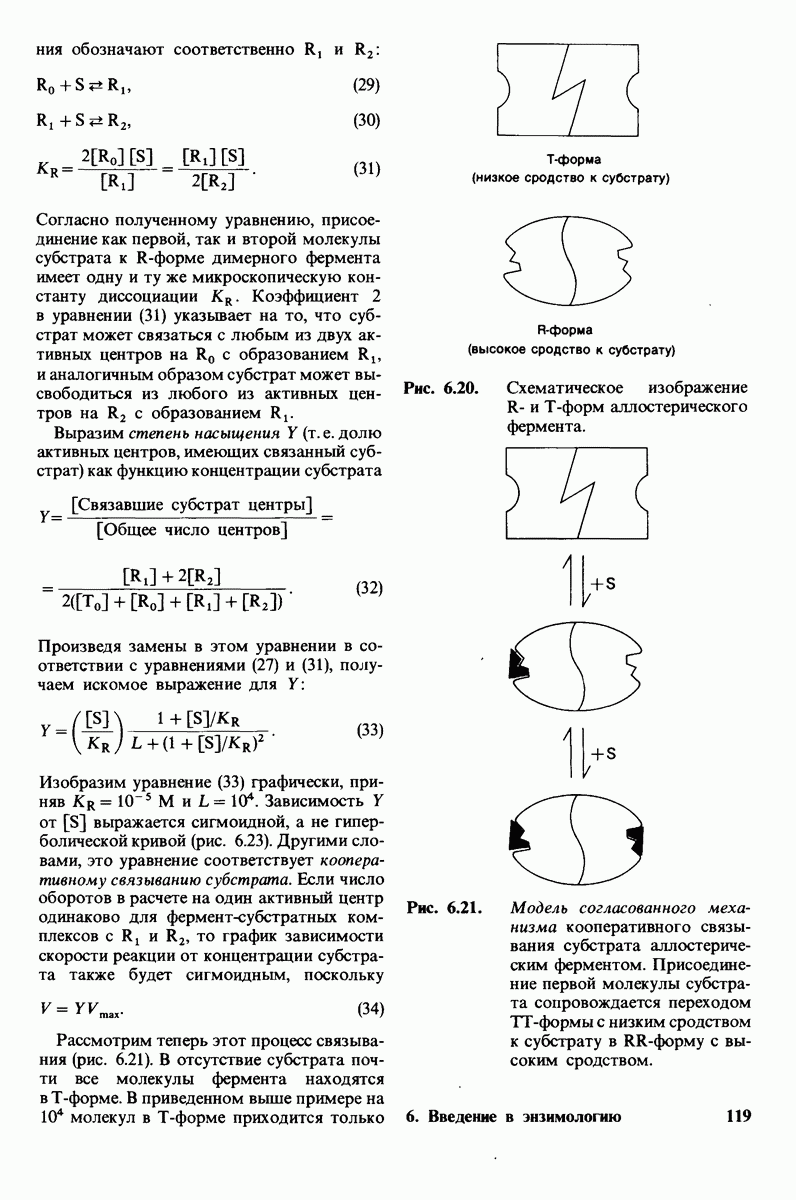
гидролизуются с крайне малой скоростью.  сильный конкурентный ингибитор фермента. Трехмерная структура лизоцима и его комплекса с три-NAG изучена на атомарном уровне. Установлено, что три-NAG занимает половину щели, идущей поперек молекулы фермента, с которым он связывается большим количеством водородных связей и вандерваальсовых взаимодействий. На основе данных о структуре комплекса лизоцима с три-NAG была построена модель, предсказывающая, как связывается с лизоцимом его эффективный субстрат
сильный конкурентный ингибитор фермента. Трехмерная структура лизоцима и его комплекса с три-NAG изучена на атомарном уровне. Установлено, что три-NAG занимает половину щели, идущей поперек молекулы фермента, с которым он связывается большим количеством водородных связей и вандерваальсовых взаимодействий. На основе данных о структуре комплекса лизоцима с три-NAG была построена модель, предсказывающая, как связывается с лизоцимом его эффективный субстрат 
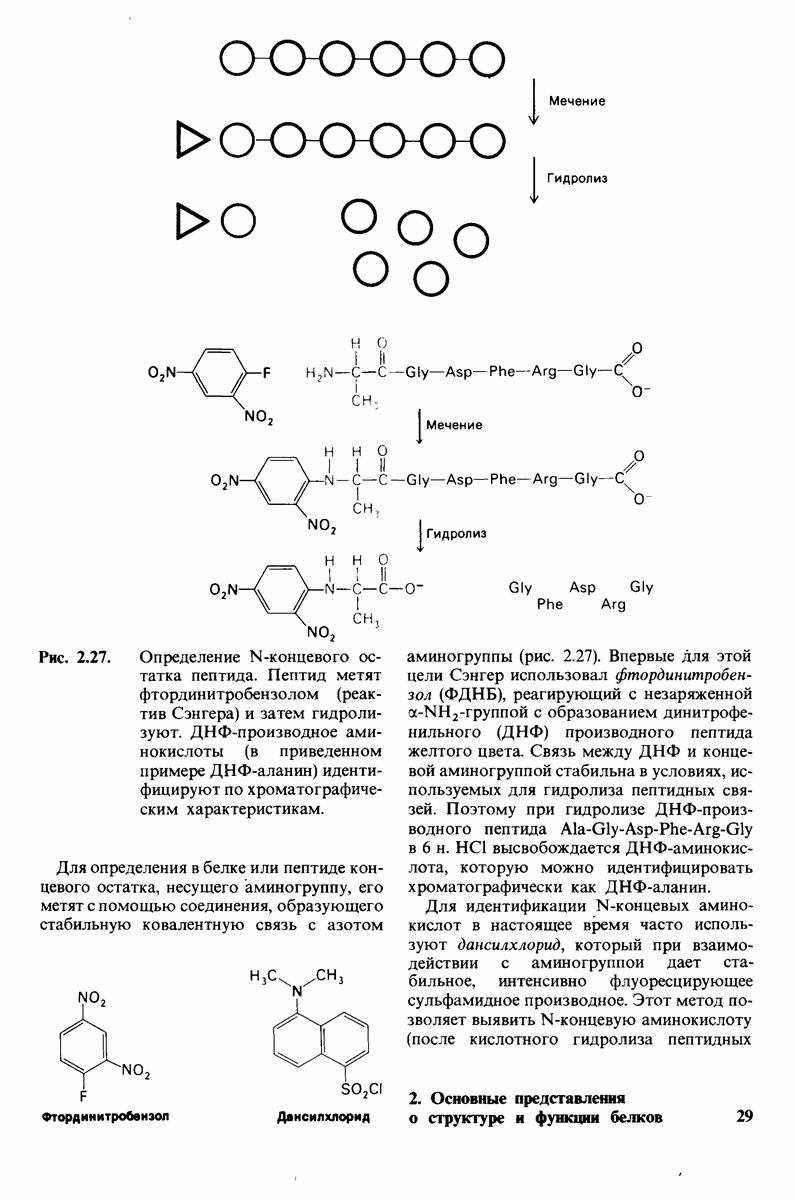
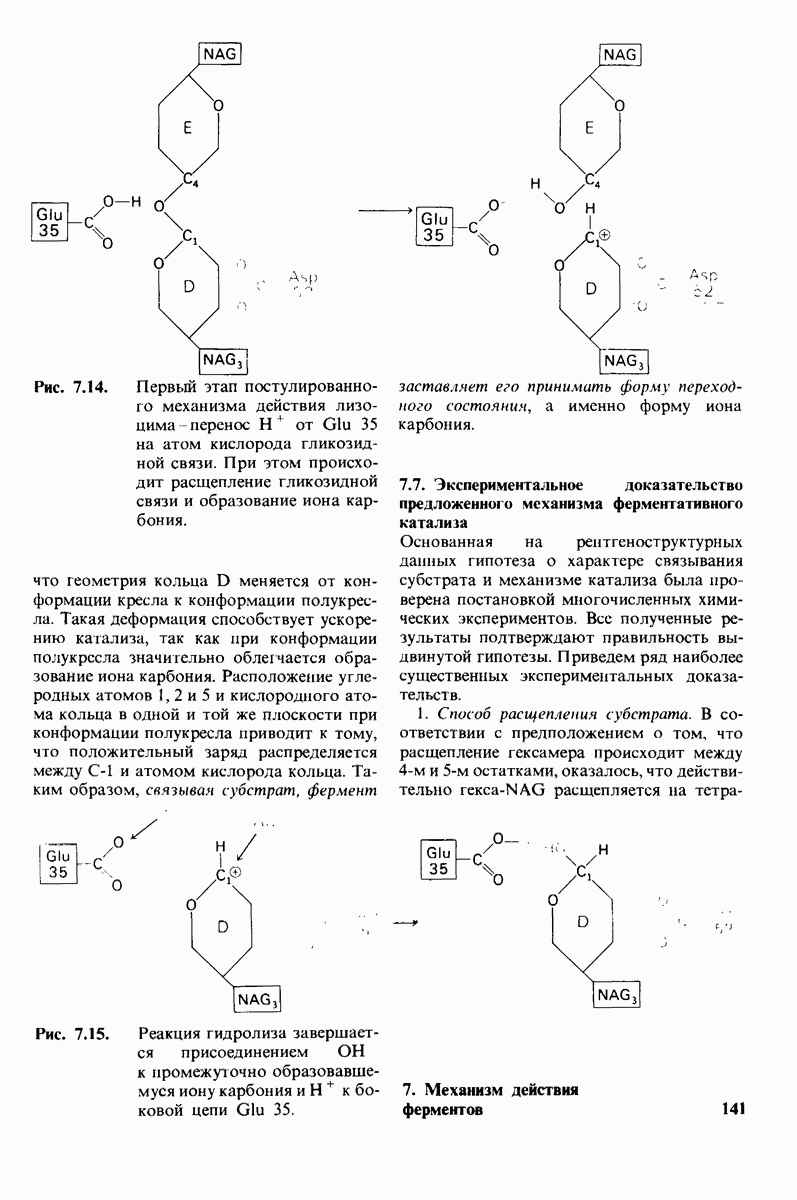
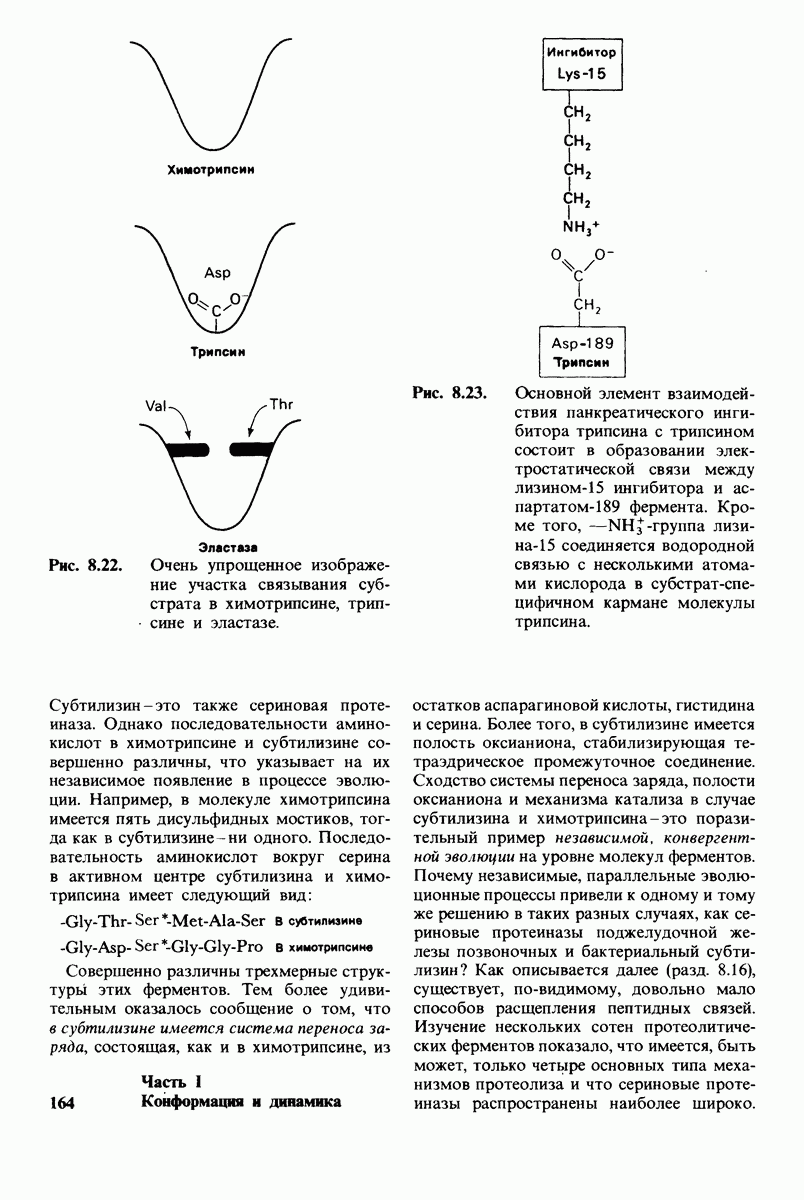
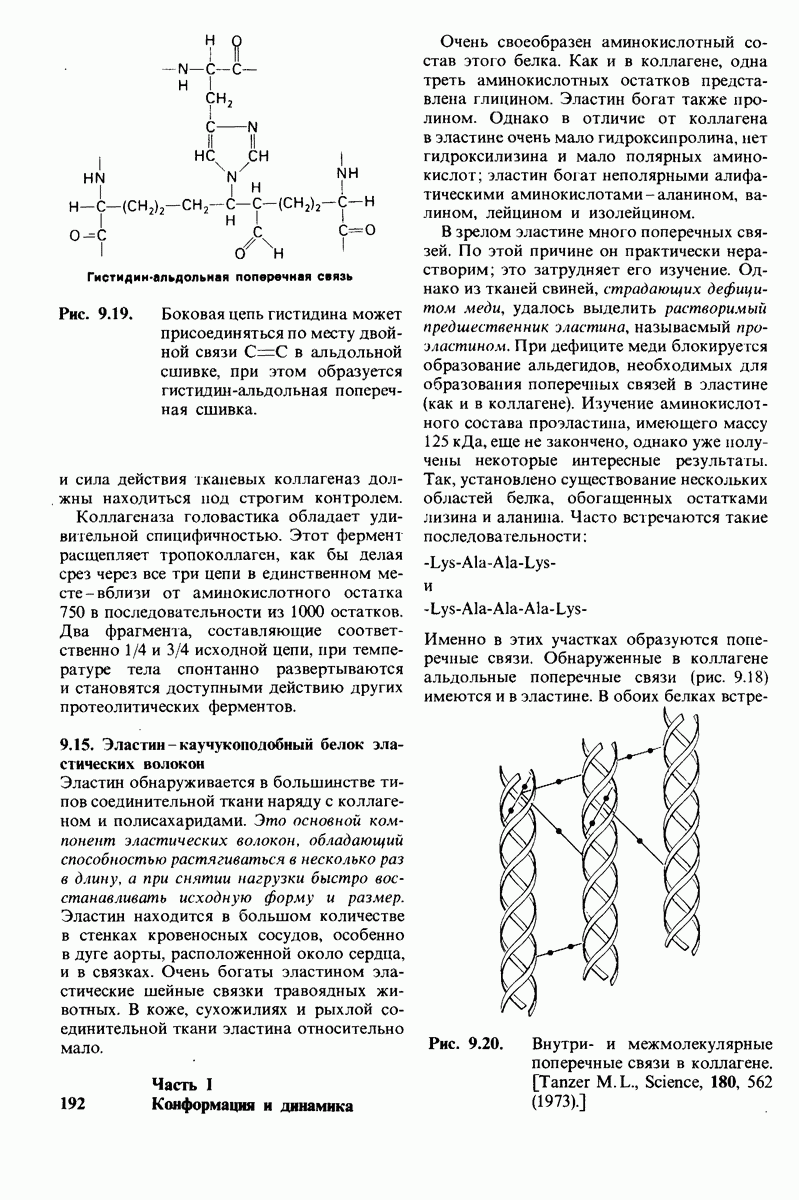
Предложена гипотеза механизма каталитического действия лизоцима; суть гипотезы состоит в следующем. Первое; критическими группами для катализа являются неионизированная карбоксильная группа остатка глутамата-35 и карбоксилат-ион аспартата-52. Обе группы расположены на расстоянии около  от гидролизуемой гликозидной связи, а именно связи между остатками
от гидролизуемой гликозидной связи, а именно связи между остатками  гексамерного субстрата. Второе: глутамат-35 отдает
гексамерного субстрата. Второе: глутамат-35 отдает  связи между
связи между  кольца D и гликозидным атомом кислорода, что приводит к разрыву этой связи.
кольца D и гликозидным атомом кислорода, что приводит к разрыву этой связи.  кольца D становится положительно заряженным; такая переходная форма называется ионом карбония. Третье; ион карбония реагирует с
кольца D становится положительно заряженным; такая переходная форма называется ионом карбония. Третье; ион карбония реагирует с  -группой растворителя, а глутамат-35 присоединяет водород, возвращаясь в исходную протонированную форму. После того как продукты реакции удаляются от фермента в результате диффузии, лизоцим готов к новому каталитическому циклу. Четвертое; скорость катализа значительно увеличивается под действием двух факторов, способствующих промежуточному образованию иона карбония: это электростатический фактор, а именно близость отрицательно заряженной боковой цепи аспартата-52 и геометрический фактор, состоящий в том, что кольцо D деформируется, приобретая конформацию полукресла, что приводит к распределению положительного заряда иона карбония между
-группой растворителя, а глутамат-35 присоединяет водород, возвращаясь в исходную протонированную форму. После того как продукты реакции удаляются от фермента в результате диффузии, лизоцим готов к новому каталитическому циклу. Четвертое; скорость катализа значительно увеличивается под действием двух факторов, способствующих промежуточному образованию иона карбония: это электростатический фактор, а именно близость отрицательно заряженной боковой цепи аспартата-52 и геометрический фактор, состоящий в том, что кольцо D деформируется, приобретая конформацию полукресла, что приводит к распределению положительного заряда иона карбония между  и атомом кислорода углеводного кольца.
и атомом кислорода углеводного кольца.
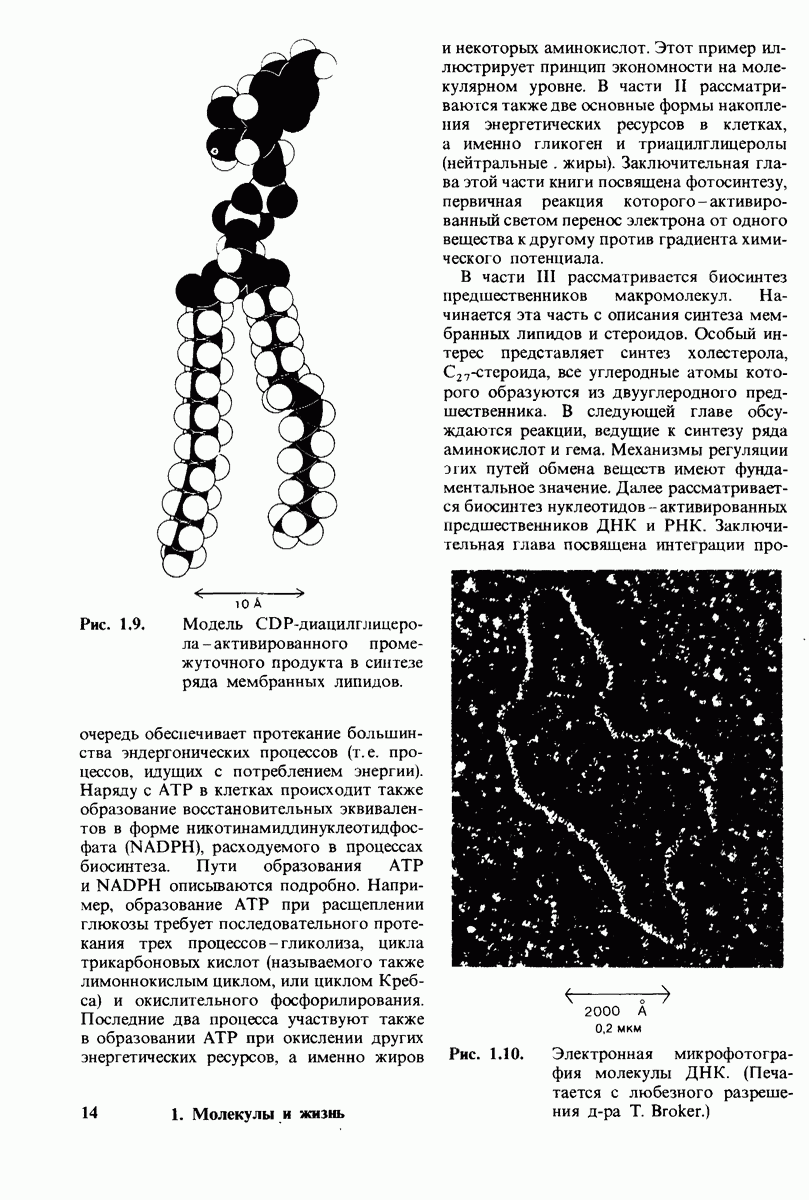
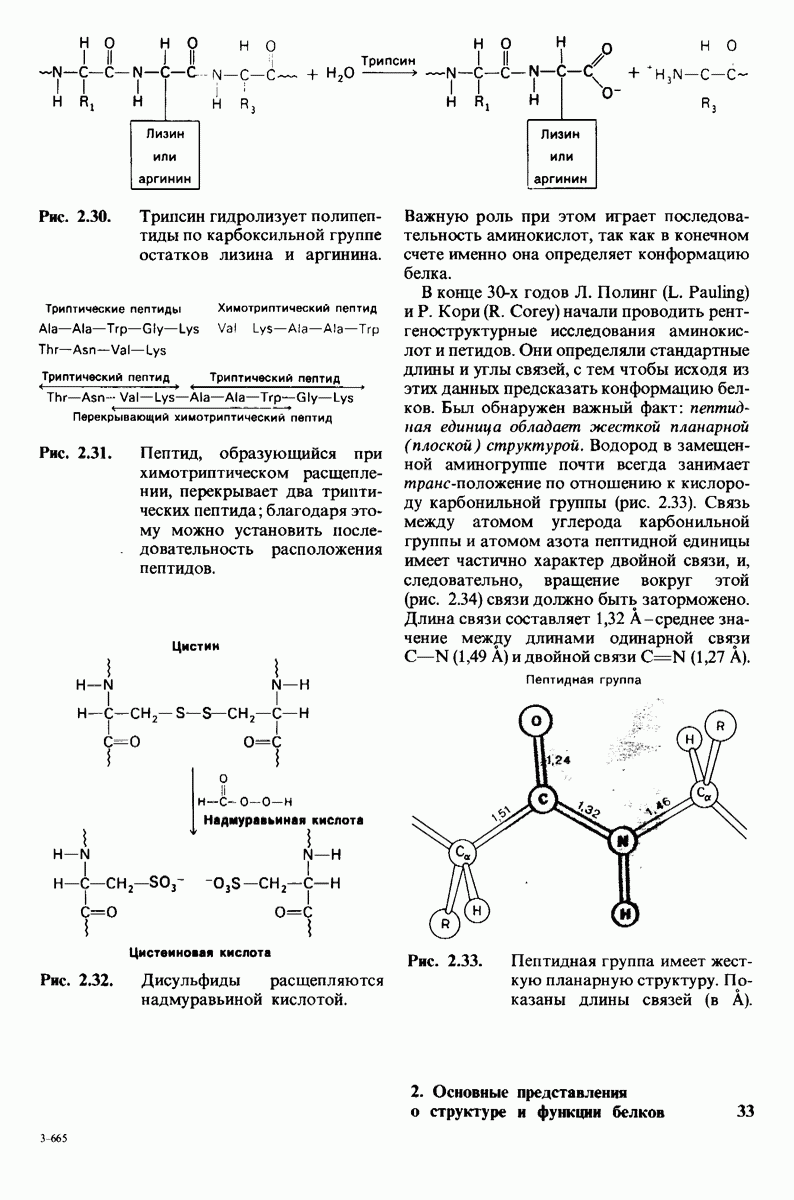
Карбоксипептидаза А, пищеварительный фермент, расщепляющий  -концевой пептид в полипептидах, служит примером фермента, в основе каталитического действия которого лежит совершенно иной механизм. Структура этого фермента и его комплекса с глицилтирозином - аналогом субстрата - раскрыта на уровне атомного разрешения. Связывание глипилтирозина вызывает большие структурные изменения в области активного центра, в результате которых эта область теряет воду и становится гидрофобной. Пример карбоксипептидазы А иллюстрирует ту важную роль, которую играет в катализе индуцированное соответствие формы фермента форме субстрата. Другая примечательная особенность данного фермента состоит в том, что в его активном центре содержится ион цинка, имеющий существенное значение для катализа. Карбонильный атом углерода расщепляемой пептидной связи поляризуется цинком и становится более чувствительным к нуклеофиль-ной атаке. Здесь мы видим пример индукции смещения электронов в субстрате. При катализе карбоксипептидазой А молекула воды, активированная глутаматом-270, непосредственно атакует карбонильную группу расщепляемой пептидной связи; одновременно тирозин-248 отдает протон на
-концевой пептид в полипептидах, служит примером фермента, в основе каталитического действия которого лежит совершенно иной механизм. Структура этого фермента и его комплекса с глицилтирозином - аналогом субстрата - раскрыта на уровне атомного разрешения. Связывание глипилтирозина вызывает большие структурные изменения в области активного центра, в результате которых эта область теряет воду и становится гидрофобной. Пример карбоксипептидазы А иллюстрирует ту важную роль, которую играет в катализе индуцированное соответствие формы фермента форме субстрата. Другая примечательная особенность данного фермента состоит в том, что в его активном центре содержится ион цинка, имеющий существенное значение для катализа. Карбонильный атом углерода расщепляемой пептидной связи поляризуется цинком и становится более чувствительным к нуклеофиль-ной атаке. Здесь мы видим пример индукции смещения электронов в субстрате. При катализе карбоксипептидазой А молекула воды, активированная глутаматом-270, непосредственно атакует карбонильную группу расщепляемой пептидной связи; одновременно тирозин-248 отдает протон на  этой связи, и в результате происходит гидролиз.
этой связи, и в результате происходит гидролиз.
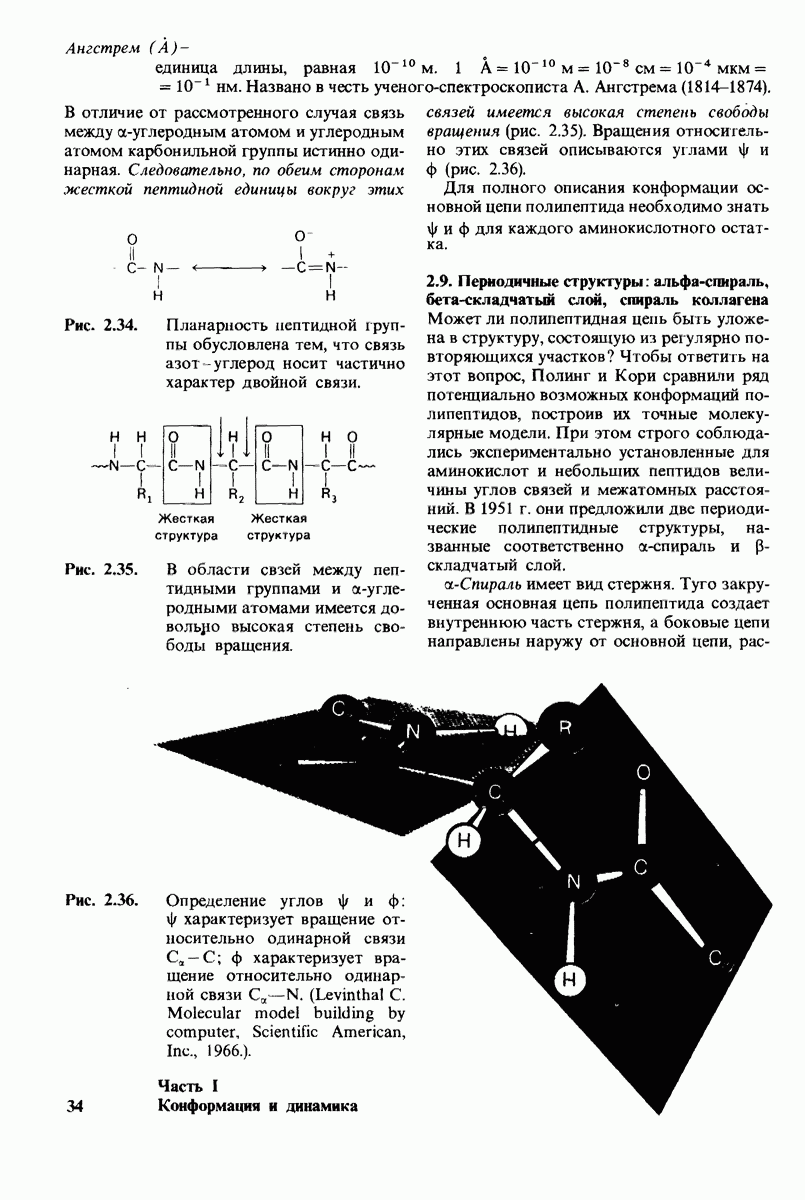
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
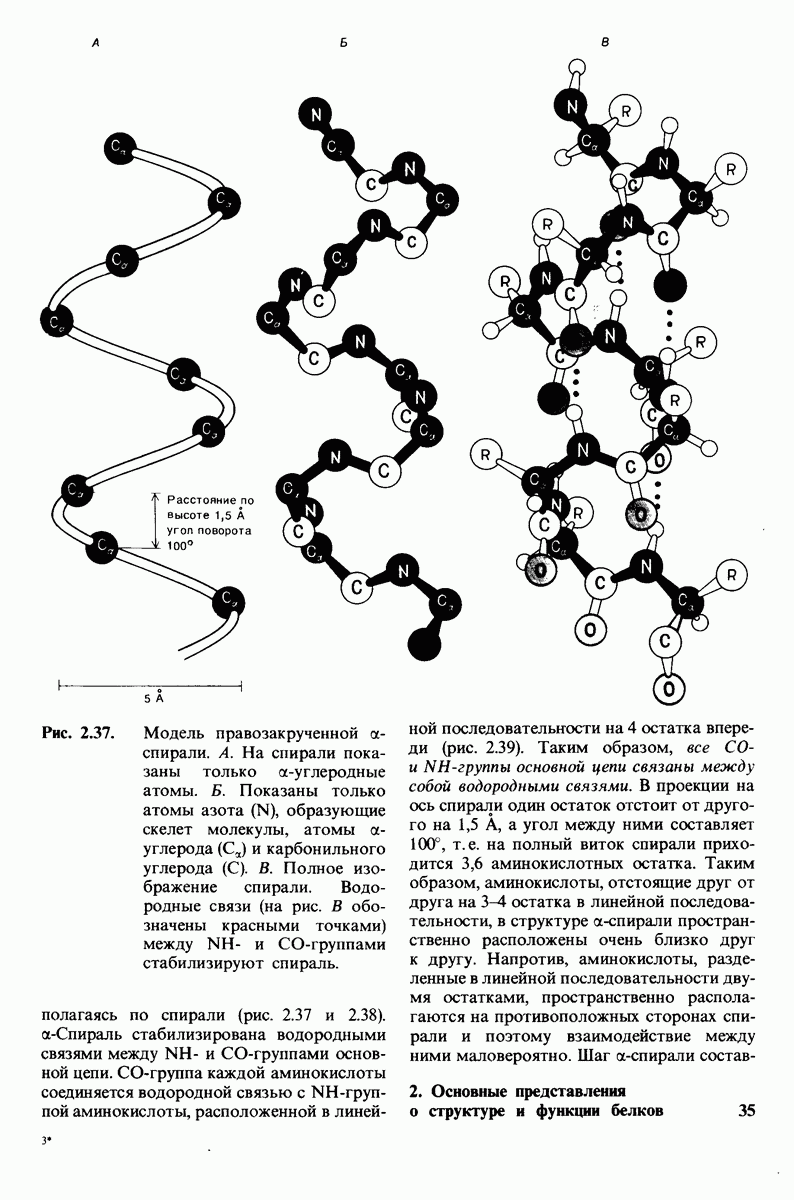
(см. скан)