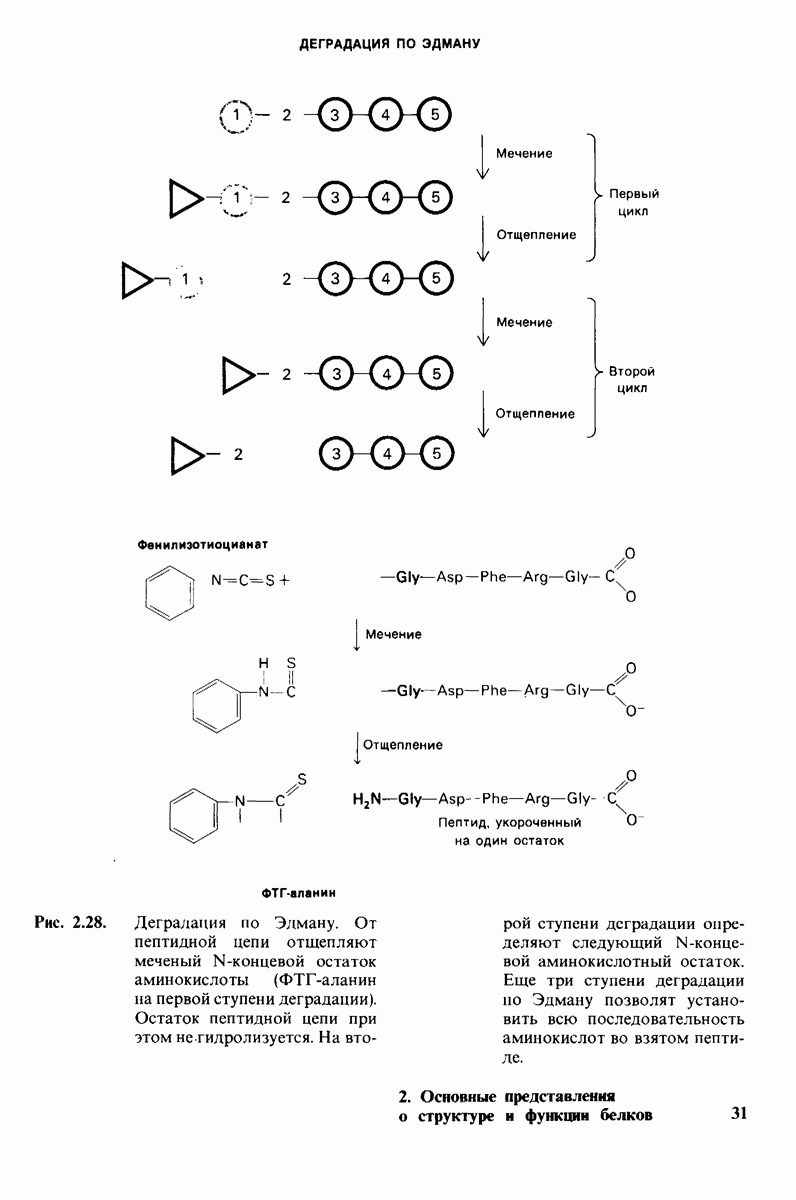
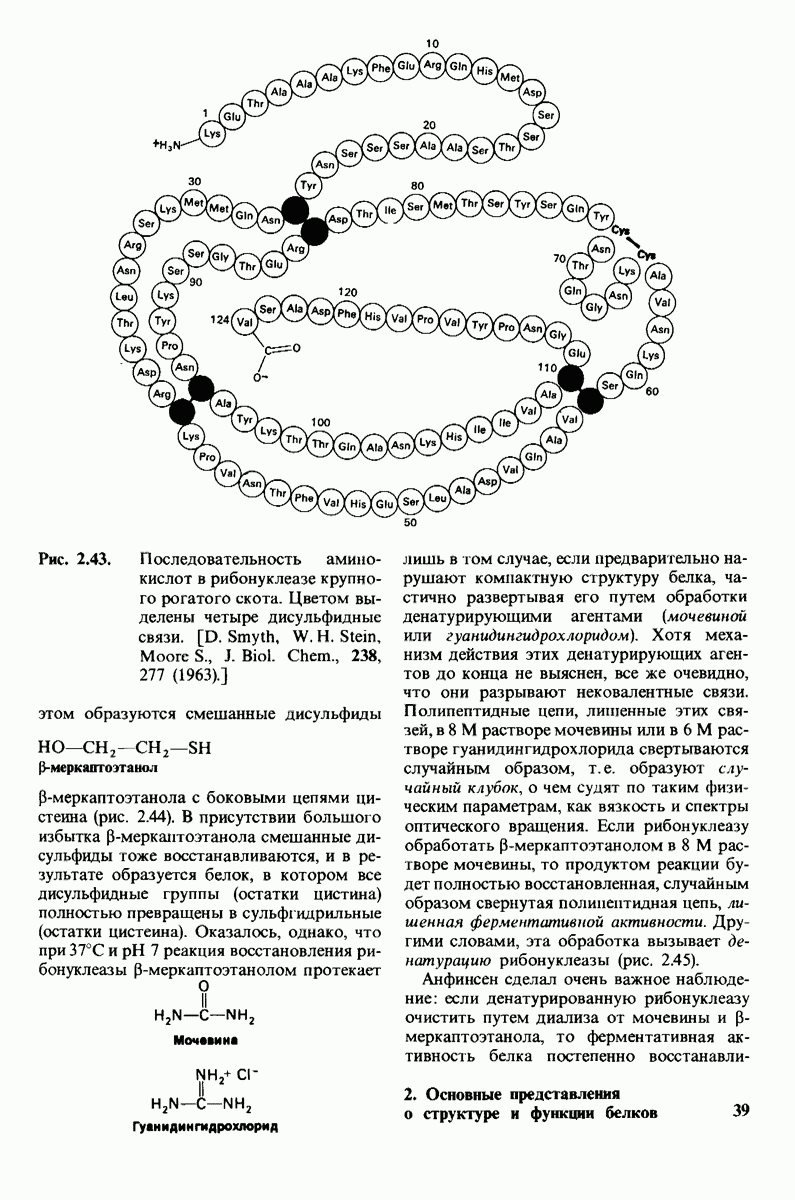
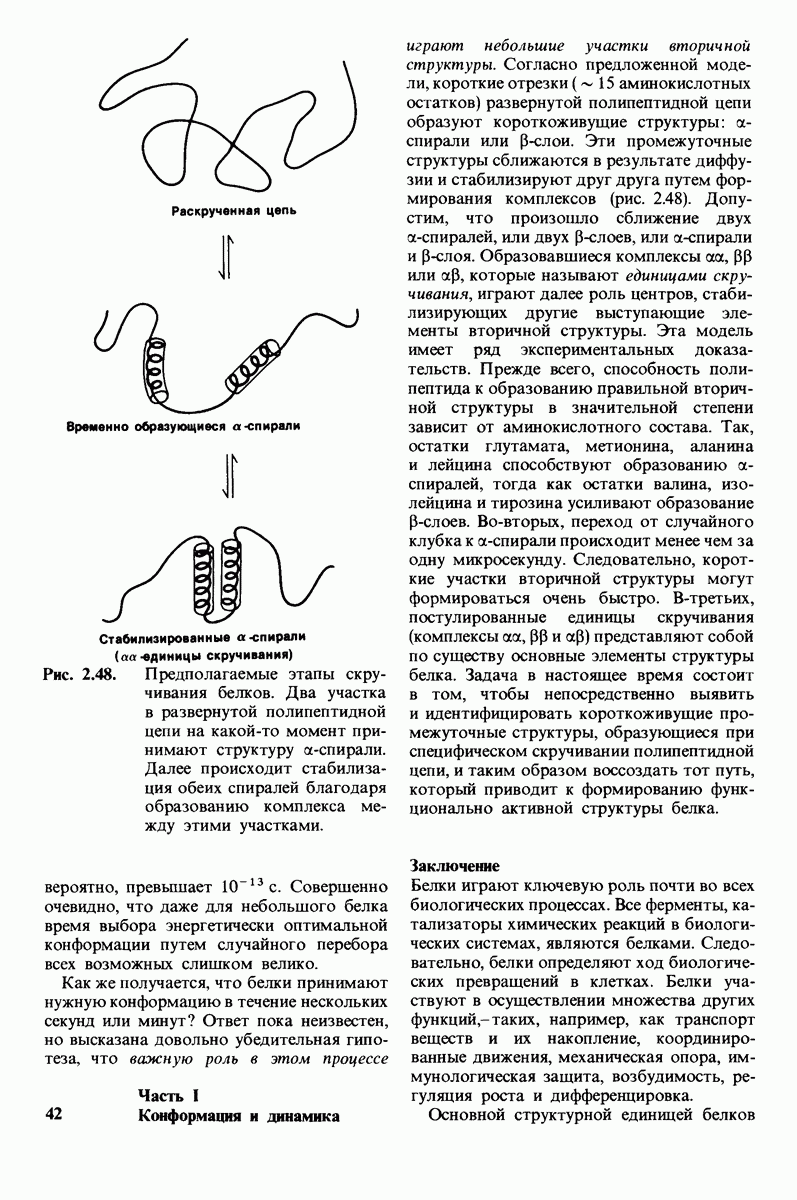
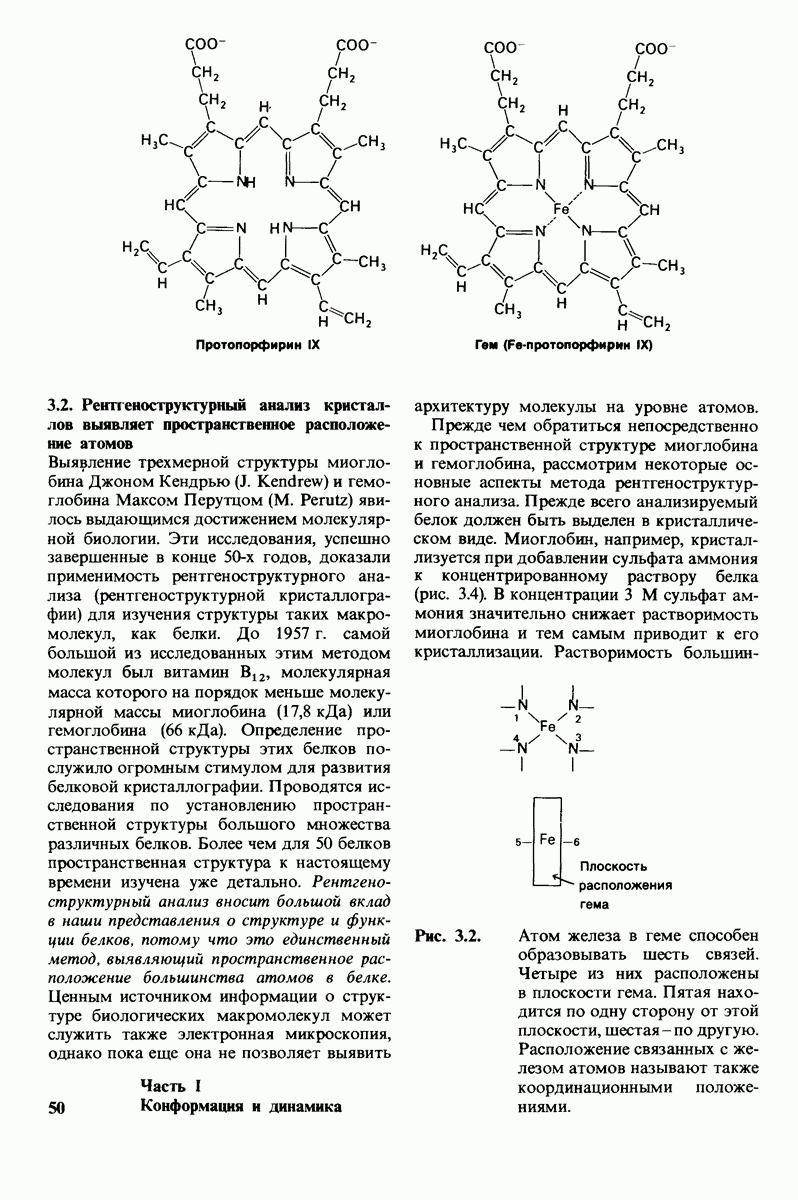
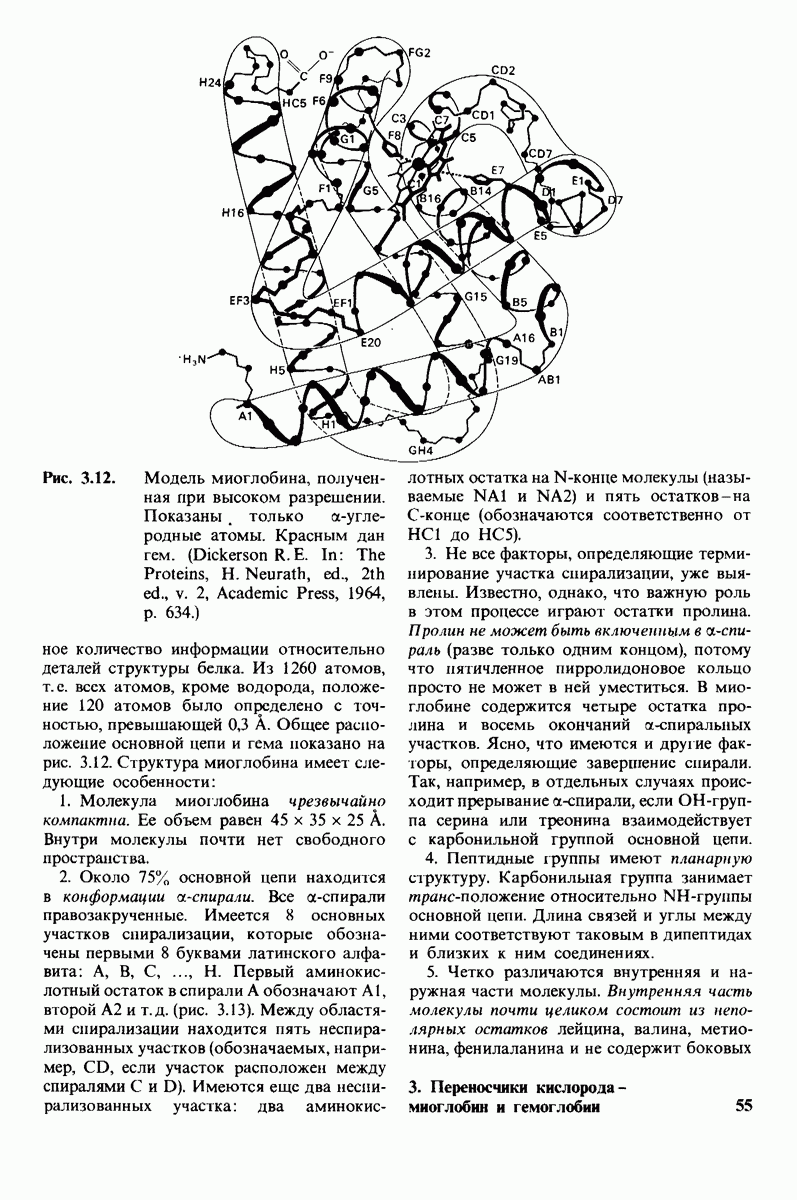
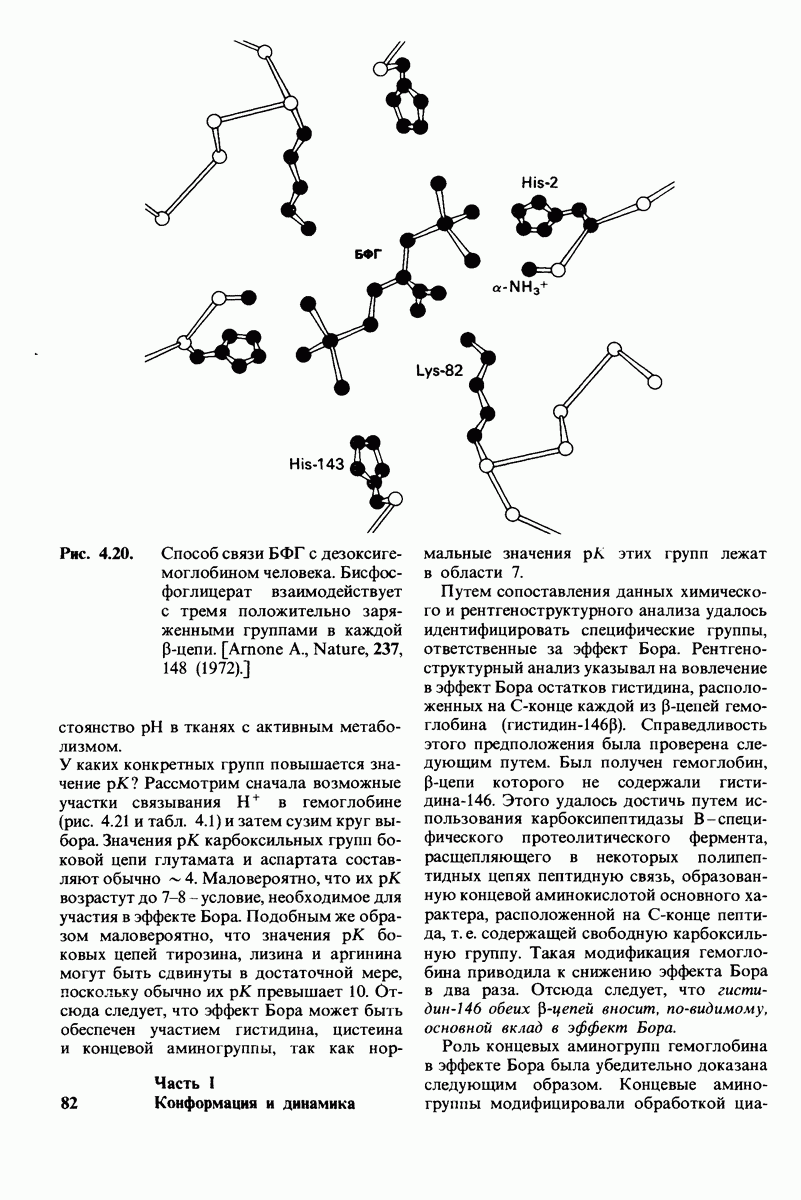
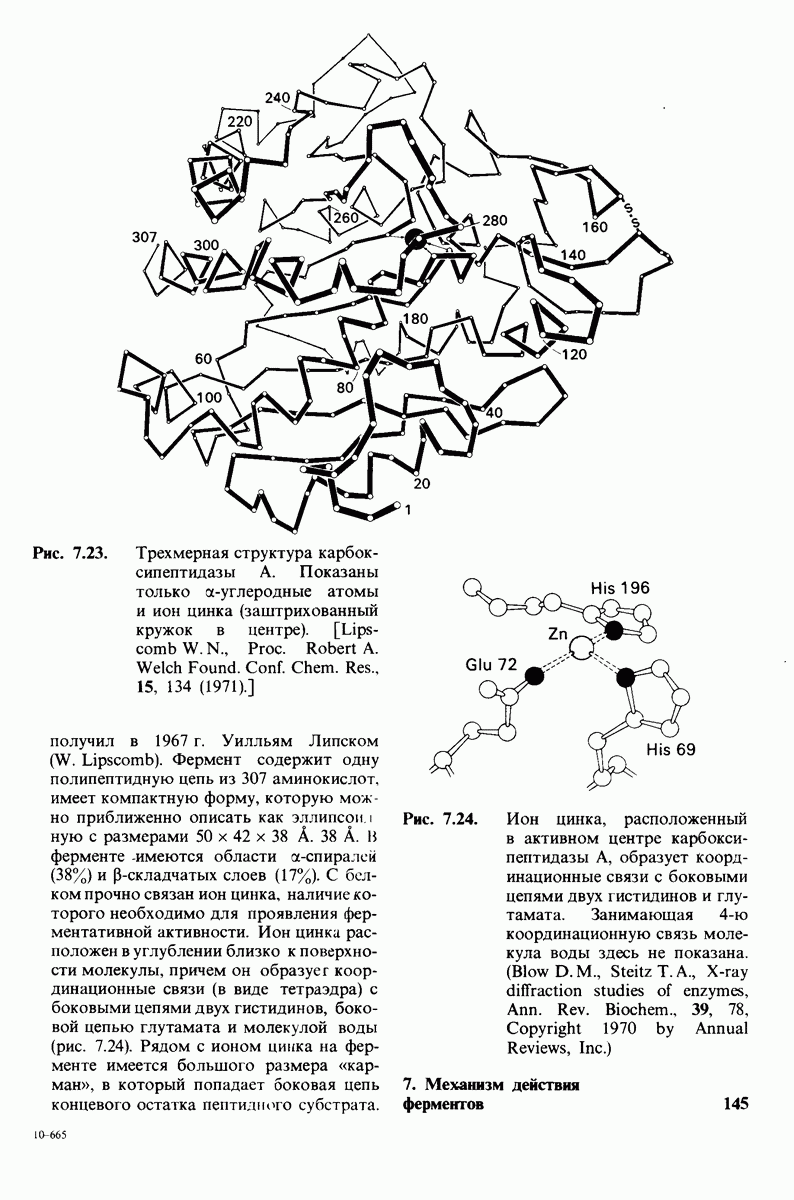
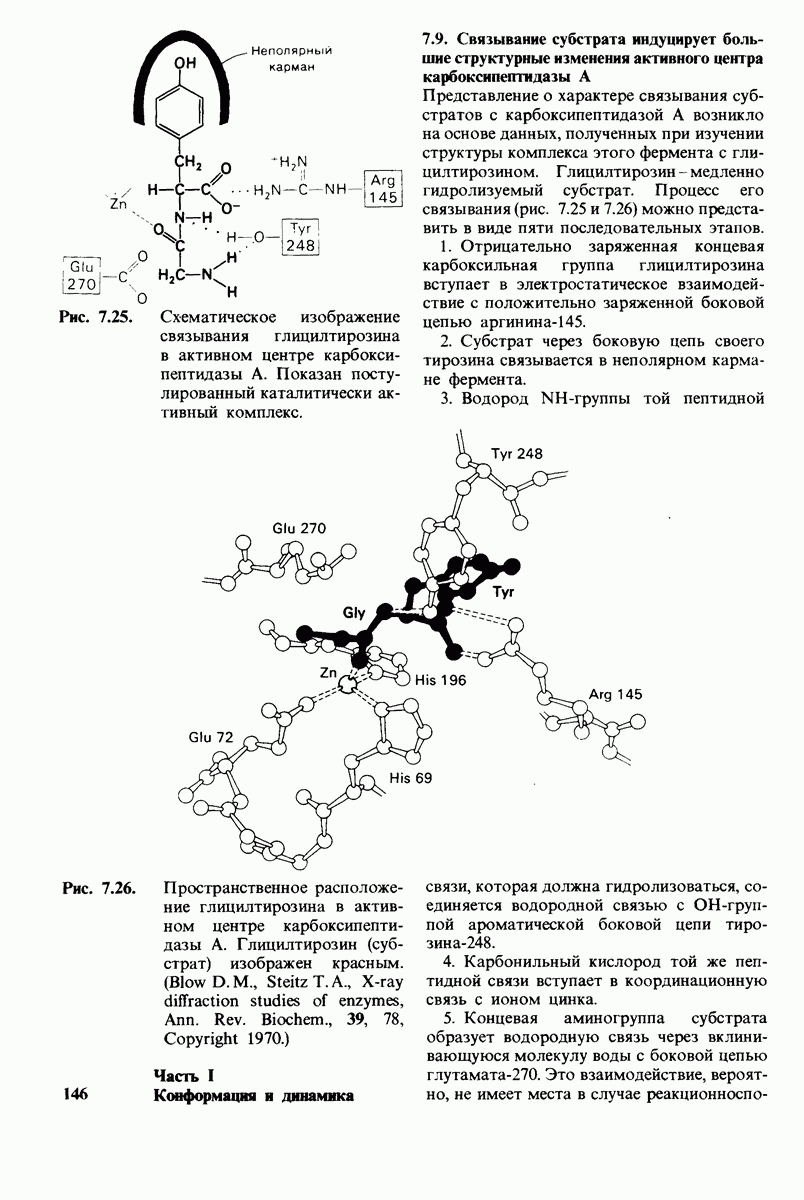
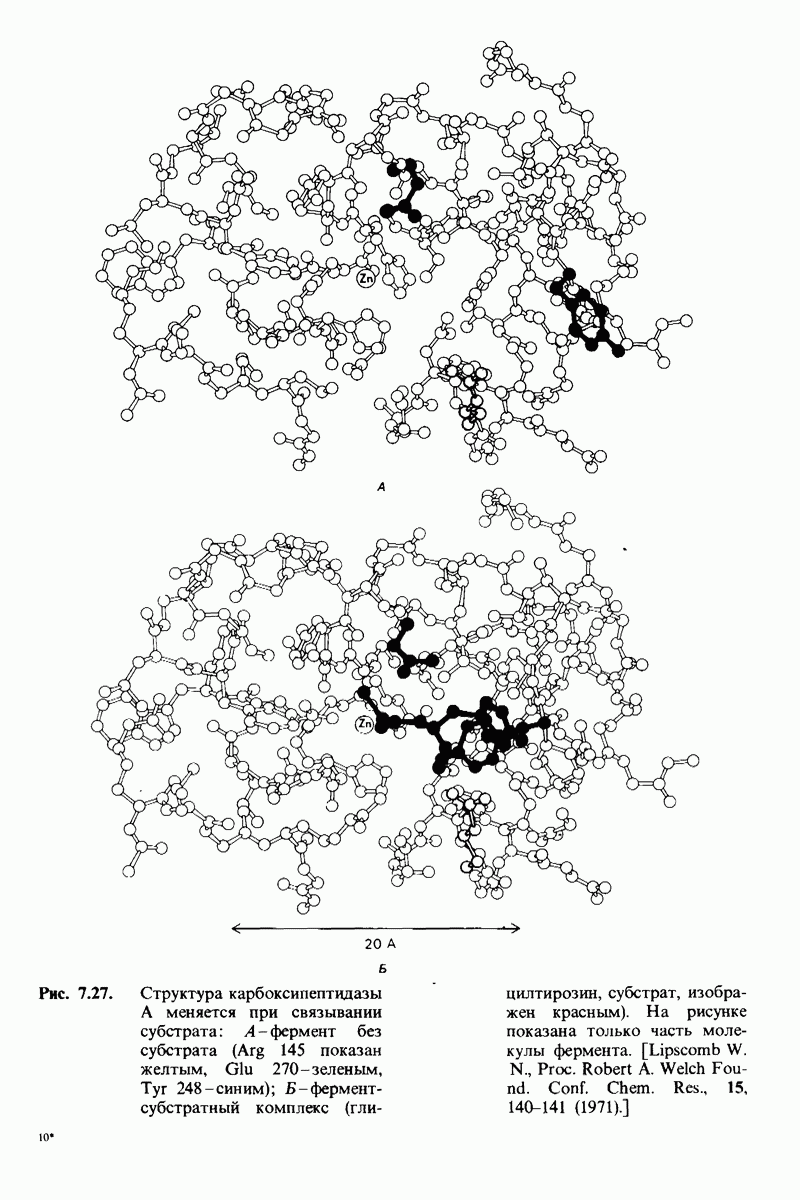
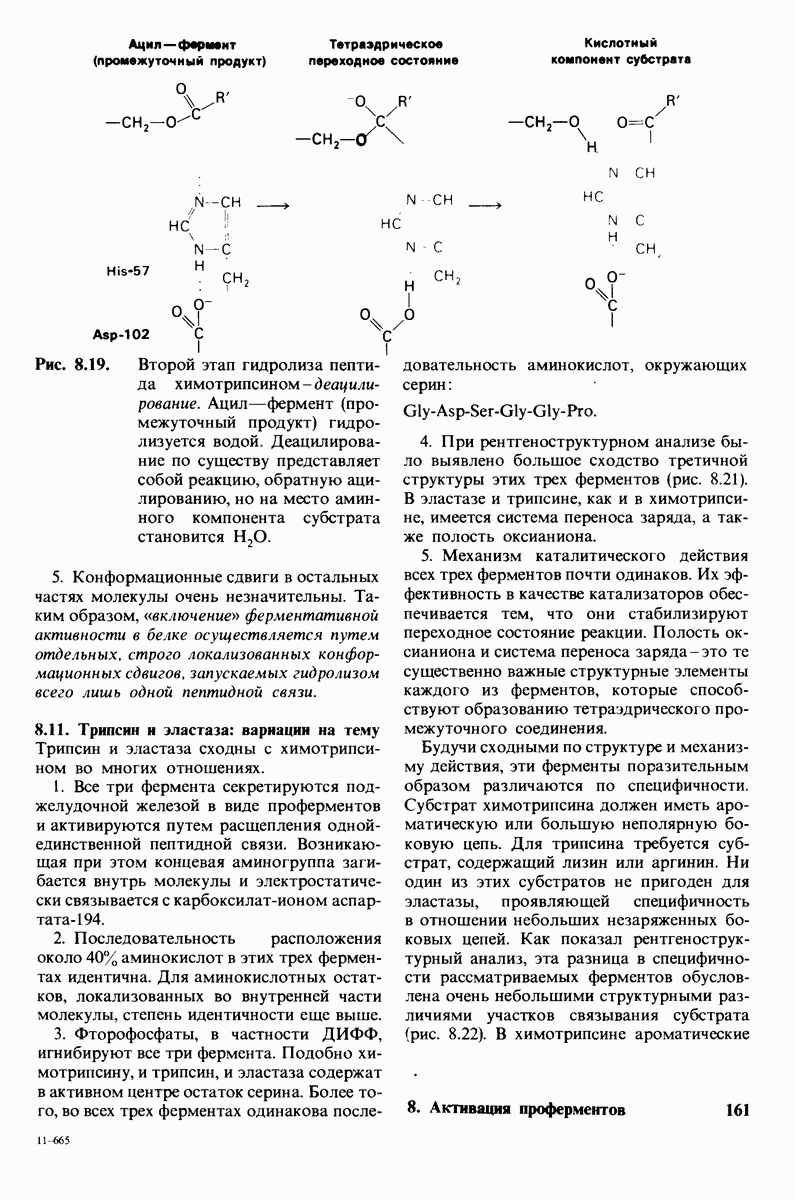
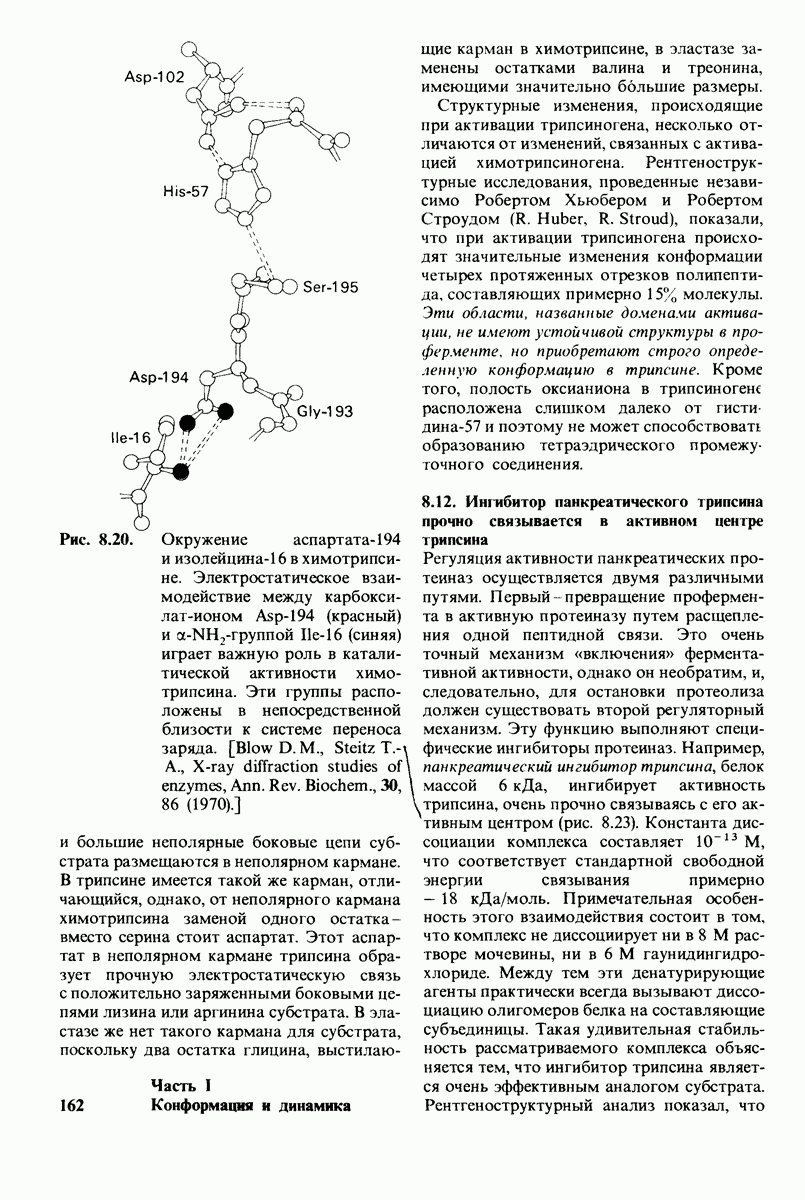
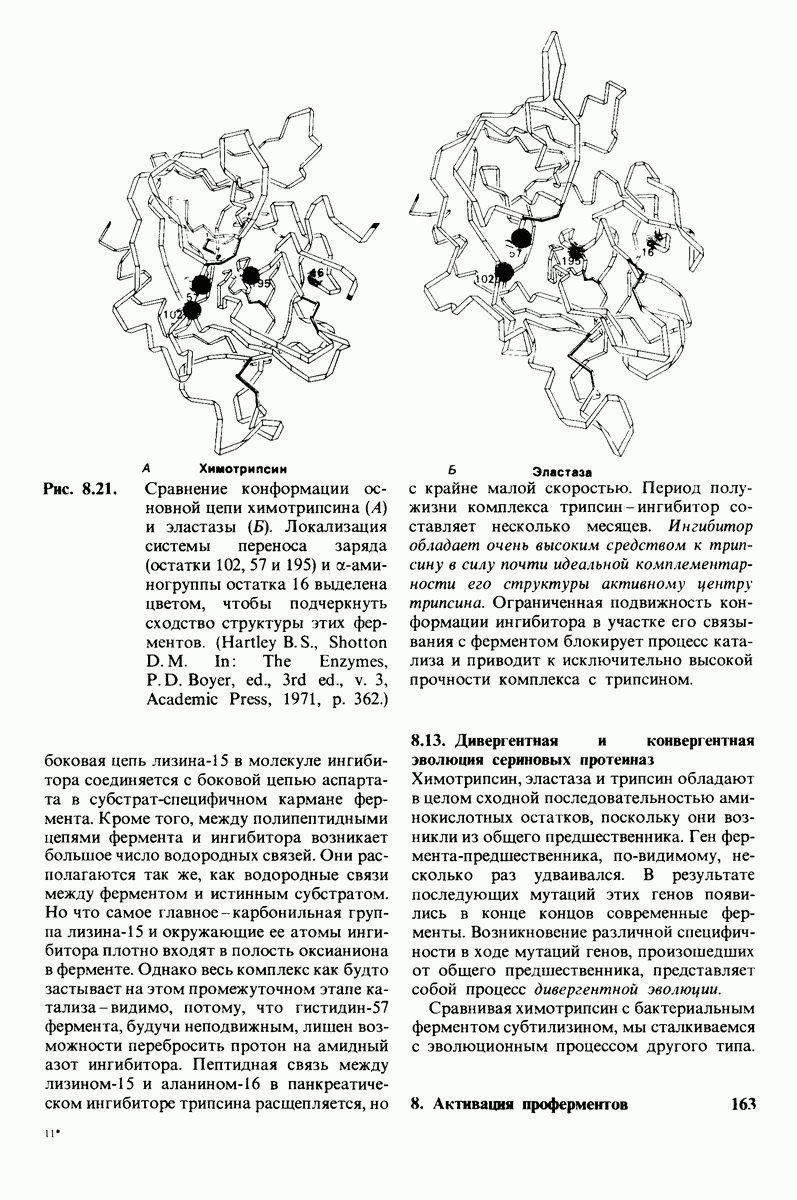
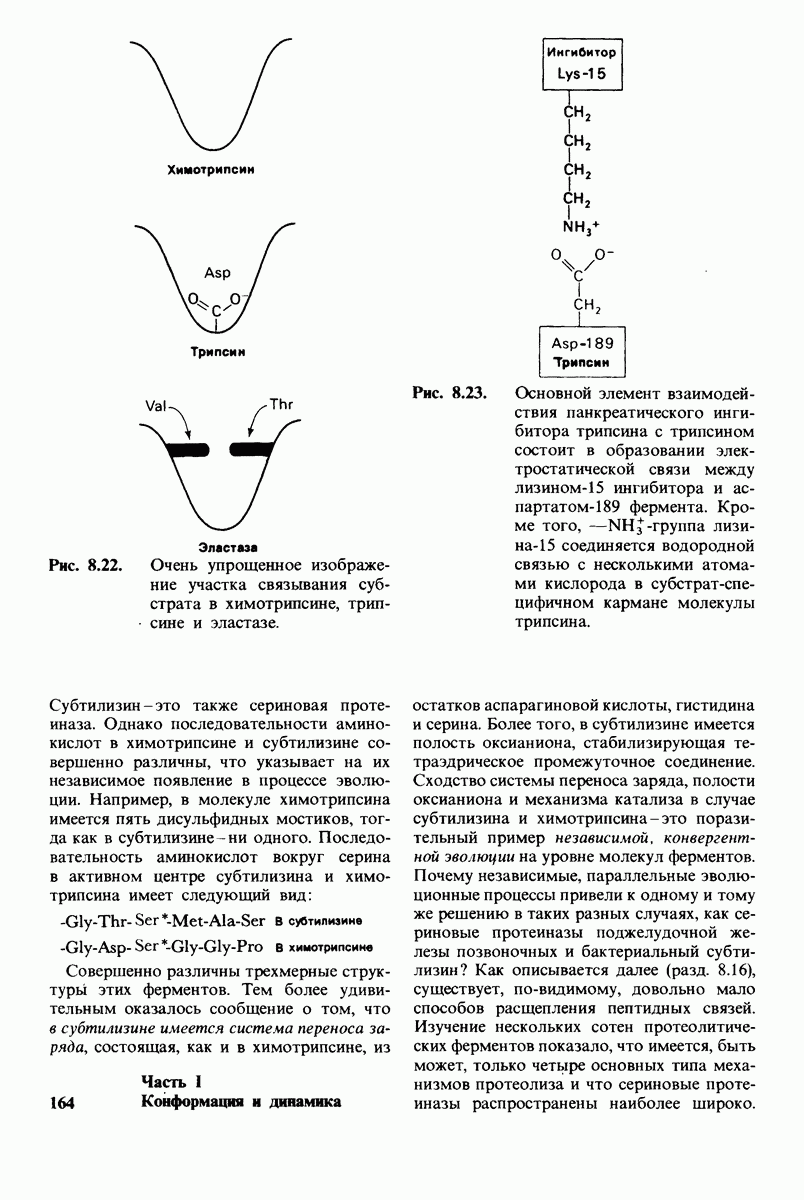
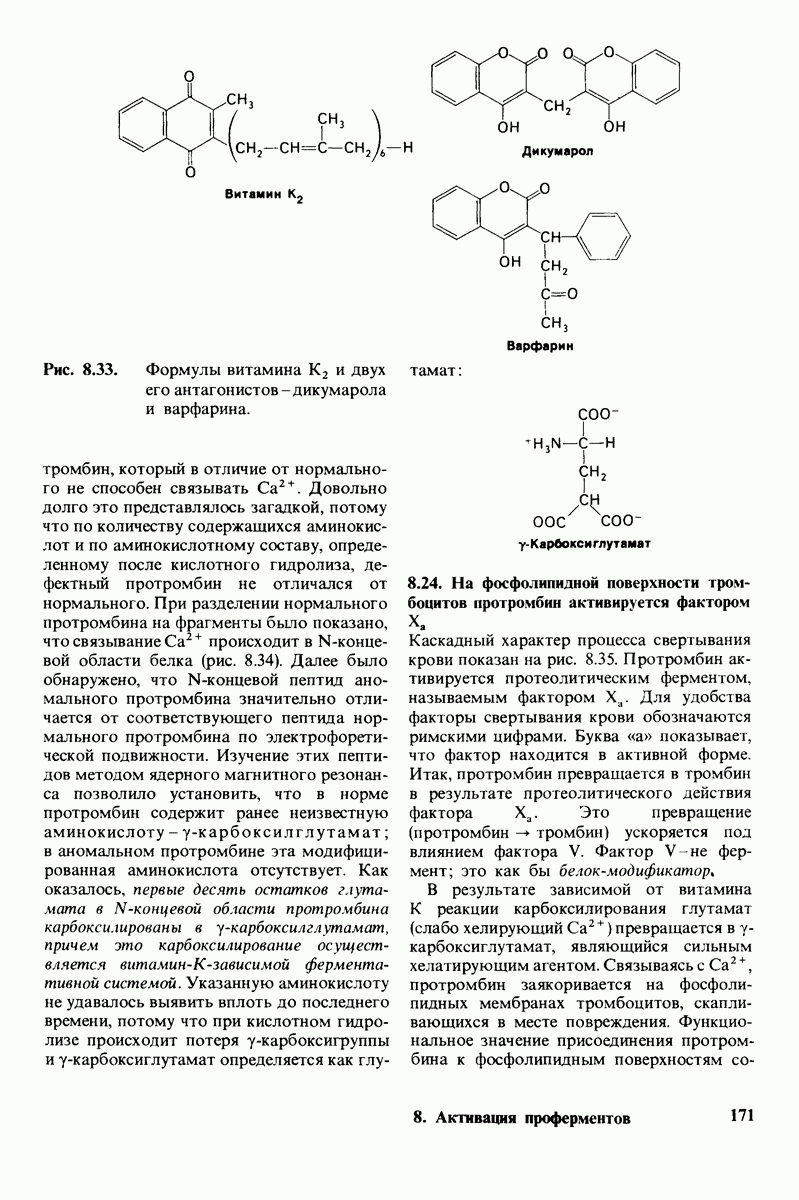
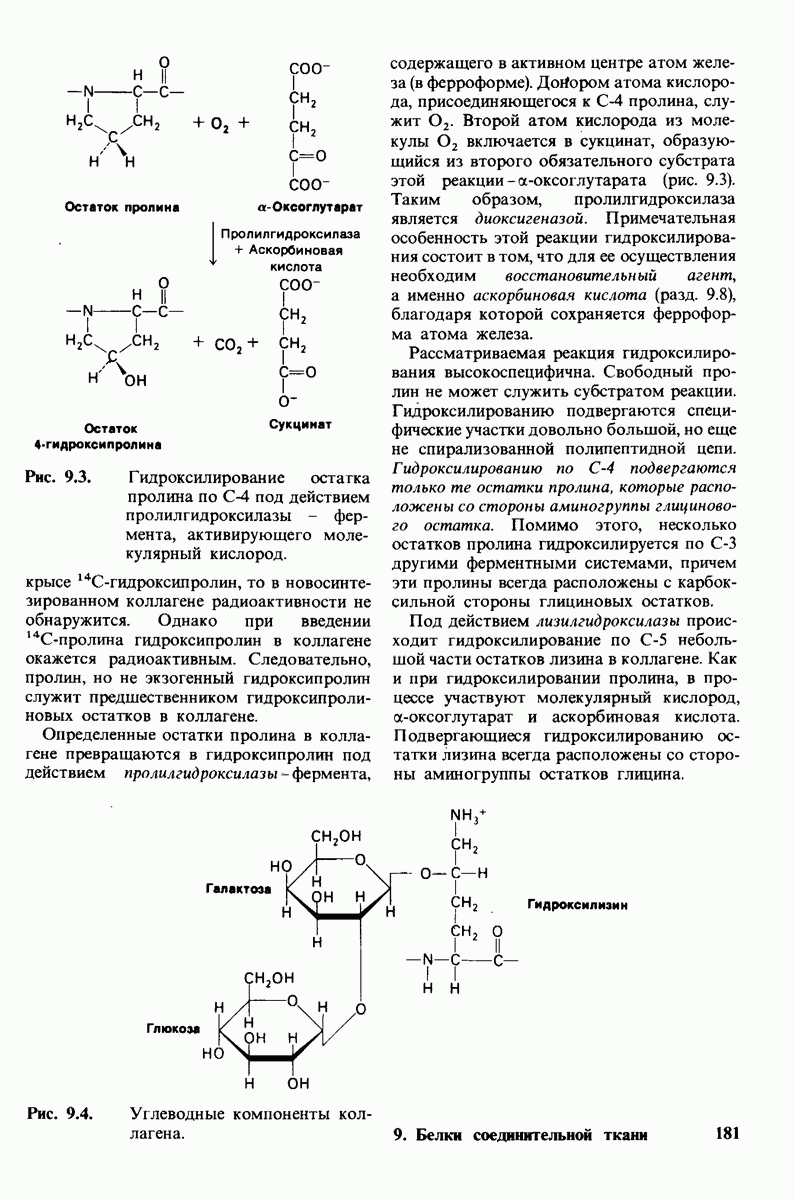
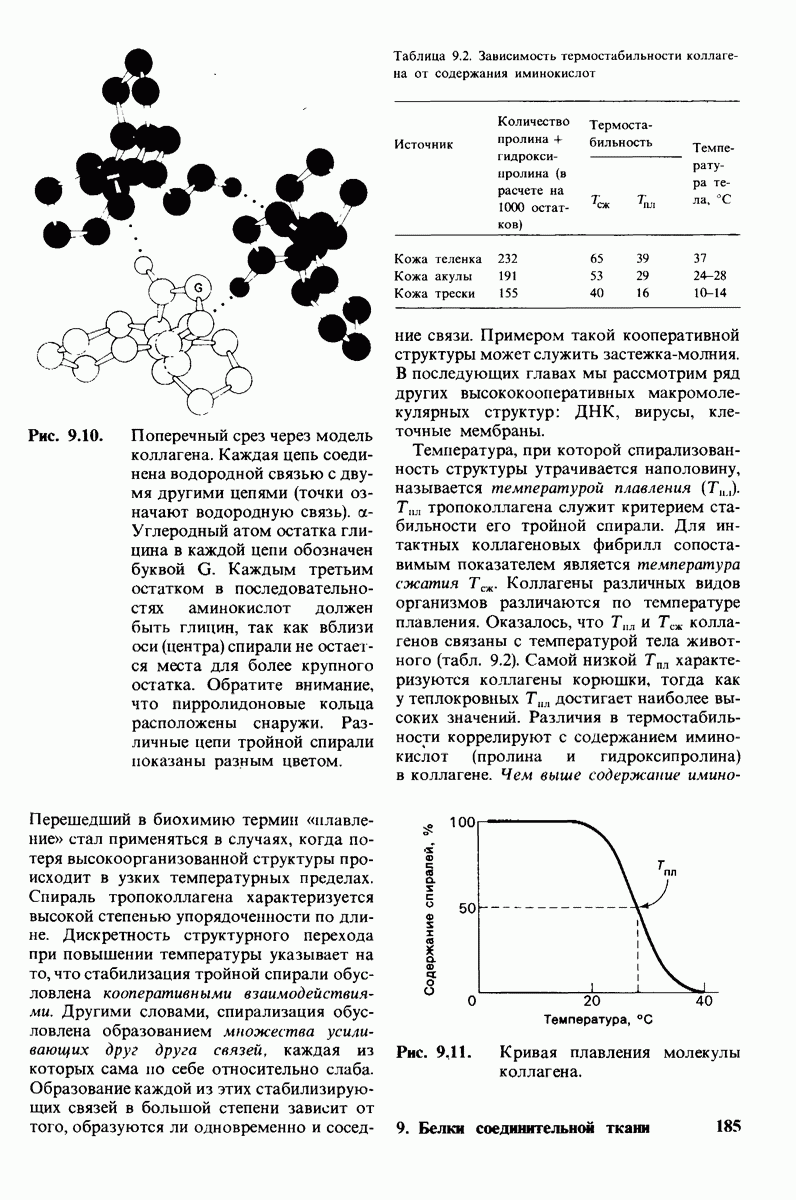
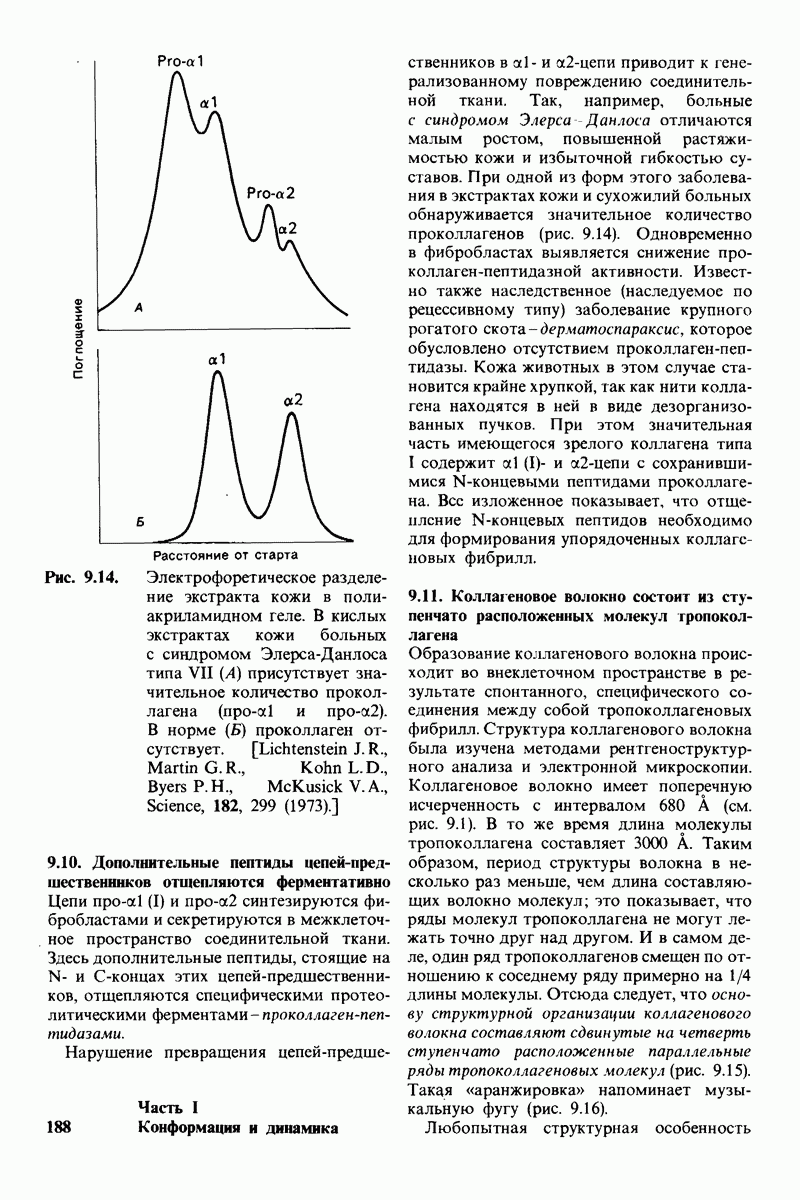
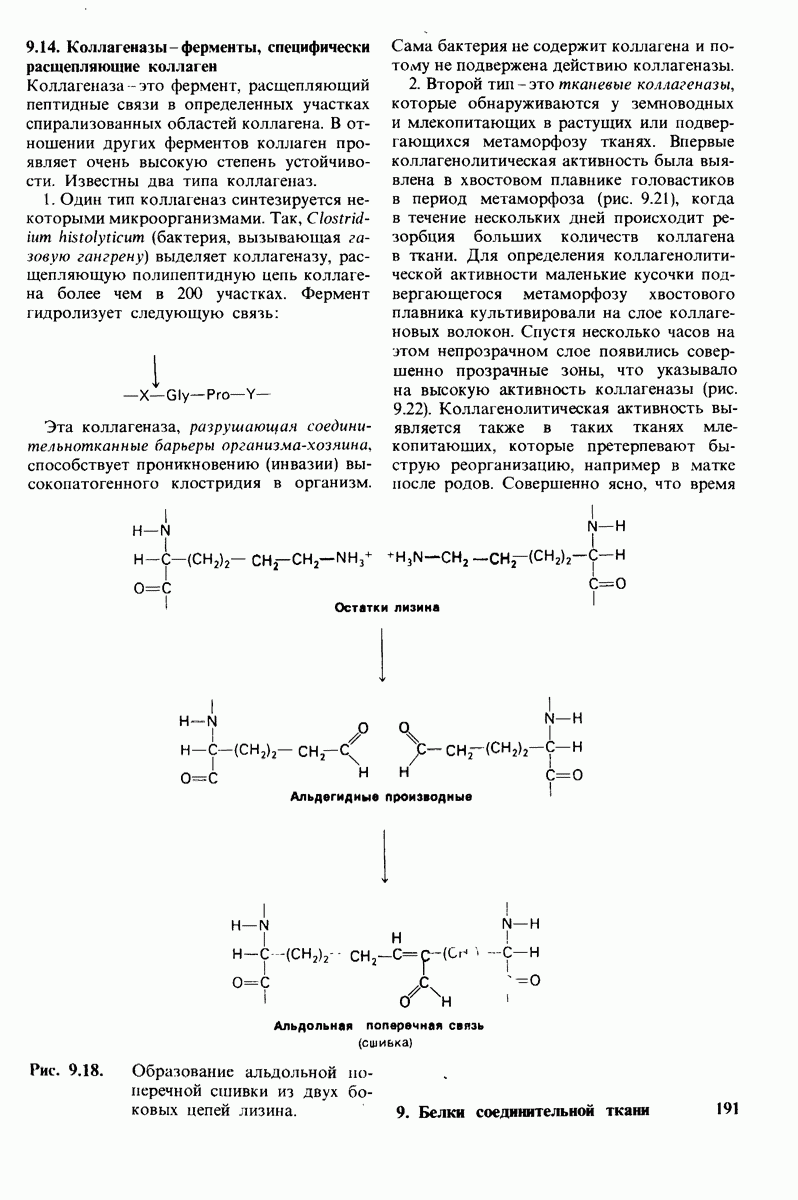
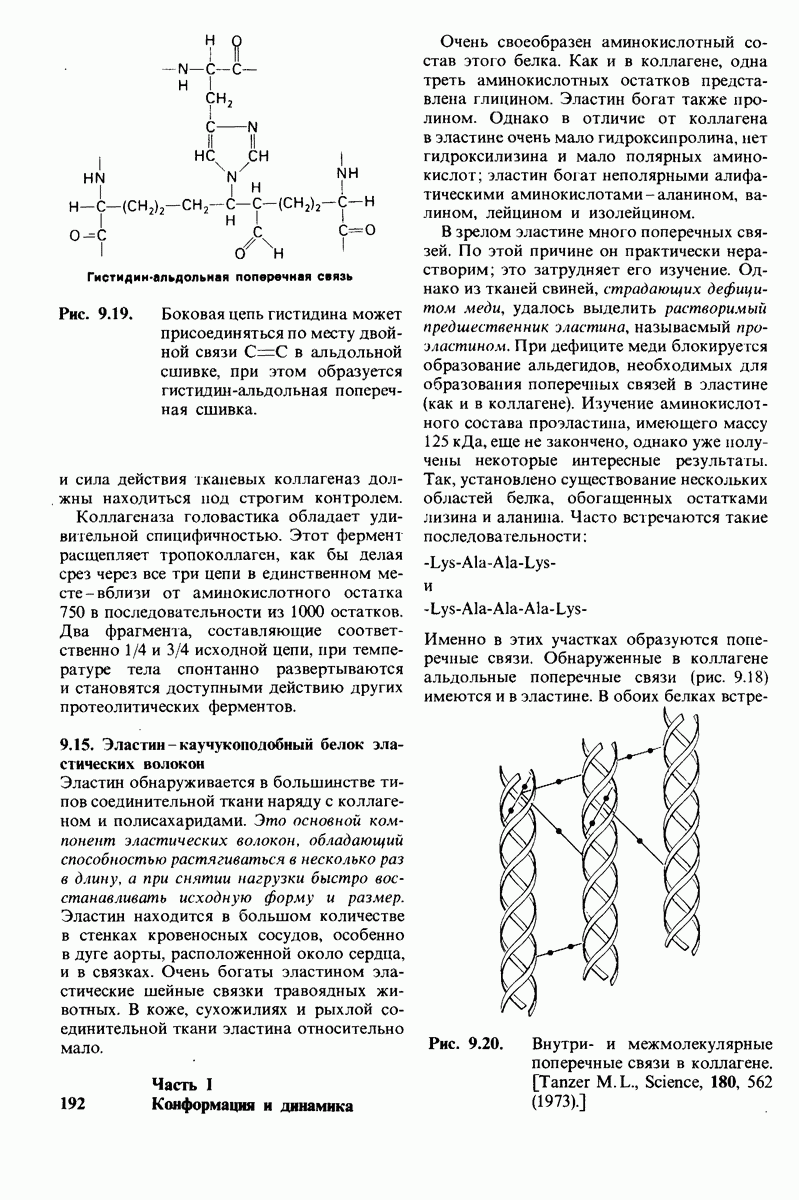
6.17. Согласованный механизм аллостерических взаимодействий
Изящную и четкую модель кинетики аллостерических ферментов предложили в 1965 г. Жак Моно, Джефри Уайман и Жан-Пьер Шанже (1 Monod,J, Wyman, J.-P.Chan-geux). Используя их подход, рассмотрим аллостерический фермент, состоящий из двух идентичных субъединиц, каждая с одним активным центром. Допустим, что субъединицы могут находиться в двух конформациях -  Конформация
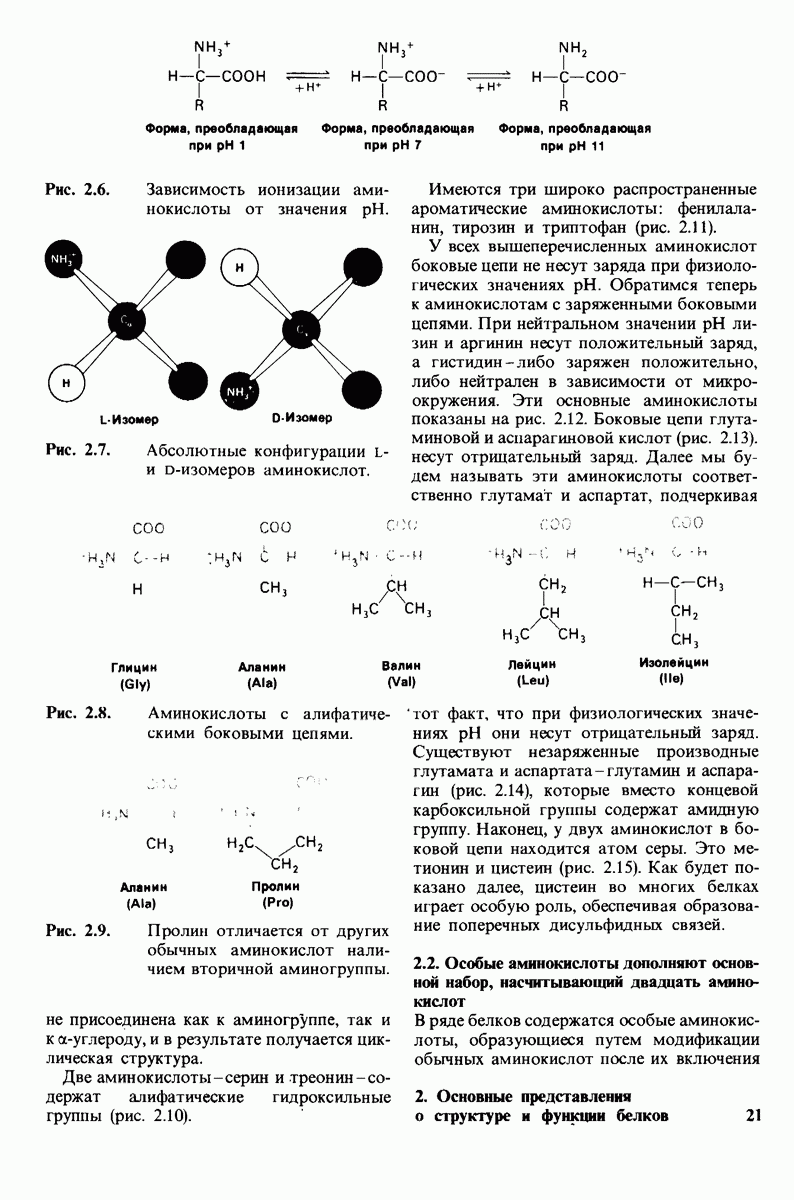
Конформация  релаксированная) обладает высоким сродством к субстрату, тогда как конформация
релаксированная) обладает высоким сродством к субстрату, тогда как конформация  -низким сродством (рис, 6.20). Вспомним, что так же обозначались две формы четвертичной структуры гемоглобина (разд. 4.10), Формы
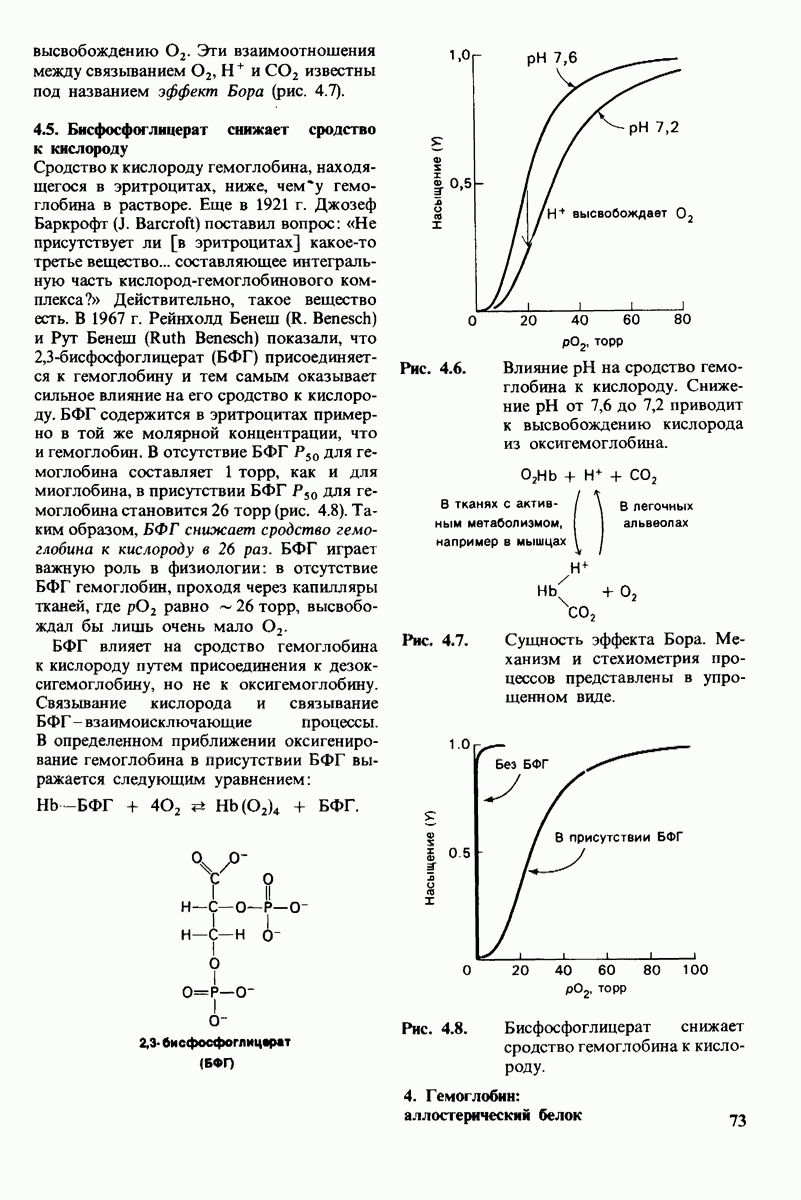
-низким сродством (рис, 6.20). Вспомним, что так же обозначались две формы четвертичной структуры гемоглобина (разд. 4.10), Формы  могут переходить одна в другую. В данной модели делается важное допущение, что для сохранения симметрии димера обе субъединицы должны находиться в одном и том же конформационном состоянии. Таким образом разрешены состояния
могут переходить одна в другую. В данной модели делается важное допущение, что для сохранения симметрии димера обе субъединицы должны находиться в одном и том же конформационном состоянии. Таким образом разрешены состояния  и не разрешено состояние
и не разрешено состояние  Символами
Символами  обозначают разрешенные состояния в отсутствие субстрата и L-соотношение их концентраций:
обозначают разрешенные состояния в отсутствие субстрата и L-соотношение их концентраций:

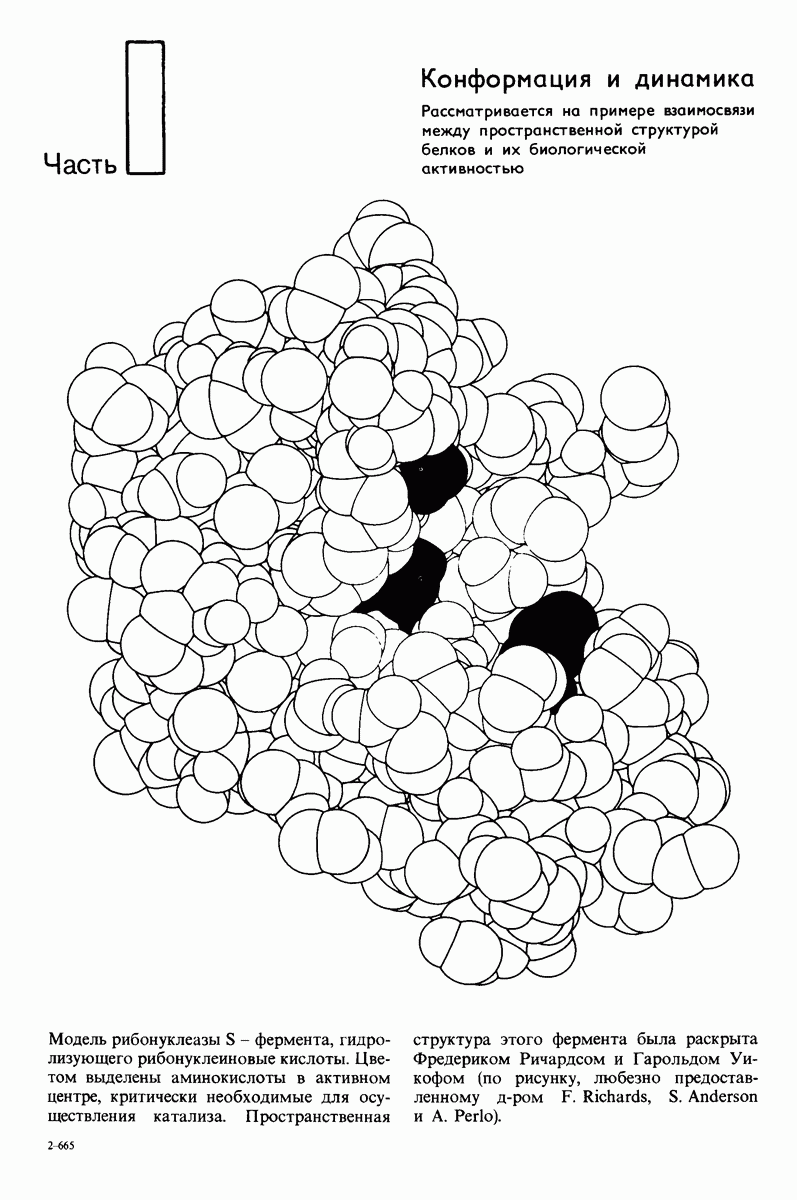
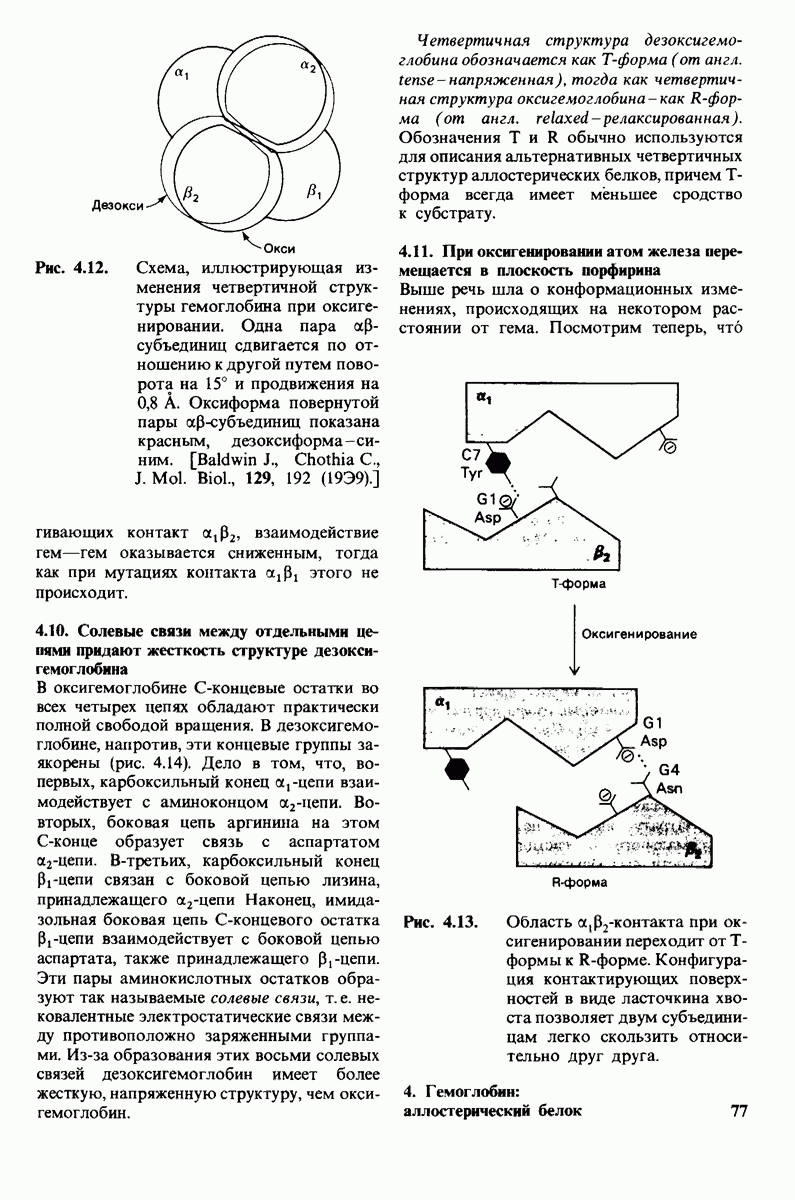
Чтоб упростить рассуждения, допустим, что субстрат не присоединяется к  -форме фермента. В R-форме димер может связать одну или две молекулы субстрата; эти
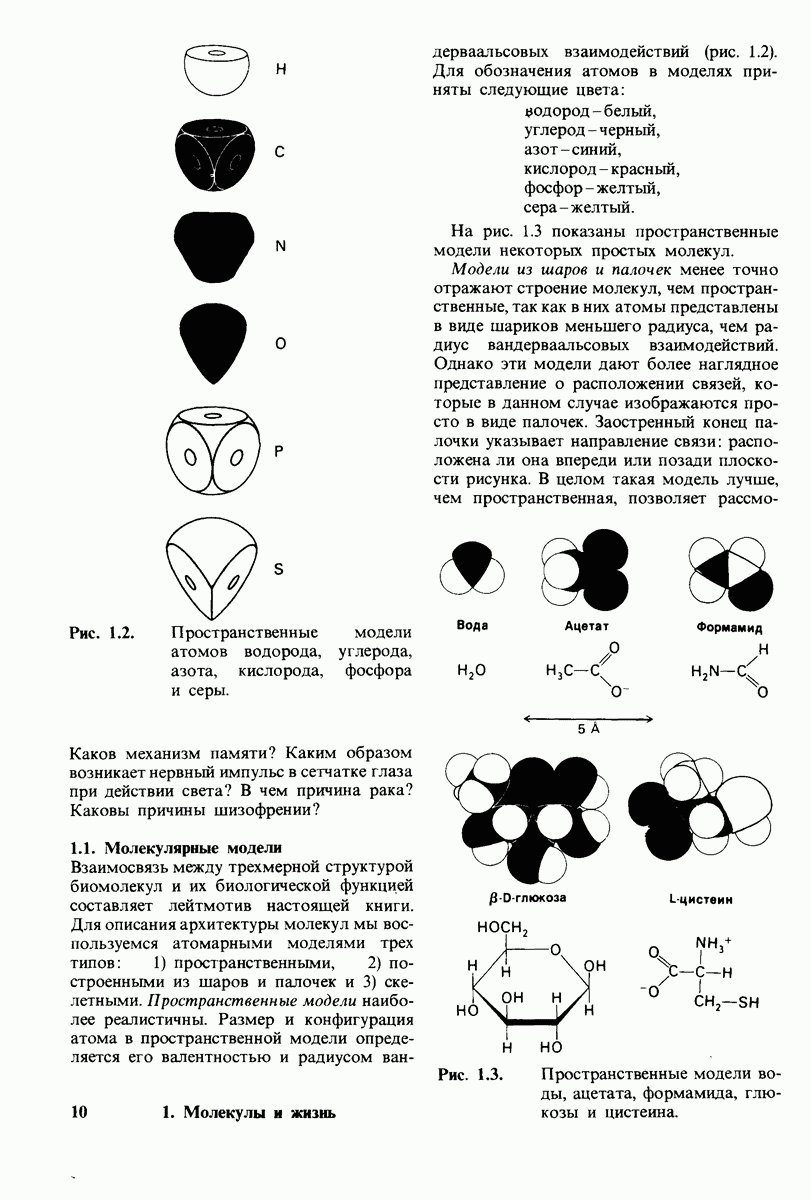
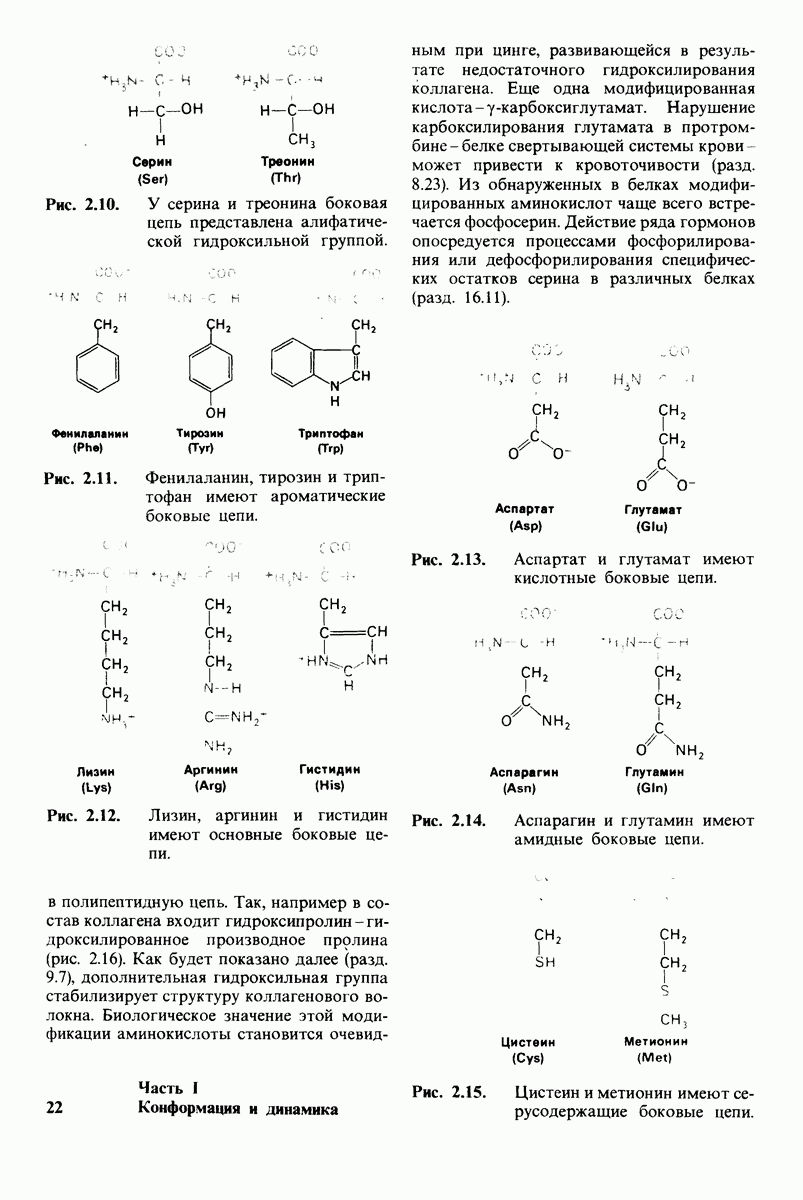
-форме фермента. В R-форме димер может связать одну или две молекулы субстрата; эти
состояния обозначают соответственно 

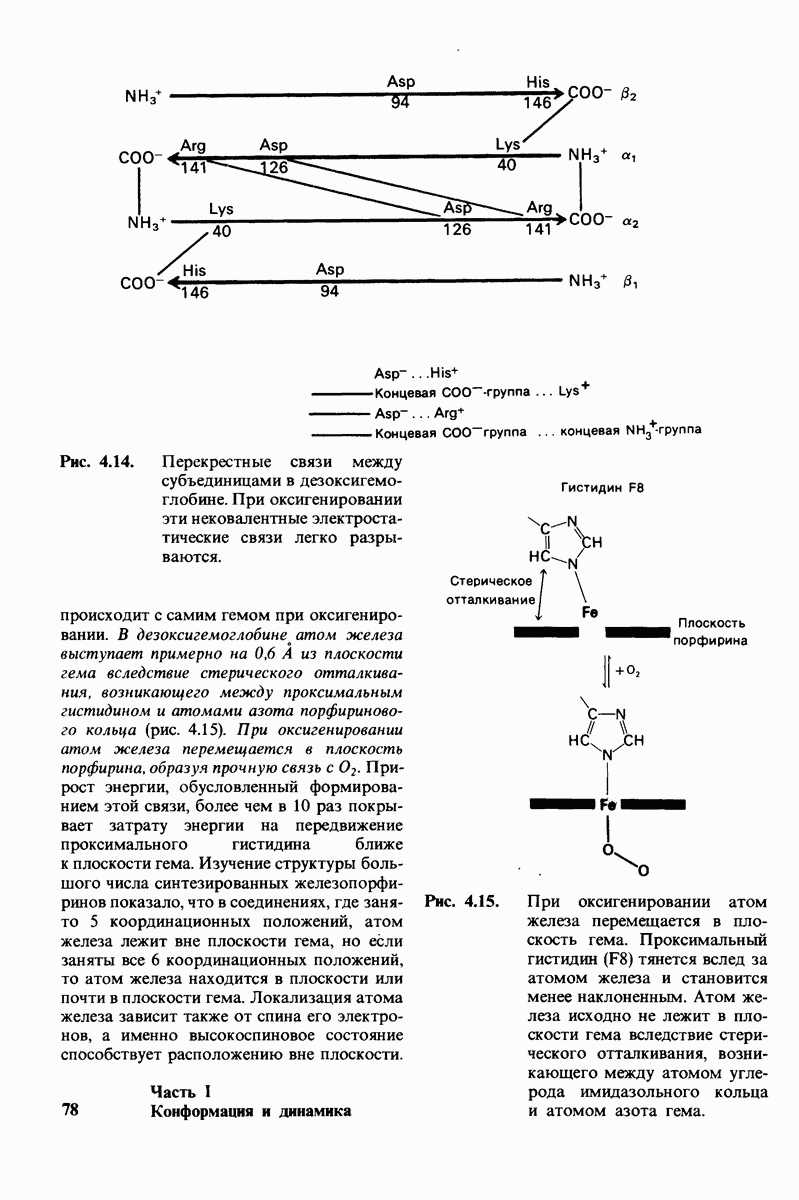
Согласно полученному уравнению, присоединение как первой, так и второй молекулы субстрата к R-форме димерного фермента имеет одну и ту же микроскопическую константу диссоциации  Коэффициент 2 в уравнении (31) указывает на то, что субстрат может связаться с любым из двух активных центров на
Коэффициент 2 в уравнении (31) указывает на то, что субстрат может связаться с любым из двух активных центров на  с образованием
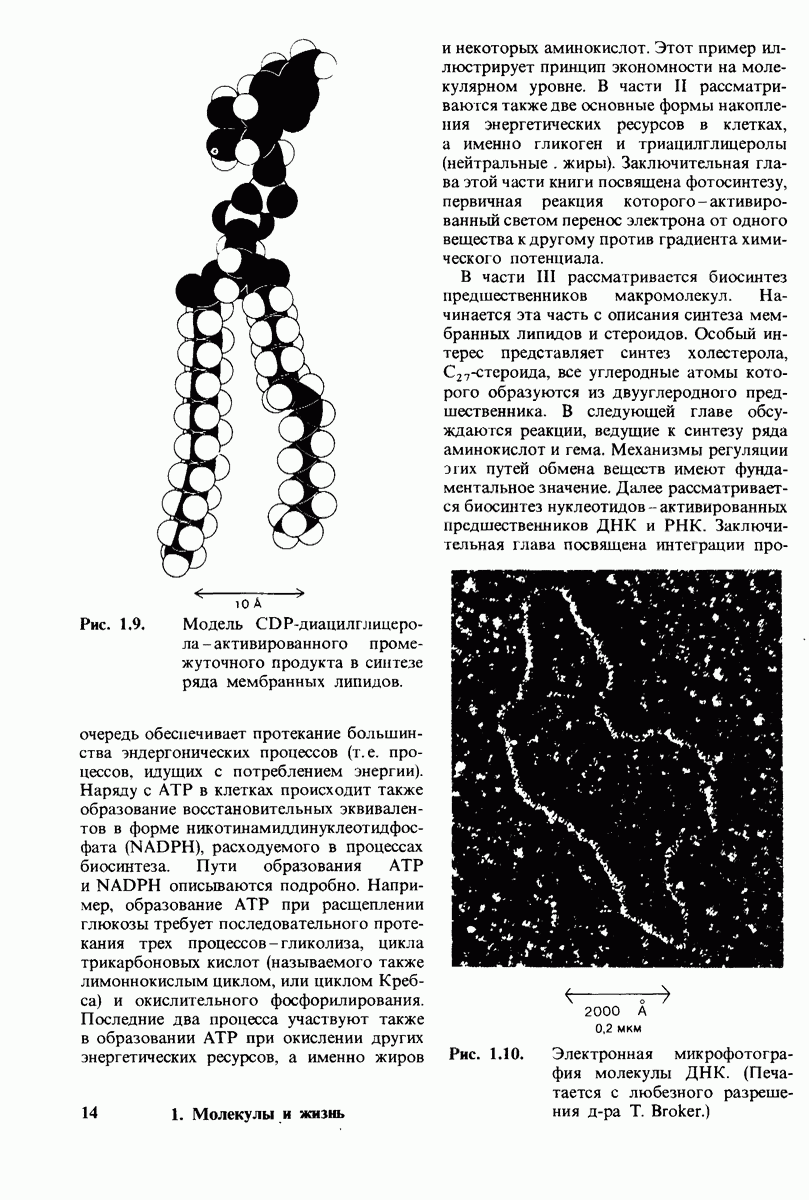
с образованием  и аналогичным образом субстрат может высвободиться из любого из активных центров на
и аналогичным образом субстрат может высвободиться из любого из активных центров на  с образованием R
с образованием R
Выразим степень насыщения У (т.е. долю активных центров, имеющих связанный субстрат) как функцию концентрации субстрата

Произведя замены в этом уравнении в соответствии с уравнениями (27) и (31), получаем искомое выражение для У:

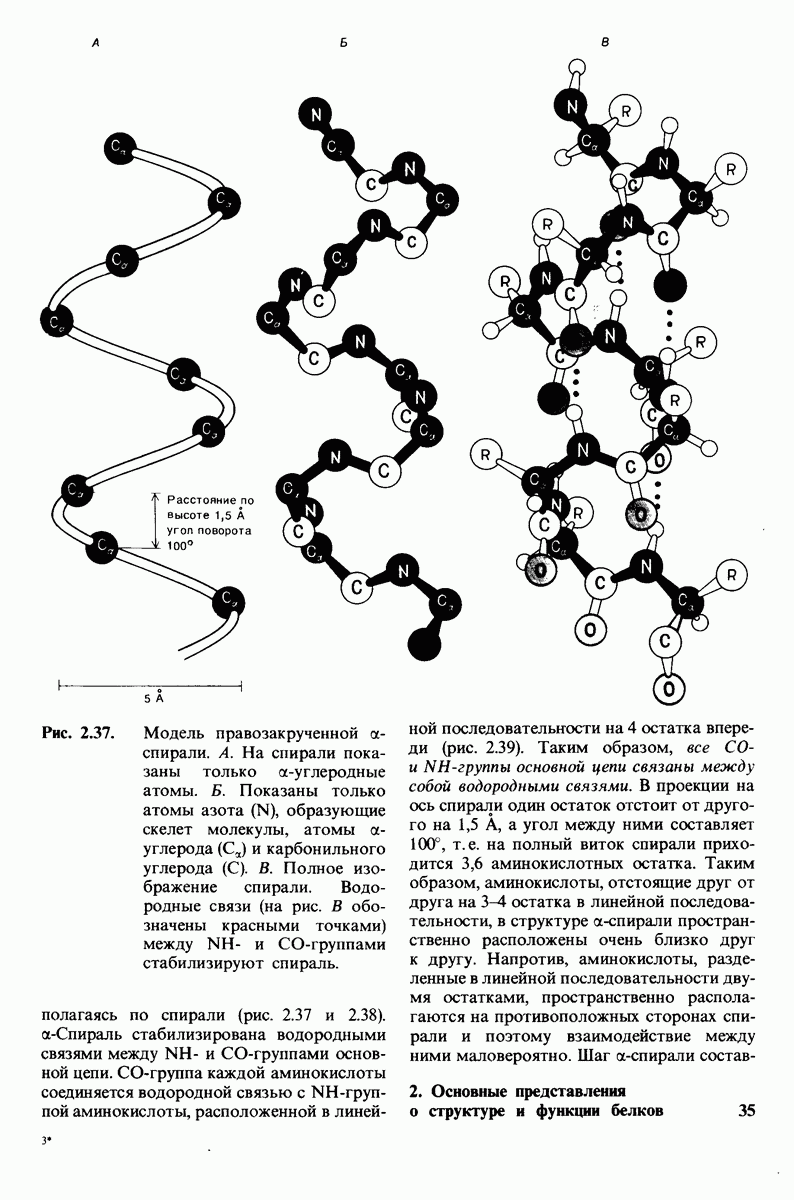
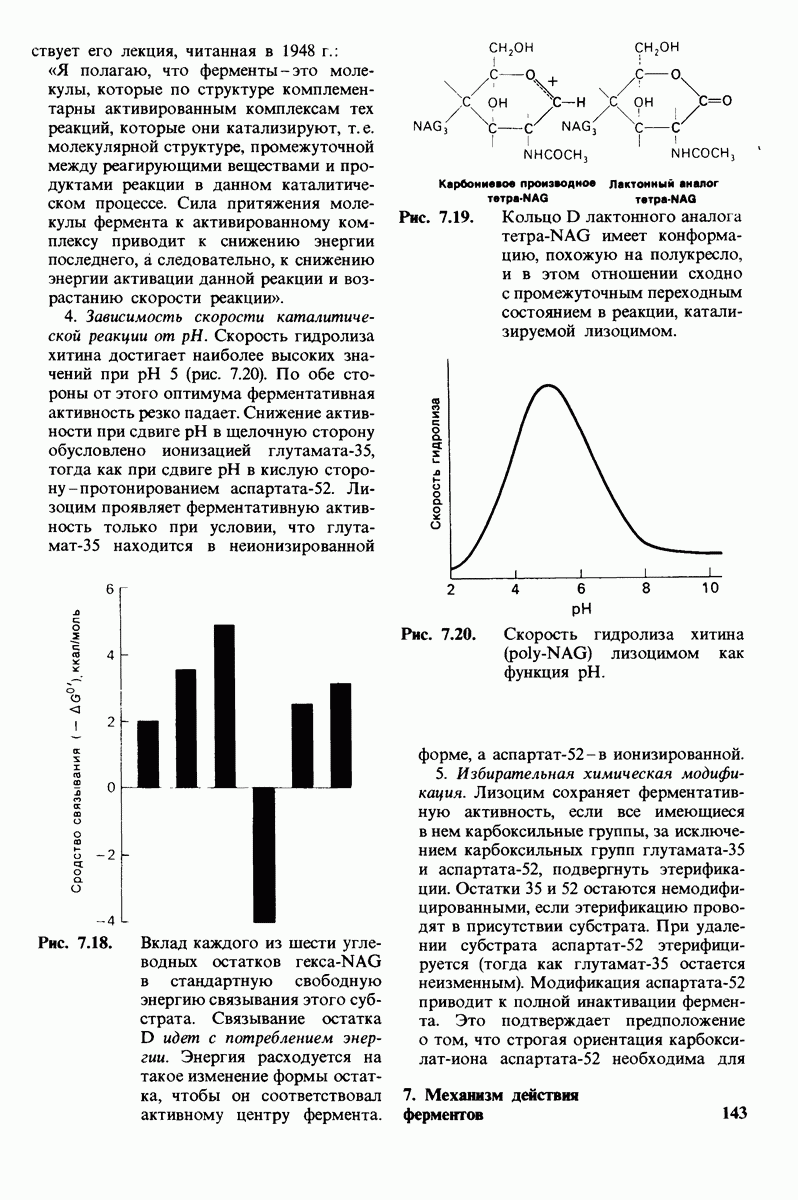
Изобразим уравнение (33) графически, приняв  Зависимость
Зависимость  от
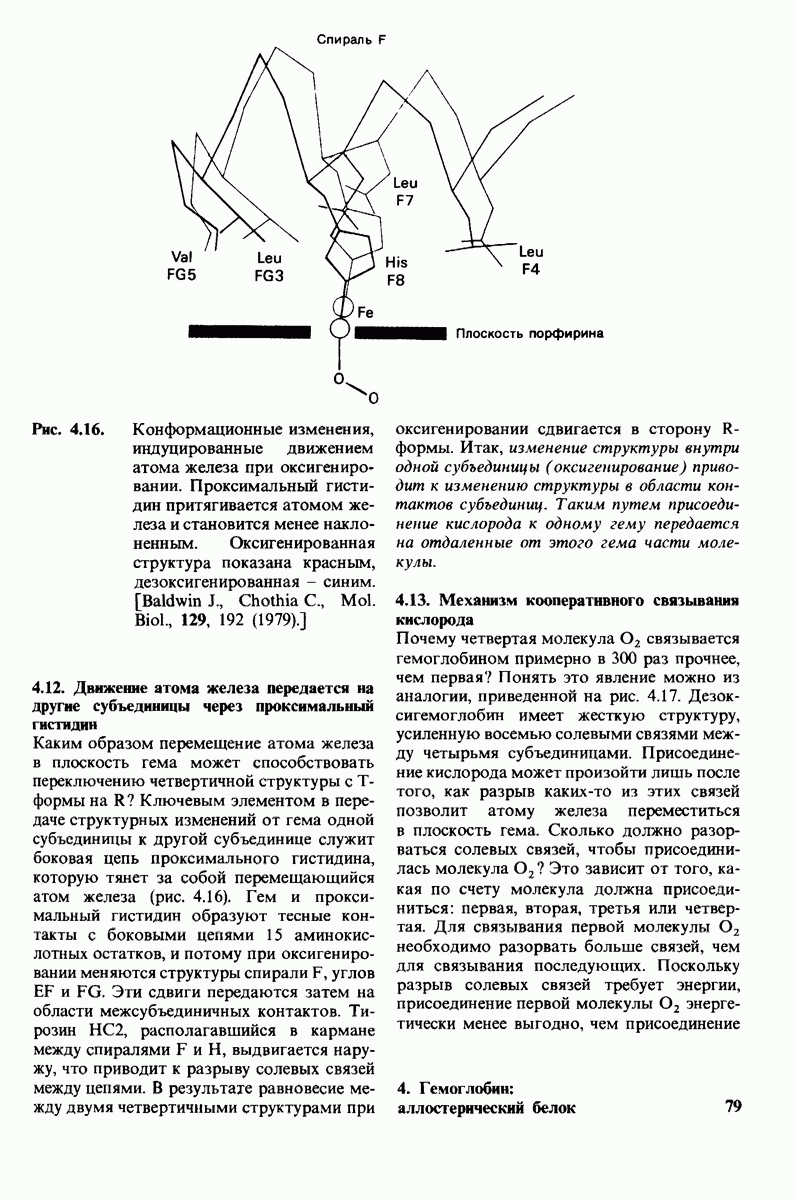
от  выражается сигмоидной, а не гиперболической кривой (рис. 6.23). Другими словами, это уравнение соответствует кооперативному связыванию субстрата. Если число оборотов в расчете на один активный центр одинаково для фермент-субстратных комплексов
выражается сигмоидной, а не гиперболической кривой (рис. 6.23). Другими словами, это уравнение соответствует кооперативному связыванию субстрата. Если число оборотов в расчете на один активный центр одинаково для фермент-субстратных комплексов  то график зависимости скорости реакции от концентрации субстрата также будет сигмоидным, поскольку
то график зависимости скорости реакции от концентрации субстрата также будет сигмоидным, поскольку

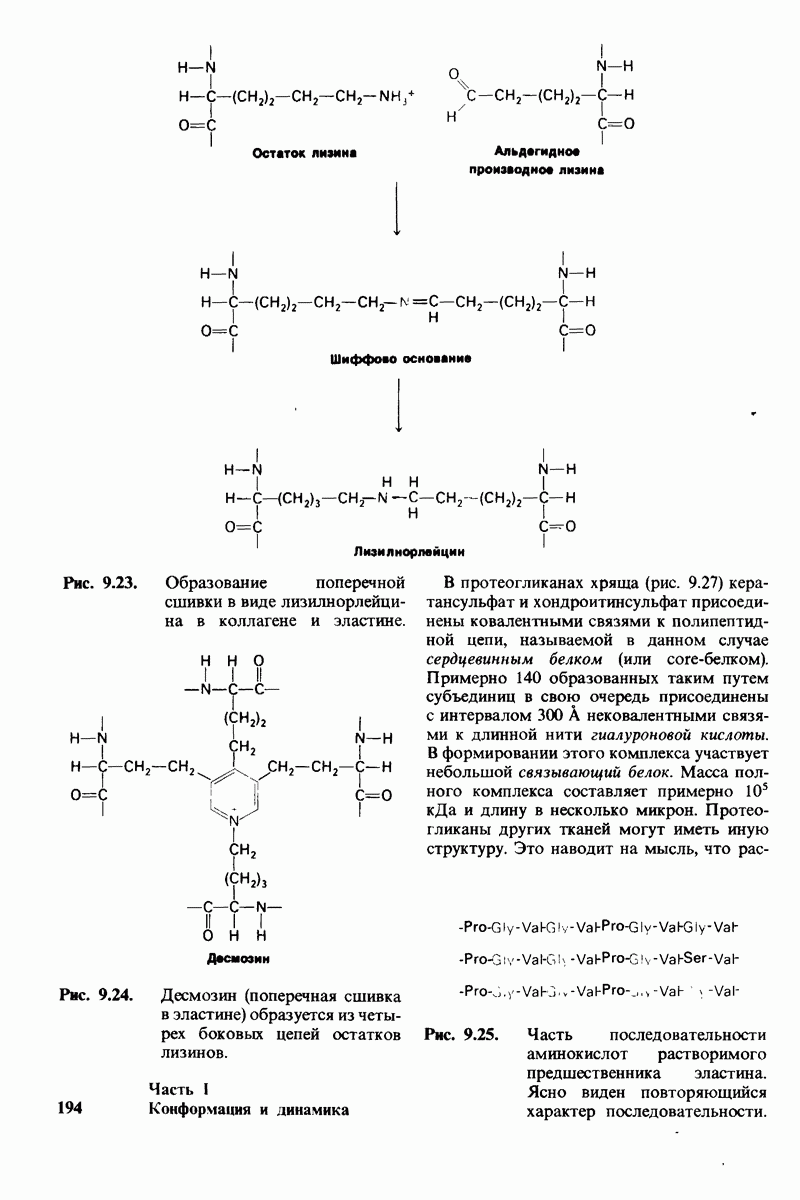
Рассмотрим теперь этот процесс связывания (рис. 6.21). В отсутствие субстрата почти все молекулы фермента находятся в  -форме. В приведенном выше примере на 104 молекул в
-форме. В приведенном выше примере на 104 молекул в  -форме приходится только
-форме приходится только

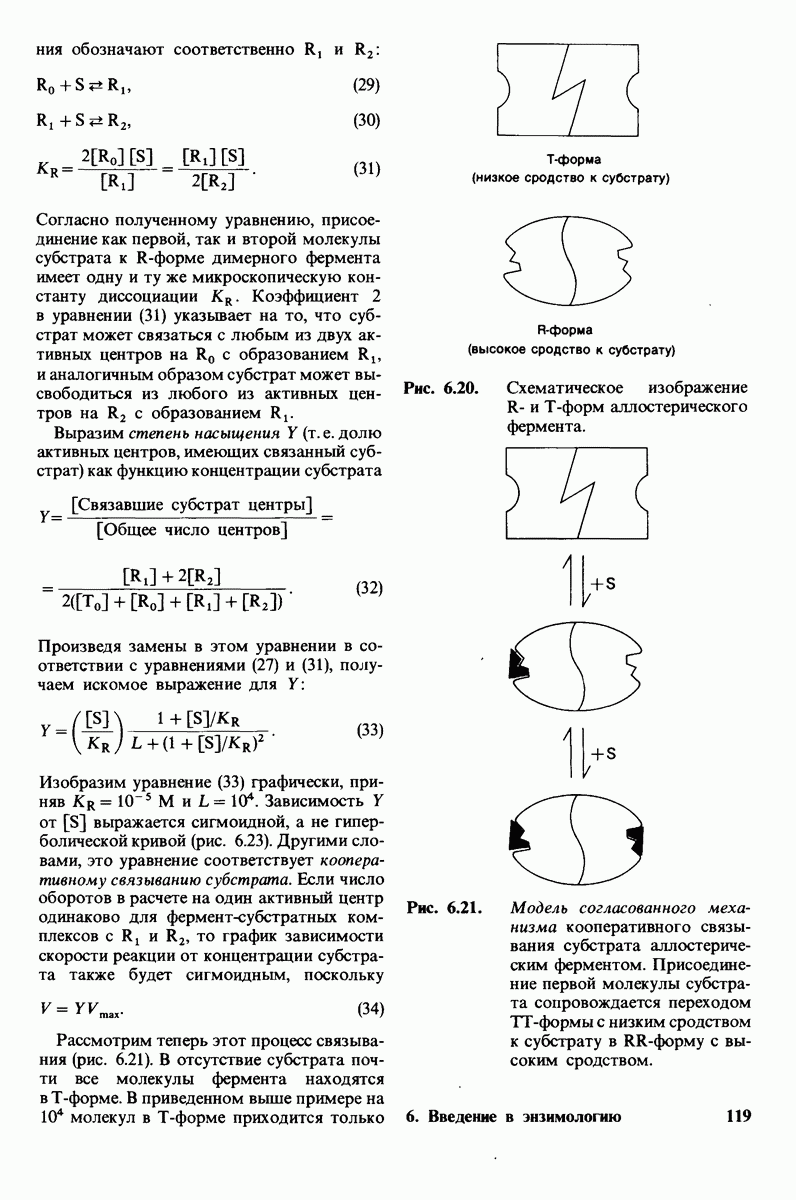
Рис. 6.20. Схематическое изображение  и
и  -форм аллостерического фермента.
-форм аллостерического фермента.

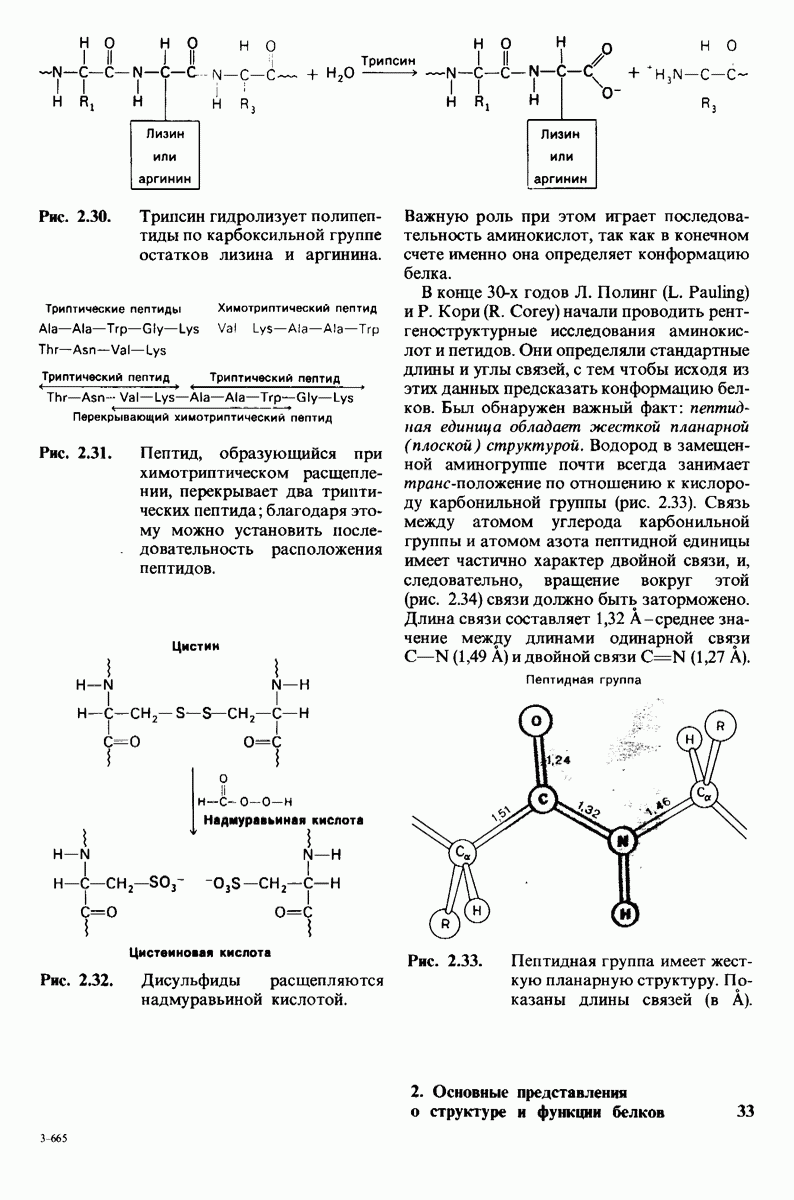
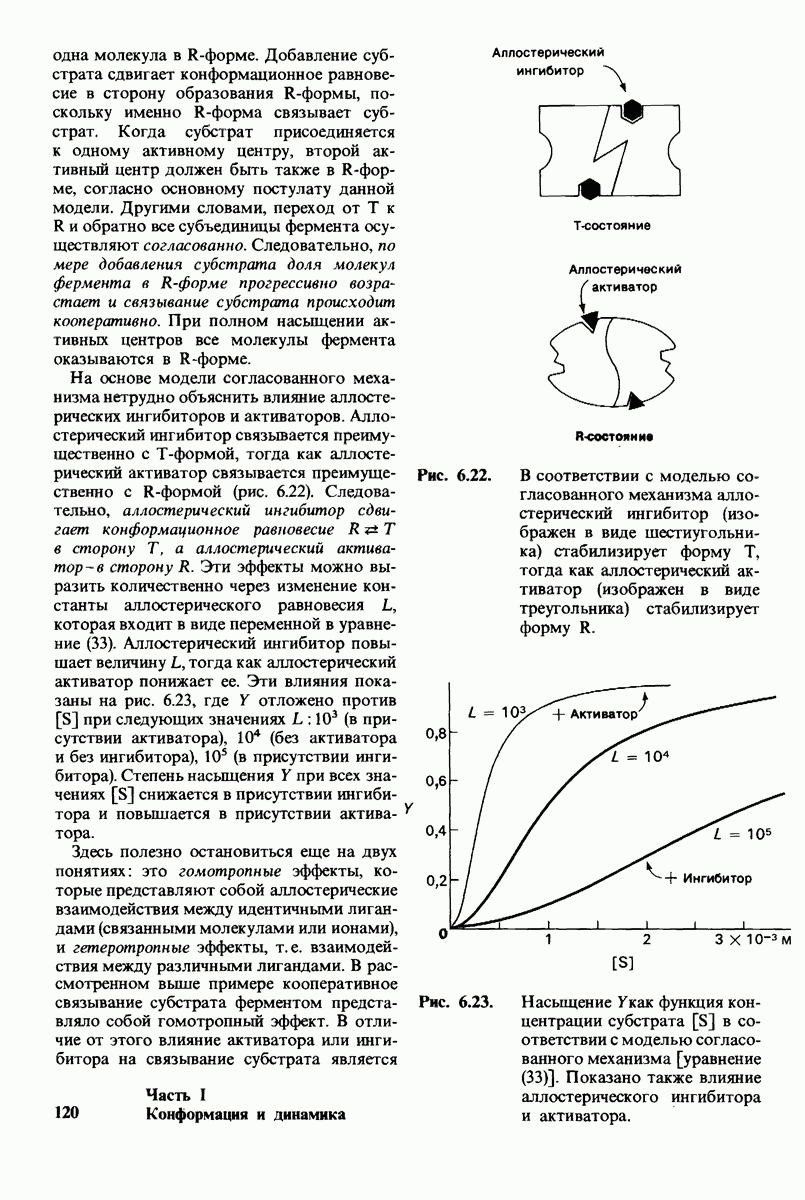
Рис. 6.21. Модель согласованного механизма кооперативного связывания субстрата аллостерическим ферментом. Присоединение первой молекулы субстрата сопровождается переходом ТТ-формы с низким сродством к субстрату в RR-форму с высоким сродством.
одна молекула в R-форме. Добавление субстрата сдвигает конформационное равновесие в сторону образования R-формы, поскольку именно R-форма связывает субстрат. Когда субстрат присоединяется к одному активному центру, второй активный центр должен быть также в R-форме, согласно основному постулату данной модели. Другими словами, переход от  к R и обратно все субъединицы фермента осуществляют согласованно. Следовательно, по мере добавления субстрата доля молекул фермента в R-форме прогрессивно возрастает и связывание субстрата происходит кооперативно. При полном насыщении активных центров все молекулы фермента оказываются в R-форме.
к R и обратно все субъединицы фермента осуществляют согласованно. Следовательно, по мере добавления субстрата доля молекул фермента в R-форме прогрессивно возрастает и связывание субстрата происходит кооперативно. При полном насыщении активных центров все молекулы фермента оказываются в R-форме.
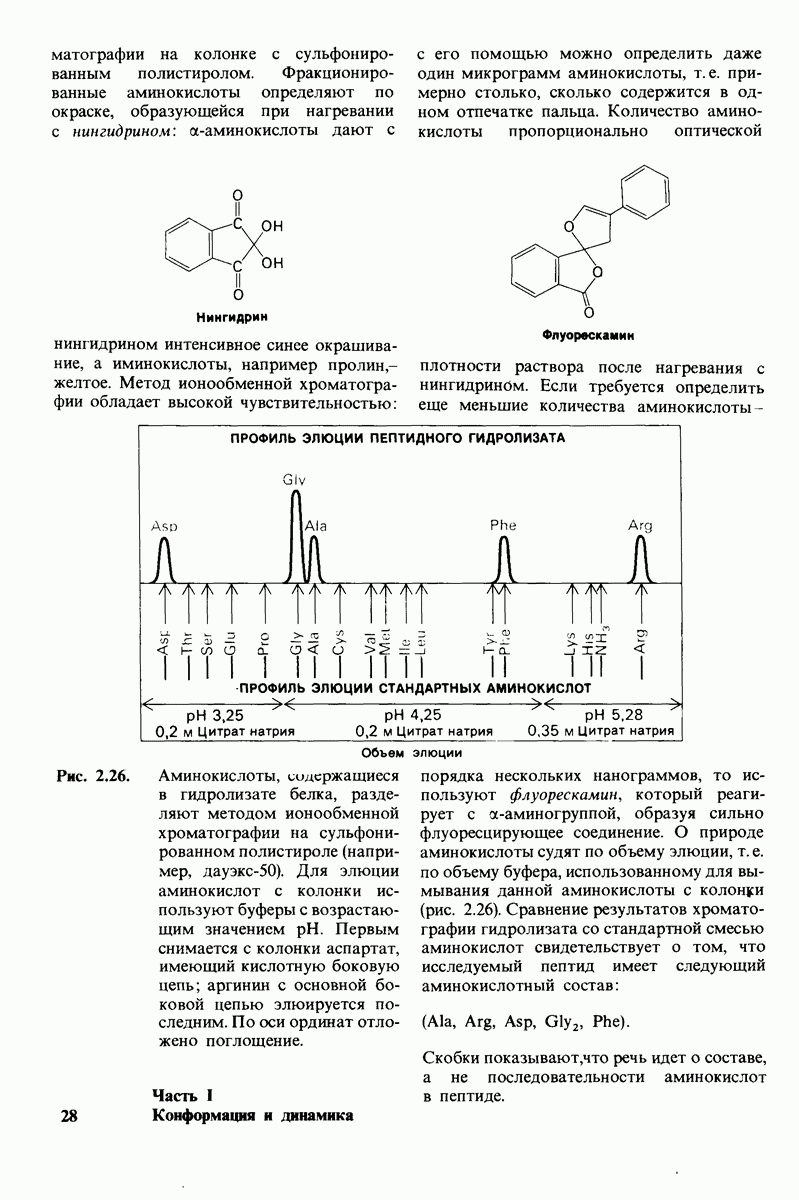
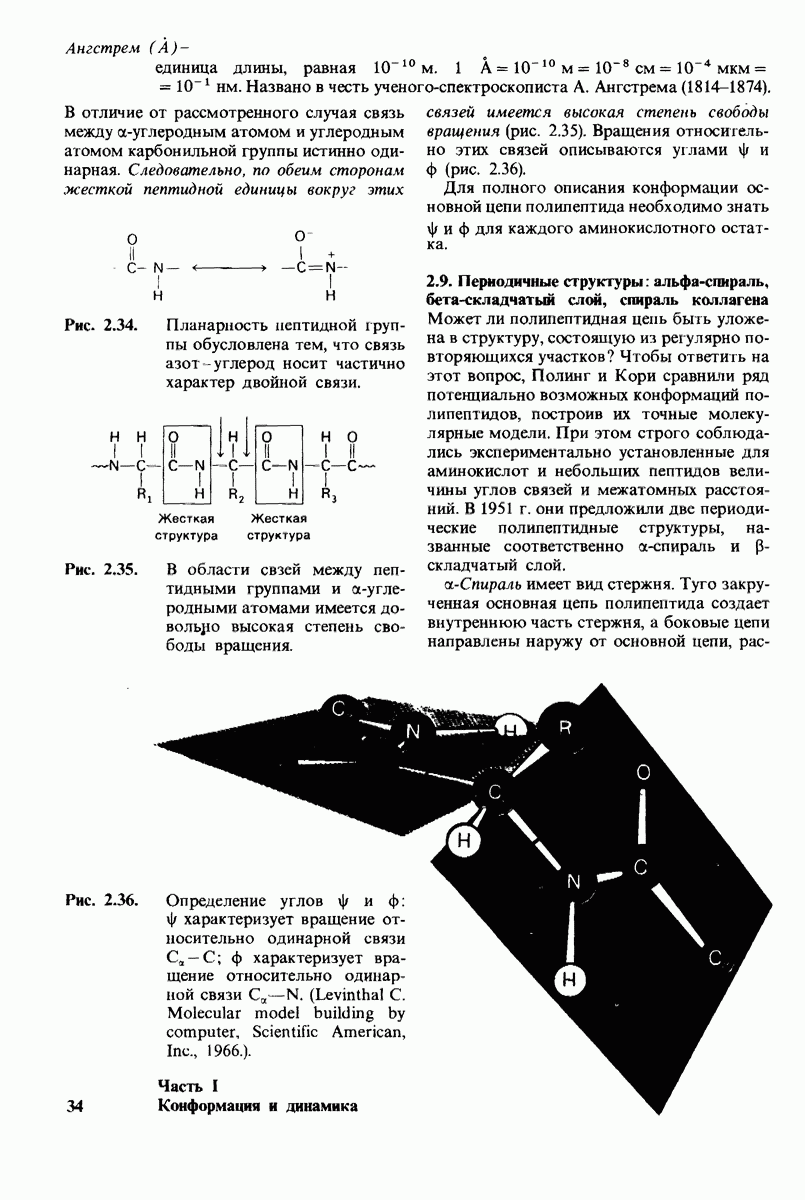
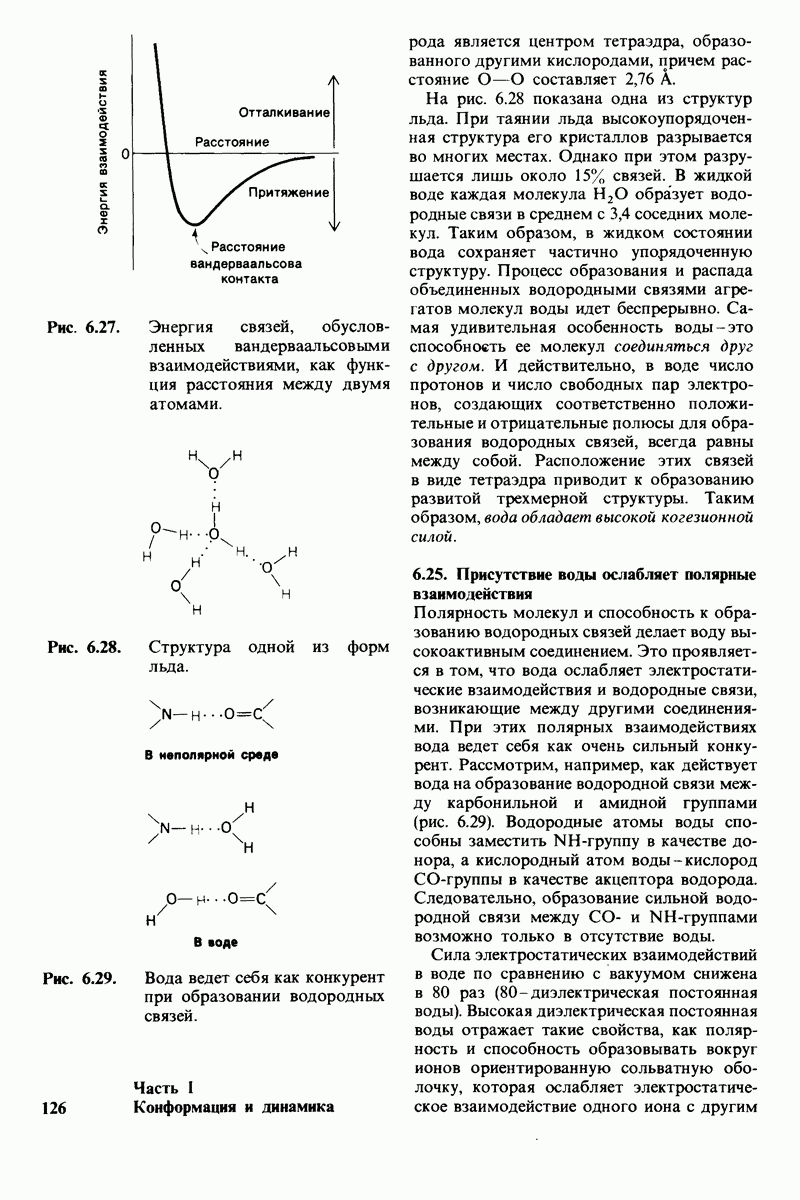
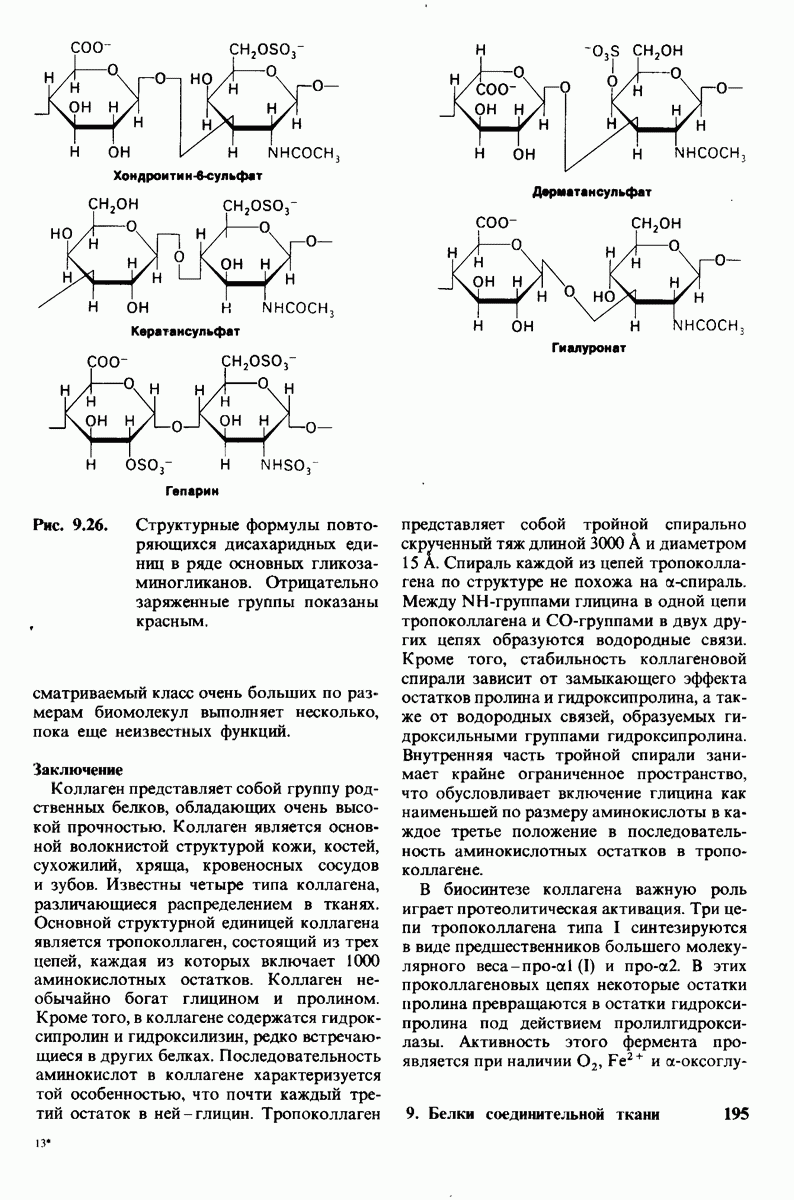
На основе модели согласованного механизма нетрудно объяснить влияние аллостерических ингибиторов и активаторов. Аллостерический ингибитор связывается преимущественно с  -формой, тогда как аллостерический активатор связывается преимущественно с R-формой (рис. 6.22). Следовательно, аллостерический ингибитор сдвигает конформационное равновесие
-формой, тогда как аллостерический активатор связывается преимущественно с R-формой (рис. 6.22). Следовательно, аллостерический ингибитор сдвигает конформационное равновесие  в сторону
в сторону  а аллостерический активаторов сторону
а аллостерический активаторов сторону  Эти эффекты можно выразить количественно через изменение константы аллостерического равновесия
Эти эффекты можно выразить количественно через изменение константы аллостерического равновесия  которая входит в виде переменной в уравнение (33). Аллостерический ингибитор повышает величину
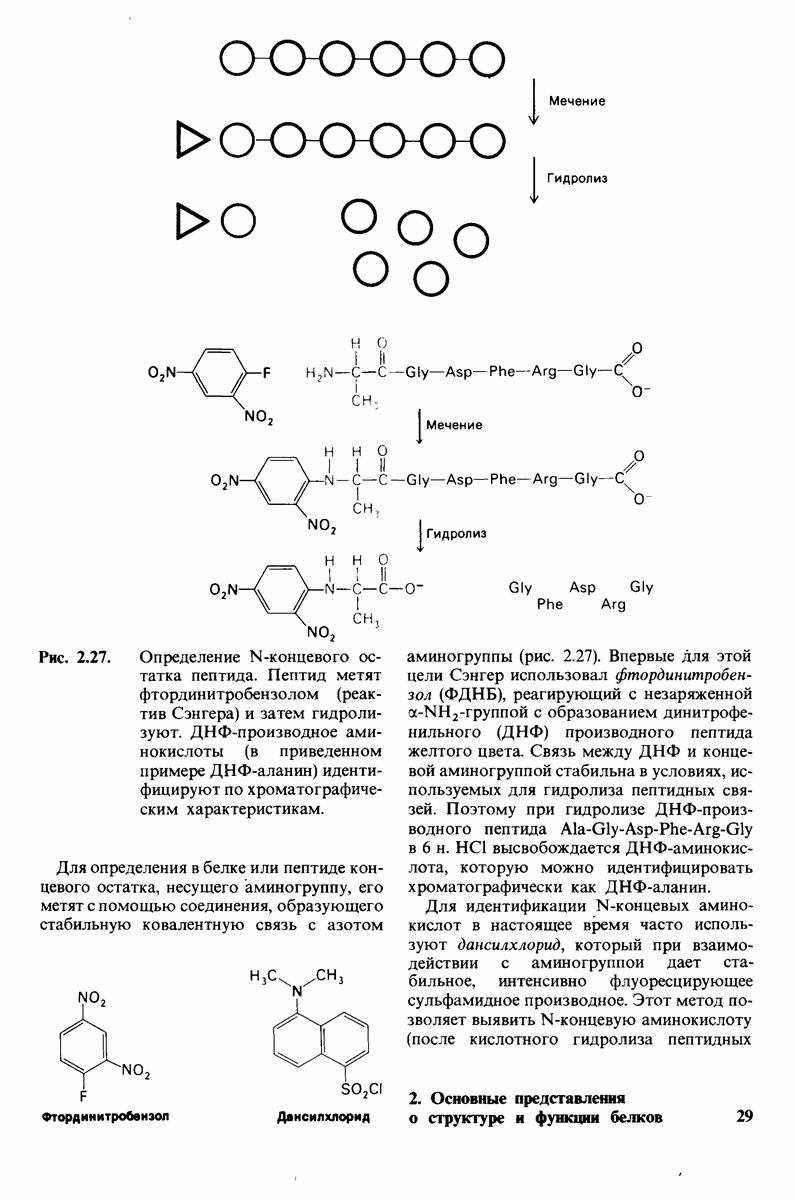
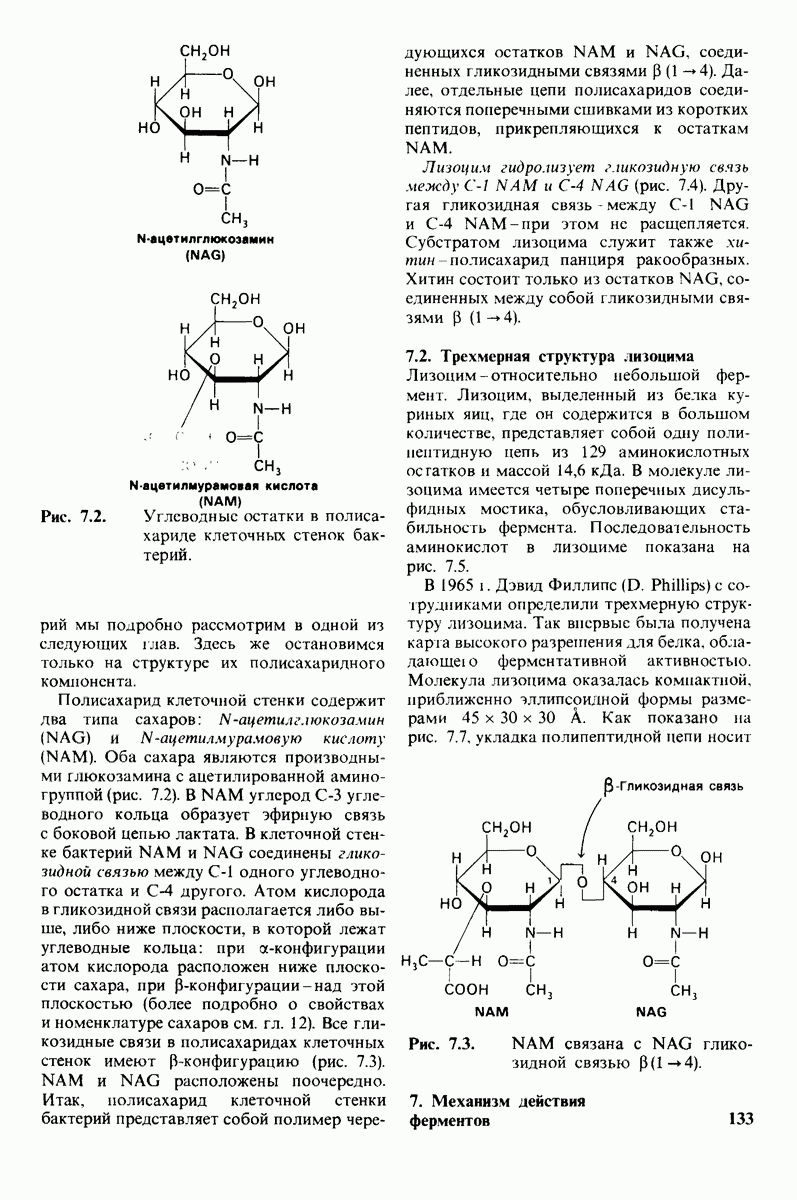
которая входит в виде переменной в уравнение (33). Аллостерический ингибитор повышает величину  тогда как аллостерический активатор понижает ее. Эти влияния показаны на рис. 6.23, где
тогда как аллостерический активатор понижает ее. Эти влияния показаны на рис. 6.23, где  отложено против
отложено против  при следующих значениях
при следующих значениях  (в присутствии активатора),
(в присутствии активатора),  (без активатора и без ингибитора),
(без активатора и без ингибитора),  (в присутствии ингибитора). Степень насыщения
(в присутствии ингибитора). Степень насыщения  при всех значениях
при всех значениях  снижается в присутствии ингибитора и повышается в присутствии активатора.
снижается в присутствии ингибитора и повышается в присутствии активатора.
Здесь полезно остановиться еще на двух понятиях: это гомотропные эффекты, которые представляют собой аллостерические взаимодействия между идентичными лигандами (связанными молекулами или ионами), и гетеротропные эффекты, т.е. взаимодействия между различными лигандами. В рассмотренном выше примере кооперативное связывание субстрата ферментом представляло собой гомотропный эффект. В отличие от этого влияние активатора или ингибитора на связывание субстрата является

Рис. 6.22. В соответствии с моделью согласованного механизма аллостерический ингибитор (изображен в виде шестиугольника) стабилизирует форму  тогда как аллостерический активатор (изображен в ввде треугольника) стабилизирует форму
тогда как аллостерический активатор (изображен в ввде треугольника) стабилизирует форму 

Рис. 6.23. Насыщение  как функция концентрации субстрата
как функция концентрации субстрата  в соответствии с моделью согласованного механизма [уравнение (33)]. Показано также влияние аллостерического ингибитора и активатора.
в соответствии с моделью согласованного механизма [уравнение (33)]. Показано также влияние аллостерического ингибитора и активатора.
гетеротропным, поскольку в этом случае взаимодействие происходит между молекулами разного типа. При согласованном механизме аллостерических взаимодействий гомотропные эффекты всегда положительны (кооперативны), а гетеротропные - либо положительны, либо отрицательны.