
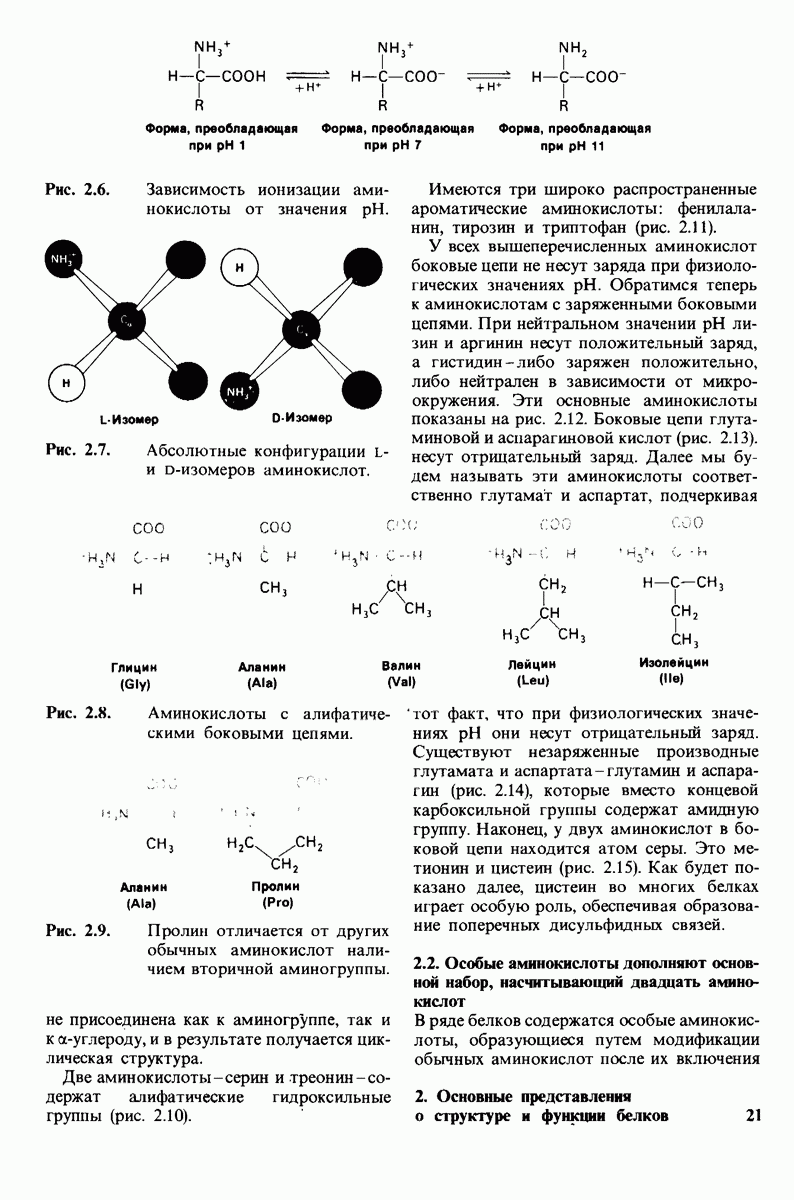
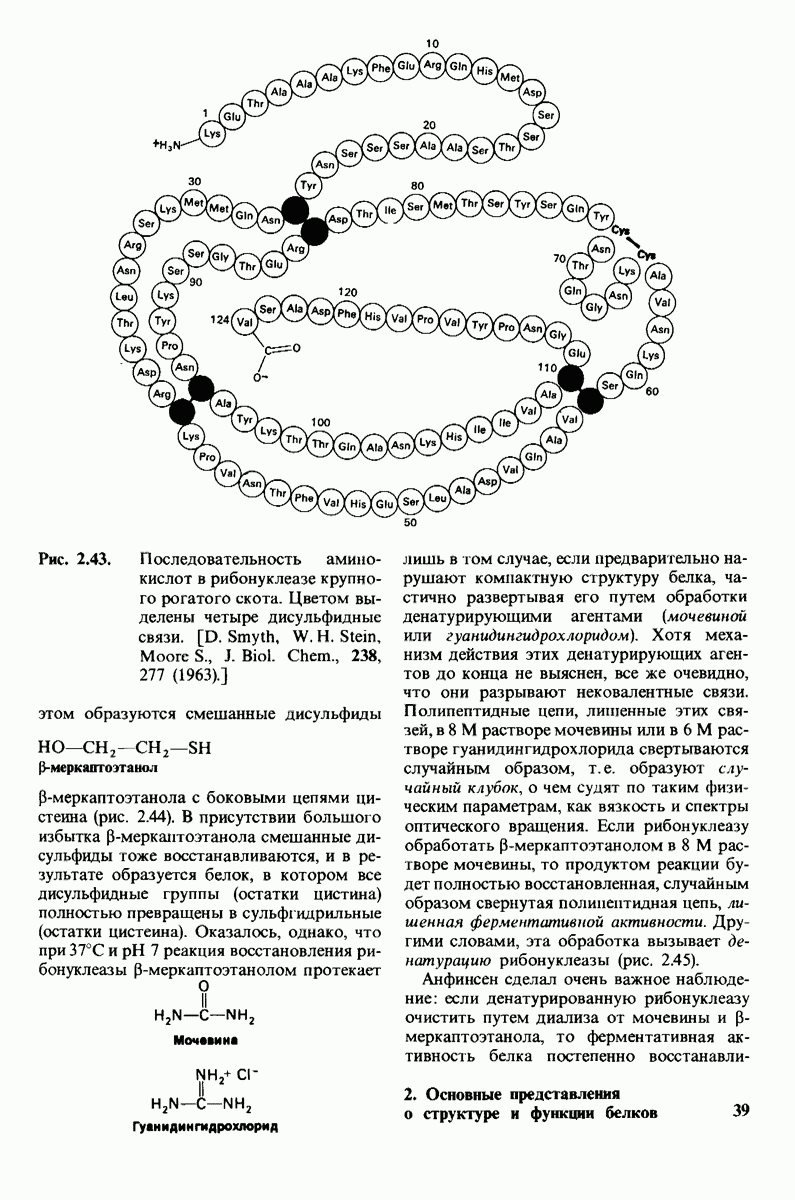
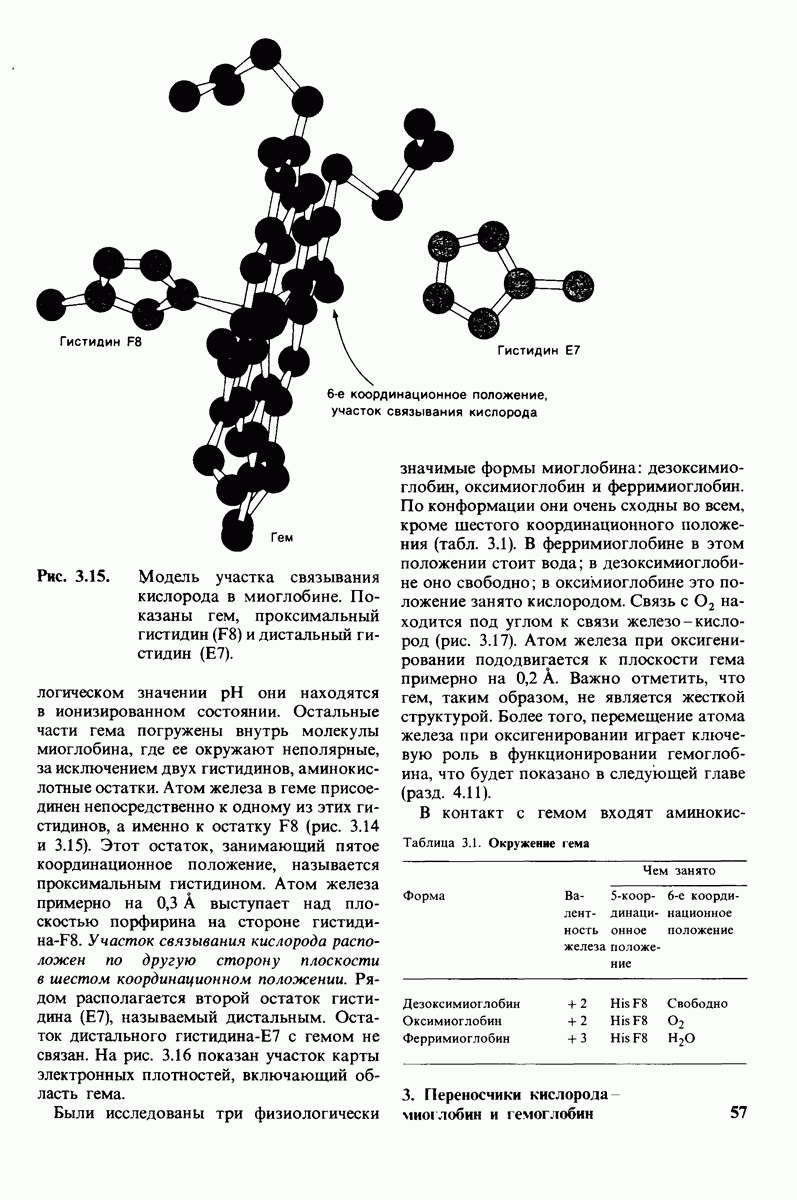
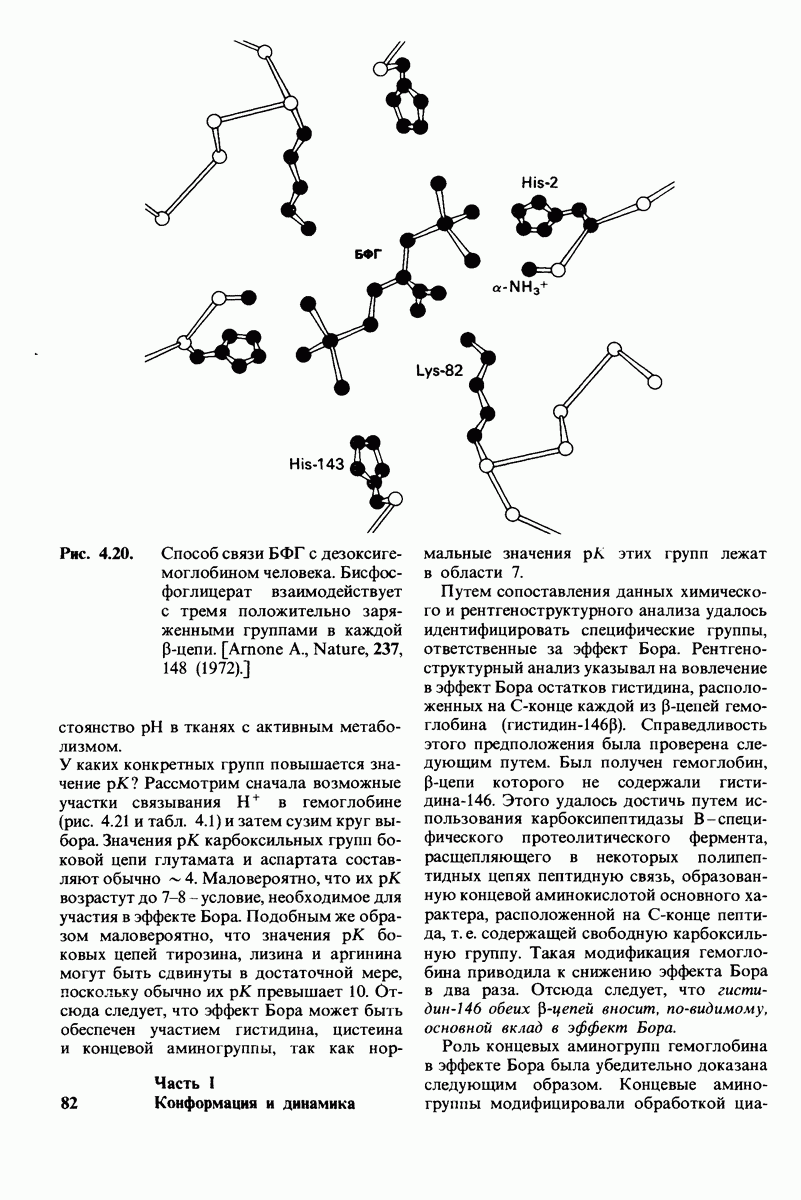
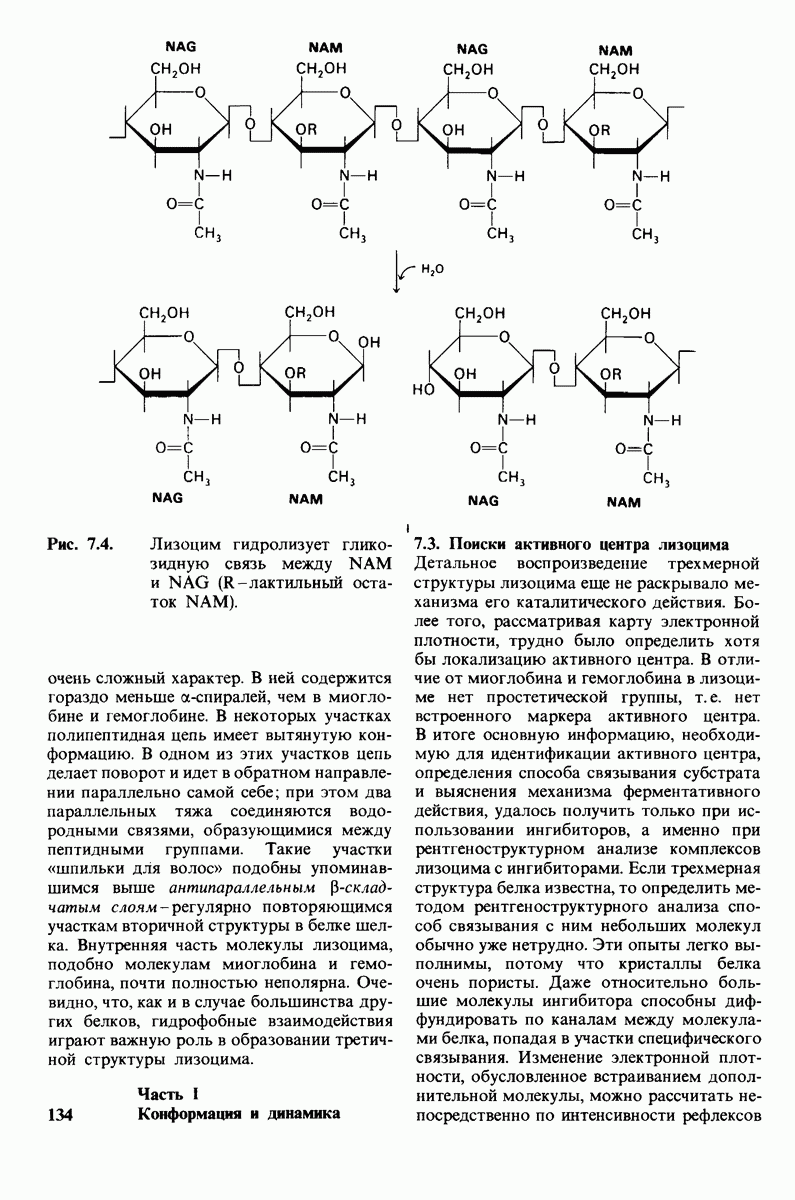
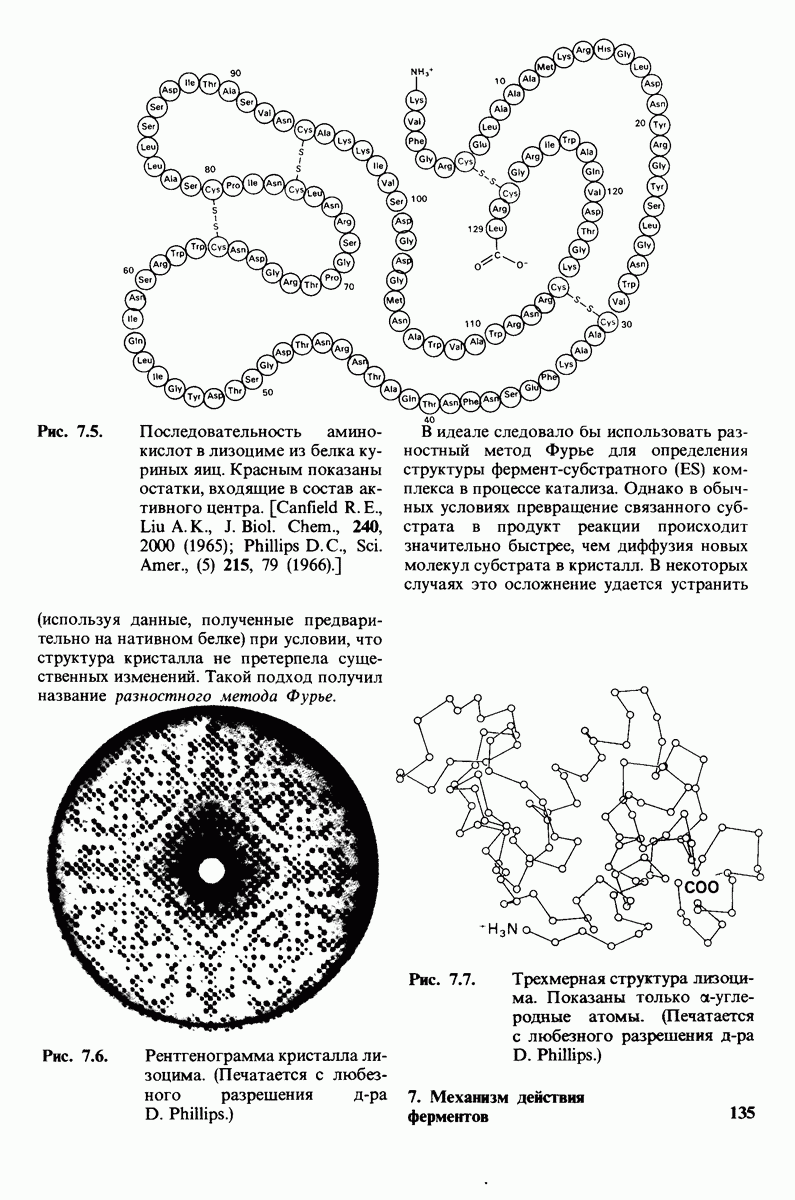
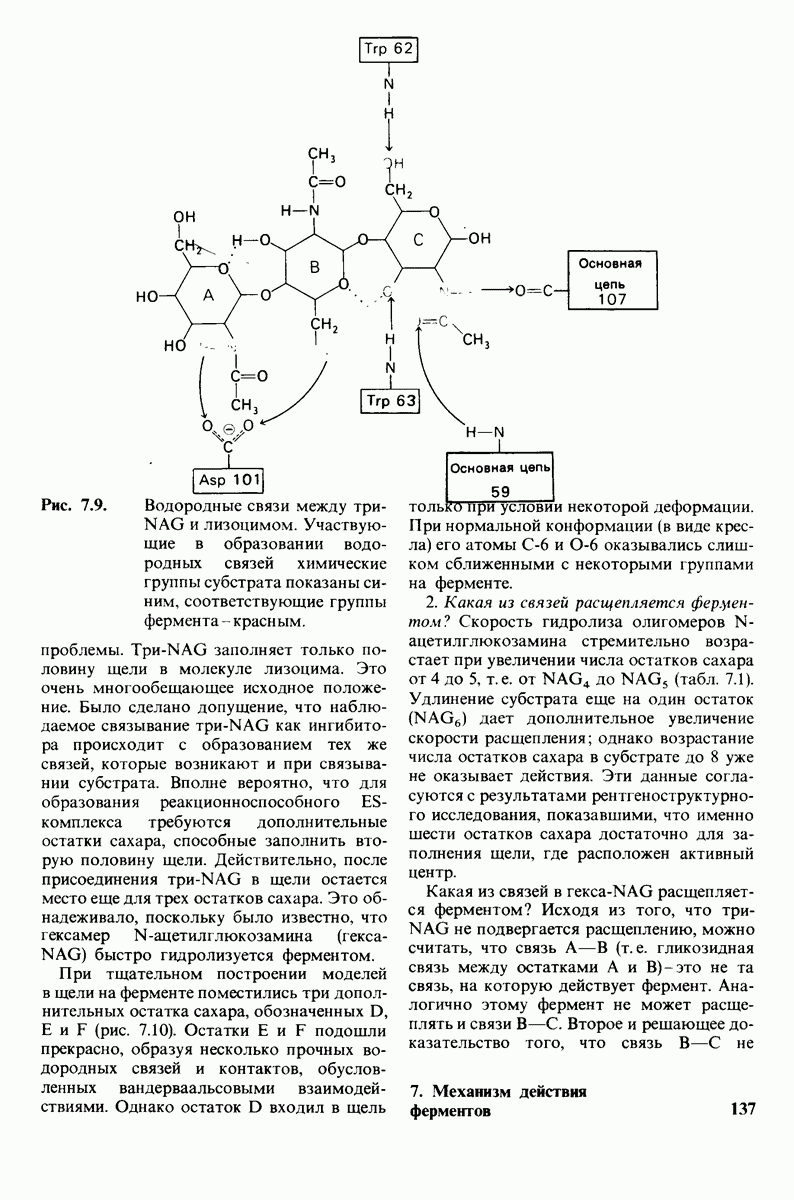
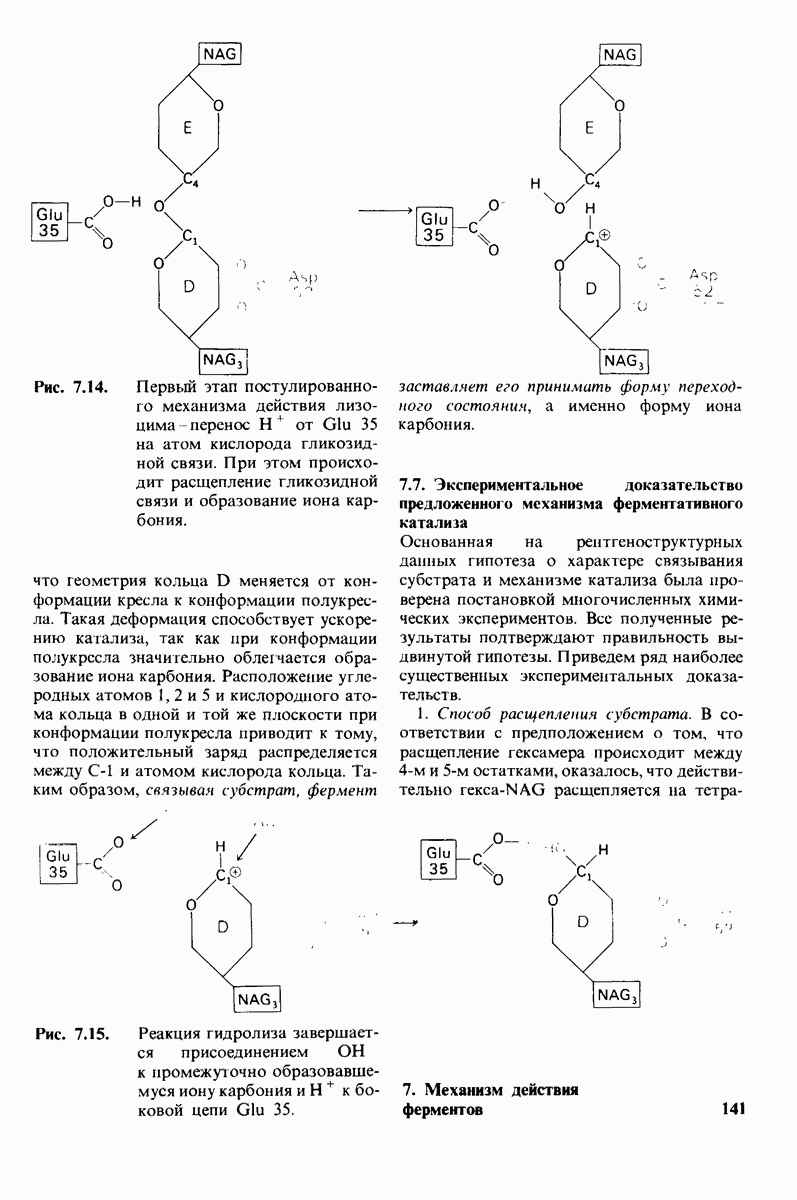
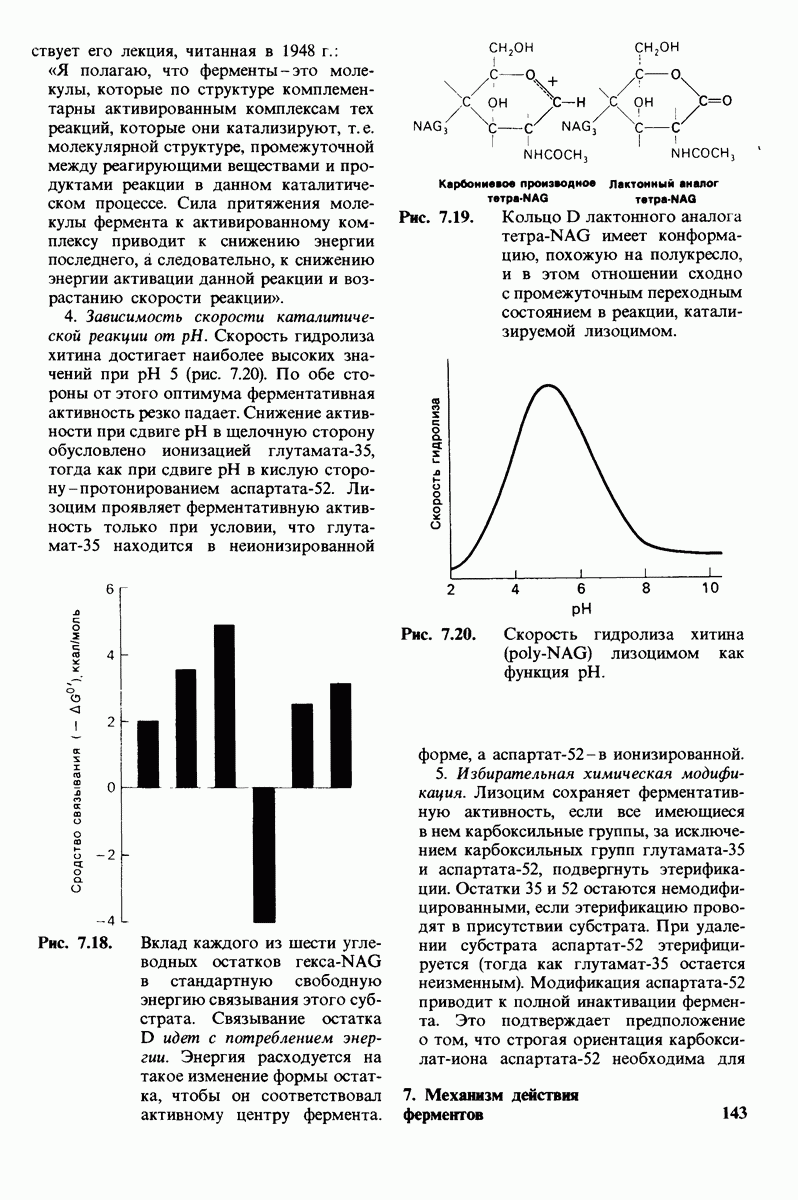
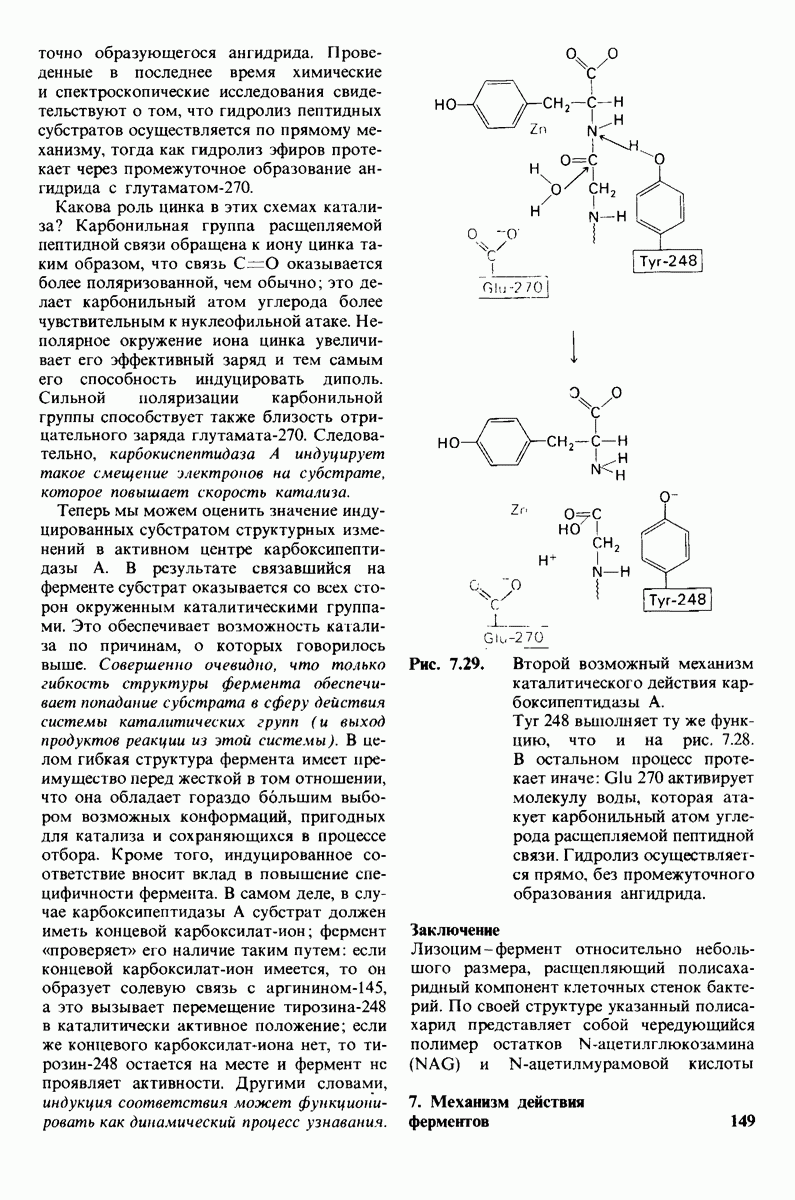
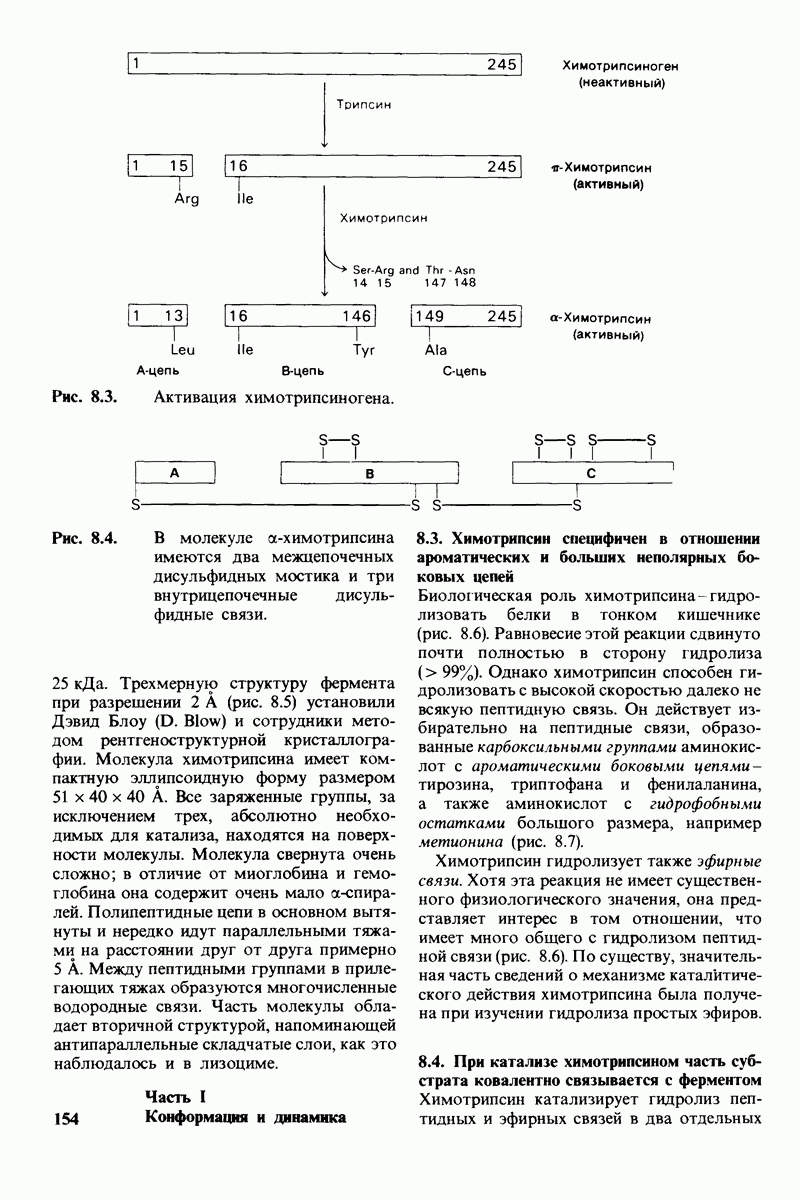
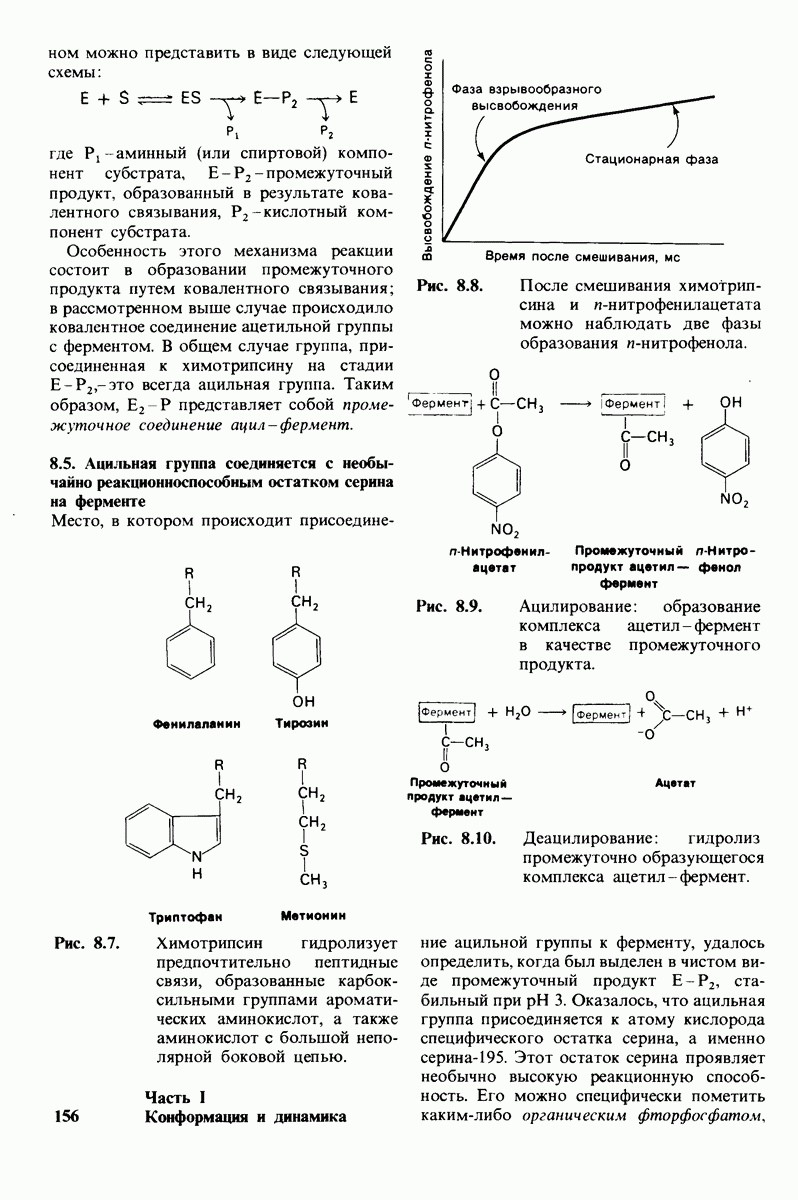
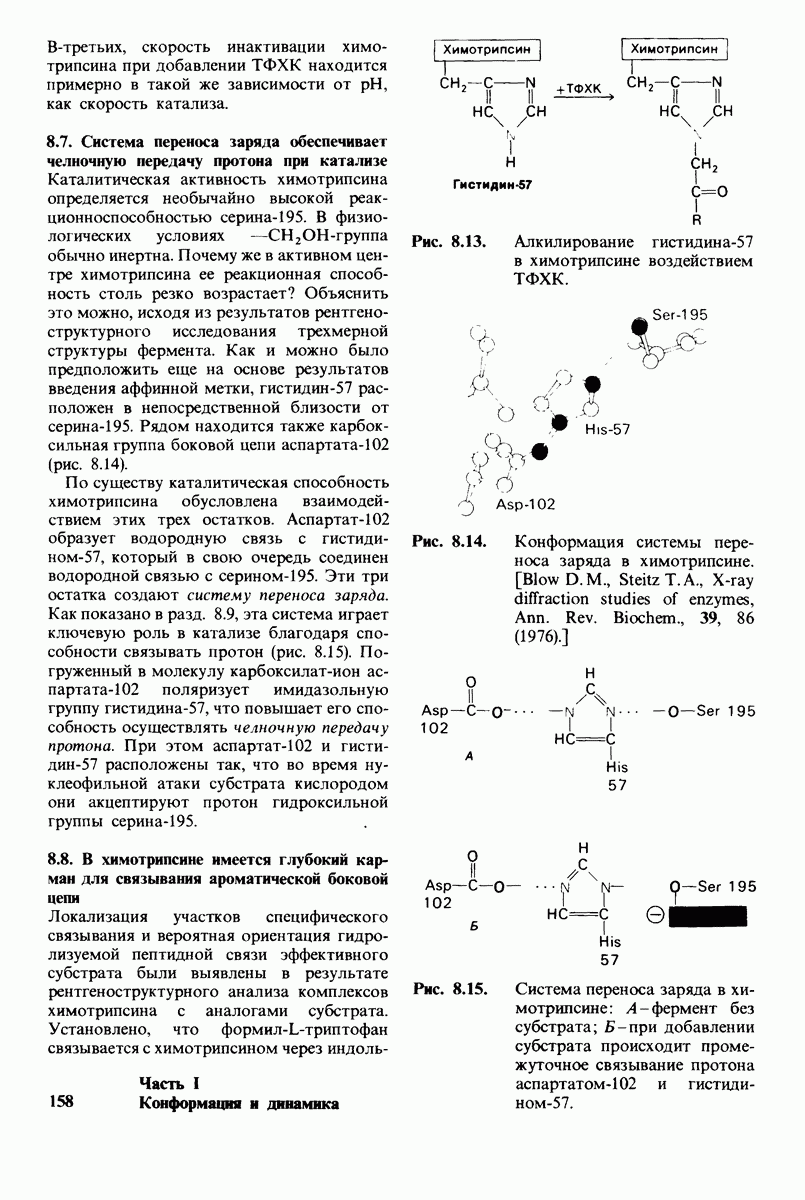
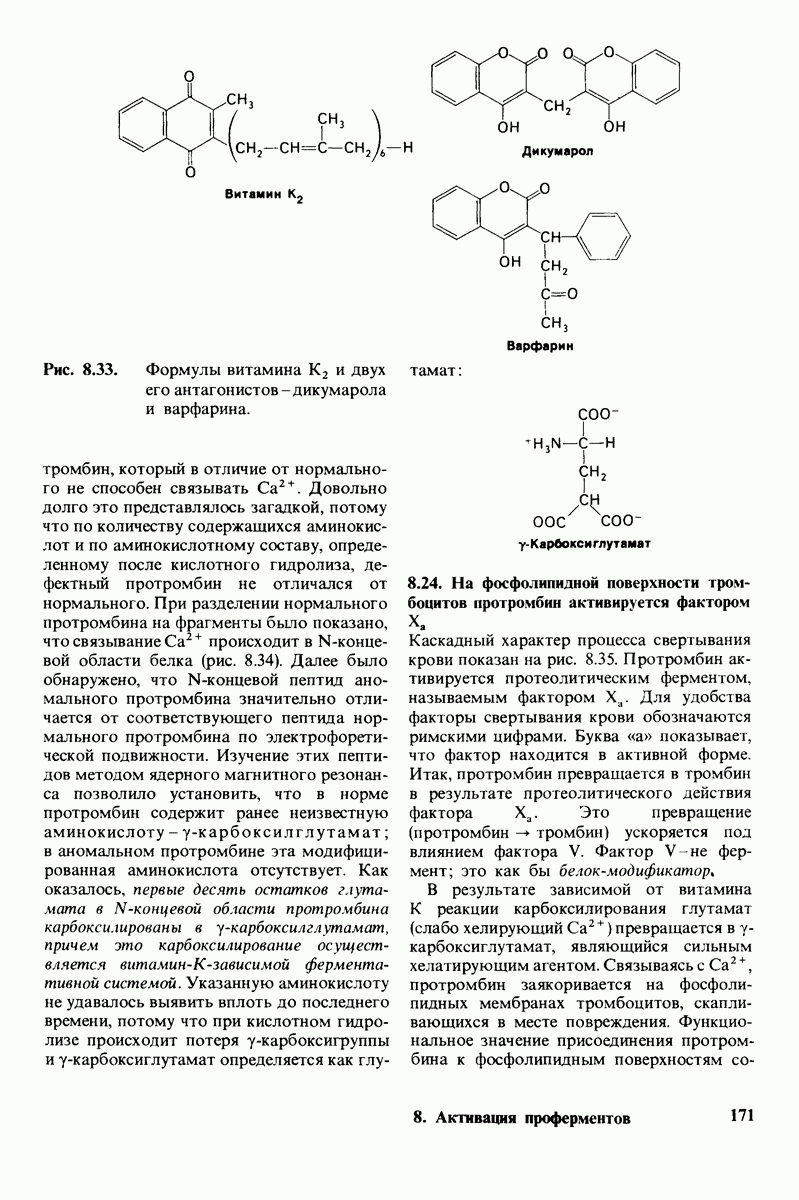
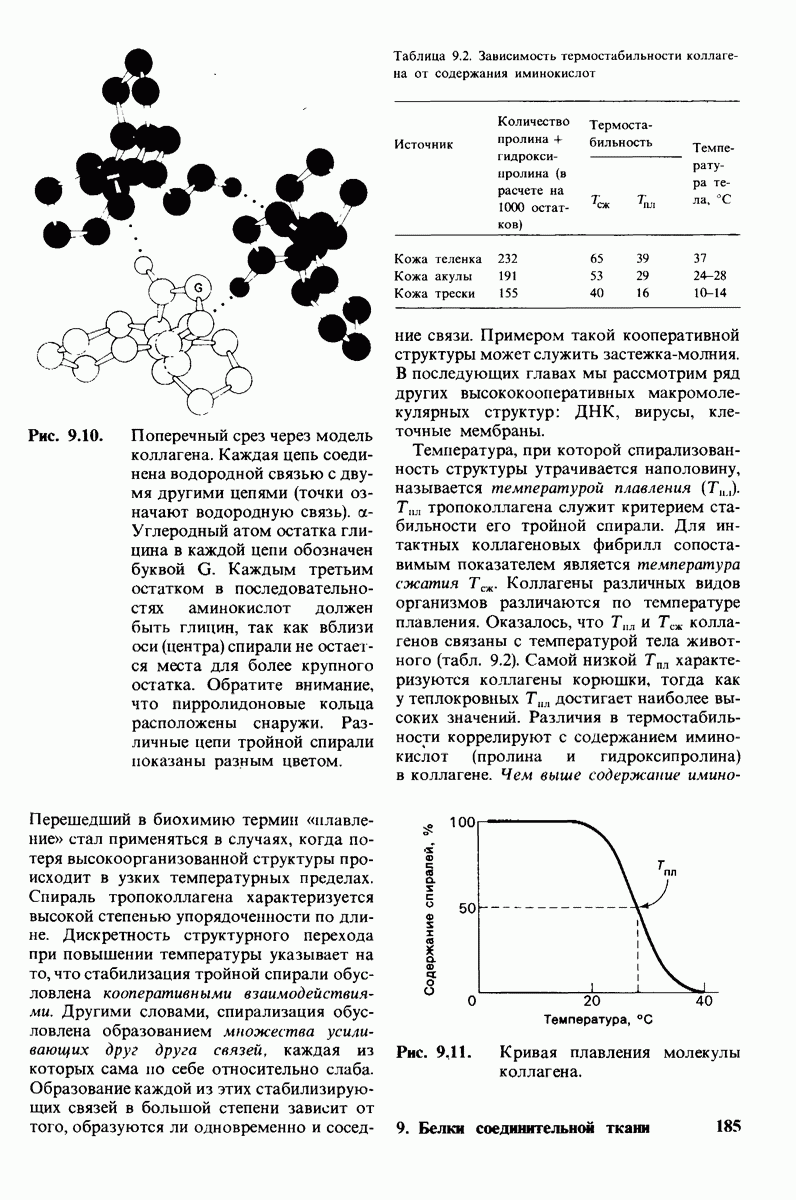
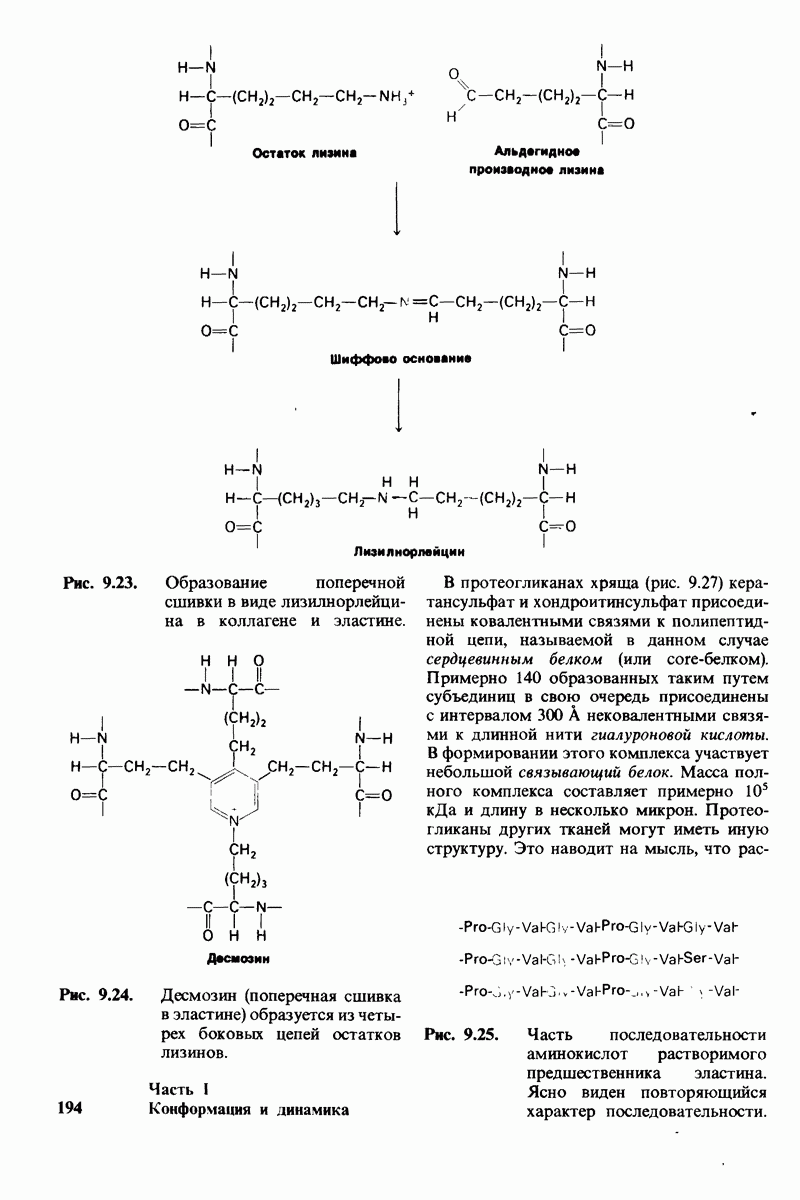
Рис. 4.20. Способ связи БФГ с дезоксигемоглобином человека. Бисфосфоглицерат взаимодействует с тремя положительно заряженными группами в каждой  -цепи. [Arnone A., Nature, 237, 148 (1972).]
-цепи. [Arnone A., Nature, 237, 148 (1972).]
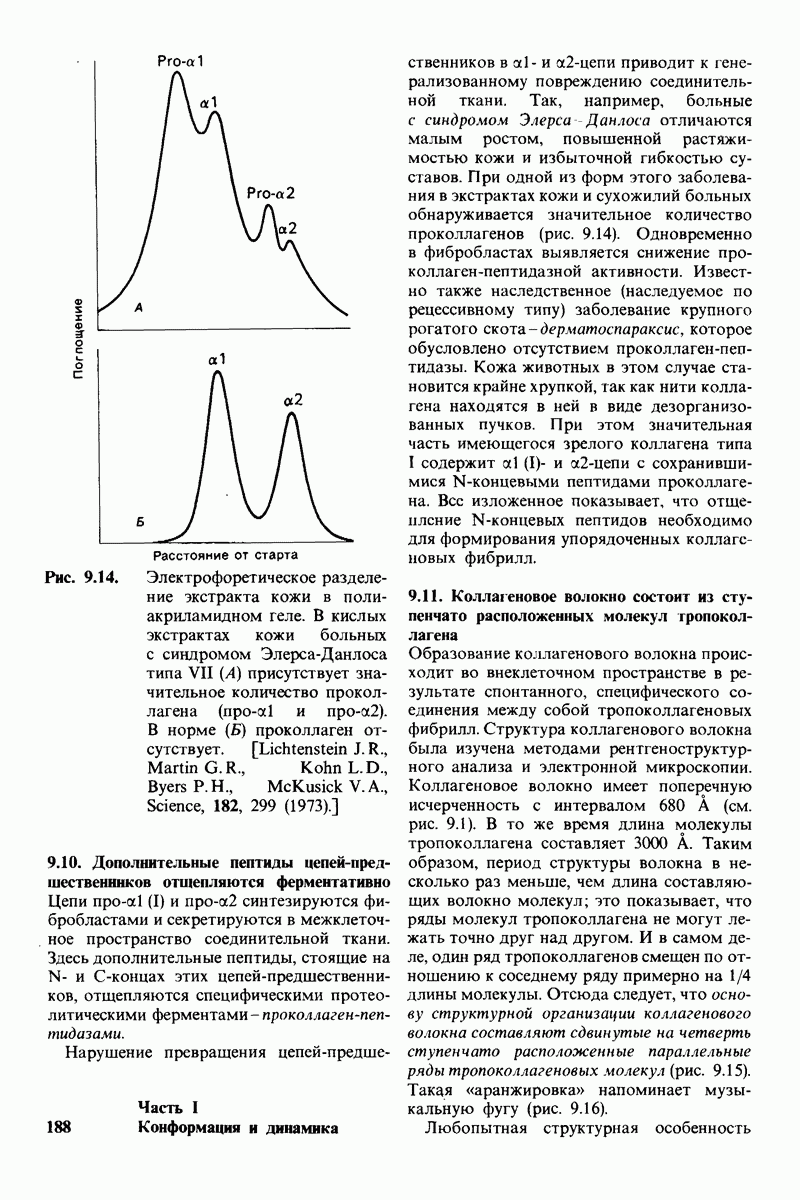
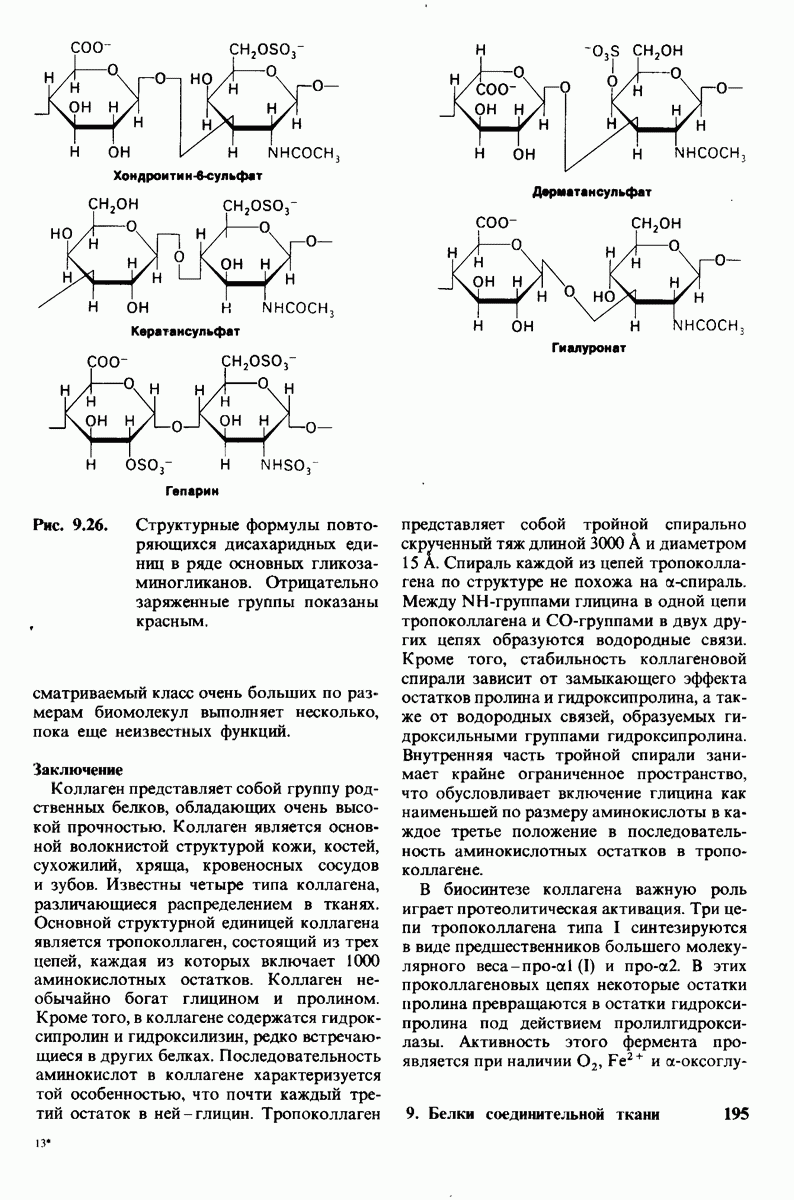
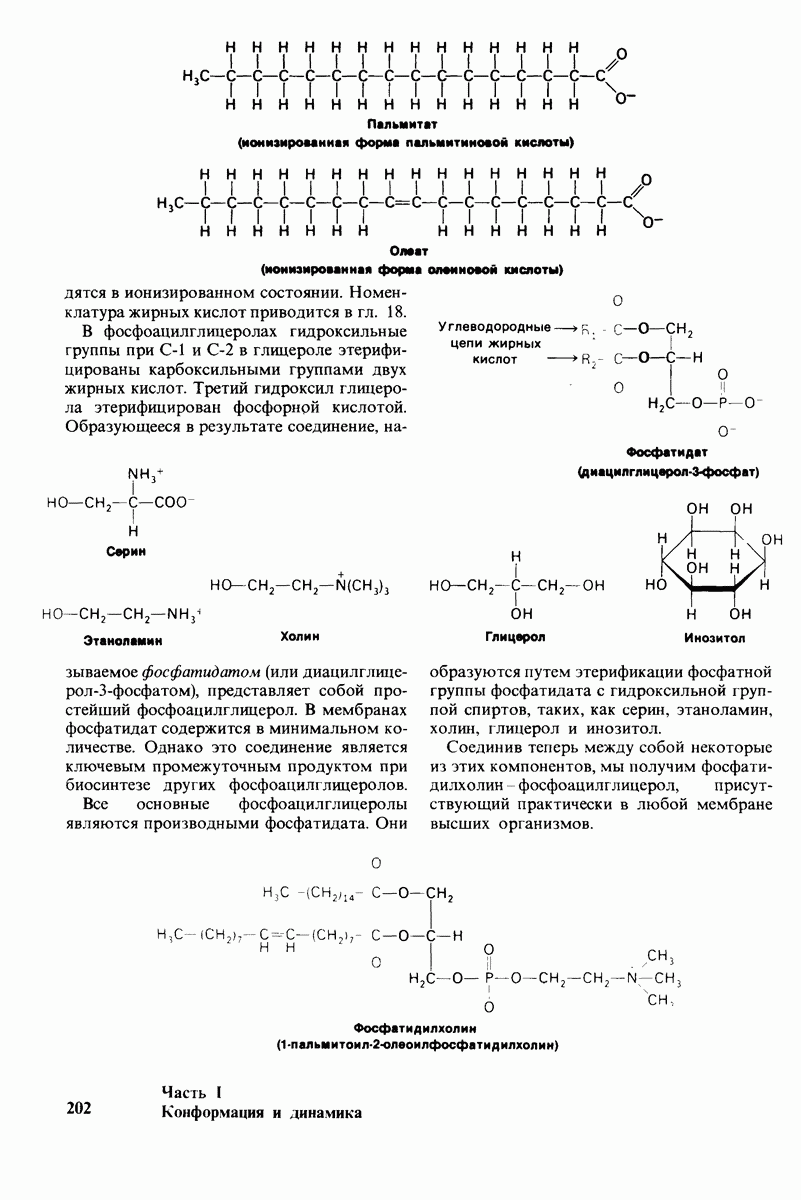
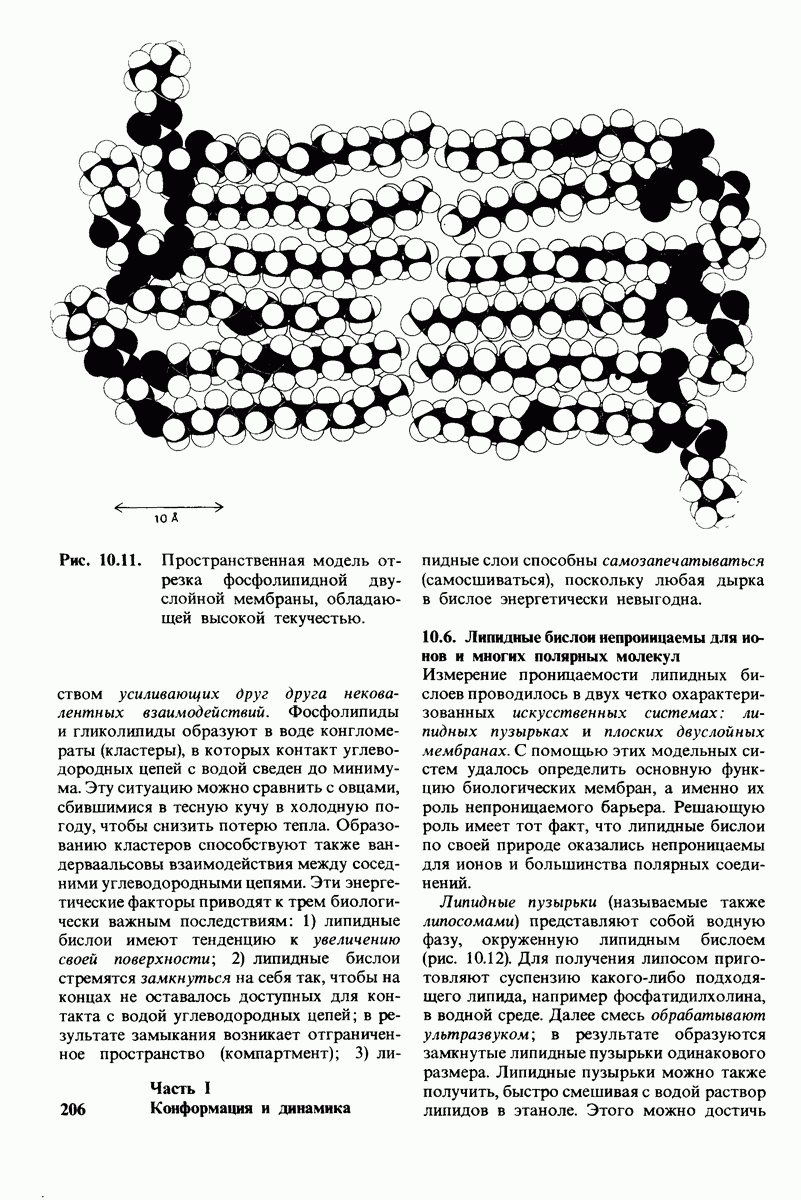
постоянство рН в тканях с активным метаболизмом.
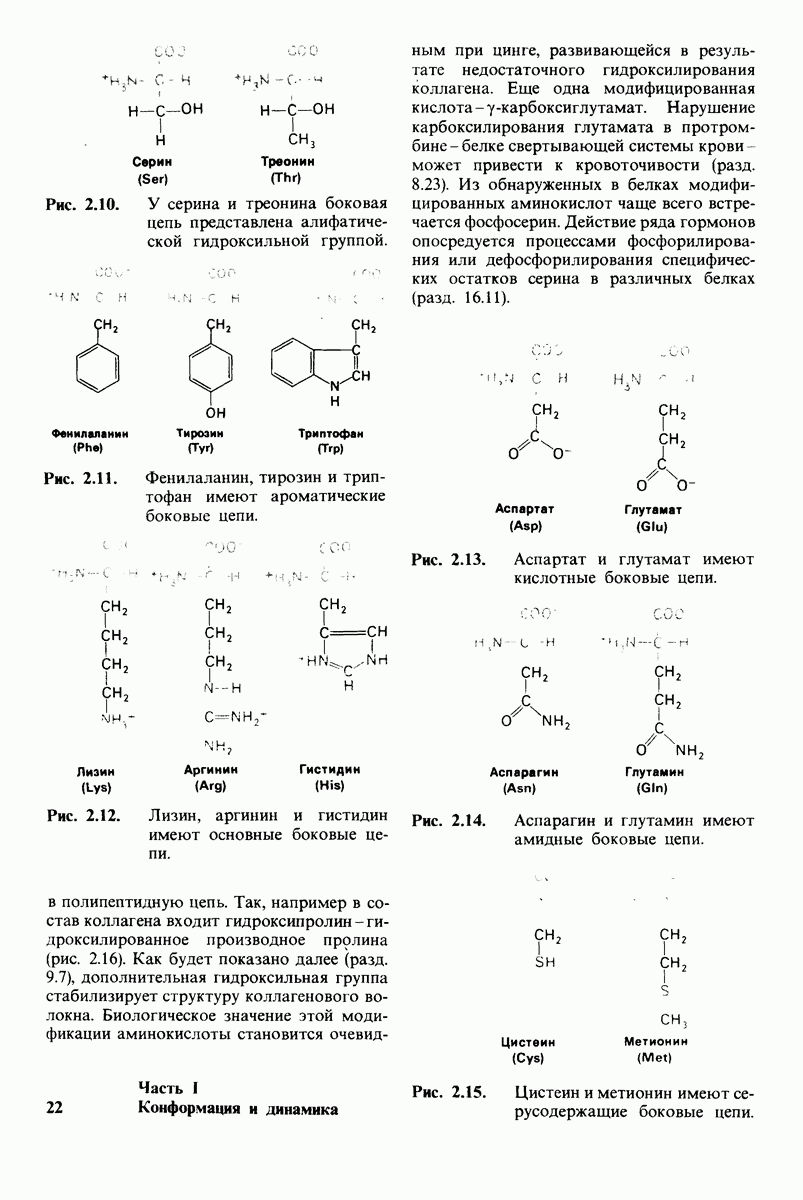
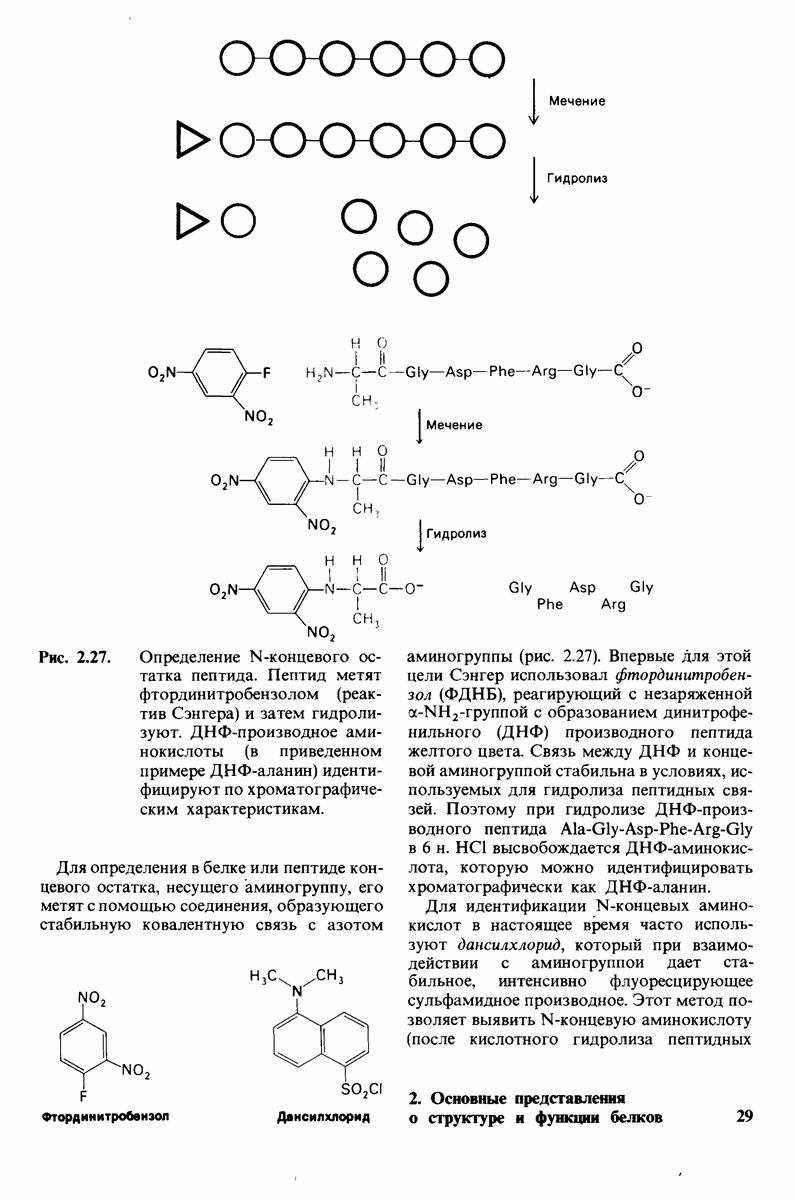
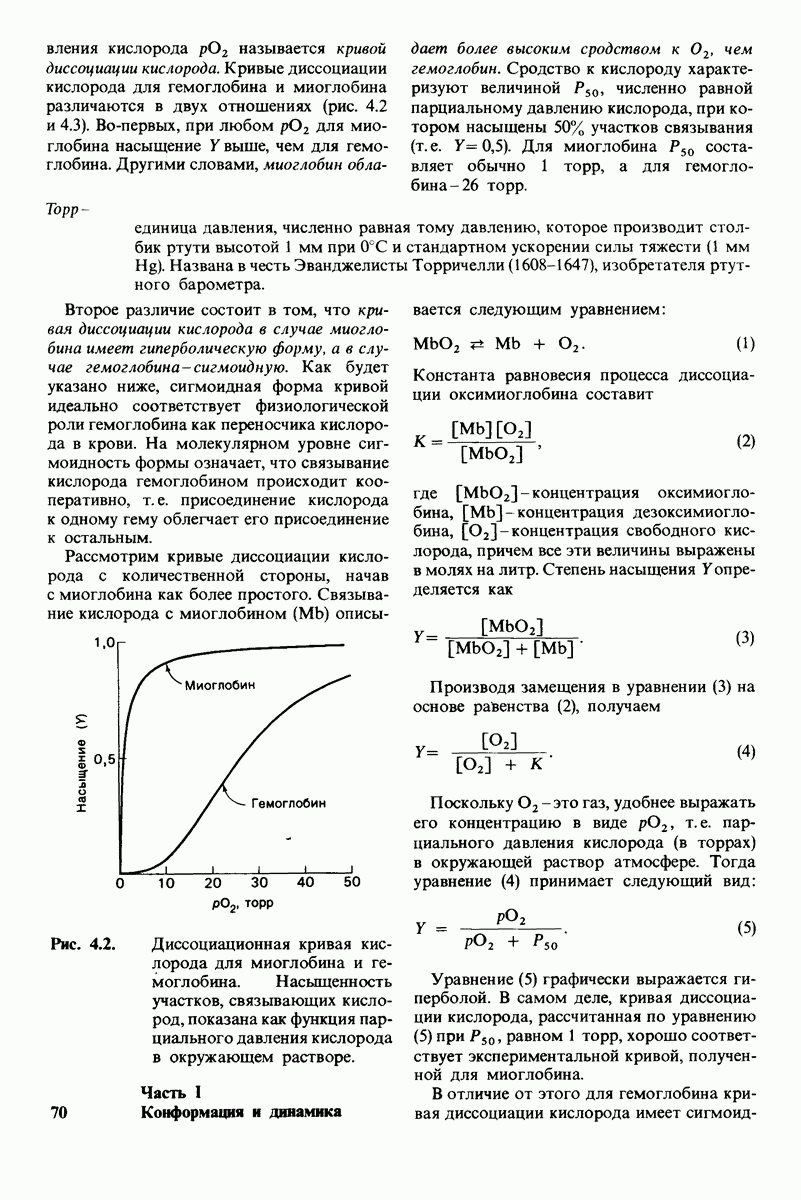
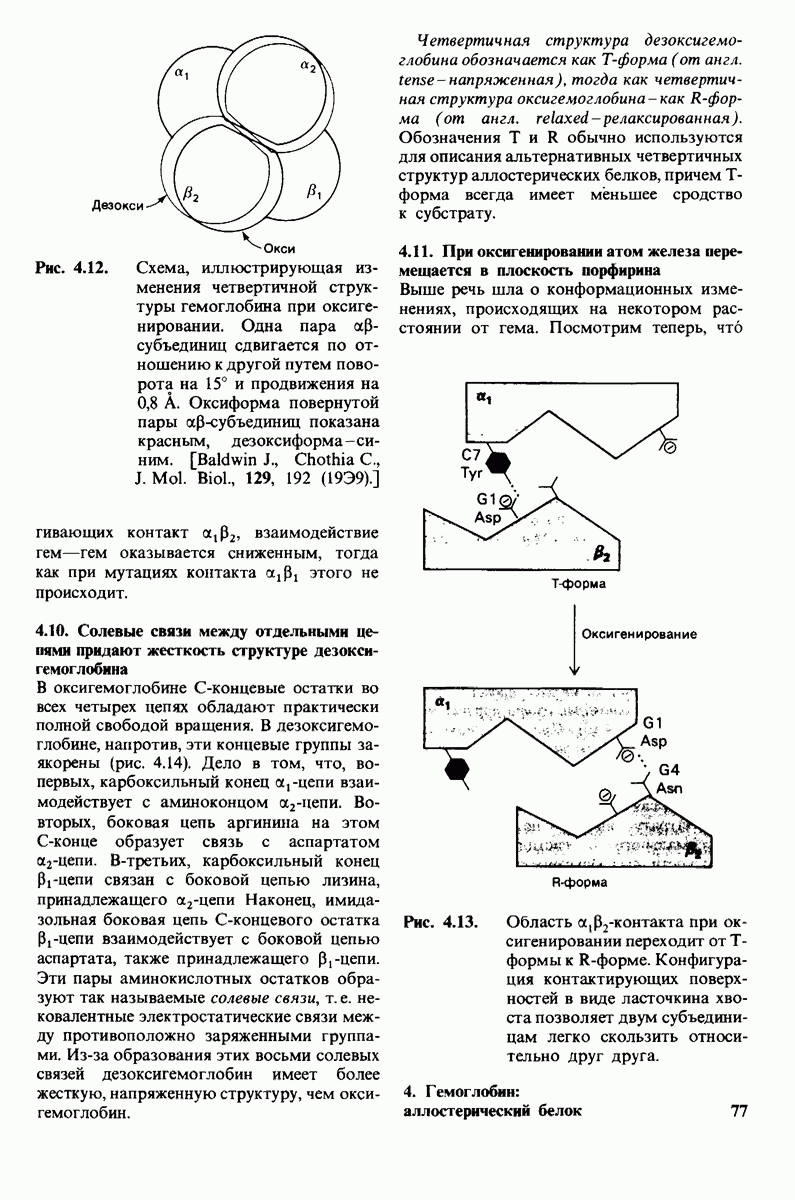
У каких конкретных групп повышается значение рХ? Рассмотрим сначала возможные участки связывания Н+ в гемоглобине (рис. 4.21 и табл. 4.1) и затем сузим круг выбора. Значения рК карбоксильных групп боковой цепи глутамата и аспартата составляют обычно ~ 4. Маловероятно, что их рХ возрастут до 7-8 - условие, необходимое для участия в эффекте Бора. Подобным же образом маловероятно, что значения рХ боковых цепей тирозина, лизина и аргинина могут быть сдвинуты в достаточной мере, поскольку обычно их рХ превышает 10. Отсюда следует, что эффект Бора может быть обеспечен участием гистидина, цистеина и концевой аминогруппы, так как нормальные значения рК этих групп лежат в области 7.
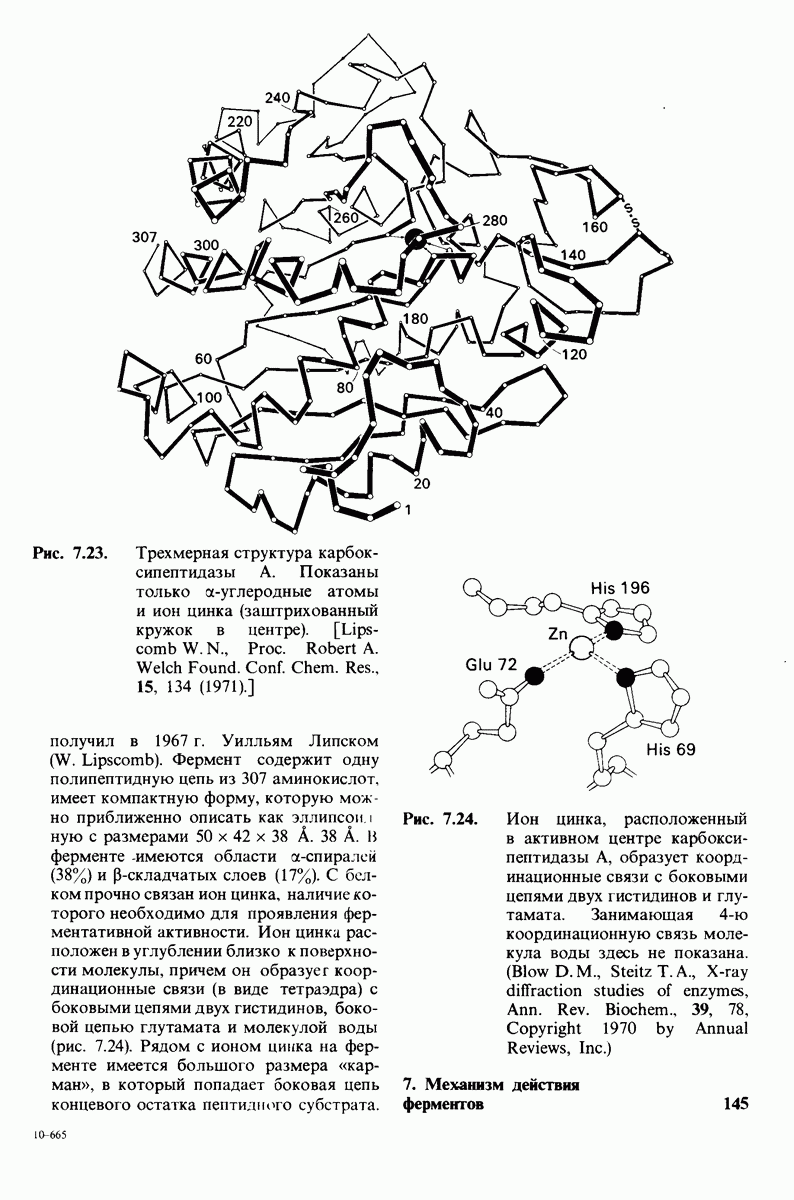
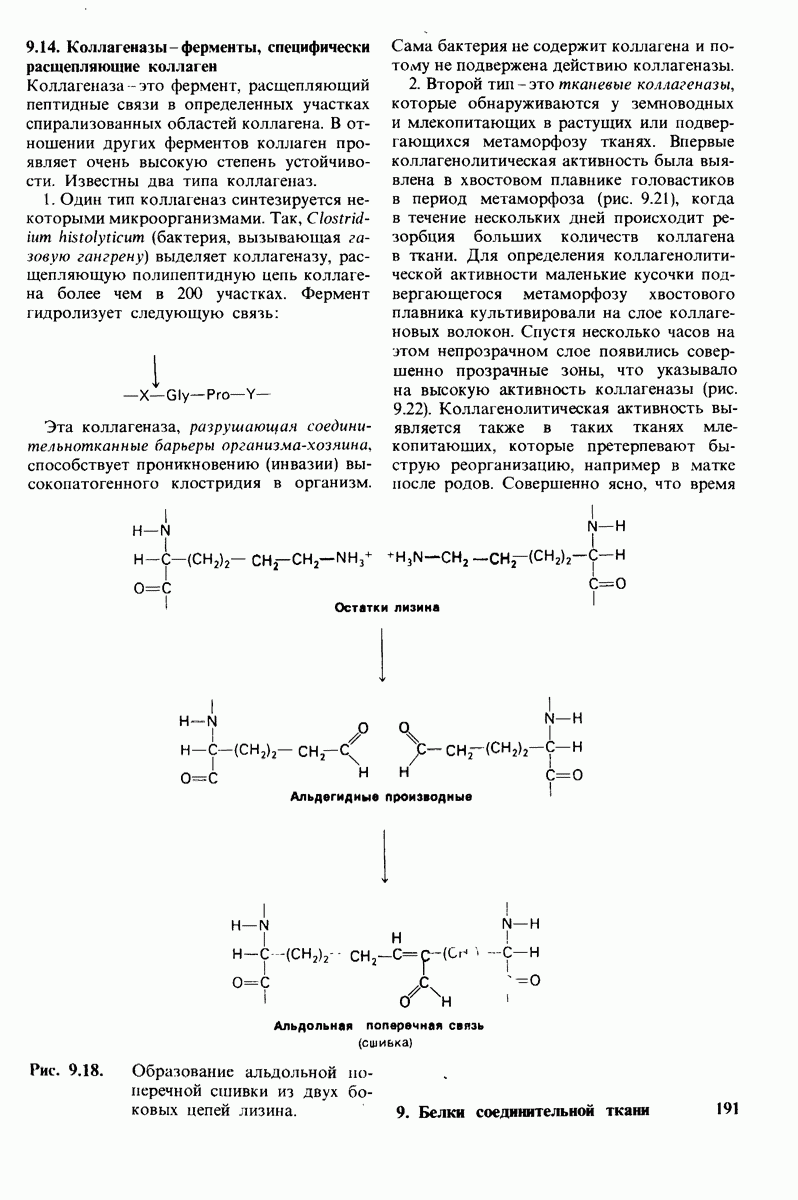
Путем сопоставления данных химического и рентгеноструктурного анализа удалось идентифицировать специфические группы, ответственные за эффект Бора. Рентгено-структурный анализ указывал на вовлечение в эффект Бора остатков гистидина, расположенных на С-конце каждой из  -цепей гемоглобина (гистидин-146Р). Справедливость этого предположения была проверена следующим путем. Был получен гемоглобин,
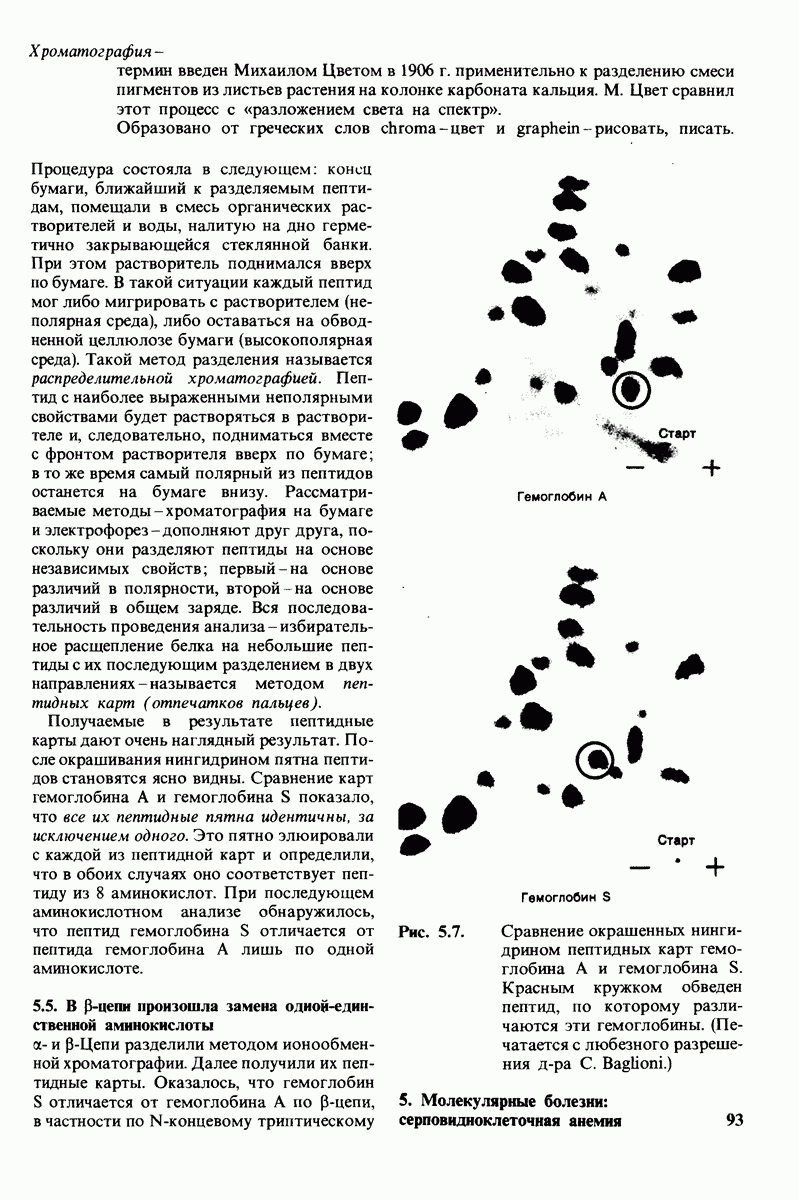
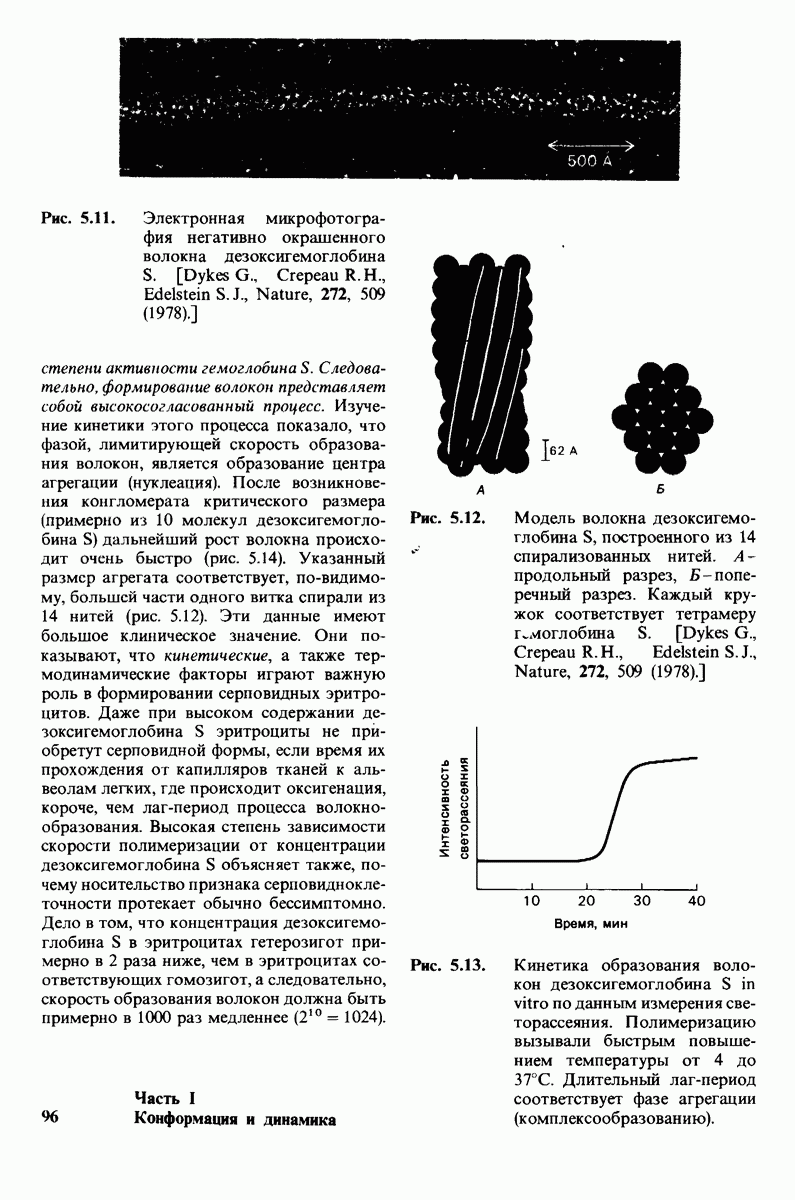
-цепей гемоглобина (гистидин-146Р). Справедливость этого предположения была проверена следующим путем. Был получен гемоглобин,  -цепи которого не содержали гистидина-146. Этого удалось достичь путем использования карбоксипептидазы В-специфического протеолитического фермента, расщепляющего в некоторых полипептидных цепях пептидную связь, образованную концевой аминокислотой основного характера, расположенной на С-конце пептида, т. е. содержащей свободную карбоксильную группу. Такая модификация гемоглобина приводила к снижению эффекта Бора в два раза. Отсюда следует, что гистидин-146 обеих $-цепей вносит, по-видимому, основной вклад в эффект Бора.
-цепи которого не содержали гистидина-146. Этого удалось достичь путем использования карбоксипептидазы В-специфического протеолитического фермента, расщепляющего в некоторых полипептидных цепях пептидную связь, образованную концевой аминокислотой основного характера, расположенной на С-конце пептида, т. е. содержащей свободную карбоксильную группу. Такая модификация гемоглобина приводила к снижению эффекта Бора в два раза. Отсюда следует, что гистидин-146 обеих $-цепей вносит, по-видимому, основной вклад в эффект Бора.
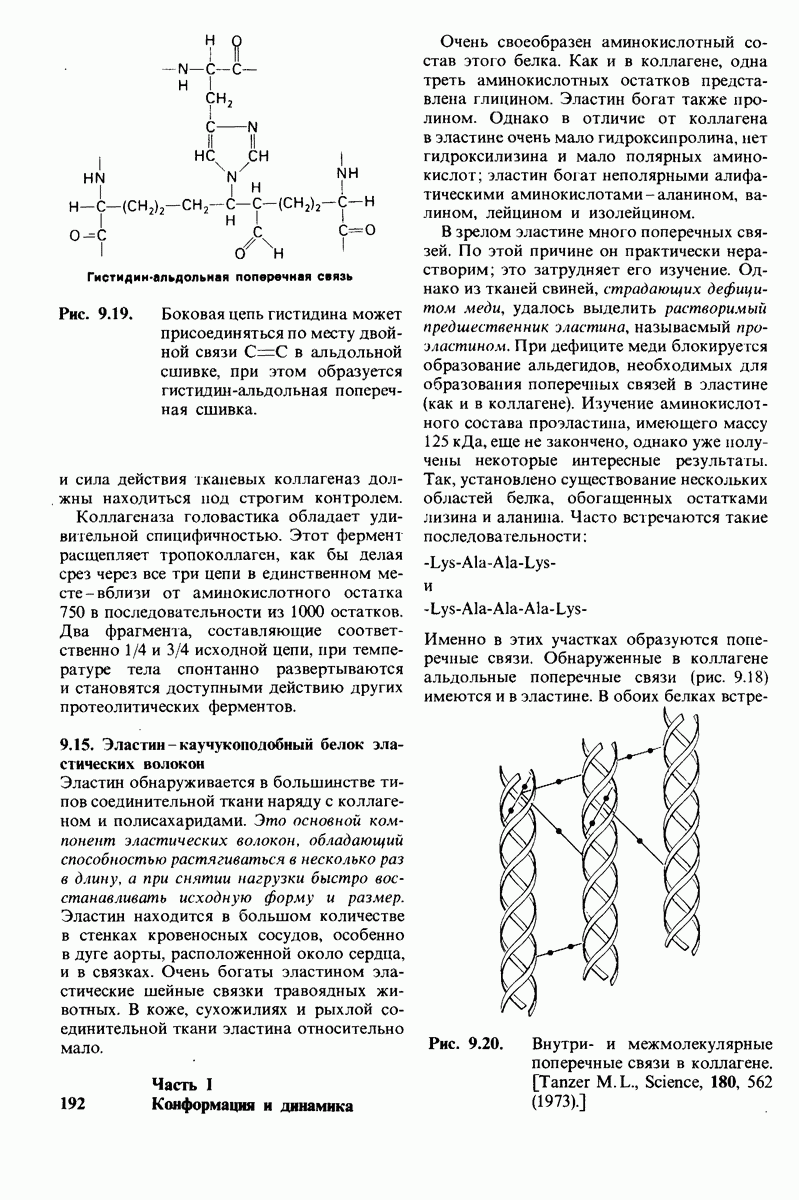
Роль концевых аминогрупп гемоглобина в эффекте Бора была убедительно доказана следующим образом. Концевые аминогруппы модифицировали обработкой
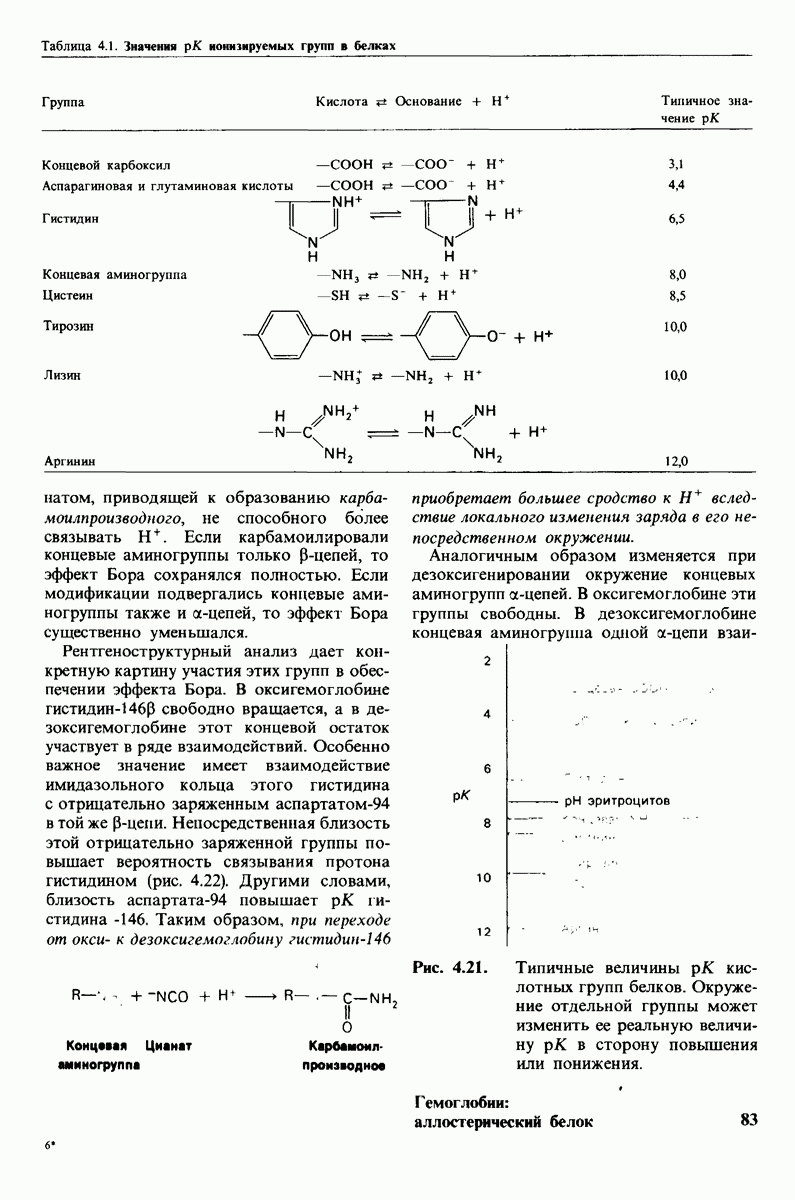
Таблица 4.1. (см. скан) Значения рК ионизируемых групп в белках
цианатом, приводящей к образованию карбамоилпроизводного, не способного более связывать  Если карбамоилировали концевые аминогруппы только
Если карбамоилировали концевые аминогруппы только  -цепей, то эффект Бора сохранялся полностью. Если модификации подвергались концевые аминогруппы также и
-цепей, то эффект Бора сохранялся полностью. Если модификации подвергались концевые аминогруппы также и  -цепей, то эффект Бора существенно уменьшался.
-цепей, то эффект Бора существенно уменьшался.
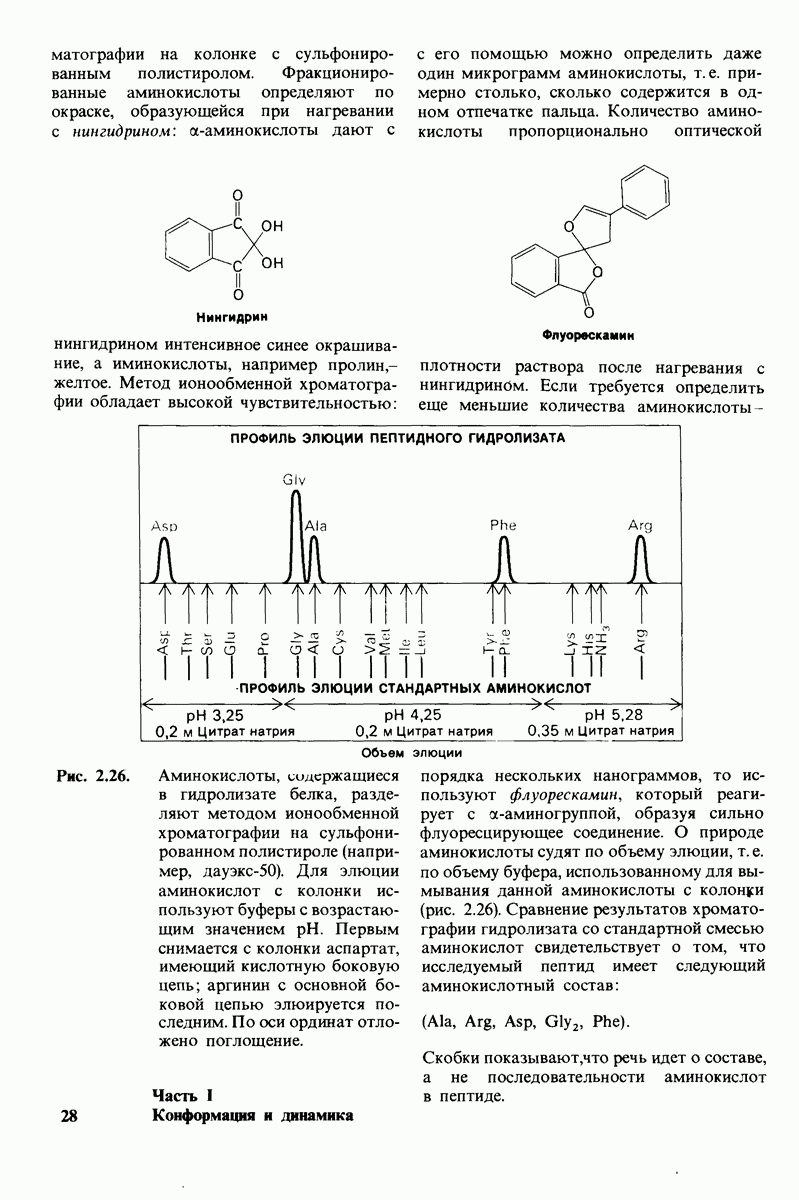
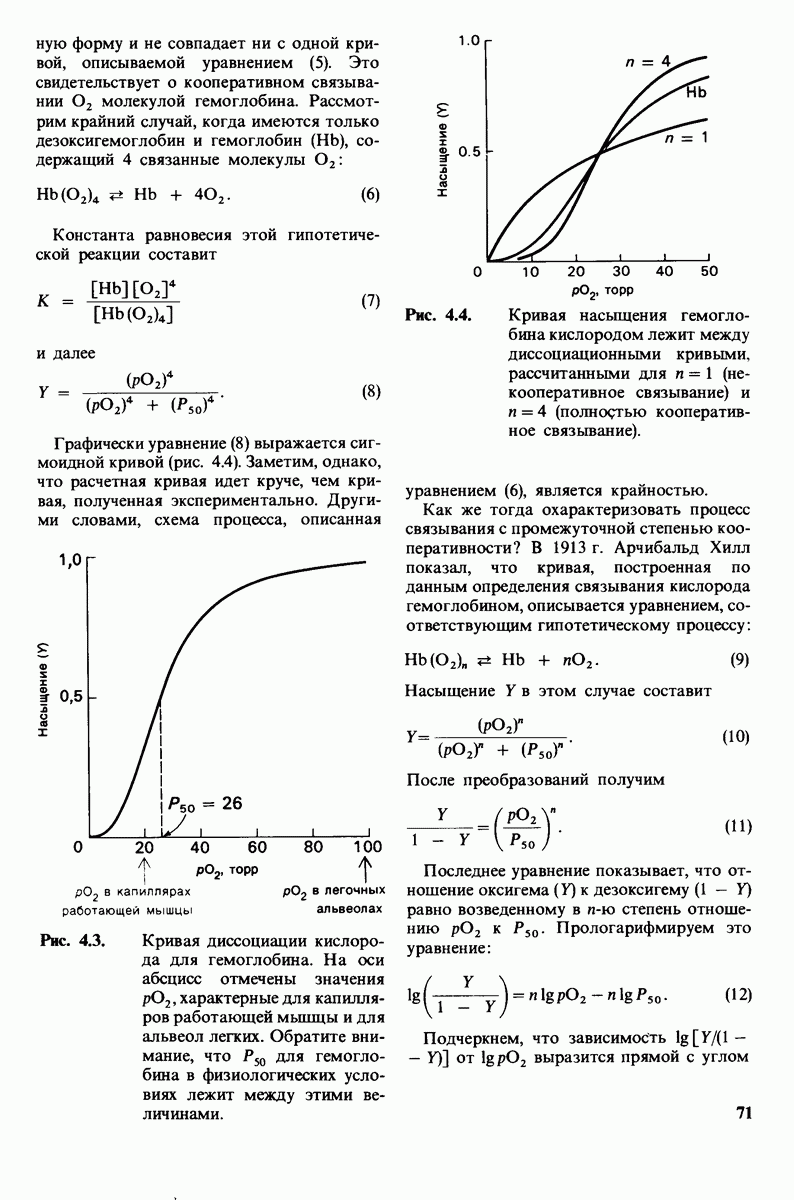
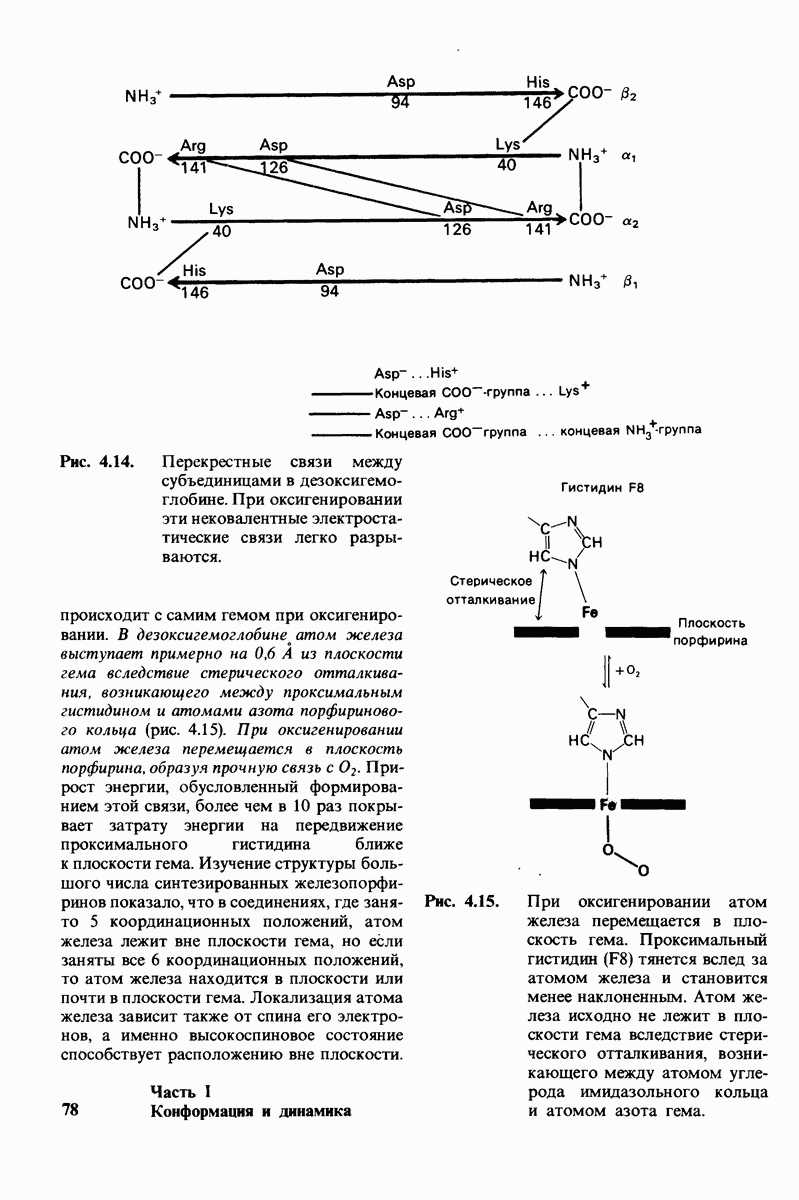
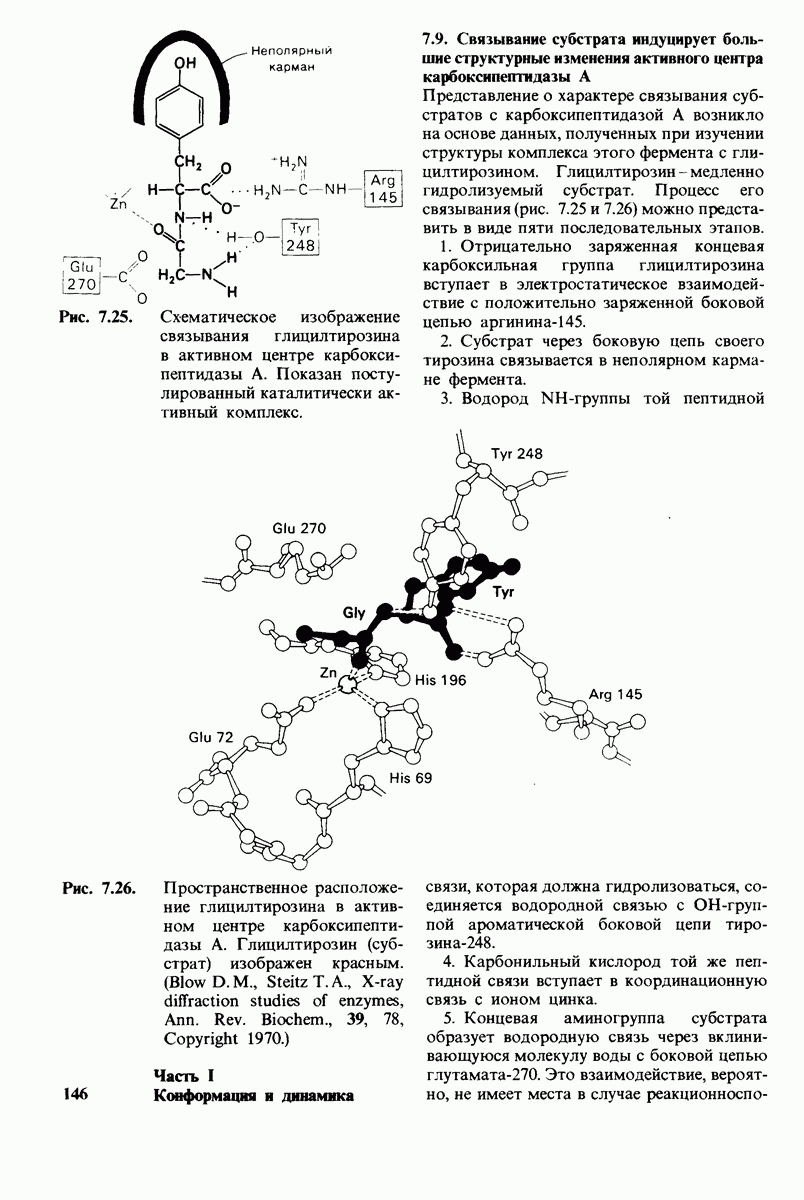
Рентгеноструктурный анализ дает конкретную картину участия этих групп в обеспечении эффекта Бора. В оксигемоглобине  свободно вращается, а в дезоксигемоглобине этот концевой остаток участвует в ряде взаимодействий. Особенно важное значение имеет взаимодействие имидазольного кольца этого гистидина с отрицательно заряженным аспартатом-94 в той же
свободно вращается, а в дезоксигемоглобине этот концевой остаток участвует в ряде взаимодействий. Особенно важное значение имеет взаимодействие имидазольного кольца этого гистидина с отрицательно заряженным аспартатом-94 в той же  -цепи. Непосредственная близость этой отрицательно заряженной группы повышает вероятность связывания протона гистидином (рис. 4.22). Другими словами, близость аспартата-94 повышает рК гистидина -146. Таким образом, при переходе от окси- к дезоксигемоглобину гистидин-146
-цепи. Непосредственная близость этой отрицательно заряженной группы повышает вероятность связывания протона гистидином (рис. 4.22). Другими словами, близость аспартата-94 повышает рК гистидина -146. Таким образом, при переходе от окси- к дезоксигемоглобину гистидин-146

приобретает большее сродство к  вследствие локального изменения заряда в его непосредственном окружении.
вследствие локального изменения заряда в его непосредственном окружении.
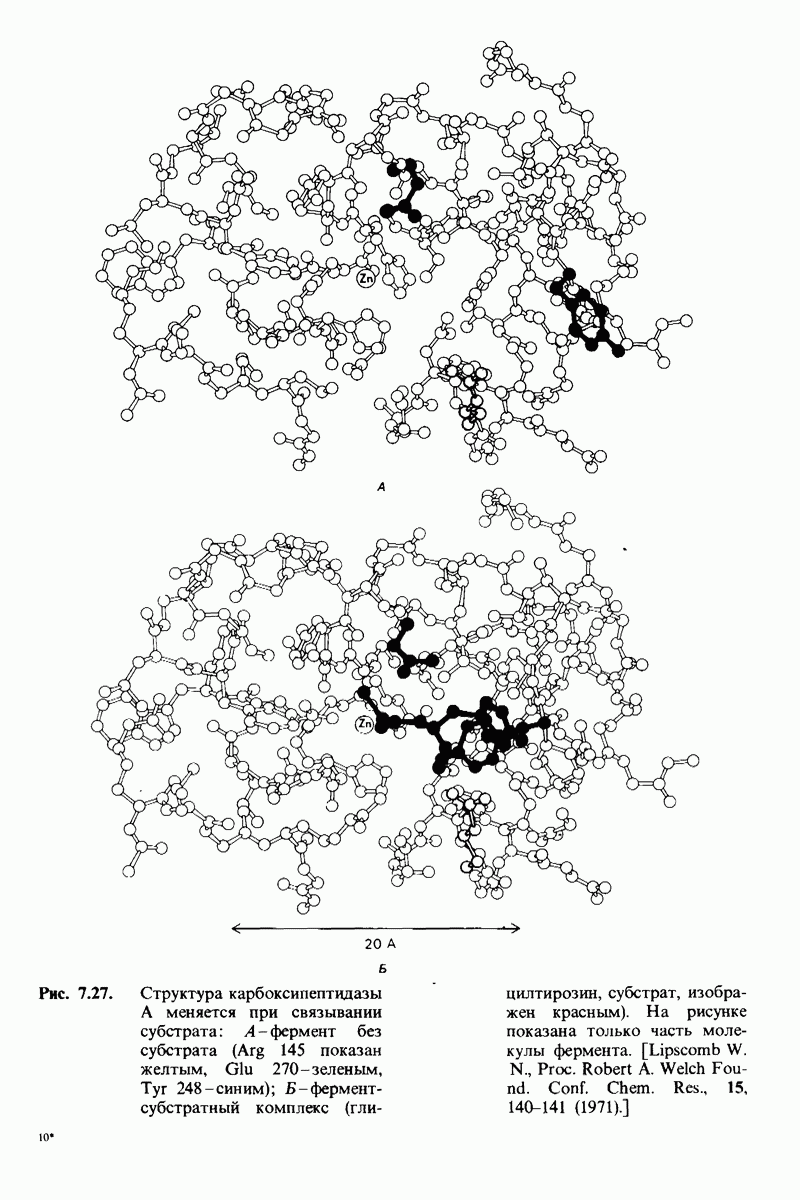
Аналогичным образом изменяется при дезоксигенировании окружение концевых аминогрупп  -цепей. В оксигемоглобине эти группы свободны. В дезоксигемоглобине концевая аминогруппа одной
-цепей. В оксигемоглобине эти группы свободны. В дезоксигемоглобине концевая аминогруппа одной  -цепи
-цепи

Рис. 4.21. Типичные величины рК кислотных групп белков. Окружение отдельной группы может изменить ее реальную величину рК в сторону повышения или понижения.

Рис. 4.22. Аспартат-94 повышает рК гистидина-146 в дезоксигемоглобине, но не в оксигемоглобине. Близость отрицательного заряда на аспартате-94 способствует протонированию гистидина-146 в дезоксигемоглобине.
взаимодействует с концевой карбоксильной группой другой  -цепи. Близость отрицательно заряженного карбоксилированного остатка в дезоксигемоглобине повышает сродство этой концевой аминогруппы к
-цепи. Близость отрицательно заряженного карбоксилированного остатка в дезоксигемоглобине повышает сродство этой концевой аминогруппы к  Данные рентгеноструктурного анализа свидетельствуют о том, что в эффекте Бора участвует и третья группа, а именно гистидин-122 в
Данные рентгеноструктурного анализа свидетельствуют о том, что в эффекте Бора участвует и третья группа, а именно гистидин-122 в  -цепи.
-цепи.
Итак, окружение трех пар групп, связывающих протоны (концевая аминогруппа и два гистидина) в окси- и дезоксигемоглобине неодинаково. В дезоксигемоглобине это локальное окружение заряжено более отрицательно. В результате при высвобождении кислорода эти группы связывают 



















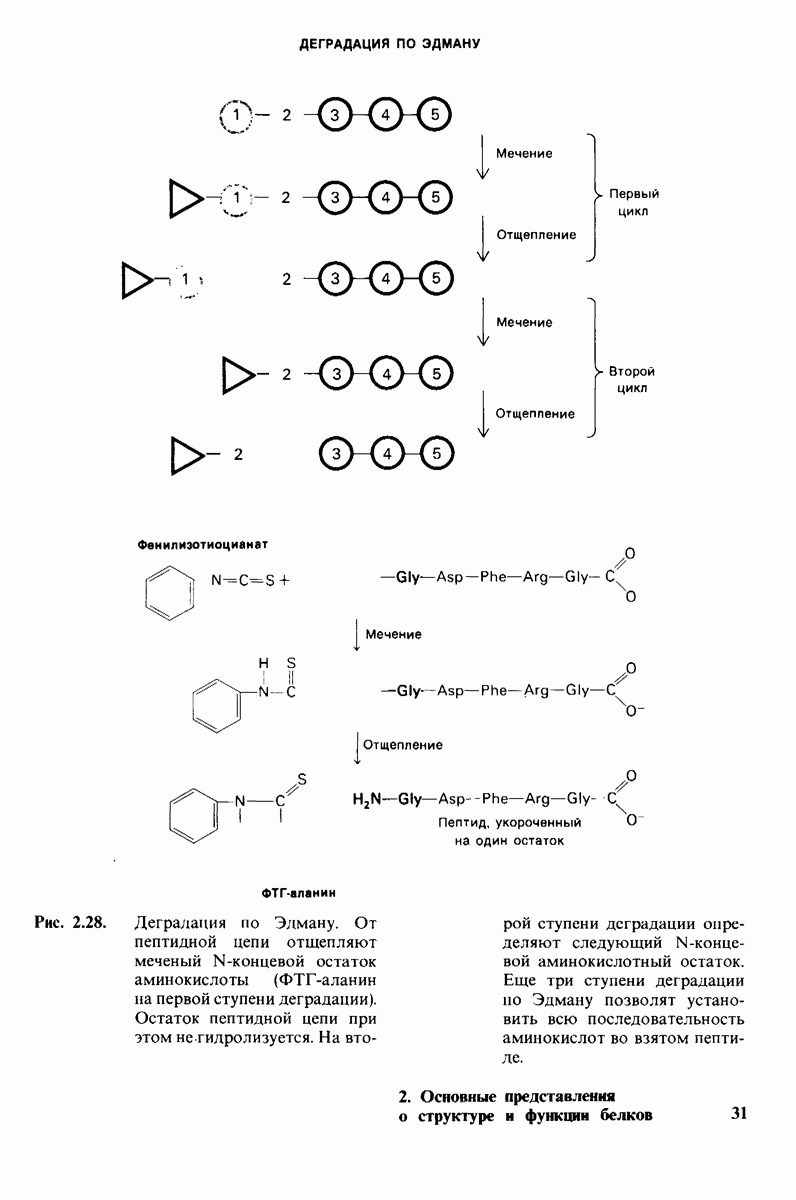

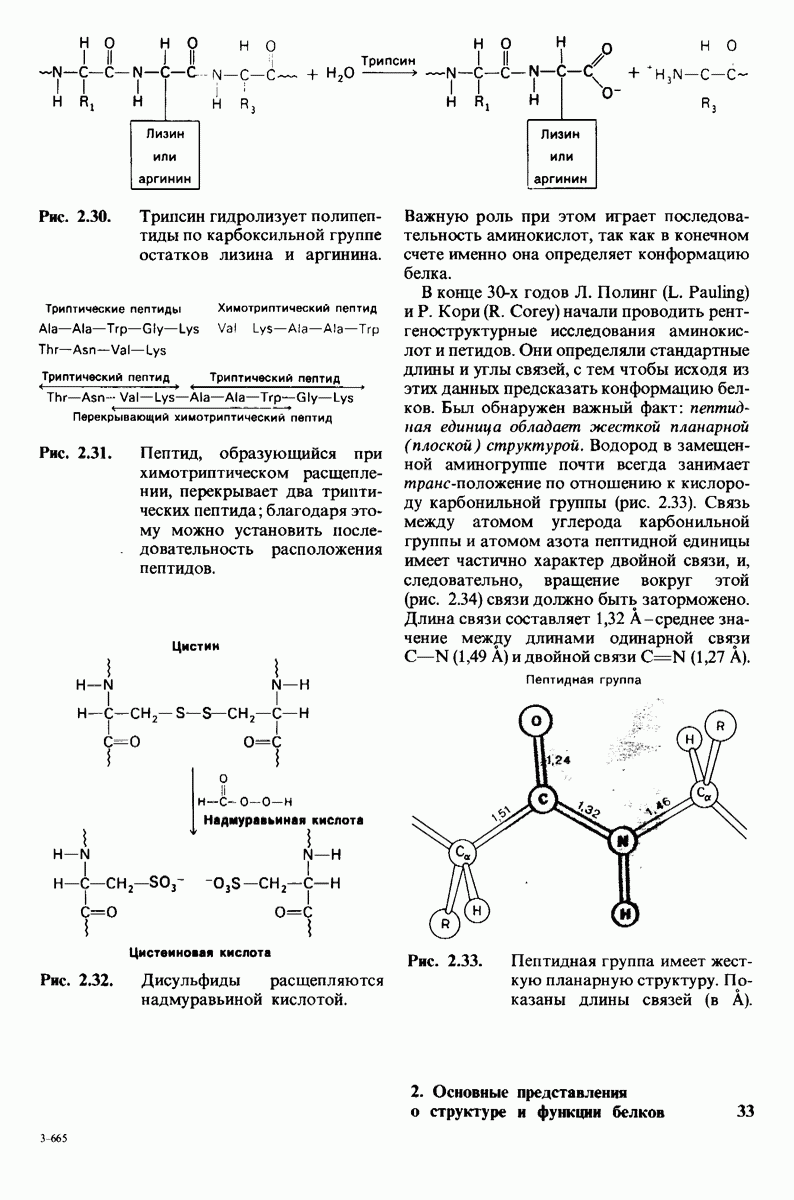
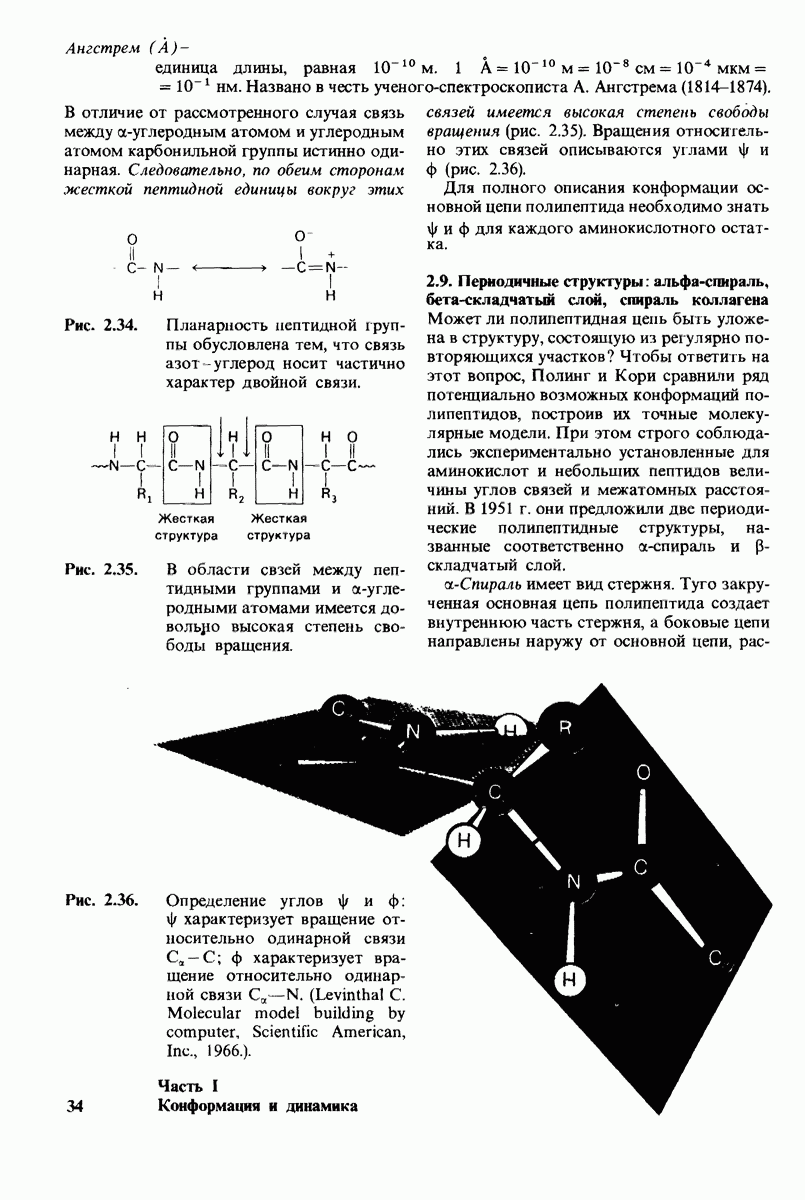
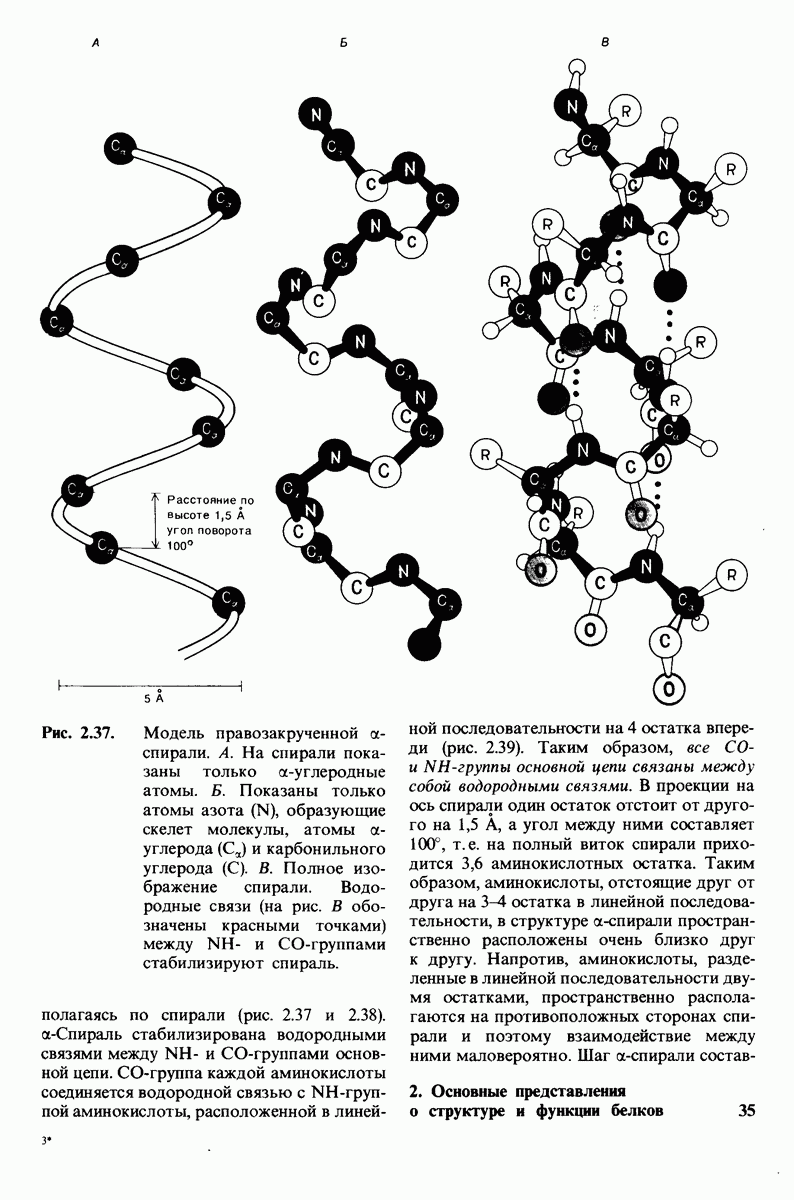


















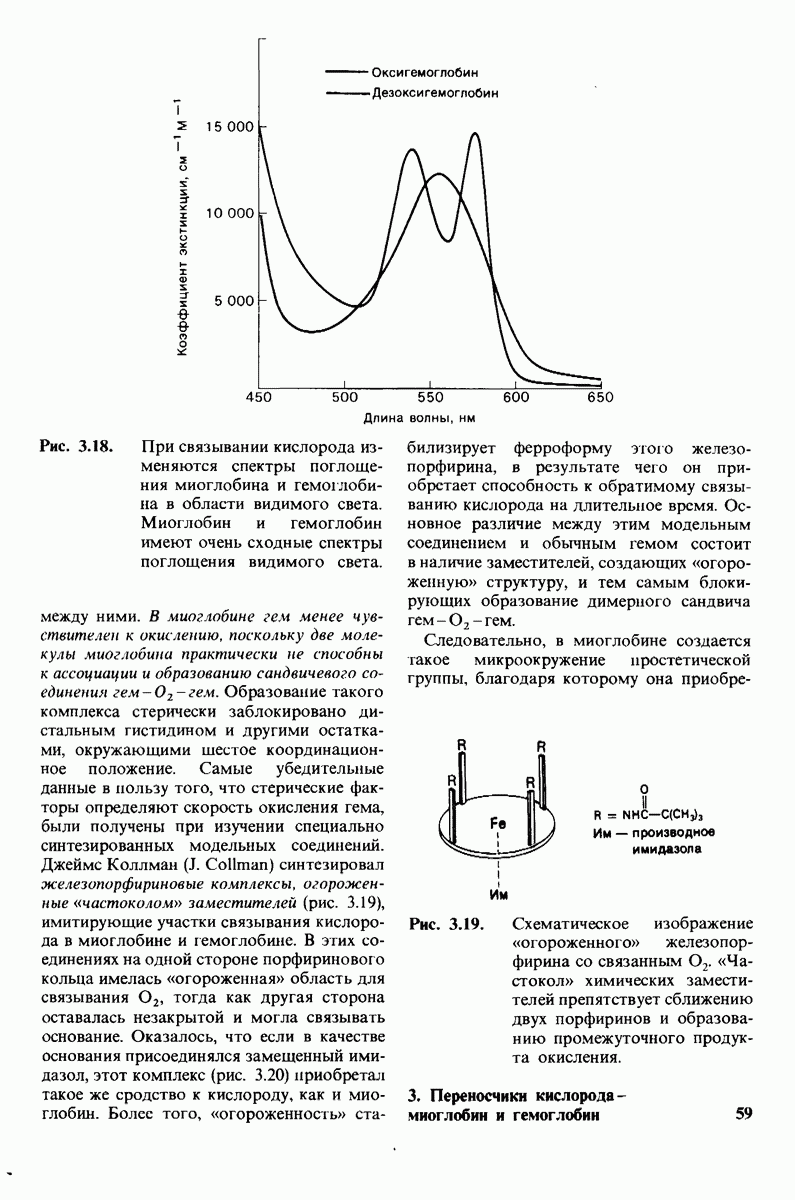







































































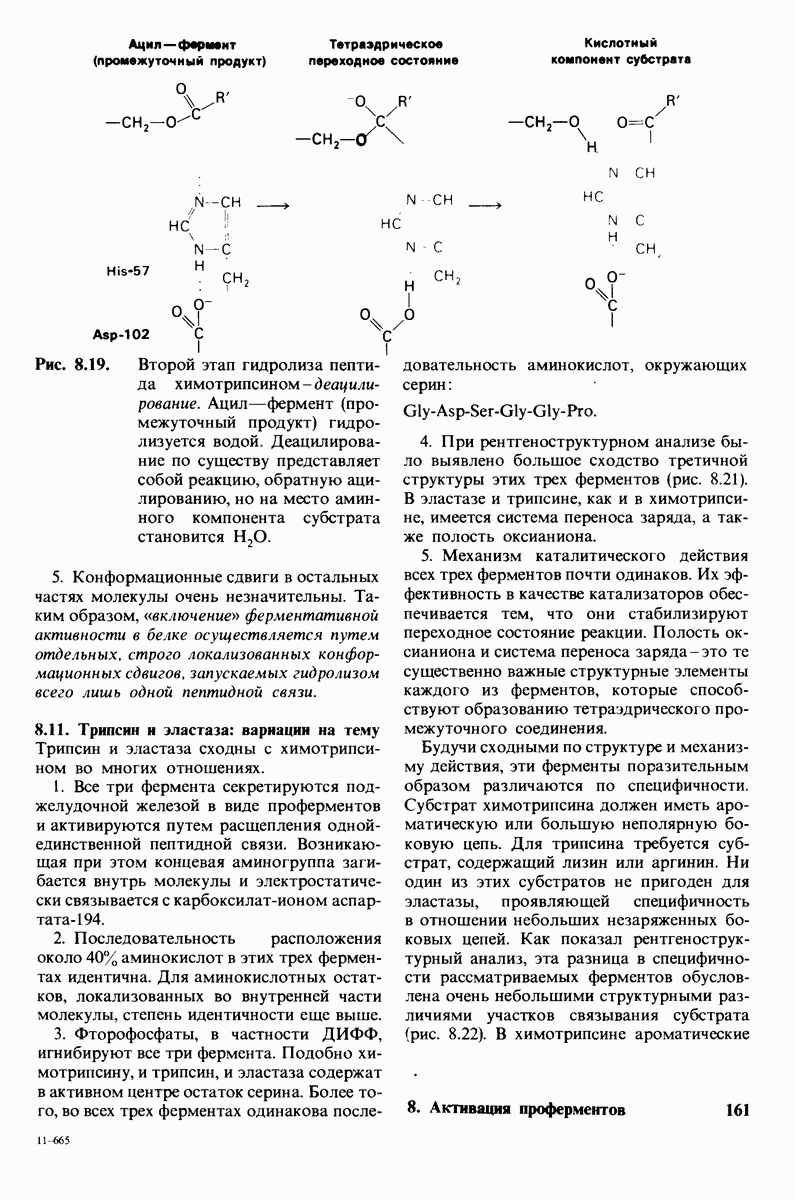
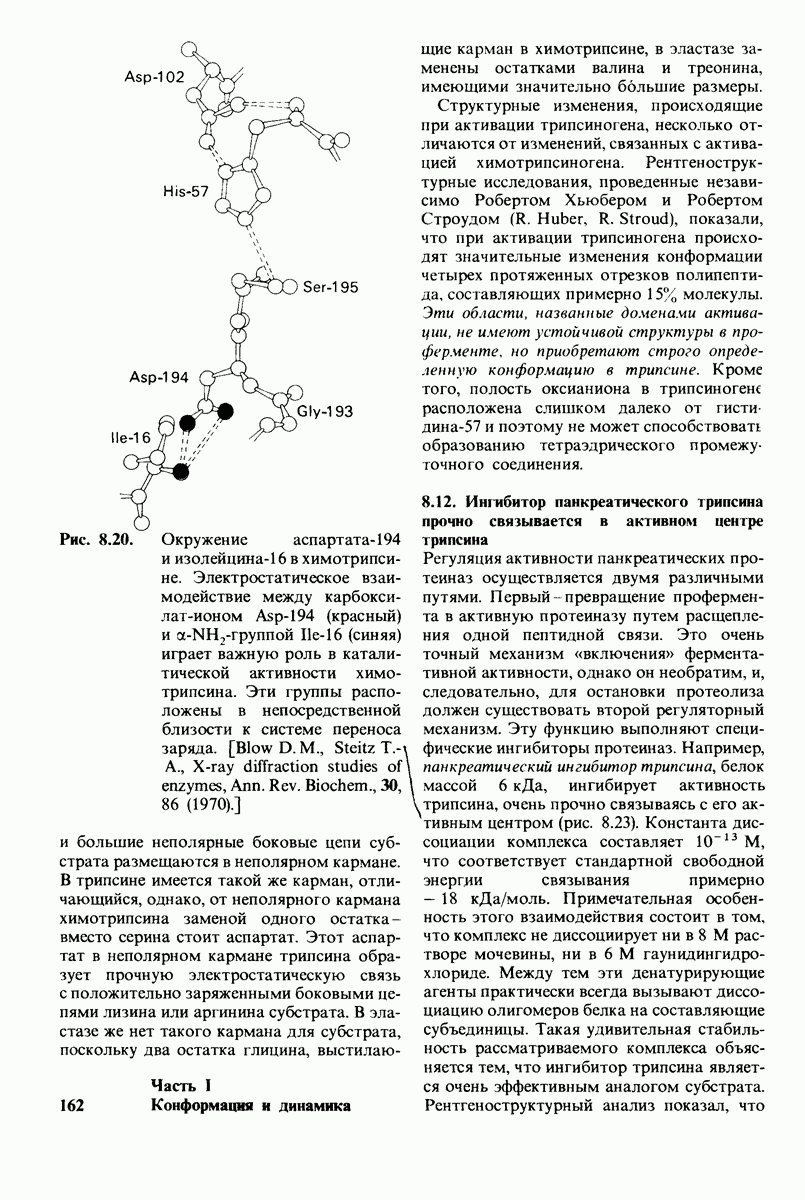
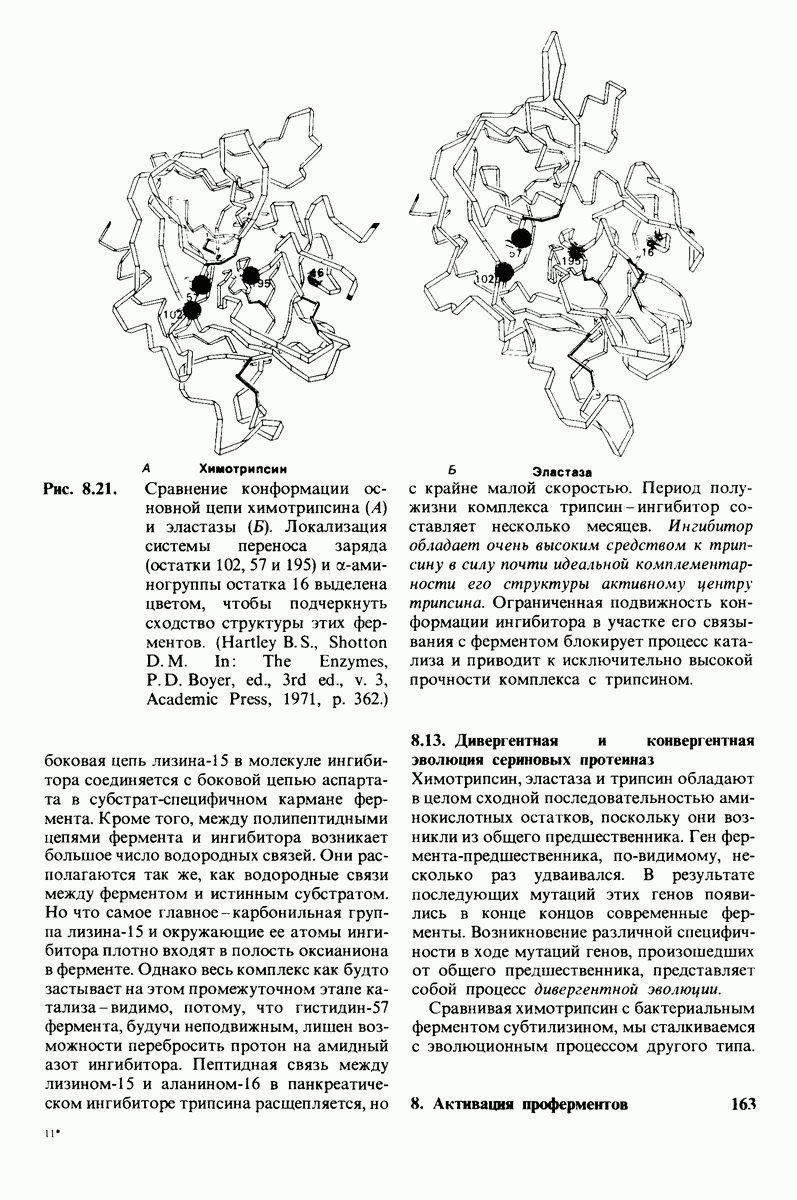
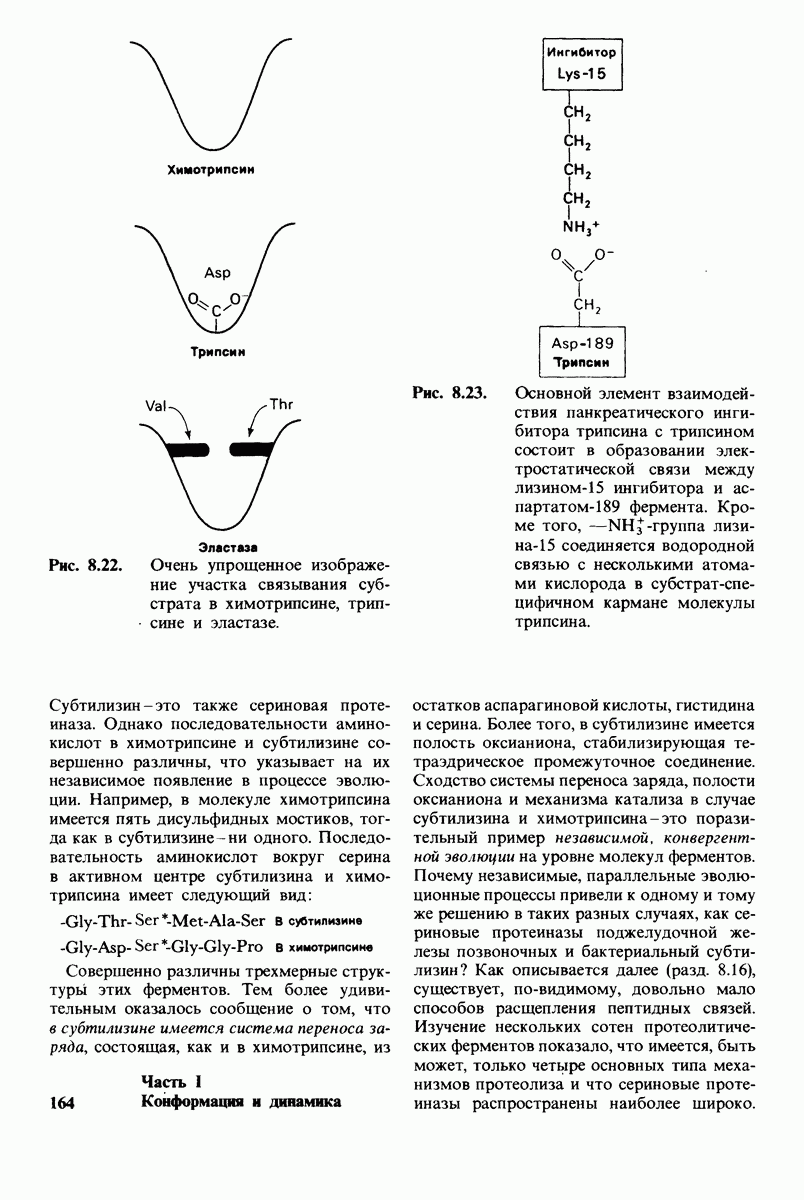

















































 В частности, при превращении из окси- в дезоксиформу должно увеличиваться значение
В частности, при превращении из окси- в дезоксиформу должно увеличиваться значение  отдельных групп, поскольку увеличение рК означает более сильное связывание
отдельных групп, поскольку увеличение рК означает более сильное связывание  Гемоглобин связывает около
Гемоглобин связывает около  в расчете на одну высвободившуюся молекулу
в расчете на одну высвободившуюся молекулу  Это присоединение
Это присоединение  позволяет поддерживать
позволяет поддерживать
